suppressing effect of tannic acid on the frequencies of mutagen-induced sister-chromatid exchanges...
TRANSCRIPT

Mutation Research, 213 (1989) 195-203 195 Elsevier
MUT 04769
Suppressing effect of tannic acid on the frequencies of mutagen-induced sister-chromatid exchanges in mammalian cells *
Yfi F. Sasaki, Hisako Imanishi, Toshihiro Ohta, Mie Watanabe, Kyomu Matsumoto and Yasuhiko Shirasu
Institute of Enoironmental Toxicology, Suzuki-cho 2-772, Kodaira, Tokyo 187 (Japan)
(Received 19 December 1988) (Revision received 23 February 1989)
(Accepted 23 February 1989)
Keywords: Tannic acid; UV; Suppressing effect; SCEs; Excision repair; XP cells
Summary
The effects of tannic acid (m-galloyl gallic acid) and 7 of its analogues on the frequencies of sister-chromatid exchanges (SCEs) were investigated in cultured Chinese hamster cells. SCEs induced by UV-light or mitomycin C (MMC) were suppressed by post-treatment with tannic acid and 5 of its analogues. These effects were independent of the extension of the cell cycle. The compounds which showed an SCE-suppressing effect have a common structure of 3 neighboring hydroxy or methoxy groups substituted on the phenyl group in benzoic acid or ester.
These decreasing effects of tannic acid were observed in the G 1 phase but not in the S or G 2 phase of the cell cycle and a greater decline of the frequencies of UV-induced SCEs during liquid holding was seen in the presence of tannic acid. However, cells irradiated with X-rays were not influenced by tannic acid. In cells from a xeroderma pigmentosum (XP) patient, a Fanconi's anemia (FA) patient, and a normal human embryo, MMC-induced SCEs were also decreased by post-treatment with tannic acid. Tannic acid reduced the SCE frequencies in UV-irradiated FA and normal human cells but not in UV-irradiated XP cells.
Our results suggest that tannic acid modifies DNA-excision repair and that the decrease in the amount of unrepaired DNA damage might cause the reduction of induced SCEs.
Recently a number of antimutagens which sup- press the rate of mutations have been detected as a result of active screenings using microorganisms (Kada et al., 1986). Different modes of action of antimutagens in induced mutagenesis have been
* This paper is dedicated to the memory of Prof. Tsuneo Kada, National Institute of Genetics, Mishima (Japan).
Correspondence: 3(-fi F. Sasaki, Institute of Environmental Toxicology, Suzuki-cho 2-772, Kodaira, Tokyo 187 (Japan).
proposed in the field of bacterial genetics. For example, Ohta et al. (1983a,b,c, 1986a,b, 1988) previously demonstrated that naturally occurring flavorings such as vanillin, e thylvanil l in , anisaldehyde, cinnamaldehyde and coumarin had antimutagenic effects on bacterial cells presum- ably by promoting a recombinational repair sys- tem, while the antimutagenic effect of 5-fluoro- uracil was due to the inhibition of an SOS-repair system. Shimoi et al. (1985) also reported that tannic acid (m-galloyl gallic acid) and gallic acid,
002%5107/89/$03.50 © 1989 Elsevier Science Publishers B.V. (Biomedical Division)

196
hydrolysates of tannin which are widely present in plants and components of green tea, exhibited antimutagenic activities, probably by promotion of the excision-repair system.
On the other hand, we have been investigating the effects of the above antimutagenic factors in mammalian cells. Tannic acid exhibited a sup- pressing effect on the frequencies of chromosome aberrations induced by mitomycin C (MMC) and UV-light. Furthermore, UV-irradiated cells were shown to have a greater colony-forming ability in the presence of tannic acid (Sasaki et al., 1988). Vanillin showed suppressing effects on the num- ber of MMC-induced chromosome aberrations in vitro (Sasaki et al., 1987a) and in vivo (Inouye et al., 1988) while sister-chromatid exchange (SCE) formation was enhanced by the post-treatment with vanillin (Sasaki et al., 1987a,b). Accordingly, it may be interesting to know the effect of tannic acid on SCE formation. Hence, we have analyzed the SCE frequencies in cultured Chinese hamster cells and human cells. To find the relationship of biological effects and chemical structures, we also investigated the effects of 7 analogues of tannic acid on the frequencies of induced SCEs. Xero- derma pigmentosum (XP) is an autosomal reces- sive genetic disease with a high frequency of skin cancers resulting from exposure to sunlight. XP cells exhibit an increased sensitivity to UV and seem to have a defect in DNA repair of UV-in- duced damage (Cleaver, 1968; Arlett and Leh- mann, 1978). Fanconi's anemia is a genetic disease
with an increased sensitivity to cross-linking agents and defective in cross-linking excision repair (Fujiwara et al., 1977; Fujiwara, 1982). SCE stud- ies using cells from a normal human embryo and from DNA repair-defective patients were per- formed in order to get insight into the possible effect of tannic acid on excision repair in cultured mammalian cells.
