ternary coordination complex between human serum albumin, copper
TRANSCRIPT

TIIE Jounx~. OP BIOLOGICAL CHEIIISTRP Vol. 246, No. 19, Issue of October 10, pp. 5938-5943, 1971
Printed in U.S.A.
Ternary Coordination Complex between Human Serum
Albumin, Copper (II), and L-Histidine*
(Received for publication, March 22, 1971)
SHOW-JY LAU AND BIBUDHENDRA SARKAR
From The Division of Biochemistry, The Research Institute of the Hospital for Sick Chilclren, Toronto, and The Depadnent of Biochemistry, University of Toronto, Toronto, Ontario, Canacla
SUMMARY
Physicochemical studies of the human serum albumin- Cu(II)-L-histidine (HSA-Cu(II)-L-His) ternary complex at pH 7.5 indicate several interesting features.
1. The dissociation constants of the HSA-Cu(II)-L-His ternary complex and the HSA-Cu(I1) binary complex are 1.38 X 1O-22 and 6.61 x lOen, respectively.
2. The absorption spectrum of the ternary complex system has characteristics similar to those of the NHz-terminal pep- tide (1-24) of bovine serum albumin in the presence of Cu(I1) and L-His. The computed spectrum of HSA-Cu(II)-L-His has a X,,, at 540 nm, a shift of 15 nm toward red from that of HSA-Cu(I1) (X,,,, = 525 nm). 3. The calculated moles of protons liberated in the forma-
tion of HSA-Cu(II)-L-His from HSA and Cu(II)-L-His2 are 0.28. While this may originate from an a-amino group, the number is much less than what would be expected if a peptide nitrogen of HSA were involved in the binding.
4. Cu(I1) in the ternary complex is most likely bound to both HSA and L-His. The possible binding mode changes from all nitrogen, as shown in HSA-Cu(II), to either a mixture of nitrogen and oxygen or an additional involvement of imidazole nitrogen.
5. Equilibria, existing in the ternary complex system, sug- gest that the ternary complex may play an important role in the biological transport of Cu(I1).
It is known that a fraction of Cu(I1) in human blood plasma is bound to human serum albumin (2, 3). This fraction is in rapid equilibrium with Cu(I1) in tissues (2) and is considered to be the transport form of Cu(I1) in blood. There is also an amino acid bound fraction of Cu(II) in serum consisting mostly of Cu(II)-L- His& complex which is in equilibrium with the fraction bound to
* Part of the data included in this paper have been presented before the XIIth International Conference on Coordination Chemistry, Sydney, 1969 (1). This work was supported by grants from the John A. Hartford Foundation of New York and The Medical Research Council of Canada.
‘The abbreviations used are: L-His, L-histidine; HSA, human serum albumin; BSA, bovine serum albumin; Asp fragment, NHz.- terminal peptide (l-24) of BSA; n-Thr, n-threonine. See also “List of Symbols” in Reference 17.
HSA (3, 4). The evidence has recently been presented for the existence of a t.ernary coordination complex between HSA, Cu(I1) and L-His (5). The importance of ternary coordination com- plexes has long been recognized in biological chemistry and many investigators have contributed significantly in this area. Klotz and Ming (6) and Gurd (7) have demonstrated that metal ions may mediate the binding of small molecules to proteins. Malm- Strom (8) later studied the ternary complex formation between enolase, metal ion, and nn-2-phosphoglycerate. Since the origi- nal proposal of a bridge structure between enzyme, metal, and substrate by Hellerman and Stock (9), this hypothesis has been put forward to explain the mechanism of action of many enzymes (10-12). Recently, important contributions have been made by Xldvan and Cohn (13) to our understanding of the formation of the enzyme-metal-substrate bridge structures. In order to understand fully the ternary complexes involving enzymes and proteins, several laboratories have begun to study the low molecu- lar weight ternary complexes of amino acids and peptides (14-23).
This communication deals with the physicochemical studies of HSA-Cu(II)-L-His by equilibrium dialysis, spectrophotometry, and titration. The formation of the ternary complex is also shown between the 24-residue NH,-terminal peptide of BSA (24), Cu(II), and L-His.
