target and differentiation terapy - giuseppe di bella · inibizione:ras (certa) inibizione enos...
TRANSCRIPT

Target and Differentiation Terapyin oncologia
Prof. Maurizio PianezzaDott. Stefano Cucumazzo

Cancro: malattia genetica acquisita dalle cellule somatiche
Conferma della definizione: aberrazioni cromosomiche
Biologia molecolareCitogenetica
Copyright Fondazione Raphael - Melide (CH)

Cellule tumorali: deviazione del normale valore diploide dei cromosomiPloidia Combinazioni MorfologiaMonosomie delezioneTrisomie duplicazionePolisomie inversione
traslocazioneMSR: regioni cromosomiche amplificate all’interno del braccioDM: regioni cromosomiche amplificate all’esterno del braccio
Copyright Fondazione Raphael - Melide (CH)

BANDE CROMOSOMICHE
71 su 329Riarrangiamenti tumore-associati
Copyright Fondazione Raphael - Melide (CH)

La citogenetica consente di individuare due classi di geni implicati nello sviluppo e progressione tumorale
Proto – oncogenesTumor Suppressors Genes
Copyright Fondazione Raphael - Melide (CH)

Attivazione dei proto-oncogeni1° meccanismo:Aumento proteine del gene per iperespressione del gene stessoDelezione del dominio regolatorio
2° meccanismoTrasposizione per traslocazionecromosomica
3° meccanismoAmplificazione genica
Copyright Fondazione Raphael - Melide (CH)

attivazioneProto-oncogene Oncogene
Differenti compartimenticellulari
recettore fattore di sistemi di sistemi di direttocrescita trasduzione trascrizione su proteine
del segnale del ciclocellulare
bersaglio
Copyright Fondazione Raphael - Melide (CH)

Inattivazione dei Tumor Suppressor Genes1° evento
Si inattiva uno dei due alleliLinea germinale: tumori “ereditari”Mutazione somatica: tumori comuni
2° eventoMutazione somatica che inattiva la restante parte allelica (allele selvaggio)
La regione cromosomica che contiene l’allele selvaggio viene eliminata attraverso:
Non disgiunzione Mancata Ricombinazione mitoticaConversione genica
Copyright Fondazione Raphael - Melide (CH)

Eventi genetici nella genesi dei tumori solidi
Eventi genetici distinti:Attivazione oncogeniInattivazione tumor suppressors genes
Lesioni genetiche multiple: PERDITA ALLELI CHE SI VALUTA CON MARKER D.N.A. CONTROPARTE MOLECOLARE ALLELOTIPO
Copyright Fondazione Raphael - Melide (CH)

Cambiamenti genetico molecolari: ploidia del tumore
Tumori solidi aneuploidia instabilità genomica
Progressione dimalattia
Aumento numerico dei cromosomi
Ipometilazione
Gene di Divisione cellulare
Modello ComputerizzatoDi Shackney
Non-disgiunzionemitotica
Cloni iperdiploidii
Cloni ipodiploidi
Perdita di cromosomiTetraploidizzazioneCrescita cellulareDivisione cellulareGrowningPromotingAberrazione cromosomicaCopyright Fondazione Raphael - Melide (CH)

Aneuploidia Attivazione oncogenica
Doppia e sequenziale sovraesposizionenon disgiunzione del cromosoma
Copyright Fondazione Raphael - Melide (CH)

Cromosoma eucarioteCROMATINA
Complesso DNAProteine CromosomicheRNA Nucleare
EUCROMATINA ETEROCROMATINA
geneticamente attiva geneticamente inattiva
Copyright Fondazione Raphael - Melide (CH)

Struttura del DNA
Copyright Fondazione Raphael - Melide (CH)

