structural and interaction studies of bacterial ...191936/fulltext01.pdf · and interaction studies...
TRANSCRIPT

S t ruc tura l and In te rac t ion S tud ies o f Bac te r ia l Po lysacchar ides by NMR Spec t roscopy
E V A - L I S A N O R D M A R K

© Eva-Lisa Nordmark 2004 ISBN 91-7265-971-8 pp 1-50 Doctoral disser ta t ion Department of Organic Chemistry Stockholm Universi ty Pr inted in Sweden by Akademitryck AB Valdemarsvik 2004
2

A B S T R A C T AN INTRODUCTION TO bacterial polysaccharides and the methods for structural determination are described in the first two parts of the thesis.
In a structural elucidation of bacterial polysaccharides NMR experiments are important as is component analysis. A short description of immunochemical methods such as enzyme immunoassays is included. Two NMR techniques used for interaction studies, trNOE and STD NMR, are also discussed.
The third part of the thesis discusses and summarizes the results from the included papers. The structures of the exopolysaccharides produced by two lactic acid bacteria are determined by one- and two dimensional NMR experiments. One is a heteropolysaccharide produced by Streptococcus thermophilus and the other a homopolysaccharide produced by Propionibacterium freudenreichii . The structure of an acidic polysaccharide from a marine bacterium with two serine residues in the repeating unit is also investigated. The structural and immunological relationship between two O-antigenic polysaccharides from Escherichia coli strain 180/C3 and O5 is discussed and investigated. Finally, interaction studies of an octasaccharide derived from the Salmonella enteritidis O-antigen and a bacteriophage are described which were performed with NMR experiments.
3

4

”Jag var ga len . Al la böcker har jag ä lskat , tunna, t jocka , fu la , snygga,
f ranska band, som en marskat t , pr imi t iv t , en bokl ig kväl l skat t som i månens sken får käns lorna i brand”
T A G E D A N I E L S S O N
5

6

T A B L E O F C O N T E N T S
L I S T O F A B B R E V I AT I O N S
L I S T O F PA P E R S
1. I N T R O D U C T I O N Bacteria in General - 11, Lactic Acid Bacteria - 14, Pseudoalteromonas aliena - 14, Escherichia coli - 15, Salmonella and Bacteriophage Interactions - 16
2. M E T H O D S F O R S T R U C T U R A L
D E T E R M I N AT I O N Component Analysis - 17, Methylation Analysis - 18, Immunochemical Methods of Analysis - 19, NMR Experiments for Structural Elucidation - 20, NMR Experiments for Interaction Studies - 23
3. R E S U LT S A N D D I S C U S S I O N S Structural Studies of three Different Polysaccharides (PAPERS I-III) - 26, Structural and Immunochemical Relationship Between two O-antigenic Polysaccharides (PAPER IV) - 36, Interaction Studies by NMR Spectroscopy (PAPER V) – 42
C O N C L U S I O N S
A C K N O W L E D G E M E N T S
PA P E R S I - V
7

L I S T O F A B B R E V I AT I O N S LPS l ipopolysaccharide EPS exopolysaccharide EAEC enteroaggregat ive Escherichia col i GLC gas- l iquid chromatography GC-MS gas chromatography mass spectrometry SDS-PAGE sodium dodecyl sulphate - polyacrylamide gel
e lectrophoresis ELISA enzyme-l inked immunosorbent assay COSY correlated spectroscopy TOCSY total correla t ion spectroscopy HSQC heteronuclear s ingle quantum coherence HMBC heteronuclear mult iple bond correlat ion BS-CT-HMBC band select ive constant t ime HMBC NOESY nuclear Overhauser effect spectroscopy T-ROESY transverse rotat ing-frame Overhauser effect
spectroscopy t rNOE t ransferred nuclear Overhauser effect STD saturat ion t ransfer di fference J scalar coupl ing constant δ chemical shif t
8

L I S T O F PA P E R S
PAPER I: Structural Studies of an Exopolysaccharide Produced by Streptococcus thermophilus THS Eva-Lisa Nordmark, Zhennai Yang, Eine Huttunen and Göran Widmalm Biomacromolecules , 6 (2005) in press
PAPER II: Structural Studies of the Exopolysaccharide Produced by Propionibacterium Freudenreichii ssp. Shermanii JS Eva-Lisa Nordmark, Zhennai Yang, Eine Huttunen and Göran Widmalm Biomacromolecules , 6 (2005) in press PAPER III: Structure of an Acidic Polysaccharide from the Marine Bacterium Pseudoalteromonas aliena Type Strain KMM 3562T Containing two Residues of L-Serine in the Repeating Unit Eva-Lisa Nordmark, Andrei V. Perepelov, Alexander S. Shashkov, Evgeny L. Nazarenko and Göran Widmalm in manuscr ipt PAPER IV: Structural Elucidation of the O-antigenic Polysaccharide from the Enteroaggregative Escherichia coli Strain 180/C3 and its Immunochemical Relationship with Escherichia coli O5 and O65 Fel ipe Urbina, Eva-Lisa Nordmark, Zhennai Yang, Andrej Weintraub, Flemming Scheutz and Göran Widmalm submit ted PAPER V: Interaction Studies of a Salmonella enteritidis O-antigen Octasaccharide to Phage P22 Tail Spike Protein by NMR Spectroscopy and Molecular Modeling Eva-Lisa Nordmark, Robert Eklund, Andrej Weintraub, Robert Seckler and Göran Widmalm in manuscr ipt
9

