studies on the respiration of sea-urchin spermatozoa
TRANSCRIPT

^ Exp. Biol. (1963), 40, 573-586
ith 6 text-figures
Printed in Great Britain
STUDIES ON THE RESPIRATION OFSEA-URCHIN SPERMATOZOA
V. THE EFFECT OF P0Oa
BY H. MOHRI*
Stazione Zoologica, Naples, Italy
AND I. YASUMASU
Biological Institute, College of General Education, University of Tokyo,Komaba, Meguroku, Tokyo, Japan
(Received 11 March 1963)
INTRODUCTION
It has long been known that sea-urchin spermatozoa remain immotile in the testisor semen and are activated only following shedding into sea water or dilution withsea water. Up to the present, many efforts have been made to clarify the nature andthe causes of such dilution phenomena or of the initiation of movement and the follow-ing factors have been reported to be responsible, although some of them shouldbe excluded in view of present knowledge (cf. Rothschild, 1951, 1956a; Bishop,1962):
(1) CO2 narcosis. Cohn (1918) considered the narcotic effect of COa, low O2 con-centration and low pH as the environmental conditions responsible for the lack ofmovement in semen of Arbacia punctulata, and that dilution with sea water reversedthese conditions with consequent activation of the spermatozoa. According to Gray(1928), however, the explanation in terms of CO2 'is only partially correct at the best,and quite erroneous' (p. 342), because dilute sperm suspensions of Echinus esculentusand Psammechinus miliaris respire at a higher rate, per unit number of spermatozoa,than dense ones, when the O2 uptake is measured by the Warburg direct methodwhich necessitates the continuous removal of respiratory CO2. Gray's finding hasrepeatedly been confirmed by many other workers with several sea-urchin species(Rothschild, 19486,1950, 1956a; Rothschild & Tuft, 1950; Rothschild & Tyler, 1954;Barron, Gasvoda & Flood, 1949; Mohri, 1956a), and has been called Dilution Effect,Dilution Effect II or Respiratory Dilution Effect. He also reported that dilute sus-pensions consume more oxygen in active life than dense ones (Dilution Effect I).Since then, CO2 narcosis has long been left out of consideration from the above-mentioned reasoning (cf. Rothschild, 19486), although Runnstrom, Tiselius &Lindvall (1945) once suggested that CO2 content of dense suspensions probably was asufficient inhibitor of sperm motility in Strongylocentrotus droebachiensis and Echino-cardium cordatum. Recently, however, the effect of CO2 has again been taken into
* On leave from the Biological Institute, College of General Education, University of Tokyo, Tokyo,Japan, as recipient of a scholarship from the Italian Government (1962-63). The travelling expenseswere granted by the Japanese Government.
37 Exp. Biol. 40, 4

574 H. M O H R I AND I. YASUMASU
consideration in relation to dilution phenomena both by Rothschild (1956a, b)and by Mohri & Horiuchi (1961) from somewhat different standpoints as will beseen below.
(2) O2 tension. Rothschild (1948a) suggested that E. esculentus spermatozoa do notmove in undiluted semen because of the lack of O2. The Oa tension in the semen wasless than one-tenth of that in air, and the spermatozoa were able to move even in undilutedsemen if placed in a gas chamber filled with pure O2. The fact that sea-urchin spermato-zoa are unable to move under anaerobic conditions has been reported by other workers(Harvey, 1930; Barron, 1932). Furthermore, the O2 uptake of dense sperm suspensionswas altered by changes in O2 tension, but that of dilute suspensions was not affected byvarying POa. This was explained by Rothschild (19486) as the inability of a significantfraction of the spermatozoa to respire in dense suspensions through lack of O2. Inthe same paper, he described how CO, a potent inhibitor of cytochrome oxidase, hasa greater inhibitory action on dilute than on concentrated sperm suspensions. Mohri(19566) also observed a similar action of sodium azide, another inhibitor of cytochromeoxidase. In a following paper, Rothschild (1950) showed that Dilution Effect I, theincreased total O2 uptake of dilute suspensions, is observed only when the O2 satura-tion in dense suspensions during manometric experiments is not satisfactory. On theother hand, if this condition is satisfactory, rather dense suspensions consume more O2
during their active life. The problem of O2 saturation, therefore, must always be bornein mind. Since Dilution Effect II or the Respiratory Dilution Effect occurs evenwhen the O2 saturation seems to be satisfactory (Rothschild & Tuft, 1950), some otherfactors than O2 tension must be involved in this effect. At any rate it seems certainthat O2 tension is an important factor in inducing sperm motility after dilution ofsemen with sea water.
(3) Allelostasis. This idea was presented by Gray (1928) in his paper dealing withthe first quantitative analysis of Dilution Effect. He concluded that the activatingeffect of sea water must be mechanical, the spermatozoa becoming surrounded by anincreased free space for movement on dilution. In other words, spermatozoa have anallelostatic effect on the activity of their neighbours in undiluted semen or in densesuspensions. According to Rothschild (personal communication) the effect is ex-plained more precisely as follows. In a dense suspension the velocity fields of differentspermatozoa will interfere with each other so that any particular spermatozoon will bein a medium with a higher viscosity than that of the medium without spermatozoa.The activity of spermatozoa is reduced in media of unusually high viscosity. Asdescribed above, however, spermatozoa can move in undiluted semen provided the O2
supply is sufficient. It is difficult, therefore, to explain the cause of the Dilution Effectmerely in terms of allelostasis, although, when activated by O2, spermatozoa in semendo not move so vigorously as after dilution (Rothschild, 19486).
