symbiotic nitrogen fixation and establishment of six montana
TRANSCRIPT

SYMBIOTIC NITROGEN FIXATION AND ESTABLISHMENT OF
SIX MONTANA NATIVE LEGUMES SPECIES
by
Sarah Jean Metcalf
A thesis submitted in partial fulfillment of the requirements for the degree
of
Masters of Science
in
Land Rehabilitation
MONTANA STATE UNIVERSITY
Bozeman, Montana
May 2005

© COPYRIGHT
By
Sarah Jean Metcalf
2005
All Rights Reserved

ii
APPROVAL
Of a thesis submitted by
Sarah Jean Metcalf
This thesis has been read by each member of the thesis committee and has been found to be satisfactory regarding content, English usage, format, citations, bibliographic style, and consistency and is ready for submission to the College of Graduate Studies.
Jeff Jacobsen
Approved for the Department of Land Resources and Environmental Sciences
Jon Wraith
Approved for the Department of College of Graduate Studies
Bruce McLeod

iii
STATEMENT OF PERMISSION TO USE
In presenting this thesis in partial fulfillment of the requirements for a master
degree at Montana State University-Bozeman, I agree that the Library shall make it
available to borrowers under the rules of the Library. If I have indicated my intention
to copyright this thesis by including a copyright notice page, copying is allowable only
for scholarly purposes, consistent with “fair use” as prescribed in the U.S. Copyright
Law. Requests for permission for extended quotation from or reproduction of this
thesis in whole or in part may be granted only by the copyright holder.
Sarah Metcalf
May 05, 2005

iv
ACKNOWLEDGEMENTS
First of all, I would like to acknowledge the Natural Resources Conservation
Service, the MSU Foundation Seed Program as well as the Department of Land
Resources and Environmental Sciences at MSU for funding this project. The
following people have helped to make this project a valuable experience for me. Jeff
Jacobsen helped me find this project; he has turned my life path in a whole new
direction, for which I will always be grateful. Clain Jones, my primary advisor,
worked very closely with me and was always around to help me solve problems. Tim
McDermott has been a wonderful mentor and provided me with opportunities too
numerous to list and Lina Butero-Griffiths, from his lab, who was very patient in
teaching protocols. Bill Grey has also given so much of his time and laboratory space
to help me with the pathogen aspects of this project. His enthusiasm for research and
discovery kept me going numerous times. I also want to thank Mark Majerus and
Larry Holzworth; both have given me great advice, encouragement and assistance over
the last two years. Dave Wichman and Bernie Schaff were appreciated for their
assistance in maintaining my field sites. I would also like to thank Aaron Mugaas,
Deirdre Willie, Ann McCauley, Teresa Warne, Ryan Weikel and Cole Germann for
their assistance in the laboratory and field. Special thanks to my family, especially my
parents who have been very supportive both financially and spiritually throughout this
project. Finally, I am thankful for my son Shea who inspired me to obtain this degree
though without whom completion would have been much easier.

v
TABLE OF CONTENTS
1. INTRODUCTION ...........................................................................................................1 Objectives ........................................................................................................................1 Background......................................................................................................................1 Materials and Methods.....................................................................................................1 Factors Effecting Nodulation and Legume Growth.........................................................4 Nodules ............................................................................................................................8 N2 Fixation in the Field....................................................................................................9 2. GREENHOUSE STUDIES: SOIL INOCULATION AND FUNGICIDE EFFECTS ON NODULATION AND BIOMASS.........................................................................12 Introduction....................................................................................................................12 Materials and Methods...................................................................................................12 Trial I: Effects of soil origin and N content on nodulation and biomass.......................12 Materials and Methods Trial 1...................................................................................13 Introduction................................................................................................................13 Seeding.......................................................................................................................15 Data Collection ..........................................................................................................15 Trial I Results..................................................................................................................16 Germination and emergence .....................................................................................16 Biomass.....................................................................................................................16 Nodulation.................................................................................................................19 Pathogenic Observations and Isolations ...................................................................20 Trial II: Effects of soil moisture content on nodulation and biomass..............................22 Materials and Methods..............................................................................................22 Introduction...............................................................................................................22 Seeding......................................................................................................................24 Data Collection .........................................................................................................24 Trial II Results .................................................................................................................24 Biomass.....................................................................................................................24 Nodulation.................................................................................................................25 Trial III: Effects of fungicide coating on nodulation, biomass and plant levels..............29 Materials and Methods..............................................................................................29 Introduction...............................................................................................................29 Seeding......................................................................................................................29 Data Collection .........................................................................................................30 Trial III Results ................................................................................................................30 Emergence and Plant Survival ..................................................................................30 Nodulation and Biomass ...........................................................................................32 Trial IV: Effects of fungicide drench on nodulation and biomass...................................36

vi
TABLE OF CONTENTS – Continued
Materials and Methods...........................................................................................36
Introduction............................................................................................................36 Seeding...................................................................................................................36
Data Collection ......................................................................................................37 Trail IV Results.............................................................................................................38 Biomass..................................................................................................................38 Root Structure, Nodules and Fungicide .................................................................39 N-content................................................................................................................41 Trial V: Effects of inoculation and fungicide application on nodulation and biomass ..............................................................................................................................42
Materials and Methods...........................................................................................42 Introduction............................................................................................................42 Seeding...................................................................................................................43
Trial V Results ..............................................................................................................44 Introduction............................................................................................................44 A. canadensis Combined Treatments....................................................................44
A. canadensis Nodulation ......................................................................................45 Inoculation .............................................................................................................46 Greenhouse Studies Summary ......................................................................................48 3. FIELD RESEARCH .....................................................................................................50 Introduction...........................................................................................................50 Materials and Methods..........................................................................................51 Site Preparation and Design..................................................................................51 Seeding..................................................................................................................52 Data Collection .....................................................................................................52 Field Research Results..................................................................................................55 Site Comparisons ..........................................................................................................55 Bridger ..........................................................................................................................56 Legumes...............................................................................................................56 Combined Biomass ..............................................................................................59 Grasses .................................................................................................................60 Summary ..............................................................................................................62 Moccasin .......................................................................................................................63 Legumes...............................................................................................................64 Combined Biomass ..............................................................................................65 Grasses .................................................................................................................66 Summary ..............................................................................................................67 Post Farm .....................................................................................................................67 Legumes...............................................................................................................68 Combined Biomass ..............................................................................................68 Grasses .................................................................................................................69

vii
TABLE OF CONTENTS – Continued
Summary ......................................................................................................................71 N Content .....................................................................................................................71 N2 fixation and Mass Balance......................................................................................76 Field Study Summary ..................................................................................................79 4. LEGUME LAND MANAGEMENT...........................................................................82 Introduction............................................................................................................82 Species Specific Information .................................................................................83
A. canadensis ....................................................................................................85 Dalea spp ...........................................................................................................86 H. boreale ..........................................................................................................87 L. argenteus........................................................................................................90 V. Americana......................................................................................................91
Technical Management Practices for Field Establishment of Native Legumes ....93 5. SUMMARY AND CONCLUSIONS ............................................................................99 REFERENCES CITED....................................................................................................102 APPENDICES .................................................................................................................111 APPENDIX A: PRELIMINARY GROWTH CHAMBER EXPERIMENTS ................112 APPENDIX B: INOCULATED AND NON-INOCULATED RESULTS FROM GREENHOUSE TRIAL V ..............................................................................................127

viii
LIST OF TABLES Table Page 1. Plant species seed stock and germination information ..................................................13 2. Field soil collection areas and characteristics................................................................14 3. Fungicide information and dilutions..............................................................................29 4. Rhizobia and legume inocula information .....................................................................43 5. Grass species and seed information ...............................................................................50 6. Field site information.....................................................................................................51 7. Native legume characteristics ........................................................................................84 A1. Six Legume and Ten Rhizobia Species.....................................................................102 A2. Plant survival in pouches AI .....................................................................................114 A3. Treatments and surviving plants in Trial AIII ..........................................................117 A4. Analysis of plant tissues and nutrient solution from growth pouches ......................121

ix
LIST OF FIGURES Figure Page 1. Emergence of all six native legumes in Geyser soil ......................................................17 2. Emergence of all six native legumes in Red Bluff soil..................................................18 3. Biomass from Trial I in Geyser and Red Bluff soil .......................................................18 4. Nodule mass from Trial I in Geyser and Red Bluff soil ................................................21 5. Nodule number from Trial I in Geyser and Red Bluff soil............................................22 6. Biomass from Trial II in Bridger and Post Farm soil ....................................................25 7. Biomass from Trial II in Bridger soil 60-80% FC soil moisture ...................................26 8. Biomass from Trial II in Post Farm soil 60-80% FC soil moisture ...............................26 9. Nodule mass from Trial II in Bridger soil 60-80% FC soil moisture ............................27 10. Nodule mass in Post Farm soil 60-80% FC soil moisture ...........................................28 11. Emergence of D. candida with fungicide seed coats in Geyser and Red Bluff soil ....31 12. Final plant number of D. candida with fungicide seed coats in Geyser and Red Bluff soil ............................................................................................................................31 13. Nodule mass of D. candida with fungicide seed coats in Geyser and Red Bluff soil .33 14. Photo-Nodules from D. candida metalaxyl vs. no fungicide.....................................34 15. Nodule number of D. candida with fungicide seed coats in Geyser and Red Bluff soil......................................................................................................................................34 16. Biomass of D. candida with fungicide seed coats in Geyser and Red Bluff soil ..............................................................................................................35 17. First harvest biomass of A. canadensis and L. argenteus with fungicide drench ........39 18. Second harvest biomass of A. canadensis and L. argenteus with fungicide drench....40 19. Photo- L. argenteus root condition when grown in Post Farm soil, fungicide vs. no fungicide treatment ............................................................................................................40

x
LIST OF FIGURES-Continued Figure Page 20. Second harvest N concentration of biomass from A. canadensisand L. argenteus with fungicide drench ........................................................................................................42 21. Biomass from A. canadensis with fungicide drench and inoculation..........................45 22. Nodule number of A. canadensis with fungicide drench and inoculation ...................46 23. Nodule mass from A. canadensis with fungicide drench and inoculation...................47 24. Total N concentration of A. canadensis with fungicide drench and inoculation .........48 27. Field emergence of legumes in 2003 season................................................................57 28. Combined grasses and legume biomass at all three field sites ....................................58 29. Bridger site: legume dry shoot biomass.......................................................................60 30. Bridger site: combined grass and legume dry shoot biomass ......................................61 31. Bridger site: grasses dry shoot biomass .......................................................................61 32. Photo: Moccasin site: fall 2003 defoliation by grasshoppers, Spring 2004 site regrowth .............................................................................................................................63 33. Moccasin site: legume dry shoot biomass ...................................................................65 34. Moccasin site: combined grasses and legume dry shoot biomass ...............................66 35. Moccasin site: grasses dry shoot biomass....................................................................67 36. Post Farm site: legume dry shoot biomass...................................................................70 37. Post Farm site: combined grasses and legume dry shoot biomass...............................70 38. Post Farm site: grasses dry shoot biomass...................................................................71 39. N concentration of legume shoot biomass at all three field sites.................................74 40. Average shoot tissue N content at all three field sites .................................................75 41. Average shoot tissue N content of legumes vs. grasses at all three field sites ...........75

xi
LIST OF FIGURES-Continued Figure Page 42. Total N tissue mass balance for all three field sites.....................................................77 43. Bridger site: nitrogen mass balance .............................................................................78 44. Moccasin site: nitrogen mass balance..........................................................................78 45. Post Farm site: nitrogen mass balance .........................................................................79 46. Photo: A. canadensis at Post Farm site summer 2004. Seedpod and flowering ..........85 47. Photo: D. purpurea and D. candida H. boreale from Post Farm field site. Flowering and seedpod ......................................................................................................87 48. Photo: H. boreale from the Post Farm summer of 2004..............................................88 49. Photo: V. americana from the Post Farm field sit .......................................................90 50. Photo: L. argenteus at Post Farm site. Seedpods, root nodules and full row..............92 A1. Photo: native legumes vs. soybeans in growth chamber Trial AIII ..........................122 A2. Legume dry biomass from growth chamber Trial AIII............................................123 A3. Legume NO3-N from pouch solution in the growth chamber Trial AIII ..................124 A4. Legume NH4-N dry from pouch solution in growth chamber Trial AIII..................125 A5. Total N content of legumes biomass from growth chamber Trial AIII ....................125 A6. Total N per legume species from growth chamber Trial AIII ..................................126 B1. Trial V: biomass from Dalea spp. and H. boreale with fungicide drench and inoculation.....................................................................................................129 B2. Trial V: nodule mass from Dalea spp. and H. boreale with fungicide drench and inoculation.....................................................................................................130

xii
ABSTRACT
Legume species have been known to increase soil N content and are incorporated into land restoration seed mixes in hopes of improving degraded soils and plant communities. The goals of this project were to determine effects of soil type, soil moisture, inocula and fungicide treatments on legume establishment, N2 fixation capacity and plant biomass for six individual native legume species. In potted greenhouse studies, legumes were grown in five field soils to verify nodulation without inocula. In a second phase, plant biomass was measured in two soils at two moisture contents (60 and 80% of field capacity). The efficacies of Rhizobia inocula and fungicide treatments were also tested. The greenhouse studies were used to supplement data gathered at three field sites around Montana. Native soils contained Rhizobia that were specific to many of the plant species tested and induced nodulation. Soil moisture content, inoculation and fungicide treatments had less effect on nodulation and biomass than the soil characteristics. Fungicide treatments benefited establishment of Lupinus argenteus, and had mixed effects on Astragalus canadensis and Dalea candida. Benefits were dependent on soil nutrient content, microbial biota and other characteristics. Shoot biomass at each field site also varied significantly; in general A. canadensis had the highest nodulation and biomass, while Dalea purpurea and D. candida typically had the lowest biomasses and nodulation. This research should assist land managers in selecting species to suit a wide variety of ecological conditions and land restoration scenarios.

1
CHAPTER 1
INTRODUCTION
Objectives
The purpose of this project was to determine the amount of nitrogen gas (N2)
that can be fixed each by of six native legume species. Establishment and biomass
production of these species, in different field soils, was also examined. Specifically,
the goals of the project were to determine the effect of different field soils on
nodulation, total biomass production and N levels. Soil moisture content was
examined in relation to effects on nodulation, total biomass production and N content
for the selected legumes in two soils. The extent of nodulation, total yield, total N
uptake, and changes in available soil N (NO3-N + NH4-N) in different field soils for
the selected legume species was determined. Finally the effect of seed fungicide
treatments and inoculation on emergence and plant biomass was examined.
Background
Natural and disturbed ecosystems are often nitrogen (N) limited. Leguminous
plants have the potential to increase the levels of available nitrogen (N) in soils
(Seastedt and Knapp 1993; Towne and Knapp 1996; Tlusty et al. 2004; Unkovich and
Pate 2000). This project evaluated six native legume species that have been released
or are under consideration for release by the Natural Resources Conservation Service
(NRCS) Plant Materials Program to be used as additions to current prairie restoration

2
seed mixes. It attempted to determine the ability of each species to fix nitrogen gas
(N2) and measure the change in levels of bioavailable N in field soils. Another goal
was to establish field guidelines for the use of native legumes in land restoration,
because very little is known about the physiology and ecology of Montana native
legumes.
Many factors affect a grass-legume system and there are many levels on which
grasses and legumes compete and benefit one another in a prairie system, but these
systems can be difficult to establish and maintain (Frame and Newbould 1984, 1986;
Ross et. al. 1972; Thornley et al. 1995; Schwinning and Parsons 1996a, b, 1999).
Developing field protocols will hopefully help researchers and land managers achieve
more successful land restoration and lasting plant community establishment. These
protocols can also serve to improve information about species selection and
management in order to best utilize the N2 fixation potential of these species based on
growth habit and ecological niches. This research seeks to improve the knowledge
base of legume systems in order to determine and eliminate some barriers to successful
plant establishment. Current barriers are drought and salinity sensitivity, soil borne
pathogens, pests and a lack of knowledge on native species-specific rhizobia and the
quality of inocula currently in use (Schwinning and Parsons 1996 a, b, 1999; Thornley
et al. 1995; McDermott et al. 1991; Tlusty et al. 2004).
Humans have used legumes in agriculture for thousands of years either as a
food crop for humans and livestock, or as a rotational crop to supply N to non-legume
crops. The shift from biological to industrial sources of N fertilizer in agricultural
systems did not occur until the 20th century (Smil 2001; Crews and Peoples 2004). In

3
more recent times, legumes have been incorporated back into rotational farming,
especially in organic farming systems (Crews and Peoples 2004, Stockdale et al. 2000;
Smil 2001). It has been demonstrated that symbiotic N2 fixation enhances soil fertility
and productivity as well as increases carbon sequestration and nutrient conservation
(Cromwell and Winpenny 1993; Snapp et al. 2002). Some less developed nations with
low fertility soils cannot afford synthetic fertilizers and have never stopped the practice
of utilizing symbiotic N2 fixation (Crews and Peoples 2004; Snapp et al. 2002).
Symbiotic N2 fixation can actually be so effective that some researchers have noted
that many of the same problems that arise with poor usage of commercial fertilizers
can arise with improper management of legume crops (Scott et al. 2000; William and
Donald 1957). For example, legume crops can lead to soil acidification, N leaching
and excess N runoff in surface waters at levels just as high and detrimental to human
health and the environment as synthetic fertilizers (Fillery 2001; Heltar and Porter
1989; Peoples and Baldock 2001; Ridley et al. 2004; Scott et al. 2000). Ridley et al.
(2004) also found that N build up from legumes often exceeds 100 kg N ha-1 and with
high moisture levels, losses of N through leaching can be 15-35 kg N ha-1 yr-1. It is
unlikely however, in semi-arid regions utilizing native legume species, that leaching of
N build up would be a problem.
Native legume effects on yield of grasses and other crops, nodulation N2
fixation potential and soil fertility are highly variable (Crews and Peoples 2004; Jha et
al. 1995; Zahran 1998; 2001). Depending on the environmental conditions and the
species selected, legumes can be a great source of natural N, although there is always a
need for good management practices adapted to the region and species of interest.

4
Integration, rotation and/or incorporation of legumes as green manure, are currently
being proposed as best management practices by many ecologists and agronomists in
the field (Ridley et al. 2004). There is extensive literature on the subject of N2 fixation
in legumes and their role as an N source for non-legumes; however, very little is
known or published on legumes native to the U.S., and specifically, Montana.
Factors Affecting Nodulation and Legume Growth
The extent of nodulation and amount of N2 fixed by legumes is largely related
to plant size and root mass and will vary depending on the species of plant and the
strain of rhizobia which it hosts (Abel and Erdman 1985; Berg et al. 1988; Ham et al.
1971a; McDermott et al. 1991; Provorov et al. 2001; Tlusty et al. 2004; ). Individual
rates of N2 fixation will vary according to the plant’s growth stage and its demands for
N at a given time.
N2 fixation is an energetically expensive process costing the plant 16 ATP for
every N2 molecule reduced (Bergersen 1982). The energetic expense of the symbiosis
can be detrimental both to the plant host, due to loss of photosynthates, and to the
rhizobia, which could otherwise use carbon (C) sources for growth and reproduction
(Denison and Kiers 2004b). Rhizobial strains can be symbiotic, non-symbiotic, unable
to infect a host, or even parasitic, nodulating the host plant but fixing little or no N2
(Denison and Kiers 2004b). Many strains of rhizobia, utilizing any of these strategies,
can be present in soil at the same time (Berg et al. 1988; McDermott et al. 1991;
Moawad and Schmidt 1987). The strains in this study and within a soil can belong to
species of Rhizobium and Bradyrhizobium and will be referred to as rhizobia

5
throughout this paper (Graham et al. 1999; Tlusty et al. 2004; Zakhia and de Lajudie
2001). A variety of molecular and genetic mechanisms have evolved between the
legume and the rhizobia to regulate the nodulation and N2 fixation processes (Lum and
Hirsch 2003; Provorov et al. 2002). For example, the host plant regulates the oxygen
(O2) supply to the interior of the nodule and has the ability to cut off the O2 supply to
nodules that are not fixing N2, allowing the plant to select for beneficial rhizobia
(Denison and Kiers 2004a).
Soil nutrient availability is a primary factor driving the occurrence and function
of this symbiosis. If there is adequate N for plant growth, the symbiosis will be less
likely to occur (Lum and Hirsch 2003; Streeter 1988; Unkovich and Pate 2000; Vance
2001). However, when bioavailable N is limited, the costs are outweighed by the
benefits, and symbiosis will be initiated. With low N levels, a biochemical dialogue is
initiated between the host plant and the rhizobia, causing nodulation to occur (Caetano-
Anolleseld 1988; Lum and Hirsch 2003). Phosphorus (P) is another key nutrient for
effective symbiosis; if P is limited, symbiosis will often fail (Holford 1997; Lum and
Hirsch 2003; Marschner 1995; Vance 2001). The importance of P in symbiotic N2
fixation is indicated by an accumulation of P in nodules, compared to other organs,
during P-limited conditions (Israel 1993; Schulze 2004). There is evidence suggesting
that some legumes may employ a dual symbiosis by utilizing arbuscular mycorrhizae
to facilitate the acquisition of P in low nutrient soils (Lum and Hirsch 2003; Provorov
et al. 2002; Requena et al. 2001; Zahran 2001).
Light levels and increased photosynthesis will also increase N2 fixation as
photosynthates provide energy to fuel the reaction and increase plant biomass, as well

6
as the demand for N (Crews and Peoples 2004; Unkovich and Pate 2000). Nodulation
is also affected by temperature and pH due to influences on the rhizobia survival
(Bushby 1990). Maintaining temperature and pH in optimal ranges will directly affect
the extent of nodulation in the host plant; the range of tolerance will vary according to
the strain and its origin of adaptation (Bushby 1990; Bergersen 1982; Jacot et al. 1999;
b). High or low pH can damage rhizobia as well as root hairs and hinder the infection
processes. After infection has occurred, N2 fixation will continue as long as the host
plant can tolerate the pH level (Bergersen 1982).
It has also been found that temperature and pH tolerance ranges often
correspond to the environmental conditions in which the plant-rhizobia relationship
evolved. For example, native and high altitude species such as lupine or prairie clovers
are better adapted to the extreme environmental conditions that exist at altitude and
therefore perform better at lower temperatures as compared to tropical legumes such as
soybeans (Graham 2004; Jacot et al. 1999; Kessler et al. 1990; Svenning et al. 1991).
Plants inoculated with rhizobia species adapted to high altitude have shown higher
tolerance to stress conditions such as low nutrients, extreme pH, low moisture and cold
temperatures (Jacot et al. 1999; Johnson and Rumbaugh 1986; Svenning et al. 1991).
Typically, these stresses limit root infection by reducing bacterial populations in the
root zone and therefore decreasing the ability of the rhizobia to initiate nodulation
(Kessler et al. 1990).
Soil moisture is important to rhizobia survival. Continuous water films in the
soil benefit membrane structure and function of free-living bacteria and aid in their
distribution and movement through the soil (Bergersen 1982). Infection occurs when a

7
root hair grows through a resident population of free-living bacteria. In low moisture
conditions, this infection process may be inhibited. Low soil water content has been
seen to reduce nodulation by as much as 50% as measured by acetylene reduction and
respiration rates (Sinclair et al. 1987). If the moisture content of the nodule drops
below 80% by weight, the nodules begin to suffer from water stress causing respiration
and N2 fixation to decrease (Abdel-Wahab et al. 2002). Continued water stress can
cause irreversible damage to nodules and bacteria, permanently stopping N2 fixation,
until new nodules can be formed (Abdel-Wahab et al. 2002). High soil moisture or
saturation can lead to anaerobic conditions and will also stop N2 fixation because
rhizobia are microaerophilic and need some O2 for respiration (Dension and Kiers
2004b; Yu and Rengel 1999). High moisture is not as detrimental as low moisture and
the organisms suffer no physiological damage; when moisture levels drop, N2 fixation
will recommence (Bergersen 1982). Symbiotic processes are more sensitive to
saturation in some legumes, such as M. sativa, than they are in tropical legumes, which
show little response (Ahmed and Quilt 1980; Ismail et al. 1983; Williams and
DeMallorca 1984).
Other stresses such as heavy metal toxicity (Broos et al. 2004), high salinity
and nutrient deficiency can also stop or hinder the infection process or N2 fixation
(Saxena 1991; Yu and Rengel 1999). Legumes with high tolerance to water and other
stresses have been found to have a strong ability to make osmotic adjustments (Ford
1984). Legumes that form indeterminate nodules may be better able to tolerate stresses
than legumes with determinate nodules. The indeterminate nodule has the ability to
continue growing and forming new tissue from the indeterminate end, while the

