th2 differentiation in distinct lymph nodes influences the site of
TRANSCRIPT

of January 30, 2018.This information is current as
Immune-Inflammatory ResponsesInfluences the Site of Mucosal Th2 Th2 Differentiation in Distinct Lymph Nodes
Fattouh, Tina Walker and Manel JordanaDavid Alvarez, Janine L. Arkinson, Jiangfeng Sun, Ramzi
http://www.jimmunol.org/content/179/5/3287doi: 10.4049/jimmunol.179.5.3287
2007; 179:3287-3296; ;J Immunol
average*
4 weeks from acceptance to publicationSpeedy Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
?The JIWhy
Referenceshttp://www.jimmunol.org/content/179/5/3287.full#ref-list-1
, 18 of which you can access for free at: cites 49 articlesThis article
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 30, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Th2 Differentiation in Distinct Lymph Nodes Influences theSite of Mucosal Th2 Immune-Inflammatory Responses1
David Alvarez, Janine L. Arkinson, Jiangfeng Sun, Ramzi Fattouh, Tina Walker,and Manel Jordana2
Allergic individuals rarely present with concurrent multiple-organ disease but, rather, with manifestations that privilege aspecific site such as the lung, skin, or gastrointestinal tract. Whether the site of allergic sensitization influences the local-ization of Th2 immune-inflammatory responses and, ultimately, the organ-specific expression of disease, remains to bedetermined. In this study, we investigated whether both the site of initial Ag exposure and concomitant Th2 differentiationin specific lymph nodes (LNs) privileges Th2 memory responses to mucosal and nonmucosal sites, and whether this restrictionis associated with a differential expression in tissue-specific homing molecules. In mice exposed to Ag (OVA) via the peri-toneum, lung, or skin, we examined several local and distal LNs to determine the site of Ag-specific proliferation and Th2differentiation. Whereas respiratory and cutaneous Ag exposure led to Ag-specific proliferation and Th2 differentiationexclusively in lung- and skin-draining LNs, respectively, Ag delivery to the peritoneum evoked responses in gut-associated,as well as distal thoracic, LNs. Importantly, only mice that underwent Th2 differentiation in thoracic- or gut-associated LNsmounted Th2 immune-inflammatory responses upon respiratory or gastric Ag challenge, respectively, whereas cutaneousTh2 recall responses were evoked irrespective of the site of initial sensitization. In addition, we observed the differentialexpression of gut homing molecules (CCR9, �4, �7) in gut-associated LNs and, unexpectedly, a universal induction ofskin-related homing molecules (CCR4, CCR10) in all LNs. These data suggest that the site of initial Th2 differentiation anddifferential homing molecule expression restricts Th2 immune-inflammatory responses to mucosal, but not cutaneous,tissues. The Journal of Immunology, 2007, 179: 3287–3296.
T he skin and mucosal surfaces of the respiratory andgastrointestinal (GI)3 tract are continuously exposed toenvironmental allergens and, thus, are likely to play an
important role in allergic sensitization. In the appropriate im-munological context, allergens are captured by tissue-residentdendritic cells (DCs) and transported to the local draininglymph node (LN) where they are presented to naive T cells (1,2). Upon cognate interaction with Ag and MHC class II, naiveT cells undergo activation, proliferation, and differentiation intoCD4� Th2 cells (3). In turn, these Ag-experienced CD4� Th2cells prime B cells for IgE class-switching (4) and, then, sub-sequently traffic to the target organ where they execute theirimmune effector program (5). Indeed, the presence of allergen-
specific IgE and activated CD4� Th2 cells is a defining featureof a number of allergic diseases (6).
Although allergic diseases share a common systemic patho-genesis, allergic individuals rarely present with concurrent mul-tiple-organ disease but, rather, with manifestations at particularsites (7), such as the lung (allergic asthma), skin (allergic der-matitis), and GI tract (food allergy). This heterogeneity insin-uates that the site of initial allergen encounter may influence theorgan-specific expression of disease. In this regard, recent ev-idence suggests that the site of initial Ag exposure (8, 9) and, inparticular, the site of concomitant T cell priming in specificdraining LNs (10 –12), may imprint the ensuing immune effec-tor program with tissue-selective tropism (13–15). Indeed, ithas been shown that DCs from either skin- or gut-associatedlymphoid tissues (GALT) selectively induce skin- (CCR4�,CCR10�) or gut-(�4�7
�, CCR9�) tropic T cells that preferen-tially traffic to the skin or gut, respectively (16 –22). In thecontext of allergic disease, whether the site of initial allergenexposure privileges memory Th2 immune responses to specificmucosal and nonmucosal sites remains unknown. Moreover,whether this restriction is also associated with a differentialexpression in tissue-specific homing molecules depending onthe site (i.e., LN) of Th2 differentiation has not beendetermined.
We and others have previously shown that mice initially sen-sitized via the airways (23) or skin (24) mount Th2 immune-inflammatory responses in the lung or skin, respectively. Like-wise, it has been shown that allergic responses in the GI tractcan be induced in animals initially sensitized via the GALT(25). Yet, whether prior sensitization to Ag via one site is suf-ficient to allow for the generation of Th2 immune-inflammatoryresponses upon Ag re-exposure at any given site remains
Department of Pathology and Molecular Medicine, Division of Respiratory Diseasesand Allergy, Centre for Gene Therapeutics, McMaster University, Hamilton, Ontario,Canada
Received for publication May 4, 2006. Accepted for publication June 26, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Canadian Institutes for Health Research (CIHR).D.A. and R.F. were supported by CIHR Doctoral Research Awards. M.J. holds aCIHR-Canada Research Chair.2 Address correspondence and reprint requests to Dr. Manel Jordana, Department ofPathology and Molecular Medicine, Division of Respiratory Diseases and Allergy,Centre for Gene Therapeutics, McMaster University, Michael G. DeGroote Centre forLearning and Discovery, Room 4013, 1200 Main Street, West, Hamilton, Ontario,L8N 3Z5, Canada. E-mail address: [email protected] Abbreviations used in this paper: GI, gastrointestinal; DC, dendritic cell; LN, lymphnode; GALT, gut-associated lymphoid tissue; Ad, adenoviral; PB, peripheral blood;BAL, bronchoalveolar lavage; GG, gene gun.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

controversial (26 –28). In this study, we investigated whetherboth the site of initial Ag exposure privileges Th2 immune-inflammatory responses to mucosal and nonmucosal sites, andwhether this restriction is associated with a differential expres-sion in tissue-specific homing molecules. Using murine modelsof Th2 sensitization targeting the peritoneum (29, 30), airways(23), or skin (24), we show that the expression of Th2 immune-inflammatory responses in mucosal, but not cutaneous, tissuesis dependent on both the site of initial Ag exposure and con-comitant Th2 differentiation.
Materials and MethodsAnimals
Female BALB/c (6–8 wk old) were purchased from Charles River Labo-ratories and housed in specific pathogen-free conditions with a 12-h light-dark cycle. Experiments described in this study were approved by the An-imal Research Ethics Board of McMaster University.