EXPERIMENTAL PROCEDURE
Materials-Crystalline HSA was obtained from Hoechst Phar- maceutical Company and used without further purification. The assumed molecular weight was 69,000. The Asp fragment of BSA was a generous gift from Dr. Theodore Peters, Jr.2 The radioisotope G7Cu(II), specific activity at least 0.5 mCi of G7Cu(II) per pg of Cu(II), was prepared (25) by irradiation of natural zinc with Bremsstrahlung from the linear accelerator at the University of Toronto. L-[14C]His was obtained from Schwarz BioResearch. L-His free base ([ar];’ + 39.9” f 0.5”) was obtained from Nutritional Biochemicals. Dialysis mem- brane was obtained from Visking Company. All other reagents were of analytical grade and the water was deionized.
Equilibrium Dialysis-The extent of binding was measured by the equilibrium dialysis technique. The dialysis cells were made of rigid acrylic plastic. Six separate chambers provided for si-
2 Preparation P2 was prepared by Dr. Theodore Peters, Jr., as in Reference 24. Trichloroacetic acid was removed by Sephadex G-10 rather than ether at the final step. The calculated molecu- lar weight was 2802 on the basis of the amino acid composition determined by two analyzer runs with 24-hour hydrolysis.
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of October 10, 1971 S.-J. Lau and B. #arka? 5939
multaneous dialysis experiments within one unit. Each half- cell had a capacity of 5 ml, and was provided with an opening for introducing or withdrawing of solutions. The membrane was placed in between two cells with two pieces of perforative Parafilm as lining to seal the compartments. The equilibrium dialysis was carried out in 0.1 M N-ethylmorpholine-HCl buffer at pH 7.53 and ionic strength 0.16. Three different stock solu- tions were prepared: (a) 1% HSA in buffer, (b) CuClz in lVa M
HCl, a,nd (c) L-His in buffer. The dialysis cells were assembled with 4 ml of solution on both sides. One half-cell contained So- lution b mixed with WuC12 and Solution a. The molar ratio of Cu(I1) to HSA was kept approximately 1. The other half- cell contained various known amounts of Solution c and L-[14C]His. The total molar ratio of L-His to HSA ranged from about 0.3 to 10.0. Similar experiments were carried out involv- ing HSA and L-His alone with a molar ratio of L-His to HSA ranging from 0.01 to 4.0.
Equilibrium was obtained by using an automatic shaker for 4 days at 6”. Radioactivity of YJu(II) and 14C was measured both before and after dialysis. A well type scintillation counter equipped with the pulse height analyzer was used for the assay of 67Cu(II) activity. Thecounts were corrected for decay on the basis of the 60-hour half-life of 67Cu(II). After allowing suffi- cient time for Wu(I1) to decay, the radioactivity of r*C was de- termined in a liquid scintillation counter.
Absorption Spectra-The visible absorption spectra of the Cu- (II) complexes were obtained with a Cary model 15 recording spectrophotometer thermostated at 25’. The cell path was 1 cm. Spectra were corrected for the slight visible absorption of HSA in the absence of Cu(I1). All solutions for the spectral measurements were prepared in 0.16 M KCl.
Proton Displacement-Titrations were performed at 25” and pH 7.50 on a Corning model 12 research pH meter. To 25 ml of 1 y. HSA solution, prepared in 0.16 M KC1 and adjusted to the required pH value, was added either CuC12 in lOma M HCl or Cu(II)-L-His2 solution at pH 7.50. The protons released were then titrated with 0.01 M NaOH in a sealed system. The molar ratio of CuCh or Cu(II)-L-Hiss to HSA remained approximately 1.
RESULTS AND DISCUSSION
Dissociation of HSA-Cu(II)+His and HSA-Cu(II)-The dissociation of the ternary and the binary complexes can be for- mulated asa:
8 The equilibria of Cu (II) (M) with L-His (HaA) in various molar ratios have been investigated by potentiometry in aqueous solu- tion (0.15 M NaCl, 25”). The species detected in the system were MHA, MA, MH&, MHA,, MA2, MH-IAN, MH-IA, and MzH-ZAZ, and their distribution was obtained over the pH range 1 to 11. The major species in the system at pH 7.5 was shown to be MA2. Minute amounts (<O.l’%) of two hydroxylated species (MH-IA and MzH-& are present at this pH, but their presence in the ternary system is considered negligible. Thus, the hydroxylated species are not included in the equilibrium equation. The presence of multiple species existing simultaneously was taken into account to interpret the structures of Cu(II)-~-His species by physical and chemical means. The mode of metal coordination in MA, was shown to be 0 (carboxyl), N (imidazole), and N (amino) involve- ment in both L-histidines. The species MA2 (log pl02 = 18.46) has a X,, = 640nm and an hmaX = 86 (T. P. A. Kruck and B. Sar- kar, manuscript in preparation).
