sumario - aida-itea.org · indicadores del suelo y el cultivo. los resultados mostraron que en...
TRANSCRIPT

Sumario
Producción Vegetal
Evaluación de algunos indicadores microbiológicos en dos suelos arroceros de la pro-vincia de Camagüey, CubaAssessing of some microbial indicators in two cuban rice soilsP. Chaveli, L. Font, B.J. Calero, P. López, A.M. Francisco, R.D. Caballero, M. Valenciano 3
Eficiencia agronómica de seis sistemas de formación con la variedad de melocotón‘Merrill O’Henry®’ (Prunus persica (Batsch))Agronomical performance of cv. ‘Merrill O’Henry®' (Prunus persica (Batsch)) on sixtraining systemsR. Núñez, I. Iglesias, R. Montserrat, S. Alegre 13
El control fotobiológico de malas hierbas: revisión de una técnica controvertidaThe photocontrol of weeds: a review on a controversial techniqueA. Cirujeda, A. Taberner 27
Producción Animal
Análisis del progreso genético obtenido en el esquema de selección de la raza ovinamanchegaAnalysis of the genetic progress obtain in the selection program in Manchega sheepbreedJ.J. Jurado, M. Serrano, Mª D. Pérez-Guzmán 41
Parámetros cinéticos en eyaculados bovinos de toros de raza frisona y rubia gallegaMotion characteristics in bovine ejaculates from holstein and rubia gallega bullsR. Muiño, M. Fernández, A.I. Peña 55


P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12 3
Evaluación de algunos indicadores microbiológicos en dossuelos arroceros de la provincia de Camagüey, Cuba
P. Chaveli*, L. Font*, B.J. Calero**, P. López*, A.M. Francisco*, R.D. Caballero*,M. Valenciano*
*Estación Experimental de Suelos Camagüey. Cacocúm #11, Reparto Puerto Príncipe, Camagüey. E-Mail: [email protected].**Instituto de Suelos. Ministerio de la Agricultura, Apto, 8022, C.P. 10800, Capdevila Boyeros, La Habana, Cuba.
ResumenLa zona Sur de la provincia de Camagüey, se dedica al cultivo del arroz. En estos suelos se muestransíntomas de degradación con la consiguiente disminución de rendimientos provocados principalmen-te, además de otros efectos ambientales, por la salinidad. Esto perjudica las condiciones de vida de lamicroflora edáfica por lo que cualquier alteración de su actividad genera indicadores que pueden serutilizados para identificar el estado de degradación del suelo. El estudio se desarrolló en dos tipos desuelos, Ferrasol y Vertisol. El muestreo se realizó a una profundidad de 0-20 cm. Como índices de eva-luación se utilizaron la actividad respiratoria y celulolítica, la capacidad nitrificadora además de otrosindicadores del suelo y el cultivo. Los resultados mostraron que en ambos tipos de suelos, existe unestado microbiológico potencialmente degradativo, lo que trae consigo un deterioro de la fertilidad ypor tanto un desequilibrio del agroecosistema de estos suelos potencialmente salinos. Estas condicio-nes de deterioro y empobrecimiento del suelo constituyen un medio desfavorable para la mayoría delos microorganismos.
Palabras claves: Indicadores microbiológicos, degradación del suelo, salinidad, cultivo del arroz
SummaryAssessing of some microbial indicators in two cuban rice soilsThe South area of the Camagüey province, Cuba is dedicated to rice crops. These soils show degrada-tion symptoms with the rising fall of the agricultural yields and affectations of the environmentcaused mainly by the salinity. This harms the life conditions of the edafic microflora hence any alter-ation of its activity indicators can be used to identify the degradation state. This can be used to pro-mote a sustainable development of the soil resource. For this study two types of soils were selected:Ferrasol and Vertisol. The sampling was carried out till a depth of 0-20 cm. As evaluative indexes thebreathing activity, celulolitic and nitrifying capacity and other indicators of the soil and the cultivationwere used. The results showed that in both types of soils, a potentially degrading microbiologic statewas identified, this causes deterioration of the fertility and therefore an imbalance of the agroecosys-tem of these soils potentially saline. These conditions of deterioration and impoverishment of the soilconstitute an unfavorable environment for most of the microorganisms.
Key words: Microbial indicators, soil degradation, salinity, rice cultivation

4 P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12
Introducción
La degradación del suelo fue definida porMatlock (1976) como un proceso insidiosoque puede ser expresado como el efectoacumulativo de una serie de acciones, quepueden o no ser evidentes tales como lareducción y desaparición de la vegetación,mayor tasa de escorrentía y menor infiltra-ción de las precipitaciones, que conducen ala erosión creciente del suelo y la pérdida dela fertilidad.
La degradación inducida por el hombre,ocasiona la destrucción del potencial bioló-gico de los recursos naturales por el mal usoy manejo de los mismos (Ortiz et al., 1994),lo que trae como consecuencia la genera-ción de procesos degradativos del mediofísico, económico y social de las poblacionesinvolucradas y su entorno.
En la actualidad, en Cuba la degradación delos suelos es la principal causa de la disminu-ción de los niveles de producción por uni-dad de tierra cultivada, a diferencia de losaños 40 cuando la causa más importante delos bajos rendimientos eran las malezas(Durán, 1998).
Algunos de los procesos naturales y/o antró-picos que determinan la degradación delsuelo son la erosión, salinización, acidifica-ción, contaminación y el empantanamiento(Febles y Miranda, 1988). Otros factores queinfluyen en la degradación son el deteriorode la fertilidad debido a los bajos conteni-dos de materia orgánica, baja capacidad decambio catiónico, daños por salinidad yaltos contenidos de sodio, predominio deiones tóxicos y manto freático próximo a lasuperficie (Galvez et al., 1998).
La degradación de los suelos por efecto dela salinización se encuentra entre los pro-blemas más apremiantes en la crisis alimen-taria mundial. Para satisfacer las demandasactuales de producciones agrícolas, ha sido
necesario abordar cambios en la agricultu-ra, de extensiva a intensiva. Esto ha poten-ciado el uso de zonas muy frágiles desde elpunto de vista ecológico, que sin un ade-cuado manejo, se han degradado aún másrápidamente (Francisco, 2000).
La zona Sur de la provincia de Camagüey seencuentra dedicada al cultivo del arroz. Enestos, suelos el monocultivo desde hace másde 50 años es una de las causas fundamen-tales de su degradación y el principal cau-sante de su baja fertilidad (Navarro, 1998).Según estudios realizados en está regiónpor Francisco (2000), se han comprobadosíntomas de degradación del suelo y empo-brecimiento con la consiguiente caída de losrendimientos agrícolas. Las afecciones delentorno provocadas por la salinidad, sodici-dad y empantanamiento, incrementado porlos efectos adversos de la deforestación dellitoral costero y subcostero, mal drenaje ymanejo no diferenciado, coincide con lascausas de los procesos de degradación enCuba descritos por Socarrás (1998). Todosestos factores tienen un marcado efecto enlas regiones tropicales (Hernández et al.,1998) e influyen además en la disminuciónde la fertilidad, la sabanización y la destruc-ción mecánica y agrotécnica del suelo.
Para cuantificar los componentes lábiles dela materia orgánica (MO) se han propuestoun gran número de métodos, dentro de losque se encuentran los métodos de fraccio-namiento físico y los métodos biológicos.Estos últimos se basan en el análisis de lapoblación microbiana, que resulta ser elcomponente más activo y sensible al impac-to externo dentro del suelo y que define suscaracterísticas, especialmente en lo referen-te a su fertilidad, interviniendo en los pro-cesos de descomposición de residuos, reci-clado de nutrientes y transformaciones dela MO del suelo (Schnürer et al., 1985;Collins et al., 1992; Lobkov, 1999).

Las bases biológicas del manejo agrícolasegún Swift (1999), han permanecido hastahace poco como el equivalente de un esta-do pre-Mendeliano, ya que el conocimientode genética, fisiología, bioquímica y ecolo-gía de los organismos del suelo se encuen-tran poco explotadas para propósitos prác-ticos.
Los microorganismos del suelo pueden servistos como entes primariamente responsa-bles en el mantenimiento de la calidad delsuelo y por lo tanto el funcionamiento delos mismos puede servir como un marcadorbiológico sensible para la comprensión deun sistema dado. De esta manera Powlsonet al. (1987) señalan que los cambios en lapoblación microbiana provee evidenciasanticipadas de los cambios en la calidad delsuelo más que cualquier indicador de la cali-dad de la materia orgánica.
La determinación de indicadores microbio-lógicos es utilizada para evaluar el efecto delas prácticas de manejo agrícola sobre la fer-tilidad del suelo, y parecen reflejar con mássensibilidad los cambios en su calidad, quelas determinaciones químicas de carbono ynitrógeno (Franzluebbers et al., 1995; Beyer,1995). Estudios realizados por Chaveli et al.(2002) demostraron la eficacia de estos indi-cadores en la evaluación del estado degra-dativo del suelo en agroecosistemas poten-cialmente erosionados y el efecto de lasmedidas de conservación de suelo aplicadas.
La respiración es uno de los indicadores másantiguos y es frecuentemente usada paracuantificar la actividad microbiana delsuelo. El uso de este indicador microbiológi-co unido a otros como la respiración induci-da por sustratos, la nitrificación y la descom-posición de la celulosa, han permitidoestimar la actividad de la biomasa y comoésta es influenciada por propiedades físicasy químicas o prácticas de manejo agrícola(Campbell et al., 1992; Calero et al., 1999;
Font, 1999; Font et al., 2002; Chaveli et al.,2002; Font et al., 2003; Chaveli et al., 2003).
De esta forma la actividad microbiana delsuelo constituye una medida fundamentalde importancia ecológica, puesto que poruna parte representa el nivel de actividadbiológica, involucrando el componente lábilde la MO y por otro, integra los factores delmedio ambiente y su influencia en la misma(Zagal et al., 2002).
El objetivo planteado en este trabajo fue lautilización de indicadores microbiológicos,para evaluar el efecto del manejo agrícolaempleado para el cultivo del arroz en dostipos de suelos en la zona Sur de la provin-cia de Camagüey.
Materiales y métodos
El estudio se desarrolló en la zona Sur de laprovincia de Camagüey, en la Unidad Básicade Producción Cooperativa (UBPC) “RodolfoRamírez Esquivel” área de producción dearroz perteneciente al municipio de Florida,provincia de Camagüey. Los suelos que pre-dominan en esta región son los Ferrasoles yVertisoles (USDA, 1994) y se encuentranafectados por la salinidad con una conducti-vidad eléctrica media en extracto de satura-ción promedio de 3 dS·m-1, clasificándosecomo moderadamente salinos.
En ambos tipos de suelo el manejo utilizadofue de siembra en seco con mejoramiento,que consiste en 13 labores mecanizadas depreparación de suelo sin lámina de agua y laaplicación de residuos de cosechas (5-8kg·m-2 de masa verde) 60 días antes de lasiembra. Se utilizó para el estudio la varie-dad de arroz J- 104 (Unión del Arroz, 2001)de ciclo medio de crecimiento y la densidadde siembra fue de 15 a 20 x 105 plantas/ha.
P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12 5

El muestreo se efectuó cuando el suelo seencontraba sin el cultivo establecido a laprofundidad de 0-20 cm, tomando muestrascompuestas en cada uno de los campos,durante tres años consecutivos. Las mues-tras para analizar se secaron al aire y sepasaron por un tamiz con malla de 2 mm(Calero et al., 1999).
Se utilizaron como índices evaluativos larespiración basal (RB), la respiración al aña-dir una fuente carbonada y nitrogenada defácil degradación (RIC, RICN), la capacidadnitrificadora real y potencial del suelo (NR yNP) y la actividad celulolítica (DC).
La RB se determinó utilizando el sistema defrasco cerrado propuesto por Isemeyer(1952), por humedecimiento de 25 g desuelo al 60% de la capacidad máxima deretención de humedad, determinada segúnForster (1995) y la determinación del CO2 alcabo de 24 horas de incubación a 30 ºC. Lassoluciones utilizadas fueron las propuestaspor Dommerguez (1960). Para la determina-ción de la RIC, se aplicó glucosa a razón de8,4 mg/g de suelo para los Ferrasoles y10mg/g de suelo para los Vertisoles en solu-ción acuosa y para la medida de la respira-
ción inducida con carbono y nitrógeno(RICN), el nitrógeno requerido para satisfa-cer la demanda de la biomasa microbianaen función del carbono se calculó conside-rando una relación C/N media para la bio-masa no superior a 10 y asumiendo que sóloel 40% del C que se añade pasa a formarparte de la biomasa microbiana (Kelley yStevenson, 1985).
La capacidad nitrificadora (NR) se determi-nó por incubación de 25 g de suelo durante15 días según Bolotina y Abramova (1968).La extracción de nitrato se realizó con K2SO4al 0,01% y se utilizó ácido disulfofenólicopara el desarrollo del color. Para la nitrifica-ción potencial (NP) se adicionó sulfato deamonio al 1% respecto al peso del suelo. Elgrado de descomposición de la celulosa(DC) se evaluó en tubos de ensayo utilizan-do tiras de papel de celulosa según Szegui(1988).
La tabla 1 recoge las principales característi-cas químicas y los rendimientos de las cose-chas realizadas en la campaña correspon-diente al momento de iniciar el estudio, enlos Ferrasoles y Vertisoles.
6 P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12
Tabla 1. Características químicas y rendimientos del cultivo en los dos tipos de suelos antes decomenzar el estudio
Table 1. Soil chemistry characteristics and crop yields in both soil types before beginning the study
pH (KCl) M.O. P2O5 K2O Rdtos% mg 100 g s-1 t·ha-1
Ferrasol 5,6 1,47 4,49 12,70 3,51Vertisol 5,5 1,77 9,14 11,09 2,92
Resultados y discusión
Los resultados obtenidos en la caracteriza-ción del estado microbiológico del Ferrasolindican que la tecnología de manejo utiliza-
da ha provocado el establecimiento de unazona microbiológicamente activa. Como seaprecia en la figura 1 la RB, índice de activi-dad potencial básica (Alef y Nannipieri,1995), es alta y muestra valores similares e

incluyo mayores a los obtenidos por otrosautores en este tipo de suelo (Alfonso et al.,1998; Calero et al., 1999); lo cual denota lapresencia de un mayor número de microor-ganismos viables y de reservas energéticascapaces de sustentar el metabolismo de losmismos, debido a que la cantidad de CO2desprendido expresa la microflora presenteen estado de latencia la cual se activa utili-zando el carbono (C) y nitrógeno (N) bioló-
gicamente disponible en las estructurasorgánicas del suelo, una vez que al mismose le proporciona las condiciones óptimasde temperatura y humedad después desecado al aire (Calero et al., 1999); por con-siguiente la actividad respiratoria es consi-derada un indicador directo de la actividadmicrobiana e indirecto de los contenidos deMO del suelo (Mattew y Daviel, 1990; Leitaet al., 1995; Cheng et al., 1997).
P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12 7
Figura 1. Comportamiento de la actividad respiratoria en el suelo Ferrasol. Figure 1. Behavior of the breathing activity in the Ferrasol soil.
La alta población microbiana existente en elsuelo, unido al bajo contenido de materiaorgánica determinado químicamente (tabla1), no adecuado en este tipo de suelo, indi-ca un estado microbiológico potencialmen-te degradativo ya que los microorganismos,al necesitar material orgánico para su meta-bolismo, ataca la fracción más estable deéste, lo cual provoca una degradación pro-gresiva y lenta de la materia orgánica nati-va del suelo. Estos resultados demuestranque el manejo al cual ha sido sometido estesuelo no ha sido el mas adecuado
Cuando la respiración es inducida por sus-tratos, puede identificar la porción de labiomasa microbiana que está activa bajo
concentraciones conocidas de C y N (Ander-son, 1982), sin que la misma se encuentrelimitada por dichas reservas. Estos indicado-res se incrementaron con respecto a la RB(figura 1), o sea, existe una respuesta de lamicroflora a la adición de fuentes externasde C fácilmente degradable, como la gluco-sa (RIC) y de N (RICN). Este comportamientoindica la presencia de un mayor número demicroorganismos, pero los mismos seencuentran limitados por deficientes reser-vas energéticas en la MO.
La capacidad nitrificadora (NR y NP) indicala capacidad microbiológica del suelo paradesarrollar los procesos de mineralizaciónde las formas orgánicas y amónicas de
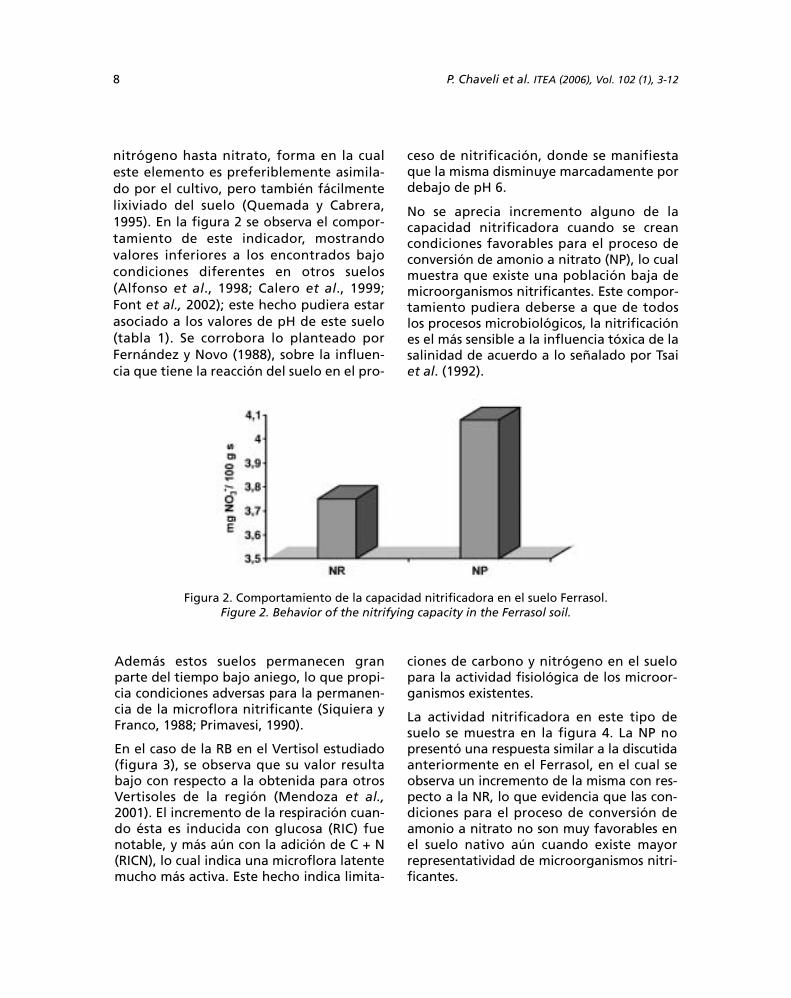
nitrógeno hasta nitrato, forma en la cualeste elemento es preferiblemente asimila-do por el cultivo, pero también fácilmentelixiviado del suelo (Quemada y Cabrera,1995). En la figura 2 se observa el compor-tamiento de este indicador, mostrandovalores inferiores a los encontrados bajocondiciones diferentes en otros suelos(Alfonso et al., 1998; Calero et al., 1999;Font et al., 2002); este hecho pudiera estarasociado a los valores de pH de este suelo(tabla 1). Se corrobora lo planteado porFernández y Novo (1988), sobre la influen-cia que tiene la reacción del suelo en el pro-
ceso de nitrificación, donde se manifiestaque la misma disminuye marcadamente pordebajo de pH 6.
No se aprecia incremento alguno de lacapacidad nitrificadora cuando se creancondiciones favorables para el proceso deconversión de amonio a nitrato (NP), lo cualmuestra que existe una población baja demicroorganismos nitrificantes. Este compor-tamiento pudiera deberse a que de todoslos procesos microbiológicos, la nitrificaciónes el más sensible a la influencia tóxica de lasalinidad de acuerdo a lo señalado por Tsaiet al. (1992).
8 P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12
Figura 2. Comportamiento de la capacidad nitrificadora en el suelo Ferrasol.Figure 2. Behavior of the nitrifying capacity in the Ferrasol soil.
Además estos suelos permanecen granparte del tiempo bajo aniego, lo que propi-cia condiciones adversas para la permanen-cia de la microflora nitrificante (Siquiera yFranco, 1988; Primavesi, 1990).
En el caso de la RB en el Vertisol estudiado(figura 3), se observa que su valor resultabajo con respecto a la obtenida para otrosVertisoles de la región (Mendoza et al.,2001). El incremento de la respiración cuan-do ésta es inducida con glucosa (RIC) fuenotable, y más aún con la adición de C + N(RICN), lo cual indica una microflora latentemucho más activa. Este hecho indica limita-
ciones de carbono y nitrógeno en el suelopara la actividad fisiológica de los microor-ganismos existentes.
La actividad nitrificadora en este tipo desuelo se muestra en la figura 4. La NP nopresentó una respuesta similar a la discutidaanteriormente en el Ferrasol, en el cual seobserva un incremento de la misma con res-pecto a la NR, lo que evidencia que las con-diciones para el proceso de conversión deamonio a nitrato no son muy favorables enel suelo nativo aún cuando existe mayorrepresentatividad de microorganismos nitri-ficantes.

La actividad celulolítica (DC) expresa lapotencialidad de la microflora para degra-dar compuestos carbonados complejos. Elcomportamiento de este indicador enambos tipos de suelo se muestra en la tabla2. Este índice tuvo una mayor degradaciónde esta fuente carbonada en el Ferrasolaunque sus valores son muy inferiores a losreportados por varios autores en ese mismoperíodo de evaluación (Font, 1999; Font etal., 2002; Chaveli et al., 2002).
En sentido general, existe degradación de latierra en ambos suelos estudiados debido a
un deterioro de la fertilidad, no sólo comoconsecuencia de los bajos contenidos demateria orgánica como índice de degrada-ción biológica (Gálvez, et al., 1998), sinotambién desde el punto de vista de su esta-do y actividad microbiológica. El cultivocontinuado del arroz unido a un manejoinadecuado de las aguas de riego, tecnolo-gías de manejo no acordes a sus condicionesfísicas, químicas y biológicas, los procesos desalinización y la deforestación del litoralcostero, han llevado a que en ambos tiposde suelos se hayan agotado elementosenergéticos que no permitan la expresión
P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12 9
Figura 3. Comportamiento de la actividad respiratoria en el suelo Vertisol. Figure 3. Behavior of the breathing activity in the Vertisol soil.
Figura 4. Comportamiento de la actividad respiratoria en el suelo Vertisol. Figure 4. Behavior of the breathing activity in the Vertisol soil.

de la microflora presente, resultando bajala población microbiana con respecto a lareportada en estos tipos de suelos bajo dife-
rentes condiciones de manejo (Alfonso etal., 1998; Calero et al., 1999; Font, 1999;Font et al., 2002; Chaveli et al., 2002).
10 P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12
Tabla 2. Comportamiento de la actividad celulolítica de ambos tipos de suelosTable 2. Celulolitic activity in both types of soils
Suelo DC (%) DC (%) DC (%) vDC/ 7días 14 días 21 días día(%)&
Ferrasol 13,29 23,41 57,29 2,73Vertisol 1,91 5,3 34,37 1,67
&Velocidad de descomposición de la celulosa por día.
Conclusiones
Los suelos Ferrasol y Vertisol cultivados conarroz, reflejan la degradación potencial desu estado microbiológico, lo que trae consi-go un deterioro de la fertilidad y por tantoun desequilibrio del agroecosistema.
El manejo inadecuado de los suelos, afecta-dos entre otros factores por la salinidad,constituyen un medio desfavorable para lamayoría de los microorganismos, principal-mente los nitrificadores.
Bibliografía
Alef K, Nannipieri P, 1995. Methods in appliedsoil microbiology and biochemistry. 575 pp.Ed. Academic Press Inc. San. Diego (EEUU).
Alfonso CA, Calero B, Morejón L, Somoza V, Gue-rrero A, Monedero M, 1998. Rehabilitación deFerrasol degradados por compactación delSur de la Habana. Memorias del 16 Cong.Inter. de la Ciencia del Suelo. Vol. III. 561 pp.Montpellier, (Francia).
Anderson J, 1982. Soil respiration. Methods ofsoil analysis (segunda edición) 9 pp. Part. 2,
Ed. Agron. Monog., Amer. Soc. Agron, Madi-son. (EEUU).
Beyer L, 1995. Soil microbial biomass and organicmatter composition in soils under cultivation.Biol. Fertil. Soils. 19: 197-202.
Bolotina NI, Abramova EA, 1968. Método paradeterminar la capacidad nitrificadora delsuelo. [en ruso]. Agrojimia. 4: 136- 141.
Calero B, Guerrero A, Alfonso CA, Somoza V,Camacho E, 1999. Efecto residual de la fertili-zación mineral sobre el estado microbiológicodel suelo. La Ciencia y el hombre. Vol XI 33:89-94.
Campbell CA, Brandt SA, Biederbeck VO, Zent-ner RP, Schnitzer M, 1992. Effect of crop rota-tions and rotation phase on characteristics ofsoil organic matter in a Dark Brown Cherno-zemic Soil. Can. J. Soil Sci. 72: 403-416.
Chaveli P, Font L, Gandarilla JE, Calero B, Valen-ciano M, 2002. Estudio microbiológico de unsuelo Pardo sin Carbonatos propenso a la ero-sión para un manejo agrícola sostenible. Cen-tro Agrícola. 4: 66-71.
Chaveli P, Font L, Calero B, Francisco AM, LópezP, Caballero R, Valenciano M, 2003. Uso deindicadores microbiológicos para la evalua-ción de la degradación de suelos Oscuros Plás-