OVA sensitization protocols
Th2 sensitization via the peritoneum. Sensitization via the peritoneumroute was achieved by conventional i.p. injections of OVA as previouslydescribed (29, 30). Briefly, mice were sensitized twice, 5 days apart, by i.p.injection of 8 �g of OVA (Sigma-Aldrich) adsorbed overnight at 4°C to 4mg of aluminum hydroxide (Sigma-Aldrich) in a total volume of 0.5 ml ofsterile PBS.Th2 sensitization via the airways. Sensitization via the airways wasachieved by exposure to aerosolized OVA in the presence of exogenousGM-CSF expressed via adenoviral (Ad) gene transfer to the airway, aspreviously described (23). Briefly, a replication-deficient human type 5 Advector encoding murine GM-CSF (Ad/GM-CSF) was delivered intrana-sally to isoflurane-anesthetized mice at a dose of 3 � 107 PFU 1 day beforeOVA aerosolization. Over the next 10 consecutive days (days 0–9), micewere placed in a Plexiglass chamber (10 cm � 15 cm � 25 cm) andexposed for 20 min daily to aerosolized OVA (grade V protein, 1% w/v in0.9% saline; Sigma-Aldrich). The OVA aerosol was generated by a Ben-net/Twin nebulizer at a flow rate of 10 L/min. We have previously shownthat only the concurrent exposure of OVA with Ad/GM-CSF is able togenerate Th2 sensitization, and not exposure to OVA alone, Ad/GM-CSFalone, or OVA with an empty Ad vector (23).Th2 sensitization via the skin. Sensitization via the skin was achieved byparticle bombardment of plasmid OVA-coated gold particles onto the skinusing the Helios-GG (Bio-Rad), as previously described (24). In brief, micereceive three nonoverlapping deliveries of gold particles coated with OVA-expressing plasmids, totaling 1 �g of plasmid per inoculation (i.e., 0.33 �gof DNA per delivery) onto the ventral abdominal skin on three occasionsat weekly intervals. The DNA-coated particles were prepared as describedpreviously and discharged at a helium pressure of 350 lb/inch (2). Theplasmids used were a modified version of pcDNA3.1� (Invitrogen LifeTechnologies) containing the cDNA for OVA under control of the humanCMV intermediate-early promoter. By this approach, we previously re-ported that delivery of plasmids expressing OVA leads to OVA-specificTh2 immunity, but not luciferase cDNA or empty control plasmids, ordelivery of gold particles alone (24).
Splenocyte and LN cell culture
Spleens or LNs (axillary, inguinal, mesenteric, deep cervical, super cervi-cal, thoracic, or Peyer’s patches) were excised at various time points fol-lowing Ag sensitization via the skin, peritoneum, or respiratory mucosaand placed into sterile tubes containing sterile HBSS on ice. Spleens/LNswere triturated between the ends of sterile frosted slides and the resultingcell suspension was filtered through 40-�m nylon cell strainers (Falcon;BD Biosciences), then washed at 1200 rpm for 10 min at 4°C. RBCs werelysed from spleen suspensions by adding 1 ml of ACK lysis buffer (0.5 MNH4Cl, 10 mM KHCO3, and 0.1 nM Na2EDTA (pH 7.2–7.4)) for 1 min.Splenocytes and dispersed LN cells were washed with HBSS and thenresuspended in RPMI 1640 medium supplemented with 10% FBS (Sigma-Aldrich), 1% L-glutamine, 1% penicillin/streptomycin (Invitrogen LifeTechnologies), and 0.1% 2-ME (Invitrogen Life Technologies). Cells werecultured in medium alone or with 40 �g of OVA/well and seeded at 8 �105 cells/well (spleen) or 5 � 105 cells/well (LN) in a flat-bottom, 96-wellplate (BD Biosciences). Following 120 h of culture incubation, superna-tants were harvested and stored at �20°C for cytokine analysis.
In vitro LN cell proliferation
Ag-specific in vitro proliferation was determined by a [3H]thymidine in-corporation assay. Briefly, 1 �Ci/well [3H]thymidine (PerkinElmer) wasadded to the last 18 h of a 3-day culture, in the presence or absence of OVA(40 �g/well), and proliferative responses were measured by cell uptake of[3H]thymidine. For proliferation, splenocytes and LN cells were seeded at5 � 105 cells/well. Cells were harvested using a Filtermate harvester(Packard Bioscience), quantified using TopCount NXT microplate scintil-lation and luminescence counter (Packard Bioscience), and expressed asthe mean cpm � SD of triplicate wells, where applicable.
Cytokine and Ig measurement
Cytokine/chemokine content was determined using ELISA kits purchasedfrom R&D Systems for murine IL-4, IL-5, IL-10, IL-13, IFN-�, andeotaxin. Each of these assays has a threshold of detection between 1.5 and5 pg/ml. Levels of OVA-specific serum IgE were measured using a pre-viously described Ag-capture (biotinylated OVA) ELISA method (29, 30).Units of OVA-specific IgE were determined relative to in-house standard-ized serum, obtained from mice sensitized to OVA through a conventionali.p. sensitization model for IgE standard, and expressed in units per mil-liliter relative to standard mouse sera.
OVA recall protocols
OVA recall to the gut: allergic diarrhea. Mice initially sensitized to OVAvia the skin, airways, or peritoneum, were re-exposed to OVA by theoral/GI route using a recently characterized protocol of Ag-induced allergicdiarrhea (25). Briefly, 2 wk postsensitization, mice were held in the supineposition and orally administered 50 mg of OVA protein in 250 �l of sterilePBS by gavage every other day up to seven times. Before each intragastricchallenge, mice were deprived of food for 4 h to limit the extent of Agdegradation in the stomach. Oral challenges were performed with intra-gastric feeding needles (1”, 1(1/4) mm ball diameter, 22G; Popper andSons). Unsensitized naive mice served as negative controls. Clinical scor-ing of diarrhea was assessed by visually monitoring mice for up to 1 hfollowing oral Ag challenge, by multiple observers. Mice demonstratingprofuse liquid stool were scored as positive. Seventy-two hours followingthe last (i.e., seventh) OVA challenge, mice were sacrificed and the spleenand mesenteric LNs were isolated and cultured for evidence of intact OVA-specific Th2 immunity, as detailed above. In addition, segments of thesmall intestine (jejunum) were removed, fixed in 10% formalin, embeddedin paraffin, sectioned at 3-�m thick, and stained with H&E for visualizationof eosinophils.OVA recall to the lung: allergic airways inflammation. Mice initiallysensitized to OVA via the skin, airways, or peritoneum, were re-exposed toOVA via a respiratory challenge with aerosolized OVA. Approximately 3wk postsensitization, mice were exposed to 1% OVA aerosol daily for 20min for 5 consecutive days. Unsensitized naive mice served as negativecontrols. Mice were sacrificed 72 h following the last (i.e., fifth) OVAaerosolization, and the ensuing immune response was assessed in the pe-ripheral blood (PB), bronchoalveolar lavage (BAL) fluid, and lung tissue,as described below. Moreover, thoracic LNs were removed and culturedfor evidence of intact OVA-specific Th2 immunity, as detailed above.OVA recall to the skin: late-phase cutaneous responses. Mice initiallysensitized to OVA via the skin, airways, or peritoneum, were re-exposed toOVA by intradermal injection. Approximately 3 wk postsensitization, micewere injected with 10 �g of OVA in 10 �l of sterile saline into one ear, andvehicle (sterile saline) into the opposite ear. Unsensitized naive animalsserved as negative controls. Ear thickness was measured before and atseveral time points after injection using a modified low-tension thicknessgauge (Dyer Company). Mice were sacrificed 48 h later, and ears fixed in10% formalin, then sectioned and stained with H&E.