10

1.
I N T R O D U C T I O N B a c t e r i a i n G e n e r a l BACTERIA ARE AMONG the smallest organisms on earth. They appear everywhere where life exists - in the air, in the ground and in the water as well as on larger organisms. Most of them are important for the processes of l ife and are not pathogenic as we might think. In the past they were classified as being a part of the vegetable kingdom. Today, however, they and cyanobacteria are classified together as prokaryota. They are named prokaryota because they lack cell nucleus.
Bacteria were described in 1683 by Leeuwenhoek using the first microscope. He called them “animacules”. FIGURE 1 shows Leeuwenhoek’s picture presented to the Royal Society in London. It was not until the 19t h century that they were studied in more depth by Louis Pasteur and Robert Koch.
F I G U R E 1. “Animacules” descr ibed by Leeuwenhoek in 1683.1
1 f rom Nat iona lencyk loped in , 2 (1990) 225 .
11

The total number of known bacterial species exceeds a million, but only around 3000 of them have been investigated.1 Bacteria can be beneficial or pathogenic. Strains of Streptococcus , Lactobacillus and Lactococcus are used in the food industry as probiotica and/or natural alternatives to commercial thickeners and stabilizers in milk products, yoghurt, cheese and vinegar and are beneficial to us.2 Some bacteria are always pathogenic. We also have many types of bacteria in the gut that are necessary to protect us but if they somehow are transported to other parts of our body they can become pathogenic, for example Escherichia coli .
There are at least three common methods to classify bacteria. Bacterial survival is very temperature dependent and there are three different types. Psycrophilic bacteria have a growth optimum not higher than 20 °C and do not survive temperatures over 30 °C. Mesophilic bacteria have their growth optimum at 20 – 40 °C and thermophilic bacteria have their growth optimum above 45 °C. In general, bacteria are much more sensitive to heat than to cold.
Bacteria are morphologically divided into three groups: bacillus or rod-shaped bacteria, coccus which are sphere-shaped and spirochete, the spiral-shaped bacteria that have a cork-screw type motili ty (see FIGURE 2).
bacillus coccus spirochete
F I G U R E 2. Three examples showing the different morphology of bacter ia . 3
Most bacteria are also classified with a method called Gram-
staining. This procedure was devised more than a century ago (1884) by Hans Christian Joachim Gram. The procedure involves staining of bacteria with a purple dye. Bacteria that remain
2 A .P . Laws and V.M. Marsha l l , In t . Da i ry J . , 11 (2001) 709-721 . 3 H .M. Gram, Den med ic in ske mikrob io log i og immun i t e t s laere , 1915 .
12

stained purple after the whole procedure are called Gram-positive and the others Gram-negative. The cell walls of Gram-positive bacteria have a high affinity for the dye and the Gram-negative cell walls have a very low affinity. The reason for this difference lies in the structure of the cel l walls of the bacteria. Gram-positive cell walls are about five times as rich in peptidoglycan as Gram-negative bacteria. FIGURE 3 shows the cell walls of Gram-positive and Gram-negative bacteria.
LPS
outer membrane
thin peptidoglycan layer
inner membrane
CPS and EPS
thick peptidoglycan layer
lipoteichoic acid
plasma membrane
lipoproteins
Gram-negative Gram-positive
F I G U R E 3. The difference between Gram-negat ive and Gram-posi t ive bacter ia .
The Gram-negative bacteria have both an inner and an outer
membrane. The latter contains a l ipopolysaccharide (LPS) (see FIGURE 4), which comprises three regions: the O-antigenic polysaccharide, the core and lipid A. The core is a non-repetitive oligosaccharide l inking the O-antigenic polysaccharide to lipid A. The O-antigenic polysaccharides are usually built of repeating units of 2 - 7 sugar residues. The Gram-positive bacteria possess only one cell membrane. Both Gram-positive and Gram-negative bacteria may be surrounded by an external polysaccharide called capsular polysaccharide (CPS) or exopolysaccharide (EPS).
O-antigen core lipid A
F I G U R E 4. The schematic s t ructure of an LPS.
13

Investigation of the structures of the bacterial polysaccharides are important for the understanding of many different processes, such as immune defence, parasitic infections and inflammation. In this thesis some quite different bacterial species are studied. Streptococcus thermophilus and Propionibacterium freudenreichii ssp. shermanii JS are Gram-positive, lactic acid bacteria, which are beneficial and used in the food industry. Pseudoalteromonas aliena is a Gram-negative, rod-shaped marine bacterium. Escherichia coli is also a Gram-negative rod, that is beneficial in some cases and pathogenic in others. In this case, the strain is pathogenic and causes diarrhoea.
L a c t i c A c i d B a c t e r i a LACTIC ACID BACTERIA are involved in the preparation of a large number of fermented food products, such as yoghurt. Incorporation of cultures that produce EPS in fermented products increases the viscosity, stability and water binding property of the products.4 These physical properties have a close relationship with the structure of the EPS produced by the bacteria. More than 45 different EPS producing lactic acid bacteria have been structurally investigated and a number of them produce the same EPS. Physiologically, EPS protects bacterial cells against a variety of unfavourable environmental factors, such as bacteriophages, protozoa, antibiotics and lysozyme.5 It has also been suggested that EPS is involved in bacterial adhesion and biofilm formation.6 In order to further understand the structure-function relationship of EPS, structural studies have been carried out on an EPS produced by Streptococcus thermophilus in PAPER I and an EPS produced by Propionibacterium freudenreichii in PAPER I I . P s e u d o a l t e r o m o n a s a l i e n a PSEUDOALTEROMONAS IS ONE of the largest genera within the γ-Proteobacteria and contains more than 30 species.7 These bacteria are essential components of the marine environment and produce a wide range of biologically active compounds. Pseudoalteromonas species are generally found in association
4 J .R . Broadben t , D . J . McMahon , D .L . Welke r , C . J . Oberg and S . Moineau , J . Da iry Sc i . , 86 (2003) 407-423 . 5 P . Ruas-Made ido , J . Hugenho l t z and P . Zoon , In t . Da i ry J . , 12 (2002) 163-171 . 6 A . Laws , Y . Gu and V . Marsha l l , Bio tech . Adv . , 19 (2001) 597-625 . 7 E .L . Nazarenko , N .A. Komandrova , R .P . Gorshkova , S . V . Tomsh ich , V .A . Zubkov , M. K i lcoyne and A .V . Savage , Carbohydr . Res . , 33 (2003) 2449-2457 .
14

