studies of the endoplasmic reticulum and plasma
TRANSCRIPT

J. Cell Sci. 31, 165-178 (1978)Printed in Great Britain © Company of Biologists Limited igyS
STUDIES OF THE ENDOPLASMIC RETICULUM
AND PLASMA MEMBRANE-BOUND
RIBOSOMES IN ERYTHROPOIETIC CELLS
JOSEPH A. GRASSO, ALBERT L. SULLIVAN ANDSAI-CHUNG CHANDepartment of Anatomy, University of Connecticut Health Center, Farmington,Connecticut 06032, and the Department of Medicine, Boston UniversityMedical Center, Boston, Massachusetts 02118, U.S.A.
SUMMARY
Erythropoietic cells of 5 species, including man, contain endoplasmic reticulum present asindividual cisternae or tubules scattered throughout the cytoplasm of all stages except matureRBCs. The endoplasmic reticulum is mainly agranular but occurs frequently as a variant ofgranular ER which is characterized by an asymmetrical and irregular distribution of ribosomesalong one cytoplasmic face. In most cells, the endoplasmic reticulum occurs in close proximityto mitochondria or the plasma membrane, suggesting that the organelle may be involved infunctions related to these structures, e.g. haem biosynthesis. Endoplasmic reticulum is moreabundant in early than in late erythroid cells. Its exact role in RBC development is unclear.
Since endoplasmic reticulum could account for 'plasma membrane-bound ribosomes'reported in lysed reticulocytes, studies were performed which ruled out this possibility andwhich suggested that such ribosomes were an artifact of the lysing conditions. Hypotoniclysis in less than 20 vol. of magnesium-containing buffers yielded ghosts variably contaminatedby ribosomes and other structures. Lysis of reticulocytes in 20-30 vol. of magnesium-freebuffer or homogenization of whole cells or crude membrane fractions in hypotonic bufferremoved virtually all contaminating ribosomes from the purified membrane fraction.
INTRODUCTION
Plasma membrane fractions prepared by osmotic haemolysis of reticulocytescontain ribosomes which have been reported to support synthesis either of globin(Woodward et al. 1973; Brennessel & Goldstein, 1975) or of non-globin proteinspredominantly (Burka, Schreml & Kick, 1967; Schreml & Burka, 1968; Bulova &Burka, 1970). These results imply the existence of a ribosomal population which isbound to the plasma membrane (Burka et al. 1967; Schreml & Burka, 1968; Bulova &Burka, 1970) and which is functionally distinct from the more numerous 'free'ribosomes in the cytoplasm of erythroid cells (Burka et al. 1967; Schreml & Burka,1968; Bulova & Burka, 1970). However, morphological studies of intact erythroidcells have not revealed plasma membrane-bound ribosomes, creating a discrepancywhose structural basis has not been fully explored and which has served to stimulatethe present studies.
In initial studies, erythroid cells from several species were found consistently to
Please direct all correspondence to: Joseph A. Grasso, Ph.D., Department of Anatomy,University of Connecticut Health Center, Farmington, Connecticut 06032, U.S.A.

166 J. A. Grasso, A. L. Sullivan and S.-C. Chan
contain a variant form of granular endoplasmic reticulum which was topographicallyassociated with both mitochondria and the plasma membrane. Its structure anddistribution in the cytoplasm led us to consider the contribution of this organelleto the presence of membrane-bound ribosomes in plasma membrane fractions. Thepurpose of this report is to describe the endoplasmic reticulum in erythropoieticcells. In addition, these studies show that plasma membrane-bound ribosomesreported in lysed mammalian reticulocytes do not arise from co-sedimentation ofgranular endoplasmic reticulum with the plasma membrane fraction but may be acontaminant of the membrane fraction induced during the process of osmotic haemo-lysis.
MATERIALS AND METHODS
Electron microscopy
Human bone marrow samples were obtained from normal volunteers or from patients inthe course of studies concerned with sickle cell anaemia and various sideroblastic anaemias. Inaddition to human cells, erythropoietic elements from rat blood and bone marrow, cat bonemarrow, rabbit foetal liver, and newt blood have also been examined. Samples were fixedat room temperature in Karnovsky's fixative (Karnovsky, 1965) prepared in o-i M cacodylate(pH 6-9) or o-i M phosphate (pH 7-2^7-6) buffers. After postosmication in 1 % OsO4, tissueswere dehydrated in a graded series of ethanol or acetone at — 20 °C (50-100%) and embeddedin Epon 812. Sections were stained with uranyl acetate and lead citrate and examined in anAEI-6B or Philips 300 electron microscope.
In this report, the term 'early erythroid' stages designates pronormoblasts, basophilic,and early polychromatic normoblasts. ' Late erythroid' stages refers to late polychromatophilic-and acidophilic-normoblasts, and reticulocytes.