Collection and measurement of specimens
Following OVA recall to the lung, mice were sacrificed and PB was col-lected by retro-orbital bleeding. Total white blood cell counts determinedin a blinded manner using a hemocytometer. Serum was obtained by cen-trifugation after incubating whole blood for 30 min at 37°C, and stored asaliquots at �20°C. Smears were prepared for differential cell analysis. Inaddition, BAL was performed as previously described (30). Briefly, thelungs were dissected, the trachea was cannulated with a polyethylene tube(BD Biosciences), and the lungs were lavaged twice with PBS (0.25 mlfollowed by 0.2 ml); �0.3 ml of the instilled fluid was consistently recov-ered. Total BAL cell counts were determined in a blinded manner using ahemocytometer. Each BAL sample was centrifuged, and the supernatantwas stored at �20°C for cytokine and chemokine analysis. The cell pelletwas resuspended in PBS and smears were prepared by cytocentrifugation
3288 SITE OF Th2 DIFFERENTIATION EVOKES MUCOSA-SPECIFIC RESPONSES
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
(Shandon) at 300 rpm for 2 min. PB and BAL smears were stained with theProtocol Hema 3 stain set (Fisher-Scientific). Differential cell counts of PBand BAL smears were determined in a blinded manner from at least 300–500 leukocytes using standard hemocytological criteria to classify the cellsas neutrophils, eosinophils, or mononuclear cells. Lung tissue was removedand fixed in 10% formalin and embedded in paraffin. Sections, 3-�m thick,were stained with H&E (for visualization of leukocytes and histopatholog-ical features) or periodic acid-Schiff (for detection of goblet cells and mu-cus production).
Isolation of mRNA and preparation of cDNA samples
At various time points during skin, respiratory, or i.p. sensitization to OVA,various LNs (thoracic, mesenteric, inguinal, axillary, or cervical) or Pey-er’s patches were pooled from three to four mice and stored in RNAlater(Ambion) at �20°C until processed. Total RNA was extracted from LNsusing a Polytron homogenizer with TriPure isolation reagent (Roche).Genomic DNA was then removed from the samples using the DNA-free kit(Ambion). Finally, RNA was reversed transcribed to cDNA using theRETRO-script kit (Ambion) with random decamers used as primers.
Gene expression by real-time quantitative PCR
For real-time quantitative PCR, Assays-on-Demand Gene Expression As-says containing two unlabeled PCR primers and a FAM-labeled, MGB-quenched TaqMan probe for either CCR4, CCR8, CCR9, CCR10, �4 in-tegrin chain, �7 integrin chain, or fucosyltransferase-VII were purchasedfrom Applied Biosystems. The housekeeping gene 18S rRNA were pur-chased from Applied Biosystems. PCR was run using TaqMan UniversalPCR mix (Applied Biosystems) on the ABI Prism (7900HT) SequenceDetection System and analyzed using the Sequence Detector software (ver-sion 2.1). Samples were run in triplicate with �1 �g of cDNA was addedper well as a single-plex PCR. Genes were expressed relative 18S rRNAand further normalized to background (i.e., naive) levels.
Data analysis
Data are expressed as mean � SEM, unless otherwise noted. Statisticalanalysis was performed using SigmaStat software (SPSS). Results wereinterpreted using ANOVA followed by Fisher’s least significant differencepost hoc test analysis, unless otherwise indicated. A p value of �0.05 wasconsidered statistically significant.
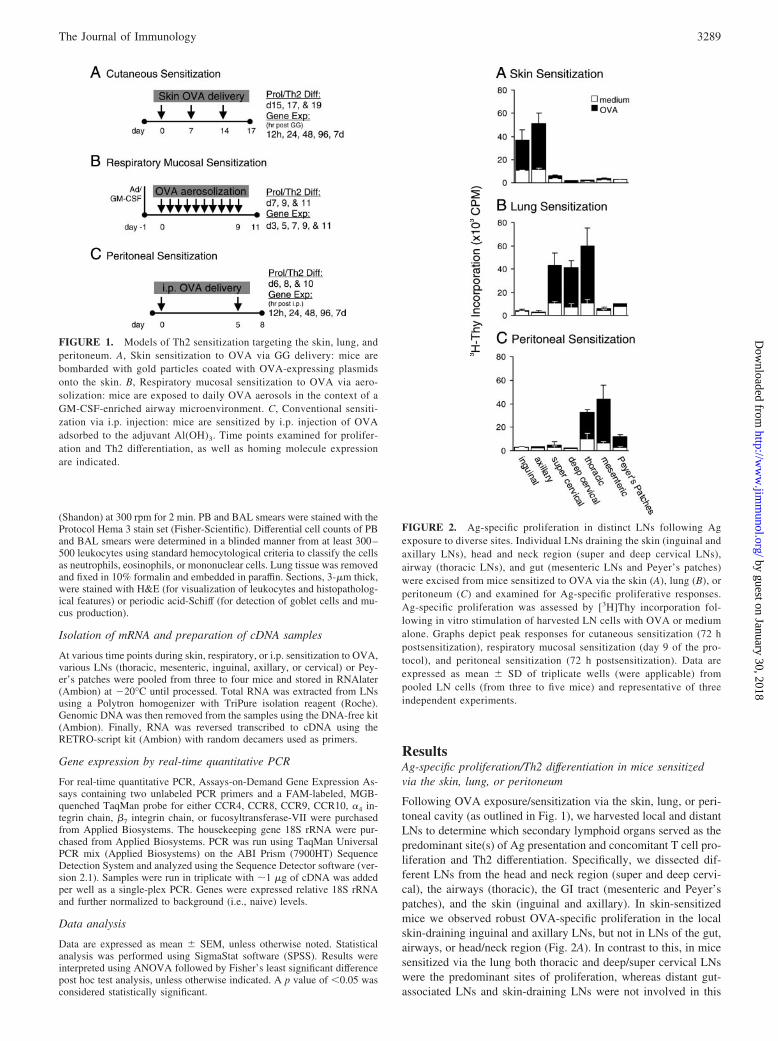
ResultsAg-specific proliferation/Th2 differentiation in mice sensitizedvia the skin, lung, or peritoneum
Following OVA exposure/sensitization via the skin, lung, or peri-toneal cavity (as outlined in Fig. 1), we harvested local and distantLNs to determine which secondary lymphoid organs served as thepredominant site(s) of Ag presentation and concomitant T cell pro-liferation and Th2 differentiation. Specifically, we dissected dif-ferent LNs from the head and neck region (super and deep cervi-cal), the airways (thoracic), the GI tract (mesenteric and Peyer’spatches), and the skin (inguinal and axillary). In skin-sensitizedmice we observed robust OVA-specific proliferation in the localskin-draining inguinal and axillary LNs, but not in LNs of the gut,airways, or head/neck region (Fig. 2A). In contrast to this, in micesensitized via the lung both thoracic and deep/super cervical LNswere the predominant sites of proliferation, whereas distant gut-associated LNs and skin-draining LNs were not involved in this
FIGURE 1. Models of Th2 sensitization targeting the skin, lung, andperitoneum. A, Skin sensitization to OVA via GG delivery: mice arebombarded with gold particles coated with OVA-expressing plasmidsonto the skin. B, Respiratory mucosal sensitization to OVA via aero-solization: mice are exposed to daily OVA aerosols in the context of aGM-CSF-enriched airway microenvironment. C, Conventional sensiti-zation via i.p. injection: mice are sensitized by i.p. injection of OVAadsorbed to the adjuvant Al(OH)3. Time points examined for prolifer-ation and Th2 differentiation, as well as homing molecule expressionare indicated.