(4) K ions. A reduction in K ions through dilution was regarded by Schlenk &Kahmann (1938) as playing a significant role in initiating motility in rainbow troutspermatozoa. In fact, the concentration of K ions in seminal plasma of E. esculentusis about four times as high as in sea water (Rothschild, 1948 a). Since, however,dilution with seminal plasma instead of sea water has no adverse effect on spermactivity (Gray, 1928; Hayashi, 1945, 1946; Rothschild, 1948a, b), this factor could beexcluded from the cause of the Dilution Effect in sea-urchin spermatozoa, although it

Respiration of sea-urchin spermatozoa. V 575
has been reported that spermatozoa are completely immotile in isotonic (0-5 M) KClsolution (Moriwaki, 1958).
(5) Androgamone I. This hypothesis was put forward by German workers (Hartmann,Schartau & Wallenfels, 1940). They obtained a factor called Androgamone I bymethanol extraction of dried sperm of Arbacia pustulosa, and ascribed the lack ofmovement in semen and the senescence of spermatozoa after dilution to this sub-stance. They believed that the substance is present in seminal plasma. The existenceof such a substance in seminal plasma obtained after centrifugation of semen wasreported by Southwick (1939) with Echinometra subangularis. The role of Andro-gamone I in sea-urchin semen was soon questioned by Runnstrom et al. (1945),although they proved that this factor is responsible for the lack of movement inundiluted salmon semen. The foregoing fact that no special inhibitory substances arecontained in seminal plasma also denies the proposed role of Androgamone I. Thishypothesis, therefore, should be abandoned in view of the present knowledge.
(6) Trace metals, Cu andZn. Rothschild & Tuft (1950) showed that Dilution Effectoccurs when small amounts of Cu or Zn ions are added to a dense sperm suspension.In dilute suspension, however, there is no Dilution Effect when these ions are added.The effect of these ions is counteracted by adding diethyldithiocarbamate, a chelatingagent. From these facts, they came to the conclusion that Dilution Effect is at leastin part caused by these metals, particularly Cu, in sea water. Mohri (1956a) andUtida & Nanao (1956a) also confirmed the stimulating action of Zn ions on O2
uptake of dense suspensions in Pseudocentrotus depressus and Hemicentrotuspulcherrimus.It has been reported, on the other hand, that both the addition of chelating agents suchas glycine (Tyler & Rothschild, 1951) or versene (Rothschild & Tyler, 1954) and theprevious removal of heavy metals from sea water by shaking with dithizone solution(Utida & Nanao, 1956a) nullify the Dilution Effect. Furthermore, Mizuno (1956) in-dicated that sea-urchin spermatozoa show a tendency to take up Zn ions from thesurrounding medium during incubation.
As a possible mechanism by which trace metals exert their Dilution Effect, Mohri(19566) tentatively suggested that the reaction rate of the cytochrome-cytochromeoxidase system is increased by adding these metals as a result of the masking ofsoluble SH groups which are believed to regulate the oxidation rate of cytochrome c(Barron, Nelson & Ardao, 1948). In fact, the cytochrome-cytochrome oxidase systemattains its full action only in dilute suspensions (Mohri, 19566; cf. Rothschild, 19486).Rothschild (1956a), on the other hand, proposed that trace metals cause uncouplingof oxidation and phosphorylation which he considered as being responsible for theDilution Effect. This assumption is based on the fact that 2,4-dinitrophenol (DNP)stimulates the O2 uptake of dense suspensions more than dilute ones, and that theeffect of DNP is reduced in the presence of versene. Similar results were obtained byMohri (1956a) with DNP and by Nelson (1948) with usnic acid, another uncouplerof oxidation and phosphorylation. It is known that DNP enhances the adenosinetri-phosphate (ATP) dephosphorylating activity in mitochondria, thus bringing abouta reduction in P/O ratio. It is still uncertain whether ATPase in sperm mid-piece,i.e. in mitochondria, of sea-urchin spermatozoa is affected by DNP, although ATP-splitting activity of whole spermatozoa is somewhat accelerated by this agent (Mohri,J958). In connexion with the action of trace metals it is suggestive that ATP-splitting
37-2

576 H. MOHRI AND I. YASUMASU
activity of the sperm' head' (head proper plus mid-piece) is activated by Zn ions (Mohri,1958). On the other hand, extensive studies on the motility-inducing mechanism instarfish spermatozoa have been made by Japanese workers (Fujii, Utida, Mizuno &Nanao, 1955; Mizuno, 1956; Kinoshita, 1956a, b; Utida & Nanao, 1956a, b). Theyshowed that histidine and other chelating agents induce sperm motility in starfish andin some other marine forms and conceived that the release of Zn from spermatozoais responsible for the sperm activity. This assumption, however, needs further factsto support it, and is not applicable at least to the Dilution Effect in sea-urchin sperma-tozoa, where the sperm activity is reduced by chelating agents and raised by tracemetals. A more recent experiment by Rothschild (19566) throws some doubts on therole of trace metals in the Respiratory Dilution Effect, because the effect can still beobserved when the suspending medium is 0*05 M borate-buffered sea water with1 o~3 M versene.
Besides having action on O2 uptake, several amino acids and other chelating agentsproduce a prolongation of the life-span of sea-urchin spermatozoa (Tyler & Atkinson,1950; Tyler & Rothschild, 1951; Tyler, 1953; Rothschild & Tyler, 1954; Mohri,19566). The results are explained by the removal of trace metals, Cu and Zn, in seawater, which are harmful to the spermatozoa, although they induce RespiratoryDilution Effect described above.
