tectal pathways of regenerating goldfish optic axons after ... · the pathways take in eacnh case...
TRANSCRIPT

Development 102, 479-488 (1988)Printed in Great Britain © The Company of Biologists Limited 1988
479
Tectal pathways of regenerating goldfish optic axons after nasal or
temporal half retinal removal
MARTIN F. HUMPHREY1* and CLAUDIA A. O. STUERMER2
1 Max-Planck-lnstitut fur Hirnforschung, Frankfurt/M, FRG2Friedrich-Miescher-Laboratorium der Max-Planck-Gesellschaft, Tubingen, FRG
* Present address for correspondence: Miami Project to Cure Paralysis, 1600 N.W. 10th Avenue, R-48, Miami, Florida 33136, USA
Summary
The tectal pathways of regenerating goldfish opticaxons are abnormal but not random. The relativeproportion of temporal axons is highest in rostraltectum (65 %) drops in midtectum (31 %) and is verylow in caudal tectum (4 %). By contrast, nasal axonsproceed into caudal tectum and are therefore rela-tively evenly distributed throughout the tectum. Inthis study, we have tested whether temporal axons areconfined to rostral tectum by the presence of nasalaxons in caudal tectum or whether they have apreference for rostral tectum regardless of otheraxons. We similarly tested whether nasal axons wouldgrow preferentially into caudal tectum in the absenceof temporal axons.
At the time of optic nerve section either the nasal ortemporal half retina was removed. Either 35 or 70days after nerve section, the regenerating optic axonswere labelled with HRP and both their pathways anddistribution determined in DAB-reacted tectal whole-mounts.
In the absence of nasal axons, the relative density oftemporal axons in rostral, mid and caudal tectum was70 %, 28 % and 2 %, respectively. The correspondingvalues for nasal axons, in the absence of temporalaxons, were 30 %, 40 % and 30 %, respectively. Thus,the overall distribution of nasal and temporal axons inthe half retinal regenerates was similar to that ofwhole retinal regenerates, demonstrating that theretinotopic preferences of the axons were not depen-dent upon interaxonal interactions.
Thus, nasal and temporal axons obviously discrimi-nate between rostral and caudal tectum despite path-way disorganization and the absence of axons from theopposite hemiretina. This is consistent with axonalgrowth being under the influence of positionalmarkers in tectum.
Key words: retinotectal, topographic projections,positional information, axonal regeneration.
Introduction
The pathways laid down by axons in locating theirtarget region provide clues about the growth strategyused for locating that region (Fujisawa et al. 1982).Recent anatomical studies in goldfish, in which thepathways of retinal axons to their target sites in theoptic tectum have been investigated, have providednew information concerning the pathway organiz-ation of both normal axons (Cook & Rankin, 1984;Easter & Stuermer, 1984; Stuermer & Easter, 1984a)and axons regenerating after optic nerve section(Rankin & Cook, 1986; Stuermer, 1986; Stuermer &Easter, 19846). In light of these findings, the view
conveyed by Attardi & Sperry (1963) that regenerat-ing axons follow normal routes to their targets hashad to be modified.
It has been shown that regenerating axons do notrecapitulate the spatiotemporal order of their normalpredecessors but course through abnormal routes andinappropriate tectal territories (Stuermer, 1986,1988a,fr). It may therefore be possible that regenerat-ing axons locate their appropriate termination regionlargely by a trial and error mechanism rather than byfollowing positional information on the tectum.
Recent quantitative evaluations of the tectal distri-bution of axons from known retinal regions have

480 M. F. Humphrey and C. A. O. Stuermer
revealed, however, that the vast majority of temporalaxons course through the appropriate rostral tectalhalf (Stuermer, 1987a,b). Thus, their relative densitydeclined from 65 % in rostral tectum to 31 % inmidtectal regions and 4 % in caudal tectum. Nasalaxons, on the other hand, were distributed at asimilar density in rostral and midtectal regions andonly declined to 15 % caudal to their appropriatetermination region. Furthermore, axons misroutedinto inappropriate tectal territories were observed toundergo course corrections directed towards theirtarget sites (Stuermer, 1986). Thus, regeneratingaxons exhibited a clear preference for their correcttectal territories, despite their abnormal pathways.