FIGURE 2. Ag-specific proliferation in distinct LNs following Agexposure to diverse sites. Individual LNs draining the skin (inguinal andaxillary LNs), head and neck region (super and deep cervical LNs),airway (thoracic LNs), and gut (mesenteric LNs and Peyer’s patches)were excised from mice sensitized to OVA via the skin (A), lung (B), orperitoneum (C) and examined for Ag-specific proliferative responses.Ag-specific proliferation was assessed by [3H]Thy incorporation fol-lowing in vitro stimulation of harvested LN cells with OVA or mediumalone. Graphs depict peak responses for cutaneous sensitization (72 hpostsensitization), respiratory mucosal sensitization (day 9 of the pro-tocol), and peritoneal sensitization (72 h postsensitization). Data areexpressed as mean � SD of triplicate wells (were applicable) frompooled LN cells (from three to five mice) and representative of threeindependent experiments.
3289The Journal of Immunology
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

route of sensitization (Fig. 2B). Finally, Ag delivery to the peri-toneal cavity led to marked proliferation in the gut-associated mes-enteric LNs and to a lesser extent in the Peyer’s patches, as well asthe thoracic LNs, but not in the LNs draining the skin or head andneck region (Fig. 2C). The magnitude of the proliferative responsewas comparable between the principal LNs activated in each of thedifferent models, with an �4- to 6-fold increase in proliferation(i.e., ratio of OVA cpm to medium cpm), with the exception of thePeyer’s patches which consistently showed a �3-fold induction inproliferation following i.p. Ag delivery.
To determine the site(s) of Th2 differentiation, we assayedculture supernatants for the production of Th2-affiliated cyto-kines (IL-4, IL-5, and IL-13) by Ag-stimulated LN cells invitro. Consistent with the proliferation data, mice sensitized viathe skin, lung, or peritoneum resulted in robust Th2-cytokineproduction in the relevant draining LNs. Specifically, Th2 dif-ferentiation was restricted to the skin-draining LNs in skin-sensitized mice, and the thoracic and cervical LNs in respiratorymucosal-sensitized mice (Fig. 3, A and B). Moreover, in i.p.-sensitized mice Th2 differentiation occurred in the gut-associ-ated mesenteric LNs and Peyer’s patches, as well as the thoracicLNs (Fig. 3C). Examination of culture supernatants for IL-10levels resulted in a similar profile, with enhanced IL-10 pro-duction solely in local draining, but not distant, LNs in each ofthe sensitization protocols (skin sensitization: 730 pg/ml ingui-nal/axillary LNs; lung sensitization: 750 pg/ml deep cervical
LNs, 914 pg/ml super cervical LNs, and 1459 pg/ml thoracicLNs; i.p. sensitization: 2259 pg/ml thoracic LNs and 2772pg/ml mesenteric LNs).
Systemic Th2 cytokine and IgE production
In addition to characterizing the response in individual LNs duringsensitization, we also determined the contribution of the spleen
FIGURE 3. Site of initial Ag ex-posure leads to Th2 differentiation indistinct LNs. Individual LNs drainingthe skin (inguinal and axillary LNs),head and neck region (super and deepcervical LNs), airway (thoracic LNs),and gut (mesenteric LNs and Peyer’spatches) were excised from mice sen-sitized to OVA via cutaneous (A), re-spiratory mucosal (B), or peritoneal(C) routes and examined for Ag-spe-cific Th2 cytokine (IL-4, IL-5, andIL-13) production following in vitrostimulation of harvested LN cellswith OVA or medium alone. Datashow pooled cytokine levels frompooled LN cells (from three to sixmice) and representative of three in-dependent experiments.
Table I. OVA-specific serum IgE and splenocyte-derived Th2 cytokineproduction following skin, respiratory, or i.p. sensitization in micea
SensitizationOVA
Specific
Splenocyte In Vitro Recallb
IL-4 IL-5 IL-13
SiteSerumIgEc (pg/ml) (pg/ml) (ng/ml)
Skin 90 � 19 234 � 42 58 � 76 8819 � 638Respiratory 69 � 11 85 � 12 3374 � 665d 10675 � 2911Peritoneal 125 � 38 292 � 34e 3462 � 910d 18984 � 7235
a Mice were sensitized to OVA via the skin, lung, or peritoneum and sacrificed atvarious time points as indicated in Fig. 1. Data are expressed as mean � SEM; n �3–4/group.
b Cytokine levels in culture supernatants (OVA stimulated).c OVA-specific serum IgE was measured by ELISA and is expressed as units per
milliliter.d Value of p � 0.05 compared to cutaneous.e Value of p � 0.05 compared to respiratory.
3290 SITE OF Th2 DIFFERENTIATION EVOKES MUCOSA-SPECIFIC RESPONSES
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

compartment in each of the models of sensitization. To this end,we harvested splenocytes from skin-, lung-, or i.p.-sensitized miceand pulsed them with OVA in vitro to examine Th2 cytokine (IL-4,
IL-5, and IL-13) production. As shown in Table I, we observedOVA-specific Th2 cytokine production by cultured splenocytesregardless of the site of initial Ag exposure. Of note, mice
FIGURE 4. Intragastric Ag challenge elicits allergic GI responses in mice initially sensitized via the GALT. Mice were initially sensitized to OVA viathe skin, airways, or peritoneum, and subsequently challenged with OVA by intragastric gavage. Mice were challenged with OVA (50 mg) seven times,every other day, and scored for the occurrence (%) of Ag-induced allergic diarrhea (A) as assessed up to 1 h postchallenge (n � 6–8). B, Mesenteric LNswere excised 72 h after the final challenge and examined for Th2 cytokine production in vitro. Data in B represent mean � SEM (n � 3). C, RepresentativeH&E sections of jejunum tissue following intragastric challenge with OVA in unsensitized naive (i), skin-sensitized (ii), airway-sensitized (iii), ori.p.-sensitized (iv) mice. Arrowheads depict eosinophils. Original magnification of panels, �200. Original magnification of insets, X600 oil immersion. D,Th2 cytokine production by in vitro stimulated splenocytes.
3291The Journal of Immunology
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

sensitized by the i.p. route produced slightly higher levels ofTh2 cytokines and OVA-specific serum IgE than mice sensi-tized via the skin or lung. Mice sensitized to OVA via the lungalso produced high levels of Th2 cytokines compared with skin-sensitized mice; however, the levels of serum IgE were lowest.
Intragastric OVA recall in mice sensitized via the skin, lung, orperitoneum
We next investigated whether both the site of initial Ag exposureand concomitant Th2 differentiation in specific LNs privileged theensuing Th2 memory response to specific tissue compartments. Tothis end, mice were sensitized to OVA via the skin, lungs, or peri-toneum, and then re-exposed to OVA by oral gavage using a re-cently characterized protocol of Ag-induced GI inflammation andacute allergic diarrhea (see Materials and Methods; Ref. 25). De-spite unequivocal signs of Th2 immunity in mice previously sen-sitized to OVA via the skin or lung (Fig. 3, A and B), only micethat were initially sensitized to OVA via the peritoneal cavity de-veloped OVA-induced allergic diarrhea (Fig. 4A). Indeed, i.p.-sen-sitized mice started developing allergic diarrhea after the third in-tragastric OVA challenge and, by the seventh exposure, all miceshowed signs of GI dysfunction, including obvious signs of dis-comfort (data not shown). We also harvested mesenteric LN cells72 h following the last (i.e., seventh) OVA challenge and stimu-lated them in vitro with OVA to determine whether a Th2 immuneresponse was been developed (Fig. 4B). We found a pronouncedincrease in Th2 cytokine production by OVA-stimulated mesen-teric LN cells from diarrhea-positive and i.p.-sensitized mice. Im-portantly, mesenteric LN cells harvested from diarrhea-negativeand skin- or respiratory mucosal-sensitized mice also producedTh2 cytokines, but at significantly lower levels. We also examined,histopathologically, jejunum segments of the small intestine 72 hfollowing the last oral challenge, and observed a slightly greaterinflux of eosinophils in diarrhea-positive/i.p.-sensitized mice com-pared with diarrhea-negative and skin- or respiratory mucosal-sen-sitized mice, or unsensitized naive controls (Fig. 4C). Despite be-ing unresponsive to intragastric OVA challenge, splenocytesharvested from skin- and respiratory mucosal-sensitized mice pro-duced very similar, if not higher, Th2 cytokines compared withdiarrhea-positive/i.p.-sensitized mice (Fig. 4D).