(7) pH. The pH of seminal plasma has been reported to be 7-6-7-9 in A. punctulata(Hayashi, 1945) and 7-5 in E. esculentus (Rothschild, 1948a), both being lower thanthat of sea water which is about 8-2. As mentioned above, Cohn (1918) suggested thatlow pH in semen is one of the important factors inhibiting sperm activity. Thispossibility was once excluded because seminal plasma does not have an adverse effecton sperm activity and spermatozoa are motile at pH's lower than 7-3 (Rothschild,1948a). Recently, however, the problem was again taken up by Rothschild (1956a, b),using rather strong buffers such as 0-05 M glycyl-glycine to fix the pH. According tohis results, an increase of 0-26 unit from pH 7-91 to 8-17 is associated with an almosttwofold increase in O2 uptake of E. esculentus spermatozoa. As sea water has onlypoor buffering capacity, it is possible that such a difference in pH between dense anddilute sperm suspensions when sea water is used as dilution medium, is due to thepresence of more CO2 in the dense ones even with constant removal of the gas byKOH. He makes the following statement (1956a, p. 170), 'the Respiratory DilutionEffect previously observed after addition of sea water to dense suspensions... maytherefore have been "artifacts", due to the increased sperm activity at high pH's'.However, pH does also play some role in general dilution phenomena, i.e. activationof spermatozoa after dilution, especially in this species. Unfortunately, his conclusiondoes not hold for all sea-urchin species, because the O2 uptake of several Japanese sea-urchins (P. depressus, H. pulcherrimus and Anthocidaris crassispina) is little affected byvarying the pH from 7-0 to 8-5 under the same experimental conditions as those usedby Rothschild (Mohri & Horiuchi, 1961). We must expect, therefore, the presence ofsome other factors than pH essential for the Dilution Effect.
As described above, there are confusing features in the arguments about the causeof the Dilution Effect, and none of the proposed explanations is conclusive, except O2
saturation. Before going further, it is necessary to define exactly what the term ' Dilu-tion Effect' means, in order to avoid unnecessary confusion in this respect. In the

Respiration of sea-urchin spermatozoa. V 577present paper, Dilution Effect is used for general dilution phenomena, including acti-vation of respiration and motility of sea-urchin spermatozoa following dilution withsea water, and Respiratory Dilution Effect for the increase in O2 uptake per unit numberof spermatozoa, in dilute as compared with dense suspensions, when measured by theWarburg direct method, as defined by Rothschild (19566).
In our previous papers (Mohri, 1959; Mohri & Horiuchi, 1961) it was reported thatthe utilization of endogenous phospholipids during ageing of sea-urchin spermatozoais much more reduced in the absence of KOH than in its presence when this substanceis used to absorb the respiratory CO2. Since most of the O2 uptake in sea-urchinspermatozoa is accounted for by oxidation of endogenous phospholipids (Rothschild& Cleland, 1952; Mohri, 1957), and since no compensatory utilization of endogenouscarbohydrates occurs without absorption of CO2 (Mohri & Horiuchi, 1961), thereduction in phospholipid utilization should be reflected in a fall in O2 uptake. In thepresent experiments, therefore, the effect of CO2 was examined by following the O2
consumption polarographically, in parallel with observation of sperm motility.
MATERIALS AND METHODS
Most experiments were done with the spermatozoa of a Japanese sea-urchin,Anthocidaris crassisptna. Some points were checked with the spermatozoa of otherJapanese sea-urchins, Hemicentrotus pulcherrimus and Pseudocentrotus depressus andof the Mediterranean sea-urchins, Paracentrotus lividus and Sphaerechinus granularis.The semen was obtained by the KCl-injecting method as described by Moriwaki
Platinumelectrode
Gas-
Agar bridge
Fig. 1. Diagram of apparatus for measurement of O2 consumption.
(1958). One ml. of semen thus obtained contains about 5-6 x io10 spermatozoa inJapanese sea-urchins and about 4-5 x io10 spermatozoa in Mediterranean sea-urchins.Sperm suspensions were made by diluting semen with filtered sea water to 1 :100throughout the present experiment, unless otherwise mentioned. Filtered sea waterwas boiled and made up to its original volume with distilled water after cooling.
The O2 consumption of a sperm suspension was followed polarographically with avibrating platinum electrode developed by Chance & Williams (1955). The generalscheme of apparatus is presented in Fig. 1. The platinum electrode was vibrated up

578 H. MOHRI AND I. YASUMASU
and down with an amplitude of about 0-5 mm. at 100 cyc./sec. The electrode is'polarized at — o-6 V. against a calomel reference electrode. For measurements of O2
consumption, 5 ml. of sperm suspension were put into one side of the H-shaped troughof the apparatus and the platinum electrode was immersed in the suspension. Gasmixtures of the desired composition were then introduced with a glass capillary andbubbled through the suspension until the galvanometer reading reached its maximum.About 200 ml. of gas were found to be sufficient for attaining this condition.Readings were taken immediately after the bubbling of the gas mixture was stopped.The trough was covered with a few cover-glasses during the measurement to preventthe diffusion of O2 into air.
Sperm motility was observed under an ordinary microscope. The sample was takenfrom the sperm suspension in the trough to a hollow slide and covered with a cover-glass as quickly as possible with minimum disturbance. The motility was roughlyclassified into five grades: very vigorous movement (+ + + +), vigorous movement(+ + +), moderate movement (+ +), faint movement (+) and motionless (—).Sometimes, the score (+) was also used in those cases where very few spermatozoain the microscopic field were motile.
RESULTS
Effect of CO2 tension on O2 consumption. Fig. 2 shows the effects of differing pro-portions of CO2 in pure O2 on the O2 uptake of sea-urchin spermatozoa as measuredpolarographically by the decrease in Oa tension of the medium. The results wereobtained with a single sperm suspension of A. crassispina. With each gas mixture,readings were taken for 5 min. at 1 min. intervals, and then one gas mixture wasreplaced by another and so on. Each curve is corrected for the slight decrease in O2
tension which occurred by diffusion in the absence of spermatozoa. It is apparentthat a marked inhibition of Oa uptake occurs with increasing tension of CO2. With1% CO2, the O2 consumption rate was little affected, but never exceeded that with100% O2. After severe inhibition with 50% CO2, the O2 uptake of the suspension wascompletely restored to the initial rate by replacing the gas mixture again with 100 % O2.The observed effect of CO2 is not due to the reduction in the initial O2 tension in thesperm suspension caused by the mixture of O2 with CO2, because no reduction in O2
uptake with decreasing initial O2 tension was obtained in a control experiment whereCO2 was replaced by N2, although the rate was somewhat altered with concentrationsof N2 above 50 %. Similar results were obtained with the spermatozoa of other Japanesesea-urchins.
