sun, sand and sea urchins
TRANSCRIPT

vided convenient building levels, and the Rock has been extensively quarried to provide build- ing stone, and also fill to reclaim additional land from the sea. Potable water is obtained from several sources, but the most spectacular are the natural slopes surfaced so as to form rainwater catchment areas (Fig. 11). Within the Rock, some 50 km of tunnels and chambers honey- comb its interior even more extensively than its caves. It is enlightening to see in so small and accessible an area how geological factors have influenced the growth of this famous port and fortress.
Suggestions for further reading Rose, E. P. F. & Rosenbaum, M. S. 1990. Royal
Engineer Geologists and the Geology of Gibraltar. The Gibraltar Museum, 55 pp. (Reprinted from The Royal EngineersJournal, v. 103 (for 1989), pp. 142-151,248459; v. 104 (for 1990),
Rose, E. P. F. & Rosenbaum, M. S. A Field Guide to the Geology of Gibraltar. The Gibral- tar Museum, 192 pp. (To be published in 1991.)
Rosenbaum, M. S. & Rose, E. P. F. Geology of Gibraltar. Single sheet 880 x 600 mm: Side 1, cross-sections and bedrock geology map 1 : 10 000, Quaternary geology, geomorpho- logy, and engineering use of geological features
pp. 61-76, 128-144.)
maps 1:20 000; Side 2, combined bedrock/ Quaternary geology map 1:lO 000, plus 17 coloured photographshigures and explanatory text. (To be published in 1991.)
Edward P . F . Rose is Senior Lecturer in Geology at Royal Holloway & Bedford New College, and M . S . Rosenbaum is Lecturer in Engineering Geology at Imperial College, both of the University of London.
Sun, sand and sea urchins STEPHEN K. DONOVAN
Jamaica, land of holiday dreams, is also fascinating to the geologist. In particular, the fossil record is diverse, the ancient fauna containing many rarities. The fossil ochinoid fauna is especially large and varied for such a small island, with about 130 nominal species ranging f rom the late Cretaceous to the Pleistocene.
T h e conception that travellers usually have of Jamaica is of a tropical paradise with every possible convenience available to the tourist - sun, sand and sea. However, life isn’t always so rosy. Many a tourist has had his or her holiday rudely interrupted by indiscretely stepping on a dark, spiny blob on the sea floor - a regular echinoid, or sea urchin (commonly called a ‘sea egg’ in the Caribbean). Seventeen species of shallow-water echinoid are known from Jamaica, of which the least loved is the so-called black urchin, Diadema antillaturn Philippi, whose long, slender, fragile spines have punc- tured many unsuspecting feet and then broken off, leaving lengths embedded in the tender flesh.
Whilst unfortunate for the unwary tourist, this sea-egg fauna is of great interest t o , the palaeontologist with a liking for echinoderms. A day at the beach can easily degenerate into a research exercise and specimen-collec ting trip, the only equipment needed being a snorkel and face mask. Even more exciting for the geologist, however, is the large fauna of fossil echinoids that are locally extremely common in the Cre- taceous and Cenozoic rocks of the island. This fauna includes about 130 nominal species; and further, hitherto undescribed, taxa are being collected all the time. The fauna is important palaeogeographically, providing constraints on Caribbean-plate evoluuon - both systematically (it includes many taxa known only from the
GEOLOGY TODAY May-June 1991 /I01
A Fig. 11. The main east- coast water catchments, above Sandy Bay, viewed from the south. (Formed by sheets of corrugated iron fastened to wooden battens set in the smoothed surface of a natural scree breccia/ Catalan Sand slope. Hotel developments at toe of slope indicate scale.)
