studies on the biological properties of polyene antibiotics · vol. 247, ice. 6, issw of...
TRANSCRIPT

THE JOURNAL OF B~LOGICAL CHEMISTRY
Vol. 247, Ice. 6, Issw of &r& 25, pp. 1918-1929, 1972
Printed in U.S.A.
Studies on the Biological Properties of Polyene Antibiotics
EVIDENCE FOR THE DIRECT INTERACTION OF FILIPIN WITH CHOLESTEROL*
(Received for publication, July 7, 1971)
A. W. NORMAN,~ R. A. DEMEL, B. DE KRUYFF, AND L. L. M. VAN DEEPEN
From the Biochemisch Laboratorium, State University of Utrecht, Vondellaan 26, Utrecht, The Netherlands
SUMMARY
The polyene antibiotic lilipin has been shown previously to mediate changes in membrane permeability in natural and artificial biological membranes. The concept has emerged that the interaction of filipin is dependent upon the presence of sterol, preferably cholesterol. The present re- port provides strong evidence that filipin can interact with sterol in a stereochemically and stoichiometrically defined manner to produce a filipin-sterol complex or adduct.
Filipin (F) has a characteristic ultraviolet absorption spec- trum with 4 maxima. The ratio of absorbance of Peak 3, h = 320 nm, to Peak 1, X = 356 mu, is 0.7 in organic sol- vents and 0.8 in aqueous solution. Addition of cholesterol (C) causes a striking spectral change in only the aqueous solutions of filipin so that, although the absorption maxima do not shift, the ratio of absorbance of Peak 3 to Peak 1 be- comes approximately 2.5. The formation of the filipin com- plex, as defined by the spectral change, is specific for stereols. No significant effect on either the formation or breakdown of the filipin-cholesterol complex (F-C) was evident by addition of bovine serum albumin, sucrose, galactose, lecithin, cetyl alcohol, 4 M NaCI, or 4 M urea, or variation of pH from 2 to 9. Addition of the nonionic detergent, Triton X-100, or organic solvents miscible with HzO, broke the F-C complex, whereas heating to 80-90” and cooling caused a reversible breaking and reforming of the F-C complex. The spectral change also occurs when filipin is added to preparations of egg lecithin-cholesterol liposomes (84: 16 to 50:50 mole %). A study of the sterol structural requirements necessary for optimal interaction indicates that with the free steroid in HzO, the interaction is primarily hydrophobic, with the pres- ence of a cholestane ring structure and A22 double bond producing the most favorable interaction. But in the li- posomal system there is an additional requirement for a 3@OH on the steroid nucleus. The same structural require- ments are also necessary for optimal interaction of filipin with mixed lecithin-sterol monolayers.
A quantitative evaluation of the spectral change mediated by cholesterol indicated that the filipin interacted identically
* A preliminary report of a portion of this work has appeared (1).
1 To whom reprint requests should be made. Present address, Department of Biochemistry, University of California, Riverside, California 92502. A. W. Norman was sunnorted during a sabbati- cal leave by National Institutes of Heaiih Special F&owship l- F03 .Anl-13, 654.
with both the free and liposomally bound sterol from lop6 to 10e4 M antibiotic, such that a defined number of cholesterols were combined with 1 filipin molecule.
The spectral change was also evident in red cell ghost membranes and in cells and membranes obtained from Myco- plasma laidlawii grown in the presence of cholesterol. No spectral change was evident in cells or membranes obtained from M. laidlawii grown in the absence of a sterol or in the presence of epi-(3o+hydroxyl)-cholesterol. Analysis of a suspension of synthetic lecithin (L), lecithin-cholesterol (L-C), or L-C-filipin dispersions (L-C-F) by differential scanning calorimetry indicates that addition of C to L (L-C) abolishes the sharp phase transition characteristic of the conversion of lecithin from the crystalline (La) to liquid- crystalline (Lo) form. Addition of F to L (L-F) has no effect on the lecithin phase transition; but addition of F to L-C (L-C-F) causes the phase transition of the lecithin to quanti- tatively reappear.
These results suggest that filipin can interact stoichio- metrically with cholesterol and that in membrane systems this interaction is capable of altering some of the physical properties of the membrane, thus providing a basis for filipin- induced changes in membrane permeability.
The compound filipin belongs to the polyene class of antibiot- ics. To date some 40 polyene antibiotics have been identified. The biological mode of action of these compounds is particularly interesting because these compounds are toxic to fungi and some higher organisms but are without effect on bact.erial systems.
The pentaene, nonahydroxy lactone structure originally l)ro- posed by Cedar and Ryhage (2) for the filipin complex may pos- sibly have to undergo some revision. Bergy and Eble (3) showed that crystalline filipin can be resolved into four components, des- ignated Filipins I, II, III, and IV which constitute 4, 25, 53, and 18yc’,, respectively, of the original material. All four components had the characteristic pentaene chromophore. Filipin I was reported to be a heptahydroxyl, II an octahydroxyl, and III and IV nonahydroxyl compounds (4). The molecular formula and number of hydroxyl groups assigned to Filipins III and IV agree
with the structure originally proposed for the filipin complex (2). Both the conjugated double bond system and the intact macro-
1918
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of ;Llarch 25, 1972 Norman, Demel, de Kruff, and van Deenen 1919
lide ring structure are essential, since either alkaline hydrolysis of the lactone or hydrogenation of the double bonds results in loss of biological activity (5).
Filipin has been shown to mediate permeability changes in erythrocytes (6). On the basis of work by Gottlieb et al. (7-9) and Lampen et al. (10) it was established that the cell membrane was the site of interaction of filipin and that sterol, particularly cholesterol or ergosterol, was an essential membrane constituent for the filipin-induced changes in permeability. The essential requirement for sterol was confirmed by Webster and Kinsky (11) and Feingold (12) using the organism Mycoplasma laidlawii. This organism does not biosynthesize sterols, nor is sterol re- quired for normal development, but it will incorporate cholesterol into its membrane when the sterol is present in the growth media. It was found (11, 12) that only A!Z. Zaidlawii grown in the presence of cholesterol was sensitive in terms of growth to the polyene ant.i- biotic, filipin.
Subsequently the necessary requirement of cholesterol for filipin interaction has been confirmed in a number of other systems. Filipin at < lop5 M is capable of (a) interacting with monolayers of pure sterol or mixed monolayers of sterol and lecithin to cause increases in surface pressure (13, 14) ; (b) interacting with sterol- containing liposomes to cause changes in permeability to several low molecular weight molecules (15) ; (c) lowering the electrical resistance and mediating a selective permeability for Ca++ in bimolecular lipid filmscontaining lecithin and cholesterol (16, 17); (d) mimicking many aspects of dietary vitamin Ds administra- tion on intestinal calcium transport when it is incubated in vitro with intestinal mucosal tissue from vitamin D-deficient chicks (18, 19); and (e) acting as a larvicidal agent in Musca domestica and blocking the uptake of dietary cholesterol in the house fly (20).
In all these diverse systems, although there is a strong implica- tion of a direct interaction between filipin and the sterol to form some type of complex, there is no direct evidence to confirm the existence of such an association. The results reported in this communication establish that filipin interacts directly with free or liposomally bound sterol in a precise stoichiometric and stereo- chemically dependent manner to form a filipin-sterol complex. Evidence is also presented for the existence of this complex in several natural membrane systems.
MATERIALS AKD NETHODS
Chemicals--The filipin used in this report was generously sup- plied by Dr. G. B. Whitfield of the Upjohn Company, Kalamazoo, Michigan. Three lots of filipin have been employed: Lots U-5956, 9098BBC94A, and 8393-DEG-11-8. Each lot was stated by the manufacturer to be approximately 93 to 96% pure on the basis of ultraviolet absorption. Similar results were ob- tained with each filipin preparation.
Stock solutions of filipin were prepared by dissolving 1 to 5 mg in 1.0 ml of dimethyl formamide, which were stored in the dark at -5O. This stock solution was normally used for only 1 week.
Lipids- I- Oleo yl - 2 - stearoyl - sn - glycero - 3 - phosphorycholine (18: l/18:0 lecithin) was synthesized by the method of van Deenen and de Haas (21). Egg lecithin was isolated and puri- fied according to established procedures (22). The purity of both lipids was ascertained by thin layer chromatography em- ploying solvents of chloroform-methanol-water (65: 35 :4, v/v/v).
Steroids-The steroids were obtained from the following sources: ergocalciferol (vitamin Dz) and dihydrotachysterolz
(Phillips-Duphar, Weesp, The Netherlands) ; cholest-5-ene-3/3-ol (cholesterol), cholest-5,7-diene-3P-ol (7-dehydrocholesterol), cholest-5,7,22-triene-24-methyl-3/3-ol (ergosterol), cholert-4-en- 3-one, and 5@-cholestan-S&o1 (cholestanol) (Fluka 4G, Buchs, Switzerland) ; cholest+en-3-one, 5a-cholestan&one, and cholest-5,22-dien-24.ethyl-3P-ol (stigmasterol) (Koch-Light Laboratories, Colnbrook, Buchs, England) ; SD-cholestan-3fl-ol (coprostanol) (K and K Laboratories, Hollywood, California) ; cholest-7-en-3P-ol (lathosterol) and androstane-3P-ol (Ikapharm, Ramat-gan, Israel); cholest-B-nor-5-en3&ol (B-norcholesterol) (Dr. J. Joska, Institute of Organic Chemistry and Biochemistry, Prague, Czechoslovakia) ; cholest-5-en-3a-ol (epicholesterol) and cholesterol-acetate (Mann Biochemicals, New York). The sterols were checked for purity by silica gel thin layer chro- matography using solvents of chloroform-acetone (98:2) and if necessary purified.