K DHSA-C,,(U)-L-His 3 HSA-Cu(II)-L-His ~ 0)
HSA-Cu(I1) f Cu(II)-n-His2 + L-His f 2 HSA + Cu(I1)
and
K=‘HsA-cn(I,)
HSA-Cu(I1) \ ’ HSA + Cu(I1) (2)
Therefore,
K DHSA-Cu(II)-L-Hi.
[HSA-Cu(II)]]Cu(II)-L-Hisz][L-His][HSA]*[Cu(II)] (3) = IHSA-Cu(II)-n-His18
and
lHSAllCu(II)l KDHSA-C~(II) = ~HSA-C~(II)I (4)
The KD values were determined from a set of equations with the derived parameters from the equilibrium dialysis data utiliz- ing 67Cu(II) and L-[14C]His double labeling. Let
Total Cu (II) concentration = [Cu (II)] L Total HSA concentration = [HSAlt Total L-His concentration = [n-Hislt
Counts of 67Cu(II) before dialysis = a Counts of 6%~ (II) after dialysis (aqueous portion) = b Counts of 67Cu(II) after dialysis (protein portion) = c Counts of L-[i4C]His before dialysis = d Counts of L-[W]His after dialysis (aqueous portion) = e Counts of L-[W]His after dialysis (protein portion) = f
Therefore, the following equations can be set up:
[Cu(II)] + [Cu(II)-L-His21 N- [Cu(II)-L-His21
= % [Cu(II)I, (5)
[HSA-Cu(II)-L-His] + [HSA-Cu(II)]
= c+ [Cu(II)It (6)
[HSA-Cu(II)-L-His] = ‘y [L-Hish
Then,
[HSA-Cu(II)] = e [Cu(II)lt - ‘5 [n-HisIt
[HSA] = lHS-41~ - ‘4 lCu(II)l,
(7)
(8)
(9)
[n-His] = [n-His]( [l - 91 - 2 % lCu(II)], (10)
and [Cu(II)] can be calculated according to the known stability constant of Cu(II)-L-His2.a
The equilibrium dialysis data are presented in Table I. The average value of the dissociation constant for HSA-Cu(II)-n-His is 1.38 x 10-n and that for HSA-Cu(I1) is 6.61 x lO-‘r.
E$ect of ~-His on Spectra of HSA-Cu(II) and, Asp Fragmmt- Cu(II)-The spectral characteristics of HSA-Cu(I1) at pH values 5.50, 6.50, 8.00, and 10.03 are shown in Fig. 1A. The spectra of the Cu(II)+Hisz system taken at about the same
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from
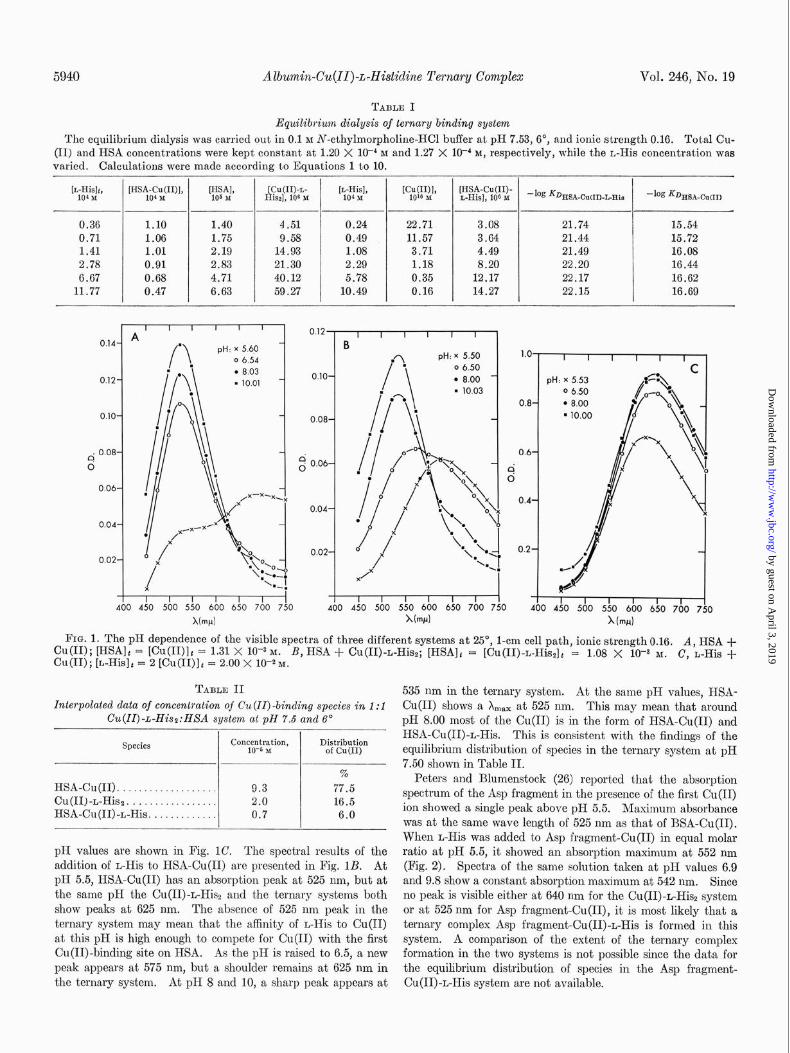
5940 Albumin-Cu(II)-L-Hi&dine Ternary Complex Vol. 246, No. 19
TABLE I
Equilibriwn dialysis of ternary binding system
The equilibrium dialysis was carried out in 0.1 M N-ethylmorpholine-KC1 buffer at pH 7.53, 6”, and ionic strength 0.16. Total Cu- (II) and HSA concentrations were kept constant at 1.20 X 1OV M and 1.27 X lo+ M, respectively, while the L-His concentration was varied. Calculations were made according to Equations 1 to 10.
