supplemental information an iterative, bimodular nonribosomal peptide synthetase …€¦ · ·...
TRANSCRIPT

1
Chemistry & Biology, Volume 20
Supplemental Information
An Iterative, Bimodular Nonribosomal Peptide
Synthetase that Converts Anthranilate
and Tryptophan into Tetracyclic Asperlicins
Xue Gao, Wei Jiang, Gonzalo Jiménez-Osés, Moon Seok Choi, Kendall N. Houk, Yi Tang,
and Christopher T. Walsh
Inventory of Supplemental Information Table S1: List of primers used during this study; Figure S1, related to Figure 2: Michaelis-Menten plots of ATP-[32P]PPi exchange assay for AspA. Figure S2: SDS-PAGE gels of the heterologously expressed proteins in this study. Figure S3, related to Figure 3: Extracted ion mass chromatograms [M+H]+ =407 from AspA in vitro reactions. Figure S4, related to Figure 3: In vitro reconstitution of the Asperlicin C, D and 1 production by using dissected AspA_M1 and AspA_M2 proteins. Figure S5, related to Figure 4: The first module of AspA iteratively utilizes two molecules of Ant. Figure S6, related to Figure 5: The T2CT di-domain fragment of AspA generates Asperlicin C and D from exogenous Ant-Ant- L-Trp-SNAC. Figure S7, related to Figure 4 and 5: No cyclic Ant-Ant dimer was observed when incubate AspA_A1-T1-C2 with Ant and ATP. Figure S8, related to Figure 7: Three alternative mechanisms for the formation of 6,11-macrocycle precursor of asperlicins C and D. Figure S9, related to Figure 7: Minimum energy pathways for the nucleophilic addition of neutral aniline to the thioester.

2
Expression and purification of intact AspA and dissected proteins. The verified AspA
expression plasmid was retransformed into S. cerevisiae strain BJ5464‐NpgA for protein
expression. Yeast cells harboring the AspA expression plasmid were grown in yeast extract
peptone dextrose medium with 1% (w/v) dextrose at 28 °C for 72 hours. The yeast cell
pellets were harvested by centrifugation 2500 g for 20 min at 4 °C, and resuspended in 20
mL lysis buffer (50 mM NaH2PO4 pH 8.0, 0.15 M NaCl, 10 mM imidazole). The yeast cells
were lysed by sonication on ice followed by centrifugation (35,000 g, 60 min, 4 °C).
The AspA_M1, AspA_M2, AspA_T2CT and AspA_CT expression plasmids were transformed
into E. coli BL21 (DE3). The resulting E. coli cells were grown in 500 mL Luria–Bertani (LB)
medium with 35 mg L‐1 kanamycin at 37 °C until A600 reach to 0.4~0.6 and 60 µL 1 M
Isopropylthio‐β‐D‐galactoside (IPTG) was added to induce protein expression at 16 °C for
overnight. The E. coli cell pellets were harvested by centrifugation (3750 rpm, 15 mins,
4 °C) and resuspend in 30 mL lysis buffer (50 mM Tris‐HCl, 2 mM EDTA, 2 mM DTT, 500
mM NaCl, 5 mM imidazole, pH 7.9). The resuspended buffer was subjected to sonication on
ice to disrupt E. coli cell membranes followed by centrifugation (14,000 rpm, 30 min, 4 °C).
The supernatant were incubated with Ni‐NTA agarose for at least 2 hours at 4°C. The
protein/resin mixture was then loaded onto a gravity flow column and washed by buffer A
(50 mM Tris‐HCl, 500 mM NaCl, pH 7.9) with a stepwise increasing concentration of
imidazole. The target proteins were eluted by using buffer A with 250 mM imidazole.

3
Table S1. List of primers used during this study Primer name Sequence (5’‐3’)
AspA_P1_F atggctagcgattataaggatgatgatgataagactagtatgggttcctacaacgccaa
AspA_P1_R caccattccgtcagcccatg
AspA_P2_F catggctcaatagtagggtcc
AspA_P2_R gccattttcagccgatctaa
AspA_P3_F gtgtggaaactgaagacaaggtc
AspA_P3_R cagatcaacacgggtgatga
AspA_P4_F gacgaagccgttggaccctt
AsPA_P4_R ttcgctttgtccacaatcca
AspA_P5_F ttgcgatgggatgtaaactg
AsPA_P5_R tggtcgacatgctgtagcca
AspA_P6_F atgtctcatgcacaatatga
AsPA_P6_R tcatttaaattagtgatggtgatggtgatgcacgtgttgatatccattcaatgcat
AspA_CT_NcoI_F aaaaaaccatggtgggcagccatcatgactcgtca
AspA_CT_EcoRI_R ttttttgaattctcagtggtggtggtggtggtgttgatatccattcaatgcatg
AsPA_C2A2T2CT_F atcaactatcaactattaactatatcgtaataccatatgcaggatatctttcactgcatg
AspA_T2_NcoI_F aaaaaaccatggtgaactacatatcaaaccaaagg
AspA_P1_NdeI_F atcaactatcaactattaactatatcgtaataccatatgggttcctacaacgccaa
AsPA_A1T1C2_R tcatttaaattagtgatggtgatggtgatgcacgtgtcgctcagtacatagattgag

4
Figure S1: Michaelis‐Menten plots of ATP‐[32P]PPi exchange assay for AspA with (A) Ant,
(B) L‐Trp, and (C) benzoic acid as substrates.

