studies of an anionic trypsinogen and its active enzyme from … · anionic and cationic...
TRANSCRIPT

THE Jou~rinr. OF BIOLOGICAL CHEMISTRY Vol. 246, No. 2, Issue of January 25, pp. 508616, 1971
Printed in U.S.A.
Studies of an Anionic Trypsinogen and Its Active Enzyme from Porcine Pancreas*
(Received for publication, May 4, 1970)
PETER VOYTEK~ AND ERLAND C. GJESSING
From the Departnzent of Biochemistry, University of Vermont College of Medicine, Burlington, Vermont 05401
SUMMARY
An anionic trypsinogen has been identified and isolated in a state of high purity from porcine pancreatic tissue and juice extracts. The anionic trypsinogen is present in large amounts and accounts for about half of the total potential tryptic activity. The kinetics of the conversion of the anionic trypsinogen to active enzyme is autocatalytic, yielding an anionic trypsin which is less anionic than the zymogen. K, values and specific activities of anionic trypsin toward esters and anilides of arginine and lysine were found to be very similar to those of cationic trypsin. Soybean trypsin inhibitor and diisopropyl phosphorofluoridate are potent inhibitors of anionic trypsin.
A previous publication (1) from this laboratory showed that extracts of acetone powders from porcine pancreas contained large amounts of a hitherto unreported endopeptidase having substrate specificities similar to trypsin. The endopeptidase and its zymogen precursor have a low isoelectric point as indi- cated by their elution from a DEAE-cellulose column with a de- creasing pH gradient and by their electrophoretic properties on starch and acrylamide gels. The enzyme was tentatively called protease A. Further investigations of protease A, based upon the hydrolysis of several synthetic trypsin substrates as well as proteins, have amply confirmed its trypsin-like nature. Because of their anionic behavior and the trypsin-like properties of the active enzyme, protease A and its zymogen are called anionic trypsin and anionic trypsinogen, respectively.
The anionic nature of this new enzyme is in marked contrast to the trypsins which have been isolated from porcine pancreas by several investigators in the past decade. Buck et al. (2), as well as Travis and Liener (3), have by free boundary electrophoresis determined the isoelectric point of porcine cationic trypsin to be 10.3 and 10.7, respectively. Charles et al. (4) have reported that porcine trypsinogen has an isoionic point of 7.5. The behavior of the isolated porcine trypsinogen and trypsin, on carboxy- methyl cellulose, as reported by the above investigators is in ac- cord with the basic or cationic properties of the zymogen and ac- tive enzyme.
* This investigation was supported by Grant AM-02024 from the United States Public Health Service.
$ Present address, Department of Pharmacology, Yale Univer- sity School of Medicine, New Haven, Connecticut 06510.
This communication describes the isolation of a highly purified preparation of the anionic trypsinogen, its conversion to active anionic trypsin, and some of its physical and kinetic properties as compared to cationic trypsinogen.
EXPERIMENTAL PROCEDURE
Materials
Crystallized soybean trypsin inhibitor was purchased from Worthington. Crystalline porcine trypsin was a gift from Dr. P. J. van Melle from Armour. DEAE-cellulose reagent grade (Lot 1623) and capacity 1.10 meq per g was purchased from the Brown Company. Cellex CM (carboxymethyl cellulose), having a capacity of 0.7 meq per g, was purchased from Bio-Rad. BAEE,’ BANA, tosyl-AME, ATEE, DFP, and poly-n-lysine (abt. 30n) were bought from Mann. Hippuryl-PL was obtained from Cycle Chemicals. Caproyl salicylate was prepared in this laboratory as previously described (1). lys-NA and protamine sulfate were supplied by Nutritional Biochemicals. Twice crystallized rabbit hemoglobin was purchased from Pentex. Poly-n-lysine (mol wt 105,000) was obtained from Pilot Chemi- cals.
Enzyme Assays
The rates of hydrolysis of the synthetic substrates BAEE and tosyl-AME, used for determining tryptic activity (5), ATEE, used for the determination of esteroproteolytic enzyme activity (I), and hippuryl-PL, used for the determination of carboxypep- tidase activity (6) were followed in a Radiometer pH-stat under nitrogen. The initial concentration of all substrates was 0.01 M
and the volume of the reaction mixture was 3 ml. CaCls was present at a concentration of 0.05 M. The titrant was 0.02 M
NaOH. Hydrolysis of tosyl-AME was carried out at pH 6.6 and the hydrolysis of ATEE and hippuryl-PL at pH 8.0. The hydrolysis of the trypsin substrates BANA and lys-NA was fol- lowed in a Gilford model 2000 multiple sample absorbance re- corder at 410 rnp according to Erlanger, Kokowski, and Cohen (7). The assays for BANA and lys-NA were carried out at pH 8.15 and 8.65, respectively. Stock solutions for both substrates were made up in dimethyl sulfoxide. Final concentrations in the reaction mixtures were 0.086 M Tris-HCl buffer, 0.043 M Cac&, 0.95 x lO+ M BANA, and 9.6% dimethyl sulfoxide, and 0.1 M
1 The abbreviations used are: BAEE, benzoyl-n-arginine ethyl ester; BANA, benzoyl-nn-arginine-p-nitroanilide; tosyl-AME, p-tosyl-L-arginine methyl ester; ATEE, N-acetyl-L-tyrosine ethyl ester; DFP, diisopropyl phosphorofluoridate; hippuryl-PL, hippuryl phenyllactate; lys-NA, L-lysine p-nitroanilide.
508
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 25, 1971 P. Voytelc and E. C. Gjessing 509
Tris-HCI buffer, 0.05 M CaC12, and 7% dimethyl sulfoxide for the TABLE I lys-NA assay. All activities are expressed as micromoles of sub- Analyses of porcine anionic trypsinogen and comparison strate hydrolyzed per min. Specific activity is expressed as units with porcine cationic trypsinogen of activity per mg of protein. Concentrations of anionic and _ _ I cationic trypsins were calculated from the extinction coefficients given in Table I. Hydrolysis of proteins was followed in a pH- stat under nitrogen. The amount of base (0.04 M NaOH) needed to maintain a constant pH of 8.0 was used as a measure of cleavage of peptide bonds. All assays were conducted at 25”.
Amino acid 2% sidueP
r
re
4mino aciC esidues pel 23,526 g of
protein
Literature values for cationic trypsinogen
Qualitative Determinations
Anionic trypsinogen, cationic trypsinogen, and soybean tryp- sin inhibitor could easily be detected in the chromatography elu- tion fractions by using the BANA spot tests as described by McConnell and Gjessing (1). Esteroproteolytic zymogen could also be determined qualitatively by using the caproyl salicylate spot test (1). These tests are capable of detecting enzyme ac- tivity of solutions containing less than 10 pg of enzyme per ml.