Preparation of reticulocyte membranes
Adult rats were made anaemic by 4 daily injections (06 ml) of 2 5 % acetylphenylhydrazinein mammalian saline. Three to 5 days after the last injection, the animals were bled by cardiacpuncture and blood cells were washed twice with heparinized saline. The buffy coat wasremoved during the washes. To enrich for reticulocytes, the red cell pellet was mixed byinversion with an equal volume of 8 % Ficol-Renografin® and layered over 10-15 m l of 14 %Ficol-Renografin® in a polyallomer or polycarbonate centrifuge tube. (® Trademark, E. R.Squibb & Sons, Inc., Princeton, New Jersey 08540, U.S.A.) The suspension was centrifugedat 25000 g for 10 min at 4 °C. The reticulocytes, layering in the interface between 8 and 14%solutions, were pipetted into saline and washed 3—4 times. Lysis was effected in 4—30 vol. ofvarious buffers, including 0-005 M Tris-HCl (pH 7 5 or 82), 0-005 M Tris-HCl containing0-0015 M MgClj, 0-005 M sodium phosphate buffer (pH 8), and RSB (Warner, Knopf & Rich,1963), with vigorous mixing at 4 °C or room temperature for 5 min. In some experiments,reticulocytes were homogenized in hypotonic buffer by means of 8—16 strokes in a Potter-Elvehjem homogenizer using a tight-fitting pestle. Membranes and debris were sedimentedand washed several times by centrifugation at 12000 rev/min for 10 min at 4 °C in a BeckmanJ-21B centrifuge. The resulting washed pellet {crude membrane fraction) was then processedfor electron microscopy or suspended in 10 ml of 70-74% (w/v) sucrose in lysis buffer andoverlaid by 5 ml of 54% sucrose, 8 ml of 48-5 % sucrose, 8 ml of 43 % sucrose, and 7-8 mlof buffer (Sirica, Goldblatt & McKelvy, 1975). In some experiments, the crude membranepellet was homogenized as above prior to loading on the sucrose gradient. The gradient wasspun at 66000 g for 1 h at 4 °C, using a Beckman SW27 rotor. The membrane fraction layeredbetween the 48-5 and 43 % sucrose layers was washed in several changes of lysis buffer bycentrifugation at 3000 rev/min for 15 min. The resulting pellet {purified membrane fraction)was processed for electron-microscopic visualization.

Membrane-bound ribosomes in erythroid cells 167
RESULTS
Appearance of endoplasmic reticulum in intact cells
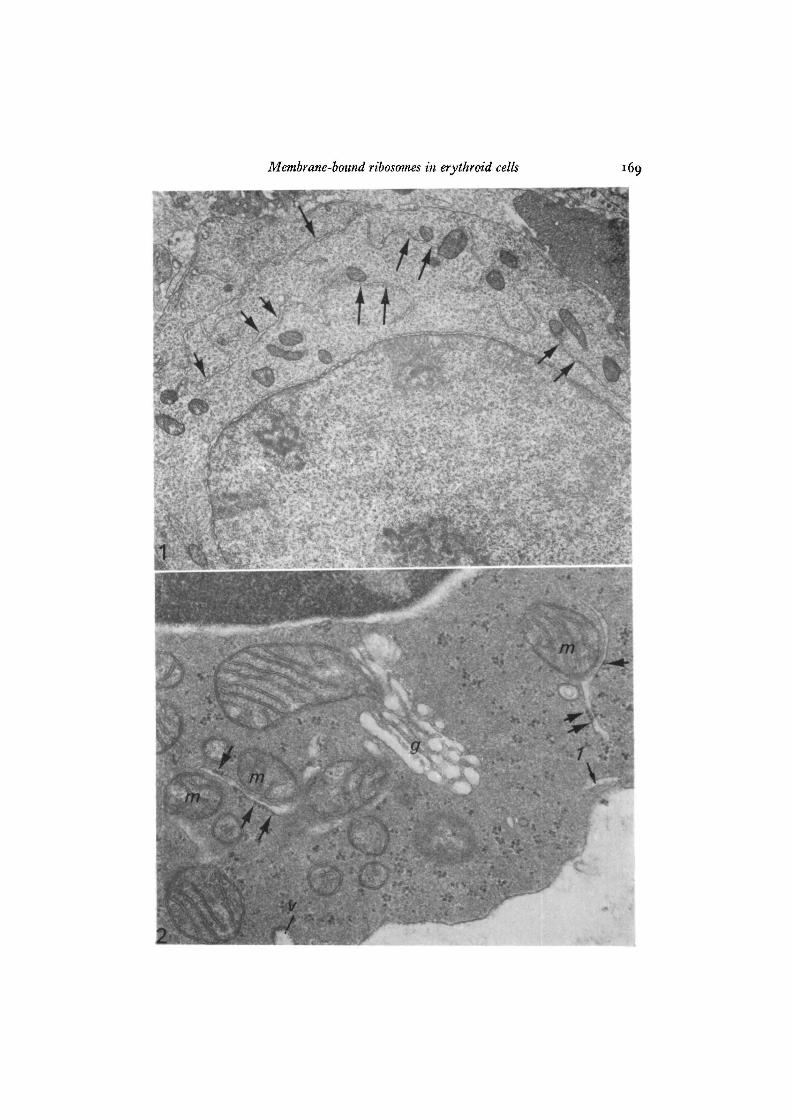
Endoplasmic reticulum (ER) was scattered as single elements in the cytoplasmof all erythroid cells except mature RBCs. It was more abundant in early erythroidstages where it occurred as vacuoles or tubules of varying length (average length inhuman cells, 0-82 /im), the longer profiles of which were tortuous and often branched(Fig. 1). In late erythroid cells (Fig. 2), ER was less abundant and visible as un-branched, relatively short tubules (average length, 0-32 /tm) or as vacuoles. Amongdifferent cells, even at the same developmental stage, ER was present in variableamounts and often was absent in sections of late erythroid cells.
