studies ofthe secretion ofdm5migu4zj3pb.cloudfront.net/manuscripts/106000/106767/...studies ofthe...
TRANSCRIPT

Studies of the Secretion of
Monkey Placental Lactogen
C. BELANGER, B. SHOME,H. FRESEN, and R. E. Myms
From the Royal Victoria Hospital, Montreal, and National Institutes of Health,Bethesda, Maryland 20014
A B S T R A C T A radioimmunoassay for monkey pla-cental lactogen (MPL) was developed to study the fac-tors controlling the secretion of MPL. The sensitivity ofthe assay was 1 ng MPL per ml. Human and monkeygrowth hormone, and human placental lactogen(HPL)showed minimal cross-reactions with MPL. MaternalMPL concentrations as measured in 40 rhesus monkeysincreased progressively throughout pregnancy to amean of 5000 ng/ml at term while umbilical vein MPLwas less than 50 ng/ml. After term delivery maternalMPL concentrations decreased rapidly with a ti of20 min.
After fetectomy but with retention of the placenta,MPL concentrations decreased by 25% reaching a pla-teau over a 6 hr period. Experimental abruption of thesecondary placenta within 1 hr produced a 50% decreasein MPL concentration. After ligation of the fetal ves-sels supplying the secondary placental disc, MPL in-creased transiently and then decreased to levels signifi-cantly below those of the control period.
These studies suggest MPL secretion is not directlycontrolled by the fetus but is sensitive to changes inplacental blood flow. The pregnant rhesus monkey servesas a useful model for investigating factors which mayregulate HPL secretion because of the close similaritybetween MPLand HPL secretion.
INTRODUCTIONNumerous studies of the concentration of human pla-cental lactogen (HPL)' have been made in normal andabnormal pregnancies in attempts to define its functionin pregnancy. However, the precise biological signifi-cance of placental lactogen during pregnancy remains
Received for publication 22 March 1971 and in revised formt30 June 1971.
'Abbreviations used in this paper: HCG, human chorionicgonadotropin; HGH, human growth hormone; HPL, humanplacental lactogen; MGH, monkey growth hormone; MtPL,monkey placental lactogen.
obscure. It is not possible to subject pregnant humansto the variety of experimental manipulations whichmight be required to explore the many factors whichinfluence HPL synthesis and secretion. Therefore, aradioimmunoassay method has been developed for mon-key placental lactogen (MPL) to study various factorswhich influence its secretion in pregnancy. If the secre-tion of monkey and human placental lactogens is similar,it may be possible to create experimental situations inmonkeys which would lead to a better understandingof the physiological role of placental lactogen in bothhuman and simian pregnancies.
The existence of placental lactogen in monkeys has al-ready been established (1-4). MPL is more closelyrelated immunologically to human growth hormone(HGH) than HPL is (4) and MPLexerts a more po-tent growth-promoting effect than does HPL. A radio-immunoassay for MPLhas been described (5) in whichantiserum to human growth hormone and HPL-'I wereused to measure MPLin placental extracts but no studiesof MPL concentrations in pregnancy have been re-ported. We developed a homologous radioimmunoassayusing guinea pig antiserum to a highly purified prepara-tion of MPLand MPL-'I as a tracer. Using this assaywe have studied a variety of factors which influence thesecretion of MPL.
METHODSAntigens. MPLwas purified according to the method of
Shome and Friesen (6) by use of term rhesus monkeyplacentas obtained through the cooperation of a number ofprimate centers. The homogeneity of the MPL preparationhas been established by a variety of conventional methods.HGHand HPL were prepared in our laboratory accordingto methods already described (7). MGHwas obtained fromrhesus monkey pituitaries. Guinea pigs and rabbits wereimmunized weekly with 1 mg of each antigen along withcomplete Freund's adjuvant for a 4 wk period. 1 wk later abooster dose of the antigen was administered intravenouslyand the antiserum was obtained 10 days later.
Iodination. Iodination was carried out according to themethod of Hunter and Greenwood (8). The MPL-`'I was
2660 The Journal of Clinical Investigation Volume 50 1971

separated from free 'I on a Sephadex G-50 (1 cm x 10 cm)using 0.05 M barbital buffer. In general the efficiency ofiodination ranged between 40 and 60%. The specific activityof the MPL-'A'I was 150-300 ,Ci/pg. Iodination damage wasconsiderable if the reaction was allowed to proceed morethan 5-10 sec after the addition of chloramine T. The frac-tion containing MPL-'I from the G-50 column was com-bined with an equal volume of male monkey serum andfractionated on Sephadex G-100 (2 cm X 40 cm) using bar-bital buffer; 5-ml fractions were collected. The MPL-l'Iwhich was damaged and aggregated emerged in the voidvolume followed by MPL-1'I and occasionally a smallamount of 'I. Portions of MPL-'I in the second peak weretested with excess antibody to MPLand only those fractionswhere 80-90% of the MPL-'I was immunoprecipitable wereused for the assay.