with marine eukaryotes in marine waters all over the world. Pseudoalteromonas species also show antibiotic effects.8
The polysaccharides of the genus Pseudoalteromonas are all acidic in character. They also contain unusual sugars, such as N-acyl derivatives of 6-deoxyaminosugars such as N-acetyl-D-quinovosamine (vide infra). Many constituent sugars in their LPS have various uncommon non-sugar substituents, such as alanine, formic acid or sulphate. The repeating units do not show any other gross structural similarities.
In PAPER I I I the structure of the polysaccharide of Pseudoalteromonas aliena type strain KMM 3562T, isolated from sea-water samples collected from the Amursky Bay (Sea of Japan, Pacific Ocean), has been elucidated.9
E s c h e r i c h i a c o l i THREE GENERAL CLINICAL syndromes result from infection with inherently pathogenic Escherichia coli strains: urinary tract infection, sepsis/meningitis and enteric/diarrhoeal disease. Diarrhoea remains one of the main sources of morbidity in today’s world and a large proportion is caused by diarrhoeagenic E. coli .10
Five pathotypes of diarrhoeagenic E. coli have been associated with diarrhoeal i l lness:11 enterotoxigenic E. coli (ETEC), enteropathogenic E. coli (EPEC), enterohemorrhagic E. coli (EHEC), enteroinvasive E. coli (EIEC) and enteroaggregative E. coli (EAEC).12 Enteroaggregative E. coli may be an emerging diarrhoeal pathogen and has been associated characteristically with persistent diarrhoea among infants,13 especially in developing countries. However, recent outbreaks and volunteer studies are suggesting that EAEC strains are also virulent in adults14 and have a global distribution.15
Studies on the structures of the polysaccharide part of E. coli l ipopolysaccharides are important in understanding the role of this
8 C . Ho lms t röm and S . Kje l l ebe rg , FEMS Microb io l . Eco l . , 30 (1999) 285 -293 . 9 E .P . Ivanova , N . M. Gor shkova , N .V . Zhukova , A .M. Lysenko , E .A . Ze lepuga , N .G. P rokofeva , V .V . Mikha i lov , D .V . N ico lau and R . Chr i s t en , In t . J . Sys t . Eco l . Microb io l . , 54 (2004) 1431-1437 . 10 S .C . C la rke , Diagnos t i c Microb io logy and In fec t ious Disease , 41 (2001) 93 -98 . 11 J .P . Na ta ro and J .B . Kape r , Cl in . Microb io l . Rev . , 11 (1998) 142 -201 . 12 M.K. Bhan , V . Khoshoo , H . Sommer fe ld t , P . Ra j , S . Sazawa l and R. S r ivas t ava , Pedr ia t r . In f ec t . D i s . J . , 8 (1989) 499-502 . 13 M.K. Bhan , P . Ra j , M.M. Lev ine , J .B . Kaper , N . Bhandar i , R . S r ivas t ava , R . Kumar and S . Sazawa l , J . In f ec t . D i s . , 159 (1989) 1061-1064 . 14 J .P . Na ta ro , D . Y ikang , S . Cookson , A . Crav ia to , S . J . Sava r ino , L .D. Guer s , M.M. Lev ine and C .O . Tacke t , J . In f ec t . D i s . , 17 (1995) 465 -468 . 15 H . I . Hupper t z , S . Ru tkowsk i , S . Aleks i c and H . Karch , Lance t , 349 (1997) 1660-1662 .
15