Other Chemicals-Bovine serum albumin (BSA) was obtained from Cal Biochemicals, Inc. (Los Angeles, California), and Triton X-100 from Wegman (Amsterdam). All other chemicals and solvents were analytical reagent grade or equivalent.
Preparation of Liposomes-Chloroform solutions of lecithin or sterol (or both) were transferred to a round bottomed flask and the solvent was removed under vacuum. The lipid mixtures were covered with 1 to 5 ml of 10 m&f Tris-acetate, (pH 2 to 9) and either vortexed for 1 min or subjected to ultrasonic radiation (Branson Sonic Power Instrument, model S-125, position 4).
Membrane Preparations---Red cell ghosts were prepared from human blood by a minor modification (23) of the procedure of Parpart (24). They were suspended (1 mg per ml) in 10 mnr Tris-acetate, pH 7.0, and sonicated for 1 min prior to use. Such preparations normally have approximately 0.20 mg of cholesterol per mg of protein.
Mycoplasma Zaidlawii B--Cells were grown in .35 to l.O-liter quantities of lipid-poor media according to the techniques of McElhaney and Tourtellotte (25). Palmitic (16:0) and oleic (18 : 1) acids were added to cell growth media, each at a final con- centration of 0.03 mM, in an appropriate amount of absolute ethanol containing 10 mg of total fatty acid per ml. Cholesterol and epicholesterol were added to individual growth media both b> (a) adding it to the growth media to a final concentrat,ionl of .OG5 mM (25 mg per liter) by the additionof aliquots of ethanol contain- ing the fatty acids and sterol at 10 mg of lipid per ml, and (b) preparing liposomes from sterol-free M. laidlawii polar lipids to which was added the sterol. Step b was accomplished by plac- ing 30 mg of polar lipids plus 15 mg of cholesterol or 30 mg of epicholesterol in 40 ml of complet,e tryptose medium (25) and sonicating 15 min in a Branson Sonic Power Instrument at maxi- mum setting, followed by centrifugation for 15 min at 17,500 rpm in a Sorvall SS-34 rotor. This supernatant was employed as a sterol rich growth medium. All the above steps were carried out under sterile conditions. In preliminary experiments it was found necessary to include the sterols in the growth media in lip- osomal form to ensure that no free sterol would precipitate dur- ing the isolation of the cells.
The cholesterol and epicholesterol content of the various mem- brane preparations was determined by the Lieberman-Burchard technique on an aliquot of the total lipid .extract of the mem-
1 Throughout this report amounts of cholesterol and other ste- roids added to H,O are reported as molar concentrations. This is only an approximation due to the low Hz0 solubility of most ste- roids.
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

F&pin-Cholesterol Interaction Vol. 247, Ko. 6
branes. The cells were grown at 37” and harvested in late log phase by centrifugation in a Sorvall refrigerated centrifuge at 10,000 rpm for 15 min in a GSA rotor. The cell pellet was washed once by resuspension in 0.2 M sucrose followed by sedi- mentation at 10,000 rpm. Membrane preparations of M. Zaid- lawii B were prepared by the procedure of van Golde et al. (26). They were suspended in 10 mM Tris-acetate, pH 7.0, prior to use. The protein determinations were carried out according to the method of Lowry et al. (27).
Xpectrophotometric Measurements-A Perk&Elmer, two wave length, double beam spectrophotometer, model 356, was used for all spectrophotometric analyses. With clear solutions it was utilized in the double beam mode, while with opaque solutions or suspensions of liposomes or membranes it was used in the split beam mode. In this latter instance appropriate base-line cor- rections were applied. The light path of the sample was usually 1.0 cm, but with concentrations of filipin above 1.6 X 10F5 M,
cuvettes with a O.l-cm light path were employed. Differential Scanning Calorimetric Measurements-A Perkin-
Elmer differential scanning calorimeter, DSC-lB, was employed to determine the presence of phase transitions in samples con- taining l-oleoyl-2-stearoyl-sn-glycero-3-phosphatidylcholine.
The samples were prepared by removing in vucuo the organic solvent from solutions of the appropriate amount of lipid. Then to 7 mg of total lipid (lecithin alone; lecithin-cholesterol, 80:20 mole %; and lecithin-cholesterol, SO:20 mole y0 + 1.1 mg of filipin) were added 35 ~1 of HzO-glycol, 1: 1 (v/v). In a separate control experiment the presence of glycol was found to have no effect on the phase transition of the synthetic lecithin. These suspensions were vortexed repeatedly over a period of several hours. Then approximately 20 ~1 of a suspension were sealed in an aluminum sample pan. The sample holder was cooled with liquid nitrogen and flushed continuously with nitrogen. A programmed heating run was made from -15” to approxi- mately 60” on Range 1, at a scan speed of 8” per min. For each sample each heating curve was repeated at least two times to ensure that all transitions observed were reversible. After dissolving the sample pans and contents in 70% perchloric acid for 48 hours, phosphorus was estimated according to the tech- niques of Fiske and Subbarow (28). The area under each phase transition curve was determined using millimeter graph paper. The results are reported as energy units per mg of lecithin.
Measurement of Xurface Pressure-Surface pressure measure- ments were carried out in a paraffin-coated glass trough, 14 cm wide X 58 cm long (total volume = 700 ml) utilizing a conven- tional Langmuir-Adam film balance. All measurements were made at room temperature. The water employed as a subphase had been distilled from alkaline permanganate and redistilled in a quartz still. The monolayers were prepared by spreading either simple sterols or mixtures of lecithin and sterol, 1: 1 molar ratio, on a surface of 560 cm2 from a chloroform solution. This area was then compressed to approximately 140 cm2 to give initial surface pressures between 2 and 30 dynes per cm. Filipin dissolved in dimethyl formamide (100 X) was injected beneath the monolayer with an Agla micrometer syringe. A stable pressure change was obtained within 2 min.
Radioactivity Measurements-The radioactivity of aqueous suspensions of [r4C]cholesterol were determined in a Packard Tri-Carb liquid scintillation counter employing a dioxane count- ing solution containing 0.3 g of dimethyl 1,4-bis[2-(5-phenyl-
oxazolyl)]benzene, 7 g of 2,5-diphenyloxazole, and 100 g of naphthalene per liter of dioxane.
RESULTS
Spectral Properties of Filipin- Fig. 1 compares the ultraviolet absorption spectra of identical concentrations of filipin dissolved in methanol or water. The wave lengths of the absorption maxima of the four peaks are essentially the same for both solvents and are identical with those reported by Whitfield et al. (29) and Lampen et al. (10). The ratio of absorbance of Peak 3 to Peak 1 is normally 0.70 to 0.73 for filipin in methanol and 0.77 to 0.84 for filipin dissolved in HzO. The latter value may increase even further, as the concentration approaches lop4 M. Using a value of 654 for the molecular weight of filipin (2) and correction for the fact that the filipin preparation em- ployed was only 96% pure, it can be determined that the ap- parent molar extinction coefficient of Peak 2 in methanol is 6.2 x 104. All concentrations of filipin reported in this paper were determined by reference to this value for filipin dissolved in methanol. The molar extinction coefficient of Peak 2 of filipin dissolved in dimethyl formamide, dimethyl sulforide, ethanol, 1-butanol, and 1,4-dioxane was within 10% of the value for filipin in methanol.
However, it is clear that in this experiment the extinction coefficient for Peaks 1 through 4 of filipin in HZ0 is only approxi- mately 607, of the respective values for filipin in methanol. Over a large number of experiments, the reduction has been found to vary from 55 to 85%. For any given solution of filipin in Hz0 the spectrum was stable for several hours. Simi- lar results were reported without comment by Lampen et al. (10). The exact amount of diminution of the spectra of equivalent concentrations of filipin in Hz0 as compared to methanol is determined in a complex manner by a number of variables which include: (a) the concentration of filipin; (b) the manner of introduction of filipin dissolved in dimethyl formamide into the water; (c) the length of time of vigorous mixing of the aqueous solution; (d) the temperature; and (e) the age of the stock solu- tion of filipin in dimethyl formamide. It is suspected that a portion of this reduction of ultraviolet absorbance of filipin in water is due to either micelle or aggregate formation.