The degree to which the above pattern of ingrowthis a consequence of interactions between the ingrow-ing axons themselves, as opposed to interactionsbetween the axons and positional cues in the tectum,is unknown. During the early period of axonalingrowth, the axons are closely associated with oneanother and so there is plenty of opportunity for themto interact (Murray, 1976; Murray & Edwards, 1982;Stuermer & Easter, 19846). It has also been shownthat axons can exclude or displace one another fromregions within the target (Meyer, 1979; Sharma &Tung, 1979). Thus it is possible that axons appropri-ate to a given tectal region prevent excessive invasionby inappropriate axons. The presence of axons appro-priate to a region may also be a stimulus for with-drawal of inappropriate branches (Fujisawa, 1981;Meyer, 1980; Schmidt & Edwards, 1983; Stuermer,
Attardi & Sperry (1963), using silver staining,provided anatomical evidence that both nasal andtemporal axons grew selectively to their appropriatetectal halves even in the absence of input from theopposite half of the retina. However, more recentHRP studies (Rankin & Cook, 1986; Stuermer, 1986;Stuermer & Easter, 1984) have shown that theirdescription of the axonal pathways was inaccurateand understaining may have been a problem. Moremodern anatomical techniques have produced a simi-lar overall picture but none were suitable for detailedaxonal tracing (Meyer, 1980; Stuermer, 1981; Meyeretal. 1985). Recent electrophysiological studies evensuggest that nasal axonal arbours are present over theentire tectum as early as 46-60 days after nerve crushfollowing temporal half retinal removal (Meyer,1987).
Therefore, we have reinvestigated the distribution,pathways and branching patterns of regeneratingnasal and temporal axons following optic nerve sec-tion and removal of their respective opposite retinalhalf. By using anterograde transport of HRP to labelthe entire nerve and also discrete bundles of axons,
we have been able to trace the pathways with greateraccuracy than previous studies.
Our results have shown that temporal axons prefer-entially course over rostral tectum, even in theabsence of nasal axons, and nasal axons proceed intocaudal tectum even when temporal axons are absent.The pathways taken in each case closely resemblethose of whole retinal regenerates (Stuermer,1987a,b). Additionally, axonal ramifications in theinappropriate tectal half did not increase in theabsence of the axons appropriate to that half.
Preliminary accounts of this study have been pub-lished in abstract form (Humphrey & Stuermer,1987).
Materials and methods
Adult goldfish, Carassius auratus, 4-6cm in length weremaintained at low density in aerated aquaria at roomtemperature (17-20°C). For all surgical procedures, thefish were anaesthetized by immersion in a 01 % aqueoussolution of tricaine methane sulphonate (MS222). A total of42 fish was operated bilaterally for this study. Thoseexamined had to meet strict criteria of complete half retinalremoval, an undamaged remaining retinal half (assessed bywhole mounting) and strong axonal labelling with lowbackground. The number examined is indicated in eachrelevant section.
Optic nerves were severed within the orbit taking carenot to cut the blood vessels supplying the eye. An incisionwas then made in the cornea, the lens was reflected fromthe eye and the retina cut with a razor blade along a verticalline adjacent to the optic nerve head. The half retina thatwas isolated from the optic nerve head was removed bygentle suction. The lens and corneal flap were then re-placed. Both temporal and nasal half removals were doneafter nerve cut. Similar removals were done without opticnerve cut to establish the location in the tectum of therepresentation of the cut retinal edge. For direct compari-son of half retinal regeneration with regeneration of thewhole retina, some whole retinal regenerates had half of theregenerate retina removed several days prior to labelling ofthe optic nerve. Thus, the representation of the half retinaafter whole retinal regeneration could also be examined.
To label the entire optic nerve a Fibrospum (Promonta)pledget soaked in a saturated solution of HRP (horseradishperoxidase, Type VI, Miles) in 0-1 M-phosphate buffer (PB)pH7-4 was applied to the distal nerve stump after severingthe nerve between the eye and initial lesion site. To ensuremaximal uptake, the nerve was severed with a sharp razorblade. Small groups of axons were labelled by damaging afew fascicles of axons within the retina using a glassmicropipette coated at the tip with HRP crystals. Followingeither procedure, animals were left for 2-3 days prior toreanaesthetization, perfusion with 0-6% saline and re-moval of the retinae and tecta which were immediatelyplaced into a solution of 0-05 % diaminobenzidine (DAB),001% hydrogen peroxide in 0-2M-PB pH7-2 for lh. Thetecta were then fixed for 2 h in 4 % glutaraldehyde in 0-2 M-

Half retinal regeneration 481
PB pH7-2 prior to flattening on a gelatinized slide followedby dehydration and mounting in Permount. The retinaewere fixed in 10 % PB formalin pH 7-4 for several days,then flattened and air dried onto 5 % gelatinized slides,dehydrated and mounted in Permount.
The quantification methods will be explained in therelevant results sections.
Results
In order to examine the overall pattern of regener-ation, the whole optic nerve was labelled in a series ofboth nasal (n = 11) and temporal (n = 6) half retinalregenerates, these were compared with control ani-mals where a half retina was removed but the nervewas not sectioned at the time of removal (n = 10).Attention was focussed on two periods of regener-ation. The first was 35-40 days after nerve sectionwhen the majority of axons should still have beengrowing towards their target regions and very fewwould have started to form terminal arborizations.The second was at 75-76 days when the tectumshould already be densely innervated and maturearbours are identifiable.
