supplemental data. kurzbauer et al. (2012). plant cell 10 ... · supplemental data. kurzbauer et...
TRANSCRIPT
Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
1
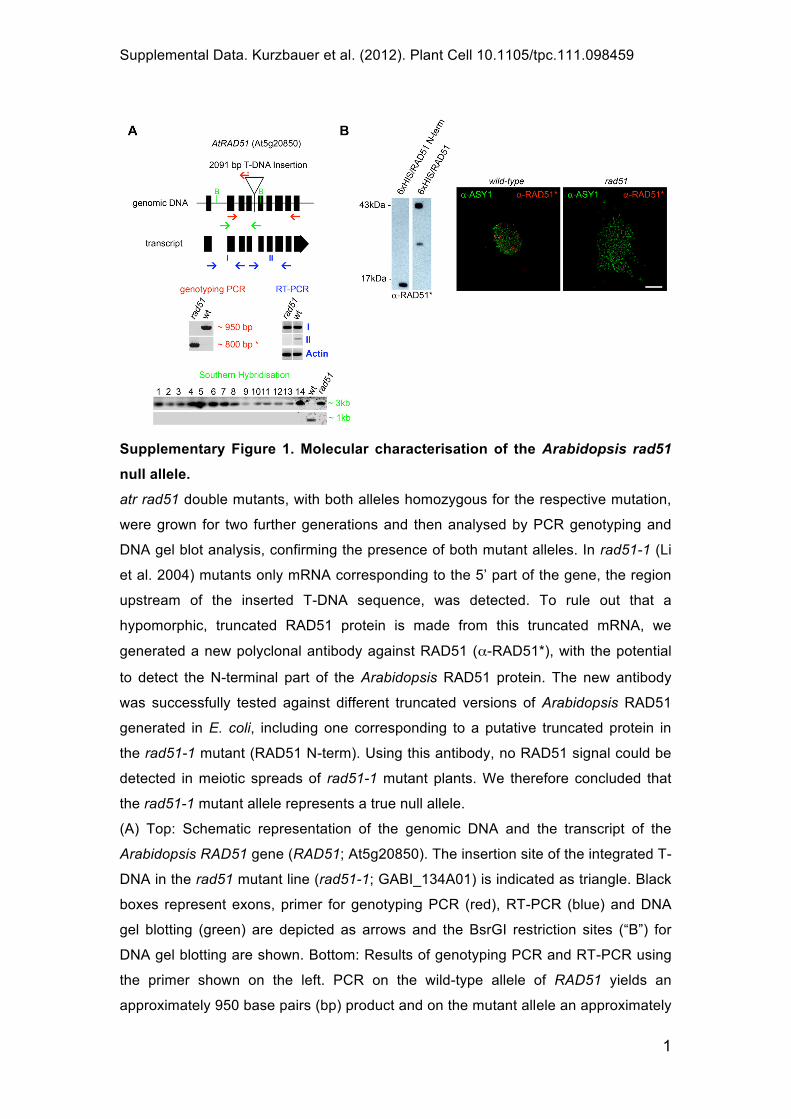
Supplementary Figure 1. Molecular characterisation of the Arabidopsis rad51
null allele.
atr rad51 double mutants, with both alleles homozygous for the respective mutation,
were grown for two further generations and then analysed by PCR genotyping and
DNA gel blot analysis, confirming the presence of both mutant alleles. In rad51-1 (Li
et al. 2004) mutants only mRNA corresponding to the 5’ part of the gene, the region
upstream of the inserted T-DNA sequence, was detected. To rule out that a
hypomorphic, truncated RAD51 protein is made from this truncated mRNA, we
generated a new polyclonal antibody against RAD51 ( -RAD51*), with the potential
to detect the N-terminal part of the Arabidopsis RAD51 protein. The new antibody
was successfully tested against different truncated versions of Arabidopsis RAD51
generated in E. coli, including one corresponding to a putative truncated protein in
the rad51-1 mutant (RAD51 N-term). Using this antibody, no RAD51 signal could be
detected in meiotic spreads of rad51-1 mutant plants. We therefore concluded that
the rad51-1 mutant allele represents a true null allele.