Proteine cromosomicheNucleosomaDNA avvolto sugli istoniH1 sigilla il DNA
Tra 2 nucleosomi: DNA detto “Linker”
Proteine IstonicheH1H2aH2bH3H4
Proteine non IstonicheEnzimiProteine di trascrizioneProteine di replicazione
Interazione proteine non istoniche che legano il DNA
SUL:Centromero: regolano DNA in mitosi e meiosievita non disgiunzione dei cromosomiCinetocromo: consentono attacco fibre del fusomitotico Telomero: evitano rimaneggiamenti cromosomi
Copyright Fondazione Raphael - Melide (CH)
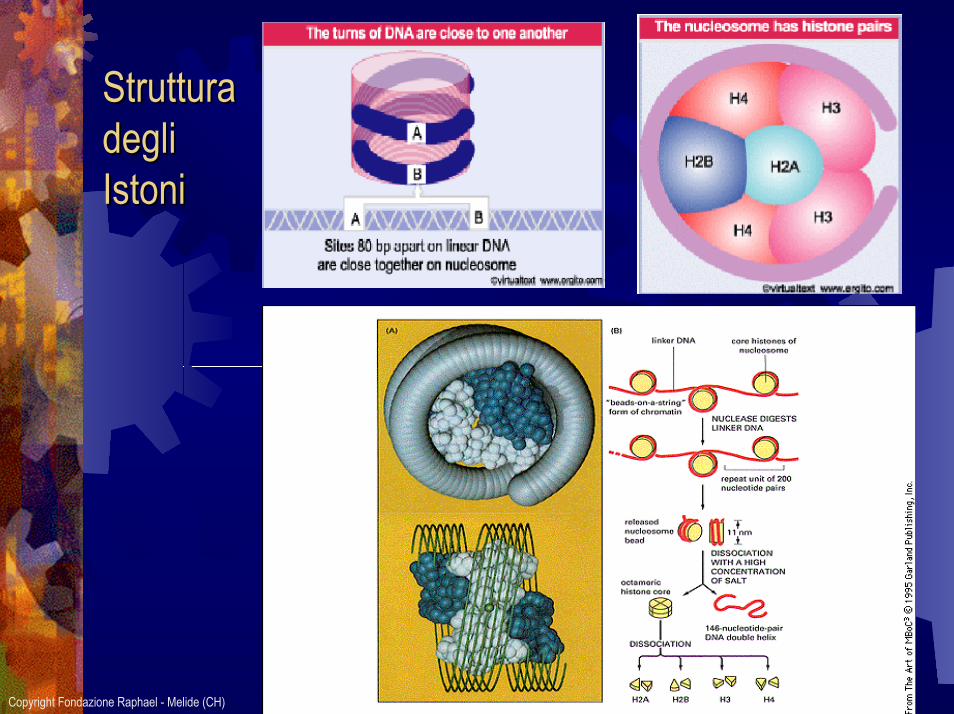
Struttura Struttura deglidegliIstoniIstoni
Copyright Fondazione Raphael - Melide (CH)

Struttura del nucleosoma
Copyright Fondazione Raphael - Melide (CH)

Copyright Fondazione Raphael - Melide (CH)

Copyright Fondazione Raphael - Melide (CH)

RECETTORE
Molecola che lega in modo specifico, definito e con affinità precisa uno o più mediatori endogeni e che, in seguito al legame, subisce una modificazione conformazionale in grado di determinare un effetto biologico
In base alla localizzazione subcellulare:1)RECETTORI DI MEMBRANA2)RECETTORI INTRACELLULARI
Copyright Fondazione Raphael - Melide (CH)