8
determinate type will have to reform new nodules in order to continue N2 fixation
(Abdel-Wahab et al. 2002). Nodule morphology has less affect on a legume’s ability
to tolerate environmental stresses that affect rhizobial survival in the soil, such as
heavy metal contamination or other toxins (Broos et al. 2004).
Nodules
The extent of nodulation is related to the combination of the legume species
and the strain or strains of rhizobia that infect it (Denison and Kiers 2004b). It is also
related to the plant’s physiology and ability to form lateral roots. Infection occurs at
the root tip where cortex cells are rapidly dividing and growing outward (Hill 1980).
No published literature has been located showing a firm correlation between nodule
size, number and potential to fix N2 in an individual plant. Hill (1980) suggested that
an inhibitory substance produced by the apex of nodules may prevent other nodules
from forming in the same region of the root. Furthermore, it is thought that larger
functioning nodules may reduce the number of nodules on the nearby root, while
ineffective nodules will not produce the inhibitory substance leading to clusters of
small nodules on regions of the root (Hill 1980).
An indicator of nodule activity is the presence of leghemoglobin (LHb), an iron
containing protein similar to human hemoglobin, which binds O2 (Becana et al. 1986;
Bergersen 1982). It facilitates O2 diffusion throughout the interior of the nodule. N2
fixation is a reduction reaction and is hindered by the presence of O2; however, the
bacteroids require O2 to maintain metabolic function (Becana et al. 1986). While
conducting studies on drought and high salinity, Abdel-Wahab et al. (2001) found LHb

9
to be indicative of N2 fixation activity, as decreases in this protein within the nodule
were correlated with decreases in acetylene reduction assays (Guerin et al. 1991;
Irigoyen et al. 1992; Nash and Schulman 1976).
N2 Fixation in the Field
The primary product of N2 fixation is NH3, which quickly complexes with H+
to form the ammonium ion (NH4+). The NH4
+ can become toxic at high levels and is
exported to the cytosol and is immediately incorporated into amino acids (Abdel-
Wahab et al. 2002). These amino acids are transported throughout the plant to be
incorporated into the tissue or used in metabolic processes; excess amino acids are also
excreted into the surrounding soil by plant roots. Field studies have shown that some
legumes take up less N from the soil after nodulation (Armstrong et al. 1994; Phillips
and DeJong 1984; Unkovich and Pate 2000). Some of the N2 fixed is excreted into the
surrounding soil as amino acids and becomes available to non-legume species (Crews
and Peoples 2004). In addition, nodules have a short life span and as they senesce,
available N is released back into the soil and becomes bioavailable or is absorbed by
the host plant (Crews and Peoples 2004). The main pathway for N transfer from
legumes to adjacent crops occurs through the decomposition of root material rather
than through root exudates or direct transfer from mycorrhizal associations (Crews and
Peoples 2004; Trannin et al. 2000).
Many studies have found that intercropping of a legume species with grasses or
other non-leguminous species can have a significant influence on biomass of the non-
legume plant (Haynes 1980; Ledgard 1991). Sengul (2003) found that incorporating

10
sainfoin or M. sativa with three grasses increased the biomass and N content of the
grasses grown in conjunction with the legumes. Grasses grown in conjunction with
sainfoin had the largest increase in biomass, N and crude protein content; however,
both intercropped groups had significantly higher biomasses than did any of the single
stands of legumes or grasses (Gökkus et al. 1999; Sengul 2003). Furthermore,
mixtures enhanced utilization of symbiotically fixed N2 as well as increasing allopathic
effects and light absorption (Pubman and Duke 1978).
Ridley et al. (2004) contend that farming systems that rely on legumes as the
sole N supply are unsustainable because of competition for nutrients and water, as well
as poor timing between the supply and demand for N between the crops. Thornley
(2001) proposed an adaptation of the Hurley Pasture Model, which is a single-species
grassland simulator. This new model for management attempts to optimize not only N
uptake by non-legume crops, but also utilizes the benefits that legumes offer in C
sequestration (Thornley 1998). This model incorporates factors of competition that
occur for grass-legume systems and attempts to aid land managers in species selection
and crop management to successfully utilize legumes as a green manure (Schwinning
and Parsons 1996a;b; 1999; Thornley et al. 1995; 2001). Ridley et al. (2004) also
contend that the benefits of legumes can be substantial when crops are properly
managed. Thornley (2001) contends that the ideal approach to harnessing the benefits
of legumes is to utilize a polyculture system. With multiple legume and grass species
there is a variety of spatial and temporal zones for nutrient and water uptake as well as
varying rates of uptake and demand for these necessities (Crews and Peoples 2004;

11
Ridley et al. 2004). The variety of legumes and their N2 fixation efficiencies as well as
species and rhizobia combinations requires further investigation.
This study looks at potential N2 fixation, establishment, individual growth
requirements and potential benefits of the selected native legume species in Montana
soils. The work was organized into greenhouse studies (Chapter 2), field research
(Chapter 3) and management implications (Chapter 4).

12
CHAPTER 2
GREENHOUSE STUDIES: SOIL INOCULATION AND FUNGICIDE EFFECTS
ON NODULATION AND BIOMASS
Background
The following greenhouse studies were designed to provide information on the
effects of soil type, moisture content, and inoculation and fungicide treatments on
seedling emergence, nodulation and biomass. In addition, data on root biomass and the
extent of nodulation were collected. These data were compared to total plant biomass
and later used to supplement the limited field data of belowground plant biomass,
which is difficult to sample. The greenhouse portion of the research consisted of five
separate trials, each targeted to test different characteristics.
Materials and Methods
Seed stock information for each species is listed in Table 1. All six species
were obtained from the Bridger Plant Materials Center (BPMC) in Bridger, Montana.
The soils used for the greenhouse studies were collected from five field sites around
Montana and the soil characteristics are listed in Table 2.

13
Materials and Methods Trial I Introduction
The overall objective of this trial was to determine the effect of soil N and
characteristics on nodulation and total biomass in all six plant species. Specifically,
this experiment tested the effects of available N levels and other factors on nodulation
and biomass in soils from Geyser, MT near Great Falls and Red Bluff Agricultural
Research Station near Norris, MT (Table 2).
Table 1. Plant species seed stock and germination information.
Plant Species Common Name Pure Seed Germination Total Viability PLS*
------------------------------------%------------------------------ Astragalus canadensis
(AC) Canada Milk Vetch 94 6 88 82
Dalea candida (DC)
Slender White Prairie Clover
98 77 99 97
Dalea purpurea (DP)
Purple Prairie Clover 100 75 91 91
Hedysarum boreale (HB)
Northern Sweet Vetch 98 51 51 50
Lupinus argenteus (LA)
Silvery Lupine 100 0 82 82
Medicago sativa (MS)
M. sativa- Excellent 10A
98 93 94 94
Vicia americana (VA)
American Vetch 100 63 87 87
*PLS- pure live seed

14
Geyser was an active grass-M. sativa pasture that had been treated with N
fertilizer leading to a high N content that was 49 mg kg-1 higher than at Red Bluff. Red
Bluff is a native rangeland soil that had never been fertilized or cultivated, aside from
periodic grazing by livestock and wildlife. L. argenteus was identified growing in the
vicinity of the soil collection area. The presence of native legumes on the site was a
good indication that the field soil contained indigenous rhizobia. All six native prairie
legumes species from Table 1 were selected. No rhizobia inocula were applied in this
trial; therefore any nodulation was the result of the indigenous rhizobia populations in
the soils.
Table 2. Field soil collection areas and characteristics.
Site Cropping History
Texture FC* EC** pH NH4-N NO3-N Olsen P K
g H2O g soil-1 dS m-1 ----------------------mg kg-1------------------
Geyser Agriculture Clay Loam
ND*** 0.45 8.0 ND 76.1 7.9 366
Red Bluff Native Range
Sandy Loam
ND 0.19 7.0 ND 27.1 8.4 450
Bridger Agricultural Research
Silty Clay Loam
0.19 0.15 8.6 3.6 20.1 32.7 564
Moccasin Agricultural Research
Silt Loam
0.23 0.13 7.3 3.4 4.1 32.8 410
Post Farm Agricultural Research
Clay Loam
0.27 0.26 8.0 3.7 48.7 61.1 508
*FC- field capacity (_Pa) **EC- electrical conductivity ***- not determined

15
Seeding
Rocks and large gravel were removed from each soil. The six species, two
soils, and three replicates led to thirty-six pots (20 cm diameter), which were washed,
dried and then filled with approximately 1.2 kg of air-dried homogenized soil. Twenty
pure live seed (PLS), untreated and unconditioned, of each species were planted 1 cm
deep in three pots of each soil type. Each pot was then watered with 50 mL of water
and placed in the greenhouse at 20-26 ºC, the optimal temperature for the Dalea spp.
seeds to germinate (McGraw et al. 2003). Sixty mL of water were applied every other
day for the first two weeks after which time watering was reduced to every 4 d with the
same volume. At 45 d the plants in each pot were thinned to 4 plants per pot. Plants
were allowed to grow for a five-month period in the greenhouse.
Data Collection
Emergence was recorded daily for 14 d beginning 5 d after planting, then
weekly for the next 1.5 months. At the initiation of flowering, plants and soil were
removed from the pots by hand, and the roots and nodules were separated from the soil
by washing through a 1 mm screen sieve with cool tap water. All live biomass was
removed from the sieve and the entire plant was washed clean and stored at 2 ºC in
airtight plastic bags.
The total wet biomass of each plant was weighed. Aboveground biomass was
separated from the belowground biomass, and each wet component was weighed
separately. The aboveground material was placed in a drying oven at 40 ˚C for 14 d;
the dry biomass was then removed and weighed. The root nodules were separated

16
from the root system using forceps and then counted and weighed. Statistical analysis
of all data was assessed using JMP 5.1 (SAS Institute Inc. Cary, NC) analysis of
variance (ANOVA).
Results and Discussion Trial I
Germination and emergence
Native species adapted to semi-arid regions often have a slow growth rate and
the seed has a high longevity in the seed bank (McGraw et al. 2003). Germination
rates in natives are often much lower than non-native species adapted to less harsh
environments. In this experiment, D. purpurea had the highest emergence rate,
followed by D. candida (Figure 1-2). The majority of viable Dalea spp. seeds
germinated within two weeks after planting. The other four species were slower to
germinate and the number of seeds germinating continued to increase for four weeks.
A. canadensis and L. argenteus had the lowest germination rates although the A.
canadensis showed more of an increase later in the study, particularly in the Geyser
soil. L. argenteus consistently had a low germination rate through most of the trials in
this study.
Biomass Two of the native species, A. canadensis and L. argenteus, had significantly
higher dry shoot biomasses in Red Bluff soil than in Geyser soil, whereas the other
four species were not significantly different between soils (Figure 3). A. canadensis
had significantly higher biomass than the other species when grown in the Red Bluff
soil, although its biomass in the Geyser soil showed less variation from the other

17
species. L. argenteus had a moderate biomass in the Red Bluff soil, but in the Geyser
soil the plants began to show symptoms of root and stem rot around the second month
of the trial and by the end of the study all of the plants had died. The Dalea spp, H.
boreale and V. americana biomasses did not differ significantly between the soils.
The Dalea spp. had the smallest biomass in both soils, not surprisingly since it was
very slow growing and had a slender morphology. Apparently, soil characteristics,
specifically nutrient content and endemic microbial communities, have major
influences on the growth responses of at least some of these species.
Figure 1. Plant emergence for six native legume species, grown in Geyser soil DAS in 2002. (DAS)=days after seeding A. canadensis (AC), D. candida (DC), D. purpurea (DP), H. boreale (HB), L. argenteus (LA), V. americana (VA).
0%
5%
10%
15%
20%
25%
30%
35%
40%
45%
0 7 14 21 28 35 42
DAS
Emer
genc
e R
ate
ACDCDPHBLAVA

18
Figure 3. Average total dry shoot biomass of the six species in Geyser and Red Bluff soils.
ed e
c db c
b
e e
cb c
b
a
0
1
2
3
4
5
6
A C D C D P H B L A V A
S p e c ie s
Dry
Sho
ot B
iom
ass
(g p
ot -1
)
G e y s e rR e d B lu ff
L S D = 0 . 6
Figure 2. Plant emergence rate for six native legume species, grown in Red Bluff soil DAS in 2002.
0%5%
10%15%20%25%30%35%40%45%
0 7 14 21 28 35 42DAS
Emer
genc
e R
ate
ACDC
DP
HB
LA
VA

19
Nodulation Soil nutrient and texture characteristics had less of an effect on plant biomass
than on nodulation and most likely N2-fixation. There was much higher nodulation in
the Red Bluff soil than in the Geyser soil. All the nodules collected had pink
pigmentation, indicating the presence of LHb, a sign that N2 fixation is probably taking
place (Abdel-Wahab et al. 2002). Many studies find a good correlation between
nodule parameters and plant production (Tlusty et al. 2004). Fresh weight may be a
better indicator of nodule functionality compared to nodule number, as active nodules
are typically heavier (Hill 1980). For example, H. boreale had a similar number of
nodules in both soils; however, the nodule biomass was much larger in Red Bluff as
was the total biomass, indicating perhaps a difference in N2 fixation amounts (Figure
4-5). However, H. boreale and L. argenteus both had much higher average nodule
weights than the other species (unpublished data). Both species are nodulated by
Bradyrhizobium, and express different nodule morphology from the other species as
well as one another.
Not only were there lower levels of nodulation in the Geyser soil, but two
species, L. argenteus and V. americana, did not form nodules at all in soil from this
site. The reduction and lack of nodulation in the Geyser soil is probably due in part to
the high N level of the soil, which is known to suppress nodulation (Schulze 2004).
This soil also had very low P content; the lack of P can drastically reduce the level of
nodulation and N2 fixation and may be a factor in the lower nodule mass in this soil
(Lum and Hirsch 2003). All the other legume species did nodulate, so it is unlikely
that the lack of nodulation in the L. argenteus and V. americana is due to soil

20
characteristics such as nutrients or pH, but rather has more to do with rhizobial
populations in that soil. There were no native legume species observed at the soil
collection site, which may be indicative of a lack of adequate species-specific rhizobia
in this soil. The Red Bluff site had native legumes present and showed much higher
nodulation in all species. Dalea spp., H. boreale and V. americana all had very low
nodulation in the Geyser soil compared to the Red Bluff soil, yet they had similar total
biomasses in both soils (Figure 3-4). This response raises the question of whether or
not the additional N provided by N2 fixation actually benefits biomass production, or
does it actually hinder it due to the additional energetic expense of the symbiosis? It
would serve to reason that biomass production would be slightly inhibited because a
portion of the energy that the plant produces would be directed toward the symbiosis,
whereas passive uptake of nutrients through the root system has a much lower
energetic cost. However, if a longer-term study had been conducted, the more
nodulated plants in the Red Bluff soil may have eventually out-competed the plants in
the Geyser soil, especially after the available soil N was depleted.
Pathogenic Observations and Isolations
The L. argenteus germinated in the Geyser soil, but the plants died during the
third month of the study, showing symptoms of disease from root and stem rot shortly
after their secondary leaves emerged. There was no sign of nodulation in this species
at any stage of plant growth in the Geyser soil. Because of the high N content in this
soil, it is unlikely that the lack of nodules was the cause of the plants’ deaths. It does
indicate, however, that the Geyser soil is probably lacking the species-specific rhizobia

21
required to establish effective symbiosis. Nodulation of the other five species was
verified within the first month of the trial. The diseased L. argenteus tissue was
isolated and several species of Fusarium spp. fungus were isolated. Although they
were not identified to the species level, this family of fungus is known to be a major
fungal pathogen leading to stem and root decay and can eventually kill the plant.
Schulze (2004) examines many mechanisms within the plant genome, as well
as the rhizobia, that regulate the symbiosis in response to physiological and
environmental stresses. Indigenous strains of rhizobia can have a large effect on plant
productivity.
Figure 4. Average dry nodule mass of all six species in Geyser and Red Bluff soils. No nodules were detected on L. argenteus and V. americana.
LS D -0 .05
e ee
cdc
decd
a
b
b
0.00
0.05
0 .10
0 .15
0 .20
0 .25
0 .30
A C D C D P HB LA V A
S p e c ie s
Nod
ule
Mas
s (g
pot
-1)
GeyserRed B luff

22
Trial II: Effects of soil moisture content on nodulation and biomass.
Materials and Methods
Introduction
In this experiment, the primary objective was to determine the effect of soil
moisture content on nodulation and total biomass for two legume species in two
different field soils. Two agricultural soils were selected from the Bridger Plant
Materials Center (BMPC) and the A.H. Post Farm, (Table 2). The four treatment
groups consisted of each of the six species grown in the two soils at moisture levels of
approximately 60% and 80% of each soil’s field capacity (FC). The soils were
submitted to the MSU Soil, Plant and Water Analytical Laboratory (SPWAL) to
measure the water holding capacity of each soil using a pressure plate at -33 kPa to
LSD-69
de ed
bc cdcd
b
a
d cd
0
100
200
300
400
500
600
AC DC DP HB LA VA
Species
Nod
ule
Den
sity
(No.
pot
-1
)
GeyserRed Bluff
Figure 5. Average nodule number from greenhouse Trial I in Geyser and Red Bluff soils.

23
approximate FC (Table 2). These values were then used to obtain the weights of each
pot at 60% and 80% of FC depending on the soil.
Seeding
Seeding followed the same protocol as listed in Trial I with the following
exceptions. An additional replica was added leading to four replicas for each
treatment. Each individual pot was weighed to make sure it was filled with exactly 1.2
kg of soil, and then water was added to bring the soil moisture to the FC of each soil,
by weight. Each pot was watered every other day to maintain FC moisture for 14 d to
allow the seedlings to properly germinate. At day 15, watering was reduced to keep
the treatments near 60% and 80% of FC by weight. Each pot was weighed every 3 d
and the appropriate volume of water was added to maintain the moisture level of each
treatment group. The plants were allowed to grow in the greenhouse for five months
during which time the soil moisture was kept between 55-65% or 75-85% FC.
Data collection
Harvest and data collection followed the same protocol as in Trial I. The data
was graphed in two different ways to better show differences between the moisture
treatments and the soil origins.
Trial II Results
Biomass
There was a high degree of variation between the species biomass between both
soil type and soil moisture treatments (Figures 6-8). There was more variation in

24
biomasses when the species were compared by soil origin (Figure 6) rather than by the
moisture content treatments; however, significant variation within species still existed
(Figure 7-8). All species but D. candida and L. argenteus had higher biomasses in the
Post Farm soil than the plants grown in the Bridger soil (Figure 6). The slower growth
in the Bridger soil could be due to several characteristics of that soil. The texture and
shrink-swell characteristics of the Bridger soil led to structure and drainage problems
in the greenhouse pots. By the end of the study much of the silt had washed out of the
pots leaving high clay content in the Bridger pots. This could have affected nutrient
uptake and benefited fungal pathogens. This soil also had a higher pH than the Post
Farm soil, which could have affected plant growth and rhizobial populations, although
endemic populations would probably be adapted.
There was some plant biomass variation within species at the different soil
moisture levels (Figure 7). Two of the six species, H. boreale and L. argenteus, had
significantly higher biomasses at the lower moisture content in the Bridger soil. There
was more variation within plant species between the different soil moisture treatments
when the plants were grown in the Post Farm soil (Figure 8). Four of the species had
larger biomasses at the upper moisture content, specifically, A. canadensis, H. boreale,
L. argenteus and V. americana.
L. argenteus germinated in the Bridger soil at both moisture level treatments,
but the seedlings in the upper moisture level showed symptoms of root decay and died
early in the study. Tissues from these seedlings were plated and species of Fusarium
spp. and Alternaria spp. were isolated. The higher moisture content may have enabled
the fungal pathogens to establish in these plants. V. americana did not survive the

25
duration of the study in either moisture treatment of the Bridger soil. There were no
pathogens isolated from this tissue, but it is possible that the pH 8 of the Bridger soil
was too high for this specie. The NRCS National Plant Database lists this specie as
having pH tolerance of 6.5-7.2. The Dalea spp. showed no significant plant biomass
differences in either soil.
Nodulation
Overall, there was little significant variation in nodule mass with soil moisture
(Figure 9-10). However, significant differences in nodule mass between species grown
in the different soils were fairly large.
d
c
d
cd
LSD-0.20bb
aba
a
cd
c
0
0.2
0.4
0.6
0.8
1
1.2
AC DC DP HB LA VA
Species
Dry
Sho
ot B
iom
ass
(g p
ot -1
) BridgerPost Farm
Figure 6. Average dry shoot biomass of all six species in Bridger and Post Farm soils. Species biomass averaged from both 60 and 80% FC treatments. A. canadensis (AC), D. candida (DC), D. purpurea (DP), H. boreale (HB), L. argenteus (LA), V. americana (VA).

26
Figure 7. Average dry shoot biomass of six species in Bridger soil at 60-80% field capacity. The L. argenteus 80% FC and V. americana of both treatments did not survive long enough in this soil to yield biomass data.
LSD-0.12
b
bc
a
bcbc
b
bc
bcbc
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
AC DC DP HB LA VASpecies
Dry
Sho
ot B
iom
ass
(g p
ot -1
)
60% Field Capacity80% Field Capacity
Figure 8. Average dry shoot biomass of six species in Post Farm soils at 60- 80% field capacity.
d
bbb c
c
d
a
a
a
b
c d
d
0
0 .2
0 .4
0 .6
0 .8
1
1 .2
1 .4
A C D C D P H B L A V A
S p e c ie s
Dry
Sho
ot B
iom
ass
(g p
ot -1
)
6 0 % F ie ld C a p a c ity8 0 % F ie ld C a p a c ity
L S D - 0 .1 5

27
L. argenteus and V. americana had substantial nodule mass in the Post Farm
soil, but showed no signs of nodulation in the Bridger soil prior to the plant
senescence. The H. boreale nodule mass showed very little variation between soils or
water contents. D. candida had significantly higher nodule mass at the upper moisture
level of the Bridger soil and it was the only nodule mass to have significant differences
between moisture treatments (Figure 9-10). There were, however, large variations
between the nodule mass within the species when grown in the different soils.
Typically, the Post Farm soil produced larger nodule mass; however, the Dalea spp.
had higher nodule masses in the Bridger soil (Figure 9-10). This may be due to the
fact that D. candida has been sown for seed production at BPMC and although it is not
planted on the study site specifically, it is possible that soil in the area would contain
substantial populations of rhizobia specific to the Dalea spp. A. canadensis nodule
mass, which did not vary with moisture, was almost ten-fold lower in the Bridger soil.
b
b
c
b
c
b
b
a
0.00
0.02
0.04
0.06
0.08
0.10
0.12
0.14
0.16
AC DC DP HB LA VA
Species
Nod
ule
Mas
s (g
pot
-1)
60% Field Capacity80% Field Capacity
LSD-0.07
Figure 9. Average nodule mass of all six species in Bridger soils at 60 and 80% field capacity.

28
Drought stress can have a large effect on nodulation and N2 fixation in many
legumes; it is possible that the greenhouse pots retained more water than expected or
perhaps lower soil moisture content is needed to see drought effects. During drought
stress, N2 fixation is often halted before other physiological processes such as
photosynthesis, transpiration, and even new tissue growth, possibly due to plant
restriction on O2 diffusion in the nodule (Sinclair et al. 1987; Durand et al. 1987). So
although the nodule number and mass did not show a large response to drought
conditions in this study, it is still possible that there was a reduction in N2 fixation,
which was not directly measured. It is also possible that because the rhizobia in the
test soils were endemic they could be better adapted to drought conditions than the
species used in other studies. This trial indicated that specific characteristics of each
soil, such as microbial community, shrink-swell potentially and nutrient content, had
d
a
bc
d
cdcd
d d
b
a
ccd
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
AC DC DP HB LA VA
Species
Nod
ule
Mas
s (g
pot
-1)
60% Field Capacity
80% Field Capacity
LSD-0.07
Figure 10. Average nodule mass of all six species in Post Farm soils at 60-80% field capacity.