[L-Hi& 104 2.5
W;-&UIN, lHS%, [Cu(II)-L- [$~~I> Will> [HSA-Cu(II)- His& 106 I L-His], 106 x -log =DHSA-CuCI)-L-His -log KDHSA-CuW
0.36 1.10 1.40 4.51 0.24 22.71 3.08 21.74 15.54
0.71 1.06 1.75 9.58 0.49 11.57 3.64 21.44 15.72
1.41 1.01 2.19 14.93 1.08 3.71 4.49 21.49 16.08
2.78 0.91 2.83 21.30 2.29 1.18 8.20 22.20 16.44
6.67 0.68 4.71 40.12 5.78 0.35 12.17 22.17 16.62
11.77 0.47 6.63 59.27 10.49 0.16 14.27 22.15 16.69
0.06-
0.04-
0.02-
I I I I I 1 I I 1
400 450 500 550 600 650 700 750
Nw) 460 4iO 560 5iO 600 650
I
FIN. 1. The pH dependence of the visible spectra of three different systems at 25“, l-cm cell path, ionic strength 0.16. cu(II); [HSA]t = [Cu(II)]r = 1.31 X lo-+ M.
A, HSA +
Cu(I1); @-HisIt = 2 [Cu(II)]t = 2.00 X 10-S M. B, HSA + Cu(II)-L-Hisz; [HSA]t = [Cu(II)-L-HissIt = 1.08 X 10Fa M. C, L-His +
TABLE II Interpolated data of concentration of Cu(IZ)-binding species in 1:l
Cu(II)+His2:HXA system at pH 7.6 and 6’ I
Species Concentration, Distribution 10-s M of Cu(I1)
% HSA-Cu(I1). . . . . . . . . 9.3 77.5 Cu(II)-L-Hisz. . . . 2.0 16.5 HSA-Cu(II)-L-His. . . . . . 0.7 6.0
pH values are shown in Fig. 1C. The spectral results of the addition of L-His to HSA-Cu(I1) are presented in Fig. 1B. At pH 5.5, HSA-Cu(I1) has an absorption peak at 525 nm, but at the same pH the Cu(II)-L-His2 and the ternary systems both show peaks at 625 nm. The absence of 525 nm peak in the ternary system may mean that the affinity of L-His to Cu(I1) at this pH is high enough to compete for Cu(I1) with the first Cu(II)-binding site on HSA. As the pH is raised to 6.5, a new peak appears at 575 nm, but a shoulder remains at 625 nm in the ternary system. At pH 8 and 10, a sharp peak appears at
535 nm in the ternary system. At the same pH values, HSA- Cu(I1) shows a X max at 525 nm. This may mean that around pH 8.00 most of the Cu(I1) is in the form of HSA-Cu(I1) and HSA-Cu(II)-L-His. This is consistent with the findings of the equilibrium distribution of species in the ternary system at pH 7.50 shown in Table II.