Activation of Trypsinogens
Anionic and cationic trypsinogen activity in the chromatog- raphy fractions was determined by taking 0.2~ml portions of each fraction and diluting to 1 ml. The activations were carried out at pH 8.0 in 0.1 M Tris-HCl buffer and 0.05 M CaC&. The acti- vator concentration (cationic trypsin) was 2 to 10 pg per ml. Aliquots of 0.05 ml were removed at different times and activity was determined by using the BANA or tosyl-AME assay. In all cases, corrections were made for hydrolysis due to activator.
Activation of Procarboxypeptidase and Esteroproteolytic Zyrrwgen
Solutions to be tested for potential carboxypeptidase and esteroproteolytic enzyme activities were adjusted to pH 8.0 with 0.2 M Tris-HCl buffer followed by the addition of 50 Mg of cat- ionic trypsin. The solutions were allowed to incubate at 5” for approximately 12 hours. Portions of 0.22 ml were then assayed with either ATEE or hippuryl-PL as substrate as described above. Under these conditions as little as 0.4 pg of either en- zyme could be detected.
Analysis of Anionic Typsinogen
Dry weight determinations for total nitrogen and the extinc- tion coefficient of anionic trypsinogen were carried out by trans- ferring portions containing 12 mg of protein to O&ml weighing bottles and removing most of the water over phosphorous pent- oxide and sodium hydroxide under reduced pressure. The tightly bound water was removed by heating the weighing bot- tles under reduced pressure over phosphorous pentoxide in an Abderhalden drying apparatus. Constant weights were obtained after 3 hours. The nitrogen content was determined by the micro-Kjeldahl method (8). The extinction coefficient (E&) of anionic trypsinogen was determined in a Beckmann spectro- photometer at 280 rnp and pH 3.0. Beer’s law was followed from zero to 1.0 absorbance unit.
Amino acid analysis of anionic trypsinogen was determined in a Beckmann-Spinco model 120 amino acid analyzer. Solutions containing 100 pg of protein were hydrolyzed according to the method of Moore and Stein (9).
The tryptophan content of the anionic trypsinogen was deter- mined according to Bencze and Schmid (10) in a Zeiss spectro- photometer.
Alanine ............ Arginine. .......... Aspartic acid. ..... Half -cystine ....... Glutamic acid ...... Glycine ............ Histidine .......... Isoleucine. ......... Leucine. ........... Lysine ............. Methionine ........ Phenylalanine ...... Proline. ........... Serine ............. Threonine. ......... Tyrosine. .......... Tryptophan. ....... Valine .............
Total. . . . . . Molecular weight. . Nitrogen (%) . E::, at 280mp. . . .
s i
,
UlOO g hroteilc
5.01 2.75
12.16 3.63 9.09 5.03 2.42 4.98 9.97 4.52 1.16 2.69 4.27 6.14 8.90 5.74 6.301 5.63
99.89
-
16.6 (17 4.1 (4) t4.8 (25 8.3 (8) 16.6 (17 30.7 (21 4.2 (4)
10.3 (10 20.7 (21 8.3 (8) 2.1 (2) 4.1 (4)
10.3 (10 16.6 (17 10.8 (21 8.3 (8) 8.0 (8)
12.4 (12
217 23,424
15.4 16.1
(3 Y (4) @)b 16 15 12 4 4 4
22 28 22 12 12 17 17 14 26 25 17
4 4 3 15 15 10 16 16 12 11 11 9
2 2 1 5 5 5
11 11 12 24 25 14 11 11 10 8 8 5 4 4 5
16 16 12
227 229 167 24,066 2 5,327 .9,288
16.! 13.!
- a Values in italics were determined after 48 hours of hydrolysis.
All other values were extrapolated to zero time. b Cationic trypsin data corrected for the release of an octapep-
tidePhe-Pro-Thr-(Asp)h-Lyswhich occurs during the activation process (4).
c Determined spectrophotometrically (10).
Ultracentrifugation Studies
The molecular weight of anionic trypsinogen was determined in the Spinco model E ultracentrifuge by the short column equi- librium technique described by Yphantis (11). The partial specific volume used for the determinations was calculated from the amino acid analysis data (Table I). In order to prevent the possibility of activation and autocatalysis of anionic trypsinogen during the centrifugation, the zymogen was first treated with in- hibitors. A 20-ml solution of purified anionic trypsinogen at a concentration of 2.5 mg per ml was treated with the irreversible inhibitor DFP at a concentration of 1 X 10m3 M at pH 7.0 for 40 min at room temperature. Soybean trypsin inhibitor was added to a final concentration of 0.02 mg per ml. The mixture was lyophilized and the dried powder was used for molecular weight determinations.
Polyacrylamicle Gel Ekctrophoresis
Discontinuous polyacrylamide gel electrophoresis was carried out in an apparatus described by Woodworth and Clark (12). The supporting medium was a 7.5% acrylamide gel with a thick- ness of & inch. A potential of 550 volts was applied for 2% hours at 5”. A Tris-glycine bufIer system was used (13). For electro-
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

510 Anionic Trypsinogen Vol. 246, No. 2
80 FRACTION NO.
160 240
FIG. 1. Protein elution profile of an acetone powder extract from a DEAE-cellulose column. Protein extract (100 ml) was added to the column at pH 7.5. The elution profile was obtained by the use of a series of sodium acetate buffers at 0.1 ,~/a. See “Experimental Procedure” for details. The pH of the fractions was determined to the nearest 0.05 pH unit at room temperature. The volume of fractions was 14 to 16 ml. Horizontal arrowa indi- cate fractions of anionic trypsinogen pooled. STI, soybean tryp- sin inhibitor.
phoretic comparison of the porcine cationic and anionic trypsins imidazole and taurine buffer systems were selected (14).
Isolation of Anionic Trypsinogen from Pancreatic Tissue Extracts
Preparation of Extra&-Sources of anionic trypsinogen were acetone powder extracts of porcine pancreas prepared for DEAE- cellulose chromatography according to McConnell and Gjessing
(1). Chromatography on DEAE-cellulose--The protein extract from
30 g of acetone powder in 100 ml of 0.01 M potassium phosphate buffer containing 0.1 M sodium acetate, 1 X 10B3 M DFP, and 1 mg per ml of soybean trypsin inhibitor at pH 7.5 was added to a DEAE-cellulose column (2 x 46 cm) containing 22 g of DEAE- cellulose (dry weight). The column was prepared as described by Peterson and Sober (15) and was equilibrated by first passing 200 ml of 1 .O M potassium phosphate buffer at pH 7.5 through the DEAE-cellulose followed by approximately 400 ml of water. The chromatography was carried out at 5-10” with flow rates of all additions maintained at 23 ml per hour. Proteins were eluted from the column (Fig. 1) by decreasing the pH of the elu- tion buffer, sodium acetate, at a constant ionic strength of 0.1:
900 ml at pH 5.5,400 ml at pH 5.3,lOOO ml of gradient from pH 5.3 to 4.9 made up of 500 ml of pH 5.3 buffer siphoned to 500 ml of pH 4.9 buffer, 430 ml at pH 4.5, and 400 ml at pH 3.3. Frac- tions containing anionic trypsinogen were pooled and concen- trated by precipitation with ammonium sulfate at 45% satura- tion at 0”. The precipitate was centrifuged at 20,000 x g for + hour, dissolved in 20 ml of cold water, dialyzed against 4 liters of ice water for 2 hours, and immediately lyophilized.