29
more effect overall on the response of individual species than actual moisture
treatments.
Trial III-Effects of fungicide coating on nodulation, biomass and plant N levels
Materials and Methods
Introduction
The objective of this trial was to assess the effect of fungicide seed coating on
the emergence and survival of a native legume. A simple experiment was designed
with five treatments. The seeds of D. candida were treated with two fungicides
(fludioxonol and metalaxyl) accompanied by a control with no fungicide application.
The germination and emergence of the treated and untreated seeds were recorded in
two soils (Geyser and Red Bluff -Table 2).
Seeding
The D. candida seeds were shaken with each diluted fungicide in a plastic bag
at the label rate (Table 3). Once all the seeds were evenly coated, they were spread flat
on a metal sheet to air dry. The soils and pots were prepared according to the protocol
listed in the Trial I seeding procedures, with the following exceptions. The seeding
Table 3. Fungicide information and dilutions.
Active Ingredient
AI* Dilution H2O Volume Applied
% mL g-seed-1
Fludioxonol 40.3 1: 499 1.20 Metalaxyl 32.3 1: 99 1.04
*Active ingredient

30
rate was increased to 50 PLS per pot, which were planted in two parallel rows 1 cm
deep. Each pot was watered with 50 mL of water and covered with plastic wrap. The
pots were then placed in a growth chamber at 20 ºC with 16 h of light and 8 h of
darkness. Plants were watered every other day with 50 mL of water for one month,
after which watering was reduced to every four days with the same volume. Harvest
again took place five months after seeding.
Data Collection
Germination and emergence were recorded every other day for 14 d and then
every 7 d until 42 days after seeding (DAS). Harvest and data analysis again followed
the protocol described in the greenhouse Trial I.
Trial III Results
Emergence and Plant Survival In the Geyser soil, emergence and survival of plants were higher for both
fungicide and control treatments when compared to the Red Bluff soil (Figure 11). Of
the fungicide treatments, metalaxyl had the largest beneficial effect on survival when
compared to the fludioxonol treatment and the control group in the number of
surviving plants (Figure 12). Both soils had a higher emergence rate when seeds were
treated with the fungicide metalaxyl, a fungicide typically used to stave off slime
molds and other pathogens that have a negative effect on seed germination and young
seedlings. Plant numbers in the fludioxonol were not significantly different from the

31
control group. Overall plant emergence and survival was higher in the Geyser soil
than in the Red Bluff soil.
Figure 11. Emergence rates of D. candida, 42 DAS in Geyser (G) and Red Bluff (RB) soils. Fungicide treatments: fludioxonol (F), metalaxyl (M), and control (C).
0
10
20
30
40
50
60
70
0 7 14 21 28 35 42
DAS
Emer
genc
e (%
)
CGCRBFGFRBMGMRB
Figure 12. Average plant number, five months after seeding D. candida in Geyser and Red Bluff soils treated with fludioxonol and metalaxyl fungicides.
c
b
c
b
a
b
0
10
20
30
40
50
60
70
Control Fludioxonol Metalaxyl
Species
Plan
t Sur
viva
l (%
)
GeyserRed BluffLSD= 12

32
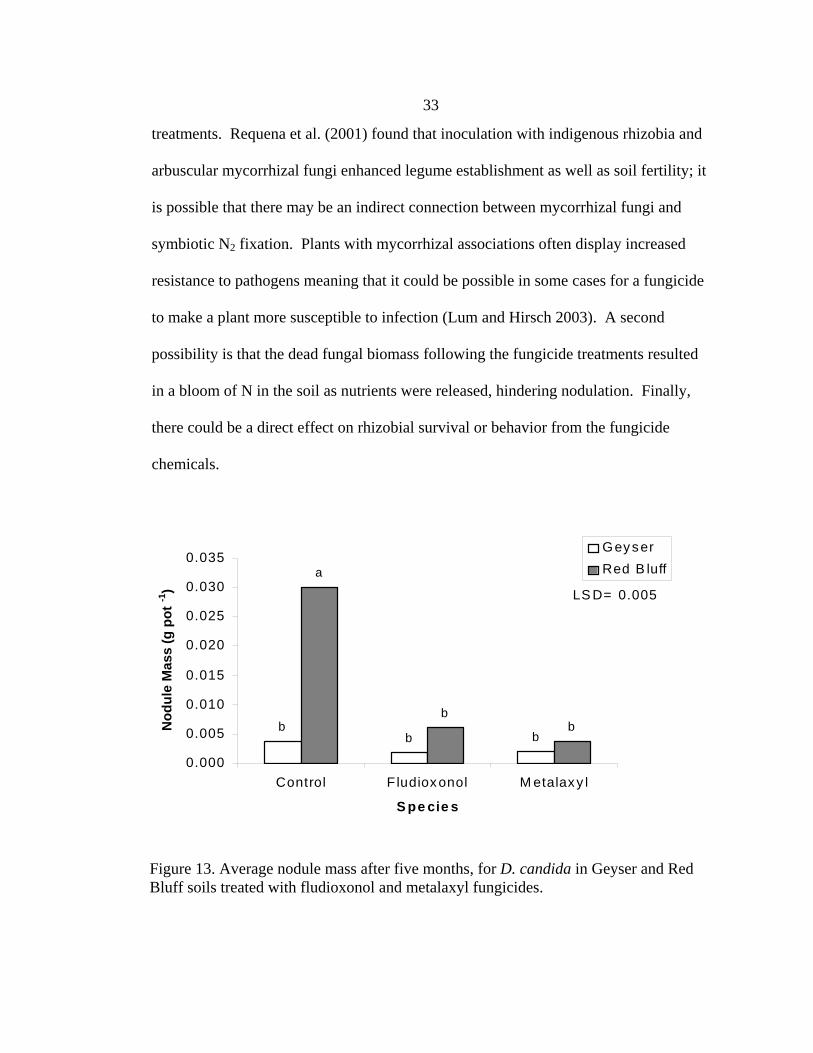
Nodulation and Biomass Nodule mass of D. candida in the Red Bluff soil was less with fungicide than in
the control (Figure 13). Nodule masses from the fungicide treatments in the Geyser soil
were not different from those in the control group. The nodule mass of the Red Bluff
control group was 0.03 g, ten-fold higher than the fungicide treatments, which had
masses between 0.003-0.006 g. However, in the Geyser soil, the control nodules were
just two times the mass in the fungicide treatments, with a mass of 0.004 g for the
control compared to 0.002 g in the treatments. Perhaps because legumes in the Geyser
soil already have difficulty forming nodules the decrease in mass due to fungicide
treatment is less substantial. Nodule numbers exhibited nearly identical results as
nodule mass (Figure 14).
Nodulation appeared to be negatively affected by fungicide treatments,
although the degree of the effect was dependent on soil characteristics. Several studies
have found that although chemical pesticides and fungicides may be effective to some
degree, they may have detrimental effects on symbiotic N2 fixation (Bagnasco et al.
1998). The nodules in the fungicide treatments were typically white in color lacking
the pronounced pink pigmentation often associated with LHb and active N2 fixation
machinery (Figure 15). It is possible that the process of symbiotic N2 fixation is
adversely affected by the two fungicide treatments; more testing should be done to
investigate other chemical and rate effects on legumes.
The proper development and function of nodules and N2 fixation appears to be
hindered by these fungicide treatments. There are several possibilities as to why these
effects are seen. All species of fungus in the soil would be killed by the fungicide
33
treatments. Requena et al. (2001) found that inoculation with indigenous rhizobia and
arbuscular mycorrhizal fungi enhanced legume establishment as well as soil fertility; it
is possible that there may be an indirect connection between mycorrhizal fungi and
symbiotic N2 fixation. Plants with mycorrhizal associations often display increased
resistance to pathogens meaning that it could be possible in some cases for a fungicide
to make a plant more susceptible to infection (Lum and Hirsch 2003). A second
possibility is that the dead fungal biomass following the fungicide treatments resulted
in a bloom of N in the soil as nutrients were released, hindering nodulation. Finally,
there could be a direct effect on rhizobial survival or behavior from the fungicide
chemicals.
Figure 13. Average nodule mass after five months, for D. candida in Geyser and Red Bluff soils treated with fludioxonol and metalaxyl fungicides.
bbb
a
bb
0.000
0.005
0.010
0.015
0.020
0.025
0.030
0.035
Control F ludioxonol M etalaxy l
S pe cie s
Nod
ule
Mas
s (g
pot
-1)
GeyserRed B luff
LS D= 0.005

34
Figure 15. Nodules from D. candida, treated with metalaxyl (left) and without fungicide treatment (right).
Figure 14. Average nodule density per plant after five months for D. candida in Geyser and Red Bluff soils treated with fludioxonol and metalaxyl fungicide.
b
bb
a
b
b
0
5
10
15
20
25
30
35
40
45
Control Fludioxonol Metalaxyl
Spe cie s
Nod
ule
Den
sity
(No.
pla
nt -1
)
GeyserRed Bluff
LSD= 12

35
In the Red Bluff soil, there was no significant variation in shoot biomass
between the fungicide treatments, but the control group had significantly higher
biomass (Figure 16). There was no significant variation within the Geyser treatments.
It is possible that the detrimental effect of the fungicide was not apparent in the Geyser
soil because infection and nodulation mechanisms were already constrained in the
Geyser soil (Figure 13). The benefits of fungicide treatments must be weighed with the
cost of possible reduction in nodule function. However, if pathogens are a problem at
a given site, fungicides may prove beneficial to the establishment of these species. If a
site does not have a history of soil pathogen problems, then fungicide application could
be detrimental to nodulation and long-term plant survival. The following trials further
examined the effects of fungicide treatments on nodulation and biomass.
Figure 16. Average dry shoot biomass of D. candida in Geyser and Red Bluff soils treated with fludioxonol and metalaxyl fungicides.
c cc
a
bcb
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
Control Fludioxonol Metalaxyl
Species
Dry
Sho
ot B
iom
ass
(g p
ot -1
)
GeyserRed Bluff
LSD=0.3

36
Trial IV: Effects of fungicide drench on nodulation and biomass
Materials and Methods Introduction
Due to the results in Trial III, further tests were designed to assess the effects of
fungicide treatments on nodulation in two native legumes (A. canadensis and L.
argenteus). These species were selected in order to test both a Rhizobium sp., which
nodulates A. canadensis, and a Bradyrhizobia sp., which nodulates L. argenteus. Soils
were collected from Bridger, Post Farm and Moccasin (Table 2). The treatment was a
soil drench, which soaks all the soil in the entire pot with label strength fungicide,
versus the seed coat, which only contacts the seed surface. This drench contained a
wide spectrum fungicide with the active ingredients etridozole (15%) and thiophanate-
methyl (35%). An equal volume of water was used as a drench for the control pots.
There were four replications of each treatment and there were two harvest times over
the course of the experiment. The objective was to assess how the effects of fungicide
on biomass and nodulation changed over time as the plants matured and the fungicide
became systemically diluted within the plant.
Seeding
The basic protocol from Trial I was followed, with the following exceptions.
During the first week in the greenhouse, the first drench application of 200 mL of
fungicide was applied to each pot at a dilution rate of 1000:1. This application was
repeated twice at 7 d intervals for a total of three applications. The control group

37
received a 200 mL water drench at each application. Each pot was watered every 3 d;
a high moisture level was used in order to promote fungal growth. The first harvest
took place at eight weeks after seeding and the second at five months.
Data Collection
Harvest followed the protocol from Trial I, but some additional data were
collected as follows. After removing biomass from the drying oven, the aboveground
biomass of each plant from the second harvest was ground and delivered to the
SPWAL for total percent N analysis of biomass using LECO combustion analysis.
Tissue samples of each treatment were collected from the roots, stems and leaves of
the plants. These tissues were surface sterilized by covering in a 10% sodium
hypochlorite (NaOCl) solution and stirring for five minutes; they were then rinsed with
sterilized DIH2O. The sterilized tissues were then placed on potato dextrose agar
plates (PDA), labeled, and allowed to grow at room temperature for a period of about 7
d. Fungi arising on the plates were identified with the assistance of Dr. Bill Grey of
MSU. Fungi suspected of being pathogenic species were isolated. Isolation was
achieved by taking a small sample from the leading edge of each fungus with a flame-
sterilized wire. This sample was placed on a sterile PDA plate. Isolates were allowed
to culture again for about 7 d at room temperature and were then identified to the
species level via microscopic observation of cellular structure.

38
Trial IV Results Biomass
The biomass from the fungicide treatments appeared to have more substantial
differences from the control during the first harvest indicating that fungicide effects
may be reduced as the fungicide is systemically diluted within the plant (Figure 17 and
18). The A. canadensis plants treated with fungicide in the Post Farm soil had higher
biomass than the control during the first harvest (Figure 17). Conversely, the biomass
of both species was higher in the Moccasin soil when there was no fungicide
application. L. argenteus biomass in the Moccasin soil decreased with fungicide.
Fungicide did not significantly affect biomass in the Bridger soil.
By the second harvest, there was also no significant variation in A. canadensis
biomass between the treatment and control groups in the Bridger soil (Figure 18).
However, the A. canadensis biomasses in the Post Farm and Moccasin soils were
higher in the control compared with the fungicide group. The L. argenteus biomass
from the second harvest in the Bridger soil was larger in the fungicide treatment than
in the control. However, this species no longer had any differences in the Post Farm or
the Moccasin soils (Figure 18). In treatment groups there may have been detrimental
chemical or salt effects from the fungicide that decreased with time. Compared to the
control group, some of the treated plants did show symptoms of chemical toxicity. The
EC of the fungicide solution was not measured, a control with a similar EC or salt
content should be used if these experiments are repeated. Fungicide treatments can
have a positive effect on seedling emergence and plant survival when fungal pathogens

39
are present and virulent in a soil. The benefits of fungicide are most pronounced in
earlier growth stages presumably because this is the period when most species are
more susceptible to disease. However, if there are no pathogens present or if fungal
species are non-virulent, the fungicide may be detrimental to plant health.
Root Structure, Nodules and Fungicide
The root structure of the treated plants from both harvests were healthier, more
flexible, and showed less discoloration and more flexibility, than the control group
which had large patches of pink and brown discoloration (Figure 19). The roots in the
control group were also very brittle and broke easily during harvest and washing.
However, the control plants were more extensively nodulated. The fungal pathogens
Fusarium spp. and Alternaria spp. were isolated from the plant tissues in both Bridger
and Post Farm soils. Specifically, F. solani and F. oxysporum were isolated from plant
tissue grown in the Post Farm soil. Both Fusarium spp. are known to be virulent
fungal pathogens that cause root and stem decay.
dd
d
cd
a b
b c
a
cd cd
b cb
cd
0
0 .1
0 .2
0 .3
0 .4
0 .5
0 .6
0 .7
A C C A C F L A C L A F
S p e c ie s a n d T re a tm e n t
Dry
Sho
ot B
iom
ass
(g p
ot -1
)
B r id g e rM o cca s inP o s t F a rm
L S D - 0 .2 2
Figure 17. Effects of a water (C) drench vs. fungicide (F) drench on average shoot biomass of A. canadensis (AC) and L. argenteus (LA) 8 weeks after seeding.

40
f
de
f f
b
ede
cd de de
bc
a
0.00.51.01.52.02.53.03.54.04.5
AC C AC F LA C LA F
Spe cie s and Tre atme nt
Tota
l Dry
Bio
mas
s (g
pot
-1)
BridgerMoccasinPost Farm
LSD- 0.65
Figure 18. Effects of a water (C) drench vs. fungicide (F) drench on average dry shoot biomass of A. canadensis (AC) and L. argenteus (LA) 5 months after seeding.
Figure 19. L. argenteus grown in Post Farm soil fungicide treatment (left) shows healthier, more robust, root system while untreated control roots had more discoloration and were brittle.

41
N Content There was little significant variation in the percentage of N within the plant
tissue in the second harvest (data not shown); however, when total N content of the
plant was calculated by multiplying the N fraction by the total biomass there were
some significant differences (Figure 20). N content of plants in the Bridger soil were
significantly lower than the levels of the plants grown in the Post Farm and Moccasin
soils.
A. canadensis showed no significant variation in N content between treatments.
However, there were significant differences in total N content of plants grown in the
different soils. Plants from the Bridger soil had the lowest N content and those from
the Post Farm soil had the highest N content. The L. argenteus plants showed similar
significant variation in N content between sites but there was more variation between
treatments in this species. Plants from the Bridger soil showed no variation in either
species. However, L. argenteus from the Moccasin and Post Farm soils had
significantly higher N content in the control treatments. These results indicate that
fungicide may indeed have a detrimental effect on nodulation and/or N2 fixation.
Fungicides have an overall beneficial effect on the condition of root systems but there
was some evidence that fungicides may be harmful to rhizobia or interfere with the
establishment of fully functioning N2 fixation mechanisms.

42
Trial V: Effects of inoculation and fungicide application on nodulation and
biomass
Materials and Methods
Introduction
The final trial of the greenhouse research was designed to test the effects of
fungicide as well as the effects of inoculation on all six species. The primary focus
was to compare the ability of rhizobial strains from a commercial inocula versus the
ability of the endemic rhizobia species to nodulate the legumes. The effect of
fungicide on nodulation and plant growth response, and the combination of both
treatments, was also evaluated.
Figure 20. Effects of a water (C) drench vs. fungicide treatment (T) drench on average shoot biomass total N content of A. canadensis and L. argenteus from the second harvest.
de dee e
b
c
d d
a
bc
cc
0 .00
0 .02
0 .04
0 .06
0 .08
0 .10
0 .12
0 .14
AC C AC T LA C LA T
Treatm ent
Shoo
t N C
onte
nt (g
pot
-1)
Bridge r
Moccas in
Pos t Fa rmLSD -0 .02

43
Seeding
In this trial, 5 cm diameter conetainers were used to add soil depth and increase
the stability of the field soils. Soils were collected from Moccasin and Post Farm
(Table 2) and available commercial and experimental inocula were obtained (Table 4).
There were a total of four treatments for each soil type with four replications. All six
species were tested with and without inocula (Table 4). A. canadensis, D. candida,
and L. argenteus were further tested with and without a fludioxonol fungicide seed
coating treatment as well as combining the fungicide with and without inocula. The
additional treatments were excluded to limit the size of the experiment.
Table 4. Rhizobia and legume inocula information.*
Plant Species Rhizobia Name/Type ID Label**
Astragalus canadensis Astragalus Spec. 1 LX1893 Dalea candida Type “M” Crownvetch/Dalea M-4 Dalea purpurea Type “M” Crownvetch/Dalea M-4 Hedysarum boreale Hedysarum spec. 2 LX1897 Vicia americana Vicia Spec. 5 LX1866 Lupinus argenteus Type “H” Lupine H-5 *Inocula obtained from Nitragin Worldwide, Brookfield, WI. **The LX ID labels are experimental and not available for commercial release.
Conetainers were washed and dried prior to filling with the pre-moistened soils.
Eight seeds were planted in each conetainer. Seedlings were thinned to one plant two
weeks after germination. Seeds in the fungicide treatment were coated as described in
Trial III. Following planting, 1 g of peat inocula (Table 4) was mixed with 1 mL of
water and the solution was pipetted over the top of each seed in the conetainer. The
seeds were then covered with 1 cm soil. Each conetainer was watered with 30 mL of
water and placed in the greenhouse to grow for 5 months. The plants were watered

44
with approximately 30 mL of water every 3 d during the growth period. Harvest
followed the procedure listed in Trial IV.
Results Trial V
Introduction
There were major differences in plant responses between soils. A. canadensis
had a much higher biomass than the D. candida, so the following discussion and
figures focus on A. canadensis only, in order to reduce variation and further investigate
the effects of the treatments. The results and a brief discussion on the inoculated vs.
non-inoculated treatments of Dalea spp., H. boreale and V. americana can be found in
Appendix B since there were not enough surviving plants to draw any significant
conclusions. The L. argenteus plants had very few survivors and were removed from
the statistical data analysis and left unpublished.
A. canadensis Biomass
The A. canadensis plants grown in the Moccasin soil generally had higher
biomass production than the plants in the Post Farm soil, although the inoculated only
treatments (CI) were similar in both soils (Figure 21). The combined fungicide and
inocula treatment in the Post Farm soil had the smallest biomass of all, and was
significantly lower compared to the fungicide treatment and the no treatment group.
The inoculation only treatment had the lowest biomass in the Moccasin soil and was
different from the non-inoculated control and fungicide treatments.