Materials and methods
Chemical Tannic acid was obtained from Wako Pure
Chemical Industries, Osaka (Japan). Gallic acid, gallic acid methyl ester, 3,4,5-trimethoxybenzoic acid, 2,3,4-tr ihydroxybenzoic acid, 2,3,4-tri- methoxybenzoic acid, 2,4,6-trihydroxybenzoic acid and shikimic acid were purchased from Tokyo Kasei Kogyo, Tokyo (Japan). The structures of these chemicals are shown in Fig. 1. Mitomycin C (MMC), methyl methanesulfonate (MMS) and 4- nitroquinoline-l-oxide (4NQO) were obtained from Kyowa Hakko Ltd., Tokyo, Eastman Kodak, New York (U.S.A.), and Tokyo Kasei Kogyo, Tokyo, respectively.
MMS, 4NQO, tannic acid and its analogues were dissolved in dimethyl sulfoxide (DMSO, Wako Pure Chemical Industries). MMC, col- chicine (Wako Pure Chemical Industries) and 5- bromodeoxyuridine (BrdUrd, Sigma Chemical, St Louis, U.S.A.) were dissolved in Hanks' balanced salt solution (pH 7.0). The final concentration of DMSO in the medium was 0.5%.
OH
H O ~ - C O - O OH
OH HO-~COOH HO~=~-COOH OH OH
Tannic acid Gallic acid
HO OH
HO-~COOH
2,3,4-rrihydroxybenzoic acid
H3CO OCH 3
H3CO ~ COOH
2,3,4-Trimethoxybenzoic acid
OH H3CO
HO-~COOCH~ H3CO@COOH OH H3CO
Gallic acid methyl ester 3,4,5-Trimethoxybenzoic acid
OH
HO~'-COOH
OH
2,4,6-Trihydro x ybenzoic acid
Fig. 1. Structures of tannic acid and its analogues.
OH 140 ~-COOH
OH Shiklmlc acid

Cells and media F7000 human cells from a normal human em-
bryo and Ce Rel cells from a Fanconi 's anemia (FA) patient were obtained from the American Type Culture Collection (ATCC). XP3OS cells from a patient with xeroderma pigrnentosum group A were obtained from Dr. Takebe, Kyoto Univer- sity, Kyoto (Japan). Chinese hamster K-1 (CHO K- l ) cells were obtained from ATCC and cloned in our laboratory. Human cells and CHO K-1 cells were grown in Dulbecco's MEM and Ham's F12, respectively, in a humidified atmosphere with 5% CO 2 at 37 ° C. The media were supplemented with 10% fetal bovine serum (FBS), 50 I U / m l penicillin G, 50 /~g/ml streptomycin sulfate and 2.5 /~g/ml fungizon. All media and antibiotics were obtained from Flow Laboratories, Inc. (U.S.A.).
SCE assay C H O K-1 cells and human cells were seeded at
a density of 0.5-1.0 x 10 6 cel ls /100-mm dish and 0.5-1.0 x 105 cel ls /60-mm dish, respectively. All treatments were done with cells in log-phase. The cells were exposed to MMC, MMS or 4NQO for 1 h, and washed twice with Hanks ' balanced salt solution. Monolayers without medium were treated with X-rays or UV using an X-ray machine (Hitachi, Tokyo) with an adjusted dosimeter (Victreen, U.S.A.), or a 10-W germicidal lamp with a UV dosimeter (Topcon, Tokyo). Treated cells were cultured in the presence or absence of tannic acid or one of its analogues for 1 or 2 cell cycles. For the analysis of SCEs, 5-bromo- deoxyuridine (BrdUrd, final concentration 5 /~M) was added 2 cell cycles before fixation. After addition of BrdUrd, the cultures were incubated in total darkness and all operations were per- formed under a red safety light. The cells were treated with colchicine for 2 h at a final con- centration of 50 /xg/ml (CHO K-1 cells) or 0.04 /xg/ml (human cells). Preparations were processed using a modified Giemsa procedure (Sakanishi and Takayama, 1977) and harlequin-stained chro- mosomes in 50 metaphases per culture were analyzed for SCEs. The SCE data were statisti- cally analyzed using Student's t-test. Mitotic in- dices were determined and given as numbers of
197
mitotic cells per 1000 cells. In addition the num- bers of cells which had completed 2 cell cycles (M2) or less than 2 cycles (M1) were determined.
Pulse treatment with tannic acid for 3 h was done in synchronized CHO K-1 cells. Following cultivation in the presence of BrdUrd for 1 cell cycle (21 h), the cells were treated with 10 ~M TN16 (Wako Pure Chemical Industries) which has a colchicine-like effect for 20 h (Sakata, 1981). Mitotic cells were collected by mitotic shake-off, seeded at a density of 0.5-1.0 x 105 cel ls /60-mm dish, irradiated with UV at 1.2 J / m 2 or treated with MMC at a final concentration of 1.0/~M for 1 h, and cultured in fresh medium containing BrdUrd. Pulse treatments with tannic acid at a final concentration of 3.0 # g / m l were performed every 3 h following irradiation or every 3 h follow- ing treatment with MMC except for the first pulse treatment. The period of first pulse treatment following treatment with MMC was 2 h. At the end of the pulse treatment with tannic acid, cells were washed twice with Hanks ' balanced salt solu- tion and recovered in medium with BrdUrd. Since the cell cycle was not prolonged in cells treated with UV (1.2 J / m 2) or MMC (1.0 /~M for 1 h), mitotic preparations were made 21 h after mitotic shake-off. The cells were treated with colchicine for 30 min. Fifty metaphases per culture were analyzed for SCEs. To determine the duration of the different cell-cycle stages (G 1, S, and G2), the following experiment was simultaneously done with the pulse-treatment study for SCEs. M-ar- rested cells which were treated with BrdUrd for 1 cell cycle (21 h) were seeded in 60-mm dishes, treated with UV or MMC and pulse-treated with tannic acid as described above. At the end of the pulse treatments with tannic acid, cells were washed twice with Hanks ' balanced salt solution and recovered in m e d i u m without BrdUrd. The replicational banding patterns on chromosomes were examined by a method described previously (Takayama et al., 1981).