Respiratory OVA recall in mice sensitized via the skin, lung, orperitoneum
Next, we determined whether mice initially sensitized via the skin,lungs, or peritoneum, were equally able to mount OVA-specificTh2 immune-inflammatory responses in the lung upon re-exposureto aerosolized OVA (Fig. 5). Mice sensitized to OVA via the lungmounted a robust inflammatory response upon respiratory OVArecall including a marked increase in total cells and eosinophils inthe BAL compartment (Fig. 5A). This was accompanied by anincrease in PB eosinophils and BAL eotaxin levels (Fig. 5, B andC), a massive peribronchial and perivascular eosinophilic infiltratein the lung tissue, and pronounced goblet cell hyperplasia and mu-cus production (Fig. 5Eiii). Likewise, mice sensitized via the peri-toneum also mounted a robust Th2 immune-inflammatory re-sponse in the airways, which was both qualitatively andquantitatively similar to mice initially sensitized via the lung (Fig.5E, iii and iv). Consistent with our previous findings (24), skin-sensitized mice failed to mount allergic responses upon OVA aero-solization. Indeed, the response elicited by skin-sensitized micewas similar to that of unsensitized naive controls (Fig. 5E, i and ii).
We also examined whether respiratory OVA recall provokedTh2 immune activity in local thoracic LNs. Indeed, mice initiallysensitized via the lung or peritoneum experienced a massive
FIGURE 5. Respiratory Ag challenge elicits allergic airways inflam-mation in mice initially sensitized via the thoracic LNs. Mice wereinitially sensitized to OVA via the skin, airways, or peritoneum, andsubsequently challenged with OVA by aerosolization. Mice were chal-lenged daily with 1% OVA aerosol for 20 min for 5 consecutive daysthen sacrificed 72 h later. A, The number of total cells, mononuclearcells and eosinophils as assessed in the BAL. B, Total circulating cellsand eosinophils in the PB. C, Eotaxin levels in BAL fluid. D, Total LNcells and Th2-cytokine production (IL-4) following aerosolized OVAchallenge. Thoracic LNs were excised 72 h postchallenge, enumerated,and stimulated in vitro for IL-4 production. Data are pooled from threeto four mice per group. E, Light photomicrographs of lung tissuestained with H&E or periodic acid-Schiff (insets) following respiratorychallenge with OVA in unsensitized naive (i), skin-sensitized (ii), air-way-sensitized (iii), or i.p.-sensitized (iv) mice. Original magnificationof panels, �50. Original magnification of insets, �400.
3292 SITE OF Th2 DIFFERENTIATION EVOKES MUCOSA-SPECIFIC RESPONSES
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

expansion in total thoracic LN cells upon respiratory OVA recall,unlike skin-sensitized mice or naive controls (Fig. 5D). UponOVA stimulation in vitro, thoracic LN cells from mice initiallysensitized via the lung or peritoneum produced enhanced levels ofTh2-associated cytokines (IL-4) compared with skin-sensitized ornaive mice (Fig. 5D).
Cutaneous OVA recall in mice sensitized via the skin, lung, orperitoneum
Next, we investigated whether OVA recall to the skin could leadto cutaneous Th2 immune-inflammatory responses irrespective ofthe site of initial Ag exposure or sensitization. To this end, micewere initially sensitized to OVA via the skin, lung, or peritoneum(or unsensitized), and 3 wk later exposed to OVA by dermal in-jection into the ear. In skin-sensitized mice, we observed a prom-inent late-phase cutaneous response as determined by a significantincrease in ear thickness following OVA, but not vehicle, injection(Fig. 6A). Moreover, this was accompanied by a marked influx ofeosinophils into the ear tissue (Fig. 6Bii). Likewise, mice initiallysensitized to OVA via the lung or peritoneum were able to mountlate-phase cutaneous responses upon dermal challenge with OVA,as indicated by an increase in ear thickness and eosinophil infil-
tration. Histopathologically we also observed a marked influx ofeosinophils into ear tissue challenged with OVA (but not saline orunsensitized naive mice challenged with OVA) in respiratory ori.p.-sensitized mice (Fig. 6B, iii and iv).
Differential expression of tissue-specific homing molecules indistinct LNs
The differential expression of unique homing molecules on variousT cell subsets is a defining feature in the establishment of tissue-selective trafficking patterns. To determine whether sensitizationvia diverse sites (skin, lung, or peritoneum) led to a differentialinduction of tissue-specific homing molecules in vivo, we exam-ined the expression of several putative skin-tropic (CCR4, CCR8,and CCR10) and gut-tropic (CCR9, �4 integrin, �7 integrin) mol-ecules. To this end, the principal draining LNs from each sensiti-zation protocol (see Fig. 1) were excised at various time points andexamined for the differential expression of tissue-homing mole-cules. In mice sensitized via the skin, we excised axillary LNs atseveral time points post the third gene-gun (GG) delivery, andobserved a selective induction of CCR4 and CCR10, but not thegut-tropic molecules (CCR9, �4 integrin, or �7 integrin; Fig. 7A).We did not observe an up-regulation in CCR8. Moreover, in mice
FIGURE 6. Skin Ag challengeelicits late-phase cutaneous Th2 im-mune-inflammatory responses. Micewere initially sensitized to OVA viathe skin, airways, or peritoneum, orunsensitized (naive) and subsequentlychallenged with OVA (or saline) byintradermal injection. A, Ear thick-ness measurements following OVA(filled symbols) or saline (open sym-bols) injection at the indicated timepoints. B, Light photomicrographs ofH&E-stained cross-sections of ear tis-sue 48 h postchallenge. All panelsrepresent sections obtained followingdermal OVA challenge in naive (i),skin-sensitized (ii), respiratory-sensi-tized (iii), or i.p.-sensitized (iv) mice.Arrows depict eosinophils. Originalmagnification of panels, �50. Origi-nal magnification of insets, �400.
3293The Journal of Immunology
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

sensitized to OVA via the lung, we also observed an induction ofskin-tropic CCR4 and CCR10, but not gut-tropic molecules, inthoracic LNs (Fig. 7B). In contrast, in mesenteric LNs of micesensitized to OVA via the peritoneum, we observed the inductionof gut-tropic molecules (both �4 and �7 integrins, as well asCCR9; Fig. 7C). Again, we also observed an increase in the ex-pression of the skin-tropic homing molecules (CCR4, CCR8,CCR10; Fig. 7C).
DiscussionIn this study, we investigated whether the localization of memoryTh2 immune responses to specific mucosal and nonmucosal siteswas dependent on both the site of initial allergen exposure andconcomitant Th2 differentiation in specific draining LNs. We showthat cutaneous and respiratory Ag delivery evoked responses ex-clusively in the skin and cervical/thoracic LNs, respectively,whereas i.p. Ag exposure induced responses both in the gut-drain-ing LNs and, importantly, also in the thoracic LNs. This latterresponse is consistent with several reports showing that the i.p.route also drains to the thoracic LNs (31). Despite differences inthe Ag delivery systems (i.e., aerosol, injection, and GG) the en-suing immune response was distinctly Th2 in nature. The differ-
ences in the levels of Th2 cytokine production between protocolsmay be attributable to kinetics, a greater number of OVA-specificT cell clones induced in one model vs the other, or whether thereare differences in the phenotype of the Th2 cells induced. It isknown, however, that the generation of Th2 differentiation can bedifferentially programmed by specific cytokines (IL-4, IL-10, andIL-13) depending on the site of sensitization (32–34).