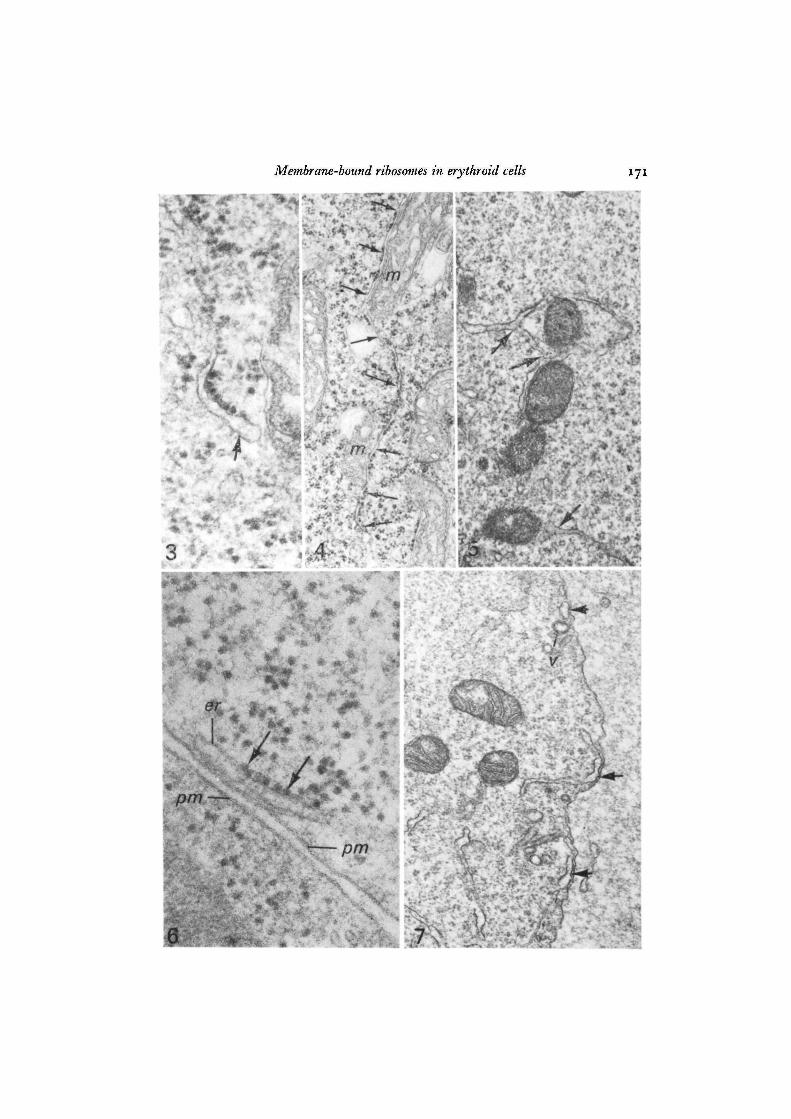
In all erythroid stages of every species, the ER consisted of mainly agranularelements and a variant form of granular ER characterized by asymmetrical andirregular distribution of membrane-bound ribosomes. Many elements of this varianttype exhibited patches of evenly spaced ribosomes attached for a variable distance toone surface of the tubular profile but absent from the opposing surface (Figs. 2, 3, 6).These asymmetrical elements resembled 'bifacial cisternae' seen elsewhere (Franke& Eckert, 1971) but differed in that the smooth membrane surface opposite theribosomes did not appear rigid or show increased electron density (Fig. 3) nor wasthere any indication that this surface was related to vesicle formation or to the Golgicomplex. Other forms of this variant type contained variable numbers of ribosomes,as few as 1-2 per profile, irregularly distributed as single ribosomes (Fig. 2), or inpatches (Fig. 4) along both surfaces of a tubular profile, with large stretches ofintervening membrane free of attached ribosomes.
While some ER profiles were distributed without any apparent relationship toother structures, a typical feature of the ER in erythroid cells was its topographicalassociation with mitochondria or the plasma membrane. Agranular and asymmetricalelements closely followed the surface contours of mitochondria (Figs. 2, 4, 5), oftenbranching to embrace them (Fig. 5). In many instances, the 2 organelles wereclearly separated by a distance of 3-5-8-0 nm (Figs. 2, 4). Occasionally, their separationwas obliterated or indistinct (Figs. 4, 5), probably as a result of their orientation withrespect to the plane of section. In associations between mitochondria and asym-metrical ER elements, the ribosomes were invariably attached to the membranesurface distal to the mitochondrion (Figs. 2, 4).