(A) Top: Schematic representation of the genomic DNA and the transcript of the
Arabidopsis RAD51 gene (RAD51; At5g20850). The insertion site of the integrated T-
DNA in the rad51 mutant line (rad51-1; GABI_134A01) is indicated as triangle. Black
boxes represent exons, primer for genotyping PCR (red), RT-PCR (blue) and DNA
gel blotting (green) are depicted as arrows and the BsrGI restriction sites (“B”) for
DNA gel blotting are shown. Bottom: Results of genotyping PCR and RT-PCR using
the primer shown on the left. PCR on the wild-type allele of RAD51 yields an
approximately 950 base pairs (bp) product and on the mutant allele an approximately

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
2
800 bp product. PCR on cDNA yields a product using primer upstream of the
insertion site (I) in wild-type and mutants but primer spanning the insertion site (II)
only give a product with wild-type and never with mutant cDNA. DNA gel blotting was
performed to non-ambiguously demonstrate the exclusive presence of the rad51
mutant allele in atr rad51 F3 plants, defined as homozygous for both mutant alleles
by PCR. In total, 77 plants were tested, 14 individuals are shown (1-14) next to wild-
type (wt) and a rad51 single mutant (rad51). The lower band represents the 1000bp
product of the restriction digest of genomic DNA with BsrGI in wild-type, the upper
band also contains the 2091bp insertion present in rad51 mutants.
(B) Immunoblot and immune-histochemical experiments performed with a newly
generated antibody against Arabidopsis RAD51 ( -RAD51*). This antibody detects
the N-terminus of Arabidopsis RAD51, gives numerous foci on spread nuclei of wild-
type PMCs and does not yield any signal on spread nuclei of mutant rad51 PMCs.
(Size bar: 10 m)

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
3
Supplementary Figure 2. A mutation in ATR suppresses the severe meiotic
defects of rad51 mutants.
(A) Meiotic spreads of pollen mother cells (PMCs) stained with 4',6-diamidino-2-
phenylindole (DAPI). About 36% of atr rad51 double mutant cells undergo meiosis
without DNA fragmentation (see as well Figures 1 and 4). The displayed metaphase
II cell shows DNA fragmentation. dmc1 mutants form univalents and DNA repair is
not compromised (4% seed formation, n=289 siliques). The same phenotype is
observed in atr dmc1 double mutants (1% seed formation, n=114 siliques). A