Fig. 3 shows the results obtained by varying PCOa in air. With 100% air, O2 uptakeproceeded at much slower rate compared with that with 100 % O2, as was expected fromthe above result that the reduction of O2 tension below 50 % brought about a slowingdown of the O2 consumption rate. Nevertheless, the inhibitory effect of CO2 wasapparent. In this case, too, the inhibition was completely reversible. The percentageinhibition with any given concentration of CO2 was almost the same in both O2 and air.
Effect of pH on O2 consumption. The increase in PCOa is necessarily accompanied bya reduction in pH, when the suspending medium is sea water. Although it has alreadybeen shown that spermatozoa of Japanese species resist the changes in pH between7-0 and 8-5 (Mohri & Horiuchi, 1961), in contrast with spermatozoa of E. esculentus

Respiration of sea-urchin spermatozoa. V 579
Pvhich are very sensitive to pH (Rothschild, 19566), actual pH determinations withglass electrodes after measurements of O2 uptake in A. crassispina gave the values y ifor 2-0% C02, 6-8 for 5 % C02 and 6-i for 10% C02. These values are a little higherthan the theoretical values for sea water, probably owing to the buffering action of thespermatozoa themselves. The values obtained with P. lividus during the winter season
100
o•5.
50
100
q"5.
50
O2100% O2 99%C O 2 1 %
O 2 97-5%CO2 2-5%
O 2 95%CO2 5%
O 2 90%CO2 10%
O 2 80%CO2 20%
O 2 50%CO2 50%
i i i I
O2 100%
Time (min.)
Fig. z. Effect of varying Poot and Pu, in Oa on O2 consumption of sea-urchin spermatozoa,Anthocidaris crassispina. Dilution, i : ioo. Temp. a8° C.
Time (min.)
Fig. 3. Effect of varying POo, in air on Oa consumption of sea-urchin spermatozoa, Anthocidariscrassispina. Dilution, i : ioo. Temp. 28° C.
were 6-9 for 2 % CO2, 6-4 for 5 % CO2 and 5-9 for 10 % CO2. It is necessary, therefore,to re-examine the influence of pH on O2 uptake, especially on the acid side beyond 7-0.In these experiments, the pH was controlled with 0-05 M histidine-HCl-NaOH buffer,
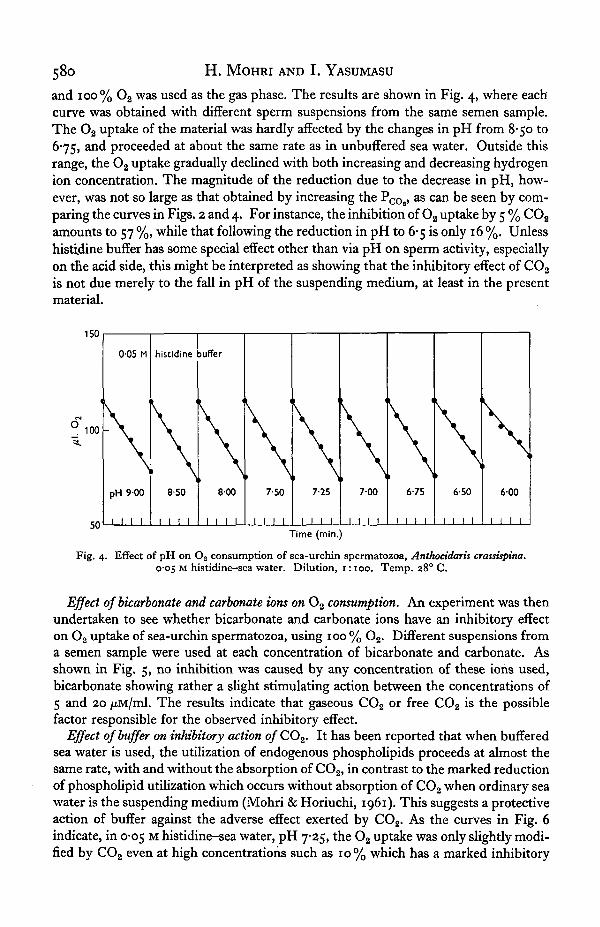
580 H. MOHRI AND I. YASUMASU
and 100% O2 was used as the gas phase. The results are shown in Fig. 4, where eachcurve was obtained with different sperm suspensions from the same semen sample.The O2 uptake of the material was hardly affected by the changes in pH from 8-50 to6-75, and proceeded at about the same rate as in unbuffered sea water. Outside thisrange, the O2 uptake gradually declined with both increasing and decreasing hydrogenion concentration. The magnitude of the reduction due to the decrease in pH, how-ever, was not so large as that obtained by increasing the Pco2>
a s c a n be seen by com-paring the curves in Figs. 2 and 4. For instance, the inhibition of Oa uptake by 5 % CO2
amounts to 57 %, while that following the reduction in pH to 6-5 is only 16 %. Unlesshistidine buffer has some special effect other than via pH on sperm activity, especiallyon the acid side, this might be interpreted as showing that the inhibitory effect of CO2
is not due merely to the fall in pH of the suspending medium, at least in the presentmaterial.
150
100 -
50
005 M
\
pH 900
I I I I
histidine
\
8-50
I I I I
)uffer
\
\
800
1 1 1 1
\
7-50
1 1 1 1
\7-25
1 1 1 1
1 1
V
\700
1 1 1 1
N6-75
1 1 1 1
1 |
\
6-50
1 1 1 1
\
600
1 1 1 1
Time (min.)