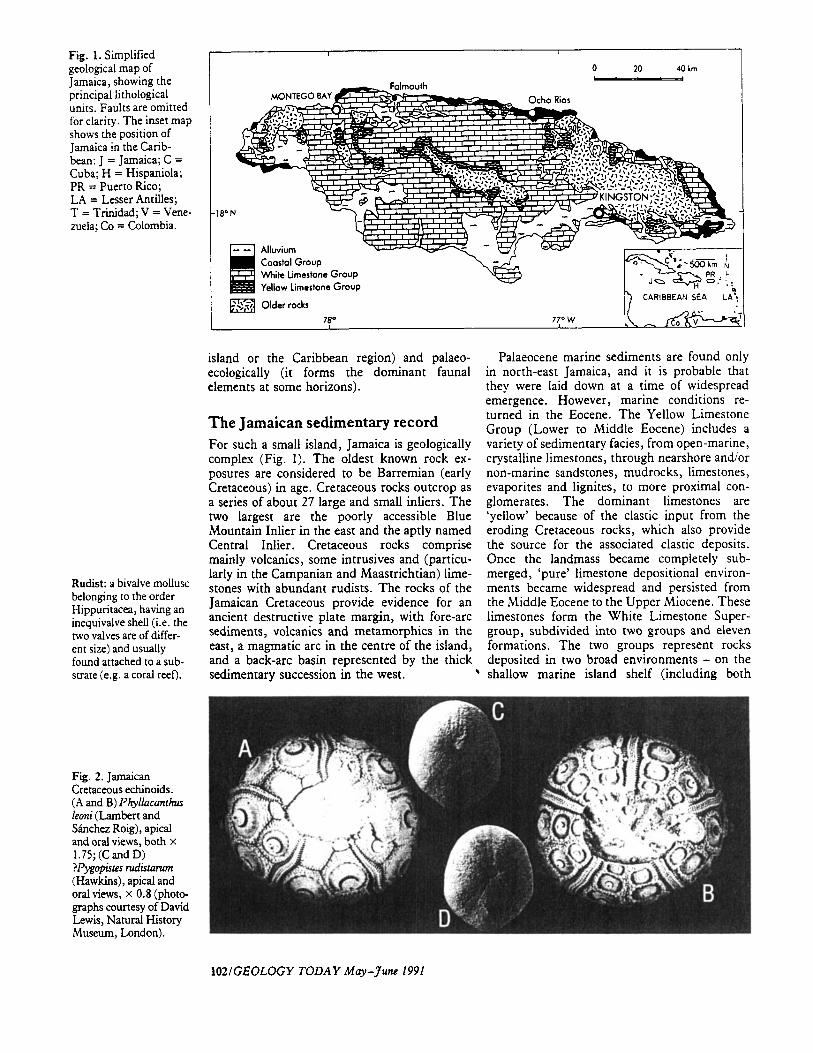
Fig. 1. Simplified geological map of Jamaica, showing the principal tithological units. Faults are omitted for clarity. The inset map shows the position of Jamaica in the Carib- bean: J = Jamaica; C = Cuba; H = Hispaniola; PR = Puerto Rico; LA = Lesser Antilles; T = Trinidad; V = Vene- mela; Co = Colombia.
Rudist: a bivalve mollusc belonging to the order Hippuritacea, having an inequivalve shell (i.e. the two valves are of differ- ent size) and usually found attached to a sub- strate (e.g. a coral reef).
Fig. 2. Jamaican Cretaceous echinoids. (A and B) Phyllacanrhus ka i (Lambert and Sinchez Roig), apical and oral views, both X 1.75; (C and D) ?Pygopistes rudisrarum (Hawkins), apical and oral views, x 0.8 (photo- graphs courtesy of David Lewis, Natural History Museum, London).
0 20 40 km - 1
- 1 P N
White Limestone Group Yellow Limestone Group
island or the Caribbean region) and palaeo- ecologically (it forms the dominant faunal elements at some horizons).