glycan in serological specificity and pathogenesis.16 In PAPER IV an O-antigenic polysaccharide from the enteroaggregative E. coli strain 180/C3 is studied.
S a l m o n e l l a a n d B a c t e r i o p h a g e I n t e r a c t i o n s THE POLYSACCHARIDE COMPONENTS of bacteria have important structural and functional roles in the life of the bacterial cell .17 A large group of bacteriophages use surface carbohydrates as receptor targets. The LPS of bacteria are their first l ine of defence against these bacteriophages. Many biological processes involve protein-ligand interactions and the biological function of a protein depend to a large extent on these interactions.18 The identification of the binding epitope of l igands is therefore important to investigate. The LPSs from the Salmonella19 serogroups are widely investigated both structurally, biochemically and genetically. The Salmonella serogroups A, B and D1
2 0 have a trisaccharide element, galactose-mannose-rhamnose, in the O-antigen in common. The fourth sugar, a 3,6-dideoxyhexose residue, is either paratose, abequose or tyvelose, respectively.20 The antigenic determinants or epitopes recognized by antibodies are the 3,6-dideoxyhexoses.21 In PAPER V, the interaction between an octasaccharide derived from the Salmonella enteritidis (serogroup D1) O-antigen and phage P22 tail spike protein is studied. 16 L .L . Mac lean and M.B. Per ry , Biochem. Ce l l B io l . , 75 (1997) 199 -205 . 17 A . Vark i , R . Cummings , J . Esko , H . F reeze , G . Har t and J . Mar th , Essen t ia l s o f G lycob io logy , (1999) 321-331 . 18 B . Meyer and T . Pe te r s , Angew. Chem. , 115 (2003) 890 -918 , Angew. Chem. In t . Ed . 42 (2003) 864-890 . 19 C .G . He l l e rqv i s t , B . L indbe rg , T . H . Svensson and A.A. L indbe rg , Carbohydr . Res . , 8 (1968) 43-55 . 20 K . Bock , M. Melda l , D . Bund le , T . Iver sen , P . J . Garegg , T . Norberg , A .A . L indberg and S .B. Svenson , Carbohydr . Res . , 130 (1984) 23 -34 . 21 H .N . Yu , C . -C . L ing and D. Bund le , Can. J . Chem. , 80 (2002) 1131-1140 .
16

2.
M E T H O D S F O R S T R U C T U R A L D E T E R M I N AT I O N C o m p o n e n t A n a l y s i s SUGAR ANALYSIS and absolute configuration determination are well established methods to determine the sugar components in the repeating unit of a polysaccharide. In the first step of the sugar analysis22, the polysaccharide is hydrolysed with acid, leading to the formation of monosaccharides, which are subsequently reduced to alditols with sodium borohydride or i ts deuterium analogue. Acetic anhydride in pyridine is then used to acetylate the alditols. They are then separated by gas-liquid chromatography (GLC). Each monosaccharide gives only a single derivative regardless of whether it existed as an α- or β-linked, pyranoid or furanoid sugar in the native polysaccharide. The relative intensities of the peaks in the chromatogram tell us roughly how many residues of each monosaccharide that are present in the repeating unit . As a rule of thumb, an error of 10 - 25% is to be anticipated in molar ratios obtained from monosaccharide analysis.23
One way of determining the absolute configuration (D or L) of the sugar residues is to derivatize them with an optically active alcohol.24, 25 This method requires authentic standards as references. Consequently, the absolute configuration of an uncommon sugar is difficult to confirm in this way.
The polysaccharide is hydrolysed and reacted with an optically active alcohol. In this case, S-(+)-2-butanol is used. The enantiomeric sugars react with the chiral butanol and form diastereomeric glycosides that have different physical properties 22 J .S . Sawardeker , J .H . S loneker and A . Jeanes , Ana l . Chem . , 37 (1965) 1602-1604 . 23 J . J iménez -Barbe ro and T . Pe te r s , NMR spec t roscopy o f G lycocon juga te s , (2003) 97 . 24 G . J . Gerwig , J .P . Kamer l ing and J .F .G . V l i egen thar t , Carbohydr . Res . , 62 (1978) 349-357 . 25 K . Leon te in , B . L indberg and J . Lönnberg , Carbohydr . Res . , 62 (1978) 359 -362 .
17

and therefore can be separated by GLC. The glycosides formed are acetylated and compared to reference substances by GLC. The retention times differ between the different sugars and also between the different D- or L-configured (diastereomeric) sugars.
M e t h y l a t i o n A n a l y s i s THE SUBSTITUTION PATTERN of the different sugar residues of the repeating unit can be determined using methylation analysis.26 The polysaccharide is treated with strong base (sodium methylsulfinylmethanide) and methyl iodide in DMSO, which methylates all the free hydroxyl groups of the polysaccharide. After this initial step, the procedure continues with the steps in the sugar analysis as described above. The substituted oxygens of the polysaccharide are identified as those hydroxyl groups that are acetylated. The substitution patterns of the sugars in the repeating unit are determined by separation on GC-MS and a characteristic mass fragmentation pattern. One complication is that 4-substituted pyranoside gives the same derivate as a 5-substituted furanoside. Fortunately, NMR can determine which one is present.
Monosaccharides in natural compounds are usually present in a cyclic hemiacetal form, in which the exocyclic hemiacetal oxygen atom is a constituent of the glycosidic linkage. However, in 1999 Vinogradov and Bock27, 28 identified a new type of l inkage between monosaccharides found in the LPS core from two serotypes of Proteus mirabilis species (FIGURE 5). The data suggested a new type of glycosidic structure with an open-chain N-acetylgalactosamine residue linked as a cyclic acetal to positions 4 and 6 of the galactosamine in the α-pyranoside form. The symbol “o” for open was proposed to be used to identify this type of sugar in abbreviated formulas.
26 S . Hakomor i , J . B iochemis t ry (Tokyo) , 55 (1964) 205 -208 . 27 E . V inogradov and K . Bock , Angew. Chem. , 111 (1999) 712-715 , Angew. Chem. In t . Ed . , 38 (1999) 671-674 . 28 E . V inogradov and K . Bock , Carbohydr . Res . , 319 (1999) 92-101 .
18