In order to block cells in the G x phase, CHO K-1 cells grown in BrdUrd-containing medium for 1 cell cycle (21 h) were incubated in an isoleucin- free medium (prepared in our laboratory) supple- mented with 10% dialyzed FBS (Gibco Laborato- ries, U.S.A.) for 36 h. Cells irradiated with UV at 2.4 J / m 2 were incubated in the presence or ab-
198
sence of tannic acid (3.0 #g /ml ) for 12 or 24 h. The cells were washed twice with Hanks' balanced salt solution and cultured in complete F12 medium
with BrdUrd and fixed 21 h later. Fifty metaphases per culture were analyzed for SCE frequencies in the first mitotic cells following G~ block.
120 ~ * * * 120 ~ , _*** 120
\ x.*** ~,~** - , . . ~ * *
i "
t J I I t I I
0 03 1 3 0 2 7 20 Tannic acid (pg/ml) Gallic acid (pg/ml)
, .... X ~ * * /
0 2 7 20 0 2 7 20 Gallic acid 3,4,5-Trimethoxy-
methyl ester (~g/ml) benzoic acid (~g/ml)
@
zu O tn
1 2 0
8 0
40
I I L I
0 2 7 2 0
2,3,4-Trihydroxy- benzoic acid (~g/ml)
120
8 0
40
, 120
80
40
" - 1 2 0
8 0
4O o
I I l l I I I I
0 2 7 2 0 0 2 7 2 0
2,3,4-Trlmethoxy- 2,4,6-Trlhydroxy-- benzoic acid ( ~ / m l l benzoic acid (~g/mll
I I ] I _
0 2 7 2 0
Sldkimic acid Irg/ml)
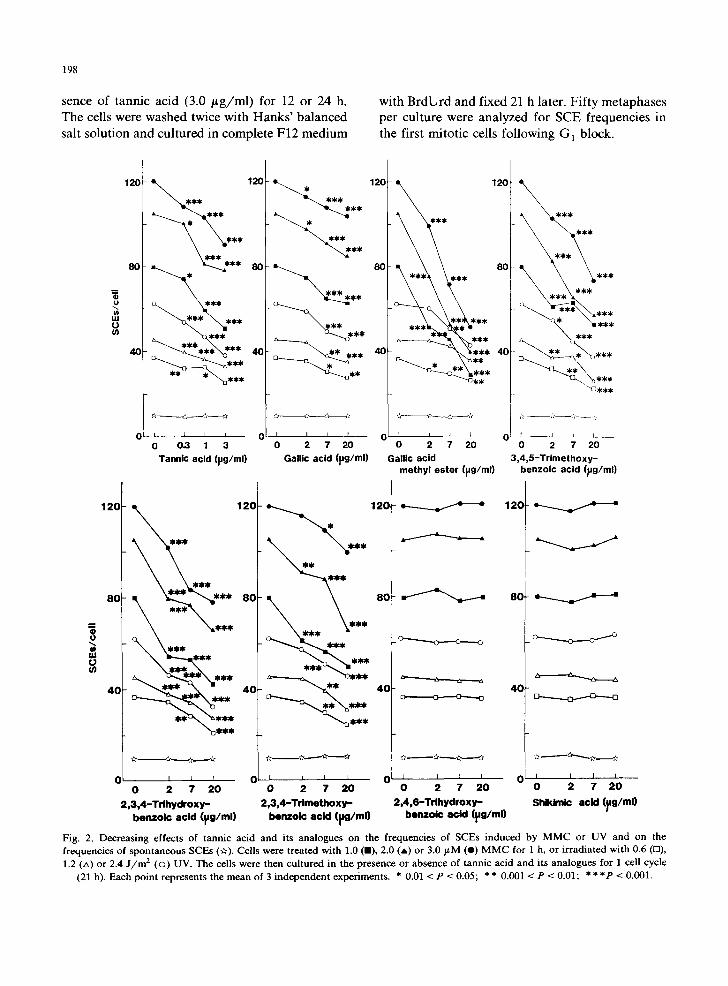
Fig. 2. Decreasing effects of tannic acid and its analogues on the frequencies of SCEs induced by MMC or UV and on the frequencies of spontaneous SCEs (if). Cells were treated with 1.0 (B), 2.0 (A) or 3.0 FM (e) M M C for 1 h, or irradiated with 0.6 (rl), 1.2 (A) or 2.4 J / m 2 (©) UV. The cells were then cultured in the presence or absence of tannic acid and its analogues for 1 cell cycle
(21 h). Each point represents the mean of 3 independent experiments. * 0.01 < P < 0.05; * * 0.001 < P < 0.01; * * * P < 0.001.