The Jamaican sedimentary record For such a small island, Jamaica is geologically complex (Fig. 1). The oldest known rock ex- posures are considered to be Barremian (early Cretaceous) in age. Cretaceous rocks outcrop as a series of about 27 large and small inliers. The two largest are the poorly accessible Blue Mountain Inlier in the east and the aptly named Central Inlier. Cretaceous rocks comprise mainly volcanics, some intrusives and (particu- larly in the Campanian and Maastrichtian) lime- stones with abundant rudists. The rocks of the Jamaican Cretaceous provide evidence for an ancient destructive plate margin, with fore-arc sediments, volcanics and metamorphics in the east, a magmatic arc in the centre of the island, and a back-arc basin remesented bv the thick
Palaeocene marine sediments are found only in north-east Jamaica, and it is probable that they were laid down at a time of widespread emergence. However, marine conditions re- turned in the Eocene. The Yellow Limestone Group (Lower to Middle Eocene) includes a variety of sedimentary facies, from open-marine, crystalline limestones, through nearshore andor non-marine sandstones, mudrocks, limestones, evaporites and lignites, to more proximal con- glomerates. The dominant limestones are ‘yellow’ because of the clastic input from the eroding Cretaceous rocks, which also provide the source for the associated clastic deposits. Once the landmass became completely sub- merged, ‘pure’ limestone depositional environ- ments became widespread and persisted from the Middle Eocene to the Upper Miocene. These limestones form the White Limestone Super- group, subdivided into two groups and eleven formations. The two groups represent rocks deposited in two broad environments - on the
sedimentary succession {n the west. ’ shallow marine island shelf (including both
1OZlGEOLOGY TODAY May-3une 1991

open-shelf and lagoonal sediments) and in deeper-water settings. Clastic deposition ceased, but volcaniclastics are interbedded with chalks of Miocene age.
Widespread limestone deposition ended with the general emergence of the island in the late Miocene. Late-Miocene-to-Holocene sedimenta- tion continued around the margins of the island, and these rocks together form the Coastal Group. Included in this sequence are both shallow-water (particularly of reefal origin) and deeper-water limestones, with clastic rocks derived from erosion of emergent strata. Con- glomerates in this sequence frequently contain limestone clasts, reworked from the White and Yellow Limestones.
Jamaican fossil echinoids Fossil echinoids are common in three intervals in the Jamaican stratigraphic record: the Campanian and Maastrichtian (Upper Creta- ceous); the Plio-Pleistocene; and, particularly, the marine rocks of the Eocene Yellow Lime- stone Group.
Some echinoids are known from pre- Campanian sedimentary rocks, but these are very rare. In contrast, about ten species have been recognized from the Campanian and Maastrichtian. Most common among these are the large, globular cidaroid Phyllacanthus leoni (Lambert and Sanchez Roig; Fig. 2A and B), the arbacioid Goniopygus supremus Hawkins, and the beautiful salenioid Heterosalenia occidentalis Hawkins (Fig. 3). All three species are well known from tests, and the spines of the first two taxa are often so common that they form mono- specific horizons.
There are also several rarities in the fauna, such as ?Pygopistes rudistarum (Hawkins). The holotype of this species (Fig. 2C and D) was originally described in 1923, the specimen having been collected from western Jamaica by the distinguished amateur, C. T. Trechmann. No further specimen was found until 1987, when Jacquie Bowen, then an undergraduate at the University of the West Indies (UWI), found a second specimen while on a field excursion. The discovery of three new late Cretaceous species (all awaiting description) in the past five years suggests that further rarities are waiting to be collected. The best areas for finding Creta- ceous echinoids are the Guinea Corn Formation of the Central Inlier and sedimentary sequences in those inliers a t the western end of the island. Cretaceous echinoids of the Blue Mountain Inlier in the east are poorly known, mainly because of the mountainous, largely inaccessible terrain.