Chromatography on Hydroxylapatite-Approximately 35 mg of the lyophiiized preparation of the anionic trypsinogen from the DEAE-cellulose column were dissolved in 10 ml of cold 0.01 M
sodium phosphate buffer at pH 6.8. The solution was made 5 X 10m3 M with DFP and kept at room temperature for 10 mm. The protein solution was filtered in order to remove a trace amount of insoluble material before adding it to a hydroxylapatite column.
0.046M O.IOOM
FRACTION NUMBER
FIG. 2. Protein elution profile of an anionic trypsinogen prepa- ration from a hydroxylapatite column at pH 6.8. The anionic trypsinogen was a preparation obtained from the DEAE-cellulose column and was equivalent to an extract from 4 to 5 g of the ace- tone powder. Vertical arrowa indicate where changes in elution buffers (sodium phosphate) were applied. Horizontal arrows indicate fractions pooled. Fractions contained 6 ml.
The packed column (0.9 x 30 cm) contained 6 g of hydroxyl- apatite (dry weight) which had been prepared according to Levin (16). The chromatography of the proteins was carried out at 5” with the flow rate of all additions at 10 ml per hour. After the protein solution had entered the column, a 30-ml linear gradient from 0.01 to 0.022 M sodium phosphate buffer at pH 6.8 was started. After the 30-ml gradient the eluate was maintained at 0.022 M until the major protein component, the anionic trypsino- gen, had been eluted. The molarity of the eluate was then raised to 0.046 M which displaced any esteroproteolytic zymogen present in the preparation. Increasing the molarity to 0.1 dis- placed an additional inactive protein peak (Fig. 2). Fractions containing the anionic trypsinogen were pooled, brought to 0”, and immediately adjusted to pH 2.5 with 0.1 N HCI. The mix- ture was dialyzed against 4 liters of 0.001 N HCl at 0” for 30 hours with six changes of fluid. The anionic trypsinogen solution could be stored at -18” for several months without losing any potential activity.
Isolation of Anionic Trypsinogen from Pancreatic Juice
Preparation of Extract-A preparation of porcine pancreatic juice proteins was obtained from Brookhaven Laboratories through the courtesy of Dr. V. N. Reinhold. The pancreatic juice of a female Yorkshire pig was collected over a period 25 days. Each day’s collection was made 1 X 1O-3 M with respect to DFP and was lyophilized. The lyophilized preparations were suspended in 2 liters of water and dialyzed against 100 liters con- taining 1 x lOpa M DFP. The protein solution was lyophiliied. The lyophilized powder was dissolved in 1 liter of water, and then 1 liter of saturated ammonium sulfate was added slowly over a period of 5 hours. After the addition of ammonium sulfate the mixture was stirred for 8 hours at 3”. The mixture was centri- fuged at 0” for 40 min at 23,000 x g and the precipitate consisted of 95% of all of the protein in the pancreatic juice and contains essentially all of the potential tryptic activity.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
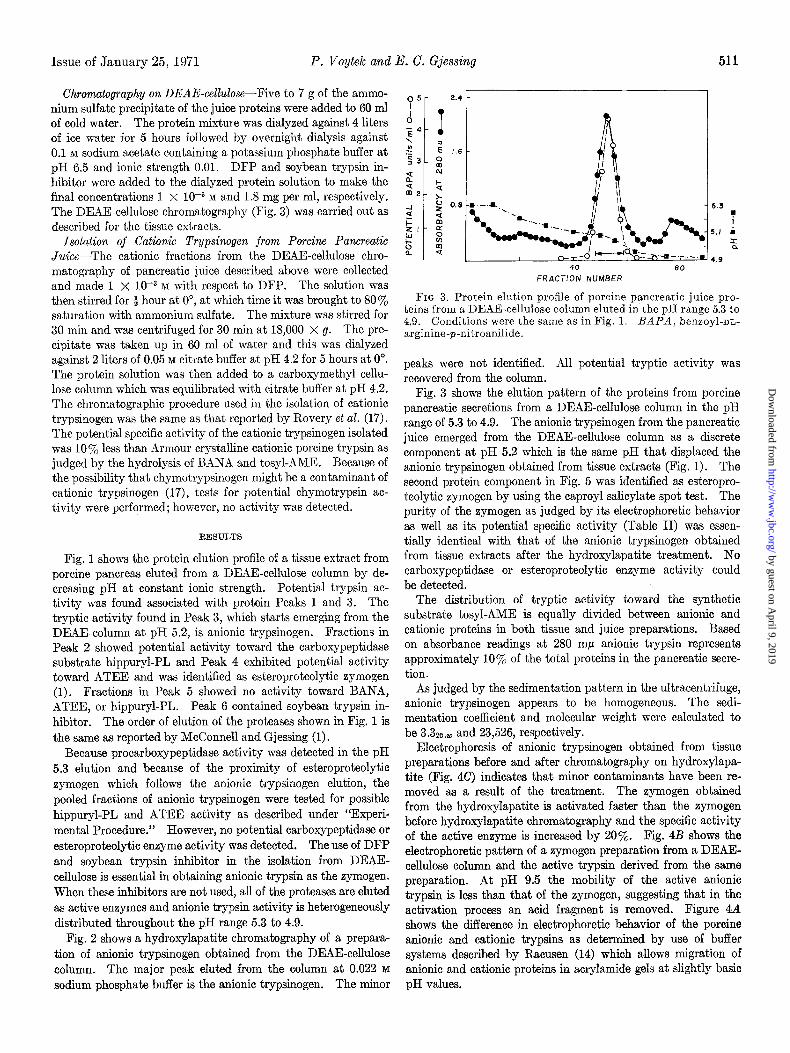
Issue of January 25, 1971 P. Voytek and E. C. Gjessing 511
Chromatography on DEAE-cellulose--Five to 7 g of the ammo- nium sulfate precipitate of the juice proteins were added to 60 ml of cold water. The protein mixture was dialyzed against 4 liters of ice water for 5 hours followed by overnight dialysis against 0.1 M sodium acetate containing a potassium phosphate buffer at pH 6.5 and ionic strength 0.01. DFP and soybean trypsin in- hibitor were added to the dialyzed protein solution to make the final concentrations 1 X lop3 M and 1.8 mg per ml, respectively. The DEAE-cellulose chromatography (Fig. 3) was carried out as described for the tissue extracts.