Next, we determined whether the site of initial Th2 differenti-ation in specific LNs influenced the localization of memory Th2immune-inflammatory responses. We found, that allergic GI re-sponses were clearly induced only in mice initially sensitized viathe peritoneum. In fact, despite evidence of systemic Th2 immu-nity in cutaneous- and respiratory mucosal-sensitized mice, thesemice were unable to develop GI manifestations upon intragastricAg challenge. Previous studies have demonstrated that oral Ag-induced diarrhea and allergic GI responses in mice are dependenton the induction of Th2 immunity including GI inflammation me-diated by IgE-primed mast cells, eosinophils, and CD4� Th2 cells(25, 35, 36). In those studies, disease-positive animals had alsobeen sensitized systemically although the precise involvement ofspecific LNs during sensitization was not examined (25). In ourstudy, the ability to mount allergic GI responses correlated not
FIGURE 7. Differential expression of tissue-specific homing molecules in skin, lung, and gut-draining LNs during sensitization. Mice were exposed toOVA via the skin, lung, or peritoneum and at various time points, RNA was isolated from the relevant skin, lung, or gut-draining LNs and examined forthe expression of gut- (CCR9, �4�7 integrins) or skin- (CCR4, CCR8, CCR10) tropic homing molecules by quantitative PCR. Relative expression of CCR4,CCR8, CCR9, CCR10, �4 integrin, and �7 integrin in skin (axillary)- (A), lung (thoracic)- (B), or gut (mesenteric)- (C), draining LNs. Data are expressedas fold increase over naive, and represent pooled means of triplicate measurements relative to housekeeping gene (18S rRNA).
3294 SITE OF Th2 DIFFERENTIATION EVOKES MUCOSA-SPECIFIC RESPONSES
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

with the existence of Th2 immunity per se, but rather with Th2differentiation having occurred initially in the gut-draining LNswhich we observed only in mice sensitized via the peritoneum.Notably, mice initially sensitized via the lung or skin, albeit un-responsive to oral Ag challenge, also showed signs of Th2 immuneresponsiveness in the mesenteric LNs. However, the levels weremarkedly lower and, perhaps, of insufficient strength (or below acertain threshold) to establish effector Th2 immunity in the GImucosa. That the levels of serum OVA-specific IgE were not sig-nificantly different in mice sensitized via peritoneum comparedwith skin- or respiratory-sensitized mice argues against diminutivesensitization in the latter groups. Moreover, while mesenteric LNcytokine responses (post-oral Ag challenge) were highest in thei.p.-sensitized and disease-positive mice (Fig. 4B), markers of sys-temic Th2 immunity (i.e., spleen-derived Th2 cytokine production)during intragastric Ag challenge were on par between all mice(Fig. 4D).
Similar to the induction of GI manifestations, airway inflamma-tion only developed in mice where Th2 differentiation had oc-curred in the thoracic, but not skin, draining LNs. Although it isnot surprising that i.p.-sensitized mice mounted allergic airwaysresponses because, as previously mentioned, thoracic LNs are in-volved during priming, it is unexpected, however, that priming viathe respiratory mucosa can prime for effector responses in the air-way but cannot lead to effector responses at the GI mucosa. Thesedata are at variance with the view of a common mucosal system(37–39), but consistent with a growing number of reports showingthat immunization at one mucosal site does automatically lead toprotective immunity in all other mucosal organs (40, 41). In con-trast to the selective expression of mucosal Th2 responses in the GItract or lung, late-phase cutaneous responses were similarly pro-voked in mice regardless of the site of initial Ag exposure, despitethe complete absence of involvement of skin-draining LNs duringi.p. or respiratory sensitization.
These data are particularly intriguing in light of recent evidencethat the tissue origin of DCs (8) and the specific LN microenvi-ronment (10–12) imprint T and B cells with tissue-specific homingmolecules (16, 19, 42, 48). In this regard, we found that gut-tropichoming molecules (CCR9 and �4�7 integrins) were differentiallyup-regulated in mesenteric LNs of i.p.-sensitized mice, but not inthe major draining LNs of skin- or respiratory-sensitized mice. Asyet, chemokine receptors/integrins specific for T cells homing tothe lung are not well defined (43). Of particular significance, wefound that regardless of site of initial Th2 differentiation, skin-tropic molecules (CCR4 and CCR10) were uniformly up-regulatedin all LNs. This is consistent with recent reports suggesting thatactivated T cells acquire skin tropism by default unless they en-counter signals from distinct DC subsets, at which point they altertheir expression pattern of homing molecules (12). The pervasiveability to mount cutaneous responses observed in our studies, de-spite no involvement of cutaneous LNs in two of the models stud-ied supports the concept of skin tropism by default. Alternatively,higher numbers of Ag-specific T cells evoked in some modelscould make available more Ag-specific clones to respond distally;a notion consistent with a “selection” view of T cell homing (49).Moreover, the use of different adjuvants during Ag delivery maypromote DC migration to distal LNs or spleen, thereby inducingdiverse homing phenotypes. Indeed, restricting Ag delivery andDC traffic to local LNs may ultimately limit pervasive responses asdiscussed below.
In our study, there are a number of variables, including the for-mulation of Ag (DNA vs protein), dose, and adjuvant (alum, GM-CSF, and gold particles) that differ depending on the protocol. Thetype of adjuvant used during Ag delivery has dramatic effects on
the migratory capacity of DCs from the site of Ag exposure tolocal and even distal LNs (44). Indeed, transcutaneous immuniza-tion strategies incorporating bacterial adjuvants like cholera toxinhas been shown to enhance migration of skin-derived DCs to im-mune-inductive sites of the gut mucosa, like the Peyer’s patches, toelicit robust mucosal CTL responses (44). It remains to be deter-mined whether the different adjuvants used in our study stimulateDC migration beyond local LNs. A number of reports have shownthat Ag sensitization via the skin primes for subsequent Th2 im-mune responses in the lung (26, 27, 34) and the systemic com-partment (45–47). It is unknown whether the Ag delivery modes inthose studies exclusively confined the Ag to specific tissues and,more importantly, LNs. For the most part, these studies did notspecifically set out to investigate the compartmentalization of Th2immunity and, consequently, a comprehensive examination of thesites of Th2 differentiation in local and distal LNs is lacking. In ourstudy, we have incorporated two models of Th2 sensitization thattarget Ag to the respiratory tract or skin. Importantly, these modelsalso generated responses in the spleen compartment, either by Agleakage into the circulation or via DC transport. Although, blood-borne Ags captured in the marginal zone of the spleen can beeventually presented to naive T cells, it is unknown whether T cellsprimed in that compartment would be imprinted with any tissue-selective homing program. It is our view that the GG Ag-deliverysystem, in particular, is better suited, but by no means perfect, tostudy the compartmentalization of immune responses, comparedwith alternatives like aerosolization, skin painting, or i.p. priming.Vast improvements in targeting and controlling the expression anddose of Ag in specific peripheral and, ideally, lymphoid tissuesthrough the use of tissue-specific and inducible promoters will pro-vide us with the necessary tools to begin to uncover the mecha-nisms underlying immune compartmentalization in vivo.