Results
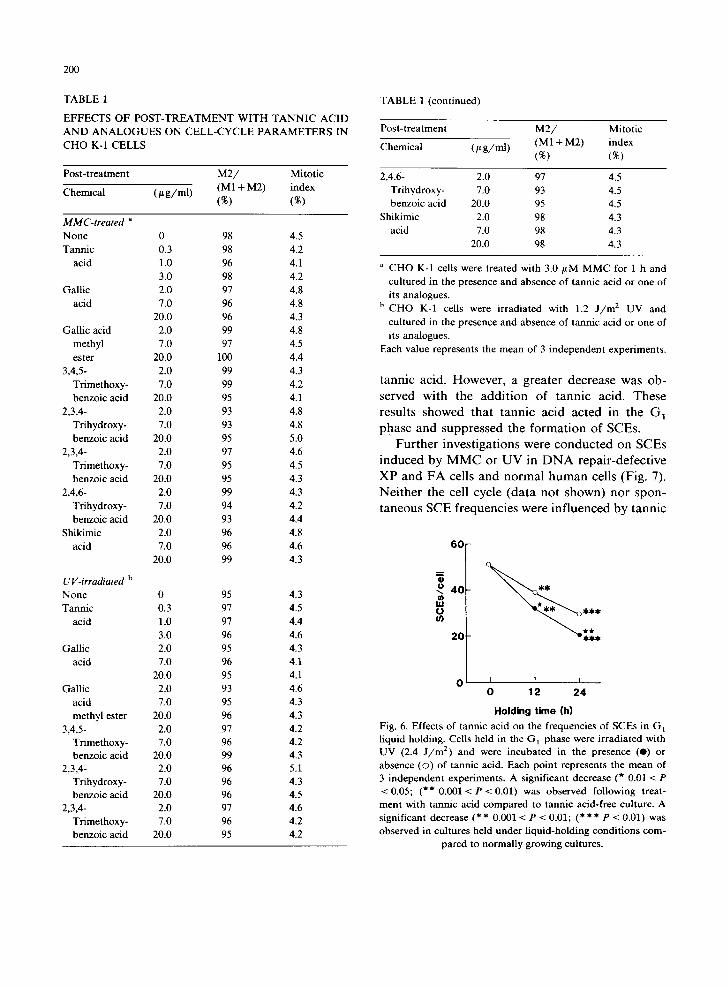
The effects of tannic acid and 7 of its analogues on MMC- or UV-induced SCEs are shown in Fig. 2. None of these chemicals affected the sponta- neous SCE frequency. Significant dose-dependent decreases in SCE frequencies were observed when cells treated with MMC or UV were cultured in the presence of tannic acid, gallic acid, gallic acid methyl ester, 3,4,5-trimethoxybenzoic acid, 2,3,4- trihydroxybenzoic acid or 2,3,4-trimethoxybenzoic acid. On the contrary, no effects on SCE frequen- cies were observed by post-treatment with 2,4,-6- trihydroxybenzoic acid or shikimic acid. As shown in Table 1, M2/(M1 + M2) and mitotic indices were not influenced by tannic acid at < 3.0 # g / m l or its analogues at __< 20/~g/ml showing that these compounds did not influence the cell cycle.
Suppressing effects of tannic acid on SCEs provoked by MMS or 4NQO were also observed (Fig. 3). Tannic acid was, however, ineffective in cells treated with X-rays (Fig. 4). As shown in Fig. 5, the frequencies of SCEs were decreased when cells were treated with 3.0 /~g/ml tannic acid in the G 1 phase but not in the S or G 2 phase. The effect of tannic acid in the G] phase was further analyzed in the liquid-holding studies. Cultured cells from rodents are known to be held in G1 by incubation in isoleucin-free medium supplemented
8O
6C ~ *** ***
o o
Ul O 4O ~ - - * * * *
20 * ~ * * *
0 I I I I 0 0.3 1 3
Tannic acid (~g/ml)
Fig. 3. Decreasing effects of tannic acid on the frequencies of SCEs induced by MMS or 4NQO. Cells were treated with 1.0 (A) or 2.0 mM (e) MMS or 0.5 (Lx) or 1.0 ~M (o ) 4NQO for 1 h. Each point represents the mean of 3 independent experi-
ments. For further explanations see Fig. 2.