Isolation of Cationic Trypsinogen from Porcine Pancreatic Juice-The cationic fractions from the DEAE-cellulose chro- matography of pancreatic juice described above were collected and made 1 X 10d3 M with respect to DFP. The solution was then stirred for 4 hour at O”, at which time it was brought to 80% saturation with ammonium sulfate. The mixture was stirred for 30 min and was centrifuged for 30 min at 18,000 X g. The pre- cipitate was taken up in 60 ml of water and this was dialyzed against 2 liters of 0.05 M citrate buffer at pH 4.2 for 5 hours at 0”. The protein solution was then added to a carboxymethyl cellu- lose column which was equilibrated with citrate buffer at pH 4.2. The chromatographic procedure used in the isolation of cationic trypsinogen was the same as that reported by Rovery et al. (17). The potential specific activity of the cationic trypsinogen isolated was 10% less than Armour crystalline cationic porcine trypsin as judged by the hydrolysis of BANA and tosyl-AME. Because of the possibility that chymotrypsinogen might be a contaminant of cationic trypsinogen (17), tests for potential chymotrypsin ac- tivity were performed; however, no activity was detected.
RESULTS
Fig. 1 shows the protein elution profile of a tissue extract from porcine pancreas eluted from a DEAE-cellulose column by de- creasing pH at constant ionic strength. Potential trypsin ac- tivity was found associated with protein Peaks 1 and 3. The tryptic activity found in Peak 3, which starts emerging from the DEAE-column at pH 5.2, is anionic trypsinogen. Fractions in Peak 2 showed potential activity toward the carboxypeptidase substrate hippuryl-PL and Peak 4 exhibited potential activity toward ATEE and was identitied as esteroproteolytic zymogen (1). Fractions in Peak 5 showed no activity toward BANA, ATEE, or hippuryl-PL. Peak 6 contained soybean trypsin in- hibitor. The order of elution of the proteases shown in Fig. 1 is the same as reported by McConnell and Gjessing (1).
Because procarboxypeptidase activity was detected in the pH 5.3 elution and because of the proximity of esteroproteolytic zymogen which follows the anionic trypsinogen elution, the pooled fractions of anionic trypsinogen were tested for possible hippuryl-PL and ATEE activity as described under “Experi- mental Procedure.” However, no potential carboxypeptidase or esteroproteolytic enzyme activity was detected. The use of DFP and soybean trypsin inhibitor in the isolation from DEAE- cellulose is essential in obtaining anionic trypsin as the zymogen. When these inhibitors are not used, all of the proteases are eluted as active enzymes and anionic trypsin activity is heterogeneously distributed throughout the pH range 5.3 to 4.9.
Fig, 2 shows a hydroxylapatite chromatography of a prepara- tion of anionic trypsinogen obtained from the DEAE-cellulose column. The major peak eluted from the column at 0.022 M
sodium phosphate buffer is the anionic trypsinogen. The minor
2.4 t
0.8 5.3 ? I
5.1 i I a
4.9
FRACTION NUMBER
FIG 3. Protein elution profile of porcine pancreatic juice pro- teins from a DEAE-cellulose column eluted in the DH range 5.3 to 4.9. Conditions were the same as in Fig. 1. BAFA, benxoyl-nn- arginine-p-nitroanilide.
peaks were not identified. All potential tryptic activity was recovered from the column.
Fig. 3 shows the elution pattern of the proteins from porcine pancreatic secretions from a DEAE-cellulose column in the pH range of 5.3 to 4.9. The anionic trypsinogen from the pancreatic juice emerged from the DEAE-cellulose column as a discrete component at pH 5.2 which is the same pH that displaced the anionic trypsinogen obtained from tissue extracts (Fig. 1). The second protein component in Fig. 5 was identified as esteropro- teolytic zymogen by using the caproyl salicylate spot test. The purity of the zymogen as judged by its electrophoretic behavior as well as its potential specific activity (Table II) was essen- tially identical with that of the anionic trypsinogen obtained from tissue extracts after the hydroxylapatite treatment. No carboxypeptidase or esteroproteolytic enzyme activity could be detected.
The distribution of tryptic activity toward the synthetic substrate tosyl-AME is equally divided between anionic and cationic proteins in both tissue and juice preparations. Based on absorbance readings at 280 rnp anionic trypsin represents approximately 10% of the total proteins in the pancreatic secre- tion.
As judged by the sedimentation pattern in the ultracentrifuge, anionic trypsinogen appears to be homogeneous. The sedi- mentation coefficient and molecular weight were calculated to be 3.3~~,~ and 23,526, respectively.
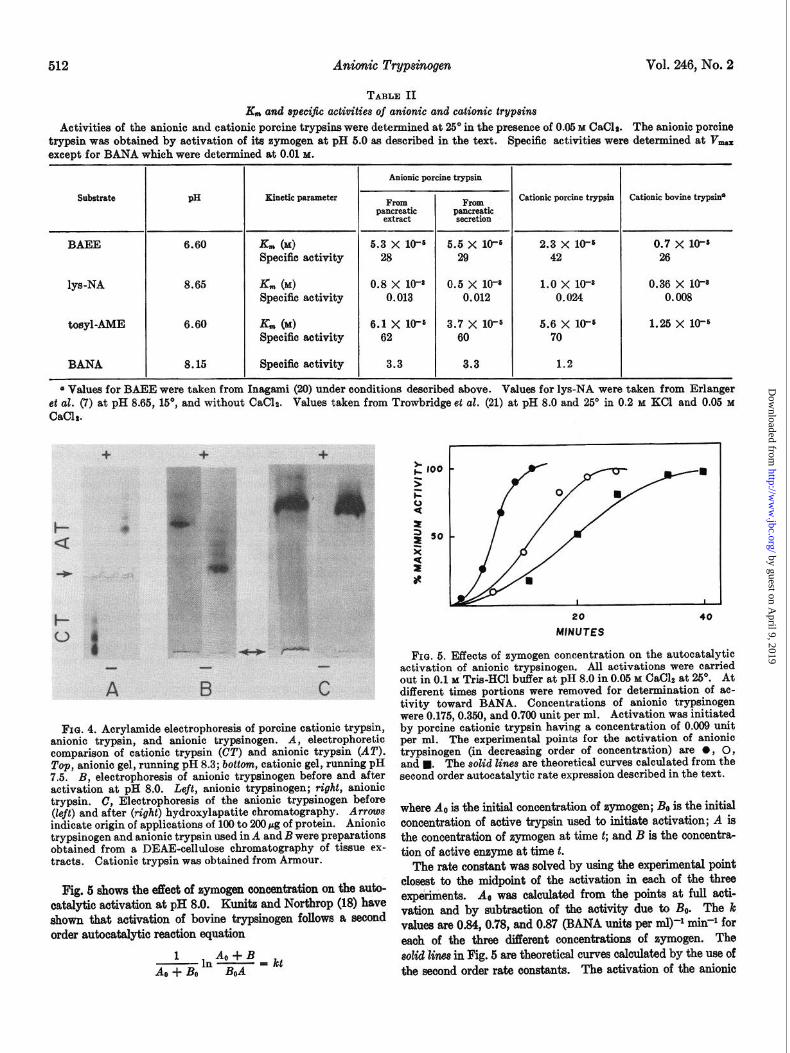
Electrophoresis of anionic trypsinogen obtained from tissue preparations before and after chromatography on hydroxylapa- tite (Fig. 4C) indicates that minor contaminants have been re- moved as a result of the treatment. The zymogen obtained from the hydroxylapatite is activated faster than the zymogen before hydroxylapatite chromatography and the specific activity of the active enzyme is increased by 20%. Fig. 4B shows the electrophoretic pattern of a zymogen preparation from a DEAE- cellulose column and the active trypsin derived from the same preparation. At pH 9.5 the mobility of the active anionic trypsin is less than that of the zymogen, suggesting that in the activation process an acid fragment is removed. Figure 4A shows the difference in electrophoretic behavior of the porcine anionic and cationic trypsins as determined by use of buffer systems described by Racusen (14) which allows migration of anionic and cationic proteins in acrylamide gels at slightly basic pH values.