Fig. 4. Effect of pH on O2 consumption of sea-urchin spermatozoa, Anthocidaris crassispina.0-05 M histidine-sea water. Dilution, 1:100. Temp. 28° C.
Effect of bicarbonate and carbonate ions on O2 consumption. An experiment was thenundertaken to see whether bicarbonate and carbonate ions have an inhibitory effecton O2 uptake of sea-urchin spermatozoa, using 100 % O2. Different suspensions froma semen sample were used at each concentration of bicarbonate and carbonate. Asshown in Fig. 5, no inhibition was caused by any concentration of these ions used,bicarbonate showing rather a slight stimulating action between the concentrations of5 and 20 /xM/ml. The results indicate that gaseous CO2 or free CO2 is the possiblefactor responsible for the observed inhibitory effect.
Effect of buffer on inhibitory action o/COa. It has been reported that when bufferedsea water is used, the utilization of endogenous phospholipids proceeds at almost thesame rate, with and without the absorption of CO2, in contrast to the marked reductionof phospholipid utilization which occurs without absorption of CO2 when ordinary seawater is the suspending medium (Mohri & Horiuchi, 1961). This suggests a protectiveaction of buffer against the adverse effect exerted by CO2. As the curves in Fig. 6indicate, in 0-05 M histidine-sea water, pH 7*25, the O2 uptake was only slightly modi-fied by CO2 even at high concentrations such as 10% which has a marked inhibitory

Respiration of sea-urchin spermatozoa. V 581
tffect in ordinary sea water. The results again support the view that free CO2 is necessaryfor the inhibitory effect, bound CO2 possessing no such effect.
Effect of CO2 tension on sperm motility. In parallel with the measurements of O2
consumption, sperm motility was examined with increasing CO2 tension. Some resultsobtained with A. crassispina are presented in Table 1. Motility decreased inversely asthe concentration of CO2 and the spermatozoa did not move at CO2 tensions above 8 %,when CO2 was mixed with pure O2. In spite of a complete cessation of movement, asmall amount of O2 is still taken up at concentrations of CO2 above 10 %, as can be seen
100 -
O
"5.
50
100 -
50
Bicarbona
0/(M
\Carbonate
0/iM
i i i i
\
e
2//M
N2/iM
1 I I I
\
\
5/<M
1 1 1 1
\
10/iM
\
WflM
I I I I
\
20/iM
\
20/^M
i i i l
\
50/<M
\
50/* M
i i i iTime (min.) " Time (mirO
Fig. s Fig. 6Fig. 5. Effect of bicarbonate and carbonate ions on O2 consumption of sea-urchin sperma-tozoa, Anthocidaris crassispina. Dilution, i : ioo. Temp. 280 C.Fig. 6. Protecting action of buffer against the inhibitory effect of CO2 on O2 consumption ofsea-urchin spermatozoa, Anthocidaris crassispina. 0-05 M histidine-sea water, pH 7-25. Dilution,1 :ioo. Temp. 280 C.
Table 1. Effect of varying PCo2 in O2 on motility of sea-urchin spermatozoa,Anthocidaris crassispina
(Dilution, 1:100. See text.)
Pco, as % of COa in gas mixture.
Medium o 2
Sea water + + + + + + + +0-05 M histidine- + + +sea water, pH 7-4
5
+ + +
8
±
from the curves in Fig. 2. Motility was regained by introducing 100% O2 after com-plete cessation. In the same table the results of observations with 0-05 M histidine-sea water, pH 7-4, are also listed. In the buffered sea water the motility was suppressed

5^2 H. MOHRI AND I. YASUMASU
a little as compared with that in ordinary sea water. As would be expected from th£results of Oa uptake measurements, however, there was no inhibition of movementeven after exposure to 10 % CO2, the spermatozoa moving rather more vigorously thanin 100% 02.
Table 2. Effect of varying PCOa in 0 2 and in air on motility of sea-urchinspermatozoa, Paracentrotus lividus
(Dilution, i : ioo. See text.)
Poo, as % of CO2 in gas mixture
Gas o 1 a 3 4 5 6 7 8 oOa + + + + + + + + + + + + + +, ++ ++, + ± - + + + +
Air + + + + + + + + , + + - - . . . . + + + +
Table 3. Effect of varying PCOa in 0 2 on motility of sea-urchin spermatozoa,Paracentrotus lividus, at different sperm densities
(See text.)
Poo, as % of CO2 in gas mixture.Sperm .• • vdensity o 2 4 6 8 10 o1:200 + + + + + + + + , + + + + + + ++ ± — + + + +1:100 + + + + + + + + ,+ + + ++ + — . + + + +
1:25 + + + + + + + + + + + , + + - . . + + + +
Tables 2 and 3 summarize the results obtained with the spermatozoa of P. lividus.When the gas phase was O2, the sperm motility was suppressed completely by about8 % CO2) as in the case of A. crassispina. When CO2 was mixed with air, on the otherhand, only 3 % was sufficient to stop the movement. The same was true in spermatozoaof other species. This might be due to the fact that there exists a minimum amount ofO2 uptake necessary for maintenance of motility, and that the O2 uptake is more easilyreduced to this level by increasing the tension of CO2 in air than in pure O2, since theO2 uptake rate is smaller in the former than in the latter, as described above. Anotheralternative is that Oa has a protecting action of some kind against the inhibitory effectexerted by CO2. The effects of PCOs, on motility in sperm suspensions of differentdensities are shown in Table 3. It is apparent that more concentrated suspensions aremore susceptible to the increasing tension of CO2. In fact, with 2 % CO2 in O2 thespermatozoa appeared to move more vigorously in a suspension of 1:25 dilution ascompared with those in a 1:200 suspension, but the movement in the dense suspensionwas abruptly stopped by increasing PC0]i to 6 %, while the spermatozoa still move in thedilute one when the gas phase contains up to 8 % CO2.