5
Figure S2: SDS‐PAGE gels of the heterologously expressed proteins in this study. AspA_ A1‐
T1‐C2 and AspA_C2‐A2‐T2‐CT were expressed from BJ5464‐NpgA. AspA_CT and AspA_T2‐CT
were expressed from BL21 (DE3). All proteins were purified by C‐terminal His6 tag.

6
Figure S3: Extracted ion mass chromatograms [M+H]+ =407 from AspA in vitro reactions. 1
mM of Ant, L‐Trp, 3 mM ATP and 5 mM MgCl2 were used in the assays. Trace i) No AspA
reaction control; and trace ii) with addition of 10 µM AspA enzyme.
7
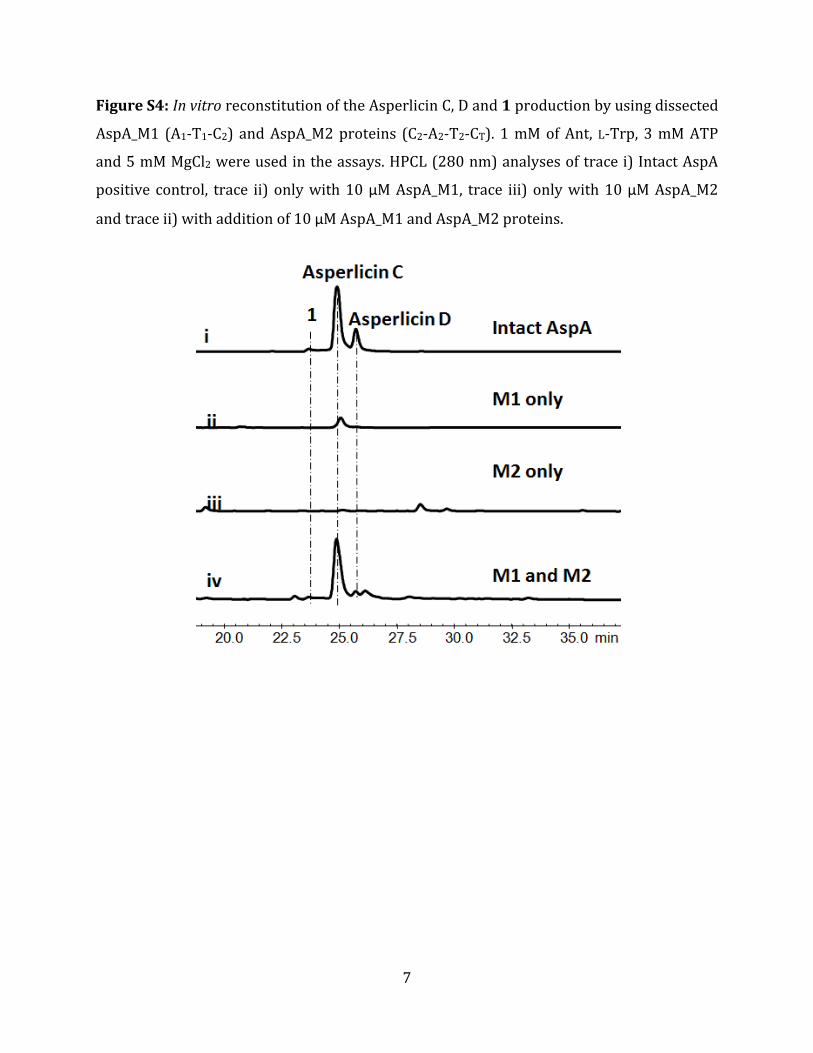
Figure S4: In vitro reconstitution of the Asperlicin C, D and 1 production by using dissected
AspA_M1 (A1‐T1‐C2) and AspA_M2 proteins (C2‐A2‐T2‐CT). 1 mM of Ant, L‐Trp, 3 mM ATP
and 5 mM MgCl2 were used in the assays. HPCL (280 nm) analyses of trace i) Intact AspA
positive control, trace ii) only with 10 µM AspA_M1, trace iii) only with 10 µM AspA_M2
and trace ii) with addition of 10 µM AspA_M1 and AspA_M2 proteins.