In summary, our data argue for the compartmentalization of Th2immune responses at mucosal sites, and the pervasive or defaultability to mount cutaneous responses. What are the physiologicalimplications of these findings? That the immune system is de-signed to compartmentalize the effector response to the site of Agentry hinges on the imprinting of particular sets of tissue-selectivehoming molecules in the local LN microenvironment. Indeed, theprecise manner by which lymphocytes selectively traffic to specifictissues may be overridden under circumstances where Ags perme-ate beyond regional immune control and drift systemically, aswould be the case for live pathogens. Such adaptation of defaultimmune surveillance patterns not only provides for a more tailoredimmune effector response, it may also be, in itself, reflective of thedanger associated with the Ag. In the context of allergic disease,perhaps the prerequisite for developing multiple organ manifesta-tions arises not through allergen exposure per se, but rather asallergic sensitization becomes increasingly systemic. It is underthese conditions where severity overwhelms the need for finesseand places allergic GI responses at the far end of the spectrum ofallergic disease.
AcknowledgmentsWe extend our thanks to Susanna Goncharova, Katherine Arias, and BobaeKim for technical assistance. We also gratefully acknowledge the secre-tarial assistance of Mary Kiriakopoulos.
DisclosuresThe authors have no financial conflict of interest.
References1. Randolph, G. J., V. Angeli, and M. A. Swartz. 2005. Dendritic-cell trafficking to
lymph nodes through lymphatic vessels. Nat. Rev. Immunol. 5: 617–628.
3295The Journal of Immunology
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

2. Cyster, J. G. 2005. Chemokines, sphingosine-1-phosphate, and cell migration insecondary lymphoid organs. Annu. Rev. Immunol. 23: 127–159.
3. Seder, R. A., and R. Ahmed. 2003. Similarities and differences in CD4� andCD8� effector and memory T cell generation. Nat. Immunol. 4: 835–842.
4. DeKruyff, R. H., T. Turner, J. S. Abrams, M. A. Palladino, Jr., and D. T. Umetsu.1989. Induction of human IgE synthesis by CD4� T cell clones: requirement forinterleukin 4 and low molecular weight B cell growth factor. J. Exp. Med. 170:1477–1493.
5. Oxenius, A., R. M. Zinkernagel, and H. Hengartner. 1998. CD4� T-cell inductionand effector functions: a comparison of immunity against soluble antigens andviral infections. Adv. Immunol. 70: 313–367.
6. Larche, M., D. S. Robinson, and A. B. Kay. 2003. The role of T lymphocytes inthe pathogenesis of asthma. J. Allergy Clin. Immunol. 111: 450–463, quiz 464.
7. Szefler, S. J. 2002. The natural history of asthma and early intervention.J. Allergy Clin. Immunol. 109: S549–S553.
8. Dudda, J. C., J. C. Simon, and S. Martin. 2004. Dendritic cell immunization routedetermines CD8� T cell trafficking to inflamed skin: role for tissue microenvi-ronment and dendritic cells in establishment of T cell-homing subsets. J. Immu-nol. 172: 857–863.
9. Campbell, D. J., and E. C. Butcher. 2002. Rapid acquisition of tissue-specifichoming phenotypes by CD4� T cells activated in cutaneous or mucosal lymphoidtissues. J. Exp. Med. 195: 135–141.
10. Dudda, J. C., A. Lembo, E. Bachtanian, J. Huehn, C. Siewert, A. Hamann,E. Kremmer, R. Forster, and S. F. Martin. 2005. Dendritic cells govern inductionand reprogramming of polarized tissue-selective homing receptor patterns of Tcells: important roles for soluble factors and tissue microenvironments. Eur.J. Immunol. 35: 1056–1065.
11. Mora, J. R., M. R. Bono, N. Manjunath, W. Weninger, L. L. Cavanagh,M. Rosemblatt, and U. H. Von Andrian. 2003. Selective imprinting of gut-hom-ing T cells by Peyer’s patch dendritic cells. Nature 424: 88–93.
12. Mora, J. R., G. Cheng, D. Picarella, M. Briskin, N. Buchanan, andU. H. von Andrian. 2005. Reciprocal and dynamic control of CD8 T cell homingby dendritic cells from skin- and gut-associated lymphoid tissues. J. Exp. Med.201: 303–316.
13. Campbell, D. J., C. H. Kim, and E. C. Butcher. 2003. Chemokines in the systemicorganization of immunity. Immunol. Rev. 195: 58–71.
14. Butcher, E. C., and L. J. Picker. 1996. Lymphocyte homing and homeostasis.Science 272: 60–66.
15. Kunkel, E. J., and E. C. Butcher. 2002. Chemokines and the tissue-specific mi-gration of lymphocytes. Immunity 16: 1–4.
16. Kunkel, E. J., J. J. Campbell, G. Haraldsen, J. Pan, J. Boisvert, A. I. Roberts,E. C. Ebert, M. A. Vierra, S. B. Goodman, M. C. Genovese, et al. 2000. Lym-phocyte CC chemokine receptor 9 and epithelial thymus-expressed chemokine(TECK) expression distinguish the small intestinal immune compartment: epi-thelial expression of tissue-specific chemokines as an organizing principle inregional immunity. J. Exp. Med. 192: 761–768.
17. Hudak, S., M. Hagen, Y. Liu, D. Catron, E. Oldham, L. M. McEvoy, andE. P. Bowman. 2002. Immune surveillance and effector functions of CCR10�
skin homing T cells. J. Immunol. 169: 1189–1196.18. Papadakis, K. A., J. Prehn, V. Nelson, L. Cheng, S. W. Binder, P. D. Ponath,
D. P. Andrew, and S. R. Targan. 2000. The role of thymus-expressed chemokineand its receptor CCR9 on lymphocytes in the regional specialization of the mu-cosal immune system. J. Immunol. 165: 5069–5076.
19. Reiss, Y., A. E. Proudfoot, C. A. Power, J. J. Campbell, and E. C. Butcher. 2001.CC chemokine receptor (CCR)4 and the CCR10 ligand cutaneous T cell-attract-ing chemokine (CTACK) in lymphocyte trafficking to inflamed skin. J. Exp. Med.194: 1541–1547.
20. Rott, L. S., J. R. Rose, D. Bass, M. B. Williams, H. B. Greenberg, andE. C. Butcher. 1997. Expression of mucosal homing receptor �4�7 by circulatingCD4� cells with memory for intestinal rotavirus. J. Clin. Invest. 100: 1204–1208.
21. Rose, J. R., M. B. Williams, L. S. Rott, E. C. Butcher, and H. B. Greenberg. 1998.Expression of the mucosal homing receptor �4�7 correlates with the ability ofCD8� memory T cells to clear rotavirus infection. J. Virol. 72: 726–730.
22. Tietz, W., Y. Allemand, E. Borges, D. von Laer, R. Hallmann, D. Vestweber, andA. Hamann. 1998. CD4� T cells migrate into inflamed skin only if they expressligands for E- and P-selectin. J. Immunol. 161: 963–970.
23. Stampfli, M. R., R. E. Wiley, G. S. Neigh, B. U. Gajewska, X. F. Lei,D. P. Snider, Z. Xing, and M. Jordana. 1998. GM-CSF transgene expression inthe airway allows aerosolized ovalbumin to induce allergic sensitization in mice.J. Clin. Invest. 102: 1704–1714.