ticos arroceros de la provincia de Camagüey.Centro Agrícola. 3: 61-66.
Cheng T, Killham K, Malcoms S, 1997. Dynamicresponse of microbial biomass respiration rateand ATP to glucose additions. Soil Biol. Bio-chem. 29(8): 1249-1256.
Collins HP, Rasmussen PE, Douglas CL, 1992. Croprotation and residue management effects onsoil carbon and microbial dynamics. Soil Sci.Soc. Am. J. 56: 783-788.
Dommerguez Y, 1960. La notion de coefficientde mineralization du carbone dans les sols. Unexample d´utilization des tecniques biologi-ques. Agron. Trop. 1(1): 54-60.
Duran JL, 1998. Degradación y manejo ecológicode los suelos tropicales con énfasis en los deCuba. Agricultura Orgánica. 1: 7-10.
Febles JM, Miranda R, 1988. La protección de lossuelos. Monografía. 148pp.Instituto Superiorde Ciencias Agropecuarias de La Habana. LaHabana. Cuba.
Fernández C, Novo R. 1988. Vida microbiana enel suelo. 1ª y 2ª Parte. 525pp. Ed. Pueblo y Edu-cación. Cuba.
Font L, 1999. Uso de indicadores microbiológicospara evaluar el efecto a largo plazo de la ferti-lización mineral en un agroecosistema citríco-la. Tesis presentada en opción al título deMaster en Ciencia. 68pp. Universidad deCamagüey, Camagüey. (Cuba).
Font L, Calero B, del Castillo A, 2002. Estadomicrobiológico del suelo, base del manejointegral de un agroecosistema citrícola. LEISA.Rev. de Agroecología. 18(3): 21-22.
Font L, Chaveli P, Calero B, Francisco AM, LÓPEZP, Caballero R, Valenciano M, 2003. Impactode algunos procesos degradativos sobre elestado microbiológico de un suelo Ferrasolcultivado de arroz. Centro Agrícola. 3: 67-72.
Forster JC, 1995. Soil physical analysis. Determi-nation of the gravimetric water content andsoil dry mass. pp. 105-116. En: Methods inapplied soil microbiology and Biochemistry. K.Alef and P. Nannipieri. (Ed). 575 pp. AcademicPress Inc. San. Diego. (EEUU).
Francisco AM, 2000. Nuevos indicadores para lacaracterización y el diagnóstico de la salinidadde los suelos de Cuba. Tesis presentada enopción al título de Master en Ciencias. 80 pp.Universidad de Camagüey. Camagüey. (Cuba).
Franzluebbers A, Zuberer D, Hon’s F, 1995. Com-parison of microbiological methods for eva-luating quality and fertility of soil. Biol. FertilSoils. 19: 135-140.
Gálvez V, Otero L, Navarro N, 1998. Reflexionessobre los suelos de la llanura sur de Pinar delRío. Agricultura Orgánica. 1: 14-15.
Hernández C, Arteaga O, Peña JL, Peña F, CancioT, León G, 1998. El establecimiento de pastoscomo alternativa eficaz contra la erosión delsuelo en la región “Escambray” AgriculturaOrgánica. 1: 21-22.
Isemeyer H, 1952. Eine einfache Methode zurBestimmung der Bodenatmung und der Kar-bonate im Boden. pp. 215-217. En: Methods inapplied soil microbiology and Biochemistry. K.Alef and P. Nannipieri. (Ed). 575 pp AcademicPress Inc. San. Diego. (EEUU).
Kelley KR, Stevenson FJ, 1985. Characterizationand extractability of immobilized 15N Fromthe soil microbial biomass. Soil Biol. Biochem.17(4): 517-523.
Leita L, de Nobly M, Muhubachova G, Mondini C,Mardual L, Zerbi G, 1995. Bioavailability andeffects of heavy metals in soil microbial bio-mass, survival during laboratory incubation.Biol. Fertil Soils. 19: 103-108.
Lobkov VT, 1999. Biodiversity in agroecosystemsas a factor optimizing the biological activityof soil. Eurasian Soil Science. 32: 664-668.
Matlock W, 1976. Segments of vicious circle:Land degradation and water resources. Deser-tification process. Problems and Perspectives.pp. 45-50. University of Tueson, Arizona.(EEUU).
Matthew R, Daviel D, 1990. Effects of conversionto organic agricultural practices on soil biota.Amer. Jour of Altern. Agric. 5(1): 26-32.
Mendoza L, Pacheco O, Vento M, Francisco A,Sánchez M, 2001. La pérdida de suelo en los
P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12 11

terrenos con relieve llano a ondulado. CentroAgrícola. 4:19-22.
Navarro N, 1998. Degradación de los suelos pro-vocado por el monocultivo en la Granja arro-cera Caribe. Agricultura Orgánica. 1: 16-17.
Ortiz ML, Ojeda A, Estrada J, 1994. Evaluación,cartografía y políticas preventivas de la degra-dación de la tierra. 161 pp. Madrid, (España).
Powlson DS, Brookes PC, Christensen BT, 1987.Measurement of soil microbial biomass provi-des an early indication of changes in total soilorganic matter due to straw incorporation.Soil Biol. Biochem. 19: 159-164.
Primavesi A, 1990. Manejo Ecológico do Solo. Aagricultura em regioes tropicais. (novena edi-ción). 549 pp. Nobel. (Brasil).
Quemada M, Cabrera M, 1995. CERES-N modelpredictions of nitrogen mineralized fromcover crop residues. Soil Sci. Soc. Am. J. 59:1059-1065.
Schnürer J, Clarholm M, Rosswall T, 1985. Micro-bial biomass and activity in an agricultural soilwith different organic matter contents. SoilBiol. Biochem. 17: 611-618.
Siqueira J, Franco V, 1988. Biotecnología do Solo:Fundamentos e Perspectivas. 36 pp: Ed. Minis-terio da Educacâo. ABEAS; Lavras: ESAL.FAEPE Brasilia (Brasil).
Socarrás A, 1998. La vida del suelo: un indicadorde su fertilidad. Agricultura Orgánica. 1: 12-13.
Swift MJ, 1999. Towards the second paradigm:integrated biological management of soil. En:Soil Fertility, Soil Biology and Plant NutritionInterrelationships. pp. 11-24. J.O. Siqueira,F.M. Moriera, A.S. López, L.R.G. Guilherme, V.Faquin, A.E. Furtini Neto, J.G. Carvalho (Eds).SBCS, Lavras. UFLA/DC. (Brasil).
Szegui J, 1988. Cellulose decomposition and soilfertility. 241 pp. Akademiai Kiadó, Budapest,(Hungria).
Tsai SM, Baraibar AVL, Romani VLM, 1992. Efeitode fatores do solo. Salinidade e pressào osmó-tica. Em: Microbiología do solo. pp. 59-72.Cardoso E.J.B.N.; Tsai, S.M. y Neves, M.C.P.(Eds). Sociedade Brasileira de Ciencia do solo.(Brasil).
Unión del Arroz, 2001. Instructivo Técnico delArroz. Segunda Edición. 119 pp. Ministerio dela Agricultura. La Habana, Cuba.
USDA, 1994. Keys to soil taxonomic. 305 pp. Soilconservation service. Sixth Edition.
Zagal E, Rodríguez N, Vidal I, Quezada L, 2002.Actividad microbiana en un suelo de origenvolcánico bajo distinto manejo agronómico.Agricultura Técnica. 62(2): 297-309.
(Aceptado para publicación el 28 de enero de2006).
12 P. Chaveli et al. ITEA (2006), Vol. 102 (1), 3-12

R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26 13
Eficiencia agronómica de seis sistemas de formación con la variedad de melocotón ‘Merrill O’Henry®’ (Prunus persica (Batsch))
R. Núñez*, I. Iglesias**, R. Montserrat**, S. Alegre**
* Colegio de Postgraduados, Campus Puebla, México.** Institut de Recerca i Tecnologia Agroalimentaries (IRTA). Estació Experimental de Lleida. AvdaAlcalde Rovira Roure, 191. 25196-LLEIDA
ResumenDurante 9 años se ha estudiado en melocotonero, variedad ‘Merrill O’Henry®’/GF-305, el efecto de 6sistemas de formación sobre el manejo, eficiencia productiva, producción, calidad, interceptación dela luz e incidencia de plagas y enfermedades. La plantación se ubica en la finca de Gimenells del IRTA-Estación Experimental de Lleida (NE-España). Los resultados obtenidos indican que la densidad deplantación ha afectado al vigor del árbol y ha condicionado la rapidez de entrada en producción. Alnoveno año de plantación, los sistemas de mayor densidad, Ypsilon y Eje, fueron los más productivos.Sin embargo el sistema Tatura, con menor densidad de plantación, presentó mayor producción que laPalmeta. Los sistemas que presentaron el valor más alto de eficiencia productiva acumulada fueron elTatura, el Vaso y la Palmeta, mientras que los sistemas con mayor densidad obtuvieron un valor másbajo. La mayor interceptación de luz correspondió al Tatura, el Ypsilon y el Vaso y la menor a la Pal-meta, el Eje y el Doble Y. Respecto a los parámetros de calidad de la fruta, sólidos solubles, firmeza yacidez, no se han dado diferencias significativas entre sistemas. Los sistemas que mostraron los mayo-res calibres fueron el Doble Y, la Palmeta y el Vaso; siendo los menores para el Ypsilon, el Eje y el Tatu-ra. El sistema-densidad afectó a la eficacia de los tratamientos fitosanitarios, presentando mayor inci-dencia de plagas y enfermedades el Tatura y los de menor afección el Eje y la Palmeta.
Palabras clave: vigor, precocidad, eficiencia, producción, calibre, calidad, interceptación de luz,enfermedades.
SummaryAgronomical performance of cv. ‘Merrill O’Henry® ' (Prunus persica (Batsch)) on six training systems The study has been carried out over a nine years period in order to evaluate the effects of 6 training sys-tems on production, quality, light interception, pest incidence and diseases by using the cv. ‘Merrill O’Hen-ry®’ on GF-305 rootstock. The experimental orchard is located at the IRTA’s Experimental Station of Lleida,plot of Gimenells (NE-Spain). The yields obtained suggested a relationship between tree density and treevigour/early bearing. Based on cumulative yields, Y-shape and Axis were the most productive, due to thehigher density. The best cumulative yield efficiency, was obtained with Tatura, Gobelet and Palmette,whereas high density systems provided lower values. Fruit quality, soluble solids contents and titratableacidity were not affected by training systems. The highest percentages of commercial fruit size wereobtained with Double Y, Palmette and Gobelet; whereas Y-shape, vertical Axis and Tatura provided thelowest values. Tatura, Y-shape and Gobelet were the most efficient on light interception. Tree shape anddensity influenced pest control management, due to the direct effect of training systems on spray effi-ciency; thus, Tatura was the most affected by diseases and Palmette and Axis the lowest ones.
Key words: vigour, precocity, efficiency, production, fruit size, quality, light, interception, diseases.

14 R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26
Introducción
El melocotonero es la tercera especie defruta dulce más producida a escala mundial,después del manzano y al peral. En los últi-mos años 30 años, la producción se ha incre-mentado en un 211 %, con un fuerte aumen-to en la última década, ocupando en el año2001 una superficie de 1.814.000 ha con unaproducción de 13,3 millones de toneladas(Carbó e Iglesias, 2002).
El cultivo del melocotonero ha experimenta-do una notable evolución en lo referido avariedades, patrones, marcos de plantacióny técnicas culturales. En las principales espe-cies frutícolas se ha dado un proceso deintensificación de las plantaciones, pues seacepta que, dentro de un cierto límite, laentrada en producción es proporcional alnúmero de plantas por hectárea o densidad.Lo anteriormente expresado es válido paracualquier especie frutal pero, en el caso par-ticular del melocotonero, es necesario teneren cuenta que es una especie muy exigenteen luz, de forma que los sombreamientosproducen secado de brotes, aborto deyemas, caídas de hojas, pérdida de calidadde la fruta y también acortan el período pro-ductivo con respecto a otras especies fruta-les (Royo y Martínez, 1992). Por otra parte,en esta especie, no se dispone de patronesenanizantes que permitan un elevado gradode intensificación, en comparación con otrascomo el manzano el peral o el cerezo. Es porello, que los sistemas de formación más utili-zados en los principales países productoresson del tipo vaso con densidades de planta-ción entre 400 y 800 árboles por hectárea.
La forma de los árboles y la densidad deplantación condicionan la rapidez de entra-da en producción e influyen sobre la pro-ducción y las necesidades de obra (Delgado,2000). La calidad de los frutos adquierecada día mayor importancia, llegando a seruno de los objetivos preferentes de la pro-
ducción. Las explotaciones frutales planifi-can su proceso productivo con la finalidadde conseguir frutos de calidad para satisfa-cer las exigencias del sector comercial y delos consumidores.
El presente trabajo tiene como finalidadvalorar en melocotonero la eficiencia agro-nómica de seis sistemas de formación parahacer la elección del sistema más adecuado,buscando: precocidad de entrada en pro-ducción, fácil manejo, alta producción ycalidad de la fruta.
Material y métodos
En enero de 1995, se estableció una parcelacon la variedad de melocotonero ‘MerrillO‘Henry®’ injertado sobre patrón GF-305 enla Finca de Gimenells del IRTA-EstaciónExperimental de Lérida, España (41.658º N,0.393º E y 248 m de altitud). El presente tra-bajo se realizó durante los años 2001, 2002y 2003. A pesar de ello y con el objeto depoder realizar un estudio global de la evo-lución de diferentes variables (vigor, preco-cidad, producción, calidad, etc.), se han uti-lizado los datos disponibles de todo elensayo, es decir desde 1995. Los sistemas deformación evaluados así como el marco deplantación, densidad por hectárea, númerode árboles por fila de la parcela elemental,así como la necesidad de la estructura deapoyo se exponen en la tabla 1. El diseñoexperimental fue en bloques al azar contres repeticiones. En cada uno de los siste-mas, la parcela elemental estuvo constituidapor tres filas de árboles de 38,5 m de largosiendo la fila central la utilizada para la rea-lización de los diferentes controles realiza-dos. La orientación de las filas de los árbolesfue en dirección Noroeste-Sureste.
Las variables determinadas fueron las si-guientes: vigor, producción, peso medio y

Tabla 1. Marco de plantación, densidad y necesidad de estructura de apoyo de los diferentes sistemasde formación evaluados
Table 1. Planting distance, density, and needs of support structure for different training systems
Sistema Marco de Densidad Parcela elemental Estructura de plantación (m) (árboles/ha) (árboles/fila) apoyo
Ypsilon 5,5 x 1,75 1.038 22 NoEje 4,5 x 1,75 1.270 22 SíVaso 5,5 x 3,5 519 11 NoDoble Y 5,5 x 3,5 519 11 NoPalmeta 4,5 x 3,5 635 11 SíTatura 5,5 x 3,5 519 11 Sí
calibre del fruto, radiación fotosintética-mente interceptada (PAR) e incidencia deenfermedades. El vigor, fue determinadoanualmente desde 1997 a 2003, y se haexpresado como sección del tronco a 20 cmpor encima del punto de injerto. La recolec-
ción se realizó a partir del 20 de agosto, endos pasadas. La eficiencia productiva o índi-ce de productividad en kg/cm2 (I.P.) se calcu-ló a partir de 1997 de forma anual y acumu-lada hasta el 2003, en base a la producción(kg/árbol) y a la sección del tronco (cm2).
R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26 15
Por cada sistema y parcela elemental, la pro-ducción se recolectó en dos pasadas, siendola primera la mas importante, estableciéndo-se como criterio de cosecha el color sobre almenos el 50% de la superficie del fruto y elcalibre superior a 75mm, por debajo de estese consideró no comercializable. De la prime-ra pasada, se tomaron 16 kg de frutos deforma aleatoria, tanto de la parte alta comode la parte baja del árbol, para determinar elpeso medio del fruto, el número de frutospor categoría comercial de calibre (<70 mm–no comercial–, 70-75, 75-80, 80-85 y>85mm), el calibre medio ponderado de losdiferentes sistemas en el período 1997-2002 yel porcentaje de producción comercial. De los16 kg de cada sistema de formación y repeti-ción se escogieron al azar 20 frutos sobre loscuales se determinaron anualmente los con-tenidos de sólidos solubles, la firmeza y laacidez titulable, en base a la metodologíapropuesta por Carbó e Iglesias (2002).
En el año 2003, noveno año de plantación, sedeterminó la incidencia de enfermedades en
la parte baja y alta de los árboles. Para ello ysobre dos árboles de cada sistema y repeti-ción, situados en el centro de la línea centralde cada parcela elemental, se contó el núme-ro de brotes afectados por Pseudomonassyringae y en cada brote afectado el númerode hojas con daños por dicha enfermedad. Elmismo procedimiento se utilizó para deter-minar los daños causados por Taphrinadeformans (Burk).
La influencia de los sistemas de formación enla interceptación de la radiación solar, sedeterminó mediante un ceptómetro SunScan SS1-UM-1.05 (Delta-T Devices Ltd Cam-bridge, UK) con 64 sensores fotodiodos dis-puestos linealmente en una lanza de 100 cmde longitud. La medición lumínica corres-pondió a la Radiación FotosintéticamenteActiva (PAR entre 400 y 700 nm). Para efec-tuar las mediciones la lanza se colocó en lamisma dirección que la fila de los árboles y seregistro el valor promedio de todos los sen-sores de la barra. Se realizaron cuatro medi-das mensuales en el período mayo-agosto de

2003. Para cada sistema, por repetición, en-tre la fila central y la adyacente en direcciónsur se marcaron 20 puntos, a una altura delsuelo de 30 cm, tal como se observa en figura1, donde se realizaron las correspondientesmedidas del PAR.
El análisis estadístico de las variables estu-diadas se realizó mediante el procedimien-to ANOVA del paquete estadístico SAS,Versión 8.1 (SAS, 2002), estableciéndose la
significación estadística para P=0,05. Cuan-do el análisis fue estadísticamente signifi-cativo para las diferentes variables evalua-das (F-test), se utilizó el test del RangoMúltiple de Duncan para la separación delas medias, estableciéndose un nivel de sig-nificación del 5 %. Las variables medidasen porcentaje se transformaron mediantearcoseno raíz cuadrada para su normaliza-ción, antes de realizar el análisis de lavarianza.
16 R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26
Figura 1. Esquema de la metodología utilizada para la determinación de la radiación interceptada porlos diferentes sistemas de formación mediante el ceptómetro. Cada punto indica un lugar de lectura.
Figure 1. Schema of methodology used to determinate the radiation intercepted by the differenttraining systems by the use of a ceptometre. Each point represents a place of measure.
Resultados y discusión
Vigor
A partir del 3er año y hasta el noveno, lossistemas de baja densidad (Vaso, Doble Y yTatura), presentaron la mayor seccióntransversal del tronco (STT), con respecto alos de mayor densidad, dándose diferen-cias significativas entre los mismos (figura
2). Ello puede ser debido a que en dichossistemas para su formación durante losprimeros años, en la poda de invierno serealizaron intervenciones más severas queen el resto de los sistemas. Dichas inter-venciones estimularon la actividad vegeta-tiva y retrasaron la entrada en producción,con el consiguiente incremento de vigor,con respecto a los sistemas de mayor den-

sidad como el Ypsilon y Eje. Destacar queen el Doble Y, para mantener la altura de
2,5 m, se utilizó la poda en verde sistemáti-camente.
R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26 17
Espada et al. (2000) evaluaron los sistemasde Vaso y Eje en melocotonero con lasvariedades ‘Jesca’, ‘Calante’, ‘CA-8’ y ‘Evai-sa’, injertados sobre GF-677, con una densi-dad de 277 y 555 árboles/ha, siendo tam-bién el vigor mayor en el vaso, por loanteriormente expuesto. Corelli y Sansavini(1991) reportan que la sección transversaldel tronco en melocotonero al cuarto año,para las variedades ‘Flavorcrest’ y ‘Rubyred’,fue mayor en Vaso (102 cm2), Palmeta (94cm2), Eje (63 cm2) y el Ypsilon (57 cm2) a unadensidad constante de 729 árboles/ha,obteniendo secciones similares a las del pre-sente ensayo.
Precocidad de entrada en producción yProducción total Acumulada
Como en otras especies, en melocotonero,la rapidez de entrada en producción ha
estado directamente relacionada con ladensidad de plantación, independiente-mente de la forma asociada a cada siste-ma, tal como se observa en la tabla 2. Elsistema en Eje, él de mayor densidad, fueel que presentó la más alta producciónacumulada en los primeros tres años(1996-1998). En el año 1999 no hubo pro-ducción debido a una fuerte granizada aprincipios del mes de mayo. En el 2000 nohubo diferencias entre sistemas, y estasfueron pequeñas en el año 2001. Mientrasen los años 2002 y 2003 los sistemas quealcanzaron la mayor producción fueron elYpsilon y el Tatura, éste último de bajadensidad de plantación, comportándose elresto de forma similar.
Al noveno año de cultivo los sistemas demayor producción acumulada fueron elYpsilon y el Eje. Sin embargo, el sistemaTatura, con menor densidad, presentó
Figura 2. Evolución de la sección transversal del tronco (cm2) de diferentes sistemas de formación enmelocotonero a la largo del período 1997-2003. Los puntos corresponden a la media de tres
repeticiones. Las barras verticales indican la LSD (P≤0,05) para cada año.Figure 2. Evolution of the trunk cross-sectional area (cm2) of several peach training systems over theperiod 1997-2003. Each point is the mean of 3 replicates. Vertical bars indicate the LSD (P≤ 0,05) for
each year.
Año
ST
T(c
m2)

mayor producción que la Palmeta, tal comose observa en la tabla 3. Es por ello que lasproducciones acumuladas en el periodo1996-2003 no han mostrado una relación
directa con la densidad de plantación, y sehan relacionado más con el volumen finalde copa de los árboles (datos no expuestos)y con la interceptación de la luz.
18 R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26
Una experiencia realizada en Francia conlas variedades ‘Manon’ y ‘O’Henry®’ con-ducidos en Doble Y y en Ypsilon; puso demanifiesto que el Doble Y presentó mejorcalidad de ramos mixtos, pero fue de maslenta entrada en producción que el Ypsi-lon, debido a una menor densidad deplantación (Delgado, 2000), resultadoscoincidentes con los obtenidos en el pre-sente ensayo. En otra experiencia similar,Blanc y Arregui (1999), evaluaron con cua-tro variedades de pavía los sistemas Vaso(marco de plantación a 5x3,5m), Doble Y(marco 5x3m), Ypsilon (marco 5x5m) y Pal-meta (marco 5x5m), resultando el Doble Yél de más rápida entrada en producción yla Palmeta el sistema más lento en entraren producción, debido probablemente aque la distancia entre árboles e hileras fuedistinta. En el presente ensayo el compor-tamiento de los sistemas ha sido diferente,dado que la Palmeta ha sido más precozque el Doble Y.
Peso medio del fruto
En la tabla 3 se expone el peso medio delfruto correspondiente a los diferentes sis-temas. En el primer año se encontraronmínimas diferencias estadísticas, siendo elsistema Ypsilon el que presentó los frutosde menor peso debido probablemente aque fue de los sistemas de mayor densi-dad. En el 2000 hubo ligeras diferencias,siendo el Ypsilon y el Tatura los que mos-traron los frutos de menor peso. En el2001 y 2002, no se encontró diferenciaestadística entre sistemas. En el año 2003en todos los sistemas, con diferencias esta-dísticas entre ellos, disminuyó el pesomedio del fruto con respecto al obtenidootros años. Los sistemas que alcanzaron elmayor peso fueron la Palmeta, el Vaso y elEje; mientras que el Tatura fue el de me-nor peso.
Marini y Sowers (2000) observaron, en lavariedad ‘Norman’ conducida en Vaso y
Tabla 2. Producciones anuales y acumuladas (kg/ha) en el período 1996-2003 de diferentes sistemasde formación en melocotonero
Table 2. Annual and cumulative yields (kg/ha) over 1996-2003 period, of different peach trainingsystems
Sistema 1996 1997 Acumulada 2000 2001 2002 2003 Acumulada2º año 3º año 1996-1998 6º año 7º año 8º año 9º año 1996-2003
Ypsilon 3.851 b 21.955 a 61.908 ab 42.243 a 41.080 a 45.703ab 60.314 a 251.248 aEje 6.890 a 22.793 a 67.985 a 43.127 a 41.454 a 42.198 b 53.860 b 248.624 aVaso 1.766 bc 20.147 a 56.379 bc 40.907 a 35.103 ab 41.929 b 50.359 b 224.677 cDobleY 1.585 c 16.128 b 45.000 e 39.035 a 29.725 b 32.555 c 45.876 b 192.191 dPalmeta 3.686 bc 16.173 b 53.331 cd 40.428 a 42.006 a 39.139 b 48.222 b 223.126 cTatura 2.104 bc 14.650 b 47.501 de 43.580 a 37.525 a 50.485 a 58.443 a 237.534 b
Valores con la misma letra en la misma columna no difieren significativamente, según el Test de Rango Múltiple deDuncan (P≤0,05).

densidades de 740 y 370 árboles por hectá-rea, que el peso medio del fruto disminuyóal aumentar la densidad, probablementedebido a una menor disponibilidad de luz.Sassella (1997) evaluó los sistemas Ypsilon yFuseto (densidad 556 árboles/ha), Palmetay Vaso (333 árboles/ha) en melocotonerocon las variedades ‘Redhaven’ y ‘SpringCrest’, utilizando el patrón GF-305, encon-trando una interacción significativa siste-ma de formación x variedad, dado que con‘Redhaven’ el Ypsilon presentó el mayorpeso medio, mientras con ‘Spring Crest’fue la Palmeta el sistema con un peso defruto superior.
Los datos obtenidos en el presente ensayomuestran que la densidad apenas afectó alpeso del fruto, dado que la disponibilidadde luz no se vio limitada en ningún caso, altratarse de densidades óptimas para cadasistema. A lo largo del período de estudio,el Ypsilon fue el sistema con frutos demenor peso, en cambio la Palmeta fue delos sistemas que produjeron frutos demayor peso. A pesar de ello dichas diferen-cias no fueron siempre significativas a lolargo de los años.
Calibre
El calibre medio ponderado de las produc-ciones de los diferentes sistemas en el perío-do 1997-2002, así como los porcentajes deproducción comercial obtenidos, se expo-nen en la tabla 4. Con respecto al calibre<70mm (no comercial), se dieron diferenciassignificativas entre sistemas, correspondien-do los mayores porcentajes a los sistemascon mayor densidad, el Ypsilon, el Eje y elTatura. No hubo diferencias en los interva-los superiores, a excepción del Ypsilon queobtuvo el porcentaje más bajo en las cate-gorías mas altas, 22,6% en 80-85 y 11,0% en>85mm. En lo que respecta al porcentaje deproducción comercializable, los sistemasque mostraron los mayores porcentajes fue-ron el Doble Y, la Palmeta y el Vaso; mien-tras que el Ypsilon, el Eje y el Tatura obtu-vieron los valores más bajos.
Caruso et al. (1998) utilizando los sistemasYpsilon y Eje con ‘Spring Lady’, densidad de1111 árboles/ha y altura de 4,5 m, obtuvie-ron con el Ypsilon los mejores resultados,especialmente en la parte media del árbol,situándose los calibres no comerciales en laparte baja. Corelli y Sansavini (1991) eva-luando los sistemas de Vaso, Palmeta, Eje e
R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26 19
Tabla 3. Peso medio del fruto (g) correspondiente a diferentes sistemas de formación demelocotonero. Valores medios anuales para el período 1998-2003
Table 3. Average fruit weight (g) of different peach trainning systems. Annual mean values obtainedover 1998-2003 period
Sistema 1998 2000 2001 2002 20034º año 6º año 7º año 8º año 9º año
Ypsilon 209,3 b 182,7 b 223,7 263,2 208,3 abEje 222,6 a 198,3 ab 224,5 269,7 213,8 aVaso 232,0 a 194,0 ab 232,6 277,6 216,8 aDoble Y 228,4 a 199,3 a 229,0 285,7 210,4 abPalmeta 232,2 a 190,6 ab 229,4 283,7 222,3 aTatura 228,8 a 182,7 b 227,6 279,6 197,4 b
Valores con la misma letra en la columna no difieren significativamente, según el Test de Rango Múltiple de Dun-can (P≤ 0,05).