Distribution of axonsIn the control fish, which had not received optic nervesection, the pattern of labelling following removal ofeither the nasal or temporal half of the retina was aswould have been expected from previous studies ofthe normal pathways (Stuermer & Easter, 1984a;Easter & Stuermer, 1984). Temporal axons wereconfined to the rostral tectum in both the superficialfibre layer (SO) and synaptic layer (SGFS). Therewas a relatively sharp boundary between labelled andunlabelled tectum located approximately halfwayalong the rostrocaudal extent of the tectum. Terminalarbours in the deepest optic termination layers pro-jected slightly farther into the denervated tectum,presumably reflecting their greater size (Stuermer,1984). Nasal axons were present in rostral tectumboth within superficial fascicles and as fibres ofpassage (the 'extrafascicular' axons of Easter &Stuermer, 1984a; Stuermer & Easter, 1984a) in thesynaptic layer. Terminal arbours of nasal axons wereconfined to the caudal tectum.
In the regenerates that had half of the retinaremoved at the time of nerve section (i.e. half retinalregenerates) the fibre order was very different fromnormal. Also, in contrast to the findings of Attardiand Sperry, temporal axons were not completelyrestricted to the appropriate rostal tectum as somehad also invaded the caudal tectum. We were particu-larly interested in comparing the relative density ofaxons in each tectal region with those previouslydetermined in whole retinal regenerates (Stuermer,
1988a) as this would indicate whether more axonsentered the inappropriate tectal half under thesecircumstances. The density and orientation of axonscrossing a standard sample area was determined at sixrepresentative tectal sites in two animals for eachdatum point (short-term nasal and temporal, long-term nasal and temporal). These sites were locatedalong lines joining the nucleus rotundus and thecaudal ends of the slit and flattened tectum(Stuermer, 1988a) and comprised a medial and lateralregion in rostral, middle and caudal tectum. At eachsite, the total number of all axons within a circulararea of tectum 136 fim in diameter was determined ata magnification of xlOOO.
The plot of relative axon density versus rostrocau-dal tectal location (Fig. 1) clearly shows that temporalaxonal density (solid lines) declined drastically from70% in rostral tectum to 28% in midtectum andfinally 2 % in caudal tectum, at both stages examined,while nasal axons maintained an even density into thecaudal tectum (rostral, 30%; mid, 40%; caudal,30%). This is very similar to whole retinal regener-ates (Stuermer, 1988a). Therefore, both nasal andtemporal axons exhibit an equally strong preferencefor their retinotopically appropriate tectal halfwhether or not the axons appropriate to the othertectal half are present.
80-
70-
60-
50-
40-
30-
10-
Rostral MidTectal region
Caudal
Fig. 1. Plot of relative axon density versus tectal region.See results for detailed explanation. The axonal densitieswithin a standard sample area in each tectal regionindicated were summed and then the density at eachlocation expressed as a percentage thus providing astandardized measure of axonal distribution. Solid linesindicate temporal half retinal regenerates while brokenlines indicate nasal half retinal regenerates. Open circlesare 76-day regenerates while filled circles are 39-dayregenerates. At both stages, the axonal density oftemporal regenerates dropped dramatically in caudaltectum while that of nasal regenerates was similarthroughout the tectum.

482 M. F. Humphrey and C. A. O. Stuermer
Fascicle formationBoth nasal and temporal axons formed fascicleswithin stratum opticum (Fig. 2A,B). Either temporalaxons left the fascicles individually in rostral tectumor the fascicles themselves curved in a mediolateralorientation, in many cases crossing the tectal equator,and therefore did not enter caudal tectum (Fig. 2B).Groups of axons making similar loops were alsofound in the synaptic layer near the middle of therostrocaudal extent of the tectum (Fig. 2C). Suchpathways are rare or absent in the normal projectionand were not described by Attardi & Sperry (1963).
In whole retinal regenerates, axons were oftenfound to exit from fascicles and cross the tectalequator in the deeper, synaptic layer apparently tocorrect for having entered via the incorrect brachium(Stuermer, 1988a). It was very evident in the currentstudy that the temporal ingrowth tecta containedmany large mediolaterally oriented fascicles crossingthe rostral tectal equator in deeper layers (Fig. 2D).Such fascicles are occasionally seen in whole retinalregenerates but the frequency appeared much higherin the temporal half retinal regenerates. Many tem-poral ingrowth fascicles abruptly changed their orien-tation to mediolateral, entered the synaptic layer as
Fig. 2. Fascicle patterns.(A) Fascicles (arrowheads) inthe stratum opticum of atemporal half retinalregenerate. (B) A lateralfascicle bending sharply into amediolateral orientation(arrows), temporal half retinalregenerate. (C) A smallfascicle of temporal axonsbending sharply in the synapticlayer of the midtectum.(D) Large fascicles crossing thetectal equator in a temporalhalf retinal regenerate. (E) Therapid termination of a fascicleof temporal axons in a wholeretinal regenerate (arrow).Note how the axonsimmediately disperse uponentering the synaptic layer.(F) The synaptic layer in therostral tectum of a wholeretinal regenerate where onlythe temporal axons werelabelled. Note the evenmeshwork of axons andabsence of fascicles.Bar, 200 jum.
an intact fascicle, which, in many cases, then crossedthe tectal equator. In contrast, the majority of fas-cicles in whole retinal regenerates came to an abruptend as they approached the tectal equator (Fig. 2E)and individual axons entered the meshwork of axonsin the synaptic layer (Fig. 2F). Nasal axons formed afascicle pattern similar to that of whole retinal regen-erates and did not have an excessive degree ofequatorial crossing. Those nasal axons that did enterthe incorrect dorsal or ventral hemitectum crossedthe equator as individuals rather than in fascicles.