45
A. canadensis Nodulation
Nodule numbers in the Moccasin soils showed no differences (Figure 22). In
Post Farm soil, the largest nodule numbers were in the non-inoculated fungicide
treatment. The combined inocula-fungicide treatment had the fewest nodules and of
all the treatments.
Nodule masses showed more differences than the other dependent variables in
this study (Figure 23). All four treatments differed in the Post Farm soil, with the
largest nodule mass in the non-inoculated, fungicide treatment, followed by the non-
inoculated, no fungicide treatment. The combined inocula-fungicide treatment had the
lowest nodule mass; apparently there are some interactions between these treatments.
In the Moccasin soil, the non-inoculated, no fungicide treatment had a higher nodule
mass than all other treatments.
Figure 21. Average total dry biomass of A. canadensis with fungicide (F) and control (C) treatments; both inoculated (I) and non-inoculated (U), in Post Farm and Moccasin soils.
aa
ab
bc
b b
bc
c
0.00.10.2
0.30.40.50.6
0.70.8
CU CI FU FI
Tre a tm e nt
Dry
Sho
ot B
iom
ass
(g p
ot-1
)
Moccas inPost FarmLSD-0.2
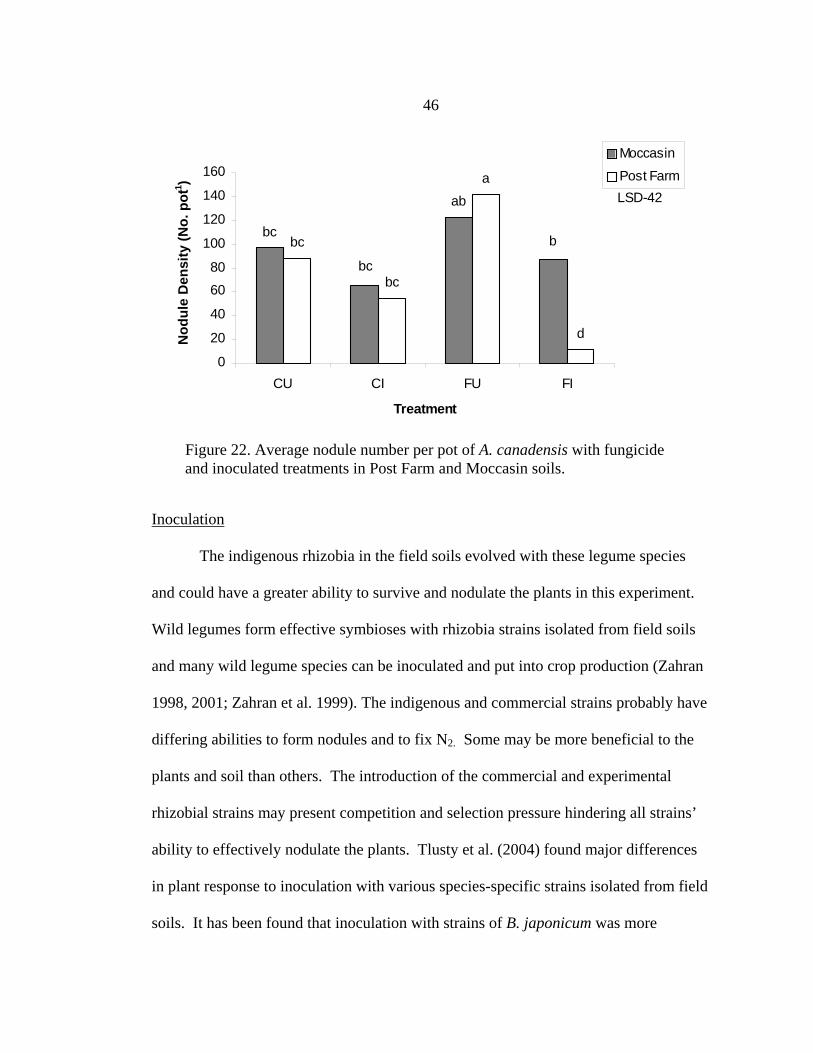
46
Inoculation
The indigenous rhizobia in the field soils evolved with these legume species
and could have a greater ability to survive and nodulate the plants in this experiment.
Wild legumes form effective symbioses with rhizobia strains isolated from field soils
and many wild legume species can be inoculated and put into crop production (Zahran
1998, 2001; Zahran et al. 1999). The indigenous and commercial strains probably have
differing abilities to form nodules and to fix N2. Some may be more beneficial to the
plants and soil than others. The introduction of the commercial and experimental
rhizobial strains may present competition and selection pressure hindering all strains’
ability to effectively nodulate the plants. Tlusty et al. (2004) found major differences
in plant response to inoculation with various species-specific strains isolated from field
soils. It has been found that inoculation with strains of B. japonicum was more
bc
ab
b
bc
bc
a
bc
d
0
20
40
60
80
100
120
140
160
CU CI FU FI
Treatment
Nod
ule
Den
sity
(No.
pot
-1)
Moccasin
Post FarmLSD-42
Figure 22. Average nodule number per pot of A. canadensis with fungicide and inoculated treatments in Post Farm and Moccasin soils.
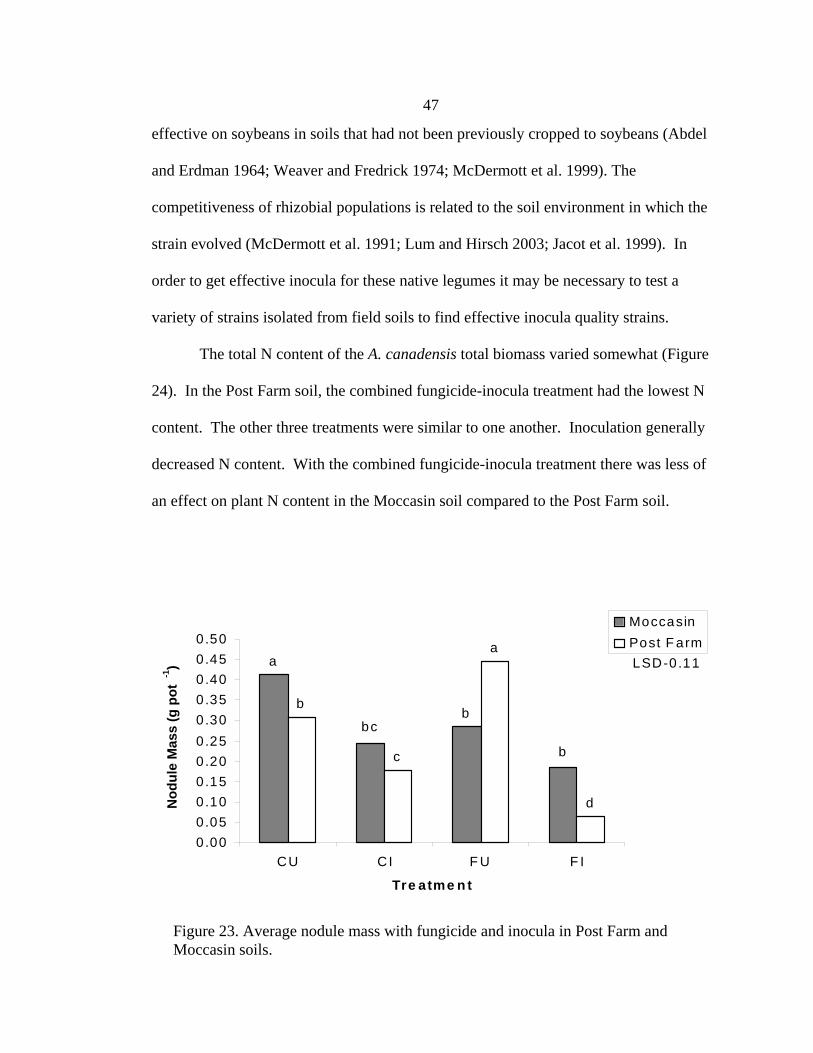
47
effective on soybeans in soils that had not been previously cropped to soybeans (Abdel
and Erdman 1964; Weaver and Fredrick 1974; McDermott et al. 1999). The
competitiveness of rhizobial populations is related to the soil environment in which the
strain evolved (McDermott et al. 1991; Lum and Hirsch 2003; Jacot et al. 1999). In
order to get effective inocula for these native legumes it may be necessary to test a
variety of strains isolated from field soils to find effective inocula quality strains.
The total N content of the A. canadensis total biomass varied somewhat (Figure
24). In the Post Farm soil, the combined fungicide-inocula treatment had the lowest N
content. The other three treatments were similar to one another. Inoculation generally
decreased N content. With the combined fungicide-inocula treatment there was less of
an effect on plant N content in the Moccasin soil compared to the Post Farm soil.
Figure 23. Average nodule mass with fungicide and inocula in Post Farm and Moccasin soils.
a
b
b
bc
b
a
c
d
0.000.050.100.150.200.250.300.350.400.450.50
CU CI F U F I
Tre atme n t
Nod
ule
Mas
s (g
pot
-1)
MoccasinPost F armLSD-0.11

48
Greenhouse Studies Summary
Soils from different origins varied in texture, nutrient content, pH and perhaps
more importantly, they apparently contained different microbial communities.
Microbial populations evolve very rapidly and because bacteria, in particular, are
limited in their movement and dispersal, it is not unreasonable to expect the genotypes
of rhizobial strains to vary significantly according to geographic location and soil
environment. These soil characteristics or origins generally had more of an effect on
biomass, nodulation and overall emergence and survival of the legume species than did
most of the other treatments in this research project. A. canadensis often had the
largest amount of biomass, as well as the highest level of nodulation, both in weight
Figure 24. Average total N content of entire A. canadensis biomass from greenhouse Trial V with fungicide and inoculation treatments in Post Farm and Moccasin soils.
a
a
ab
c
bc bcbc
d
0.000
0.005
0.010
0.015
0.020
0.025
0.030
0.035
0.040
0.045
C U C I FU FI
T re atme nt
Tota
l N C
onte
nt (g
N p
lant
-1)
Moccas inPos t Farm
LSD -0.011

49
and number, throughout all the greenhouse trials. L. argenteus, H. boreale and V.
americana followed with interchanging, but not significantly different biomass levels.
H. boreale and L. argenteus had a moderate level of nodulation, followed by V.
americana, although these species all varied depending on the field soil tested. The
Dalea spp. generally had the lowest biomass and nodulation level of the six species.
Fungicide treatments, as well as inocula treatments, had varying effects on
plant biomass and the extent of nodulation. These plant responses were highly
dependent on the soil type and microbial populations. Fungicides had inconsistent
effects on emergence and survival, but in general, they appeared to negatively affect
nodulation. Inoculation also produced some mixed responses and generally decreased
shoot biomass, nodule number and mass, again depending upon the field soil used.
Host specificity, competition, and/or soil environment outside of the species’
adaptation ranges are all factors that can affect plant response and should be
considered when using introduced or commercial strains of inocula. It is likely that the
indigenous microbial communities in a soil are of primary importance to legumes.
Symbiosis of rhizobia and fungi, both beneficial and pathogenic, influence plant
survival and production. Isolation and research on endemic strains of rhizobia from
native legume species grown in field soils could improve inocula quality and
effectiveness and may benefit plant establishment.

50
CHAPTER 3
FIELD RESEARCH
Introduction
Legume-grass ecosystems have been found to improve soil fertility, habitat
potential and forage quality; however these systems can be difficult to establish and
maintain (Ridley et al. 2004). In this field study, the establishment, biomass and N2
fixation estimate of six native legume species when seeded with native grasses (Table
5) were evaluated at three field sites. In addition, changes in the total N level of the
soil and effects of legumes on grass biomass and N content were measured. The
selected field sites included the Bridger Plant Materials Center in Bridger (BPMC), the
MSU Post Farm Agricultural Research Center in Bozeman and the MSU Agricultural
Research Station in Moccasin; all three locations are in the state of Montana (Table 6).
Table 5. Grass seed and species information Plant Species Common Name Pure Seed Germination Total Viability PLS*
------------------------------------%--------------------------- Poa secunda Sandberg Bluegrass 93 4 89 83 Elymus trachycaulus Slender Wheatgrass
98 93 93 91
Pascopyrum smithii Western Wheatgrass 93 92 92 86 Elymus lanceolatus Streambank Wheatgrass 98 99 99 97
* PLS- pure live seed

51
Table 6. Field Site Information Bridger Moccasin Post Farm
Past Crop Fallow Grasses Barley Fallow Grasses
Last legume + 10 yrs + 10 yrs 3 yrs Fertilizer No No No Pesticide Applied:
June 2004 Mustang
Applied: July and August 2004 Metathyon No
Irrigation Monthly: May-Sept 2003/2004
Sprinkled 38 mm No
1 time August apx. 2 hours
flooded Yearly Precipitation
Average (mm) 292 390 414
2003 Precipitation (mm) 169 371 345 2004 Precipitation (mm) 215 294 384 Temperature Min-Max
(ºC)
May (ºC) 0.55-16.7 3.3-16.7 1.1-18.3 June (ºC) 11.6-31.6 10-26.7 10-27.8
Materials and Methods
Site Preparation and Design
Soil characteristics were analyzed and basic soil properties at each site were
listed in Table 2. Information on the monthly temperature and precipitation amounts,
as well as site history, fertilizer and irrigation levels for each field site are included in
Table 6. Bridger and Post Farm were tilled and rolled to pack the soil prior to planting.
Moccasin was directly seeded perpendicular to stubble rows.
The treatments at all three sites consisted of alternating rows of a native grass
seed mixture (Table 5) and each legume species listed in Table 1. In addition, two
comparison treatments were used; M. sativa as a crop legume specie was alternated
with the native grass mix, and four native grass plots were seeded without legumes.
Treatments were replicated four times at each of the three field sites, in a complete

52
randomized design. Each site had 42 plots, 1.5 m by 3 m each, consisting of 35
treatments and 7 demonstration blocks in which the grasses were inter-seeded in the
same row as each of the legumes. The total area of each site was approximately 512
m2. Seeds were planted at a rate of 20 pure live seeds per linear foot totaling 200 PLS
per row in each treatment block. Grasses were seeded at 480 PLS per row; they were
inadvertently seeded at twice the normal rate.
Seeding
A four-row drill seeder was used to plant four rows spaced at 0.3 m and seeded
1.3 cm deep. The entire research block was surrounded with the grass seed mix in
order to reduce edge effects. Maintenance of the sites was minimal. Each site was
weeded several times the first season depending on the need for weed control at each
site. Weed removal took place one time at the beginning of the second season. Other
maintenance operations, such as insecticide treatment and irrigation, are listed in Table
6.
Data Collection
Following seeding, soil cores were taken with a 4 cm diameter soil auger 60 cm
deep at the Post Farm and Bridger. At Moccasin, the core depth was decreased to 30
cm due to high gravel content. Each core was placed in a 20 L plastic bucket,
homogenized, placed in a soil sample bag and stored in a cooler. The soil core and
bucket were cleaned using a bottlebrush and paper towels to remove any remaining

53
soil and prevent contamination between samples. Wet weight of each sample was
collected and the samples were dried in a soil dryer at 40˚ C for 14 d and weighed
again to calculate soil moisture content. The samples were then submitted to the MSU
SPWAL for analysis of NO3-N. NH4-N was not measured in 2004 because
concentrations in 2003 were generally very low (1 mg kg-1). Composite samples from
each site were used for routine soil analysis, consisting of texture and selected nutrient
content (Table 2).
Immediately following harvest, soil samples were again collected from each
plot as described earlier. At Moccasin the depth was again reduced to 30 cm. Each
soil sample then went through the prior procedure.
During the first season, germination numbers for both grasses and legumes
were recorded. Harvest began at the Post Farm during the third week of July, 2004.
Moccasin was harvested the fourth week of July and the Bridger site was harvested the
first week of August. Plant biomass was collected during the second season by placing
a 0.6 m by 1.5 m wooden frame over the center of the two interior rows of each plot,
again to reduce edge effects. The grasses and legumes were then clipped separately at
the ground level and the fresh biomass was placed in a 20 L plastic bucket and
weighed in the field. Each row was sub-sampled and weighed again; these sub
samples were then placed in a plant-drying oven at 40º C for 7 d. At this point, the
samples were reweighed and moisture content for each plot was calculated. Each
sample was then ground and submitted to the MSU SPWAL for analysis of total N via
LECO combustion.

54
To estimate the difference in N2 fixation between species, a N mass balance
was calculated by the following equation: N mass balance = N uptake + (soil N 2004-
soil N 2003) in the upper 60 cm (30 cm at Moccasin) of the soil profile. This formula
assumes that leaching, denitrification, mineralization, immobilization and volatilization
were fairly constant between plots. By assuming that, differences in the mass balance
were equated directly with differences in N2 fixation amounts between plots within
each site. although N uptake for below the soil sampling depth would overestimate N
fixation. N content of tissues was multiplied by biomass production to determine N
uptake. Total dry biomass N was estimated to be 1.82 x shoot N content, based on
shoot N: root N ratios from greenhouse studies. The grass-grass plots were used as a
comparison because N2 fixed in these plots should have been essentially zero in each
plot.
Nodulation was verified in the field for each species by collecting a portion of
the root mass and inspecting it for nodulation. Nodule presence and pigmentation were
recorded. Plant tissue was also collected to isolate and identify plant pathogens as
described in Trial IV of the greenhouse studies. Statistical analysis was performed
using a natural log transformation of the data means.
In order to test the accuracy of the data collection as well as the laboratory
analysis, quality control procedures were taken by splitting two field duplicates of two
plant and soil samples every sixteenth sample for each site (n=6) (Dollhopf 2000).
These duplicate samples were labeled as part of the sample sequence and submitted
with the remainder of the samples for data analysis. These splits were used to calculate
the % relative standard deviation (% RSD) for our data samples. The % RSD levels

55
for N content of the soil, dry grass shoot biomass, and dry legume shoot biomass were
18%, 11% and 8% respectively. These are fairly substantial error margins, however, it
is not uncommon to see larger errors when studying natural systems. As will be seen,
the differences between biomasses are substantial; therefore, these errors likely do not
affect the conclusions of the study.
Field Research Results
Site Comparisons Native legumes can have a fairly low germination rate; therefore, emergence
was measured two months after seeding (Figure 27). Germination at the Moccasin site
was the highest, followed by the Post Farm and finally Bridger. Moisture levels at the
time of seeding followed the same pattern with Moccasin having the highest moisture
and Bridger the lowest levels; this probably in part affected the emergence rates at each
site. A. canadensis had low rates at all three sites, although it is slightly outside of its
historical range. M. sativa typically had a higher rate of emergence than the native
legumes, although H. boreale had similar rates at the Moccasin location. According to
McGraw et al. (2003) the optimal temperature for emergence of Dalea spp. is around
25 °C, much higher than the temperatures at the time of seeding. With lower
temperatures, the time to germinate is increased and the total number of seeds to
germinate is slightly decreased; however, the difference in emergence rates for these
species showed no marked difference between 15-30 °C (McGraw et al. 2003). Being
able to germinate well across a wide variety of temperatures is an advantage that native
plant species often have (McGraw et al. 2003).

56
The Post Farm location had the most favorable environmental conditions and
was the highest producer in total biomass of all sites; both the grasses and the legumes
out-performed those at the other two sites (Figure 28). Moccasin and Bridger both had
lower moisture levels as well as insect and wildlife grazing. Plant biomass at the
Moccasin location varied the least between plots and the native legumes at this site
typically produced larger biomasses than those at Bridger, with the exception of A.
canadensis. The M. sativa was very prolific at Bridger and biomass production was
comparable to that of the Post Farm.
Bridger
Bridger had the lowest precipitation of the three sites and the annual rainfall
during the study period was well below the long-term average (Table 6). The plots
were irrigated monthly with sprinkle irrigation to deliver approximately 38 mm across
the site during each irrigation cycle.
Legumes
At two months after planting, less than 20% of the native legumes seeded at
this site had emerged (Figure 27). The emergence rates of the native species at this site
did not vary much and M. sativa had twice the average emergence (44%). M. sativa
had the highest biomass by far (Figure 29), which could be due to the fact that M.
sativa was so prolific at this site and had been previously grown in the vicinity.
Specifically, some M. sativa plants were removed from the other legume and grass
rows during the weeding sessions. M. sativa also has a deeper root system than native

57
legumes and may have been able to access nutrients and deep soil moisture (USDA-
NRCS 2004). Accumulation of N below the root zone is common in irrigated systems
due to leaching. Deeper rooted species, such as M. sativa, can take advantage of N
fixed by more shallow rooted legumes (Ridley et al. 2004).
A. canadensis had the second highest biomass, and was similar to the
biomasses produced by the grass-grass plots (Figure 29). Of the native species this
was typically the most successful during this study. D. candida, L. argenteus and H.
boreale had the next highest biomasses of the native legumes. D. candida is currently
being grown and produced in other plots at BPMC. Two species of insects were found
parasitizing the root system of this species. Many of the legume plots showed signs of
drought stress and grazing damage from insects, antelope and rabbits at the time of
harvest, which would negatively affect biomass results.
Figure 27. Legume emergence in 2003 field season 2 months after planting. A. canadensis (AC), D. candida (DC), D. purpurea (DP), H. boreale (HB), L. argenteus (LA), Grass Mixture (GR), M. sativa (MS), V. americana (VA).
ab
c
ccc
ccc
a
ab
ab
bcbc
c
ab
bcbc
ccc
c
0
10
20
30
40
50
60
AC DP DC HB LA MS VA
Species
Em
erge
nce
(%)
Bridger MoccasinPost Farm
LSD- 23

58
The H. boreale showed the highest level of water stress and plants in several of
the plots were wilted at the time of harvest. This specie was quick to recover when soil
moisture was restored, but it was also the first specie to show symptoms of wilting. It
was observed in the greenhouse that this specie was able to regenerate from
belowground biomass after extreme drought and loss of most of the original
aboveground biomass.
Nodulation was verified in five of the seven species at this site; L. argenteus
and V. americana showed no signs of nodulation although removal of an intact root
system was not possible. The NRCS lists these two plants as being adapted to pH
ranges slightly lower than that of the soil at Bridger (Table 6), which could affect the
rhizobia as well as plant growth. These are the same species that performed poorly in
Figure 28. Average total combined grasses and legume dry aboveground biomass from field sites.
d
c
d
c
d d
d
d
c c cc c
cd
c
cd
a
b
c
a
aa
b
a
020406080
100120140160
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)
Bridger Moccasin Post Farm

59
this soil during the greenhouse phases. M. sativa was also verified to have nodulation.
It is possible that rhizobia were introduced to the soil by seed or soil at some point. It
is not likely that indigenous rhizobia in Montana soils would be specific to M. sativa.
All of the species except H. boreale flowered at Bridger, although not to the
extent seen at the other two sites. Several of the Dalea spp. plants showed signs of
seed formation at the time of harvest; however, none of the other plants were close to
setting seed, possibly because harvest took place well before most of these species
typically set seed, usually in late August or early September. H. boreale was probably
unable to produce flowers due to the stress from drought and grazing. This specie is
one of the most palatable of the native species and was grazed heavily, which would
effect total biomass data collection.
Combined Biomass The biomass of the grasses plus the native legumes at Bridger showed the most
variation of the three sites (Figure 28 and 30). M. sativa plots had the highest biomass.
A. canadensis and the grass only plots had the next largest biomasses. D. candida and
L. argenteus had the third highest biomasses but were only slightly higher than the
other natives. The results at Bridger were the opposite of the results at the other two
sites, where the grass-native legume plots typically produced the largest grass biomass,
which somewhat reduced the yield in total biomass between the plots. The relative
biomass order of the combined grass-legumes treatments at the Bridger site was similar
to results of the legume biomass (Figure 29 and 30).

60
Grasses
The grasses in the A. canadensis plots had the largest biomass and were similar
to the biomass of the grasses in the grass-grass plots (Figure 31). This suggests that
this native specie had little or no influence on grass biomass production at this site. D.
candida and L. argenteus plots had the next highest grass biomass. H. boreale, D.
purpurea, V. americana and L. argenteus grasses had similar biomasses. This site was
surface irrigated and it is possible that the legume root systems remained close to the
surface in order to obtain moisture, whereas at the other sites, with no irrigation,
moisture was probably obtained deeper in the soil profile This may have resulted in
more competition between the grasses and the smaller native legumes because they
would have obtained moisture and nutrients from the same root zone.
Figure 29. Average legume dry shoot biomass from the Bridger field site. Note log scale.
c
b
d
b
cdc
a
d
1
10
100
1000
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)
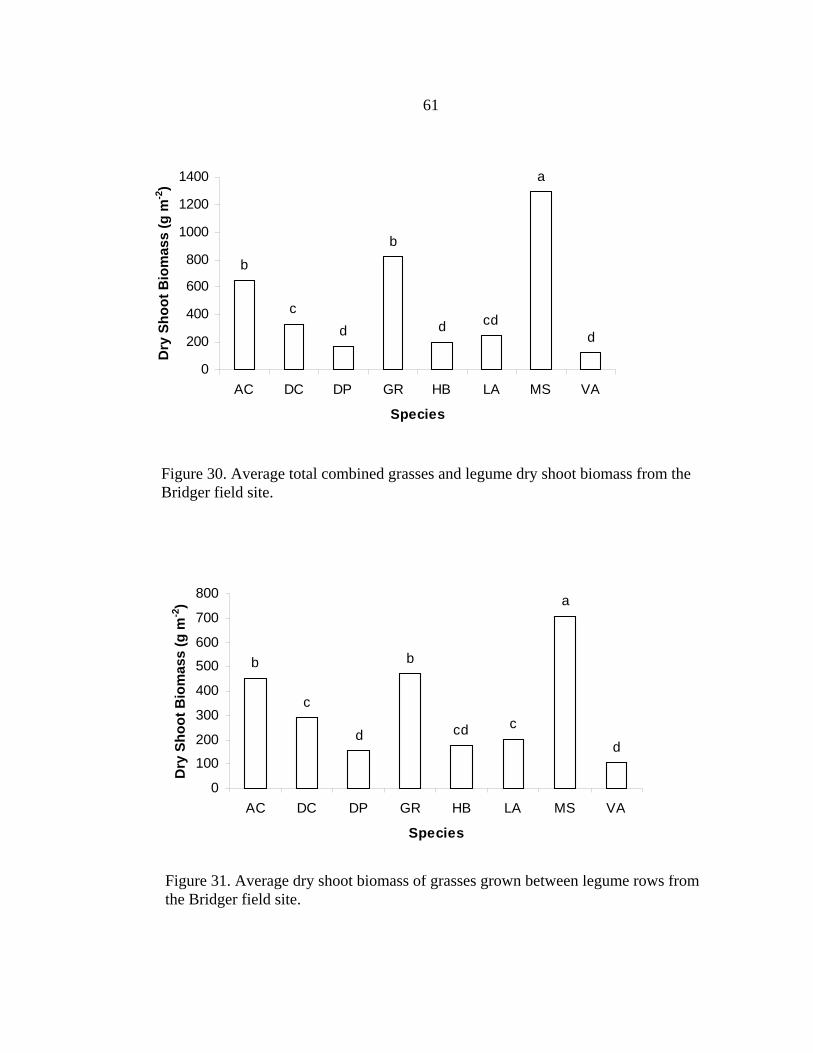
61
Figure 31. Average dry shoot biomass of grasses grown between legume rows from the Bridger field site.
c
b
d
b
cd c
a
d
0
100
200
300
400
500
600
700
800
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)
Figure 30. Average total combined grasses and legume dry shoot biomass from the Bridger field site.
d
a
cdd
b
d
b
c
0
200
400
600
800
1000
1200
1400
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)

62
The largest grass biomass grew between M. sativa rows. This was the only site
where M. sativa plots produced the largest grass biomass; at the other two sites, the
grass biomass in the native legume plots was often larger. It is possible that the deep-
rooted system of the M. sativa plants reduced competition and/or helped maintain soil
moisture and nutrient availability in the rhizosphere, which could benefit the grasses.
Summary The soil at Bridger is fine in texture and has a fairly high salinity and pH
compared to the other sites. It also had the lowest rainfall of the three sites during the
study, yet it was irrigated periodically. The combination of these environmental factors
could have led to difficulties in plant establishment. Insect damage was apparent on all
of the species at the site to varying degrees. Scat from antelope and rabbits was found
in the M. sativa, Dalea spp. and H. boreale plots and there were signs of grazing. M.
sativa was the most prolific species across the site and produced the highest legume
and grass biomasses. The natives often had smaller biomasses at this site and showed
signs of water stress and insect damage, which probably reduced biomass. The
conditions at this site were very poor; however, the survival of plants over the duration
of the study yielded valuable information on how these species respond to adverse
conditions. Many studies have noted the importance of native legume species in N-
limited, high stress conditions because their adaptive strategies and associated rhizobia
can be a primary source of N (Jacot et al. 2000; McDermott et al. 1991; Tlusty et al.
2004; Abdel-Wahab et al. 2002).