O
OO
H2NO
HO
NHAc
O
HOHO
OH
OH
O
OH
OH
HO
F I G U R E 5. Schematic picture of the open-chain element in a
core f ragment of LPS from Proteus mirabi l is O27.
The chemical stability of the acetal l inkage formed by the N-acetylgalactosamine residue does not differ very much from that of a normal glycosidic linkage. It survives acetic acid hydrolysis as well as the basic conditions during the methylation analysis, but is cleaved under the hydrolytic conditions normally used in monosaccharide analysis.
It may therefore be widely present in natural poly- and oligosaccharides. Conventional analysis of the Proteus mirabilis serotype O27 core would lead to the identification of a terminal glucose, a terminal N-acetylgalactosamine and 4,6-disubstituted galactosamine instead of the structural fragment described in FIGURE 5.
I m m u n o c h e m i c a l M e t h o d s o f A n a l y s i s THERE ARE MORE than 170 serotypes of E. coli strains.29 The serotyping is based on the reactivity of specific antibodies with cell wall components using whole bacteria in an agglutination assay. However, sometimes problems can arise. Normally, rough strains, i .e. , strains lacking the O-antigen, agglutinate spontaneously. This is also a common feature for the enteroaggregative E. coli isolates and thus they are difficult to distinguish from rough E. coli strains. An SDS-PAGE experiment
29 F .Ørskov and I . Ør skov , Can . J . Microb io l . , 38 (1992) 699-704 .
19

can, however, tell if the LPS is isolated from a rough strain or not.
One way of testing LPS for reactivity with antibodies is the enzyme-linked immunosorbent assay (ELISA). Using purified LPS and specific antibodies it is possible to predict if there are structural similarit ies between different LPS. Indirect ELISA is used in the initial state. Further investigations can be done with inhibition ELISA.
N M R E x p e r i m e n t s f o r S t r u c t u r a l E l u c i d a t i o n IN STRUCTURAL STUDIES of polysaccharides and interaction studies nuclear magnetic resonance (NMR) spectroscopy30 is widely used. In a structural elucidation of bacterial polysaccharides NMR experiments are an important complement to component analysis. Both one- and two-dimensional NMR experiments are performed. The nuclei of interest are 1H and 1 3C and in some cases 3 1P and 1 5N. The 2D-NMR experiments used in the structural studies reported in this thesis are the following: 1H,1H COSY,31 1H,1H TOCSY,32 1H,1 3C HSQC,33 1H,1 3C HSQC-TOCSY34, 1H,1 3C HSQC-NOESY,35 1H,1 3C HMBC,36 BS-CT-HMBC37 and 1H,1H NOESY.38
Three- and four-dimensional NMR spectroscopy39 have found few applications in structural determinations of carbohydrates. However, these experiments are essential for large proteins.40
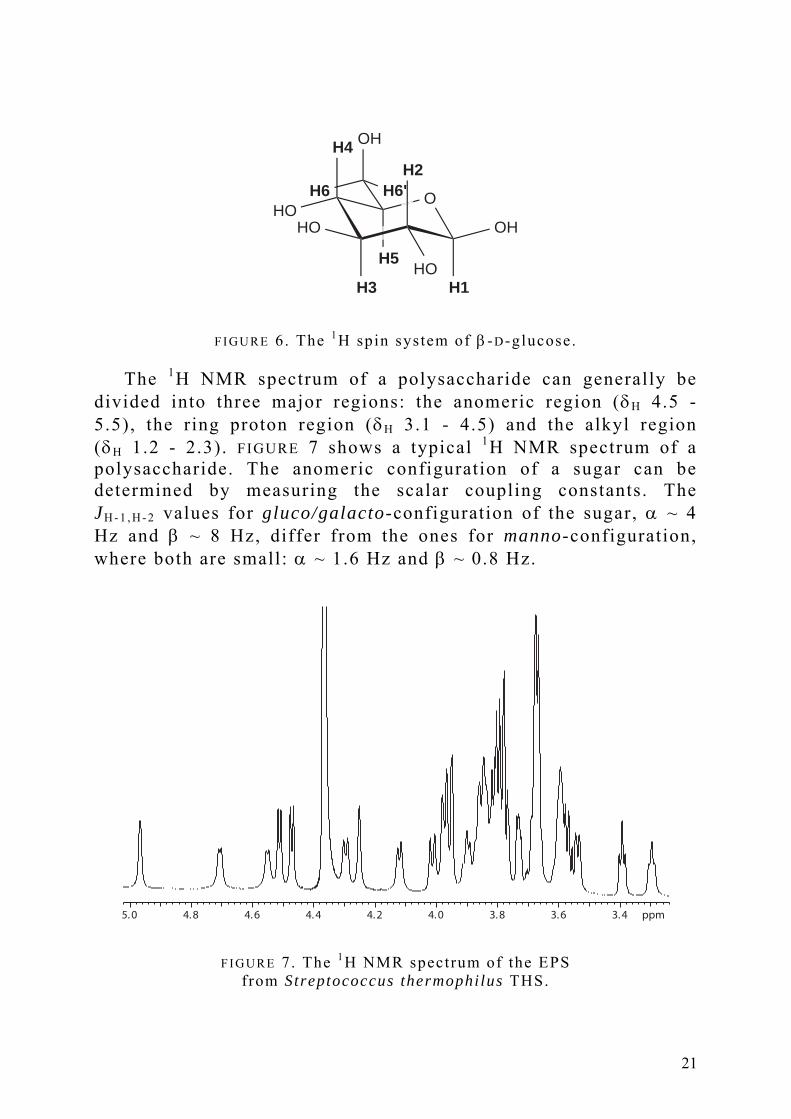
Bacterial polysaccharides consist of repeating units built up by monosaccharides connected by glycosidic linkages. This gives isolated spin systems, which are “isolated” in the sense that magnetisation must be transferred over the glycosidic linkage to connect two monosaccharide spin systems. FIGURE 6 shows the β-D-glucose spin system, which contains seven non-exchangeable protons and six carbons.
30 J . J iménez -Barbe ro and T . Pe te r s , NMR spec t roscopy o f G lycocon juga te s , 2003 . 31 A . Bax and R . F reeman , J . Magn . Res . , 44 (1981) 542 -561 . 32 L . Braunschwei l e r and R .R . Erns t , J . Magn . Res . , 53 (1983) 521 -528 . 33 L .E . Kay , P . Ke i fe r and T . Saa r inen , J . Am. Chem. Soc . , 114 (1992) 10663-10665 . 34 T . de Beer , C .W.E .M. van Zuy len , K . Hård , R . Boe lens , R . Kap te in , J .P . Kamer l ing , J .F .G . V l iegen tha r t , FEBS Le t t . , 348 (1994) 1 -6 . 35 R . Wagner and S . Berge r , Magn . Reson . Chem. , 35 (1997) 199 -202 . 36 A . Bax and M.F . Summers , J . Am. Chem. Soc . , 108 (1986) 2093-2094 . 37 T .D .W. C la r idge and I . Pé rez-Vic to r i a , Org . B iomol . Chem. , 1 (2003) 3632-3634 . 38 A . Kumar , R .R . Erns t and K . Wüth r i ch , Biochem. B iophys . Res . Commun . , 95 (1980) 1 -6 . 39 G .M Clo re and A .M. Gronenborn , Sc ience , 252 (1991) 1390-1399 . 40 J . F l aux , E . B . Ber t e l sen , A .L . Horwich and K . Wüthr i ch , Nature , 418 (2002) 207-211 .
20
O
H2
HO
HOHO
H3
H4
H5
OH
H1
H6'
OH
H6
F I G U R E 6. The 1H spin system of β -D-glucose.
The 1H NMR spectrum of a polysaccharide can generally be
divided into three major regions: the anomeric region (δH 4.5 - 5.5), the ring proton region (δH 3.1 - 4.5) and the alkyl region (δH 1.2 - 2.3). FIGURE 7 shows a typical 1H NMR spectrum of a polysaccharide. The anomeric configuration of a sugar can be determined by measuring the scalar coupling constants. The JH - 1 , H - 2 values for gluco/galacto-configuration of the sugar, α ~ 4 Hz and β ~ 8 Hz, differ from the ones for manno-configuration, where both are small: α ~ 1.6 Hz and β ~ 0.8 Hz.
ppm3. 43. 63. 84. 04. 24. 44. 64. 85. 0
F I G U R E 7. The 1H NMR spectrum of the EPS from Streptococcus thermophilus THS.
21