Recettori di membrana6 SUPERFAMIGLIE:
•RECETTORI CANALE O IONOTROPI•RECETTORI ACCOPPIATI ALLE PROTEINE G O METABOTROPI•RECETTORI CON ATTIVITA’ TIROSIN CHINASICA INTRINSECA•RECETTORI CON ATTIVITA’ GUANILATO CICLASI INTRINSECA•RECETTORI PER L’ADESIONE CELLULARE•RECETTORI PER LE CITOCHINE
RECETTORE CANALE trasduzione del segnale molto rapidaRECETTORE ACCOPPIATO ALLE PROTEINE G trasduzione del segnale più lunga e più lenta
Copyright Fondazione Raphael - Melide (CH)
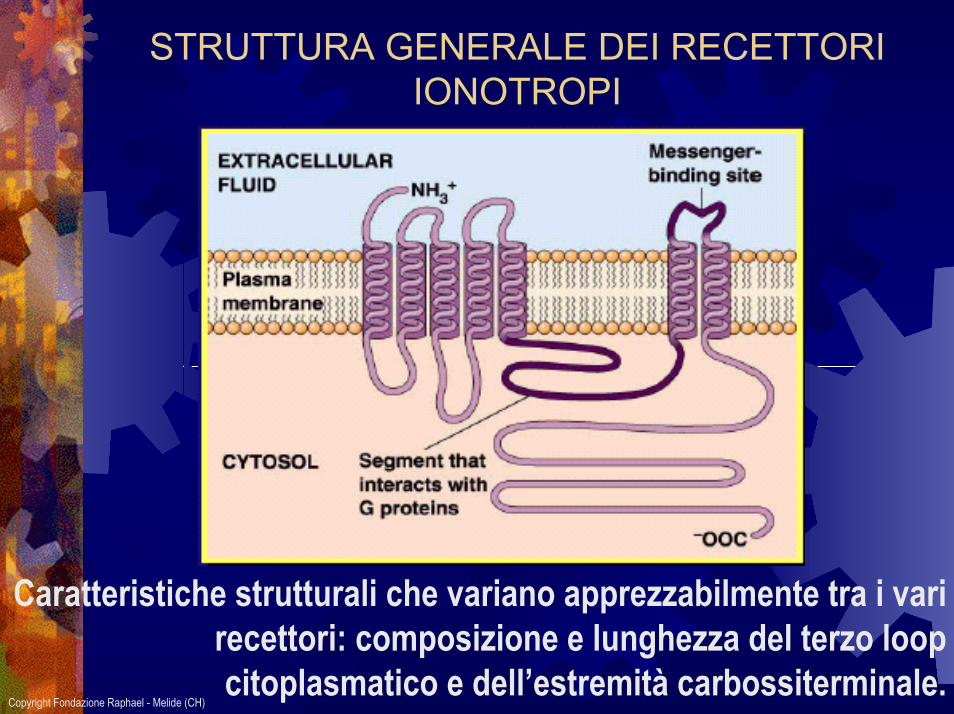
STRUTTURA GENERALE DEI RECETTORI IONOTROPI
Caratteristiche strutturali che variano apprezzabilmente tra i vari recettori: composizione e lunghezza del terzo loop citoplasmatico e dell’estremità carbossiterminale.
Copyright Fondazione Raphael - Melide (CH)

RECETTORI METABOTROPIProteine recettoriali formate da una singola catena polipeptidica che attraversa la membrana plasmaticain corrispondenza di altrettante regioni idrofobiche(regioni transmembranaTM). La catena polipeptidica si organizza spazialmente nella membrana in modo da costituire una particella globulare.Sito di legame per il neurotrasmettitore: porzioni transmembranarie o extracellulari della sequenza amminoacidica.
Copyright Fondazione Raphael - Melide (CH)

RECETTORI METABOTROPITratto di sequenza tra
TM5 e TM6: rivolto verso il citoplasma;
importante per il riconoscimento delle proteine G specifiche
con cui ciascun recettore si accoppia QUINDI responsabile
della specificità dell’effetto
dell’attivazione recettoriale.Copyright Fondazione Raphael - Melide (CH)

Struttura del recettore del fattore di crescitaepidermico EGF RECETTORI TIROSINO CHINASICI
Copyright Fondazione Raphael - Melide (CH)
Attivazione del recettore di EGF

Proteine G
Eterotrimeri costituiti da 3 subunità: α, ß, γ.α è in grado di legare il GTP e di idrolizzarlo grazie a una attività GTPasica intrinsecaLe proteine G trasferiscono informazioni dai recettori alle molecole effettrici attraverso un ciclo di attivazione-deattivazione governato dal legame e dall’idrolisi del GTP.
Copyright Fondazione Raphael - Melide (CH)

Recettori canaleo ionotropi
Recettori accoppiati alle proteine G o metabotropi
Copyright Fondazione Raphael - Melide (CH)

EFFETTORI DELLE PROTEINE G CON FUNZIONI DI CANALE IONICO
I più studiati sono i canali al calcio operati da proteina G e un numero sempre più crescente di canali al potassio
Copyright Fondazione Raphael - Melide (CH)