63
Moccasin
The annual precipitation at Moccasin during the 2003 season was substantially
higher than during the 2004 season. In July of 2003, biomass production appeared to
be high, however, in August, grasshoppers defoliated the site. Plants recovered fairly
well in 2004 and all seven legume species were able to persist and grow, although the
growth appeared to be reduced from the previous season (Figure 32). In the
greenhouse studies, it was found that approximately 50-60% of the N in legumes was
stored in the root material. It is possible that these plants were able to over winter by
utilizing stored materials in the root system and regenerating new aboveground
biomass. This regenerative ability may prove useful to land managers, as these
species seem to be able to tolerate fairly substantial grazing.
Figure 32. The Moccasin field site in the fall of 2003 following the defoliation of legumes and grasses by grasshoppers (left) and in the spring of 2004 following, regrowth of aboveground biomass at the site (right).

64
At Moccasin, the native legumes had the highest emergence rates of the three
sites (Figure 27). The H. boreale had over 40% emergence, which is higher than
expected for a native species in the field (McGraw et al. 2003). Emergence of the
other natives was between 20-30% with A. canadensis having the lowest rate of
emergence at just over 10%. M. sativa again had the highest level of emergence at
50%, but was not different from the H. boreale. The overall higher emergence at this
site may be due to two primary factors. First, Moccasin had fairly high soil moisture at
seeding, which should have benefited seed germination and emergence. Also, at this
site, seeds were sown into stubble rows, which offer some protection from
environmental elements to seeds and seedlings.
Legumes
A. canadensis did poorly compared to its growth at the other two sites and may
not respond as well as other species to heavy grazing (Figure 33). Astragalus spp. are
considered to be toxic, potentially causing illness and starvation in livestock, wildlife
and humans. These traits may indicate that the plant has adapted to grazing by
producing toxic substances to deter herbivores, rather than responding with aggressive
regrowth. This species is also considered to be adapted to fine textured soils (NRCS,
2004); perhaps the rocky soil texture at this site negatively affected its growth.
Some of the species that did poorly at the Post Farm and Bridger did very well
at this site. H. boreale had the largest biomass of the natives; it also flowered
extensively and some plants were beginning to form seed in July 2004. This specie
recovered very well from the defoliation, and sent out new shoots in response to

65
grazing. The morphology of the H. boreale at this site was very similar to that of M.
sativa. The D. candida and L. argenteus had the next highest biomass and both species
flowered and set some seed. D. purpurea, A. canadensis and V. americana had the
smallest biomass and only the D. purpurea flowered.
Combined Biomass
This site showed the least variation between species. Specifically, when total
biomass was combined, none of the native legume plots varied from one another. Only
the biomasses of the L. argenteus and V. americana were significantly lower than the
M. sativa (Figure 34). Even this response was minimal when compared to the growth
at the other two sites.
c
a
ccd
bc
c
d
1
10
100
1000
AC MS DC DP GR HB LA VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)
Figure 33. Average legume dry shoot biomass from the Bridger field site. Note log scale.

66
Grasses
The grasses in the D. purpurea and grass-grass plots had the highest biomass
production (Figure 35). A. canadensis, H. boreale and D. candida grass biomass were
similar to one another but smaller than the D. purpurea. Grasses grown with V.
americana and L. argenteus had the smallest biomass of the native legume plots. The
grasses in the L. argenteus plots were not different than the grasses from M. sativa
plots. M. sativa had a more aggressive growth rate and probably created more
competition for sunlight, water and nutrients than the native species did, resulting in
reduced grass biomass in the M. sativa plots.
Figure 34. Average of combined grasses and legume dry shoot biomass from the Moccasin field site.
b
a
b
abab
ab
abab
0
100
200
300
400
500
600
700
800
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)

67
Summary
Overall biomass production at Moccasin was fairly low for all the species
except H. boreale, which did fairly well at this site. Many of the plants, including the
grasses, were negatively impacted by the defoliation that occurred in August 2003.
This probably resulted in physiological stress within the plants, as well as permanently
removing the N contained in the shoot biomass from that season. The recovery of the
site was impressive. All the species were able to re-grow the following season,
although visually, biomass appeared to be substantially decreased. Again, adaptive
strategies of these native species lead to some interesting responses to a variety of
environmental stresses, and may warrant further study.
Figure 35. Average dry shoot biomass of grasses grown between legume rows from the Moccasin field site.
c
dcd
b
aa
bb
0
100
200
300
400
500
600
700
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)

68
Post Farm
Post Farm had the best growing conditions of the three sites over the course of
the study. Precipitation was adequate and there was very little grazing pressure at this
site. M. sativa again had the largest rate of emergence at about 40% while most of the
native species had approximately 20% emergence (Figure 27). A. canadensis had the
lowest emergence of all species. Less than 10% of the seeds emerged, which was less
than the emergence at the other two sites. This was the first site to be seeded; perhaps
the earlier time and lower nighttime temperatures decreased the seedling emergence.
Temperature can have a major effect on seed emergence, and lower temperatures
typically decrease the days to emergence, maximum total emergence and emergence
rates (McGraw et al. 2003).
Legumes
Biomass of M. sativa was much higher than that of the native legumes (Figure
36). A. canadensis, L. argenteus and H. boreale had the largest biomasses of the
native legumes, followed by V. americana and D. candida. D. purpurea had the
smallest biomass, perhaps due to the large grass biomass in these plots, which
appeared to cause a shade effect on the smaller, slower growing legumes. The growth
form of the plants at this site was slightly different than at the other two sites; the
plants at this site were much larger and fuller in most cases.
Nodules were difficult to locate at this site and the plants appeared to be much
more deeply rooted compared to the other sites. Only L. argenteus and A. canadensis
were verified to have nodulation. The soil surface was dry and hard during harvest and

69
it was difficult to obtain an intact root system. However, since all the native species
nodulated in Post Farm soil during the greenhouse studies, it is suspected that there
was nodulation in the field as well. The plots here were only irrigated once, late in the
first season. It is possible that the legumes sent roots deep into the soil in order to
access water. All seven legume species flowered at this site and were beginning to set
seed at harvest.
Combined biomass
A. canadensis, L. argenteus, M. sativa and the grass-grass rows had the highest
combined biomass of grasses and legumes at the Post Farm followed by D. candida, H.
boreale and V. americana (Figure 37). Interestingly, the native legume-grass plots
competed well with M. sativa-grass plots. The D. purpurea combined biomass at this
site was significantly lower than any of the other species. The combined biomass of
the native plots at this site were much larger than those of the other sites, indicating
that under good environmental conditions the native legumes are capable of producing
high amounts of biomass.
Grasses The grasses within the native legume rows produced close to 1000 g m-2 of dry
biomass, which was the highest grass biomass of the three sites (Figure 38). Grass
biomasses were similar when grown between native legume species and were higher
than the grass-grass and M. sativa plots. The grass-grass plots had an average biomass
of about 700 g m-2, while the M. sativa plots produced only half of the grass biomass

70
of the native legume plots. Due to the high biomass production at this site,
competition for resources was probably higher in the grass-grass and M. sativa plots.
Figure 36. Average dry above ground biomass of legumes from the Post Farm field site. Note log scale.
c
a
bb
a
d
b
c
1
10
100
1000
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)
Figure 37. Average dry above ground biomass of the combined grasses and legume from the Post Farm field site.
b
aa
b
a
c
a
b
0200400600800
10001200140016001800
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)
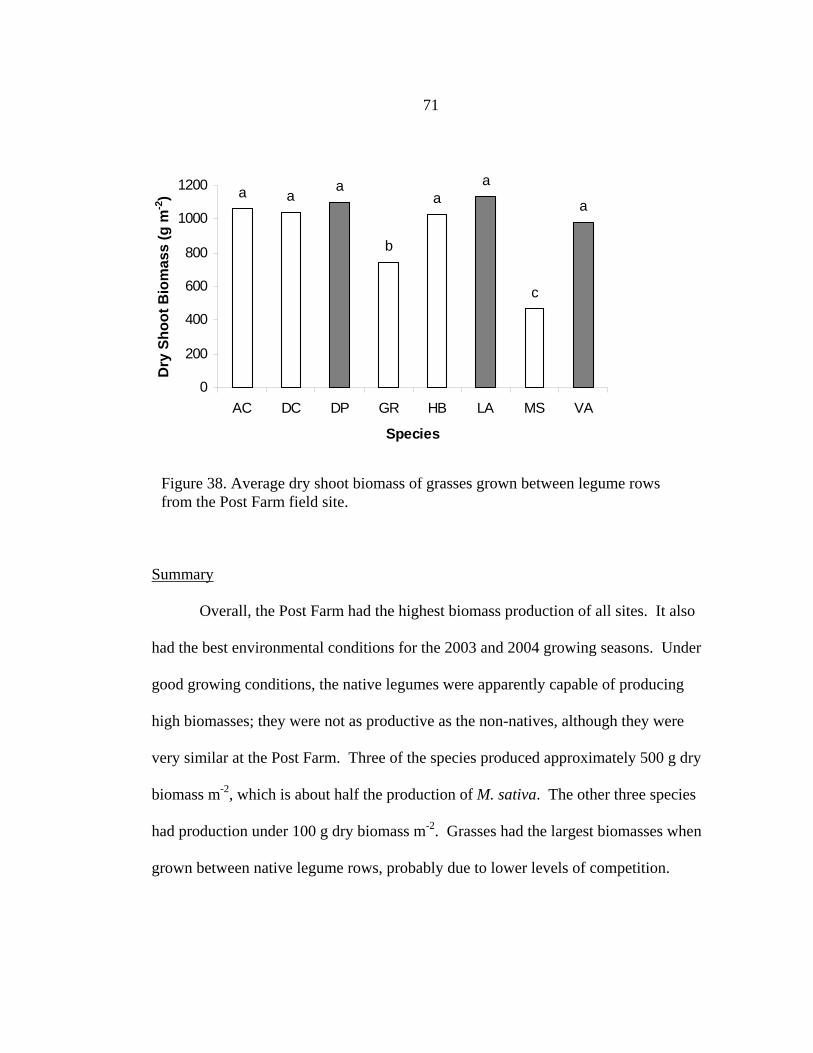
71
Summary
Overall, the Post Farm had the highest biomass production of all sites. It also
had the best environmental conditions for the 2003 and 2004 growing seasons. Under
good growing conditions, the native legumes were apparently capable of producing
high biomasses; they were not as productive as the non-natives, although they were
very similar at the Post Farm. Three of the species produced approximately 500 g dry
biomass m-2, which is about half the production of M. sativa. The other three species
had production under 100 g dry biomass m-2. Grasses had the largest biomasses when
grown between native legume rows, probably due to lower levels of competition.
Figure 38. Average dry shoot biomass of grasses grown between legume rows from the Post Farm field site.
a
c
aa
b
aa a
0
200
400
600
800
1000
1200
AC DC DP GR HB LA MS VA
Species
Dry
Sho
ot B
iom
ass
(g m
-2)

72
Nitrogen Content
The quality of a plant species as a source of N in a habitat community, as
forage or as green manure can be partially assessed by measuring the N content of the
plant tissue. From these measurements the total amount of N stored in the plant tissue
can be calculated, this information can be useful in selecting plant species for a given
environment. The N and total crude protein content of grasses has been found to be
higher in a grass-legume mixture than in single stand crops (Gökkus et al. 1999;
Sengul 2003; Tasun 1968; Whitehead 1999). There was some variation in the tissue N
content in the legumes at each site in this study (Figure 39). The legumes at Bridger
often had similar and occasionally higher N content than those at the Post Farm and
Moccasin.
At Bridger, D. purpurea had the highest tissue N content of all the species, but
was similar to M. sativa (Figure 39). L. argenteus and V. americana had the lowest
tissue N content as was expected due to lack of nodulation. The N concentrations from
the grass-grass plots at Bridger were significantly lower than the levels in the legumes
(Figure 39-40). The grasses from the M. sativa plots had the highest N concentration
while grasses grown with D. candida had the lowest (Figure 40). There were no
differences in the grasses from the other plots. The high N content of the grasses from
the M. sativa plots is likely due to the extensive root system of the species at the site.
Uptake of mineral N from soils fluctuates as factors such as plant demand, root
dispersal, surface area, volume and growth rates change; it also varies in the midst of
plant genotype and genera (Armstrong et al. 1994; Clements et al. 1993; Gregory

73
1988). N availability in the soil was high enough that the N needs of the plant could be
met without the need for N2 fixation.
The plants at Moccasin were highly variable in N content, perhaps because
there was less available N at the Moccasin site to begin with causing the plants to be
more dependent on N2 fixation (Figure 39). A. canadensis, D. purpurea and L.
argenteus had the highest N contents at this site, although the N content of M. sativa
and H. boreale were statistically similar to that of L. argenteus. D. candida and V.
americana had the lowest N contents, but were not statistically similar to one another.
The N content from the grass-grass plots at Moccasin was very low and similar to the
level at the Post Farm (Figure 39). Overall, the grasses at this site had the lowest N
content of all three sites and there was very little difference between plots (Figure 40).
The Dalea spp. at the Post Farm had significantly higher N content than the other
species, which did not differ (Figure 39). The grass-grass N content was very low, but
did not vary significantly from the grasses in the other plots (Figure 40).
The grass tissue N content varied between sites. Grasses from the M. sativa
plots at the Bridger site had the highest N content of all the grasses at each site. The
grasses had a much lower N content than the legumes and the combined average N
content of grasses from this study did not differ between legume plots (Figure 41).
This suggests that two seasons of legume growth are not sufficient to alter the N
content of the soil. The average N content of the native legumes showed no large
differences.
The N content of the grasses and soils showed no increase after two seasons of
growth, however, the N content of the legume biomass could still be beneficial. As the
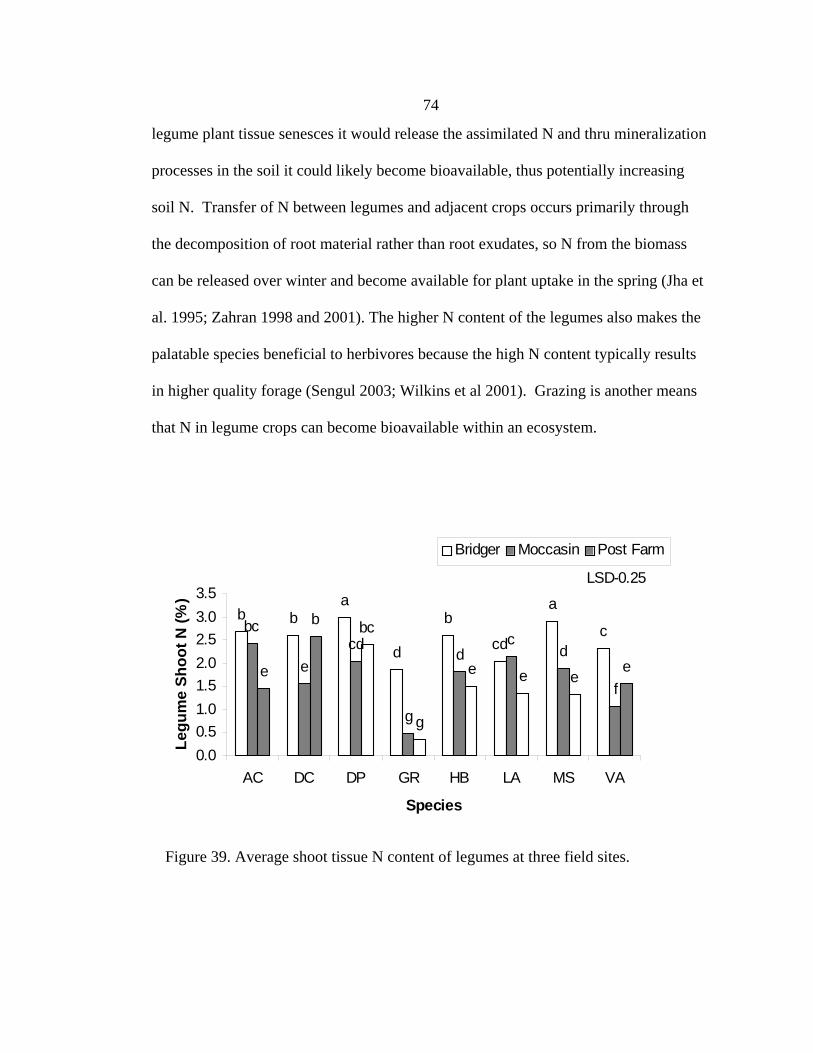
74
legume plant tissue senesces it would release the assimilated N and thru mineralization
processes in the soil it could likely become bioavailable, thus potentially increasing
soil N. Transfer of N between legumes and adjacent crops occurs primarily through
the decomposition of root material rather than root exudates, so N from the biomass
can be released over winter and become available for plant uptake in the spring (Jha et
al. 1995; Zahran 1998 and 2001). The higher N content of the legumes also makes the
palatable species beneficial to herbivores because the high N content typically results
in higher quality forage (Sengul 2003; Wilkins et al 2001). Grazing is another means
that N in legume crops can become bioavailable within an ecosystem.
Figure 39. Average shoot tissue N content of legumes at three field sites.
c
a
cd
b
d
ab b
f
dc
d
g
cde
bc
g
eeee
bcb
e
0.00.51.01.52.02.53.03.5
AC DC DP GR HB LA MS VA
Species
Legu
me
Shoo
t N (%
)
Bridger Moccasin Post Farm
LSD-0.25

75
Figure 40. Average shoot tissue N content of grasses at three field sites.
b
a
bcbbbb
deeeeeeeee
c cdc
cdcdcd
c
0.0
0.5
1.0
1.5
2.0
AC DC DP GR HB LA MS VA
Species
Gra
ss S
hoot
N (%
)Bridger Moccasin Post Farm
LSD-0.25
Figure 41. Average shoot tissue N content of legumes and grasses at three field sites.
bab
bab
c
aab ab
cc
ccccc
c
0.0
0.5
1.0
1.5
2.0
2.5
3.0
AC DC DP GR HB LA MS VA
Species
Ave
rage
Sho
ot N
(%)
Legume Grass
LSD-0.5

76
N2 fixation and N Mass Balance
N mass balances allow analysis of N changes over a season as plants grow and
senesce. Differences in the N mass balance may suggest differences in N2 fixed by the
different legumes, especially if they are higher than the grass-grass plots.
Bridger had the largest differences in N balances between species, with M.
sativa having the only positive N balance (Figure 42 and 43). This could possibly be
due to its aggressive growth at this site. With a larger root mass than the natives it
could have increased the extent of nodulation and improved this specie’s ability to fix
N2. A. canadensis had the least negative N balance at the site, indicating that it may fix
more N2 than the other native species; however, it was similar to the grass-grass plots
suggesting that N differences were more likely due to processes other than N2 fixation
(Figure 43). The Dalea spp., H. boreale and V. americana were all similar while L.
argenteus was the lowest N producer on the site but was not statistically lower than V.
americana.
The N mass balance at the Moccasin site yielded positive results for all species,
suggesting that more N2 fixation was probably occurring here than at the other sites,
although it could also be due to mineralization (Figure 42). The soil at Moccasin had
the lowest initial N concentration of the three sites; this may have increased the N2
fixation in the legumes at this site. M. sativa had a significantly larger N balance than
the native legumes. The N balances for D. purpurea and H. boreale were higher than
those of the other native species at Moccasin (Figure 44). The A. canadensis and D.

77
candida N balances were smaller than, but not statistically different from, that of H.
boreale. They also were not different than any of the other native legume species.
The M. sativa N balance was higher than for the native species at the Post Farm
(Figure 45). A. canadensis had the highest N balance of the native species at 100 kg N
ha-1. This specie also had the largest N balance of the natives at all three sites and may
be the most effective N2 fixer (Figure 42). It is realized that the differences in the N
balance could also reflect differences in N uptake from the soil below 60 cm. Deeper-
rooted species, such as M. sativa, may be capable of accessing N at a depth that the
other species cannot, resulting in higher N balances, which could indicate higher rates
of N2 fixation that are actually a result of increased deep N uptake. V. americana had
the lowest N balance at –41 kg N ha-1, although it was not different from the H.
boreale, which had an N balance near zero.
Figure 42. Total N mass balance for each plot at each sites.
g
a
g
f
de
fef
f
de
bc
decdede
cdecdecdcd
de decde
decde
ab
-ef
-400-300-200-100
0100200300400500
AC DC DP GR HB LA MS VA
Species
Plot
N B
alan
ce (k
g N
ha
-1)
Bidger Moccasin Post Farm
LSD-0.5

78
Figure 43. Bridger N mass balance.
cd
a
d
bc
b
bc
b
bc
-400-300-200-100
0100200300400500
AC DC DP GR HB LA MS VA
Species
N B
alan
ce (k
g N
ha
-1)
LSD-138
Figure 44. Moccasin nitrogen mass balance.
cd cdb
d
bc
d
a
d
020406080
100120140160180
AC DC DP GR HB LA MS VA
Species
N B
alan
ce (k
g N
ha
-1)
LSD-19

79
Field Study Summary
Striking variations can be found in total N accumulation and plant production
between individual species in any given season (Beck 1992; Schulz et al. 1999;
Unkovich and Pate 2000). This makes it difficult, if not impossible, to make direct
estimates of N2 fixation and yield in a given year. In this field study there were large
biomass variations between sites (Figure 28). Within the legumes, M. sativa had the
highest biomass at all three sites followed by A. canadensis. Grasses were better able
to compete with native legumes, whereas M. sativa was more efficient at competing
for resources. Often the biomass of the grasses was higher when they were grown in
combination with a native legume versus the M. sativa-grass plots (Figures 31, 35 and
38). At Moccasin the grass biomass from the all grass plots were similar to the grass in
Figure 45. Post Farm nitrogen mass balance.
b
b bb
b
b
a
c-100-50
050
100150200250300
AC DC DP GR HB LA MS VA
Species
N B
alan
ce (k
g N
ha
-1)
LSD-106

80
the native legume plots. At the Post Farm, the grass-grass and the M. sativa-grass plots
had the lowest grass biomass whereas at Bridger the M. sativa-grass and grass only
plots had the highest grass biomass. Biomass at the Post Farm was significantly higher
than the other two sites (Figure 46), likely due to more precipitation (Table 6).
There was no evidence of grazing by wildlife and little evidence of damage
from insects at the Post Farm, but there was some grazing by wildlife and heavy insect
damage at both Bridger and Moccasin. At Bridger and the Post Farm, A. canadensis
had the highest biomass and the highest relative estimate of N2 fixation at all three
sites. V. americana, L. argenteus and the Dalea spp. often had the lower biomasses
and lower N2 fixation estimates. V. americana and H. boreale are two of the most
palatable species; they appeared to be favored among herbivores and may have
endured heavy grazing at some sites. Any tissue that was grazed was not available for
N analysis; however, in a natural habitat any N from the legume tissue not utilized by
the herbivores is later released and becomes bioavailable in the ecosystem. H. boreale
had the highest production of the palatable natives and may prove to be important to
rangeland managers.
N uptake from the field soils was substantial over the short course of this study.
When N mass balances of the legumes were compared to those of the grass-grass plots,
it did not appear that native legumes were substantially increasing N availability. The
introduced specie, M. sativa, produced the most positive N balance at all three sites.
This species has a larger and deeper root system than the native legumes and likely has
a higher N uptake rate and demand than the native species. Due to this large root
system, M. sativa can likely take up N from deeper in the soil profile than 60 cm,

81
which was the soil sampling depth. Either of these possibilities could be expected due
to the aggressive growth of this species compared to the slower growing native species
and a low estimate of N uptake would result in a low estimate of N2 fixation.
N2 fixation estimates were largest at the Post Farm, which had higher initial N
concentration in the soil. The concentration of N within the legume tissue was much
higher than that of the grasses. However, due to the lower biomass yields, the overall
amount of N in the legumes was often lower than or comparable to the grasses. One
management strategy to harness the N content and N2 fixation potential of native
legumes is to allow the legume crop to over-winter, because small amounts of N2 are
fixed year round and as roots and nodules senesce, during the period of no growth, N
will become bioavailable for the next season’s growth (Zahran 2001). Legumes can
also be tilled into the soil and utilized as green-manure; however, a more aggressive
species such as M. sativa would be more beneficial and easier to establish for this role
versus the difficult native species (Banda et al 1994; Snapp et al. 2002). As forage,
grass-legume systems have an advantage, not only can N be transferred between
species, but these systems are often found to have increased yield and light utilization
(Brougham 1958; Haynes 1980; Ledgard 1991). The biomass production of these
systems is also very balanced for use as forage; the grasses have high production and
nutrient content in the spring whereas legumes have higher biomass production in the
summer and early fall (Brougham 1958; Mooso and Wedin 1990).