Peters and Blumenstock (26) reported that the absorption spectrum of the Asp fragment in the presence of the first Cu(I1) ion showed a single peak above pH 5.5. Maximum absorbance was at the same wave length of 525 nm as that of BSA-Cu(I1). When L-His was added to Asp fragment-Cu(I1) in equal molar ratio at pH 5.5, it showed an absorption maximum at 552 nm (Fig. 2). Spectra of the same solution taken at pH values 6.9 and 9.8 show a constant absorption maximum at 542 nm. Since no peak is visible either at 640 nm for the Cu(II)-n-His2 system or at 525 nm for Asp fragment-Cu(II), it is most likely that a ternary complex Asp fragment-Cu(II)-L-His is formed in this system. A comparison of the extent of the ternary complex formation in the two systems is not possible since the data for the equilibrium distribution of species in the Asp fragment- Cu(II)-L-His system are not available.
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of October 10, 1971 S.-J. Lau and B. Sarkar 5941
I I I I I I I I I 400 450 500 550 600 650 700 750 800
Xlw)
FIG. 2. The visible spectra of Asp fragment-Cu(II)-L-His as a function of pH. Conditions: 25”, l-cm cell path, ionic strength 0.16, [Asp fragmentI = [Cu(II)]t = [L-His]( = 2.05 X 10-4~.
Computed Spectrum of Ternary Complex-The computed spectrum of the ternary complex was obtained by subtracting the optical density due to all other visible light-absorbing species from that of the whole system as illustrated in the follow- ing equations:
= fiHsA-Cu(II)-L-His X [HSA-Cu(II)-L-His1
+ '%HSA-Cu(I1) X [HSA-Cu(II)I + e&(II)-L-Hia, (10)
X [Cu(II)-L-Hiss1 + ehHsA X [HSAI
The extinction coefficient of the ternary complex was deter- mined from the known values of the extinction coefficient of the individual species3 (27) and the concentrations obtained from the equilibrium dialysis method. It was calculated at 5 or 10 nm wave length intervals from 700 to 460 nm to constitute the spectrum. The computed spectrum of HSA-Cu(II)+His is presented in Fig. 3, along with the spectra of HSA-Cu(I1) and Cu(II)-L-Hisz, the predominant species in the Cu(II)-L-His2 system at pH 7.5. The ternary complex shows a X,, at 540 nm, a shift of 15 nm toward red from that of HSA-Cu(I1) blk3, = 525 nm). The Cu(II)-L-His2 has a X,,, at 640 nm.
Proton Displacement Studies-The number of titrable protons observed by adding Cu(I1) and Cu(II)-L-His2 to HSA at pH 7.50 is shown in Table III. When Cu(II)-L-His2 is added to HSA, there are more species formed than HSA-Cu(II)-L-His. Consequently, the proton displacement data of the ternary system shown in Table III do not reflect what has originated from HSA-Cu(II)-L-His. In order to solve the number of pro- tons displaced due only to the formation of HSA-Cu(II)+His, it is necessary to know the distribution of all of the species in the system. This was obtained by using the concentration data from equilibrium dialysis experiment (Table I) by means of interpolations to the conditions used in titration.4 The result is shown in Table II. This enabled us to calculate the contribu- tion of the individual Cu(II)-binding species to the observed proton displacement data.