Tubular elements were often seen to extend toward and approach the cell surface(Figs. 1, 7, 8). Frequently, vacuolar dilatations or tubular profiles of agranular (Figs.2, 7) or asymmetrical ER (Fig. 6) were juxtaposed to the plasma membrane, fromwhich they were separated by a distance of 5-8 nm. These structures were similar insome respects to subsurface cisterns described in nerve and muscle cells (Rosenbluth,1962; Henkart, Landis & Reese, 1976). They were found in erythroid cells of allstages.

168 J. A. Grasso, A. L. Sullivan and S.-C. Chan
Morphology of membrane fractions
Intact erythroid cells and membrane fractions prepared in various ways wereexamined by electron microscopy in an effort to determine the origin of plasmamembrane-bound ribosomes reported in mammalian reticulocytes (Burka et al.1967; Bulova & Burka, 1970; Woodward et al. 1973; Morrison & Lingrel, 1975).High-magnification microscopy revealed no evidence of ribosomes attached to theplasma membrane of intact reticulocytes (Fig. 8) or of earlier erythroid cells (Figs.2, 6). While elements of endoplasmic reticulum containing ribosomes were oftenobserved near the plasma membrane of intact cells (Figs. 6, 8), the frequency andconcentration of these elements in the region of the plasmalemma was too low toaccount for the proportion of membrane-bound ribosomes (~ 20% of total) reportedin reticulocytes (Burka et al. 1967; Schreml & Burka, 1968; Bulova & Burka, 1970).These observations indicated that plasma membrane-bound ribosomes in membranepreparations had no visible structural correlate in intact cells, either as ribosomeslinked to the plasmalemma or as granular endoplasmic reticulum.
Lysis of reticulocytes in less than 20 vol. of hypotonic buffer or in buffers con-taining magnesium yielded membrane fractions or 'ghosts' whose ribosomal contentvaried considerably. Single ribosomes and ribosomal aggregates were found at ornear the plasma membrane (Fig. 10) and often in clumps at some distance from theplasmalemma as well (Fig. 9). Some ghosts were devoid of ribosomes and consistedof membrane only. Vacuoles, mitochondria, dense bodies, or endoplasmic reticulumwere occasionally visible (Fig. 9) but clearly the bulk of ribosomal material in thesemembrane fractions was derived from free ribosomes, not from granular ER. Amongdifferent experiments utilizing osmotic haemolysis under the conditions described,the proportion of ghosts containing ribosomal material ranged from ~ 10 to 20% topractically all of the ghosts examined by electron microscopy.
Repeated washing of these membrane pellets with hypotonic buffer or bufferslacking magnesium did not remove ribosomes or other structures contained in themembrane pellet. However, if homogenized subsequent to lysis, the resultant mem-brane fractions exhibited virtually no ribosomes (Fig. 11), although ribosomesassociated with endoplasmic reticulum were not removed by this procedure (Fig. 11 a).
Lysis of reticulocytes in 20-30 vol. of hypotonic buffer free of magnesium resultedin a significant reduction of ribosomal contamination and in fewer ghosts containing
Fig. 1. Human pronormoblast exhibiting tubules of endoplasmic reticulum (arrows).Note the tortuous outline of many of these profiles and their topographical associationwith mitochondria, x 12000.Fig. 2. Human polychromatophilic normoblast. Tubules of endoplasmic reticulumare juxtaposed to mitochondria (m). The asymmetrical and irregular pattern ofribosomal distribution (arrows) is evident. Note the location of ribosomes on thesurface distal to the mitochondria and the variable number of ribosomes in eachsegment. A tubular profile of endoplasmic reticulum is located close to the plasmamembrane (arrow J ) . A ferritin-containing micropinocytotic vesicle (v) and Golgielements (g) are indicated, x 40000.
Membrane-bound ribosomes in erythroid cells I f f
I \
\
/
ft
m
\

170 J. A. Grasso, A. L. Sullivan and S.-C. Chan
ribosomes. Virtually all ribosomes present in these preparations could be removedby homogenization of the membrane pellet in hypotonic buffer. Similarly, homo-genization of intact cells in magnesium-free hypotonic buffer produced membranesessentially free of ribosomes (Fig. 12).
DISCUSSION
The endoplasmic reticulum in erythropoietic cells of several species is identicalin its structure and distribution. It characteristically occurs as agranular elementsand as a variant form of granular endoplasmic reticulum to which ribosomes areattached in an asymmetrical and irregular pattern. In a cell type primarily engaged inintracellular accumulation of haemoglobin, the endoplasmic reticulum is neitherabundant nor as organized as in secretory cells but is distributed as single profileswhich are most often associated with mitochondria or the plasma membrane. Itslocalized distribution may account for its variable occurrence in sectioned cells andfor previous descriptions stressing its absence or presence only in trace amounts(Grasso, Swift & Ackerman, 1962; Orlic, 1970; Bessis, 1973) since the plane ofsection may exclude heterogeneously distributed elements. Endoplasmic reticulum isrelatively more abundant in early than in late erythroid cells but persists even inreticulocytes (Sullivan, Grasso & Weintraub, 1976; this report). Its consistentpresence and specific topographical distribution in erythroid cells of diverse speciessuggest that endoplasmic reticulum may play a significant role in events associatedwith RBC development.