199
60
4O
2O
0 t I 1 I I I 0 113 159 225 318450
X-rays (R)
Fig. 4. Absence of decreasing effects of tannic acid on SCE frequencies induced by X-rays. Cells were irradiated with X-rays and cultured with (*) or without (©) tannic acid for 1 cell cycle. Each point represents the mean of 3 independent
experiments.
with dialyzed FBS (Tobey and Ley, 1970). Fig. 6 shows the effect of tannic acid on SCE frequencies in UV-irradiated G 1 cells during liquid holding. The frequencies of UV-induced SCEs gradually decreased during liquid holding in the absence of
80
=600 * ~ O ***
\ o 4 o
• * * * ~ / °
20 Preparation UV-irradiation
I MMC-treatment Tannic acid-treatment [~[ 0 , I , o , ,
0 3 6 9 12 15 18 21 Time after tannic acid-treatment (h)
Fig. 5. Effects of tannic acid at different cell stages on the frequencies of SCEs induced by UV or MMC. Synchronized cells were irradiated with 1.2 J / m x UV (©) or treated with 1.0 v M MMC (e) and pulse-treated with 3.0 t t g /ml tannic acid for 3 h. Each point represents the mean of 3 independent experiments with duplicate cultures. For further explanations
see Fig. 2.
200
TABLE 1
EFFECTS OF POST-TREATMENT WITH TANNIC ACID AND ANALOGUES ON CELL-CYCLE PARAMETERS IN CHO K-1 CELLS
Post-treatment
Chemical (/~ g /ml)
M 2 / Mitotic (M1 + M2) index (%) (%)
M M C - t r e a t e d a
None 0 98 4.5 Tannic 0.3 98 4.2
acid 1.0 96 4.1 3.0 98 4.2
Gallic 2.0 97 4.8 acid 7.0 96 4.8
20.0 96 4.3 Gallic acid 2.0 99 4.8
methyl 7.0 97 4.5 ester 20.0 100 4.4
3,4,5- 2.0 99 4.3 Trimethoxy- 7.0 99 4.2 benzoic acid 20.0 95 4.1
2,3,4- 2.0 93 4.8 Trihydroxy- 7.0 93 4.8 benzoic acid 20.0 95 5.0
2,3,4- 2.0 97 4.6 Trimethoxy- 7.0 95 4.5 benzoic acid 20.0 95 4.3
2,4,6- 2.0 99 4.3 Tfihydroxy- 7.0 94 4.2 benzoic acid 20.0 93 4.4
Shikimic 2.0 96 4.8 acid 7.0 96 4.6
20.0 99 4.3
UV- i r rad ia t ed b
None 0 95 4.3 Tannic 0.3 97 4.5
acid 1.0 97 4.4 3.0 96 4.6
Gallic 2.0 95 4.3 acid 7.0 96 4.1
20.0 95 4.1 Gallic 2.0 93 4.6
acid 7.0 95 4.3 methyl ester 20.0 96 4.3
3,4,5- 2.0 97 4.2 Trimethoxy- 7.0 96 4.2 benzoic acid 20.0 99 4.3
2,3,4- 2.0 96 5.1 Trihydroxy- 7.0 96 4.3 benzoic acid 20.0 96 4.5
2,3,4- 2.0 97 4.6 Trimethoxy- 7.0 96 4.2 benzoic acid 20.0 95 4.2
TABLE 1 (continued)
Post-treatment
Chemical (/t g /ml )
M 2 / (M1 + M2) (%)
Mitotic index (%)
2,4,6- 2.0 97 4.5 Trihydroxy- 7.0 93 4.5 benzoic acid 20.0 95 4.5
Shikimic 2.0 98 4.3 acid 7.0 98 4.3
20.0 98 4.3
a CHO K-1 cells were treated with 3.0/LM MMC for 1 h and cultured in the presence and absence of tannic acid or one of its analogues.
h CHO K-1 cells were irradiated with 1.2 J / m : UV and cultured in the presence and absence of tannic acid or one of its analogues.
Each value represents the mean of 3 independent experiments.
tannic acid. However, a greater decrease was ob- served with the addition of tannic acid. These results showed that tannic acid acted in the G~ phase and suppressed the formation of SCEs.
Further investigations were conducted on SCEs induced by MMC or UV in D N A repair-defective XP and FA cells and normal human cells (Fig. 7). Neither the cell cycle (data not shown) nor spon- taneous SCE frequencies were influenced by tannic
60
Q
o 40 U) ILl 0 (n
2 0 " - . : , ; ,
O I r i 0 12 24
Holding time (h)
Fig. 6. Effects of tannic acid on the frequencies of SCEs in G] liquid holding. Cells held in the G 1 phase were irradiated with UV (2.4 J / m 2) and were incubated in the presence (e) or absence (©) of tannic acid. Each point represents the mean of 3 independent experiments. A significant decrease (* 0.01 < P < 0.05; (** 0.001 < P < 0.01) was observed following treat- ment with tannic acid compared to tannic acid-free culture. A significant decrease (* * 0.001 < P < 0.01; (* * * P < 0.01) was observed in cultures held under liquid-holding conditions com-
pared to normally growing cultures.