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
512 Anionic Trgpsinogen Vol. 246, No. 2
TABLE II Km and specific activities of anionic and cationic trypsins
Activities of the anionic and cationic porcine trypsins were determined at 25’ in the presence of 0.05 Y CaClz. The anionic porcine trypsin was obtained by activation of its zymogen at pH 5.0 as described in the text. Specific activities were determined at I’,., exceptfor BANA which were determined at 0.01 M.
Substrate
BAEE
lys-NA
tosyl-AME
BANA
PH
6.60
8.65
6.60
8.15
Km (4 Specific activity
&I (4 Specific activity
Kn bf) Specific aotivity
Specific activity
Anionic porcine trypsin
FITJIll pancreatic
extract
5.3 x 10-s 28
0.8 x lo-” 0.013
6.1 X W6 62
3.3
FKUIl pancreatic secretion
5.5 x lo-5 29
0.5 x 10-a 0.012
3.7 x 10-z 60
3.3
Cationic porcine trypin
2.3 X lo-” 42
1.0 x 10-a 0.024
5.6 X lo-’ 70
1.2
Cationic bovine trypsi+
0.7 x 10-z 26
0.36 X 10-a 0.008
1.25 X 10-t
0 Values for BAIZE were taken from Inagami (20) under conditions described above. Values for lys-NA were taken from Erlanger et al. (7) at pH 8.65, 15’, and without CaClz. Values taken from Trowbridge et al. (21) at pH 8.0 and 25’ in 0.2 M KC1 and 0.05 M CaClt.
FIG. 4. Acrylamide electrophoresis of porcine cationic trypsin, anionic trypsin, and anionic trypsinogen. A, electrophoretic comparison of cationic trypsin (CT) and anionic trypsin (AT). Top, anionic gel, running pH 8.3; bottom, cationic gel, running pH 7.5. B, electrophoresis of anionic trypsinogen before and after activation at pH 8.0. Left, anionic trypsinogen; right, anionic trypsin. C, Electrophoresis of the anionic trypsinogen before (left) and after (tight) hydroxylapatite chromatography. Arrows indicate origin of applications of 100 to 200 pg of protein. Anionic trypsinogen and anionic trypsin used in A and B were preparations obtained from a DEAE-cellulose chromatography of tissue ex- tracts Cationic trypsin was obtained from Armour.
Fig. 5 shows the e&t of 5ymogen concentration on the auto- catalytic activation at pH 8.0. Ku&z and Northrop (18) have shown that activation of bovine trypsinogen follows a second order autocatalytic reaction equation
1 Ao + Bo
ln A* = kt BOA
20 40 MINUTES
FIU. 5. Effects of zymogen concentration on the autocatalytic activation of anionic trypsinogen. Ail activations were carried out in 0.1 aa’Tris-HCI buffer at pH 8.0 in 0.05 M CaClz at 25’. At different times portions were removed for determination of ac- tivity toward BANA. Concentrations of anionic trypsinogen were 0.175,0.350, and 0.760 unit per ml. Activation waz initiated by porcine cationic trypsin having a concentration of 0.003 unit per ml. The experimental points for the activation of anionic trypsinogen (in decreasing order of concentration) are 0, 0, and n . The solid lines are theoretical curves calculated from the second order autocatalytic rate expression described in the text.
where A0 is the initial concentration of symogen; Bo is the initial concentration of active trypsin used to initiate activation; A is the concentration of zymogen at time t; and B is the concentra- tion of active ensyme at time t.
The rate oonstant wae solved by using the experimental point closest to the midpoint of the activation in each of the three experiments. A. was calculated from the points at full acti- vation and by subtraction of the activity due to Bo. The k values are 0.84,0.78, and 0.87 (BANA units per ml)* min-l for each of the three Merent concentrations of symogen. The solid lines in Fig. 6 are theoretical curves calculated by the use of the second order rate constants. The activation of the anionic
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 25, 1971 P. Voytek and E. C. Gjessing
trypsinogen obeys the second order autocatalytic reaction mecha- nism.
Fig. 6A shows that anionic trypsinogen is activated auto- catalytically at pH 8.0 and 5.0. At pH 8.0 in the presence of CaC$, activation occurs equally well when either anionic or cationic trypsin is used to initiate activation. However, acti- vation at pH 8.0 did not occur within a 40-min incubation time when either activator or CaClz was omitted from the assay mix- ture. Activation at pH 5.0 in the presence of CaC% with anionic trypsin as initiator of activation occurs as rapidly as the acti- vations carried out at pH 8.0. At pH 5.0 and with CaClz present, activation takes place more slowly when cationic trypsin rather than anionic trypsin is the initiator. At pH 5.0 activation occurs more slowly in the absence of CaC12; however, the reac- tion is faster when anionic trypsin is used to initiate activation.
A comparable study was made of the autocatalytic activation of cationic trypsinogen in the presence of CaCl* (Fig. 6B). Cationic trypsinogen is activated at pH 8.0 more quickly when anionic t.rypsin is used to initiate activation as compared to cationic trypsin. Activation of cationic trypsinogen by anionic trypsin did not follow the typical second order autocatalytic activation curve as it does when cationic trypsin is used for initiation. At pH 5.0 activation is incomplete whether anionic or cationic trypsin is used to initiate activation. No activation occurs at pH 5.0 or 8.0 when activator is omitted. A comparison of Fig. 6, A and B, shows that anionic trypsinogen is activated more rapidly than cationic trypsinogen at pH 5.0 or pH 8.0 re- gardless of whether anionic trypsin or cationic trypsin is used to initiate activation. A second order autocatalytic rate constant of 0.14 was calculated for cationic trypsinogen activated by cationic trypsin at pH 8.0 (Fig. 6B), whereas a value of 0.84 was obtained for anionic trypsinogen activation in Fig. 5. Ac- rylamide gel electrophoresis of anionic trypsin activated from its eymogen at pH 8.0 and 5.0 in the Tris-glycine buffer system (13) immediately after full activation had been attained showed no difference in migration.