To examine whether the degree of fasciculation intemporal half retinal regenerates was objectivelygreater than that of whole retinal regenerates, thefascicle pattern of an animal that had the nasal retinaremoved at the time of optic nerve cut was comparedwith that of an animal in which the whole retina hadregenerated after nerve cut but only the temporalaxons were labelled. It was clear that in the wholeretinal regenerate, while some fascicles crossed theequator at the boundary between rostral and caudaltectum, the majority of fascicles dispersed into singlefibres before reaching the equator (Fig. 3B). Incontrast, the temporal half retinal regenerate con-tained large fascicles throughout the rostral tectum

Half retinal regeneration 483
which were oriented mediolaterally and continuedacross the tectal equator (Fig. 3A). In peripheraltectum, the superficial fascicles of the whole and halfretinal regenerates were similar although there wasmore fasciculation at deeper levels in the half retinal
0-5mm
Fig. 3. Fascicular paths drawn at a magnification of xl60in a temporal retinal regenerate (A), and a whole retinalregenerate (B) in which only temporal axons werelabelled. Both were labelled at 76 days after nerve cut.Note the increased crossing of the tectal equator in thehalf retinal regenerate and also the slightly greater spreadof fascicles into caudal tectum in the whole retinalregenerate. R, rostral; C, caudal; D, dorsal; V, ventral.Boxes indicate the regions analysed in greater detail.
regenerate. Drawings done at higher magnification(x400, oil immersion) also confirmed this pattern.
Thus, in the absence of nasal axons, temporalaxons seemed to associate with one another to agreater degree within rostral tectum. Thus, manytemporal axons crossed the tectal equator both indi-vidually and in fascicles. These crossings are found inall regenerates but rarely, if ever, in normals and arenot therefore consistent with axons following normalpathways (Attardi & Sperry, 1963). Also, fewerfascicles entered caudal tectum, possibly since nonasal axons were present to lead them astray. Nasalaxons followed pathways similar to those of wholeretinal regenerates crossing rostral tectum both infascicles and via deep extrafascicular pathways.
Single axon tracing
Discrete bundles of axons from defined retinal lo-cations were labelled in order to obtain informationabout the pathways taken by individual axons andtheir topographic appropriateness (six nasal re-movals, six temporal removals). Only the early stageof regeneration was examined in this manner sincethis was when the pathways were just being estab-lished in the tectum. In two nasal and two temporalhalf retinal ingrowth tecta, all the labelled axons weredrawn. Other tecta were examined without drawingand found to confirm the growth patterns of thedrawn tecta. At 35-40 days a large number of axonshad growth cones at their leading edge and also on thetips of side branches if branching had occurred. Thisconfirmed that the pathways were at an early stage offormation.
Compilation of all traced axons revealed a patternsimilar to that of half retinal regenerates in which thewhole nerve was labelled (Fig. 4A,B). Dorsotem-poral axons had a tendency to be directed towards themidlateral tectum (Fig. 4A). Axons travelled towardsthis region in various ways. Some axons, particularlythose far from the appropriate target region (oftendue to entering the incorrect brachium), continued inthe fascicle layer without branching (Fig. 4A, smallarrows). However, near the midtectum many axonswere found whose paths were curved towards theappropriate termination region (Fig. 4A, large ar-rows). Those that had passed beyond the retinotopicregion looped back (Fig. 4A, arrowheads), demon-strating a clear preference to search in rostral tectum.Within the rostral tectal half, axons outside thefascicles had branched, both close to their retinotopicregion and at ectopic sites. In whole retinal regener-ates where only the temporal axons were labelled, themajority of labelled axons branched in rostral tectumbut a few axonal ramifications were also found incaudal tectum. In half retinal regenerates, fewertemporal axons invaded caudal tectum and no

484 M. F. Humphrey and C. A. O. Stuermer
branching axons were observed in caudal tectum.Three typical temporal axons from Fig. 4A are shownseparately in Fig. 4B for greater clarity (axons 1, 2 &
3). One of these axons apparently continued extend-ing caudally, beyond the correct termination region,while maintaining an elaborate side branch directedtowards the retinotopically appropriate region(Fig. 4B, axon 3). Possibly this axon would haveeventually withdrawn the caudally directed processbut we have no direct evidence of this. The otheraxons (Fig. 4B, axons 1 & 2) are clearly branchingelaborately in the proximity of the correct regiondespite starting from relatively inappropriate tectalregions. Due to this type of course correction, themajority of temporal axons did not enter caudaltectum.