201
o
o Ih
2 0
XP ce l l s 0 I L I
0 0 . 3 1 3 T a n n i c ac id (pg/ml)
2G
10 ~ _ ~ **
FA ce l l s 0 I I i i
0 0 . 3 1 3 Tann i c ac id (pg/ml)
2 0
15
Normal ce l l s 0 i i i I
0 0 . 3 1 3 Tannic acid ( ~g /m l )
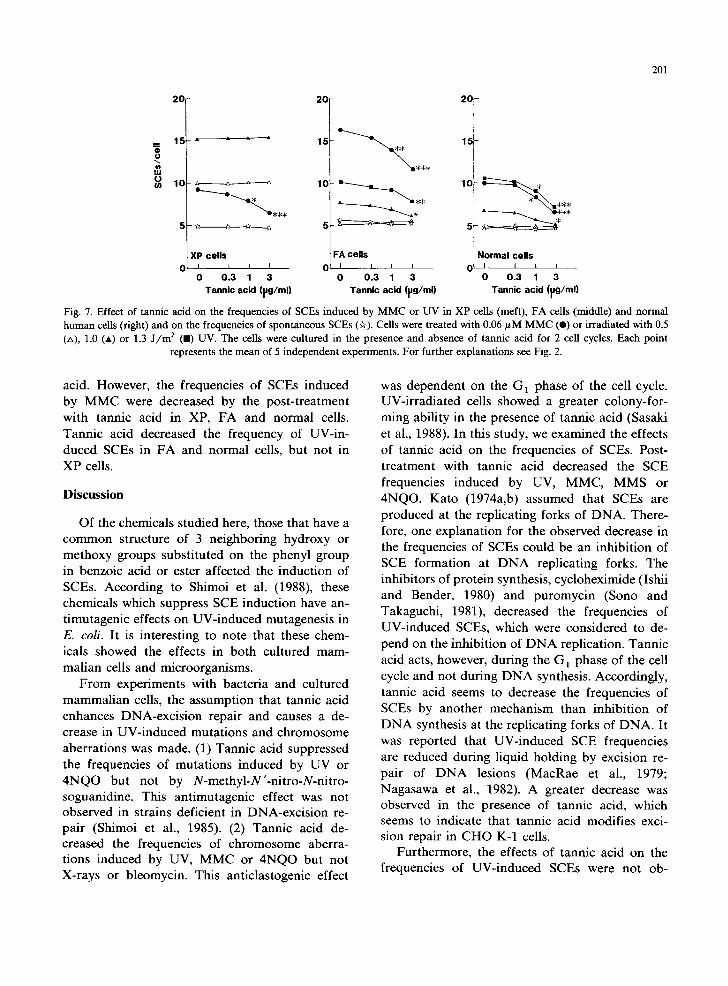
Fig. 7. Effect of tannic acid on the frequencies of SCEs induced by MMC or UV in XP cells (meft), FA cells (middle) and normal human cells (fight) and on the frequencies of spontaneous SCEs (~r). Cells were treated with 0.06 #M MMC (e) or irradiated with 0.5 (zx), 1.0 (A) or 1.3 J / m 2 ( i ) UV. The cells were cultured in the presence and absence of tannic acid for 2 cell cycles. Each point
represents the mean of 5 independent experiments. For further explanations see Fig. 2.
acid. However, the frequencies of SCEs induced by MMC were decreased by the post-treatment with tannic acid in XP, FA and normal cells. Tannic acid decreased the frequency of UV-in- duced SCEs in FA and normal cells, but not in XP cells.
Discussion
Of the chemicals studied here, those that have a common structure of 3 neighboring hydroxy or methoxy groups substituted on the phenyl group in benzoic acid or ester affected the induction of SCEs. According to Shimoi et al. (1988), these chemicals which suppress SCE induction have an- timutagenic effects on UV-induced mutagenesis in E. coli. It is interesting to note that these chem- icals showed the effects in both cultured mam- malian cells and microorganisms.
From experiments with bacteria and cultured mammalian cells, the assumption that tannic acid enhances DNA-excision repair and causes a de- crease in UV-induced mutations and chromosome aberrations was made. (1) Tannic acid suppressed the frequencies of mutations induced by UV or 4NQO but not by N-methyl-N'-nitro-N-nitro- soguanidine. This antimutagenic effect was not observed in strains deficient in DNA-excision re- pair (Shimoi et al., 1985). (2) Tannic acid de- creased the frequencies of chromosome aberra- tions induced by UV, MMC or 4NQO but not X-rays or bleomycin. This anticlastogenic effect
was dependent on the G 1 phase of the cell cycle. UV-irradiated cells showed a greater colony-for- ming ability in the presence of tannic acid (Sasaki et al., 1988). In this study, we examined the effects of tannic acid on the frequencies of SCEs. Post- treatment with tannic acid decreased the SCE frequencies induced by UV, MMC, MMS or 4NQO. Kato (1974a,b) assumed that SCEs are produced at the replicating forks of DNA. There- fore, one explanation for the observed decrease in the frequencies of SCEs could be an inhibition of SCE formation at D N A replicating forks. The inhibitors of protein synthesis, cycloheximide (Ishii and Bender, 1980) and puromycin (Sono and Takaguchi, 1981), decreased the frequencies of UV-induced SCEs, which were considered to de- pend on the inhibition of D N A replication. Tannic acid acts, however, during the G] phase of the cell cycle and not during D N A synthesis. Accordingly, tannic acid seems to decrease the frequencies of SCEs by another mechanism than inhibition of D N A synthesis at the replicating forks of DNA. It was reported that UV-induced SCE frequencies are reduced during liquid holding by excision re- pair of D N A lesions (MacRae et al., 1979; Nagasawa et al., 1982). A greater decrease was observed in the presence of tannic acid, which seems to indicate that tannic acid modifies exci- sion repair in CHO K-1 cells.