Radioimmunoassay. A double antibody radioimmunoassaywas developed according to the method of Beck, Parker, andDaughaday (9) using guinea pig antiserum to MPL andrabbit anti-guinea pig gamma globulin fraction as the secondantibody.
All dilutions were made with 2.5% bovine serum albuminin barbital buffer. 50 al of samples (standards or serumsamples) were added to 0.5 ml of the diluent along with 0.1ml of MPL-1I and 0.1 ml of the first antibody which wasdiluted 1/20,000. After incubating the samples for 72 hr at4VC, 0.1 ml of a 1/200 dilution of normal guinea pig serumand 0.1 ml of the second antibody were added followed byan overnight incubation at 4VC. The samples were counted,centrifuged at 3000 g for 30 min, the supernatants decanted,and the precipitates counted in a Packard AutogammaCounter. Whenno MPLwas present in the standards 60-70%oof the MPL-'I added was precipitated in the assays. In thehybrid assays the same procedures were employed exceptthat 0.1 ml of a 1/6000 dilution of a guinea pig antiserum toHPL was used. HPL was measured by radioimmunoassay(9) and HCGwas measured by a similar procedure usinghuman luteinizing hormone as standard (potency 4 U/mgNIH LH standard).
Fig. 1 shows that similar amounts of MPL, HPL, andHGH in the hybrid assay do not inhibit the binding ofloewo o~~MPLstd
n 96 - HGHstdMGHstd84 ) HPLstd
E 720.
60
a48,3.-
24
o d do lt 260 260 3b0 3o 460 460 Mbng/m I
FIGURE 1 The effect of varying concentrations of unlabeledMPL (0-0), HGH(--*), MGH(A-/A), andHPL (X- X) on the binding of the HGH-'I by anti-HPL serum (1:6000 dilution). The per cent of HGH-mIprecipitated is 70%o in the absence of added hormone.
0 6-x * ^ HPLE \* HGH
4 ' MGH06i As* MPL06i 3 <
,2-aX -I
1I2ba 4b O6bb do CMPL ng/ml
FIGURE 2 The effect of increasing concentrations of unla-beled MPL( * ), MGH(A-A), HGH(0-O),and HPL (X--X) on the binding of MPL-l'I by anti-MPL serum (1:16,000 dilution). The per cent of MPL-I'lprecipitated is 67%o in the absence of any added hormone.
HGH-'I to anti-HPL serum in the same manner. Hence,neither HGHnor HPL can serve as an appropriate standardfor measuring MPLconcentrations. Moreover, the inhibitionof binding of the HGH-l'I tracer to anti-HPL antibody byMGHexceeded that of MPL. Therefore, one could not ex-pect to measure low levels of MPLin the presence of MGHusing anti-HPL antibody.
Fig. 2 shows a typical standard curve characterizing thehomologous radioimmunoassay. Neither HPL, HGH, norMGHcaused significant displacement of MPL-1'I binding toits antibody unless the concentration of the three exceeded100 ng/ml. On the other hand, as little as 10 ng/ml of un-labeled MPL caused significant displacement of MPL-I'lbinding. Fig. 3 shows- that serial dilutions of a monkeyplacental extract, pregnant monkey serum, and purified MPLinhibit MPL-3'3I binding equally, whereas, there is little
Volume of MPE or PMS(ml)4 l-3 W2
10.0E6I
0.
CL4 * HGH- MGH
aHPL \£3 PMS
0. eMPE*2* MPL
1.1-7 -6 .4~ A. ".2
Quantity of unlabeled/ tIPL, HPL,lHGH, or MGH
FIGURE 3 The effect of increasing concentrations of unla-beled MPL(- *), HPL (A-A), HGH(-U),MGH (-lJ ), monkey placental extract (MPE)(0-O), and pregnant monkey serum (PMS) (A-A),on the binding of MPL-'I by anti-MPL serum (1:20,000dilution). The per cent of MPL-'I precipitated is 65%o inthe absence of any added hormone.
Secretion of Monkey Placental Lactogen 2661

ZiE0coJ0
0-0
0to.0
40 80 12D 160 200 240Elution Volume (ml)
FIGuRE 4 Absorbance of proteins (0 0) and distribu-tion of MPL (0 O) after fractionation on a column ofSephadex G-100 (100 X 2 cm) of 2 ml of monkey placentalextract (a) and 2 ml of pregnant monkey serum (b) sepa-rately. The MPL in the monkey placental extract and inpregnant monkey serum has the same elution volume (140-200 ml). Note the presence of a small but significant peak ofcross-reacting material in the monkey serum found at anelution volume 80-120 ml. In each case, a portion of thefractions containing MPL was pooled and lyophilized forexamination upon disc-gel electrophoresis.
interference in the radioimmunoassay by the three otherantigens, MGH, HGH, and HPL.