If the Cretaceous fauna is small, Jamaica's Palaeocene echinoids are virtually unknown. This is at least partly a function of the small area
Fig. 3. The Jamaican Cretaceous echinoid Heteroralenia occidentalis Hawluns, apical view, x 2.2.
of outcrop of marine Palaeocene rocks in north- east Jamaica, although no systematic search has been undertaken for echinoids in these rocks. The available evidence is limited to some frag- mentary remains (mainly spines) of a cidaroid and a diadematoid, and rare trace fossils. The diadematoid spines are significant, as the only other fossil members of this group so far found in Jamaica are much younger and come from an Ipswichian (Pleistocene) raised reef on the north coast (see below). The Ipswichian remains are presumably from the still-extant black urchin Diadema antillarum Philippi or a related taxon, but the identity of the Palaeocene species is un- known. The trace fossils, Scolicia cf. plana Ksigtkiewicz, are also important, representing the trails of heart urchins, which occur as tests in the Cretaceous and Eocene but not in the Palaeo- cene. The palaeoenvironmental setting is also different from the Cretaceous; whereas Creta- ceous echinoids are limited to shallow-water limestones and interbedded shales, the Palaeo- cene spines occur in storm beds and turbidites (and are thus possibly reworked from shallower- into deeper-water deposits), while Scolicia occurs on the top of turbiditic sandstone units.
The return of widespread marine conditions in the Eocene heralded the golden age of Jamaican echinoids (Fig. 4). In particular, echinoids thrived in the Yellow Limestone marine facies. Some of these species are unique, such as the holotype of Triadechinus multiporus Arnold and Clark (Fig. 4P), whereas others, particularly the oligopygoid Haimea ovum- serpentis (Guppy; Fig. 4C and D) and related species, are known from thousands of specimens and are extremely abundant at some localities.
Cidaroid, etc.: the terms Many of the species are of palaeogeographic cidaroid, arbacioid, interest. For example, the genus Asrerostoma salenioid, and so on, (Fig. 4A and B) is only known from three refertodifferenrf-bes species, ' divided between Cuba and Jamaica. ofechinoid having The oligopygoids Haimea and Oligopygus somewhat different (Fig. 4C-F) are more widely known, being characteristics.
GEOLOGY TODAY May-gune 1991 I103

Fig. 4. A cavalcade of Jamaican Eocene echinoids. None of the figured genera is known to have survived inKO h e Oligocene. (A and B) Asrerosroma pawsai Kier, apical and oral views; (C and D) Haimea
apical and oral views; (E and F) Oligopygus werherbyr de Loriol, apical and oral views (a Jamaican species, although this specimen is from the Crystal River Formation, Florida); (G and H) Fellius fbvearuc (Jackson), apical and oral views; (I and J) Tarphy- pygw nolabilk Arnold and Clark, apical and oral views; (K) Trochalo- soma chondra (Arnold and Clark), apical view; (L) P h y m o s m peloria Arnold and Clark, apical view; (M and N) Carib- buster dyscrirus (Arnold and Clark), apical and oral views; (0) Srene- chinus regularis Arnold and Clark, apical view, x 1 ; (P) Triadechinus mulriponu Arnold and Clark, apical view. All figures except 0 are x 0.8. All specimens (except E and F) are from the Yellow Limestone Group.
~ ~ e r p e n ~ (GUPPY),
found throughout the Caribbean and palaeo- geographically adjacent areas. Further, these three genera are all limited to the Eocene, making them useful biostratigraphic markers.
After the end of Yellow Limestone deposi- tion, in the Middle Eocene, the Jamaican echinoid fauna went into an apparent decline. Yellow Limestone deposition spanned a period of about 6.5 million years, and about 70 nominal species of echinoid have been described from deposits of this age, but only about 20 taXa have been recorded from the succeeding Eocene formations of the White Limestone Supergroup, equivalent to the last 8 million years of the epoch. This difference may be real,. reflecting extinctions that are known to have been occur- ring from the mid-Eocene to the mid-Oligocene.
On the other hand, at least part of this pattern could be an artefact, due to a failure on the part of palaeontologists to make detailed collections from these formations.