ATTIVAZIONE ED INATTIVAZIONE DELLE PROTEINE G
Le proteine G passano continuamente da una conformazione inattiva (GDP legato) ad una attiva (GTP legato). La transizione verso la forma attiva è indotta dal recettore; il ritorno alla forma inattiva è garantito
dalla proprietà della subunità αche idrolizza il GTPCopyright Fondazione Raphael - Melide (CH)

SISTEMI EFFETTORIIl segnale generato dall’interazione recettore proteina G viene tradotto in un
messaggio cellulare attraverso l’attivazione di EFFETTORI specifici:
Copyright Fondazione Raphael - Melide (CH)

Meccanismo di azione tirosino chinasico
Copyright Fondazione Raphael - Melide (CH)

Recettori a tirosin-chinasi
Copyright Fondazione Raphael - Melide (CH)

ATTIVAZIONE DELLA CASCATA DELLE MAP CHINAS
Le MAP chinasi (proteinchinasi attivate da mitogeni) rivestono un ruolo centrale in processi cellulari importanti quali la proliferazione e il differenziamento. Possono essere attivate tramite vie diverse che coinvolgono o l’attivazione di recettori per i fattori di crescita ad attività tirosin chinasica o l’attivazione di recettori associati a
proteine G. Le MAP chinasi sono fosfoproteine fosforilate da una chinasi chiamata MEK.
L’attivazione delle MAP chinasi determinano traslocazione al nucleo, con conseguente fosforilazione di proteine nucleari, soprattutto fattori di trascrizione che
regolano l’espressione di geni essenziali per la stimolazione di crescita e differenziamento cellulari. Copyright Fondazione Raphael - Melide (CH)

MECCANISMI DI TRASDUZIONE DEL SEGNALE
La TRASDUZIONE DEL SEGNALE viene definita come la capacità della cellula di modificare il proprio funzionamento in risposta a una interazione recettore-ligando Il ligando rappresenta il PRIMO MESSAGGERO.
Copyright Fondazione Raphael - Melide (CH)

EFFETTORI DELLE PROTEINE G CON FUNZIONE ENZIMATICA
I secondi messaggeri sono l’anello di congiunzione tra l’attivazione recettoriale e la generazione della risposta cellulare. Il primo messaggero è invece il neurotrasmettitoreresponsabile dall’attivazione del recettore.Dato che il secondo messaggero è in grado di diffondere nel citoplasma, esso è anche in grado di regolare l’attività di processi cellulari che hanno luogo in sedi distanti.I più studiati sono l’ADENILATO CICLASI e la FOSFOLIPASI C che generano rispettivamente i SECONDI MESSAGGERI cAMP e IP3/DAG.
Copyright Fondazione Raphael - Melide (CH)

RECETTORI ACCOPPIATI ALL’ADENILATO CICLASI
L’adenilatociclasi è un enzima ubiquitario che converte ATP in cAMP; l’attività può essere inibita o stimolata da vari recettori
associati alle proteineG. Il cAMP svolge la sua azione di secondo messaggero regolando l’attività di diverse proteine cellulari, in
particolare le proteinchinasi A (PKA). Attraverso questo meccanismo il Camp controlla una serie di
processi cellulari: reazioni metaboliche, secrezione, rilasciamento delle cellule muscolari lisce, attività di canali ionici, trascrizione di
geni specifici. Copyright Fondazione Raphael - Melide (CH)

RECETTORI ACCOPPIATI ALLA FOSFOLIPASI C
Il calcio si lega alla proteina calmodulina e il complesso CA++-calmodulina determina l’attivazione
di una serie di risposte cellulariCopyright Fondazione Raphael - Melide (CH)

STIMOLAZIONE E INIBIZIONE DELL’ADENILATO CICLASI AD OPERA DI DIFFERENTI PROTEINE G
(a)Il legame della noradrenalina al recettore ß attiva Gsche attiva l’adenilatociclasi. Questo genera cAMP che attiva l’enzima a valle protein-chinasi A
(b)Il legame della noradrenalinaal recettore α2 attiva Gi che inibisce l’adenilatociclasi Copyright Fondazione Raphael - Melide (CH)