mutation in ATRIP only partially alleviates the rad51 phenotype (4% seed formation,
n=48 siliques) and most meiocytes display DNA repair defects.
(B) Images of nuclear spreads of PMCs at early to mid-zygotene stage with RAD51
(red) and ASY1 (green) detected by immuno-fluorescence. Bars represent mean
RAD51 foci numbers per meiotic nucleus. RAD51 foci numbers are moderately but
significantly decreased in atr but not in dmc1 mutants. (Error bars represent standard
deviation; * p<0.05; Wilcoxon-Mann-Whitney test; Size bars: 10 m)
Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
4
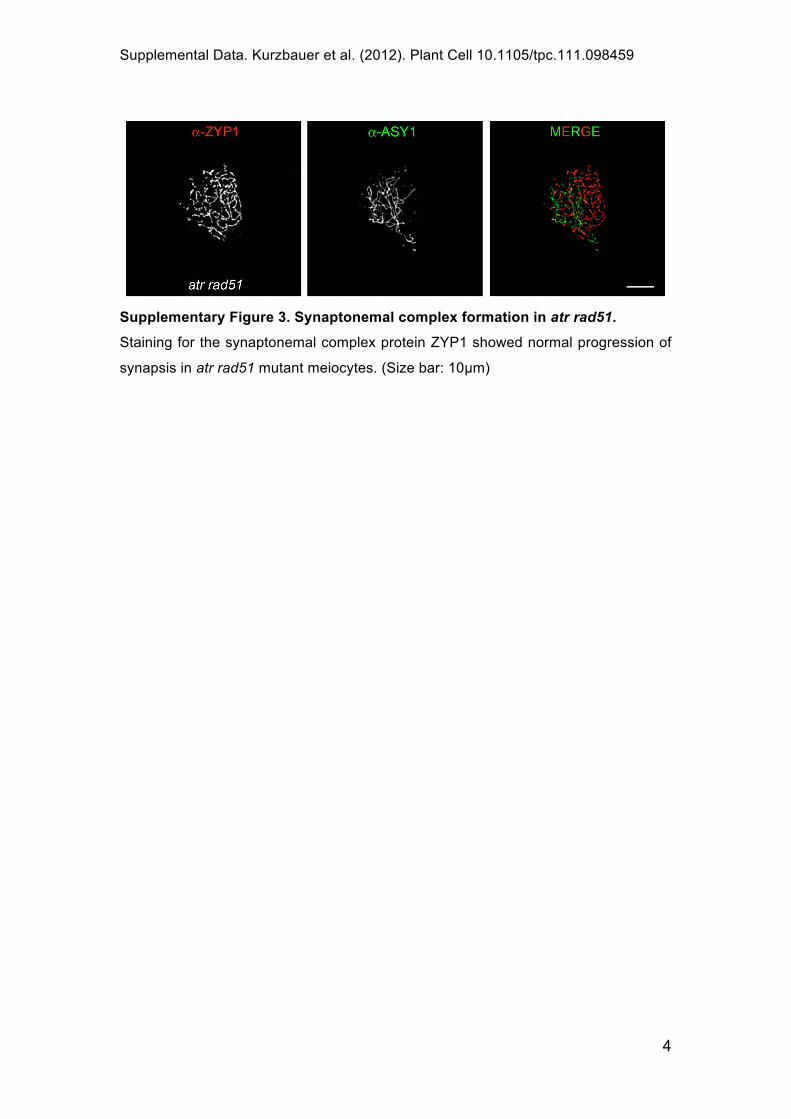
Supplementary Figure 3. Synaptonemal complex formation in atr rad51.
Staining for the synaptonemal complex protein ZYP1 showed normal progression of
synapsis in atr rad51 mutant meiocytes. (Size bar: 10 m)

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
5
Supplementary Figure 4. Antibodies and statistics.
(A) Immunoblot analysis to confirm the specificity of Arabidopsis -RAD51 and -
DMC1 antibodies that have been used for immune-histochemistry. The -DMC1
antibody detects the N-terminus of recombinant Arabidopsis DMC1 and does not
detect recombinant full-length Arabidopsis RAD51. The -RAD51 antibody detects
full-length recombinant RAD51, only shows a very weak residual binding to the
recombinant DMC1 C-terminus protein fragment and does not detect the RAD51 N-
terminus (data not shown).
(B) Monte-Carlo simulations for two further individually scored cells to determine the
expected random frequency of co-localisation of RAD51 and DMC1 foci. In all cases,
the numbers for the observed co-localising RAD51/DMC1 foci (R/D foci) were
significantly lower than the simulated numbers of randomly co-localising events.

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
6
Supplementary Figure 5. RAD51, DMC1 and H2Ax localisation in meiocytes.
Additional images of nuclear spreads of wild-type leptotene PMCs with RAD51 (red),
DMC1 (green) and H2Ax (blue) detected by immunofluorescence (please refer to
Figure 3 for picture details). (Size bar: 10 m)

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
7
Supplementary Table 1 – Inter-foci distances
The differences between observed inter-foci distances of different foci classes were
evaluated by two different methods (see below) and were found to not significantly
differ from each other (RD –RAD51 and DMC1 doublet; DD – DMC1 doublet; RR –
RAD51 doublet).