An illustration of this is given by two recent discoveries by amateur collectors, which have more than doubled the Eocene fauna from the White Limestone. Bill Schickler was puzzled by the globular fossils that occurred in great abund- ance in the flower beds around his home near Ocho Rios on the north coast. Since 1988, when parties of collectors from UWI first started in- vading Bill’s garden (with his blessing!), about 500 echinoid tests and spines have been col- lected, representing about 12 taxa and more than doubling the specific diversity of the later Eocene fauna. Further, the published map
104IGEOLOGY TODAY May-June 1991

shows Bill’s house to lie within the outcrop area of a Miocene formation, which has been shown to be incorrect by the occurrence of a typical Eocene echinoid fauna, dominated by the oligo- pygoid Haimeu (see above).
A second fauna of broadly the same age has been discovered by Dawn Scott, a distinguished Jamaican artist, from a hillside in north-east Jamaica. Whereas Bill Schickler’s garden fre- quently yields large, complete tests, whole specimens from Dawn’s locality are invariably less than 10 mm in maximum dimension. Frag- ments of larger tests are also common, but are mainly unidentifiable. Among the recognizable fauna, however, are the oldest known examples of spines from the cidaroid genus Eucidaris, which has an extant relation in the Jamaican shallow-water fauna (Fig. SB and C).
If later Eocene echinoids are rare, those of the Oligocene are almost unknown, with fewer than five species being recognized. While this rarity may be a consequence of Eocene-Oligocene extinctions, the echinoid fauna is also sparse in the Miocene of Jamaica, although it is diverse elsewhere. This suggests that our perception of Oligocene and Miocene echinoid faunas of Jamaica may be strongly influenced by the mode of preservation and an associated collecting bias. Oligocene and Miocene shallow-water lime- stones of Jamaica are crystalline and do not encourage the collection of macrofossils. As in the case of the iater Eocene, further field-work
Fig. 5. JamaicanPLio- cene and Quaternary echinoids. (A) CIypeasrer rosncew (Linnaeus), Holocene, apical view x 0.8; (B and C) Eucid~ris mbuloides (Lamarck), Holocene apical and oral views, x 0.8; (D) Encope aff. merdrupi Durham, Plio-Pleistocene, apical view, x 0.5.
may greatly clarify the true diversity of the island’s midCenozoic echinoid fauna.
Echinoids are quite plentiful in the Plio- Pleistocene deposits of the island, and new taxa continue to be discovered. Where ‘complete’ specimens are preserved, these tend to be tests of clypeasteroids (Fig. 5A and D), whose in- ternal buttressing strengthens them in life and subsequently retards post-mortem disarticu- lation. Amongst echinoid tests, that of Cfypeaster is undoubtedly one of the most robust. CZype- m e t maceus (Linnaeus; Fig. 5A) is a common component of the living echinoid fauna, and it is not surprising that it also occurs as fossils in the Plio-Pleis tocene.
A recent study at UWI by research student Carla Gordon has compared the echinoids of the last interglacial (Ipswichian) Falmouth Forma- tion, a raised reef on Jamaica’s north coast, with those living around the island at the present day. Mollusc and coral faunas from the Ipswichian of the Caribbean region are little different from those of the Holocene, and Carla has similarly identified about 50% of the extant echinoid genera of Jamaica’s shallow-water fauna in the Falmouth Formation. What is particularly novel about Carla’s analysis is that echinoid tests are almost unknown from the Falmouth Formation, and her identifications are based upon disarticu- lated ossicles (spines, test plates and lantern elements). For exampk, only one test of the cidaroid Eucidaris tribuloides (Lamarck; Fig. SB
GEOLOGY TODAY May-June 1991 I105

and C) is known from the Jamaican Pleistocene, yet spines and other elements of this taxon are abundant in the Falmouth Formation. Exten- sion of our studies of disarticulated ossicles back in time as a supplement to the collection of new test faunas (particularly from the Palaeocene and mid-Cenozoic) will add to our knowledge of the Jamaican fossil echinoids and should help develop a truer picture of the evolution of the Caribbean sea-egg fauna over the past 100 million years.
Suggestions for further reading Arnold, B. W. & Clark, H. L. 1927. Jamaican
fossil echini. Memoirs of the Museum of Com- parative Zoology at Harvard, v. 50, pp. 1-75.