4 This calculation did not take into account the effect of tem- perature on equilibrium.
&---o-- o/O x----v 4Lo
I I I I 500 540 580 620 660 700
X lwd
FIG. 3. A comparison of the absolute visible spectra of three different species at pH 7.50,25”, and ionic strength 0.16. l - 0, computed spectrum of HSA-Cu(II)-L-His; X-X, HSA-Cu(I1); O-0, Cu(II)-L-H&.
TABLE III
Proton displacement data by equimolar additions of Cu(ZZ) and Cu(ZZ)-L-H& to HSA at pH 7.60
Addition to HSA Protons displaced
Cu(I1). ....................................... 2.58 Cu(II)-L-His2 ................................. 0.67
For the formation of each mole of HSA-Cu(II)-L-His from a mixture of Cu(II)-L-Hi% and HSA, 1 mole of free L-His is liber- ated. Similarly, 2 moles of free L-His are liberated in the forma- tion of each mole of HSA-Cu(I1). In Cu(II)-L-Hisa the amino nitrogen is bonded to Cu(I1) ; hence, at pH 7.50 the L-His liber- ated from Cu(II)-L-Hiss will take up protons to protonate the newly released amino group. Meanwhile, some protons are liberated as a result of the formation of HSA-Cu(II). The actual number of protons displaced at pH 7.50 in the formation of HSA-Cu(II)-L-His can then be calculated as illustrated below:
Moles of protons taken up by L-His due to the formation of HSA-Cu(I1) = 2 X 77.501, = 1.55
Moles of protons taken up by L-His due to the formation of HSA-Cu(II)-L-His = 1 X 6% = 0.06
Then the total moles,of protons taken up = 1.55 + 0.06 = 1.61 But, the moles of protons released in the formation of HSA-
Cu(I1) = 2.58 X 77.5% = 2.00 Therefore, the net moles of protons displaced in the formation
of HSA-Cu(II)-L-His = 0.67 - (2.00 - 1.61) = 0.28
Possible Identity of Cu(II)-binding Site in HSA-Cu(II)-L-His- HSA apparently consists of a single polypeptide chain made up of approximately 565 amino acids and stabilized by about 17 disulfide bonds (28). It has been shown that the first Cu(II)- binding site in HSA involves the NHz-terminal sequence: Asp- Ala-His (29). The proposed binding site includes the a-amino nitrogen of aspartyl residue, two intervening peptide nitrogens, and the imidazole nitrogen of the histidine in position 3 (26, 3032). In the absence of Cu(II), free L-His does not bind to HSA at neutral pH (5). Others have also shown that amino acids such as glycine, leucine, and phenylalanine do not bind to albumin (33). It is unlikely that the attachment of Cu(I1) to the first binding site changes the structure of HSA in such a way
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Albumin-Cu(II) ,L-Hi&line Ternary Complex Vol. 246, No. 19
as to open up an entirely new site for a free amino acid to bind to form the ternary complex. This view is consistent with the finding that the ternary complex is also formed with t,he 24..resi- due NHz-terminal peptide of BSX. In view of this, it is most likely that Cu(I1) in the ternary complex is bound to both H&4 and L-His. Both spectral and proton displacement data are in favor of this contention.
Recent studies suggest that both L-His molecules in Cu(II)-L- His2 at p1-l 7.5 are tridentate. It is also interesting to note that ternary complex L-His-Cu(II)-L-Thr, which was isolated and crystallized in our laboratory (4) from the normal human serum and later subjected to x-ray analysis (18), was shown to have L-His in a tridentate form. The tridentatc form of L-His in HSA-Cu(II)-L-His is a possibility, alt,hough none of the results presented here entirely rule out ot’her modes of binding. The computed spectrum of the ternary complex shows a red shift from 525 nm (HSA-Cu(I1)) to 540 nm (HSA-Cu(II)-L-His). This is consistent wit,h the change of binding mode, either from all nitrogen to a mixture of nitrogen and oxygen (34) or to an additional involvement of imidazolc nitrogen3 The introduc- tion of oxygen or nitrogen (or both) in the ligand field could be from t,he carboxyl or imidazole groups (or both) of L-His.