Ypsilon, con las variedades ‘Flavorcrest’ y‘Rubired’, con una densidad de 729 árbo-les/ha; encontraron que el Ypsilon propor-cionó el mayor porcentaje de fruta comer-
cial. En el presente ensayo fue el Ypsilon elque presentó el porcentaje más bajo defruta comercial, debido probablemente a lamayor densidad de plantación.
20 R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26
Marini y Sowers (2000) observaron, con lavariedad ‘Norman’, que a medida que seincrementaba la densidad de plantacióndisminuía el porcentaje de fruta comerciali-zable. Marini et al. (1995), con la variedad‘Sweet Sue’, obtuvieron mayores calibrescon el sistema Vaso que con el Eje. En elpresente ensayo el Vaso proporcionó losmejores calibres comerciales, constatándoseque además de la densidad, tanto la formadel árbol como la carga son factores muyimportantes a considerar sobre el tamañodel fruto.
Parámetros de calidad de los frutos
Los valores medios de firmeza, contenidosde azucares solubles y acidez titulable,obtenidos a lo largo del período 1997-2002,no muestra diferencias significativas atri-buibles al efecto del sistema de formación(datos no expuestos). Estos resultados soncoincidentes a los expuestos por Hilaire y
Giauque (2002) que encontraron para elcultivar ‘O’Henry’ de 11-13º Brix y una aci-dez de 9-11 g/l, considerando a esta varie-dad de sabor equilibrado. De acuerdo a losresultados obtenidos en el ensayo, única-mente en el año 2000, los sistemas Tatura,Ypsilon y Doble Y superaron este valor.
Eficiencia productiva o índice deproductividad (I.P.)
Todos los años, a excepción de 1997, sedieron diferencias significativas entre sis-temas en lo referido a eficiencia producti-va. En general, fueron el Tatura, la Palme-ta y el Vaso, los sistemas que alcanzaronlos valores mas altos de eficiencia; siendoinferior para el Doble Y. En cuanto a efi-ciencia productiva acumulada, el sistemaTatura fue el que presentó el valor másalto; mientras el Doble Y obtuvo el valormás bajo (tabla 5).
Tabla 4. Porcentajes ponderados de la producción correspondientes a los diferentes intervalos decalibre (en mm) de los diferentes sistemas de formación y porcentajes no comercializables. Valores
medios correspondientes al período 1997-2002Table 4. Averages of yield by fruit size intervals (mm), and averages of yield corresponding to different
peach training systems. Mean values obtained over 1997-2002 period
Sistema <70 mm 70 - 75 75 - 80 80 - 85 >85mm % comercial
Ypsilon 12,7 a 22,3 31,3 22,6 b 11,0 b 87,3Eje 9,7 ab 20,3 29,6 25,7 ab 14,6 ab 90,3Vaso 7,0 bc 17,3 27,7 30,6 a 17,3 ab 93,0Doble Y 5,3 c 19,6 32,3 28,0 ab 14,6 ab 94,6Palmeta 6,3 c 17,6 29,7 25,7 ab 20,7 a 93,7Tatura 9,6 ab 18,3 27,6 24,6 ab 19,6 a 90,3
Valores con la misma letra en la columna no difieren significativamente, según el Test de Rango Múltiple de Dun-can (P≤0,05).

Los resultados obtenidos coinciden con losexpuestos por Hutton et al. (1987) que obser-varon, con la variedad ‘Yanco Queen’, unamayor eficiencia productiva del Tatura conrespecto a la Palmeta. Espada et al. (2000)evaluaron los sistemas de Vaso y Eje en lasvariedades ‘Jesca’, ‘Calante’, ‘CA-8’ y ‘Evaisa’sobre GF-677 a una densidad de 277 y 555árboles/ha, obteniendo una eficiencia pro-ductiva acumulada al octavo año de 1,32kg/cm2 para el Eje y de 1,41 para el Vaso, muyinferiores a los obtenidos en el presenteensayo.
Radiación fotosintéticamente activa (PAR)interceptada
Respecto a la radiación interceptada, se hanobservado diferencias significativas entresistemas, tal como se observa en la tabla 6,donde se han representado los porcentajesde luz interceptada (PAR) desde mayo aagosto del 2003.
El sistema de formación que interceptó elmayor porcentaje de radiación a las 10:00am hora solar del mes de mayo, fue el Tatu-ra, siendo el Ypsilon el que interceptómenos luz. Para los diferentes meses (mayo
a agosto) y de forma general se observa quea las 6:00 am hora solar, los sistemas quelograron interceptar/captar más radiación,fueron el Tatura, seguido del Eje y del Ypsi-lon; mientras que los de menor captaciónfueron el Vaso, el Doble Y y la Palmeta. Alas 8:00 am hora solar de los meses de Junio,Julio y Agosto, los sistemas que presentaronel porcentaje más alto de radiación inter-ceptada fueron el Tatura y el Ypsilon; encambio la Palmeta presentó menos capta-ción. A las 10:00 am hora solar, el sistemaque también mostró más intercepción fue elTatura, y el de menor el Eje y la Palmeta,obteniendo con el resto valores intermedios(tabla 6).
El promedio de las 10 lecturas realizadasdesde mayo hasta agosto de 2003, pone demanifiesto que los sistemas que intercepta-ron mayor radiación fueron el Tatura con82,8 %, el Ypsilon con 71,3 % y el Vaso con70,8 %; mientras que los de menor capta-ción de luz fueron el Doble Y, el Eje y la Pal-meta (tabla 6).
Los valores porcentuales de radiación inter-ceptada por los diferentes sistemas así comode la radiación directa incidente en treshoras (hora solar) de la mañana del día 6 de
R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26 21
Tabla 5. Índice de productividad anual y acumulado (kg/cm2) de diferentes sistemas de formación enmelocotonero a lo largo del periodo 1997-2003
Table 5. Annual and cumulative yield efficiency (kg/cm2) corresponding to different peach traininngsystems over 1997-2003 period
Sistema 1997 1998 2000 2001 2002 2003 I.P.3er año 4º año 6º año 7º año 8º año 9º año acumulado
Ypsilon 0,40 0,50 c 0,44 b 0,38 ab 0,38 bc 0,36 b 1,95 bEje 0,43 0,55 bc 0,43 b 0,37 ab 0,35 bc 0,35 b 1,91 bVaso 0,49 0,61 abc 0,52 b 0,40 ab 0,42 b 0,41 b 2,10 abDobleY 0,44 0,58 abc 0,51 b 0,35 b 0,33 c 0,33 b 1,78 bPalmeta 0,44 0,64 ab 0,51 b 0,48 a 0,40 bc 0,39 b 2,10 abTatura 0,47 0,69 a 0,64 a 0,47 a 0,54 a 0,51 a 2,32 a
Valores con la misma letra en la misma columna no difieren significativamente, según el Test de Rango Múltiple deDuncan (P≤0,05).

22 R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26
Tabla 6. Porcentajes de luz interceptada por los diferentes sistemas de formación, en el períodomayo-agosto de 2003 y a diferentes horas del día
Table 6. The effect of different peach training systems on light intercepted, over May-August 2003period and at different day times
Sistema Mayo Junio Julio Agosto Promedio10:00 6:00 8:00 10:00 6:00 8:00 10:00 6:00 8:00 10:00
Ypsilon 49,1 77,3b 65,3b 58,4c 83,3ab 77,0b 71,6b 87,8ab 74,7b 68,4b 71,3 bEje 58,8 78,2b 61,6b 65,1b 83,7ab 71,5bc 66,8c 86,3bc 62,5c 60,9c 69,5 bVaso 56,0 73,5b 60,5bc 68,7b 76,6c 70,7c 69,2bc 86,9bc 76,1b 69,9b 70,8 bDobleY 57,9 76,2b 63,7b 68,5b 77,6bc 72,1bc 71,8b 84,5cd 70,9b 70,6b 70,7 bPalmeta 52,3 74,2b 56,0c 60,2c 80,8bc 64,1d 59,6d 82,4d 57,9c 59,6c 64,2 cTatura 63,1 84,5a 83,4a 75,3a 89,2a 85,9a 83,6a 90,6a 85,7a 81,9a 82,8 a
Valores con la misma letra en la columna no difieren significativamente, según el Test de Rango Múltiple de Dun-can (P≤0,05).
Figura 3. Efecto de diferentes sistemas de formación en los valores de luz interceptada (µmol m2 s-1)en tres horas del día, 6 agosto de 2003. Las barras verticales indican la LSD (P≤0,05) para cada fecha y
para el promedio.Figure 3. Values of light intercepted (µmol m2 s-1) affected by different peach training systems on threetimes of 6th August 2003. The vertical bars indicate the LSD (P≤0,05) for each date and for the average.
Hora solar
µµ mo
l m2
s-1
agosto de 2003 (día soleado), se exponenen la figura 3, (ver figura 1 metodología).Se observa que fue también el Tatura el masefectivo, seguido por el Ypsilon; la menorinterceptación correspondió a la Palmeta yal Eje. Dichos resultados son similares a losexpuestos en la tabla 6 y muestran diferen-cias importantes entre sistemas, debidas a
diferencias en la distribución espacial (ocu-pación del espacio) de la copa de los mis-mos. Ello se ha traducido en capacidades deinterceptación de radiación y consecuente-mente en áreas de sombreo y eficienciasproductivas muy diferentes, mayores en alcaso de la Tatura, tal como se ha expuestoen la tabla 5.

Grossman y DeJong 1998, reportaron quela cantidad de luz interceptada en meloco-tonero se incrementa a medida que avanzala temporada, debido al crecimiento envolumen de la copa del árbol. Papasergio(1996) y Reighard y Okie (2000) mencionanque el sistema de Tatura, en el sur de Italia,y las formas en V o Y, en el sureste de losEstados Unidos, fueron las que más radia-ción interceptaron. Dichos resultados coin-ciden con los expuestos en el presenteensayo, ya que efectivamente tanto elTatura como el Ypsilon, fueron las formascon mayor interceptación, lo que estuvorelacionado un mayor cuajado de frutos(datos no mostrados) y una mejor calidadde los ramos mixtos (datos no expuestos).
Corelli et al. (2000) en melocotoneros de 5años de la variedad ‘Stark Red Gold’ sobrepatrón franco, estudiaron tres sistemas deformación: Palmeta (densidad 727 árboles/ha), Vaso retardado (densidad 545 árboles/ha) y Ypsilon (1274 árboles/ha). El sistemaque captó más luz fue el Ypsilon (68-85 %),seguido por el Vaso retardado (48-58 %) ypor la Palmeta (45- 55 %). En el presenteensayo, los sistemas se han comportado deforma similar, coincidiendo con el porcenta-je captado por el Ypsilon; en cambio la Pal-meta captó más luz de la que mencionandichos investigadores, debido probable-mente a la edad de los árboles o bien a suforma.
Incidencia de enfermedades
El sistema de formación influyó en la inci-dencia de las principales enfermedades,observándose diferencias significativas en-tre sistemas.
Daños por Pseudomonas syringae
El número de ramos mixtos y hojas daña-dos por P. syringae en abril de 2003, para
los diferentes sistemas, se expone en latabla 7. En cuanto al número de ramosdañados por árbol, los sistemas que resul-taron más afectados fueron el Tatura, elVaso y el Ypsilón, en cambio los menosdañados fueron el Eje, la Palmeta y elDoble Y. Los daños más fuertes se concen-traron en los ramos de la parte alta de losárboles, siendo el Tatura el sistema másafectado. Sin embargo a principios demayo los daños desaparecieron, debido aldesarrollo de los brotes y a las condicionesclimáticas menos favorables; ya que en eseperíodo aumentaron las temperaturas ydisminuyó la humedad.
Hilaire y Giauque (2002) señalan en el valledel Rhône (Francia) la variedad de meloco-tón ‘O’Henry’® como sensible a la bacteriaXanthomonas arborícola pv. pruni. En elensayo no se detectó la presencia de estabacteria, pero si de Pseudomonas syringae;aunque con daños y síntomas no coinci-dentes con los mencionados por dichosinvestigadores.
Aulagnier (1996) menciona que las varieda-des ‘Maycrest’, ‘Springlady’ y ‘Flavorgold’son sensibles a la bacteriosis originada porPseudomonas syringae en la región frutíco-la Rhône-Alpes (Francia). En las variedadestempranas el daño fue mayor, debido a lasbajas temperaturas y a la mayor humedadambiental, que favorece el desarrollo deeste patógeno. En ‘O’Henry’® la sensibili-dad fue menor, debido a que brota mástarde y las condiciones ambientales sola-mente favorecieron el ataque durante laprimera semana de abril. En base a ello sededuce que la afección depende principal-mente de la época de floración-inicio devegetación de la variedad en cuestión y delas condiciones ambientales del año, espe-cialmente humedad y temperatura.
R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26 23

24 R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26
Tabla 7. Número de ramos mixtos y hojas dañados por Pseudomonas syringae en la parte baja y altasegún los diferentes sistemas de formación en melocotonero año 2003
Table 7. Number of shoots and leafs damaged by Pseudomonas syringae on the top and medium partof the tree affected by different peach training systems in 2003
Sistema Número de ramos Número de ramos afectados en:
Total Parte baja Parte alta 1 hoja 2 hojas >3 hojas
Ypsilón 11 b 4 a 7 b 6 bc 4 b 1 abEje 1 c 0 b 1 c 1 c 0 c 0 cVaso 12 b 4 a 8 b 8 ab 4 ab 1 abDoble Y 4 c 4 a 8 b 2 c 1 c 0 cPalmeta 3 c 0 b 3 c 2 c 1 c 0 cTatura 20 a 3 a 17 a 11 a 6 a 3 a
Los valores seguidos de la misma letra en la columna no difieren significativamente, según el Test de Rango Múlti-ple de Duncan (P≤0,05).
Tabla 8. Número de ramos mixtos y hojas afectados por Taphrina deformans Burk., en diferentessistemas de formación en melocotonero, año 2003
Table 8. Number of shoots and leaves damaged by Taphrina deformans Burk., affected by differentpeach training systems in 2003
Sistema Número de ramos Número de ramos afectados en:
Total Parte baja Parte alta 1 hoja 2 hojas 3 hojas >4 hojas
Ypsilón 19 b 4 bc 15 b 12 bc 5 b 1 b 1 bEje 3 b 1 c 2 b 2 c 1 b 0 b 0 bVaso 33 b 9 bc 24 b 24 b 7 b 2 b 0 bDoble Y 9 b 19 b 22 b 13 bc 4 b 1 b 1 bPalmeta 6 b 2 c 4 b 5 c 1 b 0 b 0 bTatura 149 a 41 a 108 a 46 a 28 a 28 a 47 a
Los valores seguidos de la misma letra en la columna no difieren significativamente, según el Test de Rango Múlti-ple de Duncan (P≤ 0,05).
Daños por Taphrina deformans (Burk)
La afección por esta enfermedad mostródiferencias estadísticas entre sistemas. Enla tabla 8 se expone el número de ramosmixtos y hojas dañados por Taphrinadeformans (Burk ) en la primera semanade mayo del 2003, observándose una ma-yor afección en la parte alta de los árboles.
Por el número de ramos afectados, el siste-ma Tatura resultó el más dañado por estehongo, tanto en la parte alta como en laparte baja, siendo mayor el porcentaje delos síntomas para ramos que presentaronuna hoja dañada. La incidencia de estehongo se vio favorecida por las precipita-ciones que se presentaron en el mes deabril.

Aulagnier (1996) indica que las variedadesde melocotón ‘Maycrest’, ‘Springlady’ y ‘Fla-vorgold’ en Vaso fueron muy susceptibles aTaphrina deformans (Burk), detectándosetambién diferencias entre sistemas. Losresultados obtenidos en el ensayo muestranque el Vaso presentó daños por este pató-geno aunque menos intensos que los men-cionados por este autor, al tratarse de dife-rente variedad y de diferentes condicionesclimáticas.
Conclusiones
A partir de los resultados expuestos, pue-den extraerse las siguientes conclusiones.
La densidad de plantación influyó significa-tivamente en el vigor del árbol, estandoéste relacionado con la densidad de planta-ción. Independientemente del sistema deformación a mayor densidad menor fue lasección transversal de tronco y viceversa.
La densidad de plantación determinó larapidez de entrada en producción, obser-vándose una relación directa entre la densi-dad de plantación y la producción obtenidapor hectárea.
Los sistemas de mayor producción acumula-da fueron el Eje y el Ypsilon, al disponer demayor densidad de plantación. Sin embar-go, el Tatura presentó mayor producciónque la Palmeta a pesar de la menor densi-dad, debido a la mayor interceptación deradiación.
El sistema que presentó mayor eficienciaproductiva acumulada fue el Tatura, segui-do por el Vaso y la Palmeta; mientras los sis-temas de mayor densidad el Ypsilon, el Eje yel Doble Y obtuvieron valores inferiores.
Los sistemas que interceptaron los mayoresporcentajes de radiación fueron el Tatura, elYpsilon y el Vaso, correspondiendo el menor
a la Palmeta. Sin embargo este sistema pre-sentó una buena eficiencia productiva ybuen calibre.
En cuanto a la calidad de los frutos, los siste-mas que mostraron los mayores porcentajesde calibres comerciales fueron el Doble Y, laPalmeta y el Vaso; mientras que el Ypsilon,el Eje y el Tatura obtuvieron los valores másbajos.
El sistema de formación no afectó a losparámetros de calidad de los frutos: sólidossolubles, firmeza y acidez.
El sistema que presentó mayor incidencia deenfermedades fue el Tatura y los de menordaño fueron el Eje y la Palmeta. La forma yla densidad de los árboles según cada siste-ma, afectaron a la aplicación y eficiencia delos productos fitosanitarios.
Bibliografía
Aulagnier M, 1996. Pêches label rouge de laDrome. Une analyse régionale des potential-ités qualitatives. L‘arboriculture Fruitière, 497:34-41.
Blanc P, Arregui M, 1999. Pêchê de conserve. Lapavie revue au gout du jour. L‘arboricultureFruitière, 533: 33-36.
Carbó J, Iglesias I, 2002. Melocotonero. Las var-iedades de más interés. Institut de Recerca iTecnología Agroalimentàries (IRTA), Barce-lona (España), 287 pp.
Caruso T, Di Vaio C, Inglese P, Pace LS, 1998. Cropload and fruit quality distribution withincanopy of ‘Spring Lady’ peach trees trained to‘Cental Leader’ and ‘Y shape’. Acta Horticul-turae, 465: 621-628.
Corelli L, Sansavini S, 1991. Forme di allevamen-to, efficienza degli impianti e qualità dellepesche. Rivista di Frutticoltura 6: 13-24.
Corelli L, Ravaglia G, Magnanini E, 2000. Lightconversion efficiency as affected by training
R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26 25

system in peach. Dipartimento di ColtureArboree- Universita di Bologna (Italia).
Delgado M, 1992. Pêcher taille et éclaircissage,un équilibre á trouver. L‘arboricultureFruitière, 446: 32-36.
Delgado M, 2000. Technique pechers. Les formessemi-piétonnes ont la cote. L‘arboricultureFruitière, 534: 32-35.
Espada CJL, Romero SJ, Segura GJ, 2000.Adaptación de clones seleccionados de melo-cotonero ‘Calanda Tardío’ al sistema de con-ducción en Eje Central. Revista ITEA 21: 159-162.
Giauque P, 2003. Conduite du verger de pêcher.Recherche de la performance. Centre Tech-nique Interprofessionnel des Fruits etLégumes (Ctifl), 90-91.
Grossman YL, DeJong, T.M., 1998.Training andpruning system effects on vegetative growthPotential, light interception and croppingefficiency in peach trees. Journal AmericanSociety of Horticultural Science, 123 (6): 1058-1064.
Hilaire C, Giauque P, 2002. Le pêcher. CentreTechnique Interprofessionnel des Fruits etLégumes (Ctifl), 205-207.
Hutton RJ, McFadyen M, Warwick JL, 1987. Rela-tive productivity and yield efficiency of Can-ning peach trees in three intensive growingsystems. HortScience, 22(4): 552-560.
Marini RP, Sowers D, Marini MC, 1995. Tree Formand Heading at Planting affect peach tree
yield and crop value. HortScience, 30(6): 1196-1201.
Marini RP, Sowers DS, 2000. Peach fruit weight isinfluenced by crop density and fruiting shootlength but not position on the shoot. JournalAmerican Society of Horticultural Science, 119(2): 180-184.
Papasergio R, 1996. La peschicoltura nella pianadi Sibari. Rivista di Frutticoltura, 2: 51-53.
Reighard GL, Okie WR, 2000. Current peach pro-duction practices including nex Training sys-tems in the Southeastern, EEUU.pp.112-118.En: VI Simposium Internacional sobre frutalesde clima templado, Cuauhtémoc, Chihuahua(México).
Royo DJB, Martínez LT, 1992. Sistemas de forma-ción en melocotonero. Fruticultura Profesion-al, 46: 22-31.
SAS, 2002. SAS/STAT Guide for personal comput-ers Versión 8.1. SAS Institute, Inc.:Cary. N.C.
Sassella A, 1997. Influence de I´association var-iéte; porte-greffe et forme de conduite Sur lecomportement du pecher au Tessin. RevueSuisse Vitic. Arboric., 29(4): 197-202.
Wünsche J, Lakso A, Robinson L, 1996. The basesof productivity in apple production systems:The role of light interception by differentshoot types. Journal American Society of Hor-ticultural Science, 121(5): 886-893.
(Aceptado para publicación el ?? de ?? de 2006).
26 R. Núñez et al. ITEA (2006), Vol. 102 (1), 13-26
OJO falta fechade aceptación

A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40 27
El control fotobiológico de malas hierbas: revisión de unatécnica controvertida
A. Cirujeda*, A. Taberner**
* Unidad de Sanidad Vegetal. CITA. Avda. Montañana 930 50080 Zaragoza, Españ[email protected]** Servei de Sanitat Vegetal. Unitat de Malherbologia Avda. Alcalde Rovira Roure 191, 25198 Lleida,España.
ResumenEn el presente trabajo se realiza una revisión bibliográfica sobre una técnica preventiva de reducciónde la nascencia de las malas hierbas. Esta técnica ha producido resultados controvertidos debido a laabundancia de datos contradictorios en las investigaciones. El control fotobiológico consiste en sem-brar y realizar otras labores en oscuridad, o bien tapando los aperos con lonas opacas, evitando quelas semillas de las malas hierbas reciban el estímulo de luz necesario para germinar. En el presente tra-bajo se describen primero algunos aspectos de la teoría de esta técnica. Para ello, se ha revisado lafisiología de la dependencia de luz para germinar en las semillas de malas hierbas, se describe cómoesta necesidad varía según las circunstancias y se revisan las interacciones que existen entre diferentesestímulos ambientales. Respecto a la aplicación de dicha técnica, se revisan, en segundo lugar, traba-jos realizados en campo (sobre todo en el norte de Europa) y se detallan las especies de las que se haobtenido una reducción de la germinación. Finalmente, se llega a unas conclusiones sobre la posibleviabilidad de este sistema preventivo, del que se recalca su irregular eficacia difícilmente controlable,ya que está relacionada con la variabilidad de la sensibilidad entre las especies, con la humedad delsuelo y con el régimen de temperaturas o aspectos de la propia fisiología de las semillas.
Palabras clave: luz, germinación, prevención
SummaryThe photocontrol of weeds: a review on a controversial techniqueThis work presents a bibliographical review of a preventive technique to avoid weed emergence. Thistechnique has produced contradictory results in different research work and is thus a controversialmethod. The photocontrol consists in sowing and doing other labour at night or covering the toolswith a light-proof cover, avoiding that weed seeds receive the necessary light stimulus for germina-tion. Firstly some theoretical aspects are described about the physiology of light dependence of weedseed germination, which is not a constant need, and the interactions between different environmen-tal stimuli concerning light dependence. The second part is a review on the field work conductedmainly in Northern Europe and describes the weed species, whose emergence was reduced. Finally,some conclusions concerning the possible viability of this preventive technique are expressed. Themain problem seems to be the irregular efficacy, which is difficult to control as it is related to the dif-ferent light sensitivity between weed species influenced by soil moisture, temperature regime andaspects of the own seed physiology.
Key words: light, germination, prevention

Introducción
La sensibilidad por el medio ambiente y laaspiración a emplear técnicas agronómicasmás respetuosas con el mismo han idoaumentando en las últimas décadas. En pro-tección de cultivos se aboga por el controlintegrado, es decir, el uso planificado demétodos físicos, químicos y biológicos paracontrolar especies o poblaciones determina-das de malas hierbas (Powles y Matthews,1996; Hatcher y Melander, 2003) siendo eluso de productos químicos sólo una medidamás. También en el caso de la producciónecológica es necesario combinar diferentestécnicas preventivas con otras de accióndirecta sobre las hierbas, siguiendo la teoríade emplear “muchos pequeños martillos”,descrita por Liebmann y Gallandt (1997)que propone combinar numerosos métodosde baja eficacia. La técnica que se expone acontinuación podría servir como un compo-nente de un programa integrado de controlde malas hierbas (Milberg et al., 1996),siempre con la intención de ser utilizadocomo un método preventivo en combina-ción con métodos de control culturales,mecánicos o químicos (Gerhards et al., 1997;Juroszek et al., 2002).
Los efectos de la luz en la germinación delas semillas son conocidos desde hacemucho. Sin embargo, su utilización confines agronómicos ha comenzado a serinvestigada desde hace muy poco, siendo elprimer trabajo publicado sobre el fotocon-trol el realizado por el investigador alemánHartmann en 1990.
El fundamento del control fotobiológico seconcreta en preparar el lecho de siembra yen sembrar en oscuridad, para que aquellassemillas de especies no deseadas que sontrasladadas por los aperos a la superficie delsuelo, pero que vuelven a quedar enterra-das posteriormente, no reciban los instantessuficientes de luz que desencadenan la ger-
minación y no germinen. No obstante, en lasiembra nocturna (o diurna, tapando losaperos con lonas que no permitan el pasode la luz) no cabe esperar una inhibicióncompleta de la germinación, ya que por unaparte habrá semillas de especies insensiblesa la luz que germinarán, y por otra siemprequedarán semillas en capas superiores delsuelo que recibirán luz posteriormente yque podrán germinar (Hartmann y Nezadal,1990). El objetivo es reducir parcialmente lanascencia de las especies fotosensibles,teniendo en cuenta que probablementeserá necesario tomar las medidas de controlpertinentes en postemergencia.
Desde los años 90 se vienen realizando ensa-yos en campo para determinar el potencialdel control fotobiológico de malas hierbas.Estos ensayos básicamente se han realizadoen países del Norte de Europa. El primer tra-bajo fue publicado por Hartmann y Nezadal(1990), exponiendo sus experiencias de 7años en Alemania. Siguieron publicacionesde otros equipos en el mismo país (Küh-bauch et al., 1992; Klümper et al., 1996;Juroszek et al., 2002); Spiess (1995), Nie-mann y Biller (1996) y de otros países: Die-rauer (1992) en Suiza, Ascard (1993) en Sue-cia, Adamiak y Adamiak (2003) en Polonia yScopel et al. (1994) en EEUU. Se ampliaronlos estudios en campo realizando el últimolaboreo y la siembra de día, pero cubriendolos aperos con lonas opacas. Ascard (1993 y1994), Scopel et al. (1994), Gerhards et al.(1997) y Welsh (1999) observaron reduccio-nes en la densidad de hierbas germinadasen este mismo tipo de ensayos. Estos últi-mos autores también ensayaron si era posi-ble reforzar el efecto, alumbrando el espa-cio de trabajo de los aperos de laboreo conluz roja oscura. Estos trabajos han sido com-plementados por otros que estudian lafotosensibilidad de las diferentes especiesen ensayos de laboratorio: Jensen (1995),Milberg et al. (1996), Buhler (1997), Hart-
28 A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40