8
Figure S5: The first module of AspA iteratively utilizes two molecules of Ant. The holo form
of M1 (50 µM A1‐T1‐C2) was preloaded with Ant (1 mM Ant, 3 mM ATP) for 1 hour while in
parallel the T2‐CT di‐domain was converted to the holo (HS‐pantetheinyl) form via 20 µM
Sfp and 1mM CoASH for 1 hour. The two solutions were mixed on addition of 400 µM Ant‐L‐
Trp‐SNAC and incubated overnight before aliquots were analyzed by LC/MS. Extracted ion
mass chromatograms [M+H]+ =407 from the assay without T2CT di‐domain (trace i); from
incubation including holo T2CT (trace ii), and from the full length AspA starting from Ant, L‐
Trp and ATP (trace iii).

9
Figure S6: The T2CT di‐domain fragment of AspA generates Asperlicin C and D from
exogenous Ant‐Ant‐ L‐Trp‐SNAC. Reactions contained 50mM This‐HCl buffer, pH 7.5, in 100
µL. Traces (i to iii) contained 20 µM Sfp, 2mM CoASH, 100 µM Ant‐Ant‐L‐Trp‐SNAC. 50 µM
T2CT or CT was added in traces i and ii, respectively; No enzyme was added in trace iii. The
suite of asperlicins C, D and compound 1([M+H]+ = 407) are found in traces i but not ii and
iii. Extracted ion mass chromatograms [M+H]+ =407.

10
Figure S7: No cyclic Ant‐Ant dimer was observed when incubate AspA_A1‐T1‐C2 with Ant
and ATP. HPCL traces (280 nm) are shown i) no enzyme control; ii) with addition of1 mM
Ant, 3 mM ATP and 10 µM AspA_A1‐T1‐C2.
11
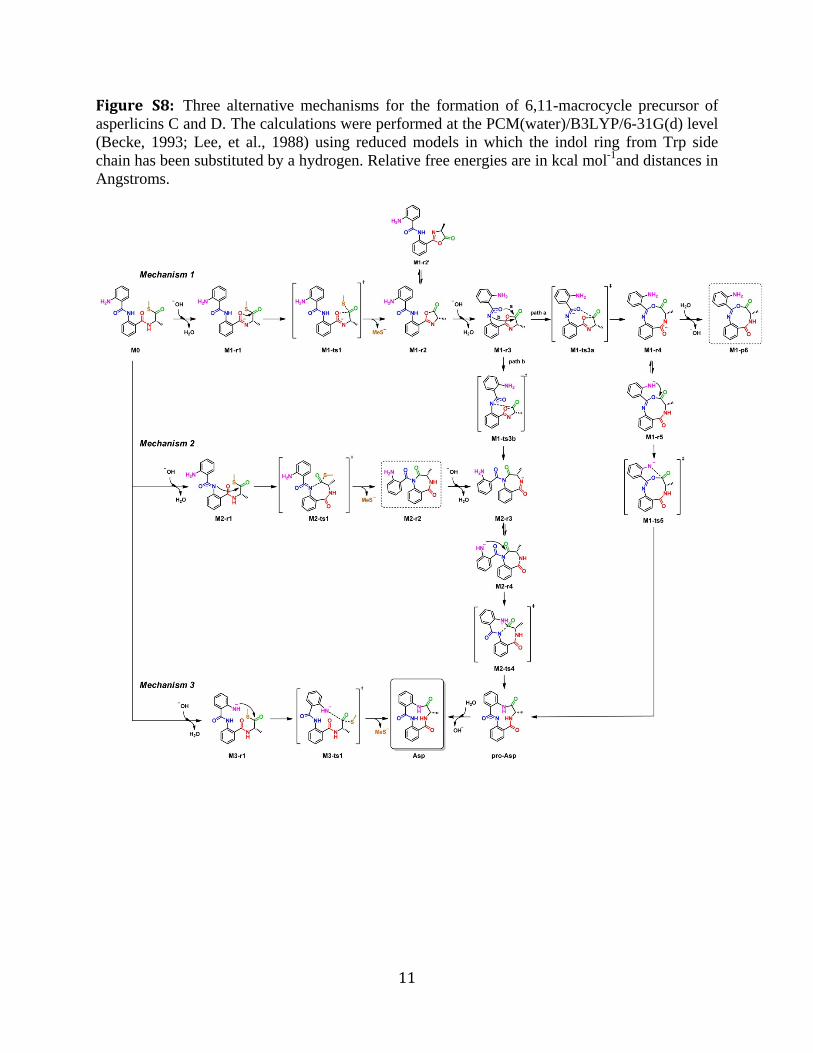
Figure S8: Three alternative mechanisms for the formation of 6,11-macrocycle precursor of asperlicins C and D. The calculations were performed at the PCM(water)/B3LYP/6-31G(d) level (Becke, 1993; Lee, et al., 1988) using reduced models in which the indol ring from Trp side chain has been substituted by a hydrogen. Relative free energies are in kcal mol-1and distances in Angstroms.