Arnold, B. W. & Clark, H. L. 1934. Some additional fossil echini from Jamaica. Memoirs of rhe Museum of Comparative Zoology ai Harvard, v. 54, pp. 139-156.
Donovan, S. K. 1988. A preliminary biostrati- graphy of Jamaican fossil Echinoidea. In: Echinoderm Biology: Proceedings of the Sixth Internarional Echinoderm Conference, Victoria, British Columbia, August 23-28, 1987 (eds R. D. Burke, P. V. Mladenov, P. Lambert
and R. L. Parsley), pp. 125-131. A. A . Balkema, Rotterdam.
Donovan, S. K. & Bowen, J. F. 1989. Jamaican Cretaceous Echinoidea. 1. Introduction and reassessment of ?Pygopisres rudistarum (Haw- kins, 1923) n. comb. Mesozoic Research, v. 2 ,
Donovan, S. K., Gordon, C. M., SchicWer, W. F. & Dixon, H. L. 1989. An Eocene age for an outcrop of the ‘Montpelier Forma- tion’ at Beecher Town, St Ann, Jamaica, using echinoids for correlation. Journal of the GeologicalSociety ofJamaica, v. 26, pp. 5-9.
pp. 57-65.
(Arnold and Clark’s monographs are still avail- able, at very reasonable cost, from the Museum of Comparative Zoology. Back issues of the Journal of the Geological Society of Jamaica can be purchased from The Editor, c/o Department of Geology, University of the West Indies, Mona, Kingston 7, Jamaica.)
Stephen K . Donovan is Senior Lecturer in the Department of Geology, University of the West Indies, Jamaica.
The local geologist 3: The responsible collector
O n c e an honourable occupation undertaken by all with more than a passing interest in natural history, collecting is sometimes considered rather sinful in the present era of conservation awareness. In the late 1970s, owning a hammer seemed to be accompanied by a sense of guilt! At that time, geological conser- vation in the UK was in its infancy; no-one knew what’ resources were available in terms of exposures, fossils and minerals, how renewable they were, or how they were being damaged. One thing was certain; it was easy to imagine the damage being done by collectors. This is hardly surprising when many of us can probably remember wandering through some remote geological wilderness only to be greeted by a geological party pouring from a coach, hammers already swinging. Like an advancing ice sheet, they’d charge down valley and over hill, shattering and plucking from every exposed rock. Unfortunately, the moraines they deposited at the end of a day’s collect- ing remain to puzzle future collectors and curators.
It’s a phenomenon seen less often these days as the austere conservation message of the late 1970s and early 1980s has sunk home. However, conservation means different things to different people, and its interpretation by geologists has appeared to depend on status. The needs of amateurs seemed to be considered subordinate to those of academics even
though field-parties of the latter showed no greater sense of responsibility in their use of sites. Profes- sional collectors also suffered their share of unfair abuse. It was with the expectation of heated debate and possible fireworks that these seemingly disparate groups met at a major conference in London in 1987 to discuss the use and conservation of palaeontological sites. The outcome of the conference turned out to be quite dramatic for completely the opposite reason. T o everyone’s surprise, it revealed broad agreement among all parties; responsible collecting, in all its forms, was now recognized as an essential part of geological conservation. The geological community at large had already accepted the need for conservation; and the Nature Conservancy Council (NCC), after a decade of research,.had gained a thorough knowledge of what the real threats to sites were.
Later, the NCC made known its policy on fossil collecting, in the Geologists’ Association Circulur (no. 883, p. 20, 1990): ‘If palaeontological sites are to continue to have scientific relevance (rather than becoming a collection of historically interesting locations), further collecting of geological specimens and their study MUST be made possible . . . Fossil collecting per se cannot, in mosr circumsiunces, be considered to be an undesirable activity, whether it is for scientific, educational or commercial purposes’
106iGEOLOGY TODAY May-June 1991