To establish the specificity of the assay further we mea-sured the distribution of MPLafter gel filtration of a crudemonkey placental extract on Sephadex G-100 (Fig. 4 a). Thecrude monkey placental extract was derived from a muchlarger extract in which 1177 g of fresh frozen placenta wereextracted in 3800 ml of 0.1 M ammonium hydroxide at pH8.6. The pH was lowered to 5.8 with HCl; the precipitatewhich formed was removed by centrifugation. To the clearsupernatant was added solid ammonium sulfate to make it50% saturated, with respect to salt. The precipitate wascollected by centrifugation, dissolved in 0.1 M ammonium
bicarbonate, and dialyzed against distilled water and lyo-philized. An amount of powder equivalent to 0.6 g of frozenplacental tissue was dissolved in 2 ml of 0.1 M ammoniumbicarbonate and applied to Sephadex G-100. The cross-react-ing material had the same elution volume as HPL and puri-fied MPL (140-200 ml). Moreover, the distribution of MPLin fractions after gel filtration on Sephadex G-100 of 2 mlof pregnant monkey serum (Fig. 4b) was similar to thatobserved in extracts of monkey placenta. However, in addi-tion to the principal peak of MPL in the elution volume160-200 ml, a small but significant peak of cross-reactingmaterial was found in the elution volume 80-120 ml. Whetherthis material represented MPLwhich -was partially adsorbedto serum proteins or an aggregate of MPL is unclear fromthese studies.
Disc electrophoresis in acrylamide gel was performed ac-cording to the method of Davis (10) using Tris buffer (pH8.2) and tetramethylenediamine to facilitate polymerization ofthe large pore gel. The sample (50-100 Al) was applied tothe column in 0.4 M sucrose and electrophoresis was carriedout at 5 ma per tube. Replicate gels were either stained with
a_L Amido Schwartz in 6% acetic acid or cut into 2 mmseg-ments which were placed in 1 ml volumes of 0.05 M barbitonebuffer (pH 8.6) and frozen and thawed once. The immuno-reactive material eluted from each segment after 3 days at4VC was estimated by radioimmunoassay. The electrophoreticdistribution of MPLin a placental extract, pregnant monkeyserum extract, and MPLstandard were determined (Fig. 5).MPL in the placental extract has an Rr which is approx-imately similar to the Rr of the MPL standard. The MPL-like material in the serum extract was resolved into two
E--JI
280
240
20
120
80
40
. PLACENTAL EXTRACT
A-sMONKEY SERUM._. MPL (STANDARD)
RfFIGURE 5 Distribution of MPLafter acrylamide-gel electro-phoresis of monkey placental extract (0 O), pregnantmonkey serum (A A), and purified MPL ( * ).Most of the cross-reacting material in both the placentalextract and the serum have an R, which is approximatelysimilar to that of MPL standard (0.8). Note the two com-ponents in the serum extract, one with an R, of 0.65 and thesecond with an Rr of 0.8.
2662 C. Belanger, B. Shome, H. Friesen, and R. E. Myers

components, one with an R, of 0.65 and the second with anRt of 0.80. In this acrylamide-gel HPL has an R, of 0.60(not shown). It appeared, therefore, that most of the cross-reacting material in both the placental extract and the serumhad similar physical and chemical characteristics as the MPLstandard.
To determine the content of HPL, HCG, and MPL, freshfrozen human and monkey placental tissue at term were ex-tracted in 0.1 M ammonium bicarbonate at room temperature(20 ml/g). The extract was centrifuged and aliquots of theclear supernatant were analyzed for the three hormones. Inaddition, 20- to 30-mg fragments of placental tissue wereincubated in Krebs-Ringer bicarbonate buffer pH 7.4 with95% 02 and 5% CO2 at 370C as described (4). At 24 hr, theconcentration of the three hormones was determined in themedia.
Animals. Pregnant rhesus monkeys were studied under avariety of circumstances at various periods of gestation. Therhesus monkeys were from the breeding colony of the Labo-ratory of Perinatal Physiology in San Juan, Puerto Rico.All blood samples were obtained without anesthesia exceptduring the course of operative procedures. Samples wereobtained from peripheral veins either in the upper or thelower limb.
Operative procedure. Monkeys were operated upon underpentobarbital general anesthesia and sterile conditions. Thetechniques used in the ligation of the fetal umbilical vesselsbridging between the primary and secondary placental discsand supplying the secondary disc with fetal blood circulationhave been described earlier (11). Experimental abruption wasproduced by making a 2.5 cm incision in the uterine wall overthe site of attachment of the secondary disc. The glovedright forefinger was inserted between the uterine wall andthe secondary disc. Separation of this disc was produced bypressing the finger between the uterine wall and the disctotally interrupting the spiral arterial and draining venoussystems. Fetectomies were performed by making a 3.5-4.0 cmincision in the uterus, avoiding the placental discs. The fetuswas delivered through this incision, the umbilical cord doublytied and severed, and the fetus discarded. The uterine incisionwas carefully repaired leaving the amniotic fluid in situ. Theanimals were returned to their cages while still anesthetized.Depot form penicillin was used 600,000 U and crystallinepenicillin 600,000 U was dissolved in the amniotic fluid beforerepair to combat infection. Without exception after fetectomy,all placentas were retained until surgically delivered at latertimes as shown in Fig. 9.