82
CHAPTER 4
LEGUME LAND MANAMENT AND ESTABLISHMENT METHODS
Introduction
In Chapter 1, the history and use of legumes as a natural source of N for crops
and soil remediation was discussed. There is a wide range of uses for legumes in order
to improve soils and increase bioavailable N for plant use. This chapter is dedicated to
land uses of legumes, the individual requirements and parameters of each species, as
well as potential benefits and drawbacks (Table 7). This will provide a reference for
land managers in order to aid in species selection and more successful plant
establishment.
M. sativa yielded higher biomasses than the natives and there may be many
cases where the most beneficial legume would be an introduced variety. However, in
land rehabilitation and habitat restoration scenarios, establishment of non-native
species is often prohibited by environmental policies. For example, introduced species
are undesirable selections for situations like National Parks or fragile habitat
restoration projects. Introduced species are often more aggressive and have a higher
demand for nutrients and water than natives, which often require less maintenance, as
they have evolved regionally and are better adapted to a semi-arid mountain
environment. In this study, the native species often had a higher or similar N
concentration to the M. sativa, meaning that they could be integrated into various
environments and utilized to improve diversity, habitat and forage quality. Legumes
assume a variety of growth forms and are typically more prostrate than grasses which

83
tend to grow more erect; in a mixture, these varied growth forms can improve light
usage and root production and minimize species competition (Haynes 1980; Mooso
and Wedin 1990; Sengul 2003). The variety in legume growth and flower morphology
can also benefit and attract many species of insects, birds and mammals. During this
study, numerous species of birds, insects and mammals were observed to have visited
and utilized the study sites.
Native legumes further benefit wildlife as forage and not only do they have a
high N and protein content but they have been shown to increase the N and crude
protein content of adjacent grasses (Brougham 1958; Haynes 1980; Ledgard 1991).
Most of the native legume species in this study showed a strong ability to re-grow
following defoliation and grazing. In the fall of the 2003 field season, grasshoppers
defoliated the plants at the Moccasin site. There were no aboveground remains of any
of the native legumes in the study area, but by spring 2004, the legumes were growing
again on the site (Figure 32), having sent out aboveground biomass from the intact root
systems.
Species Specific Information
This section will look at the individual species and go over some of their
individual characteristics. Table 7 presents basic information on the species and is
followed by a detailed section on each individual native legume species in this study.
Unless otherwise stated, information on plant characteristics not obtained from this
study was from the USDA-NRCS National Plant Database (http://plant.usda.gov).
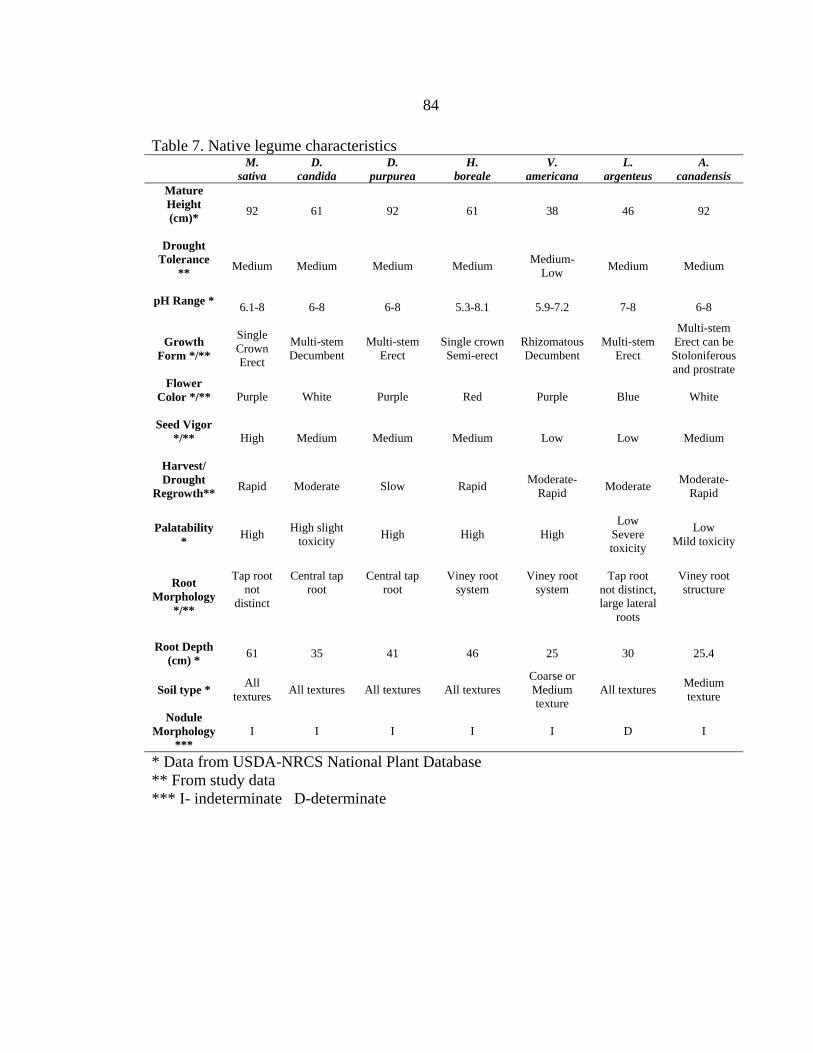
84
Table 7. Native legume characteristics
M. sativa
D. candida
D. purpurea
H. boreale
V. americana
L. argenteus
A. canadensis
Mature Height (cm)*
92 61 92 61 38 46 92
Drought Tolerance
**
Medium Medium Medium Medium Medium-Low Medium
Medium
pH Range * 6.1-8 6-8 6-8 5.3-8.1 5.9-7.2 7-8 6-8
Growth Form */**
Single Crown Erect
Multi-stem Decumbent
Multi-stem Erect
Single crown Semi-erect
Rhizomatous Decumbent
Multi-stem Erect
Multi-stem Erect can be Stoloniferous and prostrate
Flower Color */**
Purple White Purple Red Purple Blue White
Seed Vigor */**
High Medium Medium Medium Low Low Medium
Harvest/ Drought
Regrowth**
Rapid Moderate Slow Rapid Moderate-Rapid Moderate Moderate-
Rapid
Palatability * High High slight
toxicity High High High Low
Severe toxicity
Low Mild toxicity
Root Morphology
*/**
Tap root not
distinct
Central tap root
Central tap root
Viney root system
Viney root system
Tap root
not distinct, large lateral
roots
Viney root structure
Root Depth (cm) * 61 35 41 46 25 30 25.4
Soil type * All textures All textures All textures All textures
Coarse or Medium texture
All textures Medium texture
Nodule Morphology
*** I I I I I D I
* Data from USDA-NRCS National Plant Database ** From study data *** I- indeterminate D-determinate

85
A. canadensis This very hardy plant grew well and generally produced the highest biomass of
the natives throughout this study. The seed is very small like the Dalea spp. but it is
much more vigorous upon germination. Most seeds germinated within the first 14
days following planting and the seedling grew much more quickly than the other
natives in the study. The leaves have a typical pea shape, and multiple stems can reach
a height of almost one meter (Figure 46). It can be stoloniferous and prostrate, forming
new plants from underground. It has a web-like root system, which spreads throughout
the soil, and like H. boreale and V. americana, can probably be beneficial in reducing
erosion.
The flowers are large and white in color. They consist of cone shaped
flowerets that form the larger cone-like flower. The seedpods are large and upright, so
Figure 46. A. canadensis at Post Farm site Summer 2004, seedpod (left) flowering (right). Photos: S. Metcalf

86
seed production would probably not be difficult in this species. This species is
particularly attractive to birds and insects. However, it is only mildly palatable and
Astragalus spp. are actually known to be toxic to humans and animals although A.
canadensis has a lower toxicity than other Astragalus species. This species is
commonly known as “loco weed” and many livestock producers claim that after an
animal consumes it they will preferentially graze it over other feed. Many legumes
have been found to have high concentrations of certain amino acids, the type depends
on the plant species. Astragalus sp. has been used medicinally for thousands of years
as an immune system booster to be used only when the body is free of illness
(Ritchason, 1995). The toxicity of A. canadensis may have been an adaptation to
protect it from grazing.
At the Moccasin site, this species had the lowest biomass of all three sites in
2004 and was drastically reduced from the biomass it produced in 2003. This species
appears to be less capable of sending out new shoots in response to grazing compared
to the other legumes. Drought affects this species, but it is slower to show the signs of
wilting seen in the H. boreale and it recovers quickly after soil moisture is restored. It
should most likely not be planted in a drought prone areas or soils.
Dalea spp.
The prairie clovers send out a large central taproot, which can penetrate
deep into the soil and access moisture deep in the soil profile. As was seen during this
research project, the species have a fairly high drought tolerance and do well in dry
areas and well-drained soils.

87
Both species send out several stems, all of which can flower if conditions are
conducive. The D. purpurea is typically more erect than the D. candida in growth
form (Figure 47). The stalks are thin with narrow leaves. The flower is cone-shaped
and formed by many small combined flowerets. This specie is palatable to wildlife
and livestock and the flowers attract a large variety of pollinators. The D. purpurea
leaves are slightly less robust than the D. candida, although there is very little other
difference between the two species aside from flower color. Both species come from
similar seeds, which are very small, and germinated in the greenhouse studies within 7-
14 days of sowing. Germination increased with higher moisture levels during the first
month after planting. Once these plants were established, they were fairly tolerant of
low moisture levels. They also appear to be quite tolerant of low nutrient levels based
on the slow growth rate, occasionally taking two seasons to reach a mature flowering
Figure 47. D. purpurea (Left) and D. candida (Right). Photo from USDA-NRCS National Plant Database http//:plants.usda.gov.

88
stage. These species could easily be used in any land management practice, from
landscaping to wildland restoration. They would also be a good selection for crop
rotation and incorporation into grazing or hay pasture.
Due to the small seed size and slow growth rate, they have difficulty
establishing when inter-seeded with grasses and other more vigorous species.
Broadcast seeding to create a patchy landscape would be most beneficial to getting a
lasting stand of the Dalea spp. The largest specimens of these species in the field
study were at the ends of rows and in gaps where they had ample room to grow and
little competitive pressure from other plants.
H. boreale
This species has a moderately sized seed for a native species and has a
rather long germination period. Seeds in the greenhouse studies often continued to
germinate for up to three months after the planting date.
Figure 48. H. boreale from Post Farm field site summer of 2004. Flowering (left) seedpod (right).

89
H. boreale has a single crown, semi-erect growth form (Figure 48). It sends many
small roots into the soil forming a web-like structure in the soil. The root system
seems to be fairly shallow and does not appear to penetrate as deeply into the soil
profile as the taproot species. This type of root system should be extremely beneficial
in stabilizing soil and reducing surface soil erosion. It has moderate to low drought
tolerance and the leaves and stems wilt at around 55% field capacity. In the
greenhouse, it recovered quickly once soil moisture was increased and was even
capable of regenerating new aboveground biomass from the root structure, even after
the original aboveground biomass had reached permanent wilting point. No data was
located on the exact point at which the root system is no longer capable of
regenerating, but this species does possess a strong ability to regenerate. H. boreale
had the highest biomass at the Moccasin site and showed extreme tolerance to the
grazing, sending many shoots out from the root system. This native species is the most
similar to M. sativa, which is currently the one of the most utilized legumes in the
United States.
The flowers of this species are a typical pea shape and are usually red to pink in
color although there are some different varieties. The seedpod has several seeds within
it that are separately encased; this growth form may make seed harvest difficult (Figure
48). H. boreale is extremely palatable and high in protein content. In the field study,
wildlife readily grazed this species. The flowers are also very beneficial to nectivores
and pollinators; several species were observed frequenting these flowers at both the
Post Farm and Moccasin field sites.

90
L. argenteus This species is very common in Montana and can be found growing from the
open valley floors to the top of the Beartooth Pass. It has a large hard seed and
scarification from winter freeze-thaw cycles or even mild fire can improve germination
rates (Tracy and McNaughton, 1997). It had a very low germination rate and it was
often a month before the seeds germinated and many were simply not viable. This
species is moderately palatable and toxic. It can cause birth defects, such as locked
joints in the fetus of cattle and sheep if it is consumed during the spring by pregnant
animals. Again, high amino acid concentrations are suspected of hindering certain
metabolic processes.
Figure 49. L. argenteus from the Post Farm site. Seedpods (upper left), root nodules (upper right) and full row (below).

91
L. argenteus typically performs better in medium to coarse soils with fairly low
soil moisture and good drainage. It is intolerant of saturation and in moist conditions
can be highly susceptible to fungal pathogens (Saxena 1991; Cakmak and Marschner
1992; Anon 1998; Yu and Rengel 1999). The flower is a large cone shape and blue to
purple in color (Figure 49). Like many legume species, it is broken down into
flowerets and attracts many nectivores and pollinators. The plant is multi-stemmed
and erect with a large leaf and the entire plant is covered in fine silver hairs.
Although L. argenteus is very difficult to cultivate in the greenhouse, it is
remarkably hardy in the wild. It does well over a variety of habitats and elevations and
seems to establish in many harsh areas where the soils are poor. It is often a primary
colonizer in disturbed areas and is also common in open areas of climax community
forests (Tracy and McNaughton, 1996). Overall, L. argenteus is a valuable addition to
restoration seed mixes, especially in high altitude environments where species
selection is limited by extreme environmental conditions and N2 fixation is thought to
be a primary source of N (Jacot et al. 1999). It could be beneficial to further
investigate the adaptation strategies of this species and its associated bradyrhizobia, as
well as assessing N2 fixation capacities of this specie across altitudinal gradients.
V. americana This species has a medium sized seed that is round and fairly hard shelled;
scarification can improve germination. The species had a high degree of variability in
germination with some seeds germinating within seven days, but germination
continued for a two-month period. The growth form is rhizomatous and decumbent; it

92
sends out many tendril like roots and shoots that can climb and weave around other
vegetation (Figure 50).
It does not appear to harm adjacent vegetation as some vine plants can via
strangulation. The root system is diffuse and runs throughout the soil. Roots can
protrude from the soil and convert into aboveground biomass. This species has much
less drought tolerance than the other species and wilts quickly, although it can also
regenerate to some degree from the root system. In the greenhouse and growth
chamber studies, it was apparent that this specie could tolerate a higher level of soil
moisture and longer periods of saturation than the other species in the study. It can be
grown in areas that have higher precipitation and slower drainage.
V. americana is another highly palatable species that is often sought out by
livestock and wildlife. The preferential grazing of this species may make estimates of
total biomass difficult to determine, as it has an extensive root system that will
Figure 50. V. americana from the Post Farm field site. Photo: S. Metcalf

93
continue to send out aboveground biomass in response to grazing pressure. The
flowers are typical pea flowers and are small and purple in color, and the rhizomatous
growth form of this species may make seed production difficult. Birds and insects
seeking nectar often utilize this species.
Technical Management Practices for Field Establishment of Native Legumes
Legume-grass systems have been shown to enhance soil productivity and
fertility through symbiotic N2 fixation, carbon sequestration and conservation of
minerals (Snapp et al. 2002, 2002). The capacity of legumes to fix N2 symbiotically
depends on many factors and can be highly variable depending on soil and
environmental condition as well as the plant species selected. The presence of species-
specific rhizobia is another primary factor affecting N2 fixation. Soil nutrients, N
demands of the plant, moisture levels, pH and a number of other environmental factors
will influence the amount of N2 actually fixed by a certain legume. Because of all
these factors it is difficult to generalize what kind of production can be expected
from a specific legume at a site for any given year, let alone predict what kind of yield
a polyculture will produce (Beck 1992; Schulz et al. 1999; Unkovich and Pate 2000).
Regardless of inconsistencies, it is clear that native legumes are capable of forming
nodules and participating in symbiotic N2 fixation and can benefit soils in multiple
ways. Legume biomass stores a high percentage of N and as the plants senesce or are
consumed, this N will become available to other biota (Zahran 2001).
Native species are slower growing and assimilate nutrients at a slower rate than
agricultural species. Conflicting evidence has been found on legumes’ ability to

94
scavenge N before and after nodulation (Unkovich and Pate 2000). Rennie and Debutz
(1986) showed that non-nodulated and nodulated chickpeas took up the same amount
of N from the soil. Jensen (1997) found nodulated soybeans to take up less N form the
soil after nodulation, but other studies have found that nodulated N2 fixing legumes can
scavenge more N than non-nodulated legumes (George et al. 1993; Sparrow et al
1995). No studies have been found looking at the N uptake rates of these native
species. The ability of plant species to access soil N has much to do with the plant
genotype and its root growth and morphology, as well as the plant demand for N at a
given time (Clements et al. 1993; Armstrong et all 1994; Gregory 1988).
To minimize competition for resources and better utilize available N the best
strategy is careful selection of the plant species within a community (Myers et al.
1997; Crews and Peoples 2004). Selection of diverse species that have different
growth and root morphologies will improve the structure and function of a plant
community. Diversity can reduce species competition and optimize resource
utilization; it can also reduce the risk of large population losses due to pests and
disease.
For successful plant community establishment, seeding should take place in a
mosaic pattern rather than in rows. Species with a faster more aggressive growth habit,
such as grasses and M. sativa should be seeded at a lower rate than the slower growing
native legumes. Diversity will decrease the initial competition with neighboring plants
and allow the natives more space and time to reach a mature state. Broadcast seeding
is a good method of sowing seeds. This should be followed by light mulching or
raking to incorporate the seeds into the soil. Often seeding and inoculation take place

95
in the fall and both seeds and rhizobia are expected to over-winter close to the soil
surface. This can greatly reduce seedling emergence and offers a long period for
rhiziobia to persist in the soil before nodulation can occur (Graham 2004). Sowing
legume and grass seed in the early spring can increase seedling survival and decrease
the time to nodulation. The method of sowing the seeds over stubble rows at Moccasin
was highly effective in reducing weed invasion. It also may have increased seedling
emergence by providing protection against elements such as intense sun, wind, rain or
low temperatures.
Treatments such as fungicides, inocula and fertilizers can be useful in getting
plants established depending on the environmental characteristics at a site. Each of
these treatments has been shown to be either beneficial or detrimental to native legume
survival, and the use of such treatments should be determined on a site by site basis.
Fungicide treatments had a positive effect on seedling emergence and plant survival
when fungal pathogens were present and virulent in a soil. The benefits of fungicide
were most pronounced in earlier growth stages, presumably because this is the time
period when most species are more susceptible to disease. If there are no pathogens
present or if fungal species are non-virulent, the fungicides’ detrimental effects on
plant health may be more than its benefits, resulting in reduced biomass and growth
rate of plants. Fungicides do have an overall beneficial effect on the condition of root
systems, but there is evidence that fungicides may be harmful to rhizobia or interfere
with the establishment of properly functioning N2 fixation mechanisms (Bagnasco
1998). The use of fungicides in the field should be assessed at the time of seeding, as
the threat of pathogens at a given site can change each season. Once plants have

96
become established the need for fungicide is reduced as plants are more susceptible
during early growth stages.
Inoculation can be beneficial in some cases; however, in most of the soils
tested, it was unnecessary. It has been found that inoculation with indigenous rhizobia
can enhance not only plant establishment, but increase soil fertility (Zahran 2001). It
has also been found that nodulation induced by inoculation with a specific strain can be
very low in soils where endemic rhizobia are present and legumes are already known
to nodulate (McDermott et al. 1991). Inocula for native species are limited in
availability and rhizobia for most of the species cannot be obtained commercially. In
addition, many strains that are available are still experimental and not fully understood.
There are many other strains that have not yet been studied or isolated. One method of
obtaining inocula for native legumes in the field is to incorporate a native top soil
known to contain rhizobia specific to the species of interest into the seedbed as an
inocula. This method will allow for the legumes to be inoculated by species that have
adapted to the environmental conditions of the area. If a soil is lacking in species
specific rhizobia then inoculation may increase the extent of nodulation. However, in
soil that contains indigenous rhizobia the addition of inocula may create a competition
between different strains potentially hindering nodulation. Another more intensive, yet
potentially highly effective, method of obtaining inocula is to isolate rhizobia from
native plants that are well nodulated and known to be effective at N2 fixation. These
isolates can be further tested and utilized as a specific inocula; this could, however, be
a long costly process.

97
Elevated levels of N in the soil have been shown to repress nodulation, but it
has also been shown that fertilizers can increase legume production (Sengul 2003;
Unkovich and Pate 2000). The first few seasons of growth are critical to lasting plant
community establishment. If amendments such as fertilizer and fungicide benefit the
seedlings in getting established then they should be considered even if initially
nodulation is hindered. Infection and nodulation occur at the root tips, and because
plants are always generating new roots, nodulation will eventually occur as soil N is
depleted and demand for N increases with plant growth.
Controlling weeds and invasive species may be one of the most important
aspects in the first couple seasons. Many of these native plants reach a substantial size
but are slower to grow and are often out-competed by invasive species with more
aggressive growth habits. If given time to establish properly, native species will have
a better chance at lasting establishment in an area. Larger plants are better able to
compete for space and nutrients within the community and therefore have a better
chance of persisting.
Once established, these native species are hardy, require little maintenance, and
are also capable of reseeding themselves and persisting in the plant community.
Selecting species based on specific site parameters is important to successful plant
establishment and long-term survival. For example, grasses and cereal roots are
usually concentrated near the surface of the soil while legume roots grow and contract
to follow the wetting front of the soil. These differences affect N uptake (Unkovich
and Pate 2000). In some cases, a shallower rooted legume may lend more N to
adjacent grasses. However, intermixture with deeper rooted species such as H. boreale

98
or M. sativa should be utilized to access N that can build up below the root zone. Deep
rooted species are particularly useful in buffer areas of wetlands to absorb N and
prevent excessive N leaching (Crews and Peoples 2004). Legumes have also been
found to be highly beneficial in conserving N when allowed to grow over the fallow
season (Crews and Peoples 2004). They continue to fix N2 as well as scavenging some
soil N, reducing N leaching and volatilization (Crews and Peoples 2004; George et al,
1993). At planting, N from these legumes can be utilized as green-manure or they can
be mowed and grasses or other plants can be seeded over them as with other stubble
rows. The availability of N from decaying legume biomass has been found to be better
synchronized with the demand for N uptake than single and multiple applications of N
fertilizers (Becker and Ladha 1997; Gliessman 1998; Zahran 2001).
Legumes can be a great addition to a number of situations from agricultural
production to a home garden or highway median. Native legumes offer special
benefits because they are suitable for use in the Conservation Reserve Program and
plant establishment in backcountry habitat restoration. The various parameters here
are meant to help land managers with species selection and sowing methods to most
effectively establish native legumes. As with most natural systems, however, there are
no set parameters or given growth responses. There is a large degree of variability
even within a species and each plant can respond differently to certain environmental
conditions.