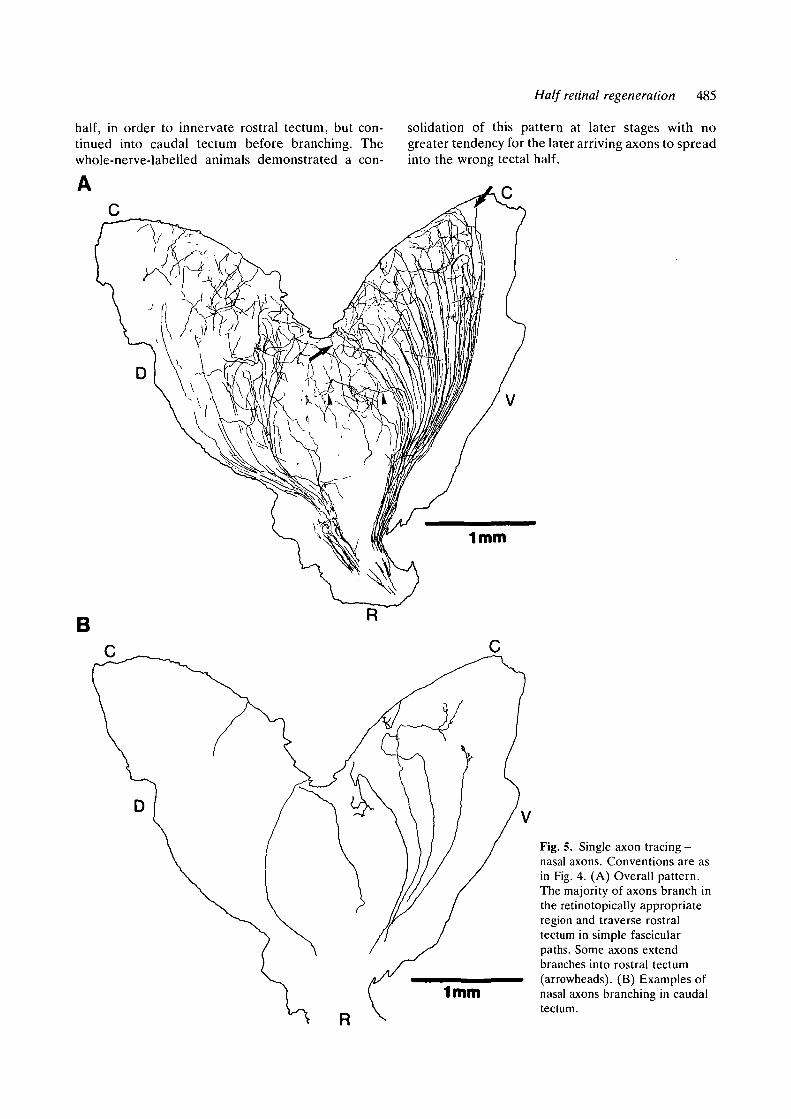
In the nasal ingrowth animals, it was evident that,although the axons had to grow through rostraltectum to reach their appropriate termination region,the majority of axons did not branch until enteringcaudal tectum (Fig. 5A,B). Only a few axons ex-tended branches into rostral tectum (Fig. 5A, arrow-heads). Fig. 5B shows the branching in caudal tectummore clearly and also examples of axons that crossrostral tectum without forming any elaborate pro-cesses. Those axons with growth cones at their tipswithin rostral tectum were usually directed towardsthe caudal tectum.
Longer term nasal and temporal half retinal regen-erate tecta were examined for the presence of maturearbours. In each case, they were only found in theretinotopically appropriate tectal half. The mor-phology of axons branching in the inappropriatetectal half was always immature.
In summary, the tectal pathways of nasal andtemporal axons were not greatly altered by theabsence of their respective partners from the otherhalf of the retina. The number of temporal axonsinvading the caudal tectum was no greater than inwhole retinal regenerates even though the caudaltectum was vacant. On the contrary, many temporalaxons made sharp course corrections to avoid theretinotopically inappropriate tectal half. Neither didnasal axons exhibit any increased tendency to enterand branch in the 'vacant' SGFS of the rostral tectal
Fig. 4. Single axon tracing - temporal axons. Thick linesindicate the tectal outline. The retinotopically appropriatetermination region for the labelled axons is indicated bythe large arrows. (A) Overall pattern. Most axons aredirected towards and branch within the retinotopicallyappropriate region. Some axons reach this region bycrossing the tectal equator because they entered thedorsal brachium (small arrows). Others turn sharply awayfrom caudal tectum (large arrowheads) and somecontinue into caudal tectum. (B) Examples of widelybranching axons (1,2) and an axon (3) continuing to growcaudally despite an elaborate side branch directedtowards the appropriate region.