mann et al. (1998), Hartmann et al. (2003).La mayoría de trabajos que se encuentranen la presente década son de revisión, resu-miendo aspectos determinados de losnumerosos ensayos realizados en la décadaanterior (p.ej. Juroszek, 2002 y Juroszek yGerhards, 2004).
Aparte de la esperada reducción de nas-cencia de las hierbas no deseadas, otroobjetivo en el manejo de la siembra o labo-res en oscuridad es el de reducir su capaci-dad competitiva respecto al cultivo (Ras-mussen, 2000 y Rasmussen, 2002). Éstepodría ser también un objetivo del controlfotobiológico, ya que, entre otros, Jensen(1992), Kühbauch et al. (1992) y Ascard(1994) encontraron un retraso de la germi-nación de las malas hierbas en parcelassembradas de noche, lo cual puede dar unaventaja competitiva a cultivos de rápidodesarrollo.
A diferencia de las malas hierbas si el cultivose siembra en oscuridad, germinará. El con-trol fotobiológico se basa en la distintafotosensibilidad que tienen las semillas dealgunas especies arvenses respecto a la quetienen las semillas de plantas cultivadas.Esto se debe a que la mayoría de cultivoshan sido seleccionados y mejorados paratener una latencia menos evidente y lamayoría de ellos germinan independiente-mente de la luz (Gerhards et al., 1997).
Otra posible aplicación del conocimientode la necesidad de luz para germinar endeterminadas especies podría ser la de esti-mular la nascencia de malas hierbas en laslabores previas, desarrollándolas a plenaluz. Sólo la última labor y la siembra se rea-lizarían de noche (Hartmann y Nezadal,1990). Otros autores proponen estimular laemergencia añadiendo luz adicional y con-trolando las plantas germinadas (Scopel etal., 1994).
La luz y la germinación
A los citados ensayos de campo cabe añadirtoda una serie de publicaciones comple-mentarias a los ensayos de campo que des-criben el efecto de la luz sobre la germina-ción de determinadas especies de plantas,siendo Lactuca sativa y Arabidopsis thalianaL. Heynh. las especies más estudiadas. Des-tacan, entre otros numerosos grupos, lasaportaciones realizadas por el mismo equi-po pionero alemán de la Universidad deErlangen-Nürnberg (Hartmann, Mollwo,Tebbe y otros), las realizadas por el equipode la Universidad de Buenos Aires (Argenti-na) compuesto por Casal, Sánchez, Ballaré,Botto y otros; el equipo de la Universidadde Wageningen (Holanda) compuesto porDerkx, Karssen, Vleeshouwers, Bowmeestery otros; y el equipo del Laboratorio de Foto-percepción y Transducción de Señales enWako City (Japón) compuesto por Shinomu-ra, Nagatani y otros.
Muchos autores describen la fotodependen-cia en numerosas especies denominadasfotoblásticas (por ejemplo Jensen, 1992;Ascard, 1994). No obstante, pronto se des-cubrió que esta necesidad de luz para ger-minar varía entre especies. Deonder et al.(1978) observaron como, de un total de 250especies analizadas, el 40% mostraba unadependencia absoluta de la luz para germi-nar, un 24% tenía una dependencia parcialy el 36% reaccionaba indiferente frente a lapresencia de la luz.
Los aspectos que pueden causar estas dife-rencias entre especies pueden ser múltiples.Por un lado, Ascard (1994) y Taiz y Zeiger(1994) encontraron una tendencia a mayorsensibilidad a la luz en semillas pequeñasfrente a mayores, confirmada también porMilberg et al., 2000, a pesar de que algunaespecie de semilla pequeña, como Matrica-ria perforata, no siguió el mismo patrón(Ascard, 1994) y de que no se encontraron
A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40 29

diferencias en otros trabajos de los mismosautores analizando hasta 44 especies desemillas distintas (Milberg et al., 1996).
La complejidad de la dependencia de la luzqueda reflejada también en el hecho quepuede variar entre poblaciones de unamisma especie (Milberg et al., 1996) e inclu-so entre distintas semillas de una mismapoblación, como lo describieron para Matri-caria inodora L. Hartmann et al. (1997).También hay unanimidad en las publicacio-nes acerca de la variación estacional de lasensibilidad a la luz. Así, Baskin et al. (2002)describen como Papaver rhoeas L. sólo res-ponde a la presencia de luz si las semillaspreviamente embebidas han pasado por unperíodo de altas temperaturas que hayaroto la latencia fisiológica.
El sistema del fitocromo en las semillas
Se considera que la mayoría de respuestas ala luz en semillas se desarrollan a través delsistema del fitocromo (Ballaré et al., 1992).Los fitocromos son una familia de proteínasnecesarias para el desarrollo vegetal queson sensibles a la luz roja y al rojo lejano(Furuya, 1989, citado por Reed et al., 1994).La existencia de al menos cinco genes dife-rentes codificando para el fitocromo enArabidopsis thaliana, especie muy utilizadapara estudios en este tema, lleva a suponerque los miembros de la familia juegan dife-rentes papeles en la fotoregulación de lagerminación de semillas (Shinomura, 1997).Estos cinco tipos de fitocromo son denomi-nados A, B, C, D y E. No obstante, no seconocen los detalles de comportamiento detodos ellos y el fitocromo A es el miembromejor caracterizado de entre la familia defitocromos en angiospermas (Casal et al.,1997).
El fitocromo es un fotoreceptor con dos for-mas foto-interconvertibles: la forma con un
máximo de absorción en el rojo (Pr) y laforma con el máximo en el rojo lejano (Pfr):
� Luz roja (654 nm)Pr��Pfr� Luz en el rojo lejano (730 nm)
La irradiación con luz de día convierte Pr enPfr (Vleeshouers et al., 1995). La fotoconver-sión de Pr hacia Pfr promueve diversos pro-cesos fisiológicos en la planta, entre ellos lagerminación. Al revés, la germinación esanulada frecuentemente tras una exposi-ción a la luz en el rojo lejano (Botto et al.,1996). Los espectros de absorción de ambasformas, no obstante, se solapan en ambasfranjas. Esto provoca que tras una irradia-ción con luz roja sólo se encuentre hasta un85% de la forma normalmente consideradaactiva Pfr y no más (Salisbury y Ross, 1992;Taiz y Zeiger, 1994). Tras una irradiación conluz roja lejana el fitocromo se revierte a laforma Pr y se puede encontrar hasta 97% dePr (Taiz y Zeiger, 1994).
La germinación y otros procesos en la plan-ta muestran tres modos diferentes de res-puesta regulados por el fitocromo: (a) larespuesta a flujo muy bajo (very-low-fluen-ce response o VLFR) caracterizada por susaturación a flujos y concentraciones de Pfrextremadamente bajas. En este caso no esposible la reversión mediante luz roja leja-na, ya que ésta por si sola establece suficien-te Pfr para inducir una proporción elevadade las respuestas VLFR. (b) La respuesta aflujo bajo (low-fluence-response o LFR) es laconsiderada clásica. Este tipo de respuestase satura con flujos medios de luz roja y esreversible por luz en el rojo lejano aplicadainmediatamente tras el pulso de luz roja.Finalmente (c) las reacciones a irradiaciónalta (high-irradiance reaction o HIR) que nosiguen la ley de reciprocidad, requieren irra-diaciones prolongadas e inhiben la germi-nación (Botto et al., 1995).
30 A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40

El sistema del fitocromo se basa en la pro-porción de las dos formas en equilibrio enlas que se puede encontrar dicho pigmento.Una VLFR típica ocurre con una relaciónPfr/Pr menor que 0,1% y la LFR a partir dePfr/Pr = 1% (a menudo 30%-85%) (Botto etal., 1995).
Investigaciones con mutantes de A. thalianacarentes de fitocromo A y/o B han permiti-do realizar comparaciones con biotipos nor-males. En esta especie se ha encontrado queel fitocromo A induce la germinación trasuna irradiación con luz de extremado bajoflujo y en un rango de longitudes amplio(desde ultravioleta, visible hasta rojo lejano,entre 300 y 780 nm) sin posibilidad de rever-sión correspondiendo a una VLFR. El fitocro-mo B, en cambio, actúa en la reacción foto-reversible clásica respondiendo a luz roja yroja lejana con niveles de flujo (LFR) cuatroveces superior al anterior (Shinomura et al.,1996 y Shinomura, 1997).
Botto et al. (1995 y 1996) concluyeron quelas VLFR son respuestas inductivas de lasque se encarga el fitocromo A. En campo,este fitocromo sería, pues, capaz de detec-tar pulsos de luz muy cortos como los queocurren al realizar un laboreo. Además, per-mitiría la germinación posterior de especiesarvenses en oscuridad siempre y cuando lascondiciones hídricas sean favorables y el cul-tivo ha sido retirado y por tanto ya no lessombree (Casal et al., 1997; Casal y Sánchez,1998).
El fitocromo B parece hacer una mayoraportación a la LFR que a la VLFR, aunqueno sea el único fitocromo que interviene enla LFR (Botto et al., 1995). Al ser capaz dedetectar tanto luz roja no filtrada por lashojas como luz que ha sido filtrada por ellassombreando el suelo, el fitocromo B proba-blemente se encargaría de detectar huecosde luz en la cubierta vegetal, estimulando la
germinación en estas situaciones (Casal etal., 1997).
Shinomura et al. (1994) citado por Botto etal. (1995) dedujeron que la inducción de lagerminación en A. thaliana está reguladaprincipalmente por el fitocromo B y secun-dariamente por el fitocromo A. Los autoresañaden que se puede tratar del fitocromo Bo de otro(s) fitocromo(s) dependiendo delas condiciones. Poppe y Schäfer (1997)encontraron, utilizando mutantes de A. tha-liana carentes de fitocromo A y B, que otrofitocromo de tipo LFR interviene en la ger-minación de las semillas aunque no pudie-ron determinar de qué otro fitocromo setrataba (C, D o E). El papel de cada uno delos fitocromos puede variar entre especies.Los resultados de Dixit y Amritphale (1996)con Celosia argentea L. apoyan su hipótesisque el fitocromo A es el principal fitocromoimplicado en la fotoregulación de la germi-nación en esta especie.
Es más, estos resultados y los de otros auto-res (Milberg et al., 1996; Bai et al., 1996)indican que el tipo de respuestas a la luzpuede ser LFR o VLFR en función de la espe-cie. Casal et al. (1997) afirman que semillasde numerosas especies muestran una res-puesta bifásica, es decir, que reaccionanpositivamente con una VLFR y LFR. Ballaréet al. (1992) modificaron que varias especiespasan de tener respuestas de tipo LFR aVLFR tras un enterrado en el suelo de formanatural. También afirman que las VLFR sonprobablemente frecuentes en semillas quehan estado durante cierto tiempo en elsuelo.
Según las observaciones de Hartmann et al.(1998), la luz de la luna y la de las estrellasfotoestimulan al fitocromo A como unaVLFR, siempre y cuando el fitocromo seencuentre en estado sensible.
La necesidad de luz, no obstante, puede sersustituída por aportes exteriores de gibere-
A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40 31

lina u otras hormonas en determinadasespecies (Salisbury y Ross, 1992; Hou y Simp-son, 1993; Plummer et al., 1995). Según elmodelo fisiológico de la germinación de Hil-horst (1993) descrito por Vleeshouwers etal. (1995), el complejo fitocromo-receptorgenera una señal que conduce a la síntesisde giberelinas, que es el último paso paraestimular la germinación en condicionesnaturales.
Variación estacional de la sensibilidad haciala luz e interacción con otros factoresambientales
La aplicación del laboreo y siembra en oscu-ridad para una reducción de la germinaciónde las especies no deseadas se enfrenta conuna gran complejidad en la fotosensibilidadde las semillas. Incluso impulsos tan sutilescomo las duraciones relativas de día y nochedurante la maduración de las semillas influ-yen en la fotolatencia de algunas especies,siendo inducida normalmente por días lar-gos (Salisbury y Ross, 1992). En Avena fatua,Hou y Simpson (1993) la humedad ambien-tal tras la cosecha de las semillas madurasde Avena fatua condiciona la respuesta alestímulo de luz roja. También la edad de lassemillas altera su estado de latencia (Bai etal., 1996).
Así, la fotosensibilidad no sólo difiere entreespecies sino que las semillas también pue-den modificarla según las condicionesambientales, como, por ejemplo, tras brevesperíodos de calor o frío (Hartmann y Moll-wo, 2002). Esto es debido a que las semillasajustan continuamente sus niveles de laten-cia a las condiciones ambientales estaciona-les (Vleeshouers et al., 1995). Derkx y Kars-sen (1994) encontraron variabilidad en lasensibilidad frente a la luz en A. thaliana, ytambién Ballaré et al. (1992) piensan que lasnecesidades de luz para germinar fluctúanestacionalmente durante el enterrado en el
suelo. Incluso la magnitud de la LFR y de laVLFR varía estacionalmente, estando tam-bién influenciadas por la profundidad en elperfil y por la cubierta del suelo (Derkx yKarssen, 1993, citados por Botto et al., 1995).
Entre los diferentes factores que influyenen las variaciones de la sensibilidad hacia laluz se encuentran la temperatura, humedady presencia de nitratos, entre otros. Vincenty Roberts (1977) estudiaron las fluctuacio-nes de temperatura y en la presencia deiones de nitrato en relación con la presenciao ausencia de luz y concluyeron que laacción de estos factores es muy compleja yraras veces uno de ellos llega a estimular lagerminación independientemente de losdemás. Vleeshouers et al. (1995) citan losestudios de Derkx y Karssen (1993) que des-criben como la sensibilidad de las semillasde Sisymbrium officinale a la luz y los nitra-tos aumentaba cuando la latencia era inte-rrumpida, pero era menor cuando ésta erainducida. En este caso los cambios en laten-cia ocurrieron aún en condiciones de abso-luta oscuridad y sin correlación con cambiosen el contenido endógeno de nitratos (Vle-eshouers et al., 1995, citan a Derkx y Kars-sen, 1993).
Por otro lado, Derkx y Karssen (1994) noencontraron una influencia de los nitratosen la germinación de A. thaliana. Juroszeket al. (2002) relacionaron los niveles de tem-peratura, humedad, nivel de nitratos en elsuelo y pluviometría antes y después de rea-lizar la siembra de 23 ensayos en los que secompararon la densidad de malas hierbasen siembra realizada en oscuridad con lasiembra realizada de día. A pesar de detec-tar reducciones notables de germinación,no se encontraron relaciones entre los fac-tores ambientales y el grado de reducciónde la germinación de las malas hierbas.Queda, pues, manifiesta la complejidad delas interacciones entre la necesidad de luz yotros factores ambientales.
32 A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40

Algunos de los factores ambientales parecenpoder sustituir la necesidad de luz para indu-cir la germinación. Por ejemplo, una tempe-ratura invernal baja puede disminuir laintensidad de luz necesaria para la germina-ción de Sisymbrium officinale (Vleeshouers etal., 1995, citan a Derkx y Karssen, 1993). Estaobservación coincide con la de Kristie y Fiel-ding (1994) que describieron cuantitativa-mente en Lactuca sativa cv. ‘Grand Rapids’como el nivel de Pfr necesario para germinaraumentaba conforme la temperatura subía.También Salisbury y Ross (1992) explicancomo la alternancia de temperaturas entre15 ºC y 25 ºC en Poa pratensis sustituyeron lafotodependencia. Hartmann y Mollwo(2000) pudieron insensibilizar y sensibilizarsemillas de L. sativa a la luz con diversos tra-tamientos de exposición a nitratos y agua atemperaturas elevadas o bajas, respectiva-mente, y demostraron que los cambios desensibilidad pueden ser muy rápidos.
Referente a la presencia de agua, numero-sos autores afirman que la posibilidad deconversión de la forma Pr a la Pfr dependede la hidratación del fitocromo (Bai et al.,1996; Gallagher y Cardina, 1997) así que esnecesaria la imbibición de la semilla parapoder reaccionar frente al estímulo de laluz. Además, Dixit y Amritphale (1996)sugieren que la síntesis de fitocromo se rea-liza durante el período de imbibición enoscuridad. La duración de la imbibicióndepende de la especie, siendo necesarias enArtemisia frigida por lo menos ocho horas(Bai et al., 1996). Para Amaranthus retrofle-xus L., en cambio, las semillas embibidasmás de 12 horas no aumentan su capacidadgerminativa con luz roja y sí lo hacen cuan-do son irradiadas menos tiempo, especial-mente entre 4 y 5 horas con la misma luz(Pita y Durán, 1986).
Gallagher y Cardina (1997) también afir-man que la fotoinducción de la germina-ción en semillas de Amaranthus retroflexus
L. está afectada por el potencial hídrico delsuelo. Encontraron tasas de germinaciónmás elevadas bajo mayores potencialeshídricos. No obstante, no se encontró unainhibición de la germinación completa enesta especie en el tratamiento de sequíaprevio a la exposición a la luz. En otrosexperimentos descritos por los mismosautores encontraron que la fotoinducciónde la germinación en estas semillas fuemenos sensible al estrés hídrico que ensemillas frescas embebidas. Es decir, lassemillas frescas necesitan antes agua queluz. Ascard (1994) cita a Bouwmeester yKarssen (1989), los cuales afirman que ladesecación de semillas embebidas en oscu-ridad induce una independencia a la luz.Ascard (1994) también comenta que el cul-tivo y siembra en oscuridad realizadossobre suelos secos apenas han llevado areducciones de nascencia de malas hierbas.
Todas estas interacciones pueden tenerimportantes efectos en la práctica. Así,Hartmann et al. (2003) encontraron que elsuelo guarda el fotoestímulo en su “memo-ria” durante diferentes períodos de tiemposegún la temperatura, siendo probablemen-te el efecto más persistente al realizar ellaboreo o siembra nocturna en invierno quesi se realiza en verano. Otra implicaciónpráctica es que un suelo seco existen menosposibilidades de que el fotocontrol sea efec-tivo, como han demostrado Juroszek y Ger-hards (2004) en varios trabajos.
Ecología de la fotodependencia
Los mecanismos de fotodependencia de lagerminación y sus complejas interaccionestienen, sin duda, una utilidad para las semi-llas. Según Vleeshouers et al. (1995) la laten-cia innata previene la germinación en perío-dos con condiciones favorables para lagerminación, pero en una época del año enla que se puede esperar que la planta origi-
A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40 33

nada de la semilla no sobreviva o no produz-ca descendencia. Bajo condiciones desfavora-bles faltarán los factores estimulantes de lagerminación y la semilla no germinará, inde-pendientemente de su estado de latencia.
La existencia de al menos dos sistemas dife-rentes para captar la luz en un amplioespectro de longitudes de onda pareceredundante. No obstante, es crucial para eldesarrollo y reproducción vegetal asegurarque el período de germinación y crecimien-to sean los óptimos. Parece ser, pues, que elsistema de los dos tipos de fitocromosaumenta la supervivencia en ambientesfluctuantes (Shinomura, 1997).
Según Casal et al. (1997) el fitocromo Aextrae la siguiente información de las seña-les de luz: a) una exposición breve a luzsolar indica que ha habido laboreo del sueloy probablemente no queden plantas en laproximidad, b) cualquier exposición a la luz(el fitocromo A reacciona en un espectro delongitudes de onda muy amplio) da infor-mación respecto a la posición de la semillaen el suelo, probablemente cerca de lasuperficie.
También Vleeshouers et al. (1995) afirmanque la calidad y cantidad de luz contribuyena que las semillas detecten su profundidadde enterrado y la ausencia de competidoresen su proximidad. Estos autores añaden queotros factores principales que permiten estasensibilidad son temperaturas alternantes ynitratos, siendo todos ellos factores estimu-lantes de la germinación.
Por otro lado, las semillas expuestas a irra-diaciones prolongadas (HIR) expresan unareacción inversa a las LFR y VLFR (Botto etal., 1995). Así, las semillas que queden en lasuperficie tras el laboreo probablemente nogerminarán, protegiéndolas de un posibledesecado debido al exceso de radiación ycalor.
Se trata, pues, de un sistema complejo, en elque las semillas de las hierbas son capacesde reconocer estímulos externos para prote-gerse de condiciones adversas y para apro-vechar las que le sean favorables.
Resultados de la siembra en oscuridadrealizados en campo
La variabilidad de los resultados descritosen la literatura son un reflejo de las comple-jas interacciones entre distintos factoresambientales que condicionan la fotosensibi-lidad. También demuestran los diferentesniveles de sensibilidad que existen entreespecies y, por tanto, los resultados de ensa-yos en los que dominan semillas de especiesmás fotosensibles son más favorables. Cuan-do se realizaron ensayos desglosando elefecto del fotocontrol para cada especie, seencontraron resultados a veces coincidentespero, también a menudo, opuestos. Las efi-cacias de control son desde sorprendente-mente favorables, con reducciones hasta100% (Scopel et al., 1994; Gerhards et al.,1997; Hartmann y Nezadal, 1990; Juroszeket al., 2002), hasta ensayos en los que inclu-so había mayor germinación en las parcelaslabradas y sembradas en oscuridad (Juros-zek et al., 2002).
No obstante, hay especies cuya nascencia hasido muy frecuentemente reducida por ellaboreo y siembra en oscuridad en los ensa-yos publicados como Chenopodium album,Stellaria media, Matricaria spp., Lamiumamplexicaule y/o L. purpureum, Sonchusspp. y Veronica arvensis o V. persicaria o V.hederifolia (tabla 1).
También se han observado reducciones enla germinación debido a la realización detareas de laboreo del suelo y siembra noc-turna en otras especies, pero con menor fre-cuencia que las anteriores, como lo son:,Sinapis arvensis (Jensen, 1992), Myosotis
34 A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40

arvensis (Jensen, 1992; Niemann, 1996),Fumaria officinalis (Gerhards et al., 1997),Papaver rhoeas (Niemann, 1996) Amaran-thus retroflexus, Solanum spp. y Alopecurusmyosuroides (Niemann, 1996; Welsh et al.,1999; Juroszek et al., 2002).
De algunas especies, que estaban presentesen menos ensayos, se dispone de resultadoscontradictorios, con respuesta a la luz enalgunos ensayos pero no en otros: Poaannua (Scopel et al., 1994; Niemann, 1996;Adamiak y Adamiak, 2003), Thlaspi arvense
A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40 35
Tabla 1. Resumen de las especies que han mostrado una reducción en su nascencia en la mayoría delos ensayos realizados. N.d.: no se presentan valores numéricos en la comunicación
Table 1. Summary of the species, which have shown a reduced emergence in most of the trials. N.d.:no numeric data are presented in that communication
Especie Autor Reducción (%)
Chenopodium album Jensen, 1992 24Ascard, 1994; 22
Niemann, 1996 52Gerhards et al., 1997 n.d.
Welsh et al., 1999 70Juroszek et al., 2002 8
Stellaria media Jensen, 1992 27Ascard, 1994 77
Scopel et al., 1994 60**Spiess, 1995 51
Welsh et al., 1999 n.d.Juroszek et al., 2002 80
Matricaria spp. Hartmann y Nezadal, 1990 100Ascard, 1994 16
Niemann, 1996 15Juroszek et al., 2002 31
Adamiak y Adamiak, 2003), 77Lamium amplexicaule Hartmann y Nezadal, 1990 100y/o L. purpureum Scopel et al., 1994 60*
Ascard, 1994 14**Niemann, 1996 86
Sonchus spp. Scopel et al., 1994 60*Gerhards et al., 1997 40
Veronica arvensis Hartmann y Nezadal, 1990 97o V. persicaria Jensen, 1992 30o V. hederifolia Ascard, 1994 14**
Scopel et al., 1994 60*Niemann, 1996 88
Juroszek et al., 2002 29
*: la reducción refiere al total de una población mixta de L. purpureum, Sonchus spp., Cerastium vulgatum, Vero-nica spp., S. media y P. annua y fue de 60%.** la reducción se refiere al total de una población mixta de V. hederifolia, Thlaspi arvense y L. purpureum y fue del 14%.

(Hartmann y Nezadal, 1990; Ascard, 1994;Spiess, 1995) y Viola arvensis (Ascard, 1994;Niemann, 1996; Adamiak y Adamiak, 2003).
Especies de las que parece que sea poco pro-bable esperar reducciones en su nascenciason: Polygonum convolvulus (Jensen, 1992;Gerhards et al., 1997; Juroszek et al., 2002),Galium aparine, Strunk et al. (1995), a pesarde que en algunos ensayos haya reducidoparcialmente su germinación (Juroszek etal., 2002), las gramíneas anuales como Seta-ria viridis, S. faberii y S. pumila así comoEchinochloa crus-galli (Buhler, 1997) y espe-cies de multiplicación vegetativa como Cir-sium arvense (Gerhards et al., 1997).
Buhler (1997) y Scopel et al. (1994), en Esta-dos Unidos, incluyeron en sus ensayos espe-cies que normalmente germinan en prima-vera y verano. Así, Buhler (1997) encontróreducciones en la germinación de C. album,Solanum ptycanthum, Polygonum pensylva-nicum y en Amaranthus spp. debido al labo-reo nocturno. También Scopel et al. (1994)encontraron reducciones de hasta cincoveces sobre la germinación de Amaranthusretroflexus y Solanum spp. Otras especies detamaño de semilla mayor (Xanthium stru-marium, Abutilon theophrasti), en cambio,no mostraron reducciones en la germina-ción. No obstante, no se dispone de másdatos sobre estas especies, por lo que la efi-cacia del control fotobiológico debe seguirsiendo estudiada.
En cuanto a la cuantía de reducción, en oca-siones fue suficiente para mantener la den-sidad de malas hierbas a un nivel inferior alumbral de tolerancia (Gerhards et al., 1997).Según la revisión realizada por Juroszek yGerhards (2004) la reducción debido allaboreo o siembra nocturnas, si ocurre,suele situarse en cerca de un 30%, por loque cabe tener en cuenta que se trata de unmétodo preventivo que debe ser combina-do con otras técnicas de manejo o control.
Aparte de realizar la siembra en oscuridadse deben tener en cuenta otros aspectosagronómicos que afectarán la eficacia delmétodo, como la capacidad competitiva delcultivo, la época de siembra, las labores rea-lizadas, etc. Por ejemplo, Gerhards et al.(1997) contabilizaron reducciones de 18%hasta 80% según el campo y la época desiembra, siendo los cereales de primaveralos más favorecidos por la técnica. Esto esdebido a que su rápido crecimiento los hacemás competitivos, de manera que en estoscasos no fue necesaria ninguna otra técnicade control. También encontraron que ger-minaron menos semillas tras un laboreoprofundo que tras uno superficial. En elcaso de Hartmann y Nezadal (1990), querealizaron ensayos durante 7 años consecu-tivos sobre una superficie en barbecho, nofue necesario utilizar herbicidas duranteeste período.
La penetración de la luz en el suelo
Niemann (1996) cree que las discrepanciasentre especies en los diferentes ensayospueden ser debidas a la profundidad desdela que germinan las diferentes semillas.Observaron que una especie de semillamediana (Veronica hederifolia) germinabadesde capas superiores del suelo cuandoéste había sido movido de noche. En los sue-los que fueron labrados de día, en cambio,las plantas germinaron también desdecapas inferiores. Probablemente, en el tra-tamiento de laboreo en oscuridad, las semi-llas sólo germinaron desde aquellas capasque recibieron luz aunque serían capaces dehacerlos desde capas inferiores si hubiesenrecibido el estímulo necesario.
Para poder contar con una reducción de lagerminación de las especies arvenses debidoal laboreo y a la siembra en oscuridad esnecesario, por un lado, que las semillas seencuentren en un estadio de fotodepen-
36 A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40