99
CHAPTER 5
SUMMARY AND CONCLUSIONS Overall this project was very successful. It looked at the individual yields of
six native legume species. Throughout the course of greenhouse and field studies,
factors affecting plant growth and nodulation were analyzed. Rough estimates of N2
fixation capacity were measured at three field sites in Montana.
The primary factors influencing legume response in this study were soil
characteristics such as nutrient content and microbial populations. Significant
variation occurred within species grown in different soils. Components such as soil
nutrient content (primarily N and P) and microbial populations within the soil had a
large effect on how a legume nodulates and, therefore, the amount of N that can be
fixed. Soil nutrient content and endemic microbial communities had major influences
on the growth responses of these species. Germination, emergence, biomass and
nodulation all responded differently depending on soil characteristics.
Agricultural species such as M. sativa produced higher biomass and had higher
estimated levels of N2 fixation when compared to native legumes. Native legumes
were slower growing, but are well-adapted to the semi-arid conditions of Montana and
may be very useful in land rehabilitation projects.
Legumes are able to regulate the rhizobial symbiosis in response to
physiological and environmental conditions. When moisture content was varied, there
was less variation in species growth response between moisture treatments than there
was with the soil origin. A. canadensis, H. boreale, and V. americana often increased

100
in biomass at higher soil moisture levels. The Dalea spp. was less responsive to soil
moisture changes.
Nodule mass was not as dependent on soil moisture and responded more to the
field soil characteristics. There were differences in nodule mass between species grown
in the different soils. This is logical, as the endemic rhizobia in the soil were
responsible for most of the nodulation in this study, and the introduced species from
the inocula in Trial V typically showed a lower degree of nodulation. Whether there
was competition between the endemic rhizobia and the introduced inocula, or the
legumes were not specific to the inocula strains, is not clear. Once again the
parameters that affect successful nodulation are extensive and highly dependent on
plant and rhizobia genotype and environmental conditions.
Fungicide treatments increased seed emergence in some cases and may have
improved plant survival, but overall, there were also many detrimental treatment
effects. Fungicides negatively affected nodulation in general and there was a
pronounced decrease in nodule number and mass depending on soil characteristics. It
is possible that some aspect of the N2 fixation mechanism could be affected by
fungicide applications and an apparent decrease in LHb in the nodules. The cost and
benefits of fungicides must be weighed in order to determine if the threat of pathogens
is worse than the losses to biomass and nodulation that can occur. A combination of
factors probably influences plant response. It is possible that fungicides will benefit
plants when there is risk from plant pathogens. However, the chemical stress to the
plant may be detrimental when there is no threat from pathogens. Application of

101
fungicides, as well as inocula, should be assessed on a site-by-site basis taking into
account specific environmental conditions and soil characteristics.
M. sativa had the highest biomass at all three sites as was expected. A.
canadensis had the strongest overall performance of the natives and did fairly well at
all of the sites and in the greenhouse. Biomasses of the grasses were often highest
when they were grown in combination with a native legume, although at Bridger, the
M. sativa-grass plots had the highest biomass. Biomass at the Post Farm was higher
than the other two sites and the estimated N2 fixation amounts were above average. N
uptake from the field soils was substantial over the course of the study. The N mass
balance showed that native legumes were not substantially increasing N availability
within the soil, but that a fair amount of N was assimilated in the biomass. M. sativa
produced much higher N levels than the native legumes at all three sites. N2 fixation
estimates were higher at sites with lower initial N concentration in the soil; symbiotic
N2 fixation may become more valuable to plants when soil N is limited. Legume
tissues have higher N concentrations than the grasses; therefore, native legumes would
provide higher protein forage for wildlife when used in habitat restoration, as long as
non-toxic species are selected.
Native legumes offer a variety of benefits to any setting. With a variety of
flower colors and growth habits, they offer diversity to seed mixes and can replace M.
sativa as a N source in areas where non-native species may be undesirable. It is hoped
that land managers can use the results of this study to optimizing species selection for a
range of land uses.

102
Literature Cited
Abdel-Wahab, A.B., Shabeb, M.S.A., Younis, M.A.M. 2002. Studies on the effect of salinity,
drought stress and soil type on nodule activities of Lablab purpureus (L.) sweet (Kashrangeeg). Journal of Arid Environments. 51:587-602.
Abel, G.H., Erdman J., 1964. Responses of Lee soybeans to different strains of Rhizobium
japonicum. Agronomy Journal. 56:423-424. Ahmed, B., Quilt, P., 1980. Effects of soil moisture stress on yield, nodulation and nitrogenase
activity of Macroptilium atropupureum cv. Siratro and Desmodium intortum cv. Greenleaf. Plant and Soil. 57:187-194.
Anon. Australian Water Technologies. 1998. Victorian Water Quality Monitoring Annual
Report. Department of Natural Resources and Environment, Victoria. Armstrong, E.L., J.S. Pate, J.S., Unkovich, M.J. 1994. Nitrogen balance of field pea crops in
southwestern Australia studied using the 15N natural abundance technique. Australian Journal of Plant Physiology. 21:553-549.
Bagnasco, P., De La Fuente, L., Gualtieri, G., Noya, F., Arias, A. 1998. Flurescent
Pseudomonas spp. as biocontrol agents against forage legume root pathogenic fungi. Soil Biology and Biochemistry. 30:1317-1322.
Banda, A.Z., Maghembe, J.A., Ngugi, D.N., Chome, V.A. 1994. Effect of intercropping
maize and closely spaced Leucaena hedgerows on soil conservation and maize yields on a steep slope at Ntcheu. Malawi Agroforest Systems. 27:17-22.
Becana, M., Aparicio, P., Sanchez-Diazo, M. 1986. Nitrogen fixation and leghaemoglobin
content during vegetative growth of alfalfa. Journal of Plant Physiology. 123:117-125. Beck, D. 1992. Yield and nitrogen fixation of chickpea cultivars in response to inoculation
with selected rhizobial strains. Agronomy Journal. 84:510-516. Becker, M., Ladha, J.K.,. 1997. Synchronizing residue N mineralization with rice N demand in
flooded conditions. In G. a. G. Cadish, K.E., ed. Driven by Nature. CAB International, UK. pp. 231-238.
Berg, R.K.J., Loynachan, T.E., Zablotowicz, R.M., Liebermann, M.T. 1988. Nodule
occupancy by introduced Bradyrhizobium japonicum in Iowa soils. Agronomy Journal. 80:876-881.

103
Literature Cited (continued) Bergersen, F.J. 1982. Root Nodules of Legumes: Structure and functions Research Studies
Press, Chicherster, NY. pp. 164. Broos, K., Uyttebroek, M., Mertens, J., Smolders, E. 2004. A survey of symbiotic nitrogen
fixation by white clover grown on metal contaminated soils. Soil Biology and Biochemistry. 36:633-640.
Brougham, R.W. 1958. Interception of light by foliage of pure and mixed stands of pasture
plants. Australian Journal of Agricultural Resources. 9:39-52. Busby, H.V.A. 1990. The role of bacterial surface charge in the ecology of root nodule
bacteria: and hypothesis. Soil Biology and Biochemistry. 22:1-9. Cabanes, D., Boistard, P., Batut, J. 200. Identification of Sinohrizobium meliloti Genes
Regulated during Symbiosis. Journal of Bacteriology. 182:3632-3637. Caetano-Anolles, G., Crist-Estes, D.K., Bauer, W.D. 1988. Chemotaxis of Rhizobium meliloti
to the plant flavone luteolin requires functional nodulation genes. Journal of Bacteriology. 170:3164-3169.
Clements, J., White, P., Buirchell, B. 1993. The root morphology of Lupinus angustfolius in
relation to other Lupinus species. Australian Journal of Agricultural Resources. 44:1367-1375.
Crews, T.E., Peoples, M. B. 2004. Legumes versus fertilizer sources of nitrogen: Ecological
tradeoffs and human needs. Agriculture Ecosystems and Environment. 102:279-297. Cromwell, E., Winpenny, J. 1993. Does economic reform harm the environment. Journal Of
International Development. 5:623-649. Denison, R.F., Kiers, K.T. 2004a. Why are most rhizobia beneficial to their plant hosts, rather
than parasitic? Microbes and Infection. 6:1235-1239. Denison, R.F., Kiers, K.T. 2004b. Lifestyle alternatives for rhizobia: Mutualism, parasitism,
and forgoing symbiosis. FEMS Microbiology Letters. 237:187-193. Dollhopf, D. 2000. Sampling Strategies for Drastically Disturbed Lands. Agronomy. 41:21-40.

104
Literature Cited (continued)
Durand, J.L., Sheehy, J.E., Minchin, F.R. 1987. Nitrogenase activity, photosynthesis, nodule water potential in soybean plants experiencing water deprivation. Journal of Experimental Botany. 38:311-321.
Fillery, I.R.P. 2001. The fate of biologically fixed nitrogen in legume-based dryland farming
systems: a review. Australian Journal of Experimental Agriculture. 41:361-381. Ford, C.W. 1984. Accumulation of low molecular weight solutes in water stressed tropical
legumes. Phytochemistry. 23:1007-1015. Frame, J., Newbould, P. 1984. Herbage production from grass/white clover swards. In D. J.
Thompson, ed. Forage Legumes. Occasional symposium no. 6., D. J. Thompson ed. British Grassland Society, Hurley, UK. pp. 15-35.
Frame, J., Newbould, P. 1986. Agronomy of clover. Advances in Agronomy. 40:1-88. George, T., Singleton, P.W., Vankessel, C. 1993. The use of nitrogen-15 natural abundance
and nitrogen yield of non-nodulating isolines to estimate nitrogen fixation by soybeans (Glycine max L.) across three elevations. Biology of Fertile Soil. 15:81-86.
Gökkus, A., Koc, A., Serin, Y., Comakli, B., Tan, M., Kantar, K. 1999. Hay yield and nitrogen
harvest in smooth brome grass mixtures with alfalfa and red clover in relation to nitrogen application. European Journal of Agronomy. 10:145-151.
Graham, P.H. 1963. Antibiotic Sensitivities of root-nodule bacteria. Australian Journal of
Bioscience. 15:557-559. Graham, P.H., Ballen, K.G., Montealegre, C., Jones, R.K., Fisher, B., Luque, E. 1999.
Characterization of rhizobia associated with Dalea spp. in natural prairies and revegetation areas of Minnesota. In E. M. a. G. Hernadez, ed. Highlights of nitrogen fixation research. Kluwer Academic/Plenum Publishers, NY. pp. 69-75.
Graham, P.H. 2004. Inoculated legumes and revegetation roadside plantings. Final Report.
Department of Transportation, St. Paul.
Gregory, P.J. 1988. Root Growth of chickpea, faba bean, lentil and pea and effects of water and salt stresses. In R. Summerfield, ed. World Crops: Cool Season Food Legumes. Kluwer Academic Publishers, Dordrecht. pp. 857-867.
Guerin, V., Trichant, J.C., Rigand, C. 1991. Nitrogen fixation (C2H2 reduction) by broad bean
(Vicia faba) nodules and bacteriods under water restricted conditions. Plant Physiology. 92:595-601.

105
Literature Cited (continued) Ham, G.E., Cardwell, V.B., Johnson, H.W. 1971a. Evaluations of Rhizobium japonicum
inoculants in soils containing naturalized populations of rhizobia. Agronomy Journal. 63:301-303.
Ham, G.E., Frederick, L.R., Anderson, I.C. 1971b. Serogroups of Rhizobium japonicum in
soybean nodules sampled in Iowa. Agronomy Journal. 63:69-72. Haynes, R.J. 1980. Competitive aspects of the grass-legume association. Advances in
Agronomy. 33:227-265. Helyar, K.R., Porter, W.M. 1989. Soil acidification, it's measurement and the processes
involved., p. 61-101, In A. D. Robinson, ed. Soil Acidity and Plant Growth. Academic Press, Sydney.
Hill, N.S. 1980. An evaluation of factors influenced by nitrogen fixation in sainfoin
(Onobrychis viciifolia Scop.). Masters of Science, Montana State University, Bozeman, MT.
Holdford, I.C.R. 1997. Soil phosphorus: its measurement and its uptake by plants. Australian
Journal of Soil Res. 35:227-239.
Johnson, D.A., Rumbaugh, M.D., Asay, K.H. 1981. Plant improvement of semi-arid range lands: possibilities for drought resistance and nitrogen fixation. Plant and Soil. 58:279-303.
Irigoyen, J.J., Emerich, D.W., Sanchez-Diaz, M. 1992. Water stress induced changes in
concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativa) plants. Pysiologia Plantarum. 84:55-60.
Isreal, D.W. 1993. Symbiotic dintirogen fixation and host-plant growth during development
and recovery from phosphorus deficiency. Plant Physiology. 88:294-300. Jacot, K.A., Luscher A., Nosberger, J., Hartwig, U.A. 1999. Symbiotic N2 fixation of various
legume species along an altitudinal gradient in the Swiss Alps. Soil Biology and Biochemistry. 32:1043-1052.
Jensen, E.S. 1997. The role of grain legume N2 fixation in the nitrogen cycling of temperate
cropping systems. RISO National Laboratory, Roskilde.

106
Literature Cited (continued)
Jha, P.K., Nair, S., Gopinathan, N.C., Babu, C.R. 1995. Suitability of rhizobia-inoculated wild legumes Argyrolobium flaccidum, Astragalus gravelens, Indigofera gangetica, and Lespedeza stenocarpa in providing a vegetational cover in an unreclaimed lime stone quarry. Plant Soil. 177:139-149.
Kessler, W., Boller, B.C., Nosberger, J. 1990. Distinct influences of root and shoot
temperature on nitrogen fixation by white clover. Annals of Botany. 65:341-346. Ledgard, S.F. 1991. Transfer of fixed nitrogen from white clover to associated grasses in
swards grazed by dairy cows, estimated using 15N methods. Plant Soil. 131:215-223. Lum, M.R., Hirsch, A.M. 2003. Roots and Their Symbiotic Microbes: Strategies to Obtain
Nitrogen and Phosphorus in a Nutrient-Limited Environment. Journal of Plant Growth Regulation. 21:368-382.
Marschner, H. 1995. Mineral nutrition of higher plants. London American Press, London.
McDermott, T.R., Graham, P.H. 1990. Competitive ability and efficiency in nodule formation
of strains of Bradyrhizobium japonicum. Applied Environmental Microbiology. 47:3035-3039.
McDermott, T.R., Graham, P.H., Ferrey, M.L. 1991. Competitiveness of indigenous
populations of Bradyrhizobium japonicum serocluster 123 as determined using root tip marking procedure in growth pouches. Plant and Soil. 135:245-250.
McGraw, R., Shockely, FW., Elam, TK. 2003. Effects of temperature on germination of 10
native legume species. Native Plants. 4:5-9.
Moawad, H., Schmidt, E.L. 1987. Occurrence and nature of mixed infections in nodules of field-grown soybeans (Glycine max). Biological Fertility Soils. 5:112-114.
Mooso, G.D., Wedin, W.F. 1990. Yield dynamics of canopy components in alfalfa-grass
mixtures. Agronomy Journal. 82:696-701. Myers, R.J.K., van Noordwijik, M., Vityakon, P.,. 1997. Synchrony of nutrient release and
plant demand: plant litter quality, soil environment and farmer management options., In G. a. G. Cadish, K.E., ed. Driven by Nature. CAB International, UK.
Nash, N.T., Schulman, H.M. 1976. Leghaemaglobins and nitrogenase activity during soybean
root nodule development. Canadian Journal of Microbiology. 54:2790-2729.

107
Literature Cited (continued) USDA-Natural Resources Conservation Service http://plant.usds.gov. National Plant Data
Base [Online]. 2005. Peoples, M.B., Baldock, J.A. 2001. Nitrogen dynamics of pastures: nitrogen fixation inputs,
the impact of legumes on soil fertility, and the contribution of fixed nitrogen to Australian farming systems. Australian Journal of Experimental Agriculture. 41:327-346.
Phillips, D., DeJong, T. 1984. Dinitrogen fixation in legumious crop plants., p. 121-132, In R. Hauck, ed. Nitrogen in Crop Production. ASA, CSSA, SSSA, Madisson, WI.
Provorov, N.A., Borisov, A.Y., Tikhonovich, I.A. 2002. Developmental Genetics and
Evolution of Symbiotic Structures in Nitrogen-fixing Nodules and Arbuscular Mycorrhizae. Journal of Theoretical Biology. 214:215-232.
Pudnam, A.R., Duke, W.B. 1978. Allelopathy in agro ecosystems. Annual Review
Phyotpathology. 16:431-451. Rennie, R., Debutz, S. 1986. Nitrogen-15 determined dinitrogen fixation in field-grown
chickpea. lentil, faba bean and field pea. Agronomy Journal. 78:654-660. Requena, N., Perez-Soils, E., Azcon-Aguilar C., Barea, P. 2001. Management of indigenous
plant-microbe symbioses aids to restoration of desertified ecosystems. Applied Environmental Microbiology. 67:495-498.
Ridley, A.M., Mele, P. M., Beverly, C. R. 2004. Legume-based farming in Southern Australia:
developing sustainable systems to meet environmental challenges. Soil Biology and Biochemistry. 36:1213-1221.
Ritchason, J.N.D. 1995. The Little Herb Encyclopedia Woodland Health Books, Pleasant
Grove, UT. pp.157 Ryden, J.C., Ball, P.R., Garwood, E.A. 1984. Nitrate leaching from grasslands. Nature.
311:50-53. SAS Institute Inc., 2004. JMP 5.1. Release 5.1. SAS Institute Inc., Cary, NC, USA. Saxana, A.K., Rewari, R.B. 1991. The influence of phosphate and zinc on growth nodulation
and mineral composition of chickpeas (Cicer arietinum L.) under salt stress. Microbiology and Biotechnology. 7:202-205.

108
Literature Cited (continued)
Schulz, S., Keatinge, J., Wells, G. 1999. Productivity and residual effects of legumes in rice-based cropping systems in a warm-temperature environment I. Legume biomass production and N fixation. Field Crops Research. 61:23-35.
Schulze, J. 2004. How are nitrogen fixation rates regulated in legumes? Journal of Plant
Nutrition and Soil Science. 167:125-137. Schwinning, S., Parsons, A.J. 1996a. Analysis of the coexistence mechanisms for grasses and
legumes in grazing systems. Journal of Ecology. 84:799-813.
Schwinning, S., Parsons, A.J. 1996b. A spatially explicit population model of stolonigerous N-fixing legumes in mixed pasture with grass. Journal of Ecology. 84:815-826.
Schwinning, S., Parsons A.J. 1999. The stability of grazing systems revisited: spatial models
and the role of heterogeneity. Functional Ecology. 13:737-747. Scott, B.J., Ridley, A.M., Conyers, M.K. 2000. Management of soil acidity in long-term
pastures of southeastern Australia: a review. Australian Journal of Experimental Agriculture. 40:1173-1198.
Seastedt, T.R., Knapp, A.K. 1993. Consequences of non-equilibrium resource availability
across multiple time scales: then transient maxima hypothesis. American Nature. 141:621-633.
Sengul, S. 2003. Performance of some forage grasses or legumes and their mixtures under dry
land conditions. European Journal of Agronomy. 19:401-409.
Sinclair, T.R., Muschow, R.C., Ludlow, M.M., Leach, G.J., Lawn, R.J., Foale, M.A. 1987. Field model analysis of effect of water deficits on carbon and nitrogen accumulation by soybean, cowpea, and black gram. Field Crops Research. 17:121-140.
Smil, V. 2001. Enriching the Earth MIT Press, Cambridge MA.
Snapp, S.S., Rohrbach, D.D., Simtowe, F., Freema, H.A. 2002. Sustainable soil management
for Malawi: can smallholder farmers grow more legumes? Agriculture Ecosystems and Environment. 91:159-174.
Somasegaran, P., Johnson, C.M., Cornforth, I.S. 1983. Handbook for rhizobia. Springer
Laboratory, NY. pp. 450. Sparrow, S.D., Cochran, V.L., Sparrow, E.B. 1995. Dinitrogen fixation by seven legume crops
in Alaska. Agronomy Journal. 87:34-41.

109
Literature Cited (continued)
Stockdale, E.A., Lampkin, N.H., Hovi, M., Keatinge, R., Lennartsson, E.K.M., MacDonald, D.W.. Padel, S., Tattersall. F.H., Wolfe, M.S., Watson, C.A. 2000. Agronomic and environmental implication of organic farming systems. Advances in Agronomy. 70:261-327.
Streeter J. 1998. Inhibition of legumes nodule formation and N2 fixation by nitrite. Critical
Reviews in Plant Sciences. 7:1-22. Thornley, J.H.M., Bergelson, J., Parsons, A.J. 1995. Complex dynamics in a carbon-nitrogen
model of grass-legume pasture. Annals of Botany. 75:79-94. Thornley, J.H.M., Bergelson, J., Parsons, A.J. 1998. Dynamic model of a leaf photosynthesis
with acclimation to light and nitrogen. Annals of Botany. 75:79-94.
Thornley, J.H.M. 2001. Simulation Grass-Legume Dynamics: a Phenomenological Submodel. Annals of Botany. 88:905-913.
Tlusty, B., Grossman, J.M., Graham, P.H. 2004. Selection of rhizobia for prairie legumes used
in restoration and reconstruction programs in Minnesota. Canadian Journal of Microbiology. 50:977-983.
Town, E.G., Knapp, A.K. 1996. Biomass and density responses in tall grass prairie legumes to
annual fire and topographic position. American Journal of Botany. 83:175-179.
Tracy, B.F., McNaughton, S.J. 1997. Elk grazing and vegetation responses following a late season fire in Yellowstone National Park. Plant Ecology. 130:111-119.
Trannin, W.S., Urquiaga, S., Guerra, G., Isbijijen, J., Cadisch, G. 2000. Interspecies
competition and N transfer in a tropical grass-legume mixture. Biological Fertility Soils. 32:441-448.
Unkovich, M., Pate, JS. 2000. An appraisal of recent field measurements of symbiotic N2
fixation by annual legumes. Field Crops Research. 65:211-228. Vance, C.P. 2001. Symbiotic nitrogen fixation and phosphorus acquisition. Plant nutrition in a
world of declining renewable resources. Plant Physiology. 127:390-397. Weaver, R.W., Frederick, L.R. 1974. Effect of inoculum rate on competitive nodulation of
Glycine max L. Merrill II. Field studies. Agronomy Journal. 66:233-236. Whitehead, D.C. 1995. Grassland Nitrogen. CAB International, Wallingford UK.

110
Literature Cited (continued) Williams, C.H., Donald, C.M. 1957. Changes in organic matter and pH in a podzolic soil as
influenced by subterranean clover and superphosphate. Australian Journal of Experimental Agriculture. 8.
Williams, P.M., DeMallorca, M.S. 1984. Effect of osmotically induced leaf moisture stress on nodulation and nitrogenase activity of Glycine max. Plant and Soil. 80:267-283.
Yu, Q., Renegel, Z. 1999. Micro deficiency influences plant growth and activities of
superoxide dimutases in narrow-leafed lupines. Annals of Botany. 83:175-182. Zahran, H.H. 1998. Structure of root nodules and nitrogen fixation in Egyptian wild herb
legumes. Biology Plant. 41:575-585. Zahran, H.H., Ahmad, M.S., Abdel-Fattah, M., Zaki, A.Y. 1999. Proceedings of the
International Symposium on Biological Nitrogen Fixation and Crop Production, Cairo. pp. 77-90.
Zahran, H.H. 2001. Rhizobia from wild legumes: Diversity taxonomy, ecology, nitrogen
fixation, and biotechnology. Journal of Biotechnology. 91:143-153. Zakhia, F., de Lajudie, P. 2001. Taxonomy of rhizobia. Agronomie. 21:569-576.