On the basis of the results depicted in Fig. 2, it seems 1ikelJ that the aqueous insolubility of filipin alone is not an explanation for its reduced ultraviolet absorption. Fig. 2 presents results of an experiment in which both Hz0 and methanol solutions of filipin, each initially 1.60 X 10-d M, were diluted 8 times suc- cessively 1: 1 with HSO or methanol. The total absorbance for Peak 1, in methanol, and Peaks 1 and 4, in HSO, was calculated taking into account the dilution factor. Filipin dissolved in methanol over the range 1 X 10e6 M to 4 X lop5 M closely follows the Beer-Lambert law, whereas neither the absorbance of Peak 1 nor Peak 4 for filipin in Hz0 followed the Beer-Lambert law over the range 1.6 X low4 M to 6 X lo-’ M. But this is not due to insolubility of the filipin in water, since as the concentration of filipin was lowered, presumably increasing its solubility, the absorbance of Peak 4 became less rather than greater, while that of Peak 1 responded in a reciprocal manner. These results obtained for filipin in water emphasize the necessity to determine the concentrations of filipin only on the basis of its absorbance in methanol.
In the course of studies concerning the partition coefficient of filipin in biphasic systems and the possible effect of cholesterol
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of March 25, 1972 Norman, Demel, cle Kru$, ancl van Deenen 1921
H20
400 350 303400 350 300 WAVELENGTH
$10.0 - .-. PEAK l(MeOH) ’
5 9.0- ., 1 --.-.I_.- .-., /
04
o--o PEAK 1 (Hz01 q
x--x PEAK 4 (H20) 1
l-i 2 O=[F] = 1.6~10-~ M
8 =[F] = 6.0~10-~ M
FIG. 1 (lejt). Spectra of filipin in methanol and water. From a stock solution of filipin in dimethyl formamide, 50 ,~l were added separately to 5.0 ml of methanol and HzO. After thorough mixing the ultraviolet absorbance spectrum was measured. The concen- tration of filipin in each sample was 1.23 X 10m5 M. The absorption maxima in methanol (MeOH) for Peaks 1 to 4 were 356,337,321, and 311 nm, respectively. In Tvater the absorption maxima for Peaks 1 to 4 were 356, 338, 320, and 311 nm.
Fro. 2 (right). Effect of dilution on filipin absorbance. Filipin, 1.60 X 10. * M, in Hz0 or methanol was diluted eight times succes- sively (1: 1) with either Hz0 or methanol. After each dilution an ultraviolet absorption spectrum was determined. For samples 0 to 3, the light path in the cuvette was 1 mm; for samples 4 to 8 it was 10 mm. The data are expressed as total absorbance present in Peak 1 (methanol) or Peaks 1 and 4 (HzO) corrected for dilution.
on this partitioning, it became apparent that the ultraviolet absorption spectrum of aqueous solutions of filipin was dramat- ically altered by the presence of cholesterol. Typical re- sults are shown in Fig. 3 where cholesterol was added to an aqueous solution of filipin. The ratio of absorbance of Peak 3 to Peak 1 is increased from approximately 0.8 in the absence of cholesterol to 2.5 in the presence of choles- terol. The extinction coefficient of Peak 4 is also sharply in- creased. A major change in the wave length of ultraviolet absorption for Peaks 1 through 4 is not evident. The extent of increase in the absorbance ratio of Peak 3 to Peak 1 has been noted to vary from 2.0 to 3.5; hoI+-ever with any given aqueous solution of filipin the ratio is quite reproducible. Also the ratio of absorbance in Peak 3 to Peak 4 of the filipin cholesterol com- plex varies from 0.8 to 1.2 from experiment to experiment. As will be discussed in detail below, this change in the spectrum of filipin is likely due to the formation in H20 of a complex or adducb between cholesterol and filipin which has a defined stoi- chiometry.
Also shown in Fig. 3 is the effect on the ultraviolet absorption spectrum of dilution of the aqueous solution of filipin-cholesterol complex to 5Oc/, water-methanol, v/v. Now the filipin exhibits an ultraviolet absorption spectrum both qualitatively and quanti- tatively identical with that of filipin in 100% methanol. The ratio of absorbance of Peak 3 to Peak 1 is approximately 0.8. Identical results have been obtained when the dilution is carried out with ethanol, I-butanol, 1,4-dioxane, and dimethyl for- mamide. Similar observations have also been described earlier by Lampen et al. (10).
Since filipin has been employed previously in studiei of anti- biotic-induced changes in the permeability of cholesterol-con-
0.4
0.3
0.2
0.1
MQOH dil.
400 350 300 400 350 300 WAVELENGTH
FIQ. 3. Spectrum of filipin-cholesterol complex. To 10 ml of a 1.19 X 10e5 M aqueous solution of filipin were added 25 ~1 of choles- terol in ethanol so that the final concentration of sterol was 10 fig per ml or 2.8 X 10e6 M. The ultraviolet absorption spectrum was measured; the absorption maxima of Peaks 1 to 4 are 356,338,320 and 310 nm, respectively. This solution was then diluted in half with methanol and the spectrum was determined again. Before methanol dilution the ultraviolet absorbance ratio of Peak 3 to Peak 1 was 2.47, and after dilution was 0.83. The spectrum ob- tained from the filipin in methanol is corrected for dilution 1:l.
a/a-“,\MO;E%
A A’ -A
'A n 50
'A 33
x/x\x- ‘----x 16
rm\. ‘• -• IO
Q-Q-6-n-d 0.5
MU- 2 4 6 I
PH FIa. 4. Interaction of filipin with egg lecithin liposomes of
differing cholesterol content at various pH levels. Egg lecithin liposomes with 0, 5, 10, 16, 33, or 50 mole ‘$& of cholesterol were prepared in 10 mM Tris-acetate as described under “Materials and Methods.” They were added to 5.0 ml of 5.0 X 1OW6 M filipin in Tris-acetate, pH 2.9. After mixing and standing for 30 min the ultraviolet absorption spectrum was determined. In each sample the molar ratio of cholesterol to filipin always exceeded 4.
taining liposomes (15, 30), experiments were carried out to as- certain whether the spectral change reported in Fig. 3 occurred upon interaction of the antibiotic with sterol in this system. Typical results are shown in Fig. 4. The content of cholesterol
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

1922 Filipin-Cholesterol Interaction Vol. 247, No. 6
.-. METHANOL o-o 1.4-DIOXANE
MOLE % ORGANIC SOLVENTIN H$
FIG. 5. Effect of organic solvents on the breakdown of the filipin-cholesterol complex. Samples of 5, 10, 20, 40, 60, 80, and 100% (v/v) methanol in Hz0 and 1,4-dioxane in Hz0 were pre- pared, all containing 3.5 X 10e6 M filipin. Cholesterol was added to each sample in 25 ~1 of ethanol so that the final concentration was 100 rg per ml or 2.6 X 10m4 M. After mixing, the ultraviolet absorpt,ion spectrum was measured and the ratio of absorbance in Peak 3 to Peak 1 was determined. A A ratio of 1.53 was calculated for the difference in ratio for the 100% Hz0 versus the 100% organic solvent. The data are expressed as a percentage of this A value for each sample as a fraction of mole y0 solvent present. Assuming that only two ultraviolet absorbing species are present this cal- culation determines the percent,age of filipin-cholesterol complex present as compared to a 100% Hz0 sample.
in the egg lecithin-containing liposomes was varied from 0 to 50 mole %;. In all cases the concentration ratio of cholesterol (in the liposome) to filipin [C12:[F] exceeded 3, so that there was always excess cholesterol potentially able to interact with the filipin. Due to the opalescent nature of the liposomal sus- pensions it was necessary to employ a split beam spectropho- tometer where the blank cuvette contained an equivalent amount of liposomes. Clearly liposomally bound cholesterol can also
mediate a change in the ultraviolet absorption spectrum of filipin which is independent of pH over 2 to 9. At 50 mole y0 cholesterol, the general shape of the spectrum and the wave length of absorption maxima are identical with that for the cholesterol-filipin complex shown in Fig. 3. It is not feasible to prepare liposomes with a mole per cent of cholesterol above 507,. But it is clear that as the mole per cent of cholesterol in the liposomal bilayer is lowered, it becomes either less ac- cessible to filipin or is bound more tightly in the liposomal mem- brane matrix thus precluding its interaction with filipin.
Factors Affecting Breakdown and Formation of Filipin-Choles- terol Complex-Fig. 5 shows the results of experiments which test the effects of addition of methanol and 1,4-dioxane on the cholesterol-mediated ultraviolet spectral change of aqueous solu- tions of filipin. The results were calculated by assuming that only two ultraviolet absorbing species were present, filipin- cholesterol which has an ultraviolet absorbance ratio of Peak 3 to Peak 1 of approximately 2.5 and filipin with a ratio of Peak 3 to Peak 1 of 0.8. The polarity of these two solvents as ap- proximated by their dielectric constant (1 ,4-diosane = 4; meth-
anol = 33) is quite different. When the data are expressed on the basis of mole per cent of solvent present, the more nonpolar dioxane is distinctly more effective in breaking the filipin-choles-
t,erol complex. Ethanol gives results intermediate between that of methanol and dioxane.