Furthermore, the effects of tannic acid on the frequencies of UV-induced SCEs were not ob-

202
se rved in D N A excis ion r epa i r -de f i c i en t X P cells,
whi le S C E s were r educed in M M C - t r e a t e d X P
cells. U V - i r r a d i a t i o n induces p y r i m i d i n e d i m e r s
a n d M M C induces base adduc t s . These D N A
les ions are c o n s i d e r e d to be r e spons ib l e for the
f o r m a t i o n o f S C E s ( K a t o , 1974b; La t t et al., 1975;
C a r r a n o et al., 1977). P y r i m i d i n e d imer s have been
r e p o r t e d to be r e p a i r e d by exc is ion repa i r and n o t
to be r epa i r ed in X P cells (Cleaver , 1968). H o w -
ever , base adduc t s p r o v o k e d by M M C are exc i sed
by a repa i r p rocess d i f f e ren t f r o m tha t of U V
d a m a g e . F A cells a re de fec t ive in the r epa i r sys-
t em of M M C les ions ( F u j i w a r a et al., 1977;
Fu j iwa ra , 1982). T h e r e f o r e ou r p r e sen t d a t a sup-
p o r t the h y p o t h e s i s tha t t ann i c ac id mod i f i e s
D N A - e x c i s i o n repa i r in c u l t u r e d m a m m a l i a n cells.
C o n s e q u e n t l y r epa i r of D N A les ions w o u l d l ead
to the o b s e r v e d dec rease in the f r equenc ie s of
SCEs . X- rays i n d u c e D N A - s t r a n d b reaks w h o s e
r epa i r is d i f f e r en t f r o m tha t o f U V d a m a g e (F r i ed -
be rg et al., 1979). I f t a n n i c ac id m o d i f i e s exc is ion
repai r , ou r o b s e r v a t i o n tha t the f r equenc ie s o f
X - r a y - i n d u c e d S C E s were no t i n f l u e n c e d by t a n n i c
ac id in C H O K-1 cel ls w o u l d be wel l exp la ined .
T a n n i c ac id is a hyd ro ly t i c p r o d u c t o f t a n n i n
tha t is wide ly p re sen t in p lants , i n c l u d i n g g reen
tea. A n a n t i c a r c i n o g e n i c p o t e n t i a l of g reen tea has
b e e n r e p o r t e d (Ogun i and Nasu , 1988). O u r pres-
en t d a t a migh t h a v e s o m e re l a t ion wi th the an t i -
c a r c i n o g e n i c ef fec t o f g reen tea.
Acknowledgements
T h e au tho r s are g ra te fu l to Prof . H i r a k u Takebe ,
K y o t o Unive r s i ty , K y o t o ( Japan) , for suppl~,ing
cells f r o m pa t i en t X P 3 O S , and to K a y o k o Sh imoi ,
S h i z u o k a Co l l ege o f P h a r m a c e u t i c a l Sc ience ,
S h i z u o k a ( Japan) , for her v a l u a b l e advice .
References
Arlett, C.F., and A.R. Lehmann (1978) Human disorders show- ing increased sensitivity to the induction of genetic damage, Annu. Rev. Genet., 12, 95-115.
Carrano, A.V., L.H. Tompson, D.G. Stetka, J.L. Minkler, J.A. Mazrimas and S. Fong (1977) DNA crosslinking, sister chromatid exchange and specific-locus mutation, Mutation Res., 63, 175-188.
Cleaver, J.E. (1968) Defective repair replication in xeroderma pigmentosum, Nature (London), 218, 652-656.
Friedberg, E.C., U.K. Ehmann and J.l. Williams (1979) Hu- man disease associated with defective DNA repair, Adv. Radiat. Biol., 8, 85-174.
Fujiwara, Y. (1982) Defective repair of mitomycin C crosslinks in Fanconi's anemia cells and loss in confluent normal human and xeroderma pigmentosum cells, Biochim. Bio- phys. Acta, 699, 217-225.
Fujiwara, Y., M. Tatsumi and M.S. Sasaki (1977) Cross-linking repair in human cells and its possible defect in Fanconi's anemia cells, J. Mol. Biol., 113, 634-649.
Inouye, T., Y.F. Sasaki, H. lmanishi, M. Watanabe, T. Ohta and Y. Shirasu (1988) Suppression of mitomycin C-induced micronuclei in mouse bone marrow cells by post-treatment with vanillin, Mutation Res., 202, 93-95.
Ishii, Y., and M.A Bender (1980) Effects of inhibitors of DNA synthesis on spontaneous and ultraviolet light-induced sis- ter chromatid exchanges in Chinese hamster cells, Mutation Res., 79, 19-32.
Kada, T., T. Inoue, T. Ohta and Y. Shirasu (1986) Antimuta- gens and their modes of action, in: D.M. Shankel et al. (Eds.), Plenum, New York, pp. 181-196.
Kato, H. (1974a) Possible role of DNA synthesis in function of sister chromatid exchanges, Nature (London), 252, 739-741.