DISCUSSION
The experiments reported above clearly show that CO2 inhibits both respiration andmotility of sea-urchin spermatozoa. The results well support our previous assumption,that O2 uptake of the spermatozoa should be much reduced in the absence, as com-

Respiration of sea-urchin spermatozoa. V 5 83
pared with the presence, of KOH used to absorb CO2, when the suspending medium issea water (Mohri & Horiuchi, 1961). If the spermatozoa are allowed to respire in astandard Warburg flask without KOH, much respiratory CO2 accumulates both inthe gas phase and within the spermatozoa. Although the CO2 tensions showing amarked inhibitory effect are rather high, it is conceivable that respiratory CO2 producedintracellularly is more effective than CO2 added extracellularly. In fact, the results ofan experiment tracing the utilization of endogenous phospholipids, which is consideredan index of O2 uptake in sea-urchin spermatozoa, showed that the rate of phospholipidutilization slows down greatly after about 30 min. incubation at 200 C. when therespiratory COa is allowed to accumulate (Mohri, unpublished). The available dataindicate that the inhibitory effect is exerted by gaseous or free CO2, but not by chemi-cally bound CO2. On the other hand, Laser & Rothschild (1939) reported that therespiration rate of sea-urchin eggs when the CO2 is absorbed by KOH is considerablysmaller than that without the absorption. They ascribed this reduction in O2 uptaketo the comparative absence of bicarbonate ions or to the increase in pH accompanyingthe absorption of CO2. The situation is, thus, entirely different for eggs and forspermatozoa. This difference should be due to the difference in metabolic pattern.
It is reasonable to assume that CO2 is also a factor involved in the Dilution Effectof sea-urchin spermatozoa, because the inhibitory effect of CO2 is removed by intro-ducing O2 or air. The facts that the suppressing action of CO2, especially on themotility, can be observed more easily in air than in pure O2, and that the effect is moremarked in dense sperm suspensions than in dilute ones, are also favourable to this view,although the final conclusion must await the direct measurements of CO2 tension insemen and within the cells. It appears, therefore, that CO2 tension and O2 tension(cf. Rothschild, 1948 a, b), and also pH especially in some sea-urchin species such asE. esculentus (cf. Rothschild, 1956 a, b), are the main factors responsible for theDilution Effect, i.e. the activation of spermatozoa after dilution, much as Cohn (1918)considered some forty-five years ago. It is unlikely, on the other hand, that CO2 is alsoresponsible for the Respiratory Dilution Effect, i.e. the increase in O2 uptake per unitnumber of spermatozoa following to dilution of sperm suspensions with constantremoval of respiratory CO2. At least at present, trace metals, Cu and Zn, in sea waterare the most probable factors causing this effect as described in the Introduction,although further investigation is necessary before this hypothesis can be accepted.There remains, however, the possibility that CO2 exerts its inhibitory effect at leasttemporarily within the sperm cells, since in the manometric experiments it is notpossible to obtain a true CO2 vacuum, but only a steady state in spite of the constantremoval by KOH of evolved CO2. Whether CO2 plays some role in senescence or notis obscure at present. Anyway, as correctly pointed out by Rothschild (19486, p. 366),'there is no single factor responsible for senescence, nor for the Dilution Effect'.
The question then arose concerning how and where CO2 exerts its inhibitory effecton sperm activity. In earlier reports, the effect has been called CO2 narcosis. This ismerely a description and not an explanation. Recently, Salisbury and his co-workers(Salisbury, VanDemark, Lodge & Cragle, i960) have shown that a PCOa above about2 % of an atmosphere inhibits glycolysis of bull spermatozoa. The inhibition is alsoreversible by replacement of CO2 by air or by N2. They assumed that the effect of CO2 indepressing glycolysis rests on its influence on intracellular pH, resulting from rapid

584 H. MOHRI AND I. YASUMASU
penetration of C02 across the cell membrane. This is also a possible explanation of theinhibitory effect of CO2 on the respiration of sea-urchin spermatozoa. As regards thesite of the inhibitory action of CO2 in bull spermatozoa, their results suggest that theuptake of hexose is prevented in a medium containing Na as the predominant cation.This, however, does not occur when the medium contains K in place of Na. In thiscase the inhibition seems to occur before the conversion of 1,3-diphosphoglycerate to3-phosphoglycerate. Since sea-urchin spermatozoa in contrast to mammalianspermatozoa have at best only a feeble glycolytic machinery (Rothschild, 1948 a;Mohri, 1957), and since the respiration of sea-urchin spermatozoa proceeds throughthe oxidation of endogenous phospholipids (Rothschild & Cleland, 1952; Mohri, 1957),the process of degradation and oxidation of phospholipids would involve the site ofthe inhibition by CO2. Salisbury (1959) has further shown that sulphide and sulphiteare capable of reversing the inhibition of anaerobic glycolysis by CO2, speculating thatsulphide and sulphite are produced from sulphhydryl compounds by the action ofenzymes such as cysteine desulphhydrase and that they participate in the reactionreleasing CO2 from the sperm cells. It is worth while examining whether these com-pounds are also involved in the mechanism of the Dilution Effect of sea-urchin sperma-tozoa. Preliminary experiments with these compounds, however, have brought aboutno definite results. Some other mechanism, therefore, must be involved in the DilutionEffect. In this connexion, the work by Bendall, Ranson & Walker (i960), whichshowed the inhibition of succinic dehydrogenase by CO2 in a particulate system ofRicinus endosperm, is very suggestive. Experiments along this line are now in progress.