The role of the endoplasmic reticulum in erythroid cells and the functional signifi-cance of its association with mitochondria and plasma membrane are unknown. Inview of the massive commitment of these cells to haemoglobin synthesis, it is possiblethat mitochondrial-endoplasmic reticulum complexes may represent a mechanismby which enzymes and other factors required for the intramitochondrial componentof haem biosynthesis may be supplied to mitochondria. In an analogous system inliver cells, complexes involving mitochondria and endoplasmic reticulum have been
Fig. 3. An asymmetrical element of endoplasmic reticulum (arrow) in a newt baso-philic erythroblast. A segment of evenly spaced ribosomes is attached to one surfaceof the tubule but ribosomes are absent from the opposing surface whose thicknessand density are similar to the remainder of the tubule, x 104000.
Fig. 4. Newt basophilic erythroblast exhibiting a long tubule of endoplasmic reticulum(arrows) closely associated with mitochondria (m). Note the asymmetrical and irregulardistribution of ribosomes and their polarity in the mitochondrial regions, x 29000.
Fig. 5. Endoplasmic reticulum in a human pronormoblast exhibiting branching(arrows) and an intimate association with mitochondria, x 30000.
Fig. 6. A subsurface profile of endoplasmic reticulum (er) in a rabbit polychromato-philic normoblast. Note the asymmetrical cluster of ribosomes (arrows) and theproximity of the profile to the plasma membrane (pm). x 90000.
Fig. 7. Subsurface profiles (arrows) of agranular endoplasmic reticulum in a humanpronormoblast. One of these profiles (centre) is continuous with a tubule extendingtowards the surface. A micropinocytotic vesicle (v) is shown, x 30000.
Membrane-bound ribosomes in erythroid cells 171
£#
pm
-T 'V
* > • • *
* ^ .
.'A 7

172 J. A. Grasso, A. L. Sullivan and S.-C. Chan
implicated in the synthesis and coupling of haemoproteins (Correia & Meyer, 1975)and in transfer of soluble mitochondrial proteins into mitochondria from their site ofsynthesis in the endoplasmic reticulum (Kadenbach, 1966). Hepatic <J-aminolevulinicacid synthetase, an enzyme required for an intramitochondrial step in haem synthesis,has been found outside mitochondria and interpreted to represent soluble enzymein transit from cytoplasm to mitochondria (Scholnick, Hammaker & Marver, 1969).More recently, polysomes containing mRNA coding for liver mitochondrial proteinshave been interpreted to originate from single cisternae of endoplasmic reticulumassociated with individual mitochondria (Shore & Tata, 1977a, b), complexes similarto those seen in erythroid cells. While a supportive function in haem synthesis isattractive with regard to mitochondrial-endoplasmic reticulum associations inerythroid cells, no experimental evidence has been gathered to substantiate thispossibility.
In some cells (Ericcson, 1969), including erythroblasts (Grasso, 1973), the originof autophagic vacuoles has been attributed to the endoplasmic reticulum. Whileautophagic degradation occurs in erythropoiesis, especially the late stages (Kent,Minick, Volini & Orfei, 1966), autophagy is infrequent in early erythroid cells wheremitochondrial-endoplasmic reticulum associations are numerous. Whether thisdiscrepancy implies a dual role for such associations cannot be ascertained. On theone hand, they may contribute to the haem-synthesizing function of these cells asdescribed but with the decline of haemoglobin synthesis and near-terminal matur-ation, some of these associations between mitochondria and endoplasmic reticulummay reflect formation of autophagic vacuoles.
Membrane-bound ribosomes have been reported in plasma membrane fractionsof lysed mammalian reticulocytcs (Burka, 1968; Burka et al. 1967; Woodward et al.1973; Brennessel & Goldstein, 1975; Fehlmann, Bellemare & Godin, 19750, i ;Morrison & Lingrel, 1975). Since previous morphological studies have reported noendoplasmic reticulum in intact reticulocytes, it has been concluded that these mem-brane-bound ribosomes are associated with the plasma membrane (Burka et al. 1967;Burka, 1968; Bulova & Burka, 1970) to which they are tightly bound since high salt(0-5 M KC1) fails to effect their release (Brennessel & Goldstein, 1975; Morrison &Lingrel, 1975). Functionally, 'plasma membrane-bound ribosomes' were reported tosynthesize predominantly non-globin proteins (Bulova & Burka, 1970), a feature which
Fig. 8. A representative section of a rabbit reticulocyte showing granular endo-plasmic reticulum (er) but no evidence of ribosomes attached to the plasma mem-brane (arrows), x 56coo.Fig. 9. Rat reticulocyte ghost prepared by lysis in 5 vol. of 0-005 M Tris-HCl buffercontaining 0-0015 M MgClj. Ribosomal material (arrows) occurring as single ribo-somes and massive aggregates is located not only near the plasma membrane but alsoin the interior of the ghost. Several mitochondria (m) are also present, x 56000.Fig. 10. Section from the same preparation as preceding figure showing severalreticulocyte ghosts with less contamination than in Fig. 9. Ribosomal material isaligned along the inner surface of the ghost membranes (arrows). A dense body (b)and a coated vesicle (u) are ahown. x 29000.