MPL(jpg/nI)7654-32 _
--I'
4014 6 6 6 5 7
)60 80 0 f1l 140 160Period of Gestation(doys)
FIGURE 6 Serum MPL measured in normal monkeysthroughout pregnancy. The mean of MPLconcentration and1 SD for each 20 day period is plotted. The asterisk refers tothe number of samples in each 20 day period.
5qy
-%
SPO
5a0
500
A
6
..*.:I A gII A
0 A
2( 40 60 aTime (min)
LU0
FIGURE 7 41 blood samples were obtained from eight mon-keys between 0 and 90 min, after removal of placenta atterm. Serum MPL concentrations were determined andplotted on semilogarithmic paper against the time when theblood samples were drawn. The least square regression lineof y on x was calculated: y = 4.05165 + (-0.01439) x, andplotted. The mean half-life of MPLwas found to be 20 min.
RESULTS
The maternal serum MPL concentrations in monkeysthroughout pregnancy are presented in Fig. 6. At 50days of gestation, the mean concentration of MPL was2 jtg/ml increasing gradually during pregnancy to amean of 6 gg/ml at term. The MPL concentration atterm ranged from 4 /Ag/ml to 8 /Ag/ml. It should be notedthat these serum specimens were 2 yr old when MPLmeasurements were made. The mean MPLconcentrationin this group at term is somewhat lower than in severalrecent experiments in which the MPL concentration offresh serum samples from pregnant monkeys at termwere measured.
The maximum concentration of MPL in serum fromthe umbilical vein was 12 ng/ml in seven different mon-key fetuses at term delivery.
MPLconcentrations were measured in serum samplestaken from the femoral vein before and every 15 minafter delivery by caesarian section at term of eight nor-mal pregnant monkeys. The half-time disappearance rateof MPL was calculated by the regression analysismethod. The MPL concentration declines with a half-life of approximately 20 min during the first 90 minafter removal of the placenta (Fig. 7). Only a few sam-ples were obtained at much later periods (24 and 48 hr)at which time MPLwas still detectable, suggesting thatthe disappearance rate of MPL declines more slowly atsubsequent periods. However, considerably more datawould be required to validate this suggestion.
Surgical removal of the fetus (fetectomy) was carriedout on three pregnant monkeys each on the 80th, 120th,and 150th days of gestation. The placentas of all ani-mals were retained in vivo until surgically deliveredfrom 15 to 62 days later. In each group of three animals,
Secretion of Monkey Placental Lactogen 2663
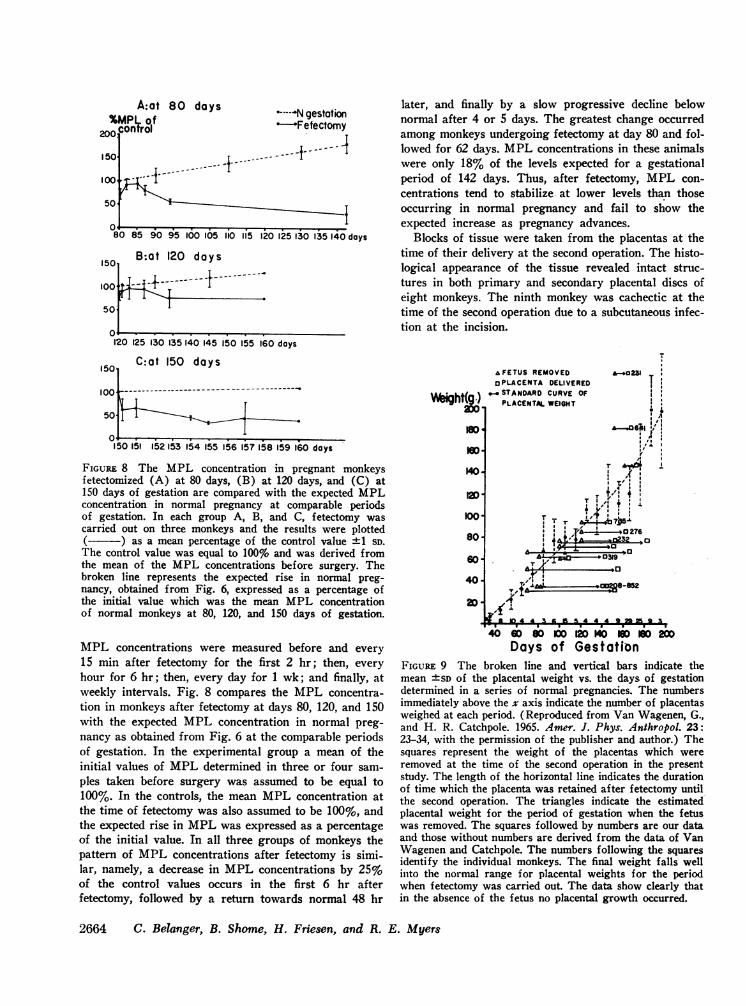
A:at 80%Mplw of
200 control
15
10
51
0o
0I00 L .-i---
80 85 90 95 100 105
1501 B:at 120 days
100
50
-- -- -- - f---i - -------
days *-----N gestation-Fetectomy
- 1-4----
1iO 115 120 125 130 135 140 days
later, and finally by a slow progressive decline belownormal after 4 or 5 days. The greatest change occurredamong monkeys undergoing fetectomy at day 80 and fol-lowed for 62 days. MPLconcentrations in these animalswere only 18% of the levels expected for a gestationalperiod of 142 days. Thus, after fetectomy, MPL con-centrations tend to stabilize at lower levels than thoseoccurring in normal pregnancy and fail to show theexpected increase as pregnancy advances.