Stability studies on anionic trypsin activated from its zymogen at pH 8.0 showed that the enzyme lost approximately 50% of its original activity at pH 2.5 and 0” within a 2+-hour incubation period. However, when preparations of the enzyme were activated at pH 5.0 and subsequently adjusted to pH 2.5 only a gradual decline in activity was observed. The stability of cationic trypsin obtained by the dierent activation procedures was not affected at pH 2.5 during incubation times of up to 96 hours at 0”. Anionic trypsinogen retained 90% of its potential activity during a 96-hour incubation at 0” and pH 2.5.
The effects of calcium on cationic trypsin have been studied by McDonald and Kunitz (19) (bovine cationic trypsin) and by Buck et al. (2) (bovine, ovine, and porcine cationic trypsin). In all species it was shown that calcium stabilizes trypsin at basic pH values. Calcium also was found to stabilize anionic trypsin at pH 8.0 at 0” and more markedly at 37”.
Buck et al. (2) have suggested that the high isoelectric point of cationic trypsinogen is due to a high concentration of amide nitrogen which masks the charge of fl- and y-carboxyl groups. It is conceivable that the anionic trypsinogen could have the same primary structure as cationic trypsinogen but with fewer carboxyl groups as amides. However, the data given in Table I indicate that the primary structures of cationic and anionic trypsinogen are different.
K, values and specific activities for porcine anionic trypsin
MINUTES
FIG. 6. A comparison of the activation of anionic trypsinogen (ATZ) and cationic trypsinogen (CTZ) at pH 8.0 and 5.0 at 25” with either anionic trypsin (AT) or cationic trypsin (CT) to initi- ate activation. Activations were carried out in 0.1 M Tris-HCl buffer and 0.05 M CaC12, or at pH 5.0 in 0.1 M sodium acetate buffer in the presence or absence of 0.05 M CaC12. A, the initial concen- tration of anionic trypsinogen in the activation mixtures was 0.18 mg per ml and the ratio of zymogen to activator was 6O:l. B, initial concentrations of cationic trypsinogen were 0 42 mg per ml and the ratio of zymogen to activator was 20: 1. At different times portions were removed for determination of activity toward BANA. Specific activity at 100% activation was 3.3 and 1.1 for anionic and cationic trypsin, respectively.
8.0 CT 0.05, 0-m 8.0 AT 0.05, v-----v 5.0 CT 0.05, o-o 5.0 AT 0.05, n -m 5.0 CT o---o 5.0 AT a---m
from tissue and juice preparations and Armour crystallized porcine cationic trypsin were calculated from Lineweaver-Burk plots (Fig. 7) with the substrates tosyl-AME, BAEE, and lys- NA. The kinetic parameters were calculated from the best linear plots determined by regression analysis and are given in Table II. The data show that, with the three substrates studied, the specific activities and K, values for the anionic trypsin are very similar to those of porcine and bovine cationic trypsins. The specific activities given for BANA were not determined at V,, since the substrate preparations contained both D and L isomers, of which the former is a reversible inhibitor
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from
Anionic Trypsinogen Vol. 246, No. 2
I 1 I I 2 6
ws~ x 10-3 M - ’
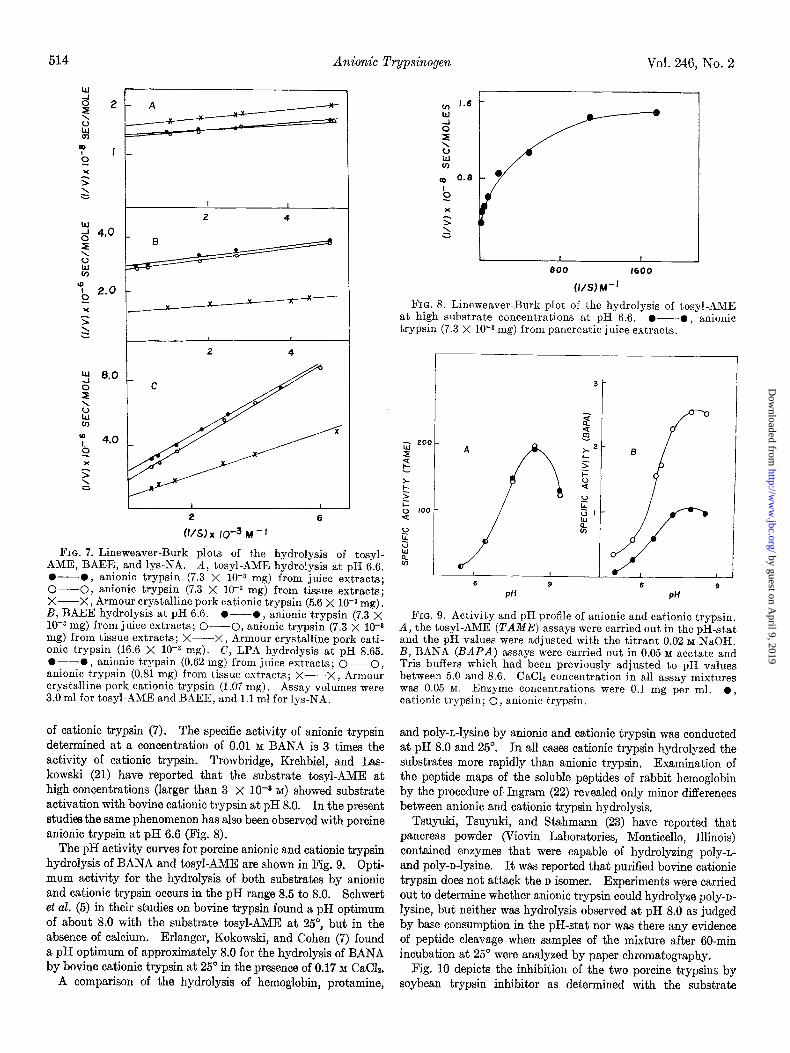
FIG. 7. Lineweaver-Burk plots of the hydrolysis of tosyl- AME, BAEE, and lys-NA. A, tosyl-AME hydrolysis at pH 6.6. O-0, anionic trypsin (7.3 X lob3 mg) from juice extracts; O-0, anionic trypsin (7.3 X lO+ mg) from tissue extracts; X-X, Armour crystalline pork cationic trypsin (5.6 X 10-B mg). B, BAEE hydrolysis at pH 6.6. O--O, anionic trypsin (7.3 X 10-s mg) from juice extracts; O--O, anionic trypsin (7.3 X 10-a mg) from tissue extracts; X-X, Armour crystalline pork cati- onic trypsin (16.6 X 10-S mg). C, LPA hydrolysis at pH 8.65. O---O, anionic trypsin (0.62 mg) from juice extracts; O-0, anionic trypsin (0.81 mg) from tissue extracts; X--X, Armour crystalline pork cationic trypsin (1.07 mg). Assay volumes were 3.0 ml for tosyl-AME and BAEE, and 1.1 ml for lys-NA.