dencia para germinar. Por otra parte, esnecesario que la luz no llegue a penetrarhasta la posición en la que han quedado lassemillas aunque hayan sido subidas a lasuperficie por los aperos pero queden ente-rradas de nuevo. Según Wooley y Stoller(1978) es posible, por lo general, que la luzdesencadene una germinación hasta los 2mm de profundidad. Sin embargo, estodependerá de las especies. Hartmann yNezadal (1990) dedujeron que la mayoríade las semillas de malas hierbas en uncampo de cultivo no son fotoestimuladaspor la luz que penetre menos de 1 cm en elsuelo. Parece que la causa principal de lagerminación está en las exposiciones cortasy repetidas que ocurren durante el laboreode día. No obstante, esto no significa quelas semillas enterradas a más de 1 cm deprofundidad ya no germinen, ya que exis-ten necesidades de Pfr en la última fase dela germinación que pueden ser satisfechaspor VLFR si las semillas están suficientemen-te cerca de la superficie del suelo.
El laboreo y siembra en oscuridad afecta-rán, pues, a aquellas semillas que muestrenfotodependencia a la luz en ese momento yque después de la siembra hayan quedado amayor profundidad de 1 cm, impidiendo sugerminación. Las semillas que queden ensuperficie tras el laboreo probablemente nogerminarán, ya que expresan una reaccióninversa a las LFR y VLFR (Botto et al., 1995).
Conclusiones: viabilidad del controlfotobiológico
Según los resultados de los ensayos denumerosos investigadores existe una granvariabilidad en la fotosensibilidad entreespecies, localidades y años. El primer factora considerar, por lo tanto, es la composicióndel banco de semillas del campo a controlar.En función de las especies cabrá esperar, de
entrada, una cierta reacción al tratamientoo no. El segundo aspecto a considerar esque la necesidad de luz para germinar varíamucho debido a factores internos de lassemillas (según su origen, su edad, etc.) ydebido a factores externos como son lasfluctuaciones de temperatura, humedad,presencia de nitratos, etc. de difícil controlpor parte del agricultor.
Por ello, la siembra o laboreo realizados enoscuridad pueden comportar reduccionesde germinación desde insignificantes hastamuy importantes, según el estado de foto-sensibilidad en el que se encuentren lassemillas. Se trata, pues, de un método decontrol de malas hierbas con efectos muydependientes de las condiciones ambienta-les y, de momento, poco previsible.
Basándonos en la literatura consultada losmejores resultados se pueden obtener ensuelos húmedos con un contenido en nitra-tos bajo y una alta proporción de especiesde semilla pequeña (Ascard et al., 1994;Juroszek y Gerhards, 2004). Parece ser queen condiciones de sequía, por ejemplo, serápoco probable que la ausencia de luz seaclave para inhibir la germinación.
Otro aspecto a tener en cuenta es que inclu-so si se obtuvieran grandes reducciones en lagerminación, el banco de semillas no se redu-ciría, ya que las semillas no germinadassiguen en el suelo. La ventaja del fotocontrolradicaría en que el banco no aumenta connuevas semillas descendientes de éstas.Como comentan diversos autores, el controlfotobiológico, igual que cualquier otra estra-tegia de control se debe combinar y/o alter-nar con otras técnicas con el fin de no provo-car una selección en el banco de semillas.
El coste de sembrar o realizar las labores enoscuridad recubriendo los aperos con lonases muy reducido y se han obtenido resulta-dos equivalentes a realizar estas operacionesen oscuridad. Por ello, puede ser recomen-
A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40 37

dable utilizar esta técnica en agriculturaecológica, ya que se deben utilizar todas lasposibilidades existentes para sumar un efec-to de control aceptable sobre las malas hier-bas. La posibilidad de que germinen másmalas hierbas por el momento no tiene basecientífica por lo que cabe esperar que reali-zando esta técnica germinen un númeroigual o inferior de plantas.
Agradecimientos
Los autores agraden la amable colaboracióndel Dr. Peter Juroszek y al Prof. Dr. KarlHartmann por proporcionar la bibliografíaeditada en revistas de divulgación nacional.
Bibliografía
Adamiak E, Adamiak J, 2003. PhotobiologischeVerunkrautungsregulierung im Winterwei-zen. Mitt. Ges. Pflanzenbauwiss. 15, 294-295.
Ascard J, 1993. Soil cultivation in daylioght witha light-proof cover on the harrow reducedweed emergence. Proceedings of the 4th Inter-national IFOAM Conference. Non-chemicalweed control, Dijon, France, 211-214.
Ascard J, 1994. Soil cultivation in darkness redu-ced weed emergence. Acta Horticulturae 372,167-177.
Bai Y, Romo JT, Hou J, 1996. Phytochrome actionin seed germination of fringed sage (Artemi-sia frigida). Weed Science 44, 109-113.
Ballaré CL, Scopel A, Sánchez RA, RadosevichS.R., 1992. Photomorphogenetic processes inthe agricultural environment. Photochemistryand Photobiology, 56 (5), 777-788.
Baskin CC, Milberg P, Andersson L, Baskin JM,2002. Non-deep simple morphological dor-many in seeds of the weedy facultative winterannual Papaver rhoeas. Weed Research 42,194-202.
Botto JF, Sánchez A, Casal JJ, 1995. Role ofphytochrome B in the induction of seed ger-mination by light in Arabidopsis thaliana.Plant Physiol., 146, 307-312.
Botto JF, Sánchez RA, Whitelam GC, Casal JJ,1996. Phytochrome A mediates the promotionof seed germination by very low fluences oflight and canopy shade light on Arabidopsis.Plant Physiol., 110, 439-444.
Buhler, 1997. Effects of tillage and light environ-ment on emergence of 13 annual weeds.Weed Technology 11, 496-501.
Casal JJ, Sánchez RA, Yanovsky MJ, 1997. Thefunction of phytochrome A. Plant, Cell andEnvironment, 20, 813-819.
Casal JJ, Sánchez RA, 1998. Phytochromes andseed germination. Seed Sci. Res. 8, 317-329.
Derkx MPM, Karssen CM, 1994. Are seasonal dor-mancy patterns in Arabidopsis thaliana regu-lated by changes in seed sensitivity to light,nitrate and gibberelin? Annals of Botany, 73(2), 129-136.
Dixit S, Amritphale D., 1996. Very low fluenceand low fluence response in the induction andinhibition of seed germination in Celosiaargentea. Seed Science Research, 6 (2), 43-48.
Gallagher RS, Cardina J, 1997. Soil water thres-holds for photoinduction of redroot pigweed.Weed Science, 45, 414-418.
Gerhards R, Kühbauch W, Juroszek P, 1997. Tiefen-grubber, Scheibenegge und zwei Krümelwal-zen. Lichtlose Bodenbearbeitung reduziertUnkrautwuchs. Mitteilungen der DeutschenForschungsgemeinschaft, 4, 14-16.
Hartmann KM, Goetz S, Market R, Kaufmann T,Schneider K, 2003. Photocontrol of weed ger-mination: Lightless tillage and variablememory of the seedbank. Aspects of AppliedBiology 69, 237-246.
Hartmann KM, Mollwo A, 2002. The action spec-trum for maximal photosensitivity of germina-tion and significance for lightless tillage. 5thEWRS Workshop on Physical Weed Control,Pisa, Italy, 70-78.
38 A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40

Hartmann KM, Mollwo A, 2000. Photocontrol ofgermination: sensitivity shift over eight deca-des within one week. Z. PflKrankh. PflSchutz,Sonderh. XVII, 125-131.
Hartmann KM, Mollwo A, Tebbe A, 1998. Photo-control of germination by moon- and star-light. Z. PflKrankh. PflSchutz, Sonderh. XVI119-127.
Hartmann KM, Krooss C, Mollwo A, 1997.Phytochrome-mediated photocontrol of thegermination of the scentless mayweed, Matri-caria inodora L. and its sensibilization bynitrate and temperature. J. Photochem. Pho-tobiol. 40, 240-252.
Hartmann KM, Nezadal W, 1990. Photocontrol ofweeds without herbicides. Naturwissenschaf-ten, 77, 158-163.
Hatcher PE, Melander B, 2003. Combining physi-cal, cultural and biological methods: propectsfor integrated non-chemical weed manage-ment strategies. Weed Research 43, 303-322.
Hou JQ, Simpson GM, 1993. Germination respon-se to phytochrome depends on specific dor-mancy states in wild oat (Avena fatua). Can. J.Bot., 71, 1528-1532.
Jensen PK, 1992. First Danish experiences withphotocontrol of weeds. Z. PflKrankh.PflSchutz, Sonderh. XIII, 631-636.
Jensen PK, 1995. Effect of light environmentduring soil disturbance on germination andemergence pattern of weeds. Ann. appl. Biol.127, 561-571.
Juroszec P, Gerhards R, Kühbauch W, 2002. Pho-tobiologische Unkrautregulierung annuellerAckerunkräuter. J. Agronomy & Crop Science188, 389-397.
Juroszek P, Gerhards R, 2004. Photocontrol ofweeds. J. Agronomy & Crop Science 190 (6),402-415.
Klümper H, Gerhards R, Kühbauch W, 1996. Ein-fluss des Lichtes auf die Keimung vonUnkrautsamen. Z. PflKrankh. PflSchutz, Son-derh. X, 1-7.
Kristie DN, Fielding A, 1994. Influence of tempe-rature on the Pfr level required for germina-tion in lettuce cv. Grand Rapids. Seed ScienceResearch, 4 (1), 19-25.
Kühbauch W, Gerhards R, Klümper H, 1992.Unkräuter durch nächtliche Bodenbearbei-tung bekämpfen? Pflanzenschutz-Praxis 1, 13-15.
Liebman M, Gallandt ER, 1997. Many little ham-mers: ecological management of crop-weedinteractions. En: Ecology in Agriculture. L.E.Jackson (Ed.), 291-343. Academic Press, SanDiego, CA, USA.
Milberg P, Andersson L, Noronha A, 1996. Seedgermination after short-duration light expo-sure: implications for the photo-control ofweeds. J. of Applied Ecology, 33, 1469-1478.
Milberg P., Andersson L., Thomson K., 2000.Large-seeded species are less dependent onlight for germination than small-seeded ones.Seed Sci. Res. 10, 99-104.
Niemann P, 1996. Unkrautbekämpfung durchLichtausschluss während der Bodenbearbei-tung. Z. PflKrankh. PflSchutz, Sonderh. XV,315-324.
Niemann P, Biller RH, 1996. Eine geeignete Tech-nik zur Umsetzung der photobiologischenBekämpfung annueller Unkräuter. Nachrich-tenblatt Deut. Pflanzenschutzd. 48 (11), 229-233.
Pita JM, Durán JM, 1986. Germinación en elgénero Amaranthus L.: II. Papel del fitocromoen A. retroflexus L. ITEA 63, 61-69.
Plummer JA, McChesney CJ, Bell DT, 1997. Germi-nation in photosensitive seeds: does phytoch-rome stimulate metabolism of GA19 and GA20to GA1? Aust. J. Plant Physiol., 24, 389-394.
Poppe C, Schäfer E, 1997. Seed germination ofArabidopsis thaliana phyA/phyB doublemutants is under phytochrome control. PlantPhysiol. 114, 1487-1492.
Powles SB, Matthews JM,1996. Integrated weedmanagement for the control of herbicideresistant annual ryegrass (Lolium rigidum).Proceedings of the Second International
A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40 39

Weed Control Congress, Copenhagen, Den-mark, 1-7.
Rasmussen K, 2002. Influence of liquid manureapplication method on weed control in springcereals. Weed Research 42, 287-298.
Rasmussen K, 2000. Can slurry injection improvethe selectivity of weed harrowing in cereals?Proceedings of the 4th reunión del Grupo deTrabajo EWRS sobre Control de Malas Hierbascon Métodos Físicos y Culturales, Elspeet, Paí-ses Bajos, 33-34.
Reed JW, Nagatani A, Elich TD, Fagan M, Chory J,1994. Phytochrome A and phytochrome Bhave overlapping but distinct functions inArabidopsis development. Plant Physiol., 104,1139-1149.
Salisbury FB, Ross CW, 1992. Photomorphogene-sis, pp. 438-463. En: Plant Physiology. N.G. Phi-lip (Ed.)., 540 pp. Wadsworth Publishing Com-pany, Belmont, California (EEUU).
Scopel AL, Ballaré CL, Radosevich SR, 1994. Pho-tostimulation of seed germination during soiltillage. New Phytologist, 126 (1), 145-152.
Shinomura T, 1997. Phytochrome regulation ofseed germination. Journal of Plant Research,110 (1097), 151-161.
Shinomura T, Nagatani A, Hanzawa H, KubotaM, Watanabe M, Furuya M, 1996. Action spec-tra for phytochrome A- and B-specific pho-toinduction of seed germination in Arabidop-sis thaliana. Proceedings of the NationalAcademy of Sciences of the United States ofAmerica, 93 (15), 8129-8133.
Spiess H, 1995. Zur Praxisrelevanz der photobio-logischen Unkrautregulierung. Beitr. 3. Wiss.-Tagung Ökol. Landbau, Kiel, 73-76.
Strunk H-B, Schliephake U, Trautz D, 1995. Pho-tobiologische Beikrantregulierung in Mais-Beitr. 3 Wiss-Tagung Ökol-Landbau, Kiel, Ale-mania, 289-292.
Taiz L, Zeiger E, 1994. Phytochrome and Photo-morphogenesis. En: Plant Physiology, 690 pp.Benjamin/Cummings Publishing Company Inc.,Redwood City, California, EEUU.
Vincent EM, Roberts EH, 1977. The interaction oflight, nitrate and alternating temperature inpromoting the germination of dormant seedsof common weed species. Seed Science andTechnology 5, 659-670.
Vleeshouers LM, Bowmeester HJ, Karssen CM,1995. Redefining seed dormancy: an attemptto integrate physiology and ecology. Journalof Ecology, 83, 1031-1037.
Welsh JP, Phillips L, Bulson HAJ, Wolfe M, 1999.Weed control strategies for organic cerealcrops. The 1999 Brighton Conference – Weeds,945-950.
Wesson G, Wareing PF, 1969. The role of light inthe germination of naturally occurring popu-lations of buried weed seeds. J. Exp. Bot, 2,402-413.
Wooley & Stoller, 1978. Light penetration andlight-induced seed germination in soil. PlantPhysiology 61, 597-600.
(Aceptado para publicación el 6 de junio de 2005).
40 A. Cirujeda, A. Taberner ITEA (2006), Vol. 102 (1), 27-40

J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54 41
Análisis del progreso genético obtenido en el esquema deselección de la raza ovina manchega
J.J. Jurado*, M. Serrano*, Mª D. Pérez-Guzmán**
*Dpto. de Mejora Genética Animal y Biotecnología. Instituto Nacional de Investigación y TecnologíaAgraria y Alimentaría (INIA). Carretera de La Coruña, Km. 7,00. 28040 Madrid, España.e-mail: [email protected], [email protected]**Centro Regional de Selección y Reproducción Animal (CERSYRA). Avda. del vino 2. 13300 Valdepeñas.Ciudad Real, e-mail: [email protected]
ResumenTras varios años de funcionamiento se analiza con detalle la tendencia genética anual obtenida en elEsquema de Selección genética de la raza ovina Manchega. Dicha tendencia (0,818±0,057 litros/año)presenta un valor inferior al de otras razas españolas y supone solo un 0,84% de la media fenotípica.
Las razones de este bajo valor es, en parte, achacable a los animales con genealogía desconocida. Ladescomposición de la tendencia genética en partes atribuibles a las vías de selección genética muestraque la vía padre de sementales no tiene apenas relevancia en la tendencia total, mientras que la víamadres de sementales es responsable del 60% de la misma. Se puede concluir que es la vía “padres desementales” la responsable de la baja tendencia total ya que los sementales testados en los primerosaños eran de calidad genética no muy elevada por tener padres poco selectos (no así sus madres, quesí presentaban un alto valor genético). El problema esta resuelto en la actualidad, ya que se exigen alos candidatos a futuros sementales un elevado índice de pedigrí.
Palabras clave: Tendencia genética, Vías de selección, Ovino lechero
SummaryAnalysis of the genetic progress obtain in the selection program in Manchega sheep breedThe realised genetic trend in the Manchega sheep breed is analysed in detail after several years ofimplementation of the selection scheme. The estimated trend (0.818±0.057 litres/year) is lower thanestimated trends in other Spanish sheep breeds schemes, being a 0.84 % of the phenotypic mean. Theestimated contributions of each path of selection indicate that the reasons for this low value are part-ly due to the fact that the contribution of the sires of sires path to the total genetic gain is negligible,while the dams of sires path is responsible for 60 % of the total gain. The low contribution of the siresof sires path is explained by the low genetic level of the sires under progeny test in the first years, dueto the low genetic merit of their own sires (although their dams were of high merit). Nowadays, thisproblem has been solved since a high pedigree index is required when selecting candidates to be siresof future generations.
Key words: Genetic trends, Selection paths,Dairy sheep

42 J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54
Introducción
En España están en funcionamiento progra-mas de mejora genética en ganado ovinode leche en la razas Latxa (Legarra et al.,2003), Churra (Merino y Olmedo, 1991),Assaf (Jiménez y Jurado, 2005a) y Manche-ga (Montoro et al. 1996). De estos, los de laraza Latxa, Churra y Manchega tienen suscomienzos en la década de los años ochen-ta. El caso de la raza Assaf es diferente yaque es una raza que fue reconocida en 2004aunque estaba implantada en España desdehace muchos años y hay varios grupos quellevan a cabo mejora genética sin coordina-ción entre ellos. El que cuenta con mayoresmedios y tiene el programa mas completoes el financiado y coordinado por la diputa-ción provincial de León, incluyendo valora-ción genética de reproductores desde 1999(Jiménez y Jurado, 2005b).
El esquema de selección genética de la razaovina Manchega (ESROM) inició su andadu-ra en el año 1986, pero no fue hasta 1990cuando se llevaron a cabo las primeras valo-raciones genéticas de reproductores y cuan-do se empezó a seleccionar reproductorespor su mérito genético. Tras algunos años defuncionamiento se empezó a calcular regu-larmente la tendencia genética obtenida(Pérez-Guzmán et al., 1997). Ahora, tras másde 14 años de funcionamiento es momentode analizar con mayor detalle el progresogenético obtenido, verificando si el rendi-miento del esquema de selección es similaral de otros que están próximos al mismo ycomprobando así el uso eficiente de losmedios empleados.
El ESROM, como la mayoría de los esquemasde selección que están desarrollándose enEspaña, esta concebido para la detección dereproductores de elevado mérito genético ysu utilización posterior de la forma más efi-ciente posible. De esta forma, se seleccionansementales y ovejas reproductoras mediante
la elección de sus progenitores, y así se hablade padres de sementales (PS), madres desementales (MS), padres de ovejas reproduc-toras (PO) y madres de ovejas reproductoras(MO), las cuatros vías de la mejora incluidasen la clásica teoría de la respuesta a la selec-ción desarrollada por Dickerson y Hazle(1944) y posteriormente por Rendel y Robert-son (1950).
La eficiencia de un esquema de seleccióndepende de que el progreso genético gene-rado en cada una de las vías de mejora seael adecuado en función de los recursos utili-zados. La vía PS es la más importante debi-do, básicamente, a la mayor capacidad delos sementales para dejar descendencia ypor tanto la elección de sementales requie-re mayor cuidado ya que su influencia en lapoblación será de mayor trascendencia. Lavía MS es la complementaria a la PS, perodebido a la menor precisión con que sesuele estimar el mérito genético de la hem-bras y a su menor capacidad para dejar des-cendencia, su importancia es algo menor. Lavía PO es la tercera en importancia por lamismas razones que la PS pero en este casolos sementales son de menor mérito genéti-co pues se necesitan más sementales paracubrir las necesidad de animales de reposi-ción en la población. Por ultimo la vía MO esla de menor importancia pues el número dehijos que una oveja deja en un rebaño sueleser bastante reducido. Como punto de refe-rencia se puede mencionar el caso del gana-do vacuno de leche en el que la vía PS supo-ne el 43% de toda la mejora, la vía PO el18%, la vía MS el 33% y por ultimo la víaMO el 6% (Groen Ab.F., 1990)
Así pues, una forma eficaz de averiguar siun esquema de selección está desarrollandotodo su potencial es analizar los progresosgenéticos realmente obtenidos por cada víay compararlos con los que se deberían obte-ner desde un punto de vista teórico, bien enotras especies o bien con los obtenidos por

J.J. Jurado et al. ITEA (2005), Vol. 102 (1), 41-54 43
otras razas ovinas explotadas en ambientesy circunstancias semejantes.
El ESROM es un clásico esquema de selec-ción que integra varios rebaños que estánconectados mediante machos de referencia,los cuales pueden ser machos mejorantes,de élite, o simplemente machos en testaje(Jurado y Serrano, 1997). La conexión de losrebaños permite una valoración genética delos animales mediante metodología BLUPque los hace comparables entre si aunqueestén en rebaños diferentes (Jurado, 1994).El objetivo de la selección es incrementar laproducción de leche por lactación y oveja yel criterio de selección es la cantidad deleche tipificada a 120 días. La estima de losvalores genéticos se lleva a cabo medianteun modelo animal con medidas repetidas ygrupos genéticos. Los parámetros genéticosutilizados para el carácter seleccionado enesta valoración son de 0.18 para la hereda-bilidad y de 0.35 para la repetibilidad(Serrano et al., 1996). También se valoranlos animales por características de calidadde la leche (porcentaje de grasa, proteína yextracto seco). La conexión de rebaños ydifusión de la mejora se lleva a cabomediante Inseminación Artificial (IA) consemen refrigerado. Los sementales delesquema están situados en el Centro Regio-nal de Selección y Reproducción Animal(CERSYRA) de Valdepeñas que actúa comoCentro de Inseminación Artificial (CIA), endonde está centralizado el control de pro-ducciones tanto de la cantidad como de lacalidad de la leche. Este centro cuenta conlas instalaciones y el personal necesariospara realizar controles de paternidadmediante marcadores genéticos (microsaté-lites).
El propósito de este trabajo es presentar unanálisis detallado de la tendencia genéticaobtenida en el ESROM, descomponiéndolaen partes atribuibles a cada vía de selección,compararla con la obtenida en otras espe-
cies y razas para el mismo carácter, observarlas discrepancias y buscar sus causas y, porúltimo, proponer soluciones prácticas.
Material y método
Para llevar a cabo el análisis antes descritose utilizó el control de producciones de laraza ovina Manchega y el registro genealó-gico utilizado para efectuar la valoracióngenética hasta Abril de 2004. En el primerose acumulan lactaciones desde 1979 y en elsegundo hay genealogías de animales naci-dos desde 1971.
La genealogía presenta información de179.695 animales perteneciente a 302 reba-ños históricamente adscritos al esquema.De estos 141.519 tienen año de nacimiento.Existen hijas de 735 machos diferentes y elnúmero de ovejas con padre conocido es de24.833 pertenecientes a 161 rebaños lagran mayoría procedentes de IA. Puestoque la valoración genética sólo tiene senti-do en aquellos rebaños que están bienconectados por machos de referencia, seconsideró que un rebaño tenía una cone-xión de calidad con los otros si había almenos 20 hijas de 5 machos diferentes. Enestas condiciones había 121 rebaños coninformación de 152.236 ovejas de la cuales24.520 tenían padre conocido. De éstas,126.093 presentaban año de nacimiento.Todos los resultados se referirán a los ani-males de rebaños conectados.
Para cada vía de selección se calculó el inter-valo generacional (LPS, LMS, LPO y LMO). Estefue definido como la media de los añostranscurridos desde el nacimiento de losreproductores y el de su progenitor.
Las medias de los valores genéticos predi-chos fueron calculadas según el año de naci-miento y para cada vía de selección. Dichasmedias se calcularon de dos forma diferen-

44 J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54
tes: ponderadas por el número de descen-dientes y no ponderadas. La media no pon-derada se puede considerar que es una esti-ma de valor genético medio de losreproductores disponibles para hacer lamejora. La media ponderada estima el valorde los reproductores realmente usados, unrepresentante típico de reproductor encada vía. (Van Tassell y Van Vleck, 1990). Porconsiguiente reproductores “disponibles”son con los que se cuentan para ser poten-cialmente utilizados como tales (se usen ono) y “usados” son los empleados verdade-ramente de entre los disponibles.
La tendencia genética de una vía de selec-ción se calcula en todos los casos como elcoeficiente de regresión de la media de losvalores geneticos predichos de los reproduc-tores nacidos un año determinado paracada vía de selección (GPS, GMS, GPO, y GMO)sobre la unidad de tiempo (T), que fuetomada como el año de nacimiento delreproductor (bGPST, bGPOT, bGMST, bGMOT). Enocasiones, la tendencia genética se expresacomo porcentaje de la media fenotípica yde la desviación típica aditiva.
De la misma forma es posible calcular la ten-dencia genética de un grupo determinadode animales, calculando el coeficiente deregresión lineal de la media de los valoresgenéticos predichos de los animales nacidoscada año en ese grupo sobre la unidad detiempo. Así es interesante averiguar la ten-dencia genética de los sementales disponi-bles y de las ovejas elegidas como madresdisponibles, así como la de las ovejas conpadre conocido frente a las de padres des-conocidos.
La tendencia genética anual de toda lapoblación se puede calcular de dos formasequivalentes:
a) Mediante el coeficiente de regresión delvalor genético medio de todas las ovejasnacidas cada año sobre la unidad de tiempo
(T) (bGOT), tomado T como año de nacimien-to de reproductor.
b) Mediante la formula sugerida por Rendely Robertson (1950):
=
o lo que es lo mismo:
Esta última forma presenta la ventaja depoder descomponer la tendencia genéticatotal en partes atribuibles a cada vía deselección, mediante las expresiones:
;
;
Todos los resultados obtenidos por estasexpresiones serian aproximados ya que suexactitud es progresiva de forma asintóticacon las generaciones.
Existen otros métodos más precisos y adap-tables a situaciones particulares como el pro-puesto por ELSEN (1993), pero que requiereun mayor esfuerzo de cálculo.
Debido a que muchos reproductores de lasvías de selección (abuelos) no tienen año denacimiento, se estimó dicho año por dosprocedimientos:
a) Calculando la media del año de nacimien-to de sus hijos y detrayendo el intervalogeneracional de dicha vía.
∆gb L
LMOG T MOMO=∑
.∆g
b L
LMSG T MSMS=∑
.
∆gb L
LPOG T POPO=∑
.∆g
b L
LPSG T PSPS=∑
.
∆gb L b L b L b L
L L L LG T PS G T PO G T MS G T MO
PS PO MS MO
PS PO MS MO=+ + +
+ + +. . . .
∆G
L∑∑
∆ ∆ ∆ ∆ ∆g
G G G G
L L L LPS PO MS MO
PS PO MS MO
= + + ++ + +

J.J. Jurado et al. ITEA (2005), Vol. 102 (1), 41-54 45
b) Detrayendo el intervalo generacional dela vía del año de nacimiento del hijo másjoven
La información utilizada en cada una de lasvías de selección se presenta separadamente:
Vía padre de sementales (PS):
Número de ovejas con padres 25.033Número de sementales diferentes 733Número de sementales con padres(abuelos) 476Número de abuelos paternos diferentes 179 Número medio de hijas por semental 34,154 Número medio de sementales porabuelo paterno 2,173 Mayor número de sementales por abuelo paterno 24
De los 179 abuelos solo 69 tienen año de nacimiento (38,5%).
Vía padre de ovejas reproductoras (PO)
Número de ovejas con madres conocidas 72.314 Número de madres diferentes 41.420 Número de madres con padres(abuelos) 5405 Número de abuelos maternosdiferentes 455 Número medio de ovejas por madres 1,745 Número medio de madres por abuelo paterno 9,822 Mayor número de madres porabuelo paterno 173
De los 455 abuelos maternos, 342 tienen año de nacimiento (75,1%)
Vía madre de sementales (MS)
Número de sementales conmadres (abuelas) 480 Número de madres de sementalesdiferentes 411
Número medio de sementales por abuela materna 1,116 Mayor número de sementales por abuela materna 3
De las 411 abuelas diferentes, 357 tiene año de nacimiento (86,8%)
Vía madres de ovejas reproductoras (MO)
Número de ovejas con madres (abuelas) 21.652 Número de abuelas maternas diferentes 14.886 Número medio de ovejas por abuela materna 1,388 Número máximo de ovejas por abuela materna 9
De las 14886 abuelas diferentes, 11091 tienen año de nacimiento (74,5%)
Resultados y discusión
En la figura 1 se presenta la tendencia gené-tica estimada de la población de rebañosconectados. Su valor es bGOT=0,818±0,057litros por año (l/a), lo que supone un incre-mento genético anual de un 0,84% de lamedia fenotípica (µP=97,46 l.) y un 5,13%de la desviación típica aditiva (σA=15,94 l.).Es claramente distinto de cero y pone enevidencia que el programa de seleccióngenética de esta raza es eficiente en modifi-car el valor genético medio de la población.No obstante, es inferior al valor señalado enotras razas españolas, como la raza Latxa(2,97 l/a) ( Legarra et al., 2003), que suponeun 2,52%. En el caso de la raza Churra esteseria de 0,96 l/a (De la fuente, 2005, (Comu-nicación personal)).
En la figura 2 se presenta la tendencia gené-tica de las ovejas con padre conocido(1,470± 0,079) y desconocido (0,395±0,043).