Blocks of tissue were taken from the placentas at thetime of their delivery at the second operation. The histo-logical appearance of the tissue revealed intact struc-tures in both primary and secondary placental discs ofeight monkeys. The ninth monkey was cachectic at thetime of the second operation due to a subcutaneous infec-tion at the incision.
120 125 130 135 140 145 I50 155 160 days
C:at 150 days15I
1001-------_~-I- Tr
50 ........
150 151 152 153 54 155 156 15?758 159 160 days
FIGURE 8 The MPL concentration in pregnant monkeysfetectomized (A) at 80 days, (B) at 120 days, and (C) at150 days of gestation are compared with the expected MPLconcentration in normal pregnancy at comparable periodsof gestation. In each group A, B, and C, fetectomy wascarried out on three monkeys and the results were plotted( ) as a mean percentage of the control value ±1 SD.The control value was equal to 100% and was derived fromthe mean of the MPL concentrations before surgery. Thebroken line represents the expected rise in normal preg-nancy, obtained from Fig. 6, expressed as a percentage ofthe initial value which was the mean MPL concentrationof normal monkeys at 80, 120, and 150 days of gestation.
MPL concentrations were measured before and every
15 min after fetectomy for the first 2 hr; then, every
hour for 6 hr; then, every day for 1 wk; and finally, atweekly intervals. Fig. 8 compares the MPL concentra-tion in monkeys after fetectomy at days 80, 120, and 150with the expected MPL concentration in normal preg-nancy as obtained from Fig. 6 at the comparable periodsof gestation. In the experimental group a mean of theinitial values of MPLdetermined in three or four sam-
ples taken before surgery was assumed to be equal to100%. In the controls, the mean MPL concentration atthe time of fetectomy was also assumed to be 100%, andthe expected rise in MPLwas expressed as a percentageof the initial value. In all three groups of monkeys thepattern of MPL concentrations after fetectomy is simi-lar, namely, a decrease in MPL concentrations by 25%of the control values occurs in the first 6 hr afterfetectomy, followed by a return towards normal 48 hr
40 8s W Mo D K 2ODays of Gestation
FIGURE 9 The broken line and vertical bars indicate themean "SD of the placental weight vs. the days of gestationdetermined in a series of normal pregnancies. The numbersimmediately above the x axis indicate the number of placentasweighed at each period. (Reproduced from Van Wagenen, G.,and H. R. Catchpole. 1965. Amer. J. Phys. Anthropol. 23:23-34, with the permission of the publisher and author.) Thesquares represent the weight of the placentas which wereremoved at the time of the second operation in the presentstudy. The length of the horizontal line indicates the durationof time which the placenta was retained after fetectomy untilthe second operation. The triangles indicate the estimatedplacental weight for the period of gestation when the fetuswas removed. The squares followed by numbers are our dataand those without numbers are derived from the data of VanWagenen and Catchpole. The numbers following the squaresidentify the individual monkeys. The final weight falls wellinto the normal range for placental weights for the periodwhen fetectomy was carried out. The data show clearly thatin the absence of the fetus no placental growth occurred.