RECETTORI ACCOPPIATI ALLA FOSFOLIPASI C
Le due vie di attivazione, calcio e proteinchinasi C (PKC), regolano svariati
processi cellulari (metabolismo,
secrezione, contrazione, attività neuronale,
proliferazione)Copyright Fondazione Raphael - Melide (CH)

AMPLIFICAZIONE DEL SEGNALE CAUSATA DALLE CASCATE DEL SECONDO MESSAGGERO
ASSOCIATO ALLE PROTEINE G
Copyright Fondazione Raphael - Melide (CH)

SCHEMA RIASSUNTIVO
Copyright Fondazione Raphael - Melide (CH)

Copyright Fondazione Raphael - Melide (CH)
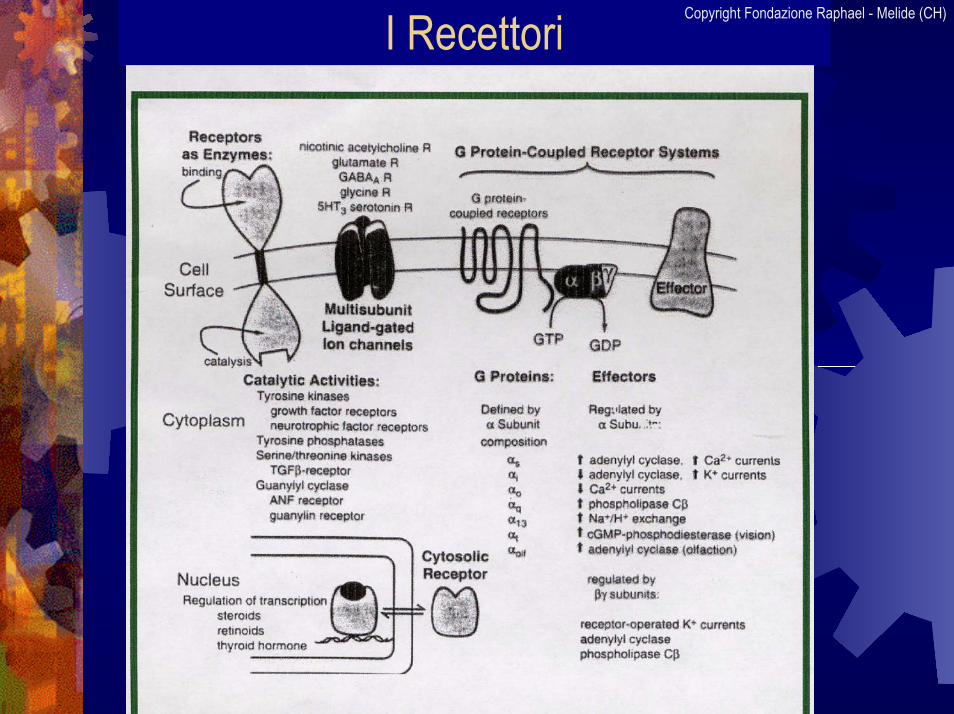
I Recettori Copyright Fondazione Raphael - Melide (CH)

Feed-back Terapeutico:differenziazione blocco proliferativo
Effettori extracellulari
Recettori diMembranaCitoplasmatica Trasduttori del
SegnaleEffettori Intracellularie secondi messaggeri
Sintesi Proteica
Recettori nucleariRNA M.T.R. Piccoli effettori
Modulazione Modulazione espressione Espressione genica genica diretta indiretta
Copyright Fondazione Raphael - Melide (CH)

FarmacodinamicaLa farmacodinamica studia gli effetti biochimici e il meccanismo d’azione dei farmaci proponendosi di:
identificare i siti di azione dei farmacidelineare le interazioni fisiche o chimiche tra farmaco e cellulacaratterizzare la sequenza completa farmaco-effettodefinire le basi per l’uso razionale dei farmaci e per il disegno di nuovi farmaci
Copyright Fondazione Raphael - Melide (CH)