Half retinal regeneration 485
half, in order to innervate rostral tectum, but con-tinued into caudal tectum before branching. Thewhole-nerve-labelled animals demonstrated a con-
solidation of this pattern at later stages with nogreater tendency for the later arriving axons to spreadinto the wrong tectal half.
Fig. 5. Single axon tracing -nasal axons. Conventions are asin Fig. 4. (A) Overall pattern.The majority of axons branch inthe retinotopically appropriateregion and traverse rostraltectum in simple fascicularpaths. Some axons extendbranches into rostral tectum(arrowheads). (B) Examples ofnasal axons branching in caudaltectum.

486 M. F. Humphrey and C. A. O. Stuermer
Discussion
The major conclusion of this study is that even whenhalf of the retina is removed, a procedure thatprevents certain interactions between axons andleaves a large area of tectum without its appropriateinnervation, both nasal and temporal axons exhibit apreference to grow through their retinotopic tectalhalf. The semiquantitative methods used allow adirect comparison with similar analyses done on nasaland temporal axons in whole retinal regenerates(Stuermer, 1988a,b). Axons exhibit similar prefer-ences during whole retinal regeneration even though,as in half retinal regenerates, they follow manyinappropriate routes through the tectum. Interaxonalinteractions cannot therefore be of major importancein this specificity since the degree of preference issimilar in these two situations.
In whole retinal regeneration, a low proportion oftemporal axons extend into the inappropriate caudaltectal half (Stuermer, 1986, 1988a,b). The presentstudy has demonstrated that the proportion of theseaxons is no greater in the absence of axons appropri-ate to the caudal tectum. In fact, fewer temporalaxons entered the caudal tectum via fascicles. Thissuggests that axons that enter the caudal tectumthrough fascicular pathways in whole retinal regener-ates do so because they are following nasal axons.Similarly, nasal axons of half retinal regenerates weredistributed as in whole retinal regenerates eventhough they had to pass through rostral tectum whichlacked its appropriate innervation. Thus, denervatedtectum appears not to be attractive to ingrowingaxons unless it is of the appropriate retinotopicspecificity.
Thus, with minor exceptions, the pattern of bothnasal and temporal axonal innervation is the samewhether they regenerate together or separately.Attardi & Sperry (1963) reached similar conclusions,however, the limitations of the techniques availableat the time led them to assume that regeneratingaxons follow their previous pathways. Numerousrecent studies have shown that, on the contrary,regenerating axons take abnormal routes through thetectum but nevertheless relocate their retinotopictermination sites (Cook, 1983; Rankin & Cook, 1986;Stuermer, 1986, I988a,b; Stuermer & Easter, 1984ft).Analysis of these pathways shows that axons do noterr randomly either in whole retinal regeneration(Stuermer, 1988a,b) or when large regions of thetectum are left without appropriate innervation (cur-rent study). Since the restriction of axons to theirappropriate tectal half is not due to competitiveinteractions between the axons and since axonalpathways are directed to their correct termination
regions despite considerable pathway disorganiz-ation, we favour the view that axons follow positionalcues present in the tectal neuropil. Studies of retino-tectal projection formation during development inXenopus and zebra fish also found no indication ofinteraxonal competition affecting the initial arboriz-ation pattern (Holt, 1984; Stuermer, 1988c). There-fore, recognition of tectal positional cues by opticaxons seems to play a major role in the initialtopography of various systems.
In half retinal regeneration, it is not possible for theage-related order of normal fascicles (Cook & Ran-kin, 1984; Easter & Stuermer, 1984; Stuermer &Easter, 1984a) to be reproduced and yet fascicleformation was very obvious in these animals,although crossing of the tectal equator was veryfrequent for temporal axons. In whole retinal regen-erates, a fascicle pattern also forms which is veryreminiscent of that in normals, however, the organiz-ation is of a much lower grade and fascicles some-times cross the tectal equator (Cook, 1983; Stuermer& Easter, 19846; Stuermer, 1986). Reformation of afascicle pattern in this study is therefore furtherevidence that the tendency to fasciculate in stratumopticum is more related to the tectal environmentthan axonal interactions.
Temporal axons of half retinal regenerates tendedto associate very closely and often remain in fascicleseven after turning in a mediolateral direction. Thisincreased fasciciilation probably reflects several fac-tors. With no nasal axons to lead temporal axonsastray they may form more obvious mediolateralfascicles. Additionally, the absence of nasal axonsintermingled with temporal axons in the fascicles maylead to a closer association which is maintained whenthe axons leave stratum opticum. Finally, there maybe an inherent tendency for temporal axons toassociate more closely which is expressed in thesecircumstances (Bonhoeffer & Huf, 1985; Gaze &Fawcett, 1983).