46 J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54
Figura 1. Tendencia genética de la raza ovina Manchega. Media de los valores aditivos estimados poraño de nacimiento de las ovejas de rebaños conectados.
Figure 1. Genetic trend in Manchega breed. Average estimated breeding values of ewes born ineach year in connected flocks.
Es bien claro que gran parte de la mejoragenética de la raza viene dada por las ove-jas hijas de machos conocidos, y que en suinmensa mayoría proceden del CERSYRA yhan tenido hijas en los rebaños medianteIA. Se podría argumentar que la metodolo-gía BLUP altera el valor de las prediccionesgenéticas de los animales cuando se desco-noce su genealogía, haciendo que éstastiendan a cero. No obstante la mayor partede las ovejas tienen información propia devarias lactaciones y en muchos casos seconoce la madre, lo cual puede hacer queesta propiedad de la metodología no modi-fique sustancialmente la conclusión ante-rior. Además la diferencia entre los valoresde ambas tendencias es estadísticamentediferente de cero.
En la figura 3 se han añadido a las líneascorrespondientes a las tendencias genéticade hijas con padres conocidos y desconoci-dos, la de la tendencia general de la pobla-ción y la de la tendencia genética de los
sementales disponibles (1,746±0,285) y demadres de ovejas disponibles (0,905±0,086)en cada año. Se observa que la línea corres-pondiente a hijas de progenitores conoci-dos transcurre entre la de sementales y lade ovejas madres, cosa por demás lógica,pero que pone en evidencia que el bajovalor de la tendencia general de la pobla-ción es en gran parte atribuible a los anima-les de genealogía desconocida.
Aunque el valor de la tendencia genéticade animales con padres conocidos (1,470)es superior a la de la población (0.818), tansolo supone un 1,5% de µP (9,22% de σA) ,lejos de los valores de otras razas. Luegono deben ser solo los animales de genealo-gía desconocida los responsables del bajonivel de respuesta genética de la raza. Laexplicación viene dada por los padres ymadres de los sementales y de las ovejas. Acontinuación se analizan las cuatro vías deselección para los animales de genealogíaconocida.

J.J. Jurado et al. ITEA (2005), Vol. 102 (1), 41-54 47
Figura 2. Tendencia genética de la raza ovina Manchega. Media de los valores aditivos estimados poraño de nacimiento de las ovejas con padre conocido ó desconocido.
Figure 2. Genetic trend in Manchega breed. Average estimated breeding values of ewes born ineach year with sires know or unknown.
Figura 3. Tendencia genética de la raza ovina Manchega. Media genética anual de las ovejas derebaños conectados, con ambos progenitores conocidos y desconocidos, así como de los sementales
y madres reproductoras.Figure 3. Genetic trend in Manchega breed. Average estimated breeding values of ewes born in
each year in connected flocks, with both parents know and unknown, rams and dams.

48 J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54
Vía padres de sementales (PS)
En la figura 4 se presentan las medias gené-ticas anuales de los padres de sementalesdisponibles cada año y los realmente usados.
Los valores de sus tendencias (0,333±1,163 y–0,207±1,795) no son diferentes de cero.Además de la nula tendencia genética seobserva una gran irregularidad en el valorgenético medio de los mismos.
Figura 4. Tendencia genética de la raza ovina Manchega. Vía padres de sementales. Medias genéti-cas por año de nacimiento de los sementales padres de machos usados y disponibles (ponderadas y
no ponderadas por el número de hijos).Figure 4. Genetic trend in Manchega breed. Selection path sires of rams. Average estimated bree-
ding values of sires of rams born in each year, actually used and potentially available (weighted andunweighted by number of sons).
Los padres de los sementales, según el fun-cionamiento del esquema de selección, sonaportados por los ganaderos. En los prime-ros años del esquema está claro que losmachos ingresados al CERSYRA no eran deuna calidad genética elevada, debido a lasreservas de algunas ganaderos ante unastécnicas nuevas para ellos y a que, paralograr su colaboración, no se era muyestricto respecto a la genealogía de estosmachos (en muchos casos desconocida). Porotra parte solamente se conoce el año de
nacimiento del 38% de estos lo que contri-buye a la imprecisión de los resultados. Enla figura 3 se observa que el valor genéticomedio de los sementales en los años 90 al93 era próximo a cero y se produjo un creci-miento continuo en años posteriores, locual debe ser atribuido a las madres dedichos sementales, como se ve más adelan-te. En la actualidad el criterio para elegirlos candidatos a futuros sementales es exi-gir un alto índice de pedigrí. Si el padre esde monta natural se exige que la abuela

J.J. Jurado et al. ITEA (2005), Vol. 102 (1), 41-54 49
paterna pertenezca al 5% superior de lashembras.
Vía madres de sementales (MS)
En la figura 5 se presentan las medias gené-ticas anuales de los ovejas disponibles y usa-das y que fueron madres de sementales. Losrespectivos valores de las tendencias son1,683±0,425 y 1,652±0,387 y sus valores
medios de +25,87 y +25,92. Está claro que,así como los ganaderos no pudieron aportarsementales con padres de calidad, si pudie-ron hacerlo con sus madres, (en la mayorparte de los casos la genealogía maternaera conocida y se tenía más información delas mismas). Esto explica que en el CIA setestaran a la postre, machos con un valorgenético medio no muy bajo. En aquellosaños iniciales la vía “padre de sementales”no fue bien utilizada.
Figura 5. Tendencia genética de la raza Manchega. Vía madres de sementales. Medias genéticas poraño de nacimiento de las madres de los sementales usados disponibles (ponderadas y no ponderadas
por el número de hijos).Figure 5. Genetic trend in Manchega breed. Selection path dams of rams. Average estimated bree-
ding values of dams of rams born in each year, actually used and potentially available (weighted andunweighted by number of sons).
Vía padres de ovejas reproductoras (PO)
En la figura 6 figuran las medias genéticasanuales de los machos disponibles y real-mente usados y que son machos que losganaderos utilizan en sus rebaños para lareposición. Los valores de sus tendencias
genéticas son 1,754±0,355 y 0,539±0,593.Aquí se pone de manifiesto que los gana-deros usaron los mejores machos con prefe-rencia a los menos buenos durante los pri-meros años y posteriormente usan todospor igual. La razón puede ser que al princi-pio se pusieron a testar machos de muy

50 J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54
diferentes calidades genéticas, utilizándoseposteriormente solo los declarados mejo-rantes. En años posteriores entraron al tes-taje machos de más calidad (hijos de mejo-rantes) y se declararon mejorantes lamayoría, usándose indistintamente. Debe
tenerse en cuenta que estos machos sontodos de IA y por eso se conoce su genealo-gía y no debe olvidarse que la mayoría delos machos en los rebaños son de montanatural y por consiguiente sus genealogíasy valores genéticos son desconocidos.
Figura 6. Tendencia genética de la raza Manchega. Vía padres de ovejas reproductoras. Media gené-tica por año de nacimiento de los padres de ovejas las ovejas reproductoras usadas y disponibles
(ponderadas y no ponderadas por el número de hijos).Figure 6. Genetic trend in Manchega breed. Selection path sires of ewes. Average estimated bree-
ding values of sires of dams born in each year, actually used and potentially available (weighted andunweighted by the number of sons).
Vía madres de ovejas reproductoras (MO)
En la figura 7 se presentan las medias genéti-cas anuales de las madres de las ovejas deja-das como reposición por los ganaderos tantodisponibles como realmente usadas. Losvalores de las tendencias son 0,898±0,123 y0,751±0,125. Estas gráficas parecen indicarque los ganaderos están usando progresiva-mente hembras de más calidad genética
como madres del recrío, como consecuencia,seguramente, de la utilización de la docu-mentación que el esquema les proporcionasobre sus animales. También se puede dedu-cir que utilizan las hembras de calidad supe-rior con más intensidad que las de calidadinferior
En la tabla 1 se presentan los intervalosgeneracionales estimados para cada una delas cuatros vías.

J.J. Jurado et al. ITEA (2005), Vol. 102 (1), 41-54 51
Figura 7. Tendencia genética de la raza Manchega. Vía madres de ovejas reproductoras. Mediagenética por año de nacimiento de las madres de las ovejas reproductoras usadas y disponibles
(ponderadas y no ponderadas por el número de hijos).Figure 7. Genetic trend in Manchega breed. Selection path dams of ewes. Average estimated bree-ding values of dams of dams born in each year, actually used and potentially available (weighted
and unweighted by the number of sons).
Tabla 1. Intervalo generacional estimado para las cuatro vias de seleccionTable 1. Estimated generational intervales for the four paths of selection
Vía de selección Intervalo generacional
Vía padre de sementales 4,429 añosVía madres de sementales 4,406 añosVía padres de ovejas 3,428 añosVía madres de ovejas 3,639 años
En la tabla 2 se presentan las tendencia gené-tica de cada vía de selección para los animalesdisponible y realmente usados, referidas a:
• Sólo los animales con año de nacimiento(Casos (1) y (2))
• Años de nacimientos faltantes estimadossegún el del hijo más joven. (Casos (3) y (4))
• Años de nacimientos faltantes estimadossegún la media del de los hijos (Casos (5) y(6)
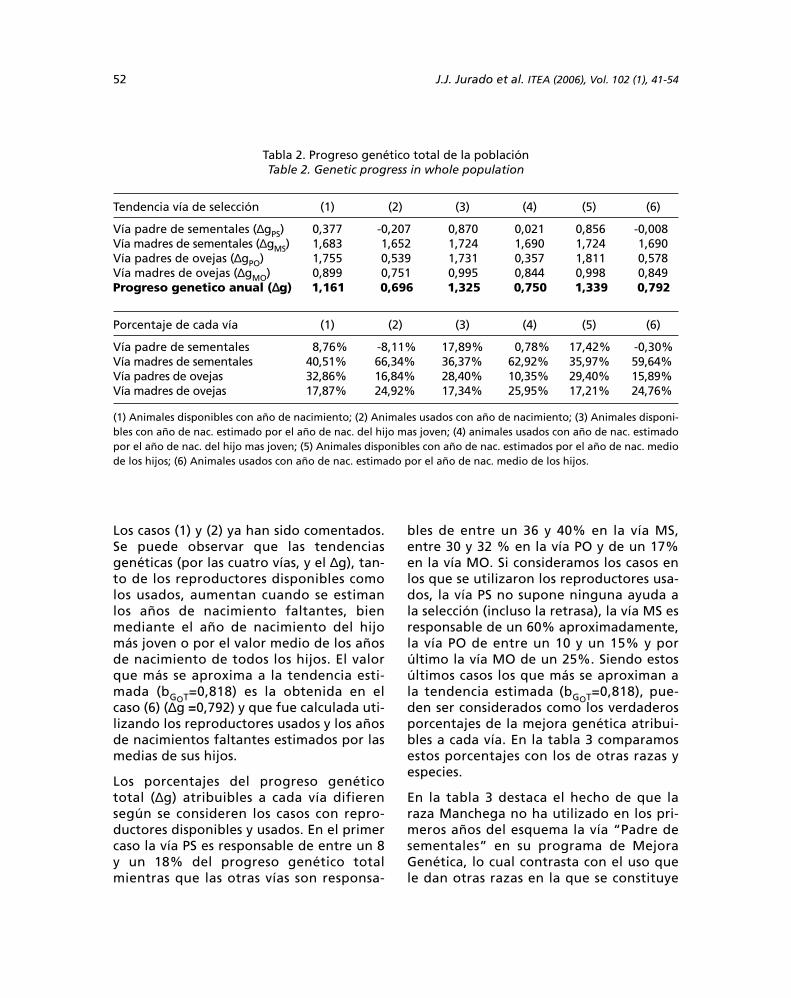
52 J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54
Los casos (1) y (2) ya han sido comentados.Se puede observar que las tendenciasgenéticas (por las cuatro vías, y el ∆g), tan-to de los reproductores disponibles comolos usados, aumentan cuando se estimanlos años de nacimiento faltantes, bienmediante el año de nacimiento del hijomás joven o por el valor medio de los añosde nacimiento de todos los hijos. El valorque más se aproxima a la tendencia esti-mada (bGOT=0,818) es la obtenida en elcaso (6) (∆g =0,792) y que fue calculada uti-lizando los reproductores usados y los añosde nacimientos faltantes estimados por lasmedias de sus hijos.
Los porcentajes del progreso genéticototal (∆g) atribuibles a cada vía difierensegún se consideren los casos con repro-ductores disponibles y usados. En el primercaso la vía PS es responsable de entre un 8y un 18% del progreso genético totalmientras que las otras vías son responsa-
bles de entre un 36 y 40% en la vía MS,entre 30 y 32 % en la vía PO y de un 17%en la vía MO. Si consideramos los casos enlos que se utilizaron los reproductores usa-dos, la vía PS no supone ninguna ayuda ala selección (incluso la retrasa), la vía MS esresponsable de un 60% aproximadamente,la vía PO de entre un 10 y un 15% y porúltimo la vía MO de un 25%. Siendo estosúltimos casos los que más se aproximan ala tendencia estimada (bGOT=0,818), pue-den ser considerados como los verdaderosporcentajes de la mejora genética atribui-bles a cada vía. En la tabla 3 comparamosestos porcentajes con los de otras razas yespecies.
En la tabla 3 destaca el hecho de que laraza Manchega no ha utilizado en los pri-meros años del esquema la vía “Padre desementales” en su programa de MejoraGenética, lo cual contrasta con el uso quele dan otras razas en la que se constituye
Tabla 2. Progreso genético total de la poblaciónTable 2. Genetic progress in whole population
Tendencia vía de selección (1) (2) (3) (4) (5) (6)
Vía padre de sementales (∆gPS) 0,377 -0,207 0,870 0,021 0,856 -0,008Vía madres de sementales (∆gMS) 1,683 1,652 1,724 1,690 1,724 1,690Vía padres de ovejas (∆gPO) 1,755 0,539 1,731 0,357 1,811 0,578Vía madres de ovejas (∆gMO) 0,899 0,751 0,995 0,844 0,998 0,849Progreso genetico anual (∆∆g) 1,161 0,696 1,325 0,750 1,339 0,792
Porcentaje de cada vía (1) (2) (3) (4) (5) (6)
Vía padre de sementales 8,76% -8,11% 17,89% 0,78% 17,42% -0,30%Vía madres de sementales 40,51% 66,34% 36,37% 62,92% 35,97% 59,64%Vía padres de ovejas 32,86% 16,84% 28,40% 10,35% 29,40% 15,89%Vía madres de ovejas 17,87% 24,92% 17,34% 25,95% 17,21% 24,76%
(1) Animales disponibles con año de nacimiento; (2) Animales usados con año de nacimiento; (3) Animales disponi-bles con año de nac. estimado por el año de nac. del hijo mas joven; (4) animales usados con año de nac. estimadopor el año de nac. del hijo mas joven; (5) Animales disponibles con año de nac. estimados por el año de nac. mediode los hijos; (6) Animales usados con año de nac. estimado por el año de nac. medio de los hijos.

J.J. Jurado et al. ITEA (2005), Vol. 102 (1), 41-54 53
como la principal vía de mejora. El valorgenético de los sementales puestos al tes-taje procede exclusivamente de las madresde los sementales. Este es muy elevado(figura 5) y su valor genético medio es de+25,9, lo cual conduce a que los sementalespuestos al testaje en el CERSYRA de Valde-peñas en los años 1999-2000 tengan un
valor genético medio de +18,6. No obstan-te su calidad podría ser superior si lospadres de los sementales tuvieran unmayor valor genético medio. Esta circuns-tancia ya se esta produciendo pues losganaderos están ofertando al CIA machosjóvenes cuyos padres son, en muchos casos,los machos mejorantes allí detectados.
Tabla 3. Importancia relativa de las diferentes vías de progreso genético para varias razas de ovino deleche y del vacuno de leche
Table 3. Relative importance of each selection paths of genetic gain in different dairy species andbreeds
Vías de selección Manchega Latxa* Lacaune** Vacuno lechero***
Vía padre de sementales 0% 40% 31% 43%Vía madres de sementales 59% 39% 39% 33%Vía padres de ovejas (vacas) 16% 11% 17% 18%Vía madres de ovejas (vacas) 25% 11% 13% 6%
*Legarra et al., 2003.**Barillet, F. 1990.***Groen Ab.F. 1990.
La importancia relativa de las vías de mejo-ra en lo que concierne a los ganaderos, osea los padres y madres de ovejas producto-ras, es de un orden semejante al de otrasrazas, aunque predominan las madres sobrelos sementales (seguramente porque estosson en su mayoría de monta natural y devalor genético no contrastado).
Conclusiones
Las conclusiones que se pueden extraer deeste trabajo son las siguientes:
1. El esquema de selección genética de laraza ovina manchega presenta una tenden-cia genética estimada en 0,818 litros/año(0,84% de la media fenotipica y un 5,13%
de la desviación típica aditiva). Es aprecia-ble pero baja en comparación con otrasrazas ovinas lecheras.
2. La mayor parte de esta tendencia es atri-buible a las ovejas con padre conocido, queen su inmensa mayoría son los machosdeclarados mejorantes en el CERSYRA deValdepeñas. La baja tendencia genéticageneral es debida en su mayor parte a losanimales de genealogía desconocida.
3. No obstante lo anterior, la tendenciagenética de las ovejas con padre conocidoes de sólo 1,470 litros/año (1,5% de µP y un9,22% de σA). Es aún baja en relación aotras razas.
4. De las cuatro vías de selección, la de“padre de sementales” presenta valores dela tendencia genética muy bajos o negati-

54 J.J. Jurado et al. ITEA (2006), Vol. 102 (1), 41-54
vos. Esto indica que dicha vía no es utilizadade forma adecuada en el esquema de selec-ción. No obstante los sementales testadosen el CIA presentan valores y tendenciapositivos, debido a la alta calidad de susmadres.
5. Los reproductores utilizados por los gana-deros (machos de monta natural y ovejasreproductoras en los rebaños) para obtenerel recrio son de alto valor genético y contri-buyen al progreso genético de la raza enproporciones parecidas a las de otras razas.
6. En los últimos años los animales testadosen el CIA son de mucha más calidad que enaños precedentes, lo que indica que se estautilizando progresivamente padres desementales de mayor valor genético.
Agradecimiento
Los autores agradecen a J.P. Smulders suayuda para contrastar los resultados de losprogramas informáticos utilizados en estetrabajo.
Bibliografía
Barillet, F, 1990. Amélioration génétique des bre-bis laitières. Seminario avanzado sobre mejorade la calidad de la producción de leche. CIHE-AM-IAMZ, Zaragoza.
Dickerson GE, Hazel LN, 1950. Effectiveness ofselection on progeny performance as a sup-plement to earlier culling in livestock. J. Agric.Res. 69: 459.
Elsen, JM, 1993. Prediction of annual geneticgain and improvement las between popula-tions. Genet Sel Evol. 25, 75: 82.
Groen AbF, 1990. Influences of production circums-tances on the economic revenue of cattle bree-ding programmes, Anim. Prod., 51, 469:480.
Jiménez MA, Jurado JJ, 2005a. La raza Assafespañola en la provincia de León. ITEA. (Acep-tada para publicación).
Jiménez MA, Jurado JJ, 2005b. Esquema de selec-ción de la raza Assaf española. ITEA. Volumenextra, nº 26 Tomo 1, 99: 101.
Jurado JJ, 1994. Evaluación genética de repro-ductores en ganado ovino de raza Manchega.Ganado Ovino. Raza Manchega., 369: 387.Ediciones Mundi-Prensa.
Jurado JJ, Serrano M, 1997. Situación actual de lamejora genética en ovino de leche: Aspectosclave. 241: 254. Ediciones Mundi-Prensa.
Legarra A, Ugarte E, Arrese F, 2003. Análisis delprogreso genético en el esquema de mejorade la raza Latxa. ITEA, vol. 99A, nº 3, 192: 202.
Merino L, Olmedo JA, 1991. Mejora genética delganado ovino de raza Churra. Hojas divulga-doras Num. 20/90 HD. Ministerio de Agricultu-ra Pesca y Alimentación.
Montoso V, Pérez-Guzmán MD, Jurado JJ, Galle-go R, Garcia-Cervigon M, Aguado MJ, GardeJJ, 1996. La selección de la raza Ovina Man-chega. Investigación Agraria en Castilla-LaMancha, 9.
Pérez-Guzman, MD, Serrano M, Montoro V, Jura-do JJ, 1997. Estima de la tendencia genéticade la raza Manchega, incluyendo gruposgenéticos. ITEA, Volumen extra, nº 18, Tomo1, 304: 396.
Rendel JM, Robertson, 1950. Estimation of gene-tic gain in milk yield by selection in a close herof dairy cattle. J. Genet. 50: 1.
Serrano M, Pérez-Guzmán, MD, MontoroV, Jura-do JJ, 1996. Genetic parameters estimationand selection progress for milk yield in Man-chega sheep. Small Rumin. Res. 23, 51-57.
Van Taseel TC, Van Vleck LD, 1990. Estimates ofgenetic selection differentials and generationfor four paths of selección. J. Dairy Sc. 74: 1078-1086.
(Aceptado para publicación el 7 de junio de 2005)

R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66 55
Parámetros cinéticos en eyaculados bovinos de toros deraza frisona y rubia gallega
R. Muiño*, M. Fernández**, A.I. Peña*
* Departamento de Patología Animal. Reproducción y Obstetricia. Facultad de Veterinaria. Universi-dad de Santiago de Compostela. 27002 Lugo.España. [email protected]** Centro de Selección y Reproducción Animal de Galicia.Xenética Fontao, S.A., Fontao-Esperante,Apdo 128. 27080 Lugo (España).
ResumenEn este trabajo se utilizaron 119 eyaculados procedentes de toros de raza Rubia Gallega y Frisona. Lacalidad del semen inicialmente se valoró en base a sus características macroscópicas, motilidad masal,motilidad individual y concentración espermática. Tras la evaluación, los eyaculados calificados como“no aptos para criopreservación” se descartaron, y los eyaculados seleccionados para congelación fue-ron analizados con un sistema CASA (Computer Assisted Sperm Analysis). El objetivo del trabajo fueestablecer valores medios, en semen fresco con calidad adecuada para la criopreservación, para unaserie de parámetros descriptores del movimiento espermático. Los valores obtenidos podrían ser utili-zados como parámetros de referencia por laboratorios que dispongan de un equipo Sperm ClassAnalyzer (software SCA®2002 Movilidad, Microptic S.L., Barcelona, España).
Palabras clave: análisis computerizado, raza, semen fresco.
SummaryMotion characteristics in bovine ejaculates from holstein and rubia gallega bullsFor this study, 119 ejaculates from Holstein and Rubia Gallega bulls were used. The semen quality wasinitially evaluated based on its macrosocopic characteristics, collective motility, individual motility andsperm concentration. After evaluation, the ejaculates scored as “not adequate for cryopreservation”were discarded whereas those selected for cryopreservation were analysed by using a CASA (Comput-er Assisted Sperm Analysis) system. The aim of the present study was to establish mean values for somemotility descriptors observed in bovine fresh semen with optimal quality for cryopreservation. The val-ues obtained in this study could be used as a reference by other institutions using a Sperm Class Ana-lyzer equipment (software SCA®2002 Movilidad, Microptic S.L., Barcelona, España).
Key words: computer analysis, race, fresh semen.
Introducción
La motilidad es sólo uno de los muchosrequisitos que ha de reunir un espermato-zoide para ser capaz de fecundar a un ovoci-to, sin embargo, ha sido y todavía es el pará-metro más utilizado para valorar la calidad
de un eyaculado o de una dosis de semenrefrigerado o congelado. Para que la migra-ción espermática a través del tracto genitalde la hembra ocurra normalmente, y sobretodo para el establecimiento de un reservo-rio espermático en el oviducto, los esperma-tozoides han tener movimiento activo; pero

56 R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66
además, la motilidad es también una mani-festación de integridad estructural y de com-petencia funcional del espermatozoide.
La motilidad espermática normalmente sevalora de forma subjetiva, este es un méto-do simple, rápido y barato. Sin embargo, esaltamente subjetivo, puesto que los resulta-dos obtenidos dependen en gran parte dela habilidad y experiencia del técnico queevalúa la muestra (Rodríguez-Martínez,2000), y además, si se trabaja con muestrasmuy concentradas se tiende a sobrestimar elporcentaje de espermatozoides móviles. Portanto, no es un método que, de forma fia-ble y repetible, permita predecir la capaci-dad fecundante de una muestra de semen(Saacke and White, 1972; Linford et al.,1976). Estos inconvenientes motivaron nu-merosos esfuerzos por parte de los investi-gadores para intentar eliminar la subjetivi-dad inherente al examen microscópico de lamotilidad (Glover, 1968; Katz and Dott,1975; Amann and Hammerstedt, 1980; Katzand Overstreet, 1981; O’Connor et al.,1981), fruto de los cuales se desarrollaronlos sistemas computerizados para el análisisde la motilidad espermática, también cono-cidos como sistemas CASA (Computer Assis-ted Sperm Analysis).
Al final del proceso, el CASA proporcionatoda una serie de datos objetivos de lascélulas móviles presentes en la muestra y lacalidad media de ese movimiento (Amann,1989; Anzar et al., 1991). Pero además, per-mite identificar la existencia de subpobla-ciones de espermatozoides con distintospatrones de movimiento (Davis et al., 1995;Holt, 1996; Abaigar et al., 1999) que coexis-ten en la misma muestra de semen, lo cuáles una visión más real que la motilidadmedia de la muestra, puesto que una mues-tra de semen es una población heterogéneade espermatozoides.
A pesar de su uso generalizado en laborato-rios de investigación y centros de I.A (insemi-nación artificial), no existe una estandariza-ción de los equipos ni de los procedimientosempleados en los análisis (Verstegen et al.,2002). Cada laboratorio utiliza un sistemaCASA diferente, con distintas especificacio-nes técnicas y en diferentes condiciones detrabajo (distintos medios de dilución, tem-peraturas, etc), lo que provoca que los resul-tados facilitados por los distintos centrossean muy dispares, y no sea posible compa-rar directamente los datos de los distintoslaboratorios (Davis et al., 1993; Tardif et al.,1997).
El objetivo del presente estudio, por tanto,fue:
a) establecer unos valores medios para unaserie de parámetros cinéticos obtenidosmediante un sistema CASA (en unas condi-ciones de trabajo definidas) para el semenfresco bovino, a partir de los resultadosobtenidos tras el análisis de eyaculadosbovinos previamente calificados, medianteevaluación subjetiva convencional, como decalidad “buena” o “muy buena” y “aptospara la congelación”.
b) evaluar posibles influencias de la raza y/ode la edad sobre los parámetros descripto-res de la motilidad espermática, mediantela comparación de los resultados obtenidosa partir del análisis de eyaculados de torosde raza Frisona y Rubia Gallega, y en ambasrazas, de toros mayores y menores de 3años.
Material y métodos
Animales
Para este estudio se utilizaron 119 toros, 61de raza Rubia Gallega y 58 de raza Frisona,de edades comprendidas entre 15 meses y