111
APPENDICES

112
APPENDIX A
PRELIMINARY GROWTH CHAMBER EXPERIMENTS

113
APPENDIX A
GREENHOUSE STUDIES
Introduction
The overall goal in this phase was to determine a base rate for symbiotic N2
fixation levels as well as the extent of nodulation for each of the six native legume
species. Each species was placed in a growth pouch containing a liquid plant nutrient
solution containing nutrients essential to plant growth, excluding nitrogen (McDermott
and Graham 1990). This allows measurement of the amount of N the plant is capable
of fixing. The experimental treatments consisted of inoculated and non-inoculated
plants.
Materials and Methods
Rhizobia inocula, which are known to be specific to five of the plant species,
were obtained from Dr. Peter Graham at the University of Minnesota. Inocula for H.
boreale were provided by Dr. Peter van Berkum via Patrick Elia with the Agricultural
Research Service in Maryland. Seedstock for each species was obtained from the
BPMC.
Preparation: Rhizobia contamination check
Each inocula strain was removed from the refrigerator and placed under the
sterile hood. Using a sterile wire loop, each species was streaked across a yeast
manatol agar (YMA) plate (Graham 1963).

114
Table A1: Six Legume and Ten Rhizobia Species Plant Species Inocula Type Dalea candida 6808, 6815, 7240, 7205 Rhizobia sp. Dalea purpurea 6808, 6815, 7240, 7205 Rhizobia sp. Hedysarum boreale 3174 Bradyrhizobia sp. Vicia americana 6005, 6101 Rhizobia sp. Lupinus argenteus 6003 Bradyrhizobia sp. Astragalus canadensis 6335, 6355 Rhizobia sp.
The inoculated plates were placed in an incubator at 27 ˚C for 5 d to allow the
colonies to grow. The plates were then removed from the incubator and the bacterial
growth patterns were analyzed. All rhizobia strains appeared to be free of
contamination based on ocular inspection of colony growth.
Seed sterilization methods
Seeds were all surface sterilized to remove any rhizobia or other microbes that
might interfere with the results of this experiment.
Method I
1. 95% EtOH for 30 seconds
2. 5% NaOCl Soln. 5 min
3. Distilled water, rinse 3 min, repeat 5 times
This method of sterilization was used for D. candida, D. purpurea, and A. canadensis.
H. boreale was sterilized following the above procedure except the NaOCl solution
was diluted to 3%, based on preliminary evidence of seed damage at 5%.
Method II
1. 95% EtOH for 30 seconds
2. Concentrated sulfuric acid 15 min

115
3. Distilled water rinse quick
4. Distilled water, rinse for 3 min, repeat 5 times
This method was used for V. americana and L. argenteus, the harder seeded species
(Somasegaran et al. 1983). The L. argenteus seeds which still showed bacterial
contamination after method II sterilization were further sterilized using method I.
Following sterilization, seeds were plated on YMA in Petri dishes and allowed
to germinate for 7-10 d, at which point the seedlings were at least 1.5 cm in length.
Any plates that showed signs of bacterial or fungal contamination were discarded. The
uncontaminated seeds were inoculated and placed in growth pouches.
Inocula preparation 15 ml of sterile yeast manatol broth (YMB) solution was added to an
Erlenmeyer flask. Using a flame sterilized wire loop a small sample of each rhizobia
strain was collected from a pure colony on the previously streaked plates. This sample
was added to the YMB solution. These solutions were incubated and shaken in a water
bath at 27º C for 4 d.
Planting and Inoculation Trial A1 Growth pouches, forceps and a 60 mL glass cylinder were autoclaved for 0.5
h. Two L of N-free plant growth solution were autoclaved for 0.75 h. All autoclaving
was conducted at 120ºC.
The following was performed under the sterile hood using aseptic technique.
Each growth pouch was filled with 50 mL of the plant nutrient solution. Two healthy
seedlings were removed with forceps from an uncontaminated germination Petri dish

116
and placed in the slit at the top of each growth pouch so that the root was in contact
with the moist paper within the pouch. 0.1 mL of each rhizobia culture was added to
10 mL of YMB solution. 1 mL of this dilution was added to the root tip of each
seedling. Each pouch received a total of 2 mL of inocula. Each legume species was
combined with each available rhizobia strain. Only one replication was used during
this initial trial. Following inoculation the pouches were placed in a growth chamber
set to cycle between 18 ºC and 30 ºC with a 16 h cycle of light and 8 h of darkness.
Nutrient solution and sterile DIH2O were added alternately to each pouch as needed to
maintain a moist contact between the growth pouch and the plant root system. The
plants were allowed to grow in the chamber for 4 months at which point they were
removed and harvested.
Data Collection The presence and quantity of nodules were recorded for each surviving pouch.
Plants were removed from the growth pouches and weighed, then placed in a drying
oven at 30˚ C for 7 d at which point they were removed and weighed again. The
minimum biomass required for laboratory analysis was not reached and the experiment
was terminated.
Results and Data Trial A1
Thirteen of the original forty plants survived. Nodulation occurred in the
Dalea spp. and the V. Americana. The other species did not show nodulation. The L.
argenteus does not tolerate saturation well and did not survive long enough to
nodulate. H. boreale showed root curling but did not form nodules indicating that the

117
rhizobia perhaps induced the initial biochemical signaling process and root curling
began but the infection process could not be completed. H. boreale did not nodulate in
either trial. There was only one experimental inocula for this plant specie and it may
not have been specific to H. boreale.
Trial AII
Introduction This experiment was designed to test a new protocol for growing native
legumes in a N-free setting. Native legumes are typically dryland species; it may be
that the saturated conditions within the growth pouches hindered the growth of these
Table A2. Plant survival in each pouch. Treatment Alive Nodules DC CONT + + + + DC6808 - - - - DC6815 - - - - DC7240 + - + - DC7205 + - + - DP CONT + + + + DP6808 - - - - DP6815 - - - - DP7240 - - - - DP7205 - - - - LA CONT - - - - LA6003 - - - - AC CONT - - - - AC 6335 + - - - AC 6355 - - - - VA CONT + + - - VA6005 + + - + VA6101 - - - - HB CONT + - - - HB3174 + - - - Each pouch has two healthy seedlings. ++: Signifies both seedlings either survived or nodulated, depending on the column. +-: One plant survived or nodulated --: Neither plant survived or nodulated.

118
species. Jars of sand were used in place of growth pouches in hopes that the reduced
moisture level would increase the growth rate of these native species. Preparation of
seeds and inocula followed the procedure outlined in Trial AI.
Planting and Inoculation Quart mason jars containing 480 g of sand and 50 ml of deionized water were
autoclaved for 2 h. Growth pouches, forceps and a 100 mL cylinder were autoclaved
for 0.5 h and 2 L of N-free plant growth solution was autoclaved for 0.75 h. All of the
following were performed under the sterile hood using aseptic technique. Four
seedlings were removed with forceps from the sterile Petri dish and were planted in the
sand in each jar. 0.1 mL of each rhizobia culture was added to 10 mL of YMB
solution. 1 ml of the diluted inocula was added to the sand at the base of each
seedling. Each jar received a total of 4 mL of inocula. Each jar received 40 mL of
growth solution and additional DIH2O was added so that the moisture level of the sand
in all jars was at FC by weight. Each jar was covered with autoclaved plastic and
ventilated with small perforations to allow for airflow, but limit contamination. Jars
were placed in a growth chamber set at 12 ºC to 24 ºC and having a 16-hour cycle of
light.
Maintenance Jars were weighed and nutrient solution was added to maintain FC until roots
reached the base of the jar at which point the soil moisture was maintained at 90% FC.
Plants were allowed to grow for four months attempting to reach the dry weight

119
required for N analysis (0.1g). Unfortunately this weight was not reached and the
experiment was terminated. No data were collected from this trial.
Trial AIII
Introduction This trial attempted to correct the problems that arose during the course of
Trial AI and AII. Native legume seeds are very small and slow growing compared to
the crop legumes that the original protocol was adapted from. Some of the plants also
had a yellowing burn patterns around the leaf edges as well as signs of stunted growth,
indicating that perhaps the nutrient solution was too concentrated for the slow nutrient
uptake of native plants. The changes made to the protocol follow.
Materials and Methods In Trial AII the growth solution evaporated rapidly in the growth chamber
allowing seedlings to dry out quickly. This time, pouches were sealed prior to
autoclaving, leaving only small openings for the seedlings. Seeds were allowed to
germinate in the Petri dishes for 14 d so that the roots were approximately 3 cm long.
The nutrient solution was diluted to one-quarter strength in Trial AII. Planting and
sterilization procedures followed those outlined in Trial AI. During inoculation the
rhizobia culture was not correctly diluted. 1ml of bacterial culture, instead of 0.1 ml,
was added to 10 ml of YMB solution. The pouches were placed in the same growth
chamber from Trial AI and allowed to grow for 3 months. After little growth increase,
0.05 mg L-1 of N was added to the main nutrient solution, in hopes of boosting the
growth rate of the seedlings. Each pouch received 10 mL of the N-free nutrient

120
solution as the liquid level reduced through evaporation. Again this experiment was
terminated due to insufficient biomass, however, the replicates were combined in order
to get N content of the plant tissues. The nutrient solutions from the pouches were
also analyzed for soluble NO3-N and NH4-N content.
Trial AIII Data Nine of the original 36 pouches survived and were submitted for N analysis
(Tables A3-A4). The individual biomass was again too small for analysis. The dry
weights for these plants ranged between 0.014 g and 0.077 g; 0.1 g are required for
testing so the replicates were combined. The N content for the four species that
survived, D. candida, D. purpurea, A. canadensis, and V. americana was measured
using Total Kjeldahl N analysis.
Trial AIII Results The L. argenteus in both the treatment and control group died shortly after
transfer to the growth pouch, apparently unable to survive in the saturated conditions
of the growth pouches. The other five species grew for the first few weeks, after which
the treatment group stopped growing. Nodulation occurred in both the treatment and
the control groups but after two months of little increase in growth, the leaves turned
yellow and began to fall off and all the plants in the treatment group died. The control
plants also formed nodules suggesting contamination may have occurred during the
daily watering process. Two nutrient solutions could be used in the future to reduce
risk of this occurring again. The improper dilution of the inocula may also have
increased contamination risk by increasing the number of rhizobia present by 10 fold.

121
Of the treatment group, only two V. americana plants survived, along with
sixteen plants from the control group. Many of the native species cannot tolerate
extended periods of saturation and therefore did not fare well in the growth pouches.
The native legumes were very slow growing in the growth chamber and H.
boreale and L. argenteus rarely survived the duration of the experiments. In fact, L.
argenteus usually died shortly after germination. This was probably because the
species has no tolerance for saturated conditions. The overall biomass and growth rate
Table A3. Treatments and surviving plants.
Treatment Alive NodulesDC2 + - 3 - DC3 + - 3 - DC4 + + 2 3 DP1 + + 2 2 DP4 + + 2 2 AC3 + + 3 4 AC4 + + 2 1 VA1 + + 2 2 VA2 + + 2 3 + +: 2 surviving plants +-: 1 surviving plant
Table A4. Laboratory analysis of plant tissues and nutrient solution from growth pouches. Plant Sp/Rx
Total Dry Weight (g)
Nodule Number
Plant Number
NO3-N (mg L-1)
NH4-N (mg L-1)
Tissue (%N)
ACC3 0.014 3 2 0.16 1.29 ACC 2.66 ACC4 0.041 3 2 0.22 0.38 DCC2 0.021 3 1 0.05 2.22 DCC3 0.021 4 2 1.09 0.49 DCC 3.23 DCC4 0.026 3 1 0.05 0.38 DPC1 0.033 4 2 0.06 0.26 DPC3 0.022 4 2 0.05 0.32 DPC 2.53 DPC4 0.014 4 2 0.05 0.45 VAC 0.077 2 2 0.14 1.46 VAO 2.20 VAT 0.101 3 2 0.06 0.46

122
of the native legumes were much less than that of the soybeans, an agricultural species
for which these protocols were originally designed. After three months in the growth
chamber soybeans were substantially larger than the native species (Figure A1).
Nitragin Inc. has a protocol set up for testing native species in the growth chamber. It
is similar to the sand jars in Trial II with slight modifications that reduce the moisture
level and alleviate the saturated conditions. If further research is performed on these
native species this method should be adapted for more successful completion of growth
chamber experiments.
Figure A1. Growth pouches containing N-free plant nutrient solution, native legumes (left) and soybeans (right) in the growth chamber. The native legume pouches were seeded 1 month prior to the soybeans and still had comparatively lower biomasses.

123
Biomass
The V. americana has the highest tolerance for saturated conditions. Once this
species germinated and established in the pouches it was quite hardy in the growth
chamber. The biomasses of the V. americana were the largest in most of the
greenhouse experiments. The other surviving species did not vary much and produced
approximately 0.03 g of dry biomass.
Nodulation and N Content
Typically the legumes in the growth chamber formed 2-4 nodules, which were
pink in color indicating LHb was present, and that N2 fixation was occurring. There
was little variation between species. Although the D. purpurea had the highest
average number of nodules, it was only slightly larger than the other species, which
0.000.010.020.030.040.050.060.070.080.090.10
AC DC DP VA
Species
Dry
Tot
al B
iom
ass
(g)
Figure A2. Average legume total dry biomass from growth chamber pouches.

124
averaged about 3 nodules per plant. N content was measured to analyze the extent to
which N2 fixation was occurring, although the extent of conclusions that can be drawn
is very limited due to the lack of adequate replicas.
D. candida solution contained more than twice the NO3 content of the other
four species (Figure A3). D. purpurea had the lowest level, but was fairly similar to
the remaining two species. There was less variation in the concentration of NH4 in the
pouches; all the species had about 1mg L-1 except for the D. purpurea, which only had
0.4 mg L-1. This species also had the highest number of nodules, the low N production
of this species may suggest that the nodule function of this legume-rhizobia
combination was not as effective as the other species. The N content of the plant tissue
did not vary much. V. americana had the largest biomass, so when the total N content
per plant was calculated it had the highest amount of N.
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
AC DC DP VA
Species
Nod
ule
Den
sity
(No.
pou
ch -1)
Figure A3. Average nodule number in growth chamber pouches.
0.00
0.05
0.10
0.15
0.20
0.25
0.30
0.35
0.40
0.45
AC DC DP VA
Species
NO
3-N
(mg
L-1)
Figure A3. NO3-N concentration in growth chamber pouches.
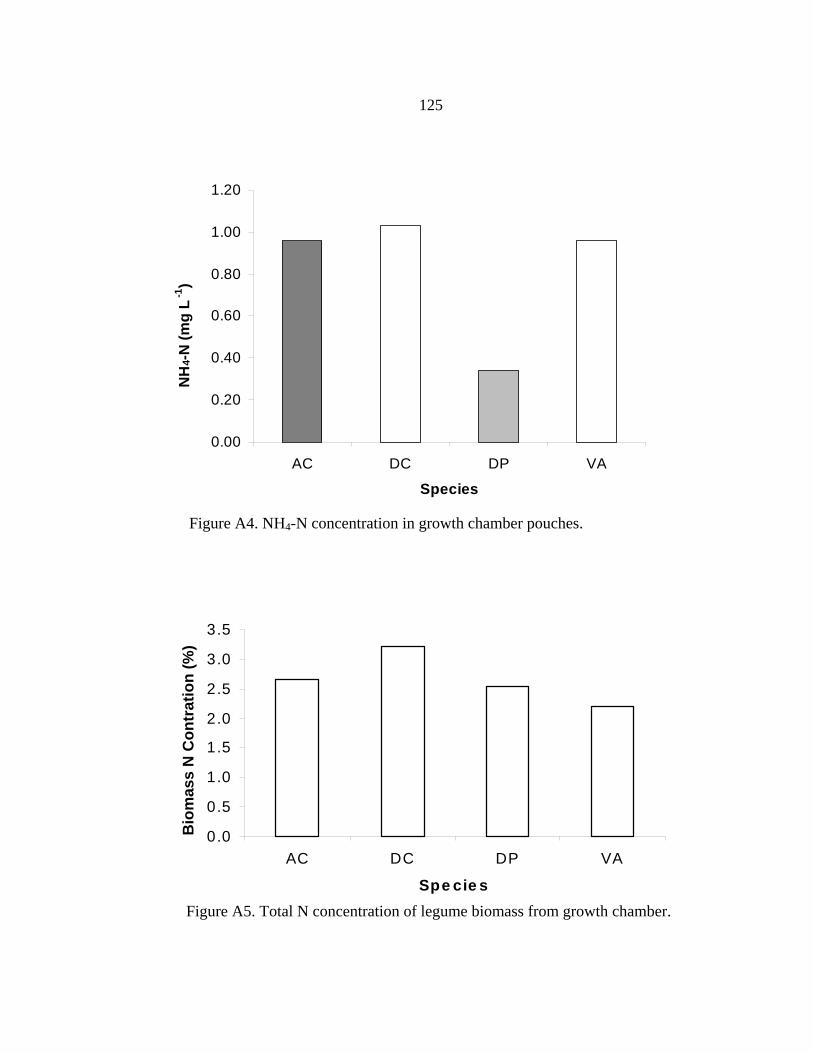
125
0.00
0.20
0.40
0.60
0.80
1.00
1.20
AC DC DP VA
Species
NH
4-N
(mg
L-1
)
Figure A4. NH4-N concentration in growth chamber pouches.
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
AC DC DP VA
Spe cie s
Bio
mas
s N
Con
trat
ion
(%)
Figure A5. Total N concentration of legume biomass from growth chamber.

126
Summary
The growth chamber experiments were highly unsuccessful, but it was apparent
that the method of testing N2 fixation potential using N-free plant nutrient solution
could be highly effective. Native species are adapted to dry, low nutrient soil and
fairly harsh environmental conditions. The proper protocol must be set up to give
these native species growing conditions that are more similar to their specialized
adaptations. Alleviation of saturated conditions and dilution of the nutrient content in
the growth solution could help these species perform better in these types of
experiments. Under the proper conditions, a more accurate measure of the true N2
fixation potential of these native species could be accomplished.
Figure A6. Average total N per legume species of legume biomass from growth chamber.
0.0000.0010.0010.0020.0020.0030.0030.0040.0040.005
AC DC DP VA
Species
Bio
mas
s N
(g N
pla
nt
-1)

127
APPENDIX B
INOCULATION RESULTS FROM GREENHOUSE TRIAL V

128
APPENDIX B
INOCULATION RESULTS FROM GREENHOUSE TRIAL V
Inoculated and Non-inoculated Results From Greenhouse Trial V
The inoculated and non-inoculated section of this had very poor plant survival
so statistics were not performed on the following data due to unequal and/or
insufficient replicas. D. candida biomass showed no response to inoculation in the
Moccasin soil (Figure B1), whereas, in the Post Farm soil, inoculation apparently
increased the shoot biomass. Biomass of D. purpurea decreased with inocula in the
Moccasin soil but slightly increased with inoculants in the Post Farm soil. H. boreale
showed no response to inoculation in the Moccasin soil but increased in biomass
substantially with inoculation in the Post Farm soil.
D. candida showed little change in nodule mass in response to inoculation in
both soils (Figure B2). The D. purpurea failed to form nodules when inoculated in the
Moccasin soil although the non-inoculated plants formed fairly large nodules. In the
Post Farm, soil inoculation may have increased the mass of the nodules in this specie.
Inoculation apparently increased H. boreale nodule weight substantially in the Post
Farm soil; however, inoculation in the Moccasin soil had essentially no effect or
perhaps an adverse effect on nodulation. The various responses to inoculation could be
due in part to competition between the introduced and endemic rhizobia, as well as the
ability of the different strains to induce nodulation in the host plants. Apparently, the
soil origin and its characteristics are of primary importance when it comes to the plant

129
growth responses to these treatments; it is likely that a combination of factors are
involved.
It is possible that fungicides will benefit plants when there is a risk of virulent
plant pathogens. However, the chemical stress to the plant may be detrimental when
there is no threat of pathogens infecting a plant. The same holds true for inoculation. If
a soil is lacking in species-specific rhizobia, then inoculation can increase the extent of
nodulation. However, the addition of inocula may create competition between the
introduced strain of rhizobia and the indigenous strain.
Figure B1. Average dry shoot biomass response of Dalea spp. and H. boreale from Trial V inoculated (I) vs. un-inoculated (U). Statistics were not performed due to unequal and/or insufficient replicas, due to poor survival.
0
1
2
3
4
5
6
7
DCI DCU DPI DPU HBI HBU
Treatment
Dry
Sho
ot B
iom
ass
(g p
ot -1
)
MoccasinPost Farm
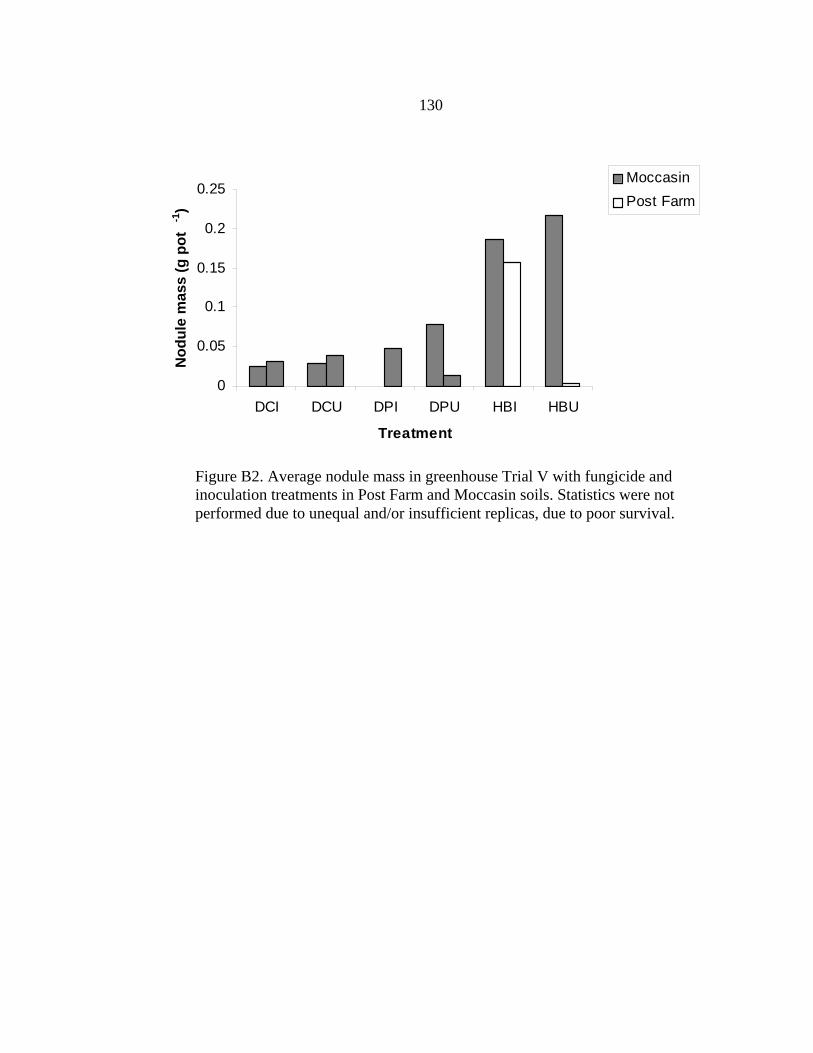
130
Figure B2. Average nodule mass in greenhouse Trial V with fungicide and inoculation treatments in Post Farm and Moccasin soils. Statistics were not performed due to unequal and/or insufficient replicas, due to poor survival.
0
0.05
0.1
0.15
0.2
0.25
DCI DCU DPI DPU HBI HBU
Treatment
Nod
ule
mas
s (g
pot
-1)
MoccasinPost Farm