The binding of the protein part to Cu(I1) in the ternary com- plex presents some interesting possibilities. It seems unlikely that the protein part of the ternary comples would bind Cu(I1) in the same manner as it does in HSX-Cu(I1) (26, 30-32), since it has also to bind L-His simult,aneously. Although no informa- tion is available concerning the intrinsic pK, of the a-amino group of HSA, it may fall around the neutral pH range (35). The protons which are displaced at pH 7.50 (Table III) come most likely from the a-amino nitrogen of HSA in the ternary complex. The displaced protons cannot come from the imida- zole group of a histidyl residue since the imidazole protons are already titrated at pH 7.50. While the participation of the imidazole nitrogen is a strong possibility, no conclusion can be derived from the available data. The quantity of protons which was displaced at pH 7.50 is much less than what one would expect if a peptide nitrogen were involved. However, these interpretations must be followed with caution, since the intrinsic pK, values of the donor groups are not known.
Studies with model ternary complexes involving glycine pep- tides, Cu(II), and glycine suggest that, in both glycylglycine- Cu(II)-glycine and diglycylglycinc-Cu(II)-glycine, the a-amino nitrogen and a peptide nitrogen are involved in Cu(I1) binding, while the glycine part of the ternary complex is shown to bind Cu(I1) with the carboxyl and amino group (17). What influence a side chain residue, such as an imidazole group, might have in the formation of the ternary complex is not known. Thus, a suitable model is lacking from which analogy can be drawn to gain any further knowledge about the ident,ity of the Cu(II)- binding site in HSA-Cu(II)-L-His.
Possible Physiological Xigni$cance of Ternary Comples-Hu- man serum contains approximately I pg of Cu(I1) per ml. Most of this is bound to ceruloplasmin, and a small fraction is bound to both albumin and amino acids (3, 4). The Cu(I1) in cernlo- plasmin is not exchangeable in viva (36). It is the albumin- and the amino acid-bound fraction of serum Cu(I1) which is ex- changeable and is considered to be the physiologically important transport form of Cu(I1) (2-4). In view of the findings pre- sented here, it seems reasonable to postulate that the following equilibria are maintained in t,he physiological st,ate:
Copper (II) + amino acid ti Cu(II)-amino acid 11 albumin
albumin-cu(IIj-amino acid 11 "
albumin-Cu(I1) + amino acid
SCHEL~E 1
Although L-His is the major Cu(II)-binding amino acid in serum (3, 4), other amino acids could also play a similar role in forming an intermediary ternary complex. The above mecha- nism5 could play an important role in the exchange of Cu(I1) between a macromolecule and a low molecular weight substance which in turn can readily be transported across the biological membrane.
REFERENCES
1. SARKAR, B., WIGFIELD, Y., LEE, M. L., KRUCK, T. P. A., AND LAU. S., in H. C. Freeman (Editor). Proceed&as of the XIIth Inteknakonal Conference on Coordination Chekisiry. Science
2.
3.
4.
Press, Merricville, Sydney, Australia, 1969, p. 126.’ BEARN. A. G.. AND KUNICEL. H. G.. Proc. Sot. EXD. Biol. Med..
88, 44 (1954).
5.
6.
7.
8. 9.
10. 11.
12.
13.
14.
15.
16.
17.
18.
NEUMANN, P. Z., AND SASS-K• RTSAK, A., 3. Clin. Invest., 46, 646 (1967).
SARICAR, B., AND KRUCK, T. P. A., in J. PEISACH, P. AISEN, AND W. E. BLUIXRERG (Editors)., Biochemistry of copper, Academic Press, New York, 1966, p. 183.
SARKAR, B., AND WIGFIELD, Y., Can. J. Biochem., 46, 601 (1968).
KLOTZ, I. M., AND MING, W.-C. I,., J. Amer. Chem. Sot., 76, 805 (1954).
GURD, F. R. N., in H. T. CLARIC (Editor), Ion transport across membranes, Academic Press, New York, 1954, p. 246.