A) Fisher's Exact Test with two distance classes between foci
nm RR RD DD
0-150 7 12 4
150-450 65 89 50
Total 72 101 54
Fisher's Exact Test
p-value
RR vs RD 0,8064
DD vs RD 0,5804
RR vs DD 0,7568
B) Mann-Whitney Test for comparison of (non-parametric) distribution
nm RR RD DD
0-50 0 0 0
50-100 0 2 0
100-150 7 10 4
150-200 6 13 13
200-250 19 13 5
250-300 11 10 8
300-350 10 14 8
350-400 11 19 6
400-450 8 20 10
Mann-Whitney Test for comparison of (non-
parametric distribution)
p-value
RR vs RD 0,1978
DD vs RD 0,06191
RR vs DD 0,4227

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
8
Supplementary Table 2 – Primer information
primer name sequence 5' - 3'
atr-2 wild-type allele
atr2_1 GGATCAAGTACTACTGACTCAG
atr2_3 CAACTCATTTTGAATATGAGAG
atr-2 mutant allele
atr2_1 GGATCAAGTACTACTGACTCAG
SALK LBa1 TGGTTCACGTAGTGGGCCATCG
rad51-1 wild-type allele
pcr_atrad51_1 GGTTCCATCACGGAGTTATATGG
pcr_atrad51_2 AGCCATGATATTCCCACCA ATC
rad51-1 mutant allele
rad51TDNA CCCATTTGGACGTGAATGTAGACAC
rad51gen GCAGGTATTGCTTCTGTTGATGTA
dmc1 wild-type allele
atdmc1_a CCTGCAATGGTCTCATGATGCATAC
atdmc1_d ACTAATCCTTCGCGTCAGCAATGC
dmc1 mutant allele
atdmc1_d ACTAATCCTTCGCGTCAGCAATGC
Feldmann LB GATGCAATCGATATCAGCCAATTTTAGAC
asy1 wild-type allele
Asy1_new_LP TCATGACATCTTGGCTGTCAG
Asy1_new_RP GTGATGGCTCAGAAGCTGAAG
asy1 mutant allele
Asy1_new_RP GTGATGGCTCAGAAGCTGAAG
SALK LBa1 TGGTTCACGTAGTGGGCCATCG
atrip wild-type allele
ATRIP_LP TACGGGATAATCTTGTGGCTG
ATRIP_RP ACCGAACAGGATGTGAAACAG
atrip mutant allele
ATRIP_RP ACCGAACAGGATGTGAAACAG
SALK LBa1 TGGTTCACGTAGTGGGCCATCG
cDNA
atrad51-E1-dn GTCCAACAACAAGACGATGAAGA
atrad51-E3-up CTTTATCGAGCTCCCGTGATCCAG
DNA gel blot
RAD51_probe_dn ATCGCGATGTGAAATAGGTGTG
RAD51_probe_up CTCCTTGATCCATGG GAAGCT
construction of expression plasmids
DMCstart103FWD7 ATAGAATTCATGTCAGTTGTAAAAATCACTACAG
DMCstartallREV8 ATACTGCAGCTAATCCTTCGCGTCAGC
DMC1stop103FWD1 CTTATAAAGAGGAAATAAGTTGTAAAAATC
DMC1stop103REV2 GATTTTTACAACTTATTTCCTCTTTATAAG

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
9
RAD51stop98FWD1 CTCCATGCTCAGTGACAGGAAATT
RAD51stop98FWD2 AATTTCCTGTCACTGAGCATGGAG
Supplementary methods
Mutant plant line
atrip/suv2-2 (SALK_077978; Sakamoto et al. 2009).
Primer information
Primer for the amplification of RAD51 cDNA were atrad51-E1-dn, binding within exon
1, together with atrad51-E3-up, binding within exon 3 and pcr_atrad51_1, binding
within exon 4, together with pcr_atrad51_2, binding within exon 8. The probe used
for the DNA gel blots was generated by PCR using the primer pair RAD51_probe_dn
and RAD51_probe_up (Supplementary Table 2).
Reverse transcription PCR
Total RNA was isolated from plants using Promega’s SV Total RNA Isolation System
according to the manufacturer’s protocol. cDNA synthesis was conducted with the
iScript cDNA synthesis kit (Biorad). Amplification of cDNA was performed with the
primer pairs given above under standard PCR conditions.