In order to obtain a good 1 3C NMR spectrum concentrated samples are required. The sensitivity is lower due to the low natural abundance (1.1%) and lower magnetogyric ratio of the 1 3C nucleus.
The 1 3C NMR spectrum can also be divided into different regions. The anomeric carbons are found at δC 95 - 110, the ring carbons at δC 50 - 85, the alkyl carbons at δC 15 - 25 and the carbonyl carbons at δC 165 - 180.
The chemical shifts of the carbon resonances for substituted positions are shifted downfield compared to the corresponding monosaccharides41 by about 6 - 10 ppm. This is used as a reliable indication of the substitution pattern.
For a complete structural elucidation one-dimensional spectroscopy does not give enough information. Two-dimensional NMR experiments are therefore also needed.
The COSY experiment shows the couplings between protons that are indirectly coupled to each other. These scalar couplings are shown as cross peaks in the spectrum, e.g. H-1 to H-2, H-2 to H-3, H-3 to H-4 and so on.
The TOCSY experiment gives more information; here it is possible to follow a complete (isolated) spin system. During the mixing time, an applied spin-lock allows the magnetization to travel through the spin system. A long mixing time gives more correlations than a short mixing time. In the case of some sugars, l ike galactose in its pyranoid form, the TOCSY experiment can unravel spins up to H-4, but no further, because of the small coupling constant between H-4 and H-5.
The HSQC experiment shows correlations between directly attached carbons and protons, with 1H frequencies on the F2 axis and 1 3C on the F1 axis. Cross peaks show which protons are directly connected to which carbons. The simplest experiment for this purpose is a 1 3C,1H HETCOR experiment, but in the case of bacterial polysaccharides there is often a l imited amount of material available (only a few mg) and the use of inverse detected experiments l ike HSQC is then necessary. The anomeric configuration of the pyranoid sugars can also be determined by measuring the scalar couplings between H-1 and C-1 in a coupled HSQC spectrum. The JC - 1 , H - 1 coupling constants are ~ 160-165 Hz for the β-form and ~ 170-175 Hz for the α-form of the sugar.
The HSQC-TOCSY experiment is a modified HSQC experiment in which, prior to acquisition, a proton spin-lock is applied.
41 P . -E . J ans son , L . Kenne and G . Widmalm, Carbohydr . Res . , 188 (1989) 169-191 .
22