of cationic trypsin (7). The specific activity of anionic trypsin determined at a concentration of 0.01 M BANA is 3 times the activity of cationic trypsin. Trowbridge, Krehbiel, and Las- kowski (21) have reported that the substrate tosyl-AME at high concentrations (larger than 3 x 10-s M) showed substrate activation with bovine cationic trypsin at pH 8.0. In the present studies the same phenomenon has also been observed with porcine anionic trypsin at pH 6.6 (Fig. 8).
The pH activity curves for porcine anionic and cationic trypsin hydrolysis of BANA and tosyl-AME are shown in Fig. 9. Opti- mum activity for the hydrolysis of both substrates by anionic and cationic trypsin occurs in the pH range 8.5 to 8.0. Schwert et al. (5) in their studies on bovine trypsin found a pH optimum of about 8.0 with the substrate tosyl-AME at 25”, but in the absence of calcium. Erlanger, Kokowski, and Cohen (7) found a pH optimum of approximately 8.0 for the hydrolysis of BANA by bovine cationic trypsin at 25” in the presence of 0.17 M CaC12.
A comparison of the hydrolysis of hemoglobin, protamine,
I t I 800 1600
(l/S~ K’
FIG. 8. Lineweaver-Burk plot of the hydrolysis of tosyl-AME at high substrate concentrations at pH 6.6. O-O, anionic trypsin (7.3 X lOma mg) from pancreatic juice extracts.
6 9 6 9 PH PH
FIG. 9. Activity and pH profile of anionic and cationic trypsin. B, the tosyl-AME (TAME) assays were carried out in the pH-stat and the pH values were adjusted with the titrant 0.02 M NaOH. B, BANA (BAPA) assays were carried out in 0.05 M acetate and Tris buffers which had been previously adjusted to pH values between 5.0 and 8.6. was 0.05 M.
CaClz concentration in all assay mixtures Enzyme concentrations were 0.1 mg per ml. l ,
cationic trypsin; 0, anionic trypsin.
and poly-n-lysine by anionic and cationic trypsin was conducted at pH 8.0 and 25’. In all cases cationic trypsin hydrolyzed the substrates more rapidly than anionic trypsin. Examination of the peptide maps of the soluble peptides of rabbit hemoglobin by the procedure of Ingram (22) revealed only minor differences between anionic and cationic trypsin hydrolysis.
Tsuyuki, Tsuyuki, and Stahmann (23) have reported that pancreas powder (Viovin Laboratories, Monticello, Illinois) contained enzymes that were capable of hydrolyzing poly-n- and poly-n-lysine. It was reported that purified bovine cationic trypsin does not attack the D isomer. Experiments were carried out to determine whether anionic trypsin could hydrolyze poly-D- lysine, but neither was hydrolysis observed at pH 8.0 as judged by base consumption in the pH-stat nor was there any evidence of peptide cleavage when samples of the mixture after 60-min incubation at 25” were analyzed by paper chromatography.
Fig. 10 depicts the inhibition of the two porcine trypsins by soybean trypsin inhibitor as determined with the substrate
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Issue of January 25, 1971 P. Voytek and E. C. Gjessing 515
mg STI/mg ENZYME
FIG. 10. The effect of the soybean trypsin inhibitor on the hy- drolysis of the substrate tosyl-AME by cationic and anionic tryp- sin. The rate of hydrolysis was determined in a pH-stat at P-H 6.6 at, 25” in the Dresence of 0.05 &X CaClz and 4.1 X 10e3 M tosvl- AME. The volume of t,he reaction mixture was 5.0 ml. 0, cati- onic trypsin (19.3 X low3 mg) ; 0, anionic trypsin (16.5 X 10e3 mg). The enzvme and inhibitor were incrlbated at DH 6.8 at 25” for 2 min prior tothe addition of substrate. Initial rates were used to cal- culate fractional activity (Vi/V) and were assumed to be propor- tional to the concentration of free enzyme in the equilibrium mixt,ure (24). The solid line was calculated by using a Ki of 2.0 X 10-S M and assuming the molecular weight of cationic trypsin to be 23,400 (3). The dashed line was calculated wit.h a K, of 7.0 X 10-g M and assuming the molecular weight for anionic trypsin to be 23,526 (Table I). The molecular weight of soybean trypsin inhibitor (5TZ) was assumed to be 18,200 (24).
tosyl-AME at pH 6.6. The theoretical curves were drawn assuming that the fractional initial velocities (Vi/V) were pro- portional to the concentration of free enzyme in equilibrium with inhibitor. The results show that soybean trypsin inhibitor is more effective in inhibiting porcine anionic trypsin than cationic trypsin. The effect of soybean trypsin inhibitor on cationic bovine trypsin has been studied by several investigators, particu- larly Green (24) and Lebowitz and Laskowski (25). Green calculated an apparent K; of 2 X lo-lo M at pH 7.8, whereas Lebowitz and Laskowski, using a novel potentiometric method for measuring protein association, determined an apparent K; of 2 x 10-g M at pH 5.75. At pH 8.0 DFP (0.01 M) completely inactivated anionic trypsin after 6-min incubation at 0”.
This is the first report of the isolation of a zymogen of an anionic trypsin from porcine pancreas. Although anionic trypsinogen and cationic trypsinogen show similar specificities toward synthetic trypsin substrates and proteins, they are quite different in isoelectric point as judged by their behavior on DEAE-cellulose and zone electrophoresis. Like cationic tryp- sinogen, anionic trypsinogen is autocatalytically converted to an active enzyme. In addition, electrophoretic comparison of anionic trypsin and its precursor suggests that in the activation process an acid peptide fragment is removed from the trypsino- gen, yielding an enzyme which is less anionic than the zymogen. Neurath and his group (26, 27), as well as Charles et al. (4), have found that autocatalytic activation of cationic trypsinogen from both the cow and pig involves the rupture of a single peptide
and the concomitant liberation of a hexa- and octapeptide, respectively. Both peptides are acidic and the active enzymes formed have isoelectric points greater than their corresponding zymogens.