Table I summarizes results testing a rariety of compounds and conditions which affect both the formation and breakdown
2 The abbreviations used are: C, cholesterol; F, filipin; F-C, flipin-cholesterol complex; L, lecithin; S, sterol.
TSBLE 1
Factors affecting formation and breakdown of jilipin-cholesterol complex
In Experiments I to III factors affecting the formation of the filipin-cholesterol complex were tested by determining the effect that the indicated compound had on the absorbance ratio of Peak 3 to Peak 1 of filipin. In “formation” experiments cholesterol was added to filipin solutions containing the indicated addition, whereas in “breakdown” experiments preformed filipin-choles- terol complex was added to the indicated component. In Experi- ment IV the experiment was carried out similarly so that the final concentration of Tris-acetate buffer was always 0.01 M. In Ex- periment V aqueous solutions of filipin or filipin-cholesterol com- plex were held at the indicated temperature for 30 min prior to spectral determination in cuvettes of the same temperature. In Experiments I to V the final concentration of filipin was 1 X 10m6 M, and cholesterol was added to a final concentration of 10 fig per ml by addition of 25 ~1 of ethanol. BSA, bovine serum albu- min.
“:,“,Z-
I
II
III
IV
V
I Sample
Breakdown Formarion
Hz0 BSA, 5 mg/ml BSA, 0.5 mg/ml Sucrose, 0.5 iv Sucrose, 0.005 M Galactose, 0.2 M Galactose, 0.005 M Egg lecithin,
0.0001 M
2.30 2.05 2.03 2.20 2.28 2.14 2.28 2.31
J&O 2.05 2.10 Triton, 0.5% 2.10 1.8’ Triton, 1.070 1.85 1 .X Triton, 2.09; 1.11 0.93
Hz0 2.31 2.11 Urea, 4.8 fi1 1.77 2.00 Urea, 2.4 M 2.10 2.74 Urea, 1.2 M 2.18 3.25 Urea, 0.6 M 2.18 3.25 NaCl, 3.2 M 2.70 3.00
NaCI, 1.G M 2.51 3.00 NaCl, 0.4 1~1 2.34 3.10
pH = 2.0 2.21 2.31 pH = 3.0 2.28 2.29 pH = 5.0 2.24 2.27 pH = 7.0 2.30 2.25 pH = 9.0 2.27 2.32
(Filipin)
4" .90 37 .95 67 1.10 87 1.18 87-37 1.25
(Filipin-Choles- terol)
2.95 2.56 1.5n
1.16 2.10
Absorbance ratio of Peak 3 to Peak 1
of the filipin-cholesterol complex. The results shown report interaction of filipin with free cholesterol. Essentially similar results have been obtained in most instances with lecithin-choles- terol (50:50 mole 7G) liposomes. The addition of bovine serum
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of March 25, 197‘2 Norman, Demel, de Kru$, and van Deenen 1923
albumin, sucrose or galactose, egg lecithin, moderate concentra- tions of urea or NaCl, or changes in pH from 2 to 9 had little effect on either the breakdown or formation of the filipin-choles- terol complex as measured by the ratio of absorbance in Peak 3 to Peak 1. Higher concentrations of urea (4.8 M) were some- what more effective in reducing the absorbance ratio of Peak 3 to Peak 1, whereas high SaCI (3.2 M) consistently enhanced the absorbance ratio of Peak 3 to Peak 1. The reason for the difference in ratio of Peak 3 to Peak 1 in the presence of urea and r\‘aCl for the formation reaction as compared to the break- down reaction is not known. The nonionic detergent, Triton X-100 at 25; (w/v), effectively prevents both the formation of the filipill-cholesterol complex and mediates its breakdown. Also
of interest is the effect of increasing the temperature (Table 1, esperimcnt V). As the temperature of the aqueous solution of filipin-cholesterol comples is raised from 37 to 87”, there is a shift in the absorbance rat,io of Peak 3 to Peak 1 from 2.5 to a value of 1.2 which is more characteristic of free filipin. However, this change is reversible. When the solution is cooled from 57 to 37”, the absorbance ratio reverts to that characteristic of the filipin-cholesterol complex. In the aggregate, these results suggest that at this level of organization, the interaction between filipin and cholesterol is not hydrophilic in nature, but is pri- marily hydrophobic. It woulcl be anticipated that a hydro- phobic interaction could be disrupted by organic solvents, non- ionic detergents, and heating.
Structural Requirements for Filipin-Cholesterol lnteraction- Table II summarizes experiments in which changes in the ab- sorbance ratio of Peak 3 to Peak 1 are detected as the structure or stereochemistry of the steroid is altered. These experiments lrere carried out in three II-ays by adding filipin to (a) aqueous dispersions of steroid in buffer or to (6) 16 mole c/ or (c) 50 mole c; of steroid in egg lecithin liposomes suspended in buffer. It
should be emphasized that all steroids were tested at identical concentrations of 5 X 10-j RI (free steroid) or 2 X lop5 M (lipo- some%) which was, respectively, a 10. and 4-fold molar excess over the filipin added. Thus, the relative abilities of the steroid to interact with the antibiotic are reported rather than the ab- solute rffect. In the case of the steroid alone, it is evident that
(a) 3P-hydroxysteroids cause a strong spectral change of filipin. (b) 3-keto steroids, cholesterol acetate, and to a lesser extent 3a-hydroxysteroids also interact with filipin; (c) a steroid nu- cleus is however required since lecithin or cetyl alcohol show little effect; and (d) rupture of the B ring to produce a seco- steroid (ergocalciferol and dihydrotachysterolJ prevents the steroid-filipin interaction. The consequences of these changes are likely a measure of the gross structural requirements neces- sary for an optimal filipin-steroid complex and are in agreement with the suggestion made above that the filipin-cholesterol in- teraction is basically hydrophobic in nature. These same struc- tural relationships are also required for the interaction of filipin with either 16 or 50 mole % sterol-lecithin liposomes. How- ever, of much more interest is the fact that several additional structural requirements are necessary for optimal filipin-lipo- somally bound sterol interaction. In the liposome it is shown that (a) conversion of the 3fl-hydroxy group to a Sa-hydroxy or a keto group has a profound effect on the ability of the steroid to mediate a spectral change on the ultraviolet absorbance ratio of Peak 3 to Peak 1 of filipin. Go cholesterol acetate is without
effect when present in the liposome. (b) Conversion of ring fus:on to the coprostnnol conformation and deletion of the side
TABLE II
E$ect of sterol structure on interaction of sterol withjilipin
Aliquots, 10 ~1, of free sterol in ethanol or of liposomally bound sterol, 25 ~1, in 10 rnM Tris-acetate, pH 7.0, were added to 5.0 ml of filipin, 5 X 10e6 M, in 10 m&l Tris-acetate, pH 7.0. After mixing and standing 30 min the spectra were taken and the ratio of ultra- violet absorbance of Peak 3 to Peak 1 was determined. In all instances the steroid(s) was present at a final concentration of 7.7 pg per ml = 2 X 10-j >\I. The liposomes were prepared as described under “Materials and Methods.”
Steroid or compound I- Free S
3P 7-Dehydrocholesterol . Cholestanol. Cholesterol. Ergosterol B-nor-Cholesterol (cholest-B-nor-5.
eri-3p-01). Stigmasterol Coprostanol Androstane-3p-ol Lathosterol
301
2.38 1.04 1.62 2.17 1.35 1.9-l 2.12 1.40 1.95 1.88 1.18 1.70
1.88 1.70 1.5G 1.52 1.75
Epicholesterol Androstane-3ol-01.
3-keto
1.63 1.50
Cholest-5-en-3-one. 501-Cholestall-3-olle Cholest-4-ell-3-one.
Other
2.50
2.27
2.03
Cholesterol-acetate. Dihydrotachysterol Ergocalciferol (vitamin DY) Cetyl alcohol. Lecithin
1.78
0.92
0.84
0.78
0.88
Filipin alone. 0.72
Absorbance ratio of Peak 3 to Peak I
0.86
1.71
0.90
0.94
0.88
0.85
0.90
0.88
0.88
0.87
0.87
2.23
0.95
1.43
1.04
0.9G 1.04
chain (androstane-S/3-01) abolishes the effect. (c) The iutro- duction of a A7 double bond (lathosterol, 7-dehydrocholesterol, ergosterol) decreases the interaction, whereas introduction of a A22 double bond (ergosterol, stigmasterol) enhances the inter- action. In some manner the stereochemistry and nature of the substituent at position 3 play a crucial orienting role in terms of the ability of the sterol in the liposome to interact with the antibiotic.