Membrane-bound ribosomes in erythroid cells
/ <'«*
10 /

174 3- A- Grasso, A. L. Sullivan and S.-C. Chan
implies their functional distinction from the more numerous free, or non-membrane-bound, ribosomes. However, subsequent studies have contradicted these findingsand report the bulk of newly synthesized protein associated with 'plasma membrane-bound ribosomes' to be globin (Woodward et al. 1973). Moreover, mRNA extractedfrom the membrane-bound ribosomal fraction resembled globin mRNA from thefree ribosomes in that it stimulated globin synthesis in a cell-free translational system(Brennessel & Goldstein, 1975; Morrison & Lingrel, 1975). Differences in relativequantity, in efficiency of stimulation of globin synthesis, and in specific activitieswere noted between the 2 respective mRNA fractions (Brennessel & Goldstein, 1975;Morrison & Lingrel, 1975) but certain differences may be attributed to partialdegradation as a result of the more vigorous extraction procedure required to releasemRNA from the membrane fraction (Brennessel & Goldstein, 1975).
Contrary to previous reports, the present studies show that reticulocytes, which areformed in response to phenylhydrazine-induced anaemia, contain elements of granularendoplasmic reticulum (Fig. 8; also see Figs. 8 and 9 in Sullivan et al. 1976). Whilethese elements account for part of the membrane-bound ribosomes in plasma mem-brane fractions, it is obvious from ultrastructural examination of osmotically lysedreticulocytes that most of the ribosomes in plasma membrane fractions are notderived from endoplasmic reticulum but represent single ribosomes and ribosomalaggregates whose presence, distribution, and relationship to the plasma membraneare highly variable. Thus, under lysing conditions similar to those employed inbiochemical studies, variable amounts of single ribosomes or large ribosomal aggre-gates are aligned along the inner surface of the plasma membrane or occur in non-membrane areas of the resultant reticulocyte residues (Figs. 9, 10). Significantly,there are no corresponding ribosomal entities, i.e. plasma membrane-bound ribo-somes or massive ribosomal aggregates, visible in intact reticulocytes or other erythroidcells.
The results of these studies suggest that the bulk, if not all, of the so-called plasmamembrane-bound ribosomes in lysed reticulocytes are artifactually generated by theprocess of osmotic haemolysis. This view is based upon several factors. First, electron-microscopic examination of the plasma membrane region in intact reticulocytes andother erythroid cells reveals no evidence of ribosomes attached to the plasma mem-brane. Second, plasma membrane-bound ribosomes contain globin mRNA (Bren-nessel & Goldstein, 1975; Morrison & Lingrel, 1975) and have been implicated inglobin synthesis (Woodward et al. 1973; Brennessel & Goldstein, 1975). Thus, theydo not appear to be functionally distinct from free or unbound ribosomes. Finally,
Fig. 11. Membrane preparation obtained by homogenization of an aliquot of thecrude membrane pellet represented in Figs. 9 and 10. The membrane fragments arevirtually free of ribosomes. x 50000.
Fig. 11 A. Profile of endoplasmic reticulum from the homogenized fraction depictedin Fig. 11. Ribosomal attachment is intact, x 42000.
Fig. 12. Rat reticulocyte membranes prepared by homogenization of intact cells in0-005 M Tris buffer. The membrane fragments are free of ribosomes. x 17000.

Membrane-bound ribosomes in erythroid cells
12C 1! I. 31

176 J. A. Grasso, A. L. Sullivan and S.-C. Chan
when reticulocytes are lysed in magnesium-containing buffers at cell: buffer ratios ofless than 1:20, the ribosomal material retained within the ghosts exhibits structuralfeatures and a distribution which is often unrelated topographically to the plasmamembrane. These features are more suggestive of ribosomal entrapment duringosmotic haemolysis, especially since a high degree of haemoglobin retention is visiblein the post-haemolytic crude membrane pellet. Correlation of these results with aconsideration of the mechanism of osmotic haemolysis suggests that lysis in less than20 vol. of hypotonic buffer and especially in magnesium-containing buffers is sub-optimal for preparation of membranes free of contamination.
Hypotonic haemolysis of erythrocytes (and presumably of reticulocytes) involvesthe transient appearance of holes in the plasma membrane through which the cellularcontents escape (Seeman, 1967; Seeman, Cheng & lies, 1973). These holes are 10-100nm wide, may or may not be confluent with one another, and are closed within 25-250 safter the onset of haemolysis by resealing of the membrane (Seeman, 1967; Seemanet al. 1973). It must be emphasized that these data were obtained with lysing mediafree of magnesium and that the presence of magnesium not only facilitates membraneresealing (Bodemann & Passow, 1972) but results in increased retention of haemo-globin (Clark & Shohet, 1976). While the size of the holes is sufficient to allow theescape of the bulk of the ribosomes, total release of ribosomes and of ribosomalaggregates may be hindered by the relatively short duration during which the holesremain open, a length of time which may be considerably less in the presence ofmagnesium. Exhaustive washing of ghosts lysed under suboptimal conditions doesnot significantly reduce the level of ribosomal contamination although release ofentrapped haemoglobin, whose diffusion radius is undoubtedly several times smallerthan that of ribosomes (Perutz et al. i960; Baker, 1967), occurs readily. Furthermore,it is not surprising that high salt washes fail to dissociate plasma membrane-boundribosomes since such conditions will not effect the reappearance of membrane holes inthe closed sacs or ghosts produced by osmotic lysis.