MALMSTR~M, B. G., Arch. Biochem. Biophys., 68, 398 (1955). HELLERMAN, L., AND STOCK, C. C., J. Biol. Chem., 126, 771
(1938). SMITH, E. L., Advan. Enzymol., 12, 191 (1951). MALMSTR~M, B. G., AND ROSENBERG, A., Advan. Enzymol., 21,
131 (1959). VALLEE, B. L., AND COLEX~N, J. E., Compr. Biochem., 12, 165
(1964). MILDVAN, A. S., AND COMN, M., Advan. Enzymol., 33, 1
(1970). SARICAR, B., AND KRUCI~, T. 1’. A., Can. J. Biochem., 46, 2046
(1967). SARKAR, B., BERSOHN, M., WIGFIELD, Y., AND CHIANG, T.-C.,
Can. J. Biochem., 46, 595 (1968). SARKAR, B., LEE, M. L., AND MARTIN, R.-P., Proc. Can. Fed.
Biol. SOL, 12, 14 (1969). MARTIN, R.-P., MOSONI, L., AND SARICAR, B., J. Biol. Chem.,
246, 5944 (1971). FREEMAN, H. C., Guss, J. M. HEALY, M. J., MARTIN, R.-P.,
NOCHHOLDS, C. E., AND SARICAR, B., Chem. Comm., 225 (1969).
19. 20.
MARTIN, R.-P., AND PARIS, K. A., Bull. Sot. Chim., 570 (1963). MARTIN, R.-P., AND PARIS, R. A., Bull. SOL. Chim., 46, 80
(1964). 21. 22.
MARTIN, R.-P., Bull. Sot. Chim., 2217 (1967). FREEMAN, H. C., AND MARTIN, R.-P., J. Biol. Chem., 244,4823
(1969). 23. MARTIN, R.-P., AND MOSONI, L., Bull. Sot. Chim., 2917 (1970).
6 The reaction kinetics in the formation of HSA-Cu(II)-L-His at pH 7.50 and 25” was attempted by using the stopped flow tech- nique. However, no observable rate could be obtained. This indicates that the formation of both IISA-Cu(II)-n-His and HSA- Cu(I1) is very rapid and beyond the detectable time range of stopped flow or that the change in optical density is not apprecia- ble, or both.
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of October 10, 1971 S.-J. Lau ancl B. Sadar 5943
24. PETERS, T., JR., AND HAWN, C., J. Biol. Chem., 242, 1566 (1967).
25. MARCEAU, N., KRUCK, T. P. A., MCCONNEL, D. B., AND ASPIN, N., Znnt.. J. Appl. R&at. Isotopes, 21, 667 (1976).
26. PETERS. T.. JR.. AND BLUAUZNSTOCK. F. A.. J. Biol. Chem.. 242. 1574 (196+). ’
I
27. APPLETON, D. W., AND SARKAR, B., J. Biol. Chem., 246, 5040 (1971).
28. HUNTER. M. J., AND MCDUFFIE, F. C., J. Amer. Chem. Sot., 81, 1400 (i949).
29. BRADSHA~. R. A.. AND PETERS. T.. JR.. J. Biol. Chem.. 244. 5582 (196b). ’
, , I
30. BRESLOW, E., J. Biol. Chem., 239, 3252 (1964).
31. SHEARER, W. T., BRADSHA~, R. A., GURD, F. R. N. AND PETERS, T., JR., J. Biol. Chem., 242, 5451 (1967).
32. BRADSHAW, R. A., SHEARER, W. T., AND GURD, F. R. N. J. Biol. Chem., 243, 3817 (1968).
33. HUNTER, M. J.. AND COMMERFORD, S. L., J. Amer. Chem. Sot., 7’7, 4856 (1955).
34. BRILL, A. S., MARTIN, R. B., AND WILLIAMS, R. J. P., in B. PULLMAN (Editor), Electronic aspects of biochemistru. Aca- demio Press, New kork, 1964, p.‘519. -
-,
35. TANFORD, C., SWANSON, S. A., AND SHORE, W. S., J. Amer. Chem. Sot., 77, 6414 (1955).
36. STERNLIEB, I.,MORELL, A. G., TUCKER, W. D., GREENE, M. W., AND SCHEINBERG, I. H., J. Clin. Invest., 40,1834 (1961).
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Show-Jy Lau and Bibudhendra Sarkarl-Histidine
Ternary Coordination Complex between Human Serum Albumin, Copper (II), and
1971, 246:5938-5943.J. Biol. Chem.
http://www.jbc.org/content/246/19/5938Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/246/19/5938.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 3, 2019
http://ww
w.jbc.org/
Dow
nloaded from