2664 C. Belanger, B. Shome, H. Friesen, and R. E. Myers
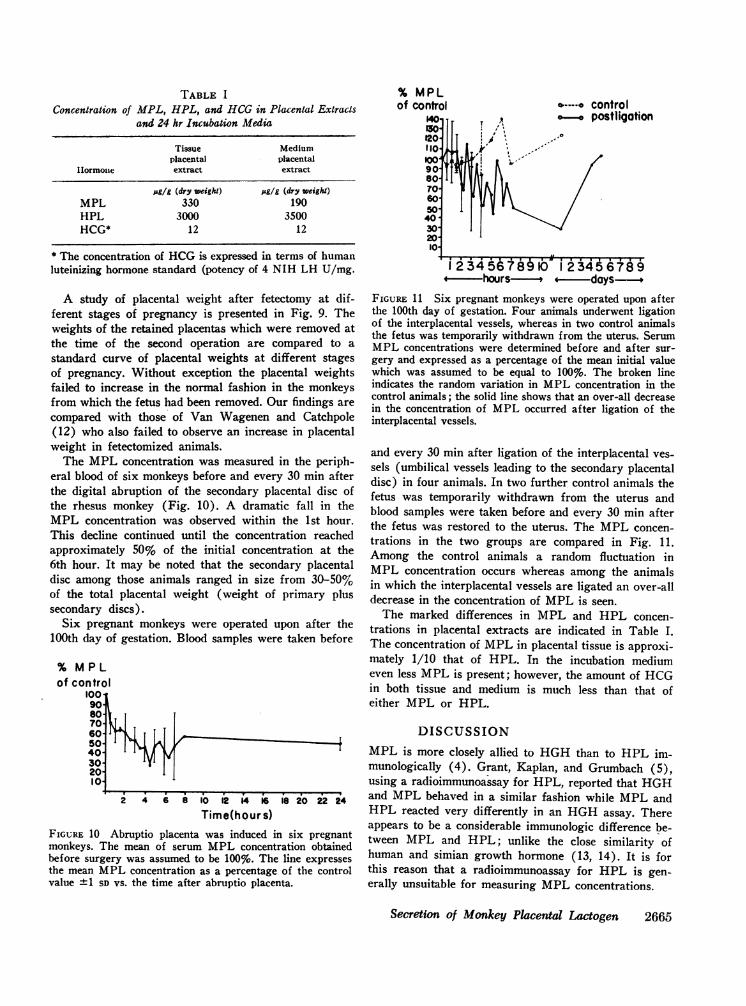
TABLE I
Concentration of MPL, HPL, and HCGin Placental Extractsand 24 hr Incubation Media
Tissue Mediumplacental placental
I1ormoue extract extract
pg/g (dry weight) ug/g (dry weight)MPL 330 190HPL 3000 3500HCG* 12 12
* The concentration of HCGis expressed in terms of humanluteinizing hormone standard (potency of 4 NIH LH U/mg.
A study of placental weight after fetectomy at dif-ferent stages of pregnancy is presented in Fig. 9. Theweights of the retained placentas which were removed atthe time of the second operation are compared to astandard curve of placental weights at different stagesof pregnancy. Without exception the placental weightsfailed to increase in the normal fashion in the monkeysfrom which the fetus had been removed. Our findings arecompared with those of Van Wagenen and Catchpole(12) who also failed to observe an increase in placentalweight in fetectomized animals.
The MPL concentration was measured in the periph-eral blood of six monkeys before and every 30 min afterthe digital abruption of the secondary placental disc ofthe rhesus monkey (Fig. 10). A dramatic fall in theMPL concentration was observed within the 1st hour.This decline continued until the concentration reachedapproximately 50% of the initial concentration at the6th hour. It may be noted that the secondary placentaldisc among those animals ranged in size from 30-50%of the total placental weight (weight of primary plussecondary discs).
Six pregnant monkeys were operated upon after the100th day of gestation. Blood samples were taken before
% MPLof contro
100190-sot80-7060-50-40-30-20-10-
2 4 6 8 10 12 14 16 iS 20 22 24
Time(hour s)FIGURE 10 Abruptio placenta was induced in six pregnantmonkeys. The mean of serum MPL concentration obtainedbefore surgery was assumed to be 100%. The line expressesthe mean MPLconcentration as a percentage of the controlvalue ±+1 SD vs. the time after abruptio placenta.
% MPLof control b-.-. control
0-. postligation
4 hours---- - - days -FIGURE 11 Six pregnant monkeys were operated upon afterthe 100th day of gestation. Four animals underwent ligationof the interplacental vessels, whereas in two control animalsthe fetus was temporarily withdrawn from the uterus. SerumMPL concentrations were determined before and after sur-gery and expressed as a percentage of the mean initial valuewhich was assumed to be equal to 100%. The broken lineindicates the random variation in MPL concentration in thecontrol animals; the solid line shows that an over-all decreasein the concentration of MPL occurred after ligation of theinterplacental vessels.
and every 30 min after ligation of the interplacental ves-sels (umbilical vessels leading to the secondary placentaldisc) in four animals. In two further control animals thefetus was temporarily withdrawn from the uterus andblood samples were taken before and every 30 min afterthe fetus was restored to the uterus. The MPL concen-trations in the two groups are compared in Fig. 11.Among the control animals a random fluctuation inMPL concentration occurs whereas among the animalsin which the interplacental vessels are ligated an over-alldecrease in the concentration of MPLis seen.
The marked differences in MPL and HPL concen-trations in placental extracts are indicated in Table I.The concentration of MPLin placental tissue is approxi-mately 1/10 that of HPL. In the incubation mediumeven less MPL is present; however, the amount of HCGin both tissue and medium is much less than that ofeither MPL or HPL.
DISCUSSIONMPL is more closely allied to HGHthan to HPL im-munologically (4). Grant, Kaplan, and Grumbach (5),using a radioimmunoassay for HPL, reported that HGHand MPLbehaved in a similar fashion while MPLandHPL reacted very differently in an HGHassay. Thereappears to be a considerable immunologic difference be-tween MPL and HPL; unlike the close similarity ofhuman and simian growth hormone (13, 14). It is forthis reason that a radioimmunoassay for HPL is gen-erally unsuitable for measuring MPL concentrations.