Thus, the size and duration of the membrane holes induced during hypotonichaemolysis may be the limiting factor in obtaining membranes free of contamination.Conditions which alter the characteristics of the transient membrane holes, e.g.lysis in presence of magnesium and at relatively high cellular concentrations, may beexpected to yield variable degrees of membrane contamination and entrapment ofcellular contents, as has been observed in the present studies. Ribosomal contami-nation can be considerably reduced by lysis in 20-30 vol. of buffer and virtuallyabolished by homogenization of whole cells or lysed ghosts in hypotonic buffer.Under the latter conditions, rupture and fragmentation of the membrane removesthe physical lestraints to ribosomal release imposed by a transiently permeable sac,but fails to dissociate true membrane-bound ribosomes such as occur in associationwith granular endoplasmic reticulum.
We cannot categorically exclude the possible existence of a small ribosomal pop-ulation whose function requires its transient attachment to the plasma membrane.Lodish and his colleagues (Lodish, 1973; Lodish & Desalu, 1973; Lodish & Small,1975) have demonstrated that reticulocyte membrane proteins are synthesized on

Membrane-bound ribosomes in erythroid cells 177
membrane-free ribosomes and have suggested that transmembrane proteins andexternal membrane proteins may require ribosomal attachment to the membrane forvectorial transport (Lodish & Small, 1975). Moreover, our data cannot explain theminor differences in protein composition of free and 'bound' reticulocyte ribosomes(Fehlmann et al. 1975 a, b). In view of the presence of endoplasmic reticulum inreticulocytes and the extent of ribosomal contamination obtained under suboptimallysis conditions, the physiological significance of membrane-bound ribosomes inerythroid cells requires re-examination under conditions which minimize the possi-bility of artifactual contamination.
These studies were supported by a grant (No. AM 19167) from the National Institute ofArthritis, Metabolic, and Digestive Diseases, National Institutes of Health. The initial studiesconcerned with ultrastructure of bone marrow in sickle cell anaemia were supported by agrant establishing the Boston Sickle Cell Center, Grant No. H L 15157 from the NationalHeart and Lung Institute, NIH. All studies involving human subjects were performed accord-ing to the guidelines of the National Institutes of Health with all procedures approved by theHuman Experimentation Committees of the involved institutions.
REFERENCES
BAKER, R. F. (1967). Entry of ferritin into human red cells during hypotonic haemolysis.Nature, Lond. 215, 424-425.
BESSIS, M. (1973). Living Blood Cells and their Ultrastructure, pp. 110-140. New York, Heidel-berg, Berlin: Springer.
BODEMANN, H. & PASSOW, H. (1972). Factors controlling the resealing of the membrane ofhuman erythrocyte ghosts after hypotonic hemolysis. J. Membrane Biol. 8, 1-26.
BRENNESSEL, B. A. & GOLDSTEIN, J. (1975). Globin mRNA from rabbit reticulocyte membrane-bound ribosomes. Biochim. biophys. Ada 378, 73-79.
BULOVA, S. I. & BURKA, E. R. (1970). Biosynthesis of nonglobin protein by membrane-boundribosomes in reticulocytes. J. biol. Chem. 245, 4907-4912.
BURKA, E. R. (1968). The distribution of RNA and ribosomes in reticulocytes. Biochim.biophys. Ada 166, 672-680.
BURKA, E. R., SCHREML, W. & KICK, C. J. (1967). Membrane-bound ribonucleic acid inmammalian erythroid cells. Biochimistry, N. Y. 6, 2840-2847.
CLARK, M. R. & SHOHET, S. B. (1976). Hybrid erythrocytes for membrane studies in sicklecell disease. Blood 47, 121-131.
CORREIA, M. & MEYER, V. A. (1975). Apocytochrome P-450: reconstitution of functionalcytochrome with hemin in vitro. Proc. natn. Acad. Set. U.S.A. 72, 400-404.
ERICCSON, J. L. E. (1969). Mechanism of cellular autophagy. In Lysosomes in Biology andPathology, vol. 2 (ed. J. T. Dingle & H. B. Fell), pp. 345-394. Amsterdam: North HollandPublishing.
FEHLMANN, M., BELLEMARE, G. & GODIN, C. (1975a). Free and membrane-bound ribosomes.II. Two-dimensional gel electrophoresis of proteins from free and membrane-bound rabbitreticulocyte ribosomes. Biochim. biophys. Ada 378, 119-124.
FEHLMANN, M., BELLEMARE, G. & GODIN, C. (19756). Free and membrane-bound rabbitreticulocyte ribosomes. Proteins from the large and the small subunits. FEBS Letters,Amsterdam 59, 8-12.
FRANKE, W. W. & ECKERT, W. A. (1971). Cytomembrane differentiation in a ciliate, Tetra-hymena pyriformis. II . Bifacial cisternae and tubular formations. Z. Zellforsch. mikrosk. Anat.122, 244-253.