The majority of nasal axons in half retinal regener-ates crossed rostral tectum within fascicles and onlyexited from the fascicles in caudal tectum, thusforming long fascicular paths in stratum opticum. Thefinding of Meyer (1987) that responses could berecorded from rostral tectum at 44-64 days in nasalhalf retinal regenerates is not readily explained by ouranatomical results. No mature arbours were found inthe rostral tectum of nasal half retinal regenerates atthese stages in this study. Some axons did, however,form immature branches and it is possible that carefulrecording would detect these.
Optic axons certainly have the capacity for wide-ranging growth in the tectum as demonstrated bytheir ability to innervate a surgically isolated tectalisland (Edwards et al. 1985), however, the majority

Half retinal regeneration 487
remain restricted to their appropriate tectal half.Much of this restriction evidently occurred in thestratum opticum, with comparatively few cases ofwide rostrocaudal growth in stratum opticum sub-sequently corrected by pathways in the synaptic layer.This suggests that some tectal positional informationmust be available in stratum opticum.
The nature of the positional information in thefascicular layer is uncertain. The ingrowing axons areunmyelinated and usually tightly clustered aroundone another (Murray, 1976; Murray & Edwards,1982; Wolburg & Bouzehouane, 1986). The fasciclesare located in the same position as the fascicles of theprevious projection and sometimes contact glial cellsor other elements of the tectal neuropil. Although alarge part of the axonal environment is composed ofother axons we have seen that elimination of asubstantial number of them did not influence thedistribution of the fascicles. It is therefore most likelythat the axons are influenced by markers on otherelements. Although most of the axons from theprevious projection have degenerated by the timethat the regenerating axons enter, it is possible thatthey are following the remnants of the previousprojection, but in a rather nonspecific way. It has alsobeen suggested that the previous projection leaves'markers' imprinted on the tectum which allowsorganization of the regenerating axons (Schmidt,1978). The role of the previous projection is currentlyuncertain. Some studies suggest that it plays a majorrole and that long-term denervation prior to regener-ation allows immediate expansion of a half retinalprojection (Schmidt, 1978). Other studies, however,show minimal effects of long-term denervation(Meyer, 1978, 1984, 1987; Busse & Stuermer, 1987).One study in particular shows that the fascicularpathways in long-term denervated tecta are similar tothose after simple regeneration suggesting that de-generating debris from the previous projection is notnecessary for these pathways to form (Busse &Stuermer, 1987). Furthermore, these axons terminateat their retinotopically appropriate sites, substantiat-ing the view that long-lasting markers inherent to thetectum guide the growing axons.
At early stages of regeneration, the axons ramifiedwidely in the synaptic layer of the tectum, havingmany fine branches with growth cones at their tips.Single axon tracing showed that, as in whole retinalregenerates, these fine arbours were not alwayslocated in the appropriate tectal region. At laterstages, only few immature branches were located inthe inappropriate tectal half and mature arbourformation was confined to the appropriate tectal half.This sequence of events is consistent with the earlybranching being a strategy for more accurately sens-ing tectal cues and thereby finding a pathway to the
appropriate termination site (Fujisawa et al. 1982;Stuermer, 1987b) rather than simply spreading tocover as much denervated tectum as possible prior toselecting appropriate branches. The axons that enterthe inappropriate tectal half in half retinal regener-ation have no axons with which to compete andtherefore competition between axons cannot be afactor in their growth out of the incorrect region.
MFH would like to thank Prof. H. Wassle for provisionof facilities and J. Sapp for typing the manuscript.
References
ATTARDI, D. G. & SPERRY, R. W. (1963). Preferentialselection of central pathways by regenerating opticfibers. Expl Neurol. 7, 46-64.
BONHOEFFER, F. & HUF, J. (1985). Position-dependentproperties of retinal axons and their growth cones.Nature, Lond. 315, 409-410.
BUSSE, U. & STUERMER, C. A. O. (1987). Navigation andtarget recognition of regenerating retinal axons in long-term denervated tecta in goldfish. Neurosci. Abstr. 13,1418.
COOK, J. E. (1983). Tectal paths of regenerated opticaxons in goldfish: evidence from retrograde labellingwith horseradish peroxidase. Expl Brain Res. 51,433-442.
COOK, J. E. & RANKIN, E. C. C. (1984). Use of alectin-peroxidase conjugate (WGA-HRP) to assess theretinotopic precision of goldfish optic terminals.Neurosci. Lett. 48, 61-66.
EASTER, S. S., JR & STUERMER, C. A. O. (1984). Anevaluation of the hypothesis of shifting terminals ingoldfish optic tectum. J. Neurosci. 4, 1052-1063.
EDWARDS, M. A., SHARMA, S. C. & MURRAY, M. (1985).Selective retinal reinnervation of a surgically createdtectal island in goldfish. I. Light microscopic analysis.J. comp. Neurol. 232, 372-385.