13 años. Todos los sementales se encontra-ban alojados en las instalaciones del Centrode Selección y Reproducción Animal deGalicia XENÉTICA FONTAO, S.A. (Fontao,Esperante, Lugo). Comenzaron su etapacomo reproductores a los 15 meses de edad,estaban clasificados como ‘aptos para lareproducción’ y libres de toda enfermedadinfecto-contagiosa.
Obtención y evaluación de los eyaculados
La extracción del semen se realizó medianteel uso de vagina artificial, y de cada toro seobtuvieron dos eyaculados, con un intervalode unos 20 min entre los dos saltos. Tras laobtención del semen, se realizó una prime-ra valoración de la calidad seminal aten-diendo a las siguientes características:
• color: normalmente entre blanco y amari-llo, dependiendo de la presencia de ß-caro-tenos.
• fluidez: se clasificó en denso, fluido oacuoso. La mayoría de las muestras presen-taban un aspecto denso o fluido. Las deaspecto acuoso se descartaron.
• volumen: determinado por lectura direc-ta en tubos graduados de 10 ml. Se calculóel volumen medio de los dos saltos.
• motilidad en masa: se puntuaron en unaescala de 0 a 5. Los eyaculados con motili-dad masal inferior a 3 se desecharon. Se cal-culó la motilidad media para los dos saltos.
• concentración espermática: determinadapor medio de un espectrofotómetro calibra-do (Photometer SDM 4, Minitüb, Tiefen-bach, Germany). Se calculó la concentraciónmedia para los dos saltos.
Una vez determinados el volumen y la con-centración, se mezcló el semen procedentede ambos saltos y se diluyó con Biociphos(IMV®, L’aigle, France) hasta una concentra-ción de 100 millones/ml. Tras la dilución del
semen se evaluó subjetivamente la motili-dad espermática individual y aquellas mues-tras con motilidad inferior al 60% se des-echaron. Las muestras con un porcentaje deespermatozoides móviles entre el 60 y el80% se consideraron de calidad ‘buena’ y siel porcentaje de motilidad era superior al85% se consideraron de calidad ‘muy bue-na’. Este criterio se utilizó con el fin de nocongelar la mezcla de eyaculados con unamotilidad subjetiva inferior al 60%, portanto, al descartarlos de la congelacióntampoco se analizaron con el CASA.
Evaluación de la motilidad espermática conun sistema CASA
Para el análisis objetivo de la motilidad,tomamos una gota de 5 µl de semen diluidopreviamente y lo diluimos aun más utilizan-do una tasa de dilución 1:5 (1 volumen desemen diluido + 4 volúmenes de Biociphos)con el fin de reducir la concentración hasta20 millones/ml, para facilitar el análisis de lamuestra con el analizador de imagen. Otros5 µl de semen diluidos a esta nueva concen-tración se depositaron entre un porta y uncubre y se evaluaron 3 campos de microsco-pio, seleccionados de la parte superior,media e inferior del portaobjetos, analizan-do aproximadamente un total de 600 esper-matozoides por muestra. Se utilizó unmicroscopio de contraste de fases (NIKON E-600, Tokyo, Japan), con Ph1 positivo, objeti-vo 10 x y platina a 37 ºC. Las imágenes ana-lógicas de los espermatozoides fuerondigitalizadas y analizadas utilizando el soft-ware SCA®2002 Movilidad (Microptic S.L.,Barcelona, España). El patrón de análisis delSCA 2002® predeterminado para la capturade las imágenes incluía los siguientes datos:número de imágenes capturadas por segun-do: 25; velocidad mínima para espermato-zoides lentos: 20 µm/s; velocidad máximapara espermatozoides lentos: 60 µm/s; velo-
R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66 57

cidad máxima para espermatozoides medios110 µm/s; conectividad (distancia máximaentre dos objetos de dos imágenes sucesivaspara que sea considerado el mismo objeto):15 µm; número mínimo de imágenes paracalcular ALH: 5.
Parámetros analizados
– Parámetros de velocidad:
• Velocidad curvilínea (VCL): es la distanciarecorrida por el espermatozoide a lo largode su trayectoria en función del tiempo.
• Velocidad rectilínea (VSL): es la distanciarecorrida por el espermatozoide entre elprimer y el último punto de su trayectoriapor unidad de tiempo.
• Velocidad media (VAP): distancia recorri-da por el espermatozoide a lo largo de sutrayectoria media por unidad de tiempo.
• Índice de linealidad de la trayectoria cur-vilínea (LIN): es la relación porcentual entreVSL y VCL.
• Índice de rectitud (STR): es la relación por-centual entre VSL y VAP.
• Índice de oscilación (WOB): es la relaciónporcentual entre VAP y VCL.
– Parámetros de angularidad y oscilaciónde la cabeza:
• Amplitud media del desplazamiento late-ral de la cabeza (ALH): es el desplazamientoque efectúa la cabeza espermática en sutrayectoria curvilínea de un lado a otro dela trayectoria media o lineal.
• Frecuencia de batido (BCF): es la frecuen-cia con la que la trayectoria curvilínea atra-viesa la trayectoria lineal o media en fun-ción del tiempo.
En función de su VCL los espermatozoidesse clasifican en estáticos (VCL<20 µm/s),móviles no progresivos y móviles progresi-
vos; y dentro de los móviles, en rápidos(VCL>110µm/s), medios (VCL entre 60 y 110µm/s) y lentos (VCL entre 20 y 60 µm/s).
Análisis estadístico
Los datos se analizaron mediante un GLM(Modelo Lineal General) utilizando el pro-grama estadístico SPSS 11.5 (SPSS Inc., Chi-cago, IL, USA), en el que las variablesdependientes (volumen, concentraciónespermática, porcentajes de motilidad yparámetros descriptores del movimiento)se consideraron en función de dos varia-bles independientes: la raza (Rubia Gallegao Frisona) y la edad (animales mayores omenores de 3 años). Las diferencias entrerazas y/o grupos de edad se consideraronsignificativas cuando P<0.05.
Resultados
Las medias (± desviaciones estándar) para elvolumen del eyaculado, la concentraciónespermática, el número total de espermato-zoides y el porcentaje de motilidad subjeti-va se presentan en las figuras 1-4. La raza, laedad o la interacción entre ambas, tuvieronefectos significativos sobre los cuatro pará-metros evaluados. Los toros de raza Frisonamayores de 3 años produjeron mayor volu-men de semen que los frisones jóvenes, yque los toros rubios de ambos grupos deedad. En la raza Rubia Gallega no se obser-varon diferencias en el volumen del eyacu-lado entre sementales mayores y menoresde 3 años. Por el contrario, la concentraciónespermática media fue mayor para la razaRubia que para la Frisona, sin observarsediferencias significativas entre grupos deedad. Mediante valoración subjetiva deleyaculado también se apreció una mayordensidad en los eyaculados de los toros deraza rubia gallega que en los de raza Friso-
58 R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66

na (resultados no mostrados). El númerototal de espermatozoides por eyaculado nodependió de la raza pero sí de la edad, sien-do superior en animales mayores de 3 años.La motilidad espermática, valorada por esti-mación subjetiva, resultó ser mayor en torosrubios que en frisones de más de 3 años,pero entre los jóvenes de ambas razas no seobservaron diferencias (figura 4).
Las medias (± desviaciones estándar) de losparámetros descriptores de la motilidadobtenidos mediante el sistema CASA se pre-sentan en las tablas 1-9. Los porcentajes deespermatozoides móviles, progresivos y noprogresivos (tabla 1), así como los de esper-matozoides rápidos y medios (tabla 2) novariaron con la raza ni con la edad de los
toros, pero se observó mayor (P<0.01) por-centaje de espermatozoides lentos en lostoros frisones que en los rubios (12.67±4.22% vs. 10.38±3.65 %). Con respecto a losparámetros de velocidad (tablas 3-8) o deangularidad y oscilación de la cabeza esper-mática (tabla 9), se observó que la raza ejer-cía un efecto significativo sobre alguno deellos (tabla 10), pero no había diferenciasentre los dos grupos de edad. En general seobservó que en toros frisones la subpobla-ción de espermatozoides clasificados comorápidos describía un movimiento de mayorvelocidad y de trayectoria más rectilínea(mayor VSL, VAP, LIN, WOB y menor ALH)que dicha subpoblación espermática en eya-culados de toros rubios.
R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66 59
Figura 1. Medias (± desviación estándar) del volumen en función de la raza y edad.Figure 1. Mean (± standard deviation) ejaculate volumes as a function of bull breed and age.
Figura 2. Medias (± desviación estándar) de la concentración de espermatozoides en función de la razay edad.
Figure 2. Mean (± standard deviation) sperm concentration in the ejaculates as a function of bullbreed and age.
Figura 1 Figura 2

60 R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66
Tabla 1. Medias (± desviación estándar) para el porcentaje de espermatozoides móviles, progresivos ono progresivos, en función de la raza y de la edad
Table 1. Mean (± standard deviation) percentages of motile spermatozoa, progressive and nonprogressive, as a function of bull breed and age
Raza Edad N Motilidad Estáticos Móviles Móviles no (años) total (%) progresivos progresivos
(%) (%) (%)
Frisona <3 29 Media (±DE) 91.04±2.98 8.96±2.98 58.59±5.64 32.45±4.77>3 29 Media (±DE) 90.79±4.20 9.21±4.20 57.28±4.59 33.51±5.48
Rubia Gallega <3 30 Media (±DE) 89.85±4.14 10.15±4.14 57.60±5.33 32.25±5.24>3 31 Media (±DE) 91.52±4.24 8.48±4.24 59.35±5.47 32.18±5.02
Figura 3. Medias (± desviación estándar) del nº total de espermatozoides en función de la raza yedad.
Figure 3. Mean (± standard deviation) sperm number in the ejaculates as a function of bull breedand age.
Figura 4. Medias (± desviación estándar) de la motilidad subjetiva en función de la raza y edad.Figure 4. Mean (± standard deviation) percentage of subjective sperm motility in the ejaculates as a
function of bull breed and age.
Figura 3 Figura 4

R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66 61
Tabla 3. Medias (± desviación estándar) para la velocidad curvilínea en la población total deespermatozoides analizados (VCL total) o en subpoblaciones de espermatozoides lentos (VCL lentos),
medios (VCL medios) o rápidos (VCL rápidos), en función de la raza y de la edadTable 3. Mean (± standard deviation) values of curvilinear velocity (VCL) for the whole sperm
population (VCL total) and for subpopulations of slow (VCL lentos), medium (VCL medios) and rapidspermatozoa (VCL rápidos) in the ejaculates, as a function of bull breed and age
Raza Edad N VCL total VCL lentos VCL medios VCL rápidos(años) (µm/s) (µm/s) (µm/s) (µm/s)
Frisona <3 29 Media (±DE) 128.89±16.90 38.69±3.26 91.35±5.94 162.20±11.86>3 29 Media (±DE) 126.01±19.95 40.81±2.25 93.51±2.80 161.01±13.05
Rubia Gallega <3 30 Media (±DE) 127.78±13.85 40.02±1.95 94.62±1.79 158.55±9.44>3 31 Media (±DE) 128.34±15.95 39.88±1.94 94.28±2.42 160.76±10.89
Tabla 4. Medias (± desviación estándar) para la velocidad rectilínea en la población total deespermatozoides analizados (VSL total) o en subpoblaciones de espermatozoides lentos (VSL lentos),
medios (VSL medios) o rápidos (VSL rápidos), en función de la raza y de la edadTable 4. Mean (± standard deviation) values of straight line velocity (VSL) for the whole sperm
population (VSL total) and for subpopulations of slow (VSL lentos), medium (VSL medios) and rapidspermatozoa (VSL rápidos) in the ejaculates, as a function of bull breed and age
Raza Edad N VSL total VSL lentos VSL medios VSL rápidos(años) (µm/s) (µm/s) (µm/s) (µm/s)
Frisona <3 29 Media (±DE) 82.14±11.79 17.33±2.99 54.24±6.63 106.69±9.10>3 29 Media (±DE) 78.57±15.06 17.63±2.88 56.59±5.34 102.21±12.86
Rubia Gallega <3 30 Media (±DE) 77.68±11.12 16.77±2.60 55.50±4.19 98.38±10.88>3 31 Media (±DE) 78.46±12.14 16.54±2.81 56.76±5.91 99.23±10.68
Tabla 2. Medias (± desviación estándar) para el porcentaje de espermatozoides rápidos (VCL >110µm/s), medios (VCL entre 60 y 110 µm/s) y lentos (VCL entre 20 y 60 µm/s), en función de la raza y de
la edadTable 2. Mean (± standard deviation) percentages of rapid (VCL>110 µm/s), medium (VCL between 60and 110 µm/s) and slow spermatozoa (VCL between 20 and 60 µm/s) in the ejaculates, as a function of
bull breed and age
Raza Edad N Rápidos Medios Lentos (años) (%) (%) (%)
Frisona <3 29 Media (±DE) 55.91±12.19 23.38±7.79 11.74±4.25>3 29 Media (±DE) 50.89±14.27 27.23±12.65 12.67±4.22
Rubia Gallega <3 30 Media (±DE) 54.98±12.07 24.59±8.64 10.28±2.82>3 31 Media (±DE) 53.37±12.43 27.77±10.43 10.38±3.65

62 R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66
Tabla 5. Medias (± desviación estándar) para la velocidad media en la población total deespermatozoides analizados (VAP total) o en subpoblaciones de espermatozoides lentos (VAP lentos),
medios (VAP medios) o rápidos (VAP rápidos), en función de la raza y de la edadTable 5. Mean (± standard deviation) values of average path velocity (VAP) for the whole sperm
population (VAP total) and for subpopulations of slow (VAP lentos), medium (VAP medios) and rapidspermatozoa (VAP rápidos) in the ejaculates, as a function of bull breed and age
Raza Edad N VAP total VAP lentos VAP medios VAP rápidos(años) (µm/s) (µm/s) (µm/s) (µm/s)
Frisona <3 29 Media (±DE) 100.24±13.82 24.73±3.06 67.99±6.15 128.48±10.03>3 29 Media (±DE) 96.96±17.48 25.40±2.70 70.42±4.21 125.09±13.54
Rubia Gallega <3 30 Media (±DE) 96.24±11.98 24.45±2.24 69.92±3.72 120.72±10.63>3 31 Media (±DE) 97.07±14.08 24.16±2.46 70.38±4.95 122.21±11.74
Tabla 6. Medias (± desviación estándar) para la linearidad en la población total de espermatozoidesanalizados (LIN total) o en subpoblaciones de espermatozoides lentos (LIN lentos), medios (LIN
medios) o rápidos (LIN rápidos), en función de la raza y de la edadTable 6. Mean (± standard deviation) values of linearity (LIN) for the whole sperm population (LIN
total) and for subpopulations of slow (LIN lentos), medium (LIN medios) and rapid spermatozoa (LINrápidos) in the ejaculates, as a function of bull breed and age
Raza Edad N LIN total LIN lentos LIN medios LIN rápidos(años) (%) (%) (%) (%)
Frisona <3 29 Media (±DE) 63.81±4.34 43.50±5.35 59.32±5.59 65.88±4.63>3 29 Media (±DE) 62.18±4.25 43.05±5.16 60.48±4.93 63.38±4.97
Rubia Gallega <3 30 Media (±DE) 60.69±4.39 41.81±5.24 58.66±4.28 61.98±5.02>3 31 Media (±DE) 60.99±3.47 41.38±5.81 60.13±5.26 61.64±3.85
Tabla 7. Medias (± desviación estándar) para el índice de rectitud en la población total deespermatozoides analizados (STR total) o en subpoblaciones de espermatozoides lentos (STR lentos),
medios (STR medios) o rápidos (STR rápidos), en función de la raza y de la edadTable 7. Mean (± standard deviation) values of straightness (STR) for the whole sperm population (STRtotal) and for subpopulations of slow (STR lentos), medium (STR medios) and rapid spermatozoa (STR
rápidos) in the ejaculates, as a function of bull breed and age
Raza Edad N STR total STR lentos STR medios STR rápidos(años) (%) (%) (%) (%)
Frisona <3 29 Media (±DE) 81.94±2.61 69.75±4.98 79.61±3.74 83.05±2.95>3 29 Media (±DE) 80.93±2.78 69.11±4.34 80.21±3.24 81.59±3.30
Rubia Gallega <3 30 Media (±DE) 80.57±2.75 68.28±4.87 79.32±2.54 81.39±3.14>3 31 Media (±DE) 80.75±2.09 68.11±5.28 80.47±3.34 81.14±2.21

Discusión
Los toros de razas lecheras se sabe que, engeneral, producen eyaculados de mayorvolumen que los de razas de aptitud cárnica(Cole y Cupps, 1984). Los resultados del pre-sente estudio claramente confirman estehecho, especialmente en el caso de torosmayores de 3 años. La concentración esper-mática, sin embargo, fue mayor en eyacula-dos de toros rubios que en los de frisones, ypor tanto, el número total de espermato-zoides del eyaculado resultó ser similar paraambas razas. Esto probablemente indica
que la diferencia entre las dos razas en elvolumen de eyaculado se debe más a unadiferencia en la cantidad de plasma semi-nal, emitido por las glándulas sexuales acce-sorias durante la eyaculación, que a unadiferencia en la producción testicular. Dehecho, el número total de espermatozoidespresentes en el eyaculado no se vio influídopor la raza, pero sí por la edad del toro.Aunque en el presente estudio no se evaluóla circunferencia escrotal o el grado de con-sistencia del parénquima testicular, sería deesperar que, en general, los sementales de
R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66 63
Tabla 8. Medias (± desviación estándar) para el índice de oscilación en la población total deespermatozoides analizados (WOB total) o en subpoblaciones de espermatozoides lentos (WOB
lentos), medios (WOB medios) o rápidos (WOB rápidos), en función de la raza y de la edadTable 8. Mean (± standard deviation) values of wobble index (WOB) for the whole sperm population
(WOB total) and for subpopulations of slow (WOB lentos), medium (WOB medios) and rapidspermatozoa (WOB rápidos) in the ejaculates, as a function of bull breed and age
Raza Edad N WOB total WOB lentos WOB medios WOB rápidos(años) (%) (%) (%) (%)
Frisona <3 29 Media (±DE) 77.81±3.38 62.19±4.03 74.38±4.07 79.27±3.51>3 29 Media (±DE) 76.76±2.99 62.12±3.88 75.29±3.46 77.57±3.48
Rubia Gallega <3 30 Media (±DE) 75.26±3.47 61.04±3.73 73.90±3.74 76.08±3.93>3 31 Media (±DE) 75.49±3.01 60.52±4.57 74.60±3.86 75.93±3.49
Tabla 9. Medias (± desviación estándar) para la amplitud media del desplazamiento lateral de lacabeza en la población total de espermatozoides analizados (ALH total) o en subpoblaciones de
espermatozoides lentos (ALH lentos), medios (ALH medios) o rápidos (ALH rápidos), en función de laraza y de la edad
Table 9. Mean (± standard deviation) values of the amplitude of the lateral head displacement (ALH)for the whole sperm population (ALH total) and for subpopulations of slow (ALH lentos), medium (ALH
medios) and rapid spermatozoa (ALH rápidos) in the ejaculates, as a function of bull breed and age
Raza Edad N ALH total ALH medios ALH rápidos BCF total BCF medios BCF rápidos(años) µm µm µm Hz Hz Hz
Frisona <3 29 Media (±DE) 4.10±0.49 3.11±0.30 4.49±0.44 8.85±0.78 7±1.03 9.48±0.91>3 29 Media (±DE) 4.11±0.44 3.09±0.19 4.63±0.32 9.24±0.50 7.37±1.35 9.86±0.57
Rubia Gallega <3 30 Media (±DE) 4.28±0.47 3.18±0.20 4.72±0.42 8.80±0.60 7.20±0.94 9.38±0.64>3 31 Media (±DE) 4.20±0.39 3.15±0.21 4.72±0.34 8.83±0.74 7.32±1.22 9.44±0.76

64 R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66
Tabla 10. Significación (valor de P) del efecto de la raza sobre algunos parámetros descriptores delmovimiento de los espermatozoides
Table 10. Significance (P values) of the breed effect on some of the sperm movement descriptors
Raza Significación
VCL medios 0.003VSL rápidos 0.006VAP rápidos 0.013LIN total 0.005LIN rápidos 0.001WOB total 0.002WOB rápidos 0.000ALH rápidos 0.024
más de 3 años presentasen un mayor des-arrollo testicular que los toros jóvenes,muchos de cuales no habrían completadosu crecimiento corporal ni la maduración desus órganos genitales. El tamaño y el pesodel parénquima testicular son parámetrosaltamente correlacionados con el númerototal de células de Sertoli (Berndtson et al.,1987), con la producción espermática (Coul-ter et al., 1976; Madrid et al., 1988), y con lacalidad del semen (Bailey et al., 1996) y fer-tilidad in vivo del semental (Elmore et al.,1976; Coulter and Foote, 1979).
En este trabajo, la motilidad individualmedia del semen fresco, determinada subje-tivamente, osciló entre el 78 y el 83%, mien-tras que el valor objetivo determinado porel CASA varió entre el 89-91%. Medianteevaluación subjetiva se tendió a subestimarel porcentaje de células móviles aproxima-damente en un 10%, y esto probablementefue debido a que los espermatozoides conmenor velocidad, que el CASA clasifica comolentos, visualmente fueron consideradosinmóviles. Aunque en semen fresco se handescrito variaciones de entre el 30 y el 60%en la estimación subjetiva de la motilidad enfunción de la experiencia del técnico (Bud-worth et al., 1988; Amann, 1989), ésta no hasido una fuente de variación en este estu-
dio, puesto que todas las muestras fueronvaloradas por la misma persona.
El porcentaje de espermatozoides móvilesde un eyaculado o de una dosis de semencongelado está correlacionado con la capa-cidad fecundante de esa muestra, pero nor-malmente explica una fracción muy peque-ña de la variación en la fertilidad de esesemen. Sin embargo, Farrell et al. (1998)observaron que si se incluían colectivamen-te varios parámetros determinados con unsistema CASA, especialmente aquellos quemostraban mayores diferencias entre toros,en una ecuación de regresión múltiple, lacorrelación con la fertilidad del semen per-mitía explicar hasta el 98% de su varianza.
Los parámetros medios descriptores delmovimiento espermático obtenidos en esteestudio con el SCA 2002® para semen frescobovino, son aplicables y repetibles en cual-quier laboratorio de análisis seminal, siem-pre y cuando se utilicen las mismas razas ygrupos de edad, así como el mismo equipode análisis y las mismas condiciones de traba-jo. Los resultados obtenidos tras el análisis delas mismas muestras de semen en distintoslaboratorios y/o con diferentes equipos pue-den ser muy dispares. Estas diferencias sondebidas a que los distintos laboratorios pue-

den utilizar distintos settings (nº de imáge-nes por segundo, límites de tamaño, límitesde velocidad para espermatozoides inmóvi-les, lentos, etc.) y distintas condiciones deanálisis (temperatura, diluyentes, tamaño demuestra), pero fundamentalmente se debena que los distintos equipos a veces utilizanalgoritmos muy diferentes para calcular elmismo parámetro cinético, con lo cuál esmuy probable que en realidad se estén calcu-lando parámetros distintos (Holt et al., 1994).Por otra parte, el usuario es una fuente devariabilidad tan o incluso más importanteque la impuesta por las diferencias técnicasexistentes entre los distintos equipos (Holt etal., 1994).
En definitiva, los valores medios de los pará-metros descriptores del movimiento esper-mático descritos en este estudio se pretendeque sirvan de consulta para los laboratoriosde análisis de semen bovino y los centros deIA siempre que dispongan de las mismasrazas y del mismo sistema CASA utilizado eneste trabajo.
Bibliografía
Abaigar T, Holt WV, Harrison RAP, Del Barrio G,1999. Sperm subpopulations in boar (Sus scro-fa) and gazelle (Gazella dama mhorr) semenas revealed by pattern analysis of computer-assisted motility assessments. Biol. Reprod. 60:32-41.
Amann RP, Hammerstedt RH, 1980. Validation ofa system for computerized measurements ofspermatozoal velocity and percentage ofmotile sperm. Biol. Reprod. 23: 647-656.
Amann RP, 1989. Can the fertility potencial of aseminal sample be predicted accurately? J.Androl. 10: 89-98.
Anzar M, Hassan MM, Graham EF, Deyo RCM,Singh G, 1991. Efficacy of the Hamilton Thornmotility analyzer (HTM-2030) for the evalua-
tion of bovine semen. Theriogenology 36:307-317.
Bailey T, Monke D, Hudson R, Wolfe D, Carson R,Riddell M, 1996. Testicular shape and its rela-tionship to sperm production in mature Hol-stein bulls. Theriogenology 46: 881-887.
Berndtson W, Igboeli G, Pickett B, 1987. Relation-ship of absolute numbers of Sertoli cells totesticular size and spermatogenesis in youngbulls. J. Anim. Sci. 64: 241-246.
Budworth RP, Amann RP, Chapman PL, 1988.Relationships between computerized meas-urements of motion of frozen thawed bullspermatozoa and fertility. J. Androl. 9: 41-54.
Cole HH, Cupps PT, 1984. Reproducción de losAnimales Domésticos. Ed. Acribia, Zaragoza,pp. 221-223.
Coulter G, Rounsaville T, Foote RH, 1976. Heri-tability of testicular size and consistency inHolstein bulls. J. Anim. Sci. 43: 10-11.
Coulter G, Foote R, 1979. Bovine testicular meas-urements as indicators of reproductive per-formance and their relationship to productivetraits in cattle. Theriogenology 11: 297-311.
Davis RO, Katz DF, 1993. Operational standardsfor CASA instruments. J. Androl. 14: 385-394.
Davis RO, Drobris EZ, Overstreet JW, 1995. Appli-cations of multivariate cluster, discriminantfunction and stepwise regression analysis tovariable selection and predictive modeling ofsperm cryosurvival. Fertil. Steril. 63: 1051-1057.
Elmore R, Bierschwal C, Youngquist R, 1976. Scro-tal circumference measurements in 764 beefbulls. Theriogenology 6: 485-494.
Farrell PB, Presicce GA, Brockett CC, Foote RH,1998. Quantification of bull sperm characteris-tics measured by computer-assisted spermanalysis (CASA) and the relation ship to fertili-ty. Theriogenology 49: 871-879.
Glover FA, 1968. Physical method of measuringthe mobility of bull sperm. Nature 219:1263.
Holt WV, Watson PF, Curry M, Holt CH, 1994.Reproducibility of computer-aided semen
R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66 65

analysis: comparison of five different systemsused in a practical workshop. Fertil. Steril. 62(6): 1277-1282.
Holt WV, 1996. Can we predict fertility rates?Making sense of sperm motility. Reprod. Dom.Anim. 31: 17-24.
Katz DF, Dott HM, 1975. Methods of measuringswimming speed of spermatozoa. J. Reprod.Fertil. 45: 263.
Katz DF, Overstreet JW, 1981. Sperm motilityassessment by video-micrography. Fertil. Ster-il. 35: 188.
Linford E, Glover FA, Bishop C, Stewart DL, 1976.The relationship between semen evaluationmethods and fertility in the bull. J. Reprod.Fertil. 47(2): 283-291.
Madrid N, Ott R, Rao D, Parret D, Vanderwert W,Willms C, 1988. Scrotal circumference, seminalcharacteristics and testicular lesions of year-ling Angus bulls. Am. J. Vet. Res. 49: 579-584.
O’Connor MT, Amann RP, Saacke RG, 1981. Com-parisons of computer evaluations of sperma-
tozoal motility with Standard laboratory testsand their use for prediciting fertility. J. Anim.Sci. 53(5): 1368-1376.
Rodríguez-Martínez H, 2000. Evaluación delsemen congelado: Métodos tradicionales y deactualidad. Topics in Bull Fertility. ChenowethPJ (ed.) International Veterinary Informationservice. Ithaca, New York. USA.
Saacke RG, White JM, 1972. Semen quality testsand their relationship to fertility. Proc. 4th
NAAB Tech. Conf. Artif. Insem. Reprod., p. 22.
Tardif AL, Farell PB, Trouern-Trend V, Foote RH,1997. Computer-Assited sperm analysis forassessing initial semen quality and changesduring storage at 5ºC. J. Dairy Sci. 80: 1606-1612
Verstegen J, Iguer-Ouada M, Onclin K, 2002.Computer assisted semen analyzers in androl-ogy research and veterinary practice. Theri-ogenology, 57: 149-179.
(Aceptado para publicación el 30 de agosto de2005)
66 R. Muiño et al. ITEA (2006), Vol. 102 (1), 55-66

PREMIOS DE PRENSA AGRARIA 2006DE LA
ASOCIACIÓN INTERPROFESIONAL PARA EL DESARROLLO AGRARIO
La Asociación Interprofesional para el Desarrollo Agrario (AIDA) acordó enAsamblea General celebrada en mayo de 1983, instaurar un premio anual de Pren-sa Agraria, con el objetivo de hacer destacar aquel artículo de los publicados enITEA que reúna las mejores características técnicas, científicas y de valor divulgati-vo, y que refleje a juicio del jurado, el espíritu fundacional de AIDA de hacer detransmisor de conocimientos hacia el profesional, técnico o empresario agrario.
El día 9 de abril de 1987, la Junta Directiva de AIDA aceptando la propues-ta del Jurado del Premio ITEA 1986 instituyó dos premios; uno para los artículospublicados en la sección de Producción Animal y otro para aquellos que aparezcanen la sección de Producción Vegetal.
Los premios se regirán de acuerdo a las siguientes
BASES
1. Podran concursar todos los artículos que versen sobre cualquier tema técnico-económico-agrario.
2. Los artículos que podrán acceder a los premios serán todos aquellos que sepubliquen en ITEA en el año 2006. Consecuentemente, los originales deberánser enviados de acuerdo con las normas de ITEA y aprobados por su Comité deRedacción.
3. El jurado estará constituido por las siguientes personas:
a) Presidente de AIDA, que presidirá el jurado.
b) Director de la revista ITEA, que actuará de Secretario.
c) Jefe del Servicio de Investigación Agroalimentaria de Zaragoza(Diputación General de Aragón).
d) Director del Instituto Agronómico Mediterráneo de Zaragoza.
e) Director de la Estación Experimental de Aula Dei.
4. Los premios serán anuales y con una dotación de 300 E cada uno.
5. Las deliberaciones del jurado serán secretas, y su fallo inapelable.
6. El fallo del jurado se dará a conocer en la revista ITEA, y la entrega del premiose realizará con motivo de la celebración de las Jornadas de Estudio de AIDA.