Apart from the problems with spermatozoa, the possible roles of CO2 in manybiological phenomena such as cell differentiation and amoeboid movement haverecently been emphasized by Loomis (1961). According to him, CO2 tension is thefirst self-produced regulator in the process of cell differentiation. In this case, too,gaseous or free CO2 is considered as the responsible factor, owing to its ability to passeasily through the cell membrane, and to its high reactivity. It is necessary, therefore,to pay much attention to this naturally occurring metabolic regulator.
SUMMARY
1. The effect of POOa on the respiration and motility of sea-urchin spermatozoawas studied on Anthocidaris crassispina. Some points were also corroborated onHemicentrotus pulcherrimus, Pseudocentrotus depressus, Paracentrotus lividus andSphaerechinus granularis.
2. It was found that any level of CO2 above 1 %, both in oxygen and in air, inhibitedthe O2 uptake of spermatozoa suspended in sea water, measured polarographicallywith a vibrating platinum electrode. The inhibitory effect paralleled the Pc0|! and wascompletely reversed by introducing oxygen or air.
3. pH variations between 8-50 and 6-75 had no influence on O2 uptake, when thepH was stabilized with 0-05 Mhistidine-HCl-NaOH. O2 uptake was, however, reducedto some extent outside this range, especially on the acid side. Although the increasein PCOa is inevitably followed by a decrease in pH, the inhibitory effect of CO2 farexceeds that caused by the reduction in pH.
4. The O2 uptake rate was little affected by the addition of both bicarbonate and

Respiration of sea-urchin spermatozoa. V 585
carbonate ions to the suspending medium, although the former had a slightlystimulating effect at certain concentrations.
5. In buffered sea water, CO2 had little influence on O2 uptake even at partialpressures as high as 10% which inhibited the bulk of O2 uptake in sea water.
6. Sperm motility was also inhibited by CO2. In this case, too, the inhibitionparalleled the PCOa and was completely reversible. The effect was more pronounced inair than in oxygen, and in dense sperm suspensions than in dilute ones.
7. These results suggest that gaseous CO2 is the factor responsible for the inhibitoryeffect. The possible role of CO2 in the dilution phenomena of sea-urchin spermatozoais discussed.
The authors are much indebted to Prof. J. Ishida and to Lord Rothschild for theirinterest and valuable suggestions. We also thank the Director and Staff of the MisakiMarine Biological Station for supplying sea-urchin materials used in this study. Oneof us (H. M.) is grateful to Dr P. Dohrn for allowing him to use the facilities of theStazione Zoologica, Naples. The technical assistance of Miss T. Abe and Mrs T.Mohri is also acknowledged. Finally, we are indebted to Dr A. Packard for his kindhelp in preparing the manuscript.
REFERENCES
BARRON, E. S. G. (1932). The effect of anaerobiosis on the eggs and sperm of sea-urchin, starfish,and Nereis, and fertilization under anaerobic conditions. Biol. Bull., Woods Hole, 62, 46—53.
BARRON, E. S. G., NELSON, L. & ARDAO, M. I. (1948). Regulatory mechanisms of cellular respiration.II. The r61e of soluble sulfhydryl groups as shown by the effect of sulfhydryl reagents on the respira-tion of sea-urchin sperm. J. Gen. Physiol. 32, 179-90.
BARRON, E. S. G., GASVODA, B. & FLOOD, V. (1949). Studies on the metabolism of action of ionizingradiations. IV. Effect of X-ray irradiation on the respiration of sea-urchin sperm. Biol. Bull.,Woods Hole, 97, 44-50.
BENDALL, D. S., RANSON, S. L. & WALKER, D. A. (i960). Effect of carbon dioxide on the oxidation ofsuccinate and reduced diphosphopyridine nucleotide by Ricinus mitochondria. Biochem. J. 76, 221-5.
BISHOP, D. W. (1962). Sperm motility. Physiol. Rev. 42, 1-59.CHANCE, B. & WILLIAMS, G. R. (1955). Respiratory enzymes in oxidative phosphorylation. I. Kinetics
of oxygen utilization. J. Biol. Chem. 217, 383-93.COHN, E. J. (1918). Studies in the physiology of spermatozoa. Biol. Bull., Woods Hole, 34, 167-218.Fujn, T., UTIDA, S., MIZUNO, T. & NANAO, S. (1955). Effects of amino acids and some chelating
substances on the motility and the oxygen uptake of starfish spermatozoa. J. Fac. Set., Univ. Tokyo,IV, 7, 335-45-
GRAY, J. (1928). The effect of dilution on the activity of spermatozoa. Brit. J. Exp. Biol. 5, 337-44.HARTMANN, M., SCHARTAU, O. & WALLENFELS, K. (1940). Untersuchungen iiber die Befruchtungs-
stoffe der Seeigel. II. Biol. Zbl. 60, 398-423.HARVEY, E. B. (1930). The effect of lack of oxygen on the sperm and unfertilized eggs of Arbacia
punctulata and on fertilization. Biol. Bull., Woods Hole, 58, 288-92.HAYASHI, T. (1945). Dilution medium and survival of the spermatozoa of Arbacia punctulata. I. The
effect of the medium on fertilizing power. Biol. Bull., Woods Hole, 89, 162-79.HAYASHI, T. (1946). Dilution medium and survival of the spermatozoa of Arbacia punctulata. II. The
effect of the medium on respiration. Biol. Bull., Woods Hole, 90, 177—87.KINOSHITA, S. (1956a). Heavy metals in the starfish spermatozoa, Asterias amurensis, with special
reference to zinc. J. Fac. Sci., Univ. Tokyo, iv, 7, 489-96.KINOSHITA, S. (19566). A zinc-containing lipoprotein obtained from the starfish spermatozoa,
Asterina pectinifera. J. Fac. Sci., Univ. Tokyo, iv, 7, 497-503.LASER, H. & ROTHSCHILD, LORD (1939). The metabolism of the eggs of Psammechinus miliaris during
the fertilization reaction. Proc. Roy. Soc. B, 126, 539-56.LOOMIS, W. F. (1961). Cell differentiation: a problem in selective gene activation through self-produced
micro-environmental differences of carbon dioxide tension. In Biological Structure and Function,vol. 11, pp. 509-25. London: Academic Press Inc. Ltd.