GRAS6O, J. A. (1973). Erythropoiesis in the newt Triturus cristatus Laur. II . Characteristics ofthe erythropoietic process. J. Cell Set. 12, 491-523.
GRASSO, J. A., SWIFT, H. & ACKERMAN, G. A. (1962). Observations on the development oferythrocytes in mammalian fetal liver. J. Cell Biol. 14, 235-254.

178 J. A. Grasso, A. L. Sullivan and S.-C. Chan
HENKART, M., LANDIS, D. M. D. & REESE, T. S. (1976). Similarity of junctions between plasmamembranes and endoplasmic reticulum in muscle and neurons. J. Cell Biol. 70, 338-347.
KADENBACH, B. (1966). Synthesis of mitochondrial proteins. Demonstration of a transfer ofproteins from microsomes into mitochondria. Biochim. biophys. Ada 134, 430-442.
KARNOVSKY, M. (1965). A formaldehyde-glutaraldehyde fixation of high osmolality for use inelectron microscopy. J. Cell Biol. 27, 137A.
KENT, G., MINICK, O. T., VOLINI, F. I. & ORFEI, F. (1966). Autophagic vacuoles in humanred cells. Am. J. Path. 48, 831-858.
LODISH, H. F. (1973). Biosynthesis of reticulocyte membrane proteins by membrane-freepolyribosomes. Proc. natn. Acad. Set. U.S.A. 70, 1526-1530.
LODISH, H. F. & DESALU, O. (1973). Regulation of synthesis of non-globin proteins in cell-free extracts of rabbit reticulocytes. J. biol. Chetn. 248, 3520-3527.
LODISH, H. F. & SMALL, B. (1975). Membrane proteins synthesized by rabbit reticulocytes.J. Cell Biol. 65, 51-64.
MORRISON, M. R. & LINGREL, J. B. (1975). Characterization of globin messenger ribonucleicacids in membrane polysomes of mouse reticulocytes. J. biol. Chetn. 250, 848-852.
ORLIC, D. (1970). Ultrastrucrural analysis of erythropoiesis. In Regulation of Hematopoiesis,vol. 1 (ed. A. S. Gordon), pp. 271-296. New York: Appleton-Century-Crofts.
PERUTZ, M., ROSSMAN, M. G., CULLIS, A. F., MUIRHEAD, H., WILL, G. & NORTH, A. C. T.(1960). Structure of hemoglobin. A three-dimensional Fourier synthesis at 5-5 A resolutionobtained by X-ray analysis. Nature, Lond. 185, 416-429.
ROSENBLUTH, J. (1962). Subsurface cisterns and their relationship to the neuronal plasmamembrane. J. Cell Biol. 13, 405-421.
SCHOLNICK, P. L., HAMMAKER, L. E. & MARVER, H. S. (1969). Soluble hepatic(5-aminolevulinicacid synthetase: end-product inhibition of the partially purified enzyme. Proc. natn. Acad.Sci. U.S.A. 63, 65-70.
SCHREML, W. & BURKA, E. R. (1968). Properties of membrane-bound ribosomes in reticulocytes.J. biol. Chem. 243, 3573-3580.
SEEMAN, P. (1967). Transient holes in the erythrocyte membrane during hypotonic hemolysisand stable holes in the membrane after lysis by saponin and lysolecithin. J. Cell Biol. 32,55^7°-
SEEMAN, P., CHENG, D. & ILES, G. H. (1973). Structure of membrane holes in osmotic andsaponin hemolysis. J. Cell Biol. 56, 519-527.
SHORE, G. C. & TATA, J. R. (1977a). Two fractions of rough endoplasmic reticulum from ratliver. I. Recovery of rapidly sedimenting endoplasmic reticulum in association with mito-chondria. J. Cell Biol. 72, 714-725.
SHORE, G. C. & TATA, S. R. (19776). Two fractions of rough endoplasmic reticulum from ratliver. II. Cytoplasmic messenger RNA's which code for albumin and mitochondrial proteinsare distributed differently between the two fractions. J. Cell Biol. 72, 726-743.
SIRICA, A. E., GOLDBLATT, P. J. & MCKELVY, J. F. (1975). Isolation and partial characterizationof a bile canalicular plasma membrane fraction from normal and regenerating rat liver.J. biol. Chem. 250, 6464-6468.
SULLIVAN, A. L., GRASSO, J. A. & WEINTRAUB, L. R. (1976). Micropinocytosis of transferrin bydeveloping red cells: an electron-microscopic study utilizing ferritin-conjugated transferrinand ferritin-conjugated antibodies to transferrin. Blood 47, 133-143.
WARNER, J. R., KNOPF, P. M. & RICH, A. (1963). A multiple ribosomal structure in proteinsynthesis. Proc. natn. Acad. Sci. U.S.A. 49, 122-129.
WOODWARD, W. R., ADAMSON, S. D., MCQUEEN, H. M., LARSON, J. W., ESTVANIK, S. M.,WILAIRAT, P. & HERBERT, E. (1973). Globin synthesis on reticulocyte membrane-boundribosomes. J. biol. Chem. 248, 1556—1561.
{Received 19 October 1977)