FUJISAWA, H., TANI, N., WATANABE, K. & IBATA, Y.(1982). Branching of regenerating retinal axons andpreferential selection of appropriate branches forspecific neuronal connections in the newt. Devi Biol.90, 43-57.
GAZE, R. M. & FAWCETT, J. W. (1983). Pathways ofXenopus optic fibres regenerating from normal andcompound eyes under various conditions. J. Embryol.exp. Morph. 73, 17-38.
HOLT, C. E. (1984). Does timing of axon outgrowthinfluence initial retinotectal topography in Xenopus] J.Neurosci. 4, 1130-1152.
HUMPHREY, M. F. & STUERMER, C. A. O. (1987). Tectalpathways of regenerating goldfish optic axons afterhalf-nasal or half-temporal retinal removal. Neurosci.Abstr. 13, 1419.
MEYER, R. L. (1978). Deflection of selected optic fibersinto a denervated tectum in goldfish. Brain Res. 155,213-227.

488 M. F. Humphrey and C. A. O. Stuermer
MEYER, R. L. (1979). Retinotectal projection in goldfishto an inappropriate region with a reversal in polarity.Science 205, 819-821.
MEYER, R. L. (1980). Mapping the normal andregenerating retinotectal projection of goldfish withautoradiographic methods. J. comp. Neurol. 189,273-289.
MEYER, R. L. (1984). Target selection by surgicallymisdirected optic fibers in the tectum of goldfish. J.Neurosci. 4, 234-250.
MEYER, R. L. (1987). Tests for relabelling the goldfishtectum by optic fibers. Devi Brain Res. 31, 312-318.
MEYER, R. L., SAKURAI, K. & SCHAUWECKER, E. (1985).Topography of regenerating optic fibers in goldfishtraced with local wheat germ injections into retina:Evidence for discontinuous microtopography in theretinotectal projection. J. comp. Neurol. 239, 27-43.
MURRAY, M. (1976). Regeneration of retinal axons intothe goldfish optic tectum. J. comp. Neurol. 239, 27-43.
MURRAY, M. & EDWARDS, M. A. (1982). A quantitativestudy of the reinnervation of the goldfish optic tectumfollowing optic nerve crush. J. comp. Neurol. 209,363-373.
RANKIN, E. C. C. & COOK, J. E. (1986). Topographicrefinement of the regenerating retinotectal projectionof the goldfish in standard laboratory conditions: aquantitative WGA-HRP study. Expl Brain Res. 63,409-420.
SCHMIDT, J. T. (1978). Retinal fibers alter tectal positionalmarkers during the expansion of the half retinalprojection in goldfish. J. comp. Neurol. Yll, 279-300.
SCHMIDT, J. T. & EDWARDS, D. L. (1983). Activitysharpens the map during the regeneration of theretinotectal projection in goldfish. Brain Res. 269,29-39.
SHARMA, S. C. & TUNG, Y. L. (1979). Interactionsbetween nasal and temporal hemiretinal fibers in adultgoldfish tectum. Neurosci. 4, 113-119.
STUERMER, C. A. O. (1981). Modified retinotectalprojection in goldfish: a consequence of the position ofretinal lesions. In Lesion-Induced Neuronal Plasticity inSensorimotor Systems (ed. H. Flohr & W. Precht).Berlin, Heidelberg: Springer-Verlag.
STUERMER, C. A. O. (1984). Rules for retinotectalterminal arborizations in the goldfish optic tectum: Awhole-mount study. J. comp. Neurol. 229, 214-232.
STUERMER, C. A. O. (1986). Pathways of regeneratedretinotectal axons in goldfish. 1. Optic nerve, tract andtectal fascicle layer. J. Embryol. exp. Morph. 93, 1-28.
STUERMER, C. A. O. (1988a). The trajectories ofregenerating retinal axons in the goldfish tectum. I. Acomparison of normal and regenerated axons at lateregeneration stages. /. comp. Neurol. 267', 55-68.
STUERMER, C. A. O. (19886). The trajectories ofregenerating retinal axons in the goldfish tectum.II. Exploratory branches and growth cones on axons atearly regeneration stages. J. comp. Neurol. 267, 69-91.
STUERMER, C. A. O. (1988c). Retinotopic organization ofthe developing retinotectal projection in zebra fishembryos. J. Neurosci. (in press).
STUERMER, C. A. O. & EASTER, S. S., JR (1984a). Rulesof order in the retinotectal fascicles of goldfish. J.Neurosci. 4, 1045-1051.
STUERMER, C. A. O. & EASTER, S. S., JR (19846). Acomparison of the normal and regenerated retinotectalpathways of goldfish. J. comp. Neurol. 223, 57-76.
WOLBURG, H. & BOUZEHOUANE, U. (1986). Comparisonof the glial investment of normal and regenerating fiberbundles in the optic nerve and optic tectum of thegoldfish and crucian carp. Cell Tissue Res. 244,187-192.
(Accepted 17 November 1987)