MIZUNO, T. (1956). Relation between zinc and sperm motility in some marine forms. J. Fac. Sci.,Univ. Tokyo, iv, 7, 477-87.

586 H . MOHRI AND I . YASUMASU
MOHRI, H. (1956 a). Studies on the respiration of sea-urchin spermatozoa. I. The effect of 2,4-di-nitrophenol and sodium azide. J. Exp. Biol. 33, 73-81.
MOHRI, H. (19566). Studies on the respiration of sea-urchin spermatozoa. II. The cytochrome oxidaseactivity in relation to the dilution effect. J. Exp. Biol. 33, 330-7.
MOHRI, H. (1957). Endogenous substrates of respiration in sea-urchin spermatozoa. J. Fac. Sci.,Univ. Tokyo, iv, 8, 51-63.
MOHRI, H. (1958). Adenosinetriphosphatases of sea-urchin spermatozoa. J. Fac. Sci., Univ. Tokyo,iv, 8, 307-15.
MOHRI, H. (1959). Change in acyl-ester content of lipid fraction in sea-urchin spermatozoa duringageing. Annot. zool.jap. 32, 170-3.
MOHRI, H. & HORIUCHI, K. (1961). Studies on the respiration of sea-urchin spermatozoa. III. Respira-tory quotient. J. Exp. Biol. 38, 249-57.
MORIWAKI, K. (1958). Changes in content of high energy phosphate esters in sea urchin spermatozoaafter dilution. J. Fac. Sci., Univ. Tokyo, iv, 8, 297-305.
NELSON, L. (1948). Usnic acid, an antibiotic, and sperm metabolism. Biol. Bull., Woods Hole, 95,286-7.
ROTHSCHILD, LORD (1948 a). The physiology of sea-urchin spermatozoa. Lack of movement in semen.J. Exp. Biol. 25, 344-52-
ROTHSCHILD, LORD (19486). The physiology of sea-urchin spermatozoa. Senescence and the dilutioneffect. J. Exp. Biol. 25, 353-68.
ROTHSCHILD, LORD & TUFT, P. H. (1950). The physiology of sea-urchin spermatozoa. The dilution effectin relation to copper and zinc. J. Exp. Biol. 27, 59-72.
ROTHSCHILD, LORD (1950). The respiration of sea-urchin spermatozoa. J. Exp. Biol. 27, 420-36.ROTHSCHILD, LORD (1951). Sea-urchin spermatozoa. Biol. Rev. 26, 1-27.ROTHSCHILD, LORD & CLELAND, K. W. (1952). The physiology of sea-urchin spermatozoa. The nature
and location of the endogenous substrate. J. Exp. Biol. 29, 66-71.ROTHSCHILD, LORD & TYLER, A. (1954). The physiology of sea-urchin spermatozoa. Action of versene.
J. Exp. Biol. 31, 252-9.ROTHSCHILD, LORD (1956a). The physiology of sea-urchin spermatozoa. Action of pH, dinitrophenol,
dinitrophenol + versene, and usnic acid on O2 uptake, y. Exp. Biol. 33, 155-73.ROTHSCHILD, LORD (19566). The respiratory dilution effect in sea-urchin spermatozoa. Vie et Milieu, 7,
405-12.RUNNSTR6M, J., TISELIUS, A. & LINDVALL, S. (1945). The action of Androgamone III on the sea-urchin
egg. Ark. Zool. 36A, no. 22, 1-25.SALISBURY, G. W. (1959). Reversal by metabolic regulators of COa-induced inhibition of mammalian
spermatozoa. Proc. Soc. exp. Biol., N.Y., IOI, 187-9.SALISBURY, G. W., VANDEMARK, N. L., LODGB, J. R. & CRAGLE, R. G. (i960). Inhibition of spermatozoan
metabolism by pCO2, pH, K ion and antibacterial compounds. Amer. y. Physiol. 198, 659-64.SCHLENK, W., Jr. & KAHMANN, H. (1938). Die chemische Zusammensetzung des Spermaliquors und
ihre physiologische Bedeutung. Biochem. Z. 295, 283-301.SOUTHWICK, W. E. (1939). Activity-preventing and egg-sea-water neutralizing substances from
spermatozoa of Echinometra subangularis. Biol. Bull., Woods Hole, 90, 177-87.TYLER, A. & ATKINSON, E. (1950). Prolongation of the fertilizing capacity of sea-urchin spermatozoa
by amino acids. Science, 112, 783-5.TYLER, A. & ROTHSCHILD, LORD (1951). Metabolism of sea-urchin spermatozoa and induced anaerobic
motility in solutions of amino acids. Proc. Soc. exp. Biol., N. Y., 76, 52-8.TYLER, A. (1953). Prolongation of life-span of sea-urchin spermatozoa, and improvement of the ferti-
lization-reaction, by treatment of spermatozoa and eggs with metal-chelating agents (amino acids,versene, DEDTC, oxine, cupron). Biol. Bull., Woods Hole, 104, 224-39.
UTIDA, S. & NANAO, S. (1956a). Effect of zinc and 2,4-dinitrophenol on the oxygen uptake of thespermatozoa of sea-urchin and other marine animals, y. Fac. Set., Univ. Tokyo, iv, 7, 505-14.
UTIDA, S. & NANAO, S. (19566). The effects of histidine on the 'free' and 'bound' phospholipid con-tents of starfish spermatozoa, y. Fac. Sci., Univ. Tokyo, iv, 7, 515-18.