FarmacomodulazionePresuppone la conoscenza di:
meccanismi biomolecolari normali e nel cancroconoscenza dei dosaggi terapeutici ottimali e i sinergismi tra di essi
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariTrastuzumab (herceptin)
Legame con ER2-NEUcodificato da gene C-ERB2Tirosino chinasicoFamiglia EGFR
MammellaStomaco
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariImatinib (Glivec)
Legame con CD117Tirosino Chinasico
Leucemia mieloide cronicaGIST
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariRituximab (Mabthera)
Legame CD20Citotossicità complemento-mediataCitotossicità cellulo-mediata
Linfoma Non-Hodgkin
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariGentuzumab Ozogamicin (Milotarg)
Legame CD33+Citotossicità come rituximab
Leucemia mieloide acuta
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariBortezomid (Velcade)
Blocco del proteosoma
Mieloma multiplo
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariGefitinib (Iressa)
Legame EGFRTirosino-Chinasico
PolmoneApparato digerente
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariCetuximab (Erbitux)
Legame EGFRTirosino ChinasicoMaggiore efficacia se su pezzo operatorio riscontro immunoistochimico di R1
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariBevacizumab (Avastin)
Legame VEGF
Tutti i tumori solidi con prevalenza di quelli ipervascolarizzati
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariSU11248 (sperimentale)
Legame VEGFLegame PDGF-R
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariBAY43-9009 (sperimentale)
Inibizione del RAFchinasiPromotore G proteins per effetto antiproliferativo ed antiangiogenetico
Tutti i tumori
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariBatimastat, Barimastat, Ag3340, BAY219566 (sperimentali)
Inibitori delle metalloproteinasi intercellulariInibizione angiogenetica
Tutti i tumori
Copyright Fondazione Raphael - Melide (CH)

Farmaci target molecolariFarnesil difosfato analoghi, CAAXtetrapeptide analogo
Inibitore della farnesil transferasi con interruzione dell’attività della proteina RAS-H, RAS-K, RAS-NBlocco del messaggio recettoriale da EGF, PDGF, GM-CSF, IL2, IL3
Copyright Fondazione Raphael - Melide (CH)

Farmaci Molecolari OrmonaliEstrogeni (Antagonisti ormonali)
Dietil-StilbestroloDietilen-estradioloFosfestrolo-tetrasodico
Progestinici Medrossi-progesterone acetato (Provera, Farlutal)Megestrolo-Acetato (Megace)Noretisterone (non registrato in Italia)
Copyright Fondazione Raphael - Melide (CH)

Farmaci Molecolari OrmonaliInibitori delle aromatasi (impediscono conversione androgeni--> estrogeni)
Aminoglutetimide NS (Orimeten)Anastrozolo NS (Arimidex)Letrozolo NS (Femara)Formestan S (Entaron)Esemestan S (Aromasin)Trilostato S (non registrato in Italia)
Copyright Fondazione Raphael - Melide (CH)

Farmaci Molecolari OrmonaliInibizione recettoriale, antagonista ormonale
Tamoxifene (Nolvadex)Toremifene (Fareston)Fulvestrant (Fasiodex)
Copyright Fondazione Raphael - Melide (CH)

Farmaci Molecolari OrmonaliAntiandrogeni, antagonisti ormonali
Ciproterone (Androcur)Flutamide (Eulexin)Bicalutamide (Casudex)
Copyright Fondazione Raphael - Melide (CH)

Farmaci Molecolari OrmonaliInibitori LHRH, analoghi ormone di rilascio
Buserelina (Suprefat)Goserelin (Zoladex)Leuprorelina (Enantone)Triptorelina (Decapeptil)
Copyright Fondazione Raphael - Melide (CH)

Somatostatina, Octreotide, Lanreotide, Vapreotide* Copyright Fondazione Raphael - Melide (CH)
Azione antiproliferativa:
Diretta Indiretta
SST1-2-3-4-5 IGFR
Antiproliferativa Antiangiogenetica Antiproliferativa Antiangiogenetica Differenziante Differenziante
FosforilazionePTP Inibizione Inattivazione ERK Inibizione(fosfotirosin fosfatasi) bFGF IGFR
Inibizione:RAS (certa) Inibizione ENOS Inattivazione SRC (certo) Attivazione PI3K-AKTSRC (incerto) Ceramide UpRegolation P27 (incerto)
espressione Ki67 (incerto)
Defosforilazione ERK Inibizione di NO Inibizione eNOS-NOG-Protein inibizione SHP-2C-SRC
Blocco Proliferativo Blocco Angiogenetico Blocco Proliferativo Blocco Angiogenetico