24. Alvarez, D., G. Harder, R. Fattouh, J. Sun, S. Goncharova, M. R. Stampfli,A. J. Coyle, J. L. Bramson, and M. Jordana. 2005. Cutaneous antigen priming viagene gun leads to skin-selective Th2 immune-inflammatory responses. J. Immu-nol. 174: 1664–1674.
25. Brandt, E. B., R. T. Strait, D. Hershko, Q. Wang, E. E. Muntel, T. A. Scribner,N. Zimmermann, F. D. Finkelman, and M. E. Rothenberg. 2003. Mast cells arerequired for experimental oral allergen-induced diarrhea. J. Clin. Invest. 112:1666–1677.
26. Spergel, J. M., E. Mizoguchi, J. P. Brewer, T. R. Martin, A. K. Bhan, andR. S. Geha. 1998. Epicutaneous sensitization with protein antigen induces local-
ized allergic dermatitis and hyperresponsiveness to methacholine after single ex-posure to aerosolized antigen in mice. J. Clin. Invest. 101: 1614–1622.
27. Saloga, J., H. Renz, G. L. Larsen, and E. W. Gelfand. 1994. Increased airwaysresponsiveness in mice depends on local challenge with antigen. Am. J. Respir.Crit. Care Med. 149: 65–70.
28. Saloga, J., and J. Knop. 2000. Does sensitization through the skin occur? Allergy55: 905–909.
29. Gajewska, B. U., F. K. Swirski, D. Alvarez, S. A. Ritz, S. Goncharova,M. Cundall, D. P. Snider, A. J. Coyle, J. C. Gutierrez-Ramos, M. R. Stampfli, andM. Jordana. 2001. Temporal-spatial analysis of the immune response in a murinemodel of ovalbumin-induced airways inflammation. Am. J. Respir. Cell Mol.Biol. 25: 326–334.
30. Ohkawara, Y., X. F. Lei, M. R. Stampfli, J. S. Marshall, Z. Xing, and M. Jordana.1997. Cytokine and eosinophil responses in the lung, peripheral blood, and bonemarrow compartments in a murine model of allergen-induced airways inflam-mation. Am. J. Respir. Cell Mol. Biol. 16: 510–520.
31. Marco, A. J., M. Domingo, J. Ruberte, A. Carretero, V. Briones, andL. Dominguez. 1992. Lymphatic drainage of Listeria monocytogenes and Indianink inoculated in the peritoneal cavity of the mouse. Lab. Anim. 26: 200–205.
32. Laouini, D., H. Alenius, P. Bryce, H. Oettgen, E. Tsitsikov, and R. S. Geha. 2003.IL-10 is critical for Th2 responses in a murine model of allergic dermatitis.J. Clin. Invest. 112: 1058–1066.
33. Ritz, S. A., M. J. Cundall, B. U. Gajewska, D. Alvarez, J. C. Gutierrez-Ramos,A. J. Coyle, A. N. McKenzie, M. R. Stampfli, and M. Jordana. 2002. Granulocytemacrophage colony-stimulating factor-driven respiratory mucosal sensitizationinduces Th2 differentiation and function independently of interleukin-4.Am. J. Respir. Cell Mol. Biol. 27: 428–435.
34. Herrick, C. A., H. MacLeod, E. Glusac, R. E. Tigelaar, and K. Bottomly. 2000.Th2 responses induced by epicutaneous or inhalational protein exposure are dif-ferentially dependent on IL-4. J. Clin. Invest. 105: 765–775.
35. Kim, S. H., H. N. Jung, K. Y. Lee, J. Kim, J. C. Lee, and Y. S. Jang. 2005.Suppression of Th2-type immune response-mediated allergic diarrhea followingoral administration of traditional Korean medicine: Atractylodes macrocephalaKoidz. Immunopharmacol. Immunotoxicol. 27: 331–343.
36. Kweon, M. N., M. Yamamoto, M. Kajiki, I. Takahashi, and H. Kiyono. 2000.Systemically derived large intestinal CD4� Th2 cells play a central role inSTAT6-mediated allergic diarrhea. J. Clin. Invest. 106: 199–206.
37. McDermott, M. R., D. A. Clark, and J. Bienenstock. 1980. Evidence for a com-mon mucosal immunologic system. II. Influence of the estrous cycle on B im-munoblast migration into genital and intestinal tissues. J. Immunol. 124:2536–2539.
38. McDermott, M. R., and J. Bienenstock. 1979. Evidence for a common mucosalimmunologic system. I. Migration of B immunoblasts into intestinal, respiratory,and genital tissues. J. Immunol. 122: 1892–1898.
39. Macpherson, A. J., and T. Uhr. 2004. Compartmentalization of the mucosal im-mune responses to commensal intestinal bacteria. Ann. NY Acad. Sci. 1029:36–43.
40. Kantele, A., M. Hakkinen, Z. Moldoveanu, A. Lu, E. Savilahti, R. D. Alvarez,S. Michalek, and J. Mestecky. 1998. Differences in immune responses induced byoral and rectal immunizations with Salmonella typhi Ty21a: evidence for com-partmentalization within the common mucosal immune system in humans. Infect.Immun. 66: 5630–5635.
41. Wu, H. Y., and M. W. Russell. 1997. Nasal lymphoid tissue, intranasal immu-nization, and compartmentalization of the common mucosal immune system.Immunol. Res. 16: 187–201.
42. Morales, J., B. Homey, A. P. Vicari, S. Hudak, E. Oldham, J. Hedrick, R. Orozco,N. G. Copeland, N. A. Jenkins, L. M. McEvoy, and A. Zlotnik. 1999. CTACK,a skin-associated chemokine that preferentially attracts skin-homing memory Tcells. Proc. Natl. Acad. Sci. USA 96: 14470–14475.
43. Campbell, J. J., C. E. Brightling, F. A. Symon, S. Qin, K. E. Murphy, M. Hodge,D. P. Andrew, L. Wu, E. C. Butcher, and A. J. Wardlaw. 2001. Expression ofchemokine receptors by lung T cells from normal and asthmatic subjects. J. Im-munol. 166: 2842–2848.
44. Belyakov, I. M., S. A. Hammond, J. D. Ahlers, G. M. Glenn, and J. A. Berzofsky.2004. Transcutaneous immunization induces mucosal CTLs and protective im-munity by migration of primed skin dendritic cells. J. Clin. Invest. 113:998–1007.
45. Strid, J., J. Hourihane, I. Kimber, R. Callard, and S. Strobel. 2005. Epicutaneousexposure to peanut protein prevents oral tolerance and enhances allergic sensi-tization. Clin. Exp. Allergy 35: 757–766.
46. Strid, J., M. Thomson, J. Hourihane, I. Kimber, and S. Strobel. 2004. A novelmodel of sensitization and oral tolerance to peanut protein. Immunology 113:293–303.
47. Strid, J., and S. Strobel. 2005. Skin barrier dysfunction and systemic sensitizationto allergens through the skin. Curr. Drug Targets Inflamm. Allergy 4: 531–541.
48. Mora, J. R., M. Iwata, B. Eksteen, S.-Y. Song, T. Junt, B. Senman, K. L. Otipoby,A. Yokota, H. Takeuchi, P. Ricciardi-Castagnoli, et al. 2006. Generation of gut-homing IgA-secreting B cells by intestinal dendritic cells. Science 314:1157–1160.
49. Davenport, M. P., M. C. Grimm, and A. R. Lloyd. 2000. A homing selectionhypothesis for T-cell trafficking. Immunol. Today 21: 315–317.
3296 SITE OF Th2 DIFFERENTIATION EVOKES MUCOSA-SPECIFIC RESPONSES
by guest on January 30, 2018http://w
ww
.jimm
unol.org/D
ownloaded from