Secretion of Monkey Placental Lactogen 2665
II
Ie) .) --7
I
-

The maternal serum concentrations of MPL andHPL are very similar; the mean for HPL at term hasbeen reported to range from 3 to 10 /hg/ml (15-17). Inour study MPLconcentrations range from 3 to 7 Ag/ml.MPL and HPL fail to cross the placental barrier be-cause the concentration of each in cord blood is less than1% of the maternal MPL or HPL concentration. Theendogenous half-lives of MPLand HPL calculated afterthe removal of the placenta are also very similar. Kaplan,Gurpide, Sciarra, and Grumbach (18) reported a bi-phasic disappearance of HPL with the half-life of thefirst phase equal to 12 min compared to an estimated 20min for MPL. The second phase of HPL had a half-life of 75 min, whereas in the case of MPL insufficientdata are available for an accurate assessment of a sec-and phase. From the half-life, plasma concentration, andvolume of distribution of MPL and HPL we can esti-mate that the production rate of MPL is 0.36 g/daycompared to 1.0 g/day for HPL. These calculations as-sume that the volume of distribution of MPL is similarto HPL (10% of body weight), that the average weightof a monkey is 10 kg, and that turnover is monoexponen-tial. In view of the lower tissue content of MPL com-pared to HPL, turnover of the tissue pool of MPL isconsiderably greater than that of HPL. Because thesecretion of MPLand HPL is similar in many ways itappears likely that MPL is a very useful model forstudying factors which regulate the secretion of HPL.
An increasing number of reports have attempted todefine the usefulness of measuring HPL concentrationsin pregnancy as an index of placental function (17, 19,20). It was felt that monitoring HPL concentrationsmight assist the obstetrician to identify fetuses at risk.Our studies on MPL suggest that this is unlikely to bethe case. Even when the fetus is removed at variousstages of pregnancy, the immediate fall in MPL con-centrations is only modest, ranging from 10 to 20%.Only when the secondary placental disc is displaced asin the experiments where an experimental abruptioplacenta is induced does a drastic fall in MPL concen-tration occur. These data suggest that a major com-promise in placental function is required before a majorchange in MPLconcentration is observed. However, theconcentration of MPL seems to be influenced to a con-siderable degree by changes in maternal placental per-fusion as shown by the fact that when the maternalaorta was constricted, resulting in a decreased intervil-lous space perfusion, a prompt decrease in MPLconcen-tration in maternal blood was observed (unpublishedobservations).
The failure of MPL concentrations to increase in theabsence of the fetus poses several interesting questions.Was the lack of increase simply a reflection of the arrestin placental development or does the fetus exert a spe-
cific role in regulating MPLsecretion? Presently avail-able data suggest that placental growth is dependentupon normal fetal growth. In our studies no evidenceof a placental weight increase after fetectomy was foundin contrast to the study by Behrman, Parer, and de-Lannoy (21) which suggested that placental weight gaincontinued if the fetectomy was carried out after 120 daysof gestation. Rather, our results agree with the findingsof Van Wagenen and Catchpole (12) who reported thatplacental weight remained constant after fetectomy.
Histological and ultrastructural changes in rhesusmonkey placenta after interruption of the fetal placentalcirculation by fetectomy or interplacental vessel ligationhave been studied by Panigel and Myers (22). Theseauthors described an early reduction and disappearanceof the cytotrophoblast after interruption of the fetal vil-lous blood circulation, suggesting that a continued supplyof some humoral or cellular constituent from the develop-ing fetus is required for its continued maintenance. Onthe other hand, the presence of abundant ergastoplasmicformation (endoplasmic reticulum) in the syncytiotropho-blast for up to 2 months after fetectomy or fetal vesselligation indicates that the morphologic components re-quired for protein synthesis, including placental hor-mones, are present in the retained placenta. These histo-logical findings in monkey placentas correlate well withthe placental changes described after fetal death in thehuman (23).
The present studies suggest that the synthesis andsecretion of MPL and HPL are similar, and therefore,studies of MPLmay prove helpful in elucidating the roleof HPL in pregnancy.
ACKNOWLEDGMENTSWe acknowledge the assistance of the following PrimateCenters in collecting placental tissue for us: Tulane Univer-sity Delta Regional Primate Research Center, Covington,La.; Washington Regional Primate Research Center, Seattle,Wash.; National Centre for Primate Biology, Davis, Calif.;Laboratories Inc., Falls Church, Va.; Southwest Foundationfor Research and Education, San Antonio, Tex.; and YerkesRegional Primate Research Center, Atlanta, Ga. We alsoacknowledge the technical help of Mrs. Klara Holmwoodand Mrs. Judy Halmagyi and the secretarial assistance ofMiss Francine Dupuis. Mr. William Rodriguez and Mr.Estaban Monel-Tonens assisted at surgery and collectedblood samples in relation to the present study.