Grazie per la cortese attenzione

Il cancro è una malattia in cui le cellule sfuggono al meccanismo di controllo della crescita indispensabile per mantenere normale lastruttura e le funzioni dei tessuti
Copyright Fondazione Raphael - Melide (CH)

Il concetto che il cancro sia una malattia genetica delle cellule somatiche ha trovato concretezza grazie agli studi di biologia molecolare e citogenetica molecolare. La prova della teoria della mutazione somatica è il frequente riscontro di aberrazioni cromosomiche nelle neoplasie solide, presentando peraltro uncariotipo molto più complesso rispetto alle neoplasie ematologichee quindi i cambiamenti cromosomici tumore specifici si rendono più difficoltosi.
Copyright Fondazione Raphael - Melide (CH)

La tumorogenesi è infatti considerata come il risultato finale di eventi genetici multipli che portano le cellule alla perdita delcontrollo della crescita ed alla metastatizzazione.
Copyright Fondazione Raphael - Melide (CH)

Nelle cellule tumorali si riscontrano spesso deviazioni dal normale valore diploide dei cromosomi. Questi cambiamenti citogeneticiinteressano l’aploidia o la morfologia dei cromosomi oppure combinazioni di entrambe le alterazioni. Le monosomie o letrisomie e polisomie appartengono al primo gruppo mentredelezioni, duplicazioni, inversioni e traslocazioni appartengono al secondo gruppo. In più certe regioni cromosomiche possono risultare amplificate e mostrarsi come regioni colorate omogeneamente all’interno del braccio cromosomico o all’esterno di esso.
Copyright Fondazione Raphael - Melide (CH)

La specificità dei cambiamenti cromosomici è dimostrata dal fatto che solo 71 delle 329 bande cromosomiche del genoma umano sono interessate da riarrangiamenti tumore associati. Inoltre molti oncogeni cellulari che sono stati localizzati su specifici cromosomi localizzano anche break-point neoplasia associati.
Copyright Fondazione Raphael - Melide (CH)

L’analisi dei cromosomi di cellule tumorali ha evidenziato che solo il 15% presenta alterazioni del cariotipo. Questo dato però, apparentemente discrepante, trova una spiegazione nella tecnica di studio: infatti è difficile ottenere preparati di buona qualità a causa di problemi come l’indice mitotico basso e crescita sproporzionata, oppure perché il cariotipo mostra alti numeri modali con molti markers cromosomici bizzarri che probabilmente riflettono l’evoluzione del cariotipo ma parallelamente mascherano gli eventi specifici principali.
Copyright Fondazione Raphael - Melide (CH)

Al momento attuale due classi di geni sono coinvolte nello sviluppo e progressione del cancro: proto-oncogeni e tumor suppressors genes
Copyright Fondazione Raphael - Melide (CH)

I proto-oncogeni sono normali geni cellulari implicati nel controllo della crescita cellulare. Le mutazioni possono convertirli in oncogeni attivati sovrintendendo in moto patologico alla crescita delle cellule tumorali. Ad oggi sono stati individuati 70 proto-oncogeni, ed il loro bersaglio è localizzato in differenti compartimenti e funzioni della cellula quale il recettore per lacrescita cellulare, i fattori di crescita cellulare, i sistemi ditrasduzione intracellulari, i fattori di trascrizione e direttamente sulla proteine che sovrintendono il ciclo cellulare.
Copyright Fondazione Raphael - Melide (CH)

La mutazione che attiva il proto-oncogene è dovuta ad un aumento delle proteine del gene per imprevedibile espressione del gene edelezione del dominio regolatorio del gene oltre, come secondo meccanismo di trasformazione, vi è la transposizione pertranslocazione cromosomica. Vi è poi l’amplificazione genica che porta ad una sovraespressione degli oncogeni.
Copyright Fondazione Raphael - Melide (CH)