Kato, H. (1974b) Photoreactivation of sister chromatid ex- changes induced by ultraviolet irradiation, Nature
" (London), 252, 552-553. Latt, S.A., G. Stetten, L.A. Juergens, G.R. Buchanan and P.S.
Gerald (1975) Induction by alkylating agents of sister chro- matid exchanges and chromatid breaks in Fanconi's anemia, Proc. Natl. Acad. Sci. (U.S.A.), 72, 4066-4070.
MacRae, W.D., E.A. MacKinnon and H.F. Stich (1979) The fate of UV-induced lesions affecting SCEs, chromosome aberrations and survival of CHO cells arrested by depriva- tion of arginine, Chromosoma, 72, 15-22.
Nagasawa, H., A.J. Fornace Jr., M.A. Ritter and J.B. Little (1982) Relationship of enhanced survival during confluent holding recovery in ultraviolet-irradiated human and mouse cells to chromosome aberrations, sister chromatid ex- changes, and DNA repair, Radiat. Res., 92, 483-496.
Oguni, I., and K. Nasu (1988) Epidemiological and physiologi- cal studies on the antitumor activity of the fresh green tea leaf, in: Tea Research Institute (Ed.), Proceeding of Inter- national Tea-Quality-Human Health Symposium, Chinese Academy of Agricultural Sciences, Hangghow, pp. 222-226.
Ohta, T., K. Watanabe, M. Moriya, Y. Shirasu and T. Kada (1983a) Antimutagenic effects of cinnamaldehyde on chem- ical mutagenesis in Escherichia coli, Mutation Res., 107, 219-227.
Ohta, T., K. Watanabe, M. Moriya, Y. Shirasu and T. Kada (1983b) Antimutagenic effects of coumarin and umbel- liferone on mutagenesis induced by 4-nitroquinoline 1-oxide or UV irradiation in E. coli, Mutation Res., 117, 135-138.
Ohta, T., K. Watanabe, M. Moriya, Y. Shirasu and T. Kada (1983c) Analysis of antimutagenic effect of cinnamalde- hyde on chemically induced mutagenesis in Escherichia coli, Mol. Gen. Genet., 192, 309-315.
Ohta, T., M. Watanabe, K. Watanabe, Y. Shirasu and T. Kada (1986a) Inhibitory effects of flavourings on mutagenesis

induced by chemical in bacteria, Fd. Chem. Toxicol., 24, 51-54.
Ohta, T., M. Watanabe, R. Tsukamoto, Y. Shirasu and T. Kada (1986b) Antimutagenic effects of 5-fluorouracil and 5-fluorodeoxyuridine on UV-induced mutagenesis in Escherichia coli, Mutation Res., 173, 19-24.
Ohta, T., M. Watanabe, Y. Shirasu and T. Inoue (1988) Post-replication repair and recombination in uvrA umuC
strains of Escherichia coli are enhanced by vanillin, an antimutagenic compound, Mutation Res., 201, 107-112.
Sakanishi, S., and S. Takayama (1977) A simple Giemsa method for the differential staining of sister chromatids with a note on the presumptive mechanisms involved, Proc. Japan Acad., 53, 143-146.
Sakata, Y. (1981) New reagents for synchronized culture, Tis- sue Cult., 7, 309-311 (in Japanese).
Sasaki, Y.F., H. Imanishi, T. Ohta and Y. Shirasu (1987a) Effects of vanillin on sister chromatid exchanges and chro- mosome aberrations induced by mitomycin C in cultured Chinese hamster ovary cells, Mutation Res., 191, 193-200.
Sasaki, Y.F., H. Imanishi, T. Ohta and Y. Shirasu (1987b) Effects of antimutagenic flavourings on SCEs induced by chemical mutagens in cultured Chinese hamster cells, Mu- tation Res., 189, 313-318.
203
Sasaki, Y.F., H. Imanishi, T. Ohta, M. Watanabe, K. Matsumoto and Y. Shirasu (1988) Suppressing effect of tannic acid on UV and chemically induced chromosome aberrations in cultured mammalian cells, Agric. Biol. Chem., 52, 2423-2428.
Shimoi, K., Y. Nakamura, I. Tomita and T. Kada (1985) Bio-antimutagenic effects of tannic acid on UV and chem- ically induced mutagenesis in Escherichia coli B/r , Muta- tion Res., 149, 17-23.
Shimoi, K., Y. Okada, I. Tomita, T. Matsunaga and O. Nikaido (1988) Effects of gallic acid derivatives on UV-in- duced mutagenesis and DNA repair in Escherichia coil B/r, J~ Radiat. Res., 29, 73.
Sono, A., and K. Sakaguchi (1981) Inhibition of protein synthesis antagonizes induction of sister chromatid ex- changes by exogenous agents, Mutation Res., 80, 121-131.
Takayama, S., T. Taniguchi and Y. Iwashita (1981) Applica- tion of the 4NA-EDTA Giemsa method for analysis of DNA replication pattern, Chromosome Inf. Serv., 31, 36-38.
Tobey, R.A., and K.D. Ley (1970) Regulation of initiation of DNA synthesis in Chinese hamster cells, J. Cell Biol., 46, 151-157.