Demel et al. (13, 14) have previously suggested on the basis of monolayer studies that a free sterol hydroxyl group was necessary for the interaction of cholesterol with filipin. In view of the results shown in Table II, an experiment was initiated to ex- amine in more detail the structural requirements for filipin-sterol interaction in monolayers of pure sterol and in mixed lecithin- sterol monolayers. Results of such an experiment are given in Figs. 6 and 7. In Fig. 6 are shown the pressure increases in monolayers of pure sterols resulting where filipin is injected into the subphase at a concentration of 6.3 x 10eQ M, whereas Fig. 7 shows the resulting pressure increases in mixed monolayers of sterol-lecithin (1: 1) when filipin is injected in the subphase. In both instances the magnitude of pressure increase was highly
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Filipin-Cholesterol Interaction Vol. 247, Iv'o. 6
STIGMASTEROL \
B ; m EGG LECITHIN
2 -m-m------q-
5 IO 15 20 25
B EIO- /*Ns. 2 G
q STIGMASTEROL
2 l CHOLESTEROL
5-
2 z
. EGG LECITHIN
3 ~-m-------,-
v)
-. CHOLEST-5-EN-3-ONE V CHOLESTAN-3-ONE A CHOLEST-4-EN-3-ONE v ERGOCALCIFEROL (VITD2)
15 20 25 IN!TIAL SURFACE PRESSURE
I r Q ERGOSTEROL / B LATHOSTEROL ~ o EPiCHOLESTEROL
o COPROSTANOL 1
A CHOLEST-5-EN-3 -ONE v CHOLESTAN - 3 -ONE A CHOLEST-4-EN-3-ONE
5 10 15 20 25 5 IO 15 20 25 INITIAL SURFACE PRESSURE
dependent upon the stereochemistry at the steroid nucleus. The highest pressure increase in pure steroid monolayers is ob- served with cholesterol and stigmasterol, although epicholesterol also shows a strong effect. The pressure increase was signifi- cantly reduced by deletion of the side chain (androstane-3-p-01), the presence of the coprostanol conformation between ring A/B, the opening of the B ring to give a secosteroid (ergocalciferol), or by the presence of a keto functional group. In mixed sterol- lecithin monolayers the highest pressure increase is also found for cholesterol and stigmasterol. Ergosterol and lathosterol show only a significant pressure increase at an initial surface pressure of 14 dynes per cm. Epicholesterol, coprostanol, or keto steroids show little effect in mixed monolayers. The chief difference in response of the pure versus mixed sterol-lecithin monolayers was apparent in the results obtained with epicholes- terol. In the pure steroid monolayer quite significant pressure increases after filipin addition were apparent with epicholesterol, whereas in the mixed lecithin-epicholesterol monolayer only small pressure increases were noted.
Mixed monolayers show the same specificity for the sterol- filipin interaction as was observed for liposomes (Table II). Be- sides the presence of a 3@-OH group, structural requirements for an intact sterol moiety are also evident. Hydrophobic interac- tions in monolayers can possibly occur by reorientation of the sterol molecules towards the interphase since the observed area changes are small (31).
Studies on Stoichiometry of F&pin-Cholesterol Complex-It was found possible to bring about a gradual increase in the ratio of
u-1 5 IO 15 20 25
FIG. 6. Effect of filipin on mono- layers of pure steroid. Monolayer-s of the indicated steroid were pre- pared as described under “Materials and Methods.” After a stable initial surface pressure was obtained filipin v-as added to the subphese at 6.3 x 10e9 15 and the increase in surface pressure measured.
FIG. 7. Effect of filipin on mixed monolayers of lecithinsteroid. Mon- olayers of egg lecithin-sterol, l:l, were prepared as described under “Materials and Methods.” After a stable initial surface pressure was ob- tained, filipin was added to the sub- phase at 6.3 X 10m9 M and the increase in surface pressure measured
absorbance of Peak 3 to Peak 1 of aqueous solutions of filipin from 0.8 to a limiting value of approximately 2.5 by the addition of increasing amounts of cholesterol. Clearly there existed a proportionality between the amount of cholesterol added and the magnitude of increase in the ratio of absorbance of Peak 3 to Peak 1. The simplest description of the system would be to assume that only two ultraviolet absorbing species are present. One species would be free filipin with an absorbance ratio of Peak 3 to Peak 1 of 0.8; the second species must be some form of filipin the ultraviolet spectrum of which is altered by virtue of an interaction with an unknown number of cholesterol molecules so that the ultraviolet absorbance ratio of Peak 3 to Peak 1 can approach in the limit a value of aproximately 2.5. Thus it was possible to estimate the relative amounts of each of these species by evaluation of the change in absorbance ratio of Peak 3 to Peak 1 and calculation of a A in the ratio of absorbance of Peak 3 to Peak 1 due to the addition of cholesterol.
A = (absorbance ratio of Peak 3 to Peak 1 in presence of sterol)
- (absorbance ratio of Peak 3 to Peak 1 in absence of sterol)
If the interaction between F and S can be written as F + nS * FSn then Aa[FSn].
A plot of A versus [cholesterol] is similar in form to that of a Langmuir adsorption isotherm. Then l/A versus l/[cholesterol] could be plotted to permit evaluation of the cholesterol concen- tration necessary to produce a one-half maximal spectral change. At this point the concentration of free filipin and filipin-sterol complex can be considered equal. This analysis was utilized to
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from
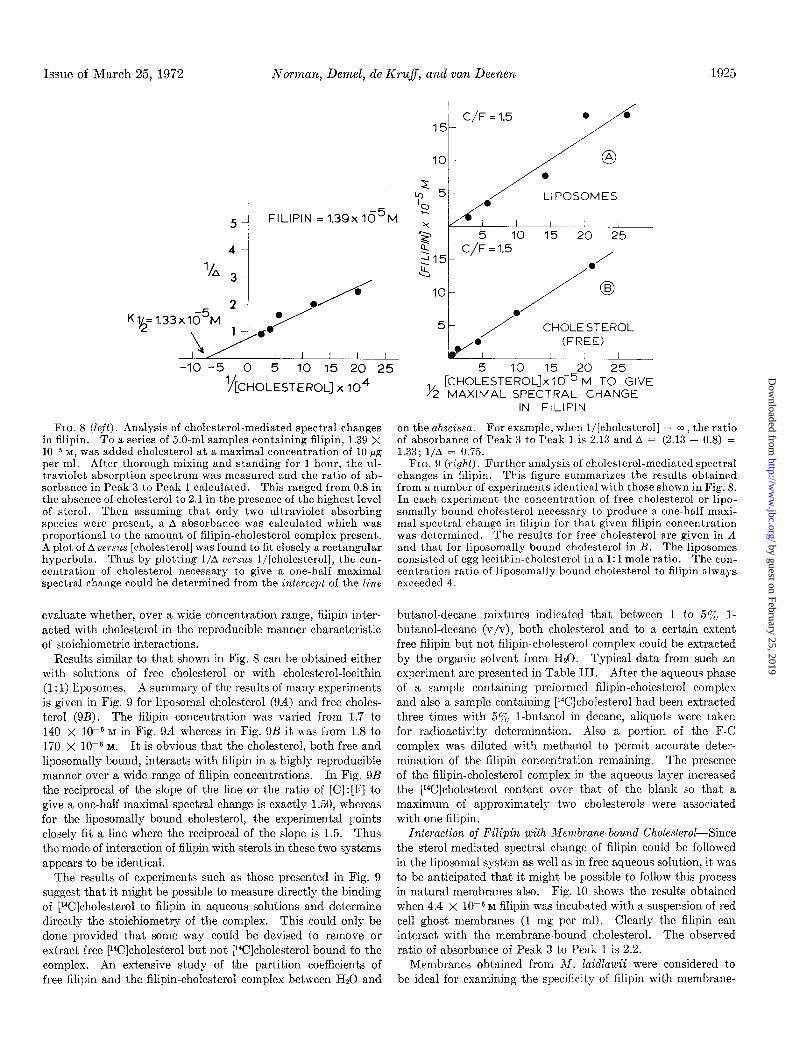
Issue of March 25, 1972 NoTman, Demel, de Kru$, and van Deenen 1925
5 FILlPIN = 1.39x 1e5M
4
‘/[CHOLESTEROL] x 10~
FIQ. 8 (left). Analysis of cholesterol-mediated spectral changes in filipin. To a series of 5.0-ml samples containing filipin, 1.39 X 10e5 M, was added cholesterol at a maximal concentration of 10 pg per ml. After thorough mixing and standing for 1 hour, the ul- traviolet absorption spectrum was measured-and the ratio of ab- sorbance in Peak 3 to Peak 1 calculated. This ranged from 0.8 in the absence of cholesterol to 2.1 in the presence of thk highest level of sterol. Then assuming that only two ultraviolet absorbing species were present, a A absorbance was calculated which was proportional to the amount of filipin-cholesterol complex present. A plot of A uersu.s [cholesterol] was found to fit closely a rectangular hyperbola. Thus by plotting l/A versus l/[cholesterol], the con- centration of cholesterol necessary to give a one-half maximal spectral change could be determined from the intercept of the line
evaluate whether, over a wide concentration range, filipin inter- acted with cholesterol in the reproducible manner characteristic of stoichiometric interactions.
Results similar to that shown in Fig. 8 can be obtained either with solutions of free cholesterol or with cholesterol-lecithin (I: 1) liposomes. A summary of the results of many experiments is given in Fig. 9 for liposomal cholesterol (9A) and free choles- terol (9B). The filipin concentration was varied from 1.7 to 140 x 1OV M in Fig. 9A whereas in Fig. 9B it was from 1.8 to 170 x 10e6 M. It is obvious that the cholesterol, both free and liposomally bound, interacts with filipin in a highly reproducible manner over a wide range of filipin concentrations. In Fig. 9B the reciprocal of the slope of the line or the ratio of [C] : [F] to give a one-half maximal spectral change is exactly 1.50, whereas for the liposomally bound cholesterol, the experimental points closely fit a line where the reciprocal of the slope is 1.5. Thus the mode of interaction of filipin with sterols in these two systems appears to be identical.