12
In mechanism 1, the C4=O carbonyl oxygen (imidate form) first attacks the thioester to generate an oxazol-5(4H)-one, followed by attack of the C11=O carbonyl oxygen (path a) and subsequent ring expansion to yield a 9-membered ring, which can then be opened to the 6,11-macrolactam. Another possible reaction pathway (path b) involves the attack of N1 to the oxazolone to yield a 7-membered ring accessible also from mechanism 2.
13
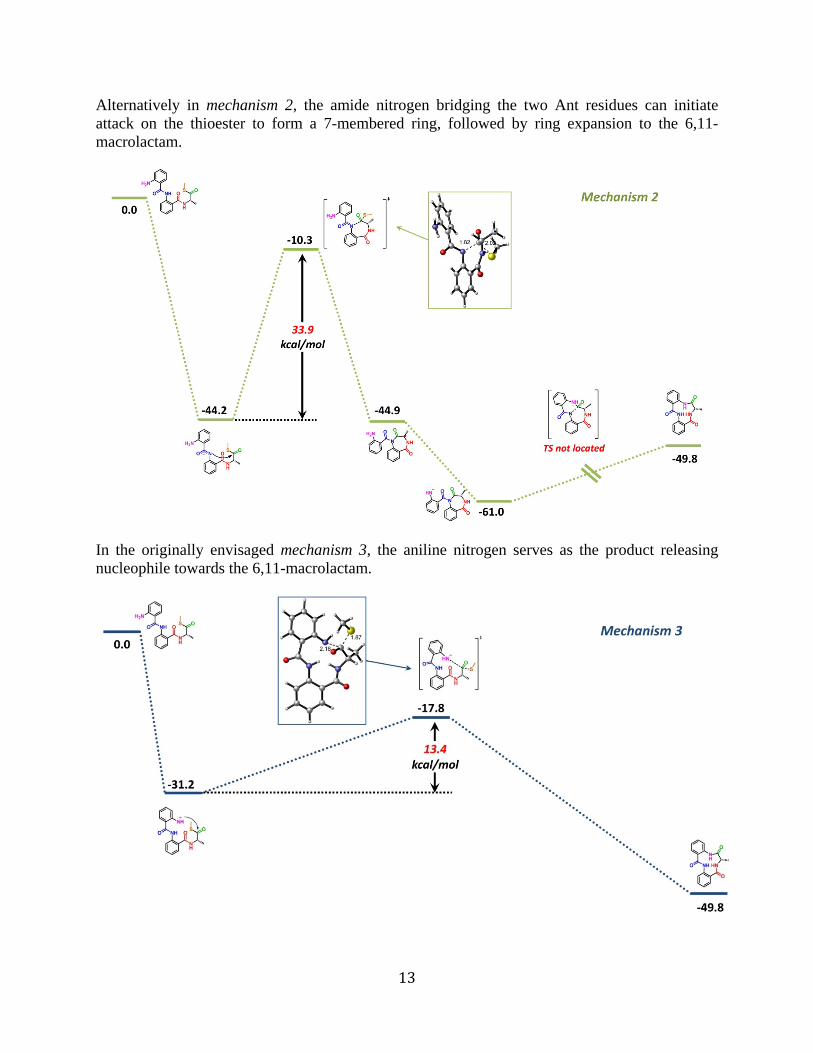
Alternatively in mechanism 2, the amide nitrogen bridging the two Ant residues can initiate attack on the thioester to form a 7-membered ring, followed by ring expansion to the 6,11-macrolactam.
In the originally envisaged mechanism 3, the aniline nitrogen serves as the product releasing nucleophile towards the 6,11-macrolactam.

14
Figure S9. Minimum energy pathways for the nucleophilic addition of neutral aniline to the thioester. All attempts to locate transition structures in these potential energy surfaces were unsuccessful either without explicit solvation (A), in the presence of four water molecules surrounding the NH2 and SMe (B) or with the assistance of explicit solvation and methylamine as a general base (C). This behavior was observed also when attempting the ring expansion of other intermediate macrocycles by nucleophilic addition of aniline to lactam carbonyls (D). The calculations were performed at the PCM(water)/B3LYP/6‐31G(d) level using reduced models in which the indol ring from Trp side chain has been substituted by a hydrogen. Relative free energies are in kcal mol‐1and distances in Angstroms.

15
References:
Becke, A.D. (1993). Density‐functional thermochemistry. III. The role of exact exchange. J. Chem. Phys. 98, 5648‐5652. Lee, C., Yang, W., and Parr, R.G. (1988). Development of the Colle‐Salvetti correlation‐energy formula into a functional of the electron density. Phys. Rev. B Condens. Matter. 37, 785‐789.