I tumor suppressor genes formano una categoria di geni di recente scoperta. La loro inattivazione per mutazione libera i meccanismi di crescita negativi da loro esercitati. Il primo risultato che si ottiene è una linea germinale (tumore ereditario) o una mutazione somatica(tumore sporadico) che inattiva uno dei due alleli del gene. Il secondo evento è una mutazione somatica che inattiva la restanteparte allelica: il cosiddetto allele selvaggio.
Copyright Fondazione Raphael - Melide (CH)

Questo è dovuto al fatto che la presenza dell’allele selvaggio nella regione cromosomica sede di mutazione provoca la sua eliminazione. Ciò avviene per non disgiunzione, mancataricombinazione mititoca o conversione genica.
Copyright Fondazione Raphael - Melide (CH)

Gli studi di genetica molecolare dimostrano come la sequenza adenoma-carcinoma nelle neoplasie colo-rettali sia caratterizzata da un sovrapporsi di eventi genetici distinti tra cui l’attivazione di oncogeni e la inattivazione di Tumor Suppressors Genes. Occorrono quindi come avviene in altri tumori, il verificarsi di lesioni genetiche multiple.
Copyright Fondazione Raphael - Melide (CH)

Per valutare correttamente e quantizzare la perdita di alleli, bisogna testare tutti i cromosomi con almeno un marker DNA, ottenendo una controparte molecolare chiamata “allelotipo”.
Copyright Fondazione Raphael - Melide (CH)

Quale parametro quantitativo della perdita multipla di alleli, è stato introdotto il termine di perdita allelica frazionala (F.A.L.) che rappresenta la parte evidenziabile di cromosoma con perditaallelica per singolo tumore.
Copyright Fondazione Raphael - Melide (CH)

La aneuploidia rappresenta il segnale tipico dei tumori solidi e nello stesso tempo la sua associazione clinico patologica della malattia. Questo sembra si a generato od accompagnato da instabilità delgenoma.
Copyright Fondazione Raphael - Melide (CH)

L’evoluzione genetica dei tumori solidi sottostà a delle forze che sono state recentemente messe a fuoco da un lavoro di Shackneyche ha sviluppato un modello computerizzato di simulazione temporale delle mutazioni che accompagnano la spontanea mutazione neoplastica. Il modello include la crescita cellulare, la divisione cellulare, la tetraploidizzazione, la perdita di cromosomi, lo sviluppo di aberrazioni strutturali cromosomiche.
Copyright Fondazione Raphael - Melide (CH)

Mentre la tetraploidizzazione non è il solo meccanismo messo in moto, la non disgiunzione mitotica può portare ad aneuploidia, rappresentando la via preferita di sviluppo di cloni ipodiploidi oiperdiploidi.
Copyright Fondazione Raphael - Melide (CH)

L’instabilità genomica certamente gioca un ruolo centrale nell’evoluzione clonale. Oltre all’aumento numerico dei cromosomi, la ipometilazione e le mutazioni di uno specifico gene coinvolto nella divisione cellulare sono le teorie a sostegno.
Copyright Fondazione Raphael - Melide (CH)

I proto-oncogeni attivati sarebbero i candidati migliori per la crescita dose-dipendente con anormalità cromosomiche per l’evoluzione di linee aneuploidi come avviene nel modello diShackney.
Copyright Fondazione Raphael - Melide (CH)

L’aumento di dosaggio del gene mutante, associato adaneuploidia, è stato identificato in una doppia e sequenziale non disgiunzione del cromosoma dell’oncogene attivato.
Copyright Fondazione Raphael - Melide (CH)

La destabilizzazione di normali cellule diploidi da parte di fattori genetici può sfociare in cloni near-diploidi o tetraploidi.
Copyright Fondazione Raphael - Melide (CH)

Entrambe queste vie possono sfociare in attivazione di oncogeni oinattivazione di Tumor Suppressors Genes. Le cellule così configurate possono crescere e moltiplicarsi come linea dominante.
Copyright Fondazione Raphael - Melide (CH)

Analogamente si è dimostrata una forte correlazione traaneuploidia ed inattivazione di Tumor Suppressors Genes.
Copyright Fondazione Raphael - Melide (CH)Copyright Fondazione Raphael - Melide (CH)