The similarities in the hydrolysis of synthetic substrates and proteins by anionic and cationic trypsin raise the question as to why the animal would synthesize two such enzymes in equiva- lent amounts. The data presented here support the possibility that anionic trypsin may play a more important role than cati- onic trypsin in initiating activation of proteolytic zymogens in the intestine. However, in order to confirm this concept more studies on activations simulating conditions in viva would be required, including comparisons between activation of other zymogens by both anionic and cationic trypsins.
In recent years reports from several laboratories have sug- gested the presence of anionic trypsin in the pancreas of species other than the cow or the pig. Prahl and Neurath (28) have made an extensive study of some of the proteolytic enzymes of the pancreas of the Spiny Pacific dogfish. The authors reported finding a trypsin in the anionic fraction eluted from a DEAE-cellulose column at pH 6.6 with a sodium succinate gradient, while no cationic trypsin appeared to be present in the same species. Croston (29) found anionic trypsin as well as cationic trypsin in extracts of the cecal tissue of the Chinook salmon. In a study of the enzymes of human pancreatic juice, Keller and Allan (30) reported the presence of an anionic trypsin while cationic trypsin appeared to be absent or present in amounts not considered significant. Marchis-Mouren (31) in a preliminary communication has reported finding an anionic trypsinogen in the pancreatic juice of the rat.
Recently, Sach and Thely (32) have discovered a hydrolase in porcine pancreas with activities toward the substrates BAEE, N-acetyl-L-lysine methyl ester, and casein. The hydrolase enzyme is anionic as judged by its electrophoretic properties and by its elution from DEAE-cellulose. The hydrolase is eluted from the anion exchanger immediately preceding the esteroproteolytic enzyme which is consistent with our observa- tions. The authors further reported that the hydrolase had a molecular weight of 32,000 to 33,000 as determined by the gel filtration technique. However, the hydrolase was inactive toward the substrate BANA and was not inhibited by soybean trypsin inhibitor.
Acknowledgment--We wish to express our appreciation to Dr. Nicholas Calvanicb of the State University of New York at Buffalo for the molecular weight and amino acid determina- tions.
REFERENCES
1. MCCONNELL, B., AND GJBSSING, E. C., J. Biol. Chem., 241, 573 (1966).
2. BUCK, F. F., \‘ITH.LY.I’I’HIL, A. J., BIER, M., ANU NORD, F. F., Arch. Biochem., 97, 417 (1962).
3. TRAVIS, J., AND LIENF:IL, I. E., J. Biol. Chem., 240,1962 (1965). 4. CHARLES, M., KOVERY, M., GUIDONI, A., AND DESNUELLE, P.,
Biochim. Biophqs. Acta, 69, 115 (1963). 5. SCH~VERT, G. I$.,- NEURATH, H., KAUFMAN, S., AND SNOICE,
J. E.. J. Biol. Chem.. 172, 221 (19481. 6. YAMAS.&, M., BROXVN, J..R., cox, ‘D. J., GREENSHI>>LDS,
It. N., WAD?:, 1%. T>., AND NEURATH, H., Biochemistry, 2, 859 (1963).
7. ERLANGER, B. F., KOI~O~SKI, N., AND COHEN, W., Arch. Bio- them. Biophys., 96, 271 (1961).
8. CAMPBELL, W. R., AND HANNA, M. I., J. Biol. Chem., 119, 1
(1937).
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

516 Anionic Trypsinoyen Vol. 246, No. 2
9.
10. 11. 12.
13. 14. 15.
16.
17.
18.
19.
MOORE, S., AND STEIN, W., in S. P. C~LOWICIC AI\TD N. 0. KAPLAN (Editors), Methods in enzymology, Vol. 6, Academic Press, New York, 1963, p. 819.
BENCZE, W. L., AND SCHMID, K., Anal. Chem., 29,1193 (1957). YPHANTIS, D. A., Ann. N. Y. Acad. Sci., 88,486 (1960). WOOD~ORTH, R. C., AND CLARK, L. G., Anal. Biochem., 18,
295 (1967). DAVIS, B. J., Ann. N. Y. Acad. Sci., 121,404 (1964). RACUSEN, D., Nature, 213, 922 (1967). PETERSON, E. A., AND SOBER, H. A., in S. P. COLOWICIC END
N. 0. KAPLAN (Editors), Methods in enzymology, Vol. 6, Academic Press, New York, 1962, p. 3.
LEVIN, O., in S. P. COI,OWICK AND N. 0. KAPLAN (Editors), Methods in enzvmologv, Vol. 5. Academic Press, New York, 1962,p.27. ” ““’ ROVERY, M., CHARLES, M., GUY, O., GUIDONI, A., AND DES-
NUELLE, P.. Bull. Sot. Chim. Biol.. 42, 1235 (1960). KUNITZ, 6f., AND NORTHROP, J. H.,‘J. ‘Gen. Physiol., 19, 991
(1936). MCDONALD, M. R., BND KUNITZ, M., J. Gen. Physiol., 26, 53
(1941).
20. INAGAXI, T., J. Amer. Chem. Sot., 84,2556 (1962). 21. TROWBRIDGE, C. G., KREHBIEL, A., AND LASKOWSKI, M., JR.,
Biochemistry, 2, 843 (1963). 22. INGRAM, V. M., Biochim. Biophys. Acta, 28,539 (1958). 23. TXTYUKI, E., TSUYUKI, H., AND ST.~HMANN, M. A., J. Biol.
Chem., 222, 479 (1956). 24. GREEN, N. M., J. Biol. Chem., 205, 535 (1953). 25. LEBOWITZ, J., AND LASICOWSKI, M., JR. Biochemistry, 1, 1044
(1962). 26. DAVIE, E. W., AND NEURATH, H., J. Biol. Chem., 212, 515
(1955). 27. DIXON, G. H., NEURATH, H., AND PECHERE, J. F., Annu. Rev.
Biochem., 27, 489 (1958). 28. PRAHL, J. W., AND NEURATH, H., Biochemistry, 6, 2131 (1966). 29. CROSTON, C. B., Arch. Biochem. Biophys., 112,218 (1965). 30. KELLER, P. J., AND ALLAN, B. J., J. Biol. Chem., 242, 281
(1967). 31. MARCHIS-MOUREN, G., P~SERO, L., AND DESNUELLE, P.,
Biochem. Biophys. Res. Commun., 13, 262 (1963). 32. SACH, E., AND THELY, M., Compt. Rend. Acad. Sci., 266, 1200
(1968).
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Peter Voytek and Erland C. GjessingStudies of an Anionic Trypsinogen and Its Active Enzyme from Porcine Pancreas
1971, 246:508-516.J. Biol. Chem.
http://www.jbc.org/content/246/2/508Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/246/2/508.full.html#ref-list-1
This article cites 0 references, 0 of which can be accessed free at
by guest on April 9, 2019
http://ww
w.jbc.org/
Dow
nloaded from