The results of experiments such as those presented in Fig. 9 suggest that it might be possible to measure directly the binding of [Wlcholesterol to filipin in aqueous solutions and determine directly the stoichiometry of the complex. This could only be done provided that some way could be devised to remove or extract free [r4C]cholesterol but not [14C]cholesterol bound to the complex. An extensive study of the partition coefficients of free filipin and the filipin-cholesterol complex between H20 and
[C~~~LESTEROLJX 10P r-4 To GIVE 1/ MAXIMAL SPECTRAL CHANGE
IN FILlPIN
on the abscissa. For example, when l/[cholesterol] = m , the ratio of absorbance of Peak 3 to Peak 1 is 2.13 and A = (2.13 - 0.8) = 1.33; l/A = 0.75.
FIG. 9 (right). Further analysis of cholesterol-mediated spectral changes in filipin. This figure summarizes the results obtained from a number of experiments identical with those shown in Fig. 8. In each experiment the concentration of free cholesterol or lipo- somally bound cholesterol necessary to produce a one-half maxi- mal spectral change in filipin for that given filipin concentration was determined. The results for free cholesterol are given in A and that for liposomally bound cholesterol in B. The liposomes consisted of egg lecithin-cholesterol in a 1: 1 mole ratio. The con- centration ratio of liposomally bound cholesterol to filipin always exceeded 4.
but.anol-decane mixtures indicated that between 1 to 5% l- butanol-decane (v/v), both cholesterol and to a certain extent free filipin but not filipin-cholesterol complex could be extracted by the organic solvent from HSO. Typical data from such an experiment are presented in Table III. After the aqueous phase of a sample containing preformed filipin-cholesterol complex and also a sample containing [W]cholesterol had been extracted three times with 5gF I-butanol in decane, aliquots were taken for radioactivity determination. Also a portion of the F-C complex was diluted with methanol to permit accurate deter- mination of the filipin concentration remaining. The presence of the filipin-cholesterol complex in the aqueous layer increased the [YJcholesterol content over that of the blank so that a maximum of approximately tm-o cholesterols were associated with one filipin.
Interaction of Filipin with Membrane-bound Cholesterol--Since the sterol-mediated spectral change of filipin could be followed in the liposomal system as well as in free aqueous solution, it was to be anticipated that it might be possible to follow this process in natural membranes also. Fig. 10 shows the results obtained when 4.4 x 10e6 M filipin was incubated with a suspension of red cell ghost membranes (1 mg per ml). Clearly the filipin can interact with the membrane-bound cholesterol. The observed ratio of absorbance of Peak 3 to Peak 1 is 2.2.
Membranes obtained from M. Zaidlawii were considered to be ideal for examining the specificity of filipin with membrane-
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

1926 F&pin-Cholesterol Interaction Vol. 247, No. 6
T.IBLE III
Association of [W]cholesterol with aqueous solutions of flipin [l”C]Cholesterol, 890 cpm per nmole, was added in 25 ~1 of etha-
nol to 5.0 ml of Hz0 or 8.0 X low6 M filipin in HZO. After thor- oughly mixing the samples, aliquots were taken for spectral meas- urement and l4C content. Then 5.0 ml of 5% 1-butanol-95yo decane (v/v) were added to each sample. After vortexing for 3 min, the samples were centrifuged at 3000 rpm for 15 min and the upper organic phase aspirated away. This procedure was re- peated five times with 0.50.ml aliquots of the aqueous layer being retained after each wash for ‘4C content determination. After the last wash, the spectra of the aqueous phase containing the filipin- cholesterol complex was determined. The ratio of ultraviolet absorbance of Peak 3 to Peak 1 was 1.89. This aliquot was then diluted with methanol to break the filipin-cholesterol complex. The final concentration of filipin in the aqueous phase was 5.7 X 1O-6 M. Under the conditions of this experiment, approximately 90% of the filipin partitions into the aqueous phase in the presence of 5c/b 1-butanol-95y0 decane.
SkUllpIe PClCholesterol
tFilipin I
--Filipin
cfm/O.50 ml Hz0
At start. _. 11,200 12,600 After Wash 1. 10,300 5,950 After Wash 3. 9,500 4,300 After Wash 5. 6,700 1,550
BASELINE 4
400 350 300 250 WAVELENGTH (NM)
FIG. 10. Interaction of filipin with red cell ghost membranes. Sonicated red cell ghost membranes, in 20 mM sucrose-10 mM Tris- acetate, pH 7.0, were obtained as described under “Materials and Methods.” Identical suspensions of this preparation were placed in the sample and reference cuvettes of a split beam spectropho- lometer. After a stable base-line was obtained, filipin was added to the sample cuvette in 10 ~1 of Tris-acetate buffer to give a final concentration of 4.4 X lOwE M.
bound sterol. This organism neither biosynthesizes cholesterol nor requires its presence in the growth media (5). However
X. laidlawii will incorporate sterol into its membranes when it is present. Thus membranes may be obtained either with or without sterol. Tn the nonsterol-containing membranes, there
.25;
.20 c
.I 5 t
.I0 c
+ EPICHOL. i
.05 -
400 350 300 350 300 400 350 300 WAVELENGTH
FIG. 11. Interaction of filipin with cells of Mycoplasma laidlawii. Cell suspensions of M. Zaidlawii grown in the absence of any sterol, or in the presence of either cholesterol or epicholesterol, were obtained as described under “Materials and Methods.” Identical suspensions of the cells in 0.20 M sucrose were placed in the reference and sample cuvettes of a split beam spectrophotom- eter. After obtaining a stable base-line, filipin was added in 10 ~1 of Tris-acetate, pH 7.0, to the sample cuvette to a final concentra- tion of 9.4 X 10e6 M.
is no spectral evidence of membrane interaction in the presence of 9.4 x 10e6 M filipin, i.e. t’he ratio of absorbance of Peak 3 to Peak 1 is 0.8. In separate experiments, when such membrane preparations have been centrifuged at 105,000 x g for 80 min so that 75% of the protein has been sedimented, 80% of the filipin absorbance remains in the supernatant. This indicates that in the absence of sterol the filipin does not associate with the membrane to a significant extent. However if exogenous cholesterol was added to this cholesterol-deficient system a spectral change was evident instantly. Now the ultraviolet absorbance ratio of Peak 3 to Peak 1 became 2.5. With choles- terol-containing membranes, as expected, the filipin int.eracted with the sterol so that the ratio of absorbance was 1.58, which is indicative of formation of a filipin-cholesterol complex. Fig.
11 illustrates the ability of filipin to interact with intact cell suspensions of M. Zaidlawii. In this experiment the cells were grown either in the absence of sterol or in the presence of choles- terol or epicholesterol. Chemical analysis of the cell prepara- tions revealed that there was 14 and 15 mole y0 of the total lipid present as cholesterol or epicholesterol, respectively. Referring
to Fig. 4 it can be seen that lecithin liposomes, 15 mole s) in sterol, have an ultraviolet absorbance ratio of Peak 3 to Peak 1 of 1.4 to 1.5 in the presence of filipin. Thus, the results obtained with cholesterol-containing intact cells is quantitatively similar to cholesterol-containing liposomes and cholesterol-containing membranes. Of much greater significance are the results ob- tained with the epicholesterol-containing cells. No spectral evidence was obtained for a filipin-sterol interaction, which is in agreement with the liposomal data presented in Table II.
Fig. 12 presents results of the differential scanning calorimetric analysis of various combinations of synthetic (18: 1 to 18:O) lecithin, cholesterol, and filipin. Pure lecithin with a fatty acid
composition of 18: 1 to 18:0 exhibits a sharp phase transition at approximately 13” characteristic of conversion of the lecithin from the L (a) crystalline phase to the L (0) liquid crystalline phase. Addition of 20 mole Yc cholesterol markedly reduces
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of &‘/larch 25, 1972 Norman, Demel, de Kruff, and van Deenen 1927
SAMPLE ENERGY UNITS PER MGLECITHIN
/:i
(18:l h8:O)PC 147
(18:l /l8:O)PC 75 + 20 MOLE% /\A CHOLESTEROL
(18 : 1/18:OIPC 132
+ FILlPIN
(18 : 1/18:O)PC + 20 MOLE%
,48
CHOLESTEROL t FILIPIN
0 20 40°C FIG. 12. Effect of filipin on the phase t,ransitions of lecithin-
cholesterol dispersions as determined by differential scanning calorimetry. Samples of synthetic l-oleoyl-2-stearoyl-sn-glycero- 3.phosphatidylcholine (18:1/18:0, PC), cholesterol, and filipin were analyzed by differential scanning calorimetry as described under “Materials and Methods.” The figure presents graphically the results obtained from samples with approximately equivalent amounts of lecithin, whereas the tabular results show the actual energy units required per milligram of lecithin. The particular synthetic lecithin employed undergoes the phase transition from the crystalline L (a) form to the liqnicrystalline L (p) form at 13”.
the magnitude of the phase transition as has been previously reported (31). Addition of filipin to pure synthetic lecithin causes only a lOGA reduction in the energy required to bring about the phase transition. Thus filipin has no significant effect on phase transition of pure lecithin. However addition of filipin to lecithin-cholesterol (80 : 20 mole ‘:i j causes a striking reversal of the effect of cholesterol on the phase transition of lecithin- cholesterol mixtures. Now the lecithin exhibits a sharp phase transition which requires as much energy as a pure lecithin solu- tion. Addition of filipin to the lecithin-cholesterol system has resulted in the formation of a filipin-cholesterol complex which apparently results in either rupture of t,he lecithin-cholesterol interaction or physical relocation of the cholesterol in the dis- persion. Now the sample exhibits :I phase transition charac- teristic of the pure lecithin. These results permit insight into some of the possible physical bases for filillin-mediated changes in membrane systems.