This research was supported by MRCMA-1862 and U. S.Public Health Service Grant HD-017/27. Dr. C. Belanger wasthe recipient of a fellowship from the Quebec Medical Re-search Council.
REFERENCES
1. Kaplan, S. L., and M. M. Grumbach. 1964. Studies ofa human and simian placental hormone with growthhormone-like and prolactin-like activities. J. Clin. Endo-crinol. 24: 80.
2666 C. Belanger, B. Shome, H. Friesen, and R. E. Myers

2. Josimovich, J. B., and D. H. Mintz. 1968. Biologicaland immunochemical studies on human placental lacto-gen. Ann. N. Y. Acad. Sci. 148: 488.
3. Friesen, H. G. 1968. Biosynthesis of placental proteinsand placental lactogen. Endocrinology. 83: 744.
4. Friesen, H. G., S. Suwa, and P. Pare. 1969. Synthesisand secretion of placental lactogen and other proteinsby the placenta. Recent Progr. Hormone Res. 25: 161.
5. Grant, D. B., S. L. Kaplan, and M. M. Grumbach.1970. Studies on a monkey placental protein with im-munochemical similarity to human growth hormoneand human chorionic somatomammotropin. Acta Endo-crinol. 63: 736.
6. Shome, B., and H. G. Friesen. 1971. Purification andcharacterization of monkey placental lactogen. Endo-crinology. 89: 631.
7. Friesen, H. G. 1965. Further purification and charac-terization of a placental protein with immunologicalsimilarity to human growth hormone. Nature (London).208: 1214.
8. Hunter, W. M., and F. C. Greenwood. 1962. Prepara-tion of iodine-131 labeled human growth hormone ofhigh specific activity. Nature (London). 194: 495.
9. Beck, P., M. L. Parker, and W. H. Daughaday. 1965.Radioimmunologic measurement of human placental lac-togen in plasma by a double antibody method duringnormal and diabetic pregnancies. J. Clin. Endocrinol.25: 1457.
10. Davis, B. J. 1964. Disc electrophoresis. II. Method andapplication to human serum proteins. Ann. N. Y. Acad.Sci. 121: 404.
11. Myers, R. E., D. Hill, A. B. Holt, R. E. Scott, E. D.Mellits, and D. B. Cheek. Fetal growth retardationproduced by experimental placental insufficiency in therhesus monkey. Body weight, organ size. In press. Biol-ogy of the Neonate.
12. Van Wagenen, G., and H. R. Catchpole. 1965. Growthof the fetus and placenta of the monkey (Macacus mu-latta). Amer. J. Phys. Anthropol. 23: 23.
13. Hunter, W. M., and F. C. Greenwood. 1964. A radio-immunoelectrophoretic assay for human growth hor-mone. Biochemn. J. 91: 43.
14. Abrams, R. L., and M. L. Parker, S. Blanco, S. Reich-lin, and W. H. Daughaday. 1966. Hypothalamic regu-lation of growth hormone secretion. Endocrinology.78: 605.
15. Samaan, N., S. C. C. Yen, H. Friesen, and 0. H.Pearson. 1966. Serum placental lactogen levels duringpregnancy and in trophoblastic disease. J. Clin. Endo-crinol. 26: 1303.
16. Kaplan, S. L., and M. M. Grumbach. 1965. Serumchorionic "growth hormone-prolactin" and serum pitui-tary growth hormone in mother and fetus at term. J.Clin. Endocrinol. 25: 1370.
17. Singer, W., P. Desjardins, and H. G. Friesen. 1970.Human placental lactogen: an index of placental func-tion. Obstet. Gynecol. 36: 222.
18. Kaplan, S. L., E. Gurpide, J. J. Sciarra, and M. M.Grumbach. 1968. Metabolic clearance rate and produc-tion rate of chorionic growth hormone-prolactin in latepregnancy. J. Clin. Endocrinol. 28: 1450.
19. Saxena, B. N., K. Emerson, Jr., and H. A. Selenkow.1969. Serum placental lactogen (HPL) levels as anindex of placental function. N. Engl. J. Med. 281: 225.
20. Genazzani, A. R., M. Aubert, M. Casoli, P. Fioretti,and J.-P. Felber. 1969. Use of human-placental-lacto-gen radioimmunoassay to predict outcome in cases ofthreatened abortion. Lancet. 2: 1385.
21. Behrman, R. E., J. T. Parer, and C. W. deLannoy, Jr.1967. Placental growth and the formation of amnioticfluid. Nature (London). 214: 678.
22. Panigel, M., and R. E. Myers. Histological and ultra-structural changes in rhesus monkey placenta followinginterruption of fetal placental circulation by fetectomyor interplacental umbilical vessel ligation. Acta Anat.
23. Fox, H. 1968. Morphological changes in the human pla-centa following fetal death. J. Obstet. Gynaecol. Brit.Commonw. 75: 839.
Secretion of Monkey Placental Lactogen 2667