This will give a 1H,1 3C correlated spectrum, but in addition to the ordinary cross peaks, there will appear TOCSY cross peaks along the 1H spin systems at different 1 3C chemical shifts. The HSQC-NOESY experiment is a corresponding modified HSQC experiment with NOESY cross peaks.
Information about the sequence of the sugars in the polysaccharide may be obtained with HMBC and NOESY experiments. HMBC shows through-bond correlations between carbons and protons separated by two or three bonds. The magnetisation is transferred over the glycosidic linkage and we can identify the 3JC , H connectivities (see FIGURE 8a).
In the NOESY experiment the magnetisation is transferred through space and protons as far as 5Å apart can be detected and give cross peaks in the spectrum. Inter-residual NOE correlations are most important for the sequence determination (see FIGURE 8b).
C
O
HO
HOHO
H
OH
C
O
HO
OHO
OH
OH
H
O
HO
HOHO
H
OH
O
HO
OHO
OH
OH
H
a) b)
F I G U R E 8. (a) 3JC , H correlat ion (HMBC) and (b) inter-res idual NOE correlat ion.
N M R E x p e r i m e n t s f o r I n t e r a c t i o n S t u d i e s BINDING INTERACTIONS between proteins and small l igands have been studied by different NMR methods and some of the most common techniques used in this area are transferred NOE (trNOE)42, 43 and saturation transfer difference (STD) NMR spectroscopy.1 8 STD is a relatively new technique although the principle behind it was first described in the early 1960s.44
When a ligand binds to a protein, the NOEs change drastically, which leads to the observation of trNOEs. The tumbling times for small or medium sized molecules (Mw < 2000 Da) - such as l igands - differ from the tumbling times for large molecules (such as proteins). The small or medium sized molecules have short
42 G .M. C lo re and A .M. Gronenborn , J . Magn . Reson . , 48 (1982) 402-417 . 43 A . Poveda and J . J imé nez -Barbero , Chem. Soc . Rev . , 27 (1998) 133-143 . 44 S . Fo r sén and R . A . Hof fman , J . Chem. Phys . , 39 (1963) 2892-2901 .
23

tumbling times and therefore show positive, small negative or no NOEs - depending on molecular weight, shape and magnetic field strength.
Large molecules however always show large negative NOEs. When a ligand binds to a protein it behaves as a part of the protein and shows similar dynamics as the protein. This gives rise to large negative NOEs in the ligand. These strong NOEs are called trNOEs, which reflect the bound state of the ligand. To establish that a l igand binds to a protein the size and sign of the NOEs must be determined.
To further study the interaction between a ligand and a protein, the STD experiment45, 46 is very informative. As the name implies, the technique is based on intermolecular saturation transfer from the protein to the bound ligand. The irradiation of the protein is performed at resonances around -0.5 or 7 ppm, where no signals from the ligand are present. This is called the on-resonance experiment. A second experiment is performed with the irradiation frequency at about 30 ppm away, where neither the protein nor the ligand have any signals. Subtraction of the on-resonance spectrum from the off-resonance spectrum gives a difference spectrum (the STD spectrum). The protons in the ligand that are closest to the protein gives the strongest STD signals and this leads to information about the epitope. Protons not interacting with the protein will not experience any transfer and therefore not give rise to any signals at all in the STD spectrum. Another name for this is group epitope mapping (GEM). The principle of the STD experiment is shown in FIGURE 9.
45 M. Mayer and B . Meyer , J . Am. Chem. Soc . , 123 (2001) 6108-6117 . 46 M. Mayer and T .L . James , J . Am. Chem. Soc . , 124 (2002) 13376-13377 .
24

receptor-ligand complex
selective saturation pulse
free ligand
increasingsaturation
kon
koff
F I G U R E 9. Schematic picture of the saturat ion t ransfer difference (STD) spectroscopy phenomenon.
25

3.
R E S U LT S A N D D I S C U S S I O N S P a p e r I : S t r u c t u r a l S t u d i e s o f a n E x o p o l y s a c c h a r i d e P r o d u c e d b y S t r e p t o c o c c u s t h e r m o p h i l u s T H S LACTIC ACID BACTERIA that produce EPS with interesting properties are a growing area of investigation. A strain of Streptococcus thermophilus THS from yoghurt has been found to produce a viscous EPS when grown in skim milk. The production of EPS may give fermented products, l ike yoghurt and cheese, improved rheological properties.47 The rheology is dependent on chemical composition, monosaccharide linkages and side-chain groups, i .e. , the structure.2 The structural investigation of the EPS produced by S. thermophilus THS shows that i t consists of a pentasaccharide repeating unit of D-glucose and D-galactose. This has not been reported earl ier for a S. thermophilus species.
When EPS from lactic acid bacteria are studied with NMR spectroscopy some problems arise. Only small amounts of material can be used (in this case 2 mg) since it is very viscous. The molecules are large, 2 - 3 MDa, consisting of more than 1000 repeating units, compared to E. coli O-antigens that contain about 25 repeating units. The samples are viscous and the signals relax rapidly, which makes it difficult and sometimes impossible to obtain results from some two-dimensional NMR experiments, for example HMBC. When the EPS is composed of similar sugars, i .e. , glucose and galactose, the signals often overlap to a large extent, and although the NMR experiments were run at a 1H frequency of 800 MHz, the signals are sometimes difficult to assign. Assignments of the spin systems were performed by homo- and heteronuclear two-dimensional NMR techniques.
47 R . Tun ie r , W.H.M. van Cas te ren , P . J . Loo i j es t e i j n , H .A. Scho l s , A .G . J . Voragen and P . Zoon , Biopo lymers , 59 (2001) 160-166 .
26