Verification of the rad51-1 mutant allele by DNA gel blot analysis
The presence of the atr-2 and rad51-1 mutant alleles in generations F3 and F4 was
verified via DNA gel blotting. Two inflorescences per plant were homogenized in
Urea Lysis buffer (0.3M NaCl, 30mM Tris-Cl pH8, 20mM EDTA pH8, 1% (w/v) N-
Lauroylsarcosine, 7M Urea) using plastic pestles (Sigma) and incubated at 50°C for
10 minutes. After extraction with PCI (Biomol) and centrifugation (5min, 14000rpm,
RT), the upper phase was recovered and DNA was precipitated with 3M NaOAc
pH5.2 and isopropanol. After washing with 70% EtOH, the pellet was air-dried and
re-suspended in ddH2O. DNA concentration was estimated on a 1% agarose gel and
500ng of DNA were digested overnight in a volume of 100 l using 20 units of BsrGI
restriction endonuclease (Fermentas). RNA was removed by treatment with 1 l
RNaseA (500 g/ml, Roche) at 37°C for 1 hour. DNA was precipitated with 3M
NaOAc pH5.2 and 96% EtOH at -20°C. After centrifugation, the pellet was washed
with 70% EtOH, dried, re-suspended and loaded onto a 0.8% agarose gel. DNA
fragments were separated at 80V. The gel was de-purinated in 0.25M HCl and after
washing, it was neutralized in 0.5M NaOH/1.5M NaCl, followed by equilibration in

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
10
0.5M Tris/1.5M NaCl pH7. DNA was transferred in 20x SSC (3M NaCl, 0.3 M
Na3Citrat) overnight to a Nylon membrane (Hybond-N, Amersham). Crosslinking was
performed in a Strata crosslinker (UV StratalinkerTM 2400, Stratagene) using the
autocrosslink program. Afterwards, the membrane was air-dried.
Probes were labeled with 50 Ci [ 32P] dATP by Klenow polymerase (10u/ l, MBI
Fermentas). Un-incorporated nucleotides were removed using the QIAquick
Nucleotide Removal Kit (QIAGEN). The membrane was pre-hybridized with Church
& Gilbert buffer (1% [w/v] BSA, 1mM EDTA, 7% [w/v] SDS, 0.5M phosphate buffer
pH7.2). After addition of the radioactively labeled probe, the membrane was
hybridised overnight at 65°C, washed twice with 2x SSC/0.1%SDS and once with 1x
SSC/0.1%SDS for 30minutes. After washing, the membrane was sealed into a
plastic bag and detection was accomplished on a Phosphor Screen (Kodak) in a
Biorad Molecular Imager FX Pro Plus.
Generation of RAD51 and DMC1 expression constructs
The coding sequence (CDS) of Arabidopsis RAD51 was cloned in frame into pET28a
(Merck) using EcoRI restriction sites. The CDS of Arabidopsis DMC1 was cloned in
frame into pET28a using the restriction sites EcoRI and XhoI. The ATPase domain
(C-terminal; amino acid residues 104-344) of DMC1 was amplified by PCR using the
primer DMCstart103FWD7 and DMCstartallREV8 with KOD DNA Polymerase
(Novagen). Each primer included either an EcoRI or a PstI restriction site. The C-
terminal domain of DMC1 was then cloned in frame into the vector pMAL-TEV/6xHIS
(this vector is a modified version of the NEB pMAL-c2 plasmid with a TEV protease
cleavage site instead of the factor Xa site and an additional 6xHIS-tag, inserted in the
SacI site upstream of the TEV site).
In order to construct the expression plasmids for the N-terminal domains of DMC1
(amino acid residues 1-103) and RAD51 (amino acid residues 1-98), respectively, in
vitro mutagenesis was conducted. The above mentioned vector pET28a/DMC1 was
utilised as a template for PCR amplification using recombinant Pfu DNA Polymerase
(Fermentas) with primer DMC1stop103FWD1 and DMC1stop103REV2. For
generating an overexpression plasmid of the N-terminal domain of RAD51, the
plasmid pET28a/RAD51 served as a template for PCR amplification using
recombinant Pfu DNA Polymerase (Fermentas) with primer RAD51stop98FWD1 and
RAD51stop98FWD2. The four primer mentioned above were designed to insert a
stop codon at the end of the N-terminal domains. After amplification, the PCR
products were incubated with DpnI for 3 hours and transformed into chemically

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
11
competent E.coli DH5alpha cells. All amplicons generated by PCR were sequenced
to confirm error-free amplification (LGC Genomics).