We have studied the interaction of filipin with sterols at three levels of organization: (a) with free sterol in aqueous solutions; (b) with sterol incorporated into lecithin-containing liposomes; and (c) with sterol incorporated in biological membranes. With free sterol we have found (Table I) that, the formation of the iilipincholesterol complex can occw in the presence of protein,
carbohydrate, phospholipid, salt, and urea, and over a range of pH levels from 2 to 9. Similarly, these same parameters do not cause the breakdown of the filipin-cholesterol complex. Thus at this level of organization, the interaction of filipin with choles- terol is primarily hydrophobic in nature and is not dependent upon hydrogen bond formation. However, organic solvents (Fig. 5), nonionic detergent,s, and high temperature (Table I) can break the sterol-antibiotic complex. The effect.iveness of organic solvents to break the complex is inversely related to their polarity or dielectric constant. Clearly no covalent bond formation has occurred between the antibiotic and sterol. Com- plex formation can occur reversibly. The filipin-sterol interac- tion is optimal (Table II, free S column) when (a) there is a steroid nucleus (compare with cetyl alcohol and lecithin); (b) the B ring of the steroid nucleus is intact (compare with ergo- calciferol and dihydrotachyst,erol) ; (c) the steroid nucleus has the cholestane rather than the coprostane conformation; (d) there is a side chain (compare with androstane-3P-ol) ; and (ej also the conformation of the hydroxyl at position 3 and the exact positioning of double bonds in the B ring have important effects.
The interaction of filipin with organized structures of sterols present in liposomes shows even a more complex picture. Al- though the effects of heat, nonionic detergents, and organic solvents and the lack of effect of changes in pH and ionic en- vironment also are applicable here, there are additional struc- tural requirements not only on t)he steroid itself but also in terms of the steroid-lecithin structure. Of central importance is the amount of sterol relative to lecithin (see Fig. 4). When there is less than 10 mole y0 of cholesterol in the liposomes, the sterol is apparently incapable of interacting with filipin. Previous workers were unable to show effects of low concentrations of filipin on monolayers (13), bimolecular lipid membranes (IQ, or in natural membranes with a low cholesterol content (32). For optimum interaction the bilayer must approach the struc- tural characteristic of a 1: 1 mole ratio of lecithinsterol. When a variety of steroids are incorporated into lecithin-containing liposomes either at 16 or 50 mole c/ steroid (see Table II), it is apparent that for optimal antibiotic-steroid interaction that there must be a 3fl-hydroxyl group. Neither epicholesterol, androstane-3-o-01, or any of the 3-keto steroids produced a significant spectral change in filipin. Also the cholestane rather than the coprostane ring or j-membered B ring (P-nor-choles- terol) is essential for optimal interaction. Further, the optimum steroid has been found to be stigmasterol, which is a steroid with a cholestane ring conformation, a 3p-hydroxyl group, A5, 22 double bonds, and AC-24 ethyl group.
When a detailed study was carried out of the steroid structural requirements necessary for optimum interaction (as defined by pressure increases) of filipin with either monolayers of pure steroids or mixed monolayers of sterol-lecithin (1: I), results were obtained which were essentially identical with those from a study of the spectral changes with free and liposomally bound steroid. That is compounds like lecithin and ergocalciferol gave little or no pressure increases, whereas steroids with a 3a-hydrosyl, 3- keto, or coprostane ring conformation have only moderate pres- sure changes, and steroids with a 3P-hydroxyl gave the largest pressure changes after filipin was added to the subphase. Pre- vious work by Demel et al. (13) with the monolayer system had emphasized the importance of the 3&hydroxyl group for filipin interaction.
At the highest level of organization, the interaction of filipin
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

1928 Filipin-Cholesterol Interaction Vol. 247, No. 6
with membrane-bound sterol has been clearly documented with the aid of the split beam spectrophotometer. The spectral change characteristic of this interaction has been shown to occur in red cell ghost membrane preparations (Fig. lo), in M. ZaidZawii membrane preparations and in cells of M. Zaidluwii (Fig. 11). The M. laidlawii system is particularly useful for these studies since this organism will only contain sterol in its membranes if sterol is present in the growth media. As shown in Fig. 11, when there is no cholesterol in the membrane, no change in the spectrum of filipin can be detected. Of even more significance is the finding that the requirement for a 3&hydroxyl group as compared to a 3cr-hydroxyl group carries over also to this level of organization. Thus although epicholesterol and cholesterol were present in the membranes, as defined by chemical analysis, in nearly identical concentrations only the membrane containing a 30.hydroxyl sterol can give any evidence of interaction with filipin. In separate experiments we have determined that the interaction of filipin with membrane-bound cholesterol does not physically remove the sterol from the membrane and that filipin does not bind or absorb to any significant extent to cholesterol-free membranes. Thus there is an excellent corre- lation between the presence of st’erol in membranes at certain minimal levels and its ability to interact with filipin.
Since filipin is a polyene compound it has a highly character- istic ultraviolet absorption spectrum. Although no significant wave length shift occurs, clearly aqueous solutions of filipin absorb ultraviolet light differently from organic solutions. Thus filipin in organic solvents follows Beer’s law over a wide range of concentrations, whereas filipin in Hz0 does not. This deviation from Beer’s law is not simply explainable in terms of insolubility of filipin since (Fig. 3) as the filipin was diluted, the absorbance of Peak 1 increased, while that of Peak 4 decreased. A more likely explanation is that due to the amphipathic properties of filipin, at higher concentrations in I&O some aggregation of micelle formation occurs, which affects the ultraviolet chromo- phores in a concentration-dependent manner.
When cholesterol is added to filipin in organic solvents no change in spectra occurs, but when the sterol is added to filipin in Hz0 a dramatic alteration in the ultraviolet absorption spectra is apparent (Fig. 3). The wave lengths of absorption maxima are surprisingly independent of the presence or absence of sterol. The alteration is best characterized by determining the ratio of absorbance in Peak 3 to Peak 1. Free filipin in Hz0 has a ratio of 0.8, whereas addition of excess sterol produces a limiting value of 2 to 3. The reasons for the variation in the limiting value of the absorbance ratio of Peak 3 to Peak 1 in the presence of sterol are not clear. Analysis of the ratio of absorbance of Peak 3 to Peak 1 has the advantage of being independent of the anti- biotic concentration; only the ratio of sterol to filipin determines the exact magnitude of this ratio (Fig. 9B).
We have exploited this spectral change to study in a detailed manner the interaction of filipin with sterol in a number of systems. On the basis of the results reported here, we believe that we have provided the evidence that filipin is capable of interacting directly with sterol in a defined manner to form a filipin-sterol complex. Lampen et al. (10) and Gottlieb et al. (9) also previously demonstrated spectral changes upon addition of cholesterol to aqueous solutions of filipin and that this alteration could be reversed by organic solvent addition. Also these work- ers noted some correlation between structure of the sterol and the ability of the sterol to block the biological action of filipin.
The addition of increasing amounts of cholesterol to filipin results in a proportionate change in ratio of absorbance of Peak 3 to Peak 1 from 0.8 up to a limiting value of 2-3. The most simple interpretation of the spectral change is that there are only two ultraviolet absorbing species present, each with a characteristic ratio of absorbance of Peak 3 to Peak 1; free filipin with a value of 0.8 and the filipin-sterol complex with a value of 2 to 3. It was possible to evaluate the concentrat’ion of sterol necessary to mediate a one-half maximal spectral change as a function of filipin concentration by preparing reciprocal plots of l/(A absorbance) versus l/[cholesterol] (Figs. 8 and 9). At this point the concentration of free filipin and filipin-sterol complex can be considered equal. As shown in Fig. 9 this n-as accomplished for both free and liposomally bound cholesterol. With the free cholesterol, a precise linear relationship existed between the concentration of cholesterol required for a one-half maximal spectral change over filipin concentrations from 1 to 1000 X lo+ M. The slope of the line was exactly 0.66, i.e. [C] :[F] = 1.5. Similar result’s were also obtained with the liposomally bound cholesterol. Again the ratio of [C] : [F] was approximately 1.5 when the cholesterol produced a one-half maximal spectral change. Collectively this is strong evidence that the mode of interaction of filipin in both these systems is identical in terms of stoichiometry between antibiotic and sterol at all concentrations of the antibiotic.