*MEJORA GENÉTICA VEGETAL 4 Oct. 04/ Zaragoza IAMZ10 Jun. 05
GESTIÓN DE RIESGOS EN LA AGRICULTURA MEDITERRÁNEA: 22-26 Nov. 04 Zaragoza IAMZ/MAPA-ENESA/SEGUROS AGRARIOS OCDE/AECI
USO DE LOS MARCADORES MOLECULARES EN MEJORA VEGETAL 17-28 Ene. 05 Cabrils IAMZ/IRTA*OLIVICULTURA Y ELAIOTECNIA 26 Sep. 05/ Córdoba UCO/CAP-JA/
31 Mayo 06 CSIC/COI/IAMZDISEÑO Y ANÁLISIS DE SERIES DE ENSAYOS: 12-23 Sep. 05 Zaragoza IAMZ/Generation MÉTODOS CLÁSICOS Y BASADOS EN QTLS Challenge ProgramAGRICULTURA DE REGADÍO SOSTENIBLE EN LA REGIÓN 14-25 Nov. 05 Zaragoza IAMZMEDITERRÁNEA: GESTIÓN DE INSUMOS Y CONTROL DE LA CONTAMINACIÓNHORTICULTURA INTENSIVA Y MEDIO AMBIENTE 16-20 Ene. 06 Zaragoza IAMZCARACTERIZACIÓN DE LA VARIABILIDAD ESPACIAL Y TEMPORAL 7-17 Mar. 06 Zaragoza IAMZ/ICARDAPARA LA PLANIFICACIÓN AGRARIA Y EL MANEJO DE CULTIVOS EN CONDICIONES MEDITERRÁNEASALIMENTOS FUNCIONALES: BASES CIENTÍFICAS Y OPORTUNIDADES 3-7 Abr. 06 Zaragoza IAMZPARA EL SECTOR AGROALIMENTARIO
PLANIFICACIÓN Y DESARROLLO DE CAMPAÑAS DE SANEAMIENTO 13-17 Sep. 04 Zaragoza IAMZ/FAO/OIEGANADEROTRAZABILIDAD DE PRODUCTOS CÁRNICOS: SISTEMAS Y TÉCNICAS 13-17 Dic. 04 Zaragoza IAMZVALORIZACIÓN DE PRODUCTOS LÁCTEOS DE OVINOS 7-18 Feb. 05 Pamplona IAMZ/Univ. Pública Y CAPRINOS EN EL ÁREA MEDITERRÁNEA. TECNOLOGÍAS de NavarraACTUALES Y PERSPECTIVAS DE MERCADOTÉCNICAS MOLECULARES EN MEJORA GENÉTICA ANIMAL 4-8 Abr. 05 León IAMZ/Univ. LeónPRODUCCIÓN ANIMAL Y GESTIÓN DEL MEDIO AMBIENTE 2-7 Mayo 05 Fonte Boa IAMZ/EZN*PRODUCCIÓN ANIMAL 3 Oct. 05/9 Jun. 06 Zaragoza IAMZ
CENTRO INTERNACIONAL DE ALTOS ESTUDIOS AGRONÓMICOS MEDITERRÁNEOSINSTITUTO AGRONÓMICO MEDITERRÁNEO DE ZARAGOZA
CIHEAM/IAMZ - Cursos 2004-05-06
CURSOS FECHAS LUGAR ORGANIZACIÓN
PR
OD
UC
CIÓ
N V
EG
ETA
LPR
OD
UCC
IÓN
AN
IMA
L
NA
TIO
NU
M
AGRARIAUNIV
ER
SITA
S
MEDITERRANENSIUM
CIHEAM
– MARKETING AGROALIMENTARIO: 03-04; 05-06; 07-08
– ECONOMÍA Y GESTIÓN DE LA ACTIVIDAD PESQUERA: 04-05; 06-07; 08-09
– ACUICULTURA: 04-05; 06-07; 08-09
(*) Cursos de Especialización Postuniversitaria del correspondiente Programa Master of Science (*marcados con asterisco en el listado). Se des-arrollan cada dos años:
– MEJORA GENÉTICA VEGETAL: 04-05; 06-07; 08-09
– OLIVICULTURA Y ELAIOTECNIA: 03-04; 05-06; 07-08
– PRODUCCIÓN ANIMAL: 03-04; 05-06; 07-08
– PLANIFICACIÓN INTEGRADA PARA EL DESARROLLO RURAL Y LA GESTIÓN DEL MEDIO AMBIENTE: 04-05; 06-07; 08-09
Se destinan primordialmente a titulados superiores en vías de especialización postuniversitaria. No obstante se estructuran en ciclos independien-tes para facilitar la asistencia de profesionales interesados en aspectos parciales del programa. Los participantes que cumplan los requisitos aca-démicos pueden optar a la realización del 2º año para la obtención del Título Master of Science. El plazo de inscripción para los cursos de MejoraGenética Vegetal, Ordenación Rural en Función del Medio Ambiente y Acuicultura finaliza el 15 de Mayo 2002. El plazo de inscripción para los cur-sos de Mejora genética vegetal, Planificación integrada para el desarrollo rural y la gestión del medio ambiente, Economía y gestión de la activi-dad pesquera y Acuicultura finaliza el 15 de Mayo 2004. El plazo de inscripción para el curso de Olivicultura y elaiotecnia finaliza el 15 de Abril2005. El plazo de inscripción para los cursos de Producción animal y Marketing agroalimentario finaliza el 15 de Mayo 2005.Los cursos de corta duración están orientados preferentemente a investigadores y profesionales relacionados en el desarrollo de sus funcionescon la temática de los distintos cursos. El plazo de inscripción para los cursos de corta duración finaliza 90 días antes de la fecha de inicio del curso.Becas. Los candidatos de países miembros del CIHEAM (Albania, Argelia, Egipto, España, Francia, Grecia, Italia, Líbano, Malta, Marruecos, Portu-gal, Túnez y Turquía) podrán solicitar becas que cubran los derechos de inscripción, así como becas que cubran los gastos de viaje y de estanciadurante el curso. Los candidatos de otros países interesados en disponer de financiación deberán solicitarla directamente a otras institucionesnacionales o internacionales.

PROGRAMAS DE COOPERACIÓN INTERNACIONAL PARA EL CONTROL 27 Sep./1 Oct. 04 Zaragoza IAMZ/MMA-DGB/DE INCENDIOS FORESTALES EN LA REGIÓN MEDITERRÁNEA FAO/AECI*PLANIFICACIÓN INTEGRADA PARA EL DESARROLLO RURAL 4 Oct. 04/ Zaragoza IAMZY LA GESTIÓN DEL MEDIO AMBIENTE 10 Jun. 05ECONOMÍA AMBIENTAL Y DE LOS RECURSOS NATURALES 24 Ene./4 Feb. 05 Zaragoza IAMZRESTAURACIÓN FORESTAL EN LA LUCHA CONTRA LA 19-25 Sep. 05 Zaragoza IAMZ/REACTION/DESERTIFICACIÓN: INNOVACIONES, CONTROL DE CALIDAD Y CE-DGInvestigaciónEVALUACIÓN DE PROYECTOSNUEVAS ESTRATEGIAS PARA LA PLANIFICACIÓN INTEGRADA 12-16 Dic. 05 Zaragoza IAMZDE LOS RECURSOS HÍDRICOS EN LA REGIÓN MEDITERRÁNEAMITIGACIÓN DE LOS EFECTOS DE LA SEQUÍA: MÉTODOS, 12-16 Feb. 06 Aleppo IAMZ/ICARDA/FAOTÉCNICAS Y ALTERNATIVAS DE GESTIÓNDIAGNÓSTICO, SEGUIMIENTO Y GESTIÓN DE LA SALUD 20-25 Feb. 06 Zaragoza IAMZDE LOS BOSQUESBALANCE DE CARBONO EN SISTEMAS AGRARIOS Y FORESTALES: 27-31 Mar. 06 Barcelona IAMZ/CREAFEL POTENCIAL DE REDUCCIÓN DE EMISIONES Y LOS COMPROMISOS DEL PROTOCOLO DE KIOTOLOS INDICADORES AMBIENTALES COMO HERRAMIENTA DE APOYO 8-12 Mayo 06 Zaragoza IAMZA LA GESTIÓN SOSTENIBLE DEL MEDIO RURALSISTEMAS DE AYUDA A LA DECISIÓN PARA EL DESARROLLO 12-16 Jun. 06 Zaragoza IAMZY LA GESTIÓN MEDIOAMBIENTAL DE ZONAS RURALES
LA ORGANIZACIÓN MUNDIAL DEL COMERCIO Y SU IMPACTO E 18-22 Oct. 04 Zaragoza IAMZ/OMCN EL COMERCIO INTERNACIONAL AGROALIMENTARIOASEGURAMIENTO DE LA CALIDAD EN EL MARKETING 7-11 Mar. 05 Zaragoza IAMZAGROALIMENTARIO: NORMATIVAS, SISTEMAS Y REPERCUSIONESEMPRESARIALESTRAZABILIDAD Y ETIQUETADO DE LOS ALIMENTOS: RESPUESTA 14-18 Mar. 05 Zaragoza IAMZA LOS REQUERIMIENTOS DE SEGURIDAD Y DE MERCADOMARKETING DE FRUTAS Y HORTALIZAS EN FRESCO 11-22 Abr. 05 Zaragoza IAMZESTRATEGIAS DE MARKETING EN EL MERCADO INTERNACIONAL 16-20 Mayo 05 Zaragoza IAMZDEL VINO*MARKETING AGROALIMENTARIO 3 Oct. 05/9 Jun. 06 Zaragoza IAMZ
DIAGNÓSTICO Y CONTROL DE ENFERMEDADES DE PECES EN 13-24 Sep. 04 Santiago de IAMZ/Univ. Santiago ACUICULTURA MARINA MEDITERRÁNEA Compostela de Compostela*ECONOMÍA Y GESTIÓN DE LA ACTIVIDAD PESQUERA 27 Sep. 04/ Barcelona Univ. Barcelona/
18 Mar. 05 MAPA/FAO/IAMZ*ACUICULTURA 6 Oct. 04/ Las Palmas de ULPGC/ICCM/IAMZ
8 Abr. 05 Gran CanariaDISEÑO Y GESTIÓN DE CRIADEROS DE MOLUSCOS BIVALVOS 14-19 Feb. 05 Zaragoza IAMZ/FAOESTABLECIMIENTO Y GESTIÓN DE AREAS MARINAS PROTEGIDAS 7-12 Mar. 05 Palma de Mallorca IAMZ/IEODE INTERÉS PESQUEROUSO DE LOS SISTEMAS DE INFORMACIÓN GEOGRÁFICA EN PESCA 13-24 Jun. 05 Zaragoza IAMZ/AECIY ACUICULTURATRAZABILIDAD DE PRODUCTOS DEL MAR: SISTEMAS Y TÉCNICAS 17-21 Oct. 05 Zaragoza IAMZ/FAOMEJORAS TECNOLÓGICAS EN ARTES DE PESCA PARA UNA GESTIÓN 6-10 Feb. 06 Zaragoza IAMZSOSTENIBLEACUICULTURA MEDITERRÁNEA Y GESTIÓN DEL MEDIO AMBIENTE 24-28 Abr. 06 Murcia IAMZ/CAA-CARMCULTIVO DE ALGAS MARINAS: TÉCNICAS, USOS Y PERSPECTIVAS 22-26 Mayo 06 Zaragoza IAMZDE DESARROLLO
CO
MER
CIA
LIZA
CIÓ
NPESC
A Y
AG
RIC
ULT
UR
AM
EDIO
AM
BIEN
TE
CURSOS FECHAS LUGAR ORGANIZACIÓN
Información e inscripción. Los folletos informativos de cada curso se editan 6-8 meses antes de la fecha de inicio. Dichos folletos, así como loscorrespondientes formularios de inscripción pueden solicitarse a la dirección del IAMZ u obtenerse directamente de la página web:
Instituto Agronómico Mediterráneo de Zaragoza
Apartado 202 - 50080 ZARAGOZA (ESPAÑA)Teléfono +34 976 716000 - Fax +34 976 716001 - e-mail [email protected]
www.iamz.ciheam.org

INFORMACIÓN PARA AUTORES
De acuerdo con los fines de la Asociación Interprofesional para el Desarrollo Agrario (AIDA) ITEA publica artículos que haganreferencia a los distintos aspectos de las ciencias agro-alimentarias, cuyo contenido sean resultados obtenidos, descripción demétodos, técnicas o materiales, presentación de nuevas ideas, etc. El envío de un artículo para que se considere su publicación enITEA implicará que el mismo no haya sido publicado o enviado para publicar en cualquier otro medio de difusión y ha de indicar-se expresamente en la carta que acompaña al artículo original. Los derechos sobre todos los artículos o ilustraciones publicadosserán propiedad de ITEA, que deberá recibir por escrito la cesión, una vez aceptado el articulo. Sólo los resúmenes de los mismospodrán ser reimpresos.
Se enviarán tres copias en una versión reciente de Word para Windows. La extensión máxima será de 25 folios de texto mecano-grafiado a doble espacio, tablas y figuras incluidos. Los artículos que superen dicha extensión serán considerados sólo excepcio-nalmente. Los autores deberán aceptar cualquier modificación editorial en la presentación de los artículos. Los artículos se remiti-rán a evaluadores expertos en el tema y el autor recibirá las indicaciones que deberán ser atendidas. En caso de desacuerdo, elautor deberá justificarlo debidamente por escrito. Una vez realizadas las correcciones, el autor enviará un sólo ejemplar mecano-grafiado y una copia en disquete o por correo electrónico. Terminado el proceso de evaluación, se enviará una carta de aceptaciónal remitente. Los autores recibirán las primeras pruebas de impresión que deberán ser revisadas y devueltas rápidamente a laRedacción. El retraso en el retorno de las pruebas determinará que el artículo sea publicado con las correcciones del Editor.
La publicación de un artículo de ITEA no implica responsabilidad o acuerdo de ésta con lo expuesto, significando solamenteque el Comité de Redacción lo considera de suficiente interés para ser publicado. Una vez publicado, los autores recibirán 25separatas del artículo. El autor podrá encargar separatas adicionales, en cuyo caso se le informará previamente del coste quecorrerá por cuenta del propio autor. Los originales serán mecanografiados en castellano y enviados a:
Sr. Director de la Revista ITEAApartado 202
50080 ZARAGOZA
Los originales deberán mecanografiarse a doble espacio en hojas de tamaño DIN A4, dejando márgenes amplios. Cada pági-na estará numerada, utilizándose hojas separadas para las referencias bibliográficas, las tablas, las figuras y los pies de figuras.
Al principio de cada trabajo se indicará:
• Título. El título no deberá incluir abreviaturas ni fórmulas químicas (excepción de los símbolos químicos para indicar isóto-pos), y se procurará que sea lo más breve posible.
• Apellido del autor o autores, precedidos de las iniciales del nombre.• Dirección postal, indicando el autor a quien se va a dirigir la correspondencia, el teléfono y su dirección de correo electrónico.
En la redacción de los trabajos se deberá prestar atención especial al estilo, puntuación, disposición lógica y jerárquica de títu-los y subtítulos y al empleo de abreviaturas, las cuáles no deberán utilizarse en el resumen; en el texto deberán aparecer juntasla abreviatura y el nombre completo la primera vez que aparezcan. Se aplicará el Sistema Internacional de Unidades
Dada la diversidad de las colaboraciones que pueden interesar a los lectores de ITEA, se mantendrá un criterio amplio encuanto se refiere al plan de redacción de los mismos. En general se procurará mantener una disposición lógica, considerandocuidadosamente la jerarquía de títulos, subtítulos y apartados. Se evitará la separación innecesaria de parte del texto en formade anexos. Todas las colaboraciones constarán en primer lugar de un resumen corto de unas 200 a 250 palabras. Dicho resumendeberá ser comprensible por sí mismo y sin referencias al artículo. Tras el resumen se incluirán las palabras clave, separadas porcomas, que sin exceder el número de seis podrán incluir el nombre científico y vulgar de las especies objeto de estudio y la téc-nica relevante utilizada. En lo posible se evitará incluir palabras que ya figuren en el título y en el resumen, tratando de utilizarsinónimos y palabras derivadas. Además, se adjuntará la traducción al inglés del título, del resumen y de las palabras clave.
En el caso de artículos experimentales, se seguirán los siguientes apartados: Introducción, Materiales y Métodos, Resultados yDiscusión. La introducción deberá explicar la finalidad del artículo. El tema se expondrá de la manera más concisa posible, utili-zando un vocabulario preciso y directo. Deberá asimismo, quedar claro qué partes del artículo representan contribuciones propiasy cuáles corresponden a aportaciones de otros trabajos. Se podrá incluir un apartado de Agradecimientos antes de la Bibliografía.
Los trabajos experimentales de extensión reducida, podrán publicarse como Nota Técnica. No excederán de 2.000 palabras,incluidos tablas o ilustraciones y deberán incluir la traducción al inglés del Título, Resumen y Palabras Clave.
Además:
• los nombres latinos de géneros, especies y nombres de variedades deben ponerse en cursiva. Los nombres de cultivaresentre comillas simples (p.ej., ‘Sugar Baby’)

• los nombres de los productos químicos deben comenzar con minúsculas (p. ej., progesterona en vez de Progesterona; sulfa-to de metilo en vez de Sulfato de Metilo).
• los nombres y fórmulas químicas se nombrarán según las normas IUPAC (p. ej., H2SO4 en vez de SO4H2) y los comerciales seescribirán con la inicial en mayúscula (p.ej., Foligón, Sincrozoo, etc.,);
• las llamadas en nota a pie de página o cuadro, que deberán ser las menos posibles y, en todo caso, hechas mediante núme-ros correlativos entre paréntesis; ejemplo (1), (2), evitando el uso de asteriscos, letras o cualquier otro signo
• el nivel de significación estadística, * = p<0,05; ** = p<0,01; ***= p<0,00l; NS = no significativo, no necesitan explicación.
Tablas y Figuras
• El número de tablas y figuras será reducido al mínimo necesario, y los datos no deben ser presentados al mismo tiempo enforma de tabla y de figura.
• Las tablas, dibujos, gráficos, mapas y fotografías deben acompañarse separadamente al texto, aunque señalando su empla-zamiento aproximado. Los dibujos, gráficos, mapas y fotografías deben incluirse como figuras. Las tablas y figuras debenllevar numeración diferente y estar citadas en el texto.
• Los encabezamientos de tablas y los pies de figura deben redactarse de modo que el sentido de la ilustración pueda com-prenderse sin necesidad de acudir al texto. Además, se deberá incluir su traducción al inglés. Los gráficos, dibujos y mapasserán presentados con la mayor calidad posible y en blanco y negro excepto en casos excepcionales que deberán ser acep-tados por la revista.
Bibliografía
Sólo deberán citarse aquellas referencias relacionadas con el trabajo o que contribuyan a la comprensión del texto. Los auto-res deben evitar las relaciones exhaustivas de referencias, salvo en el caso de revisiones bibliográficas.
Las referencias en el texto deben hacerse siguiendo los siguientes ejemplos: (Padilla, 1974), (Vallace y Raleigh, 1967), (Verga-ra et al., 1994). Los nombres de entidades u organismos que figuren como autores, por ejemplo: Dirección General de la Pro-ducción Agraria (DGPA), deberán citarse completos en la Bibliografía la primera vez. Las referencias múltiples se harán segúnse indica en el siguiente ejemplo: (Martinez et al., 1971 y 1979; Ayala y Ortega, 1977). Cuando de un mismo autor o autoresexista más de una referencia del mismo año de publicación, se distinguirán unas de otras añadiendo una letra del alfabeto, enminúscula y comenzando por a, al año de publicación (Lorenzo, 1966a; Aparisi, 1970a y b). Si las citas deben hacerse formandoparte de la frase en el texto, sólo el año de publicación figurará entre paréntesis: ... como indicaban Gómez et al. (1969) ...
Al final del trabajo y precedida del apartado Bibliografía se hará constar una lista de todas las referencias utilizadas en eltexto. Esta lista deberá ordenarse alfabéticamente según el nombre del primer autor de cada trabajo referido. En el caso deincluir varios trabajos del mismo autor, éstos se ordenarán por orden cronológico. En el caso de menciones como "Comunica-ción personal", "Resultados no publicados", éstas no se harán constar en la Bibliografía.
Ejemplos:
Artículos en revistas:
Demarquilly C, 1969. Valeur alimentaire du maïs fourrage, I.Composition chimique el digestibilité du maïs sur pied. Ann. Zoo-tech. 18: 17-32.
Herrero J, Tabuenca MC, 1966. Épocas de floración de variedades de hueso y pepita. An. Aula Dei 8: 154-167.
Tabuenca MC, 1968a. Necesidades de frío invernal de variedades de peral. An. Aula Dei 9: 1-9.
Tabuenca MC, 1968b. Necesidades de frío invernal de variedades de albaricoquero. An. Aula Dei, 9: 10-24.
Vallace JD, Raleigh RJ, 1967. Protein intake and exercise for pregnant heifers. J. Anim. Sci. 26: 931-936.
Vergara H, Gallego L, Molina A, Torres A, 1994. Efecto del nivel de reservas corporales de las ovejas en el parto y del tipo decría sobre el crecimiento de corderos de raza manchega. ITEA 90: 73-80.
Comunicaciones de Congresos:
Angel I, 1972. The use of fasciculate form (determinate habit) in the breeding of new Hungarian pepper varieties. ThirdEucarpia Meeting on Genetics and Breeding of Capsicum, 17-24, Universidad de Turín (Italia).
Capítulos de libros:
Gamborg OL, 1984. Plant cell cultures: nutrition and media, pp. 18-26. En: IK Vasil (Ed.). Cell Culture and Somatic Cell Geneticsof Plants. Vol. 1, 825 pp. Academic Press, USA
Para obtener información adicional envíe un mensaje a: [email protected]


* Si desea Ud. pertenecer a la Asociación, rellene la ficha de inscripción así como la carta para ladomiciliación del pago de la cuota de asociado y envíelas a AIDA. Aptdo. 727. 50080 Zaragoza.
El abajo firmante solicita su inscripción como miembro de la Asociación Interprofesionalpara el Desarrollo Agrario.
Apellidos................................................................................... Nombre.............................................
Dirección postal ...................................................................................................................................
Teléfono ...............................................................................................................................................
Profesión......................................... Empresa de trabajo....................................................................
Área en que desarrolla su actividad profesional ...............................................................................
CUOTA ANUAL: Firma.
❏ ITEA 30 €
FORMA DE PAGO:❏ Cargo a cuenta corriente o libreta ❏ Cargo a tarjeta ❏ Cheque bancario ❏ VISATarjeta número: ❏ MASTERCARD� � � � � � � � � � � � � � � � Fecha de caducidad: /
INSCRIPCIÓN EN AIDA
SR. DIRECTOR DE..................................................................................................................................
Muy Sr. mío:
Ruego a Vd. se sirva adeudar en la cuenta cte./libreta n.º ..........................................................que matengo en esa oficina, el recibo anual que será presentado por la “AsociaciónInterprofesional para el Desarrollo Agrario”.
Atentamente,
Firmado:
BANCO O CAJA DE AHORROS: ...........................................................................................................
SUCURSAL: ...........................................................................................................................................
DIRECCIÓN CALLE/PLAZA: ...................................................................................... N.º ......................
CÓDIGO POSTAL: .......................................................................................................
POBLACIÓN: ...............................................................................................................