Expression and purification of recombinant proteins for Immunoblot analysis
The plasmids containing full-length DMC1, full-length RAD51 or the C-terminal and
the N-terminal domains of DMC1 and RAD51, respectively, were transformed into
chemically competent Rosetta(DE3)pLysS cells (Merck). A single colony was
inoculated in 2 liters of 2xTY liquid media (16g/l peptone, 10g/l yeast extract, 5g/l
NaCl) and the culture was grown at 37°C until an OD600 of about 0.7 was reached.
1mM Isopropyl- -D-thiogalactopyranosid (IPTG) was added and after 3 hours the
cells were harvested by centrifugation (5000 g, 30', 4°C). The cell pellet was frozen
at -20°C and thawed on ice. Subsequent resuspension of the pellet in ice cold buffer
A (one gram pellet per 5ml of 50mM phosphate buffer pH 8, 500mM NaCl, 2mM beta
Mercaptoethanol, 0.05% NP-40, complete Mini EDTA-free, Roche) was followed by
sonication on ice and an additional round of centrifugation (20000 g, 30', 4°C) yielded
a precleared lysate that was incubated with Ni-beads (Profinity IMAC Ni-charged
resin, Biorad) for 1h at 4°C. The Ni-beads were washed three times with buffer A and
the recombinant 6xHIS-tagged proteins were eluted with 1M Imidazole pH8 in buffer
A. The purity and concentration of the recombinant proteins in the eluates were
analysed by SDS-PAGE using Coomassie staining and Bradford assay according to
standard techniques.
Generation of an antibody directed against Arabidopsis RAD51 ( -RAD51*)
The His-tagged full length Arabidopsis RAD51 protein was expressed and isolated as
described above with the modification that after sonication and subsequent
centrifugation the pellet and not the precleared lysate was used to purify the
recombinant RAD51 protein. To this end the inclusion bodies of the pellet were
resolubilised in buffer A containing 5M Guanidinium chloride and the protein solution
was incubated with Ni-beads for 4 hours at room temperature. After elution with 1M
Imidazole in buffer A containing 5M Guanidinium chloride, the RAD51 protein was
dialysed stepwise against decreasing concentrations of Guanidinium chloride (3M,
1.5M, 0M) in buffer A at 4°C, with buffer changes every 8 hours. After removal of
Imidazole and Guanidinium chloride the protein solution was concentrated with the
help of Vivaspin 20 10,000 MWCO PES centrifugal concentrators (Sartorius Stedim
Biotech) and gel filtration was performed using an ÄKTA FPLC system and a
Superdex 200 16/60 column (GE Healthcare Life Sciences). The fractions that

Supplemental Data. Kurzbauer et al. (2012). Plant Cell 10.1105/tpc.111.098459
12
contained the His-tagged RAD51 protein were pooled and approximately 0.5 mg of
the affinity purified recombinant protein was used for antibody production performed
by Eurogentec (Belgium).
Immunoblot analysis
Recombinant proteins (10 ng of purified protein per lane) were separated by SDS-
PAGE and blotted onto a PVDF membrane using standard techniques. The
membrane was blocked with 3% milk (Nonfat dried milk powder; AppliChem) in TBS-
T (10mM Tris/HCl pH 7.5, 150 mM NaCl, 0.05% Tween20) for 1h and incubated for
1h with 1:1000 dilution of anti-DMC1 antibody from rabbit or 1:100 dilution of anti-
RAD51 antibody from rat or 1:5000 dilution of anti-RAD51* antibody from rat in TBS-
T/3% milk. Commercially available secondary antibodies coupled to HRP and
directed against rabbit (1:2000, HP1202, Hycult biotech) or rat (1:5000, ECL Anti-rat
IgG, HRP linked AB, GE Healthcare) were used to detect the recombinant proteins
on the membrane with the help of HRP reagent (Pierce ECL Western Blotting
substrate, Thermo Scientific). The calculated molecular weight is 41,4 kDa for
6xHIS/DMC1, 41,2 kDa for 6xHIS/RAD51, 74,2 kDa for MBP/6xHIS/DMC1 C-term,
15,4 kDa for 6xHIS/DMC1 N-term and 14,6 kDa for 6xHIS/RAD51 N-term.