Preliminary results obtained from the binding of [14C]choles- terol to filipin in biphasic systems (Table III) suggest that only 2 cholesterol molecules are associated with 1 filipin molecule. A previous estimate of the stoichiometry of the F-C complex was obtained from the monolayer layer studies of Demel et al. (31). They found that 1 filipin per 100 molecules of sterol could cause a significant pressure increase. However, their results are likely “system-dependent” since the collapse pressure of cholesterol was already reached at filipin concentrations of 6 x lo-’ M.
With the knowledge of the existence of a defined complex between filipin and cholesterol and also knowing that filipin treat- ment of several membrane systems caused changes in membrane permeability to a variety of small molecules, it was of interest to ascertain whether filipin could mediate any changes in the phys- ical properties of lecithin-cholesterol dispersions. The analysis was carried out by differential scanning calorimetry using prep- arations of chemically synthesized lecithin with a defined fatty acid composition. Such lecithins exhibit sharp phase transitions over a narrow temperature range, characteristic of the conver- sion from the crystalline L (CX) to liquid-crystalline L (0) phase (33). The exact temperature of the phase transition is deter- mined by both the fatty acid composition and polar group. This transition is believed to reflect melting of the fatty acid hydrocarbon chains from an ordered to a more random structure. It was also known that the addition of cholesterol, up to 50 mole ‘$& to the synthetic lecithin abolishes the phase transition (34). Apparently incorporation of cholesterol leads to fluidization of the hydrocarbon chains.
Addition of filipin to lecithin-cholesterol dispersions caused a complete reappearance of the phase transition characteristic of pure lecithin. In a control experiment, filipin addition to pure lecithin dispersions caused no alteration in the amount of energy required for a phase transition. Filipin addition to a lecithin- cholesterol dispersion leads to formation within that dispersion of the f&pin-cholesterol complex. Formation of this complex
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

Issue of March 25, 1972 Norman, Demel, de Kru$, and van Deenen 1929
abolishes the fluidizing effect that the sterol had on the hydro- carbon chains and now they undergo the phase transition from ordered to melted. Thus filipin addition to a lecithin-cholesterol disperion has mediated a striking change in one of the physical properties of the sample.
Although it is not known definitely whether such a change in phase transition would specifically lead to a more permeable liposome bilaper, these results do provide strong support for the concept that filipin addition to sterol-containing liposomes or membranes can lead to changes in the molecular architecture in these systems. Such structural changes might provide the basis for a physical description of filipin-mediated increases in permeability of (a) liposomes to glucose and chromate (15, 30), (b) of intestinal mucosal tissue from vitamin D-deficient chicks to only calcium (18, 19), and (c) of black lipid films to primarily calcium (17).
Also Kinsky et al. (35) have previously presented electron micrographic evidence that filipin treatment of sterol-containing membranes and liposomes can cause “pit” formation in the membrane. These changes likely are further evidence of the ability of filipin to cause changes in the physical organization of sterol-containing membranes.
The present report provides evidence for the formation of a filipin-cholesterol complex with a defined stoichiometry and in- dicates how formation of such a complex in a liposomal or mem- brane system may lead to changes in the permeability of these systems. It will be of interest in the future to compare these results obtained with filipin with that of some of the other polyene antibiotics, e.g. pimaricin, nystat.in, etruscomycin, and ampho- tericin B. Each of these compounds has differing biological activities in various systems, which are believed to be dependent to varying extents on the presence of sterol (36).
Acknowledgment-The expert technical assistance of Mr. W. S. M. Geurts van Kessel is gratefully acknowledged.
REFERENCES
1. NORMAN, A. W., DEMEL, R. A., AND VAN DEEKEN, L. L. M. (1971) Fed. Proc. 30, 1282
2. CED.~R, O., AND RYH~GE, R. (1964) Acta Chem. &and. 18, 558 3. BERGY, M. E., AND EHLE, T. E. (1968) Biochemistry ‘7, 653 4. PAXDEY, R. C., AND RINEHSRT, K. L. (1970) J. Antibiot.
(Tokyo) Ser. A 23, 414 5. KINSICY, S. C., DEMEL, R. A., AND VAN DEENEN, L. L. M.
(1967) Biochim. Biophys. Acta 136, 835-843 6. KIXSI~Y, S. C. (1963) Arch. Biochem. Biophys. 102, 180
7. GOTTLIEB, D., CARTER, H. E., SLONEKER, J. Y., AND AMMAN, A. (1958) Science 128, 361
8. GOTTLIEB, D., CARTER, H. E., AND G~NDY, E. (1959) Bacterial. Proc. 1159, 114
9. GOTTLIEB, D., CARTER, H. E., SLONEI~ER, J. H., WV, L. C., AND GANDY, E. (1961) Phytopathology 61, 321
10. LAMPEN, J. O., ARROW, P. M., AND SAFFERMAN, R. S. (1960) J. Bacterial. 80, 200
11. WEBER, M. M., AND KINSKY, S. C. (1965) J. Bacterial. 89, 306- 312
12. FEINGOLD, D. S. (1965) Biochem. Biophys. Res. Commun. 19, 261-267
13. DEMEL, R. A., VAX DEENEN, L. L. M., AND KINSICY, S. C. (1965) J. Biol. Chem. 240, 2749-2753
14. D&EL, R. A., CROMBAG, F. J. L., VAN DEENEN, L. L. M., AR’D KINSICY. S. C. (1968) Biochim. Biophus. Acta 160, 1-14
15. KIXSI~Y, i. C., H.&BY; J., KINSICY, d. g., DEMEL, n. A., AXD VAN DEENEN, L. L. M. (1968) Biochim. Biophys. Acta 162, 174-185
16. VAN ZUTPHEN, H., VAX DEEKEN, L. L. M., AXD KINSKY, S. C. (1966) Biochem. Biophys. Res. Commun. 22, 393-398
17. VAN ZUTPHEN, H., DEMEL, R. A., NORMAN, A. W., AXD VAN DI~ENEN, L. L. M. (1971) Biochim. Biophys. Acta. 241, 310
18. ADAMS, T. H., WONG, R. G., AND NORMAN, A. W. (1970) J. Biol. Chem. 246, 4432-4442
19. WONG, R. G., ADAMS, T. H., ROBERTS, P. A., AND NORMAN, A. W. (1970) Riochim: Biophys. Acta 2i9, 61
20. SWEELEY. C. C.. O’CONNOR. J. D.. AND BIEBER. L. L. (1970) Chem. &al. Interactions 2; 247 ‘
~ I
21. VAN DEENEN, L. L. M., AND DE HAAS, G. H. (1964) Advan. Lipid Res. 2, 167
22. PANGBORN, M. C. (1951) J. Biol. Chem. 188, 471 23. ROELEFSON, B., DE GIER, H., AND VAN DEENEN, L. L. M. (1964)
J. Cell Comp. Physiol. 63, 233 24. PARPART, A. K. (1942) J. Cell Comp. Physiol. 19, 248 25. MCELHANEY, R. N., AND TOURTELLOTTE, M. E. (1970) Bio-
chim. Biophys. Actu 202, 120-128 26. VAN GOLDE, L. M. G., MCELHANEY, R. N., AND VAN DEENEN,
L. L. M. (1971) Biochim. Biophys. Acta 231, 245 27. LOWRY, 0. N., R&EBROUGH, N.-J., FARR, A. L., AND RANDALL,
R. J. (1951) J. Biol. Chem. 193, 265 28. FISICE, c‘. H.,.AND SUBBAROW, Y. (1929) J. Biol. Chem. 66,375 29. WHITFIELD, G. B., BROCK, T. D., AMMANN, T. D., GOTTLIF,B,
D., AND CARTER, H. E. (1955) J. Amer. Chem. Sot. 77,4799 30. SESSA, G., AND WEISSMANN, G. (1968) J. Biol. Chem. 243, 4364 31. DEMEL, It. A., VAN DEENEN, L. L. M., AND KINSKY, S. C.
(1965) J. BioZ. Chem. 240, 2749-2753 32. KINSICY, S. C., GRON~U, G. R., AND WEBER, M. M. (1965)
Mol. Pharmucol. 1, 190 33. L~DBROOKE, B. D., AND CHAPMAN, D. (1969) Chenz. Phys.
Lipids 3, 304 34. L~DBROOKE, B. D., WILLIAMS, R. M., A~VD CH‘ZPMAN, D. (1968)
Biochim. Biophys. Acta 150, 333 35. KINSICY, S. C., LUSE, S. A., AND VAN DEENEN, L. L. M. (1966)
Fed. Proc. 26, 1503 36. KINSITY, S. C. (1970) Annu. Rev. Pharmacol. 10, 119
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from

A. W. Norman, R. A. Demel, B. de Kruyff and L. L. M. van DeenenDIRECT INTERACTION OF FILIPIN WITH CHOLESTEROL
Studies on the Biological Properties of Polyene Antibiotics: EVIDENCE FOR THE
1972, 247:1918-1929.J. Biol. Chem.
http://www.jbc.org/content/247/6/1918Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/247/6/1918.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on February 25, 2019http://w
ww
.jbc.org/D
ownloaded from