Component analysis showed that the EPS from S. thermophilus THS contains only D-glucose and D-galactose, similar to other lactic acid bacteria in a molar ratio of 3:2. Methylation analysis revealed that two of the glucose residues were 4-substituted (the 1 3C glycosylation shifts show that these were residue B and D) and the third was 6-substituted (A). One galactose residue was terminal (E) and the other 3,4-substituted (C).
The H-2 chemical shifts (determined in the COSY experiment) are an indication of which sugar type is present, typically δH 3.25 and δH 3.45 for unsubstituted glucose and galactose, respectively. A , B and D are glucose residues and C and E are galactose residues.
From a coupled HSQC experiment the anomeric configuration of the sugars could be corroborated in agreement with JH - 1 , H - 2 data: JH - 1 , C - 1 ≈ 164 Hz revealed the β-configuration of residues B-E and JH - 1 , C - 1 = 172 Hz revealed the α-configuration of residue A .
In the spin system of residue A , the magnetization transfers smoothly between the protons throughout the spin system, indicating large coupling constants between all neighbouring protons and a pyranosidic sugar with gluco-configuration is determined.
The sequence of the sugars in the repeating unit of the EPS was largely determined by trans-glycosidic NOEs from a NOESY experiment with a mixing time of 50 ms. Inter-residual NOE correlations are shown in TABLE 1 and FIGURE 10.
T A BL E 1. Inter-residue correlat ions observed in the 1H,1H-NOESY spectrum of the EPS from S. thermophilus THS.a
Residue 1H (anomeric) Residue 1H NOE to
A 4.97 C 4.26 (s), 3.90 (m), 3.85 (w)
B 4.71 C 3.87 (m)
C 4.56 B 3.60 (s)
D 4.52 A 3.97 (s), 4.12 (w)
E 4.48 D 3.67 (s) a s = strong, m = medium, w = weak.
27

Residue A shows an NOE from its anomeric proton to H-4 in residue C , which indicates that residue A is l inked to position 4 in residue C . I t also shows NOEs to both H-6s in residue C . One weak (δH 4.12) and one strong (δH 3.97) NOE is present between the anomeric proton H-1 in residue D and both H-6s in residue A . A structural element consisting of three sugars, -D-A-C-, can be identified as →4)-β-D-Glcp-(1→6)-α-D-Glcp(1→4)-β-D-Galp from these results. This result rules out that residue E forms an open-chain cyclic acetal l inkage to residue C , analogous to the one found by Vinogradov and Bock. NOEs from the anomeric proton H-1 in residue C to H-4 in residue B and from H-1 in residue B to H-3 in residue C define the repeating backbone of the EPS as →4)-β-D-Glcp-(1→3)-β-D-Galp-(1→ . The terminal E residue shows a cross-peak in the NOESY spectrum between H-1 and a proton at δH 3.67. H-3 in residue B and H-3 and H-4 in residue D have this chemical shift . The only possibili ty in this case is that the NOE goes to H-4 in residue D since it is 4-substituted. This NOE is consistent with the termination of the side-chain in the repeating unit of the EPS.
4.454.504.554.604.654.704.754.804.854.904.955.00
3.50
3.60
3.70
3.80
3.90
4.00
4.10
4.20
4.30
A B C
1H /ppm
1H
/p
pm
A2
A6
C6
C6
C4
C3 C3
C5
B3
B5 B4
D3
D5
D4
D E
F I G U R E 10. Par t of the NOESY NMR spectrum with a mixing t ime of 50 ms of the EPS from S. thermophilus THS.
28

The structure was further corroborated by several correlations observed in the HMBC spectrum: H-1 in residue E to C-4 in residue D , H-1 in residue D to C-6 in residue A and C-1 in residue C to H-4 in residue B .
Thus, the structure of the EPS from S. thermophilus THS is defined below.
E D A β-D-Galp-(1→4)-β-D-Glcp-(1→6)-α-D-Glcp 1 ↓ 4 →3)-β-D-Galp-(1→4)-β-D-Glcp-(1→ C B
29

P a p e r I I : S t r u c t u r a l S t u d i e s o f t h e E x o p o l y s a c c h a r i d e P r o d u c e d b y P r o p i o n i b a c t e r i u m F r e u d e n r e i c h i i s s p . S h e r m a n i i J S IN SWISS-TYPE CHEESES, propionibacteria are used as dairy starters. The dairy propionibacteria are generally food-grade. Certain strains, Propionibacterium freudenreichii ssp. shermanii JS have been considered as potential probiotic organisms.48
Component analysis of the EPS revealed that i t only contains D-glucose. The 1H NMR spectrum, in corroboration with HSQC, showed two signals in the anomeric region, which means that the repeating unit contains two sugar residues. Both anomeric protons show JH - 1 , H - 2 ≈ 7.5 Hz (they are β-linked) and have the pyranoid ring form.
One of the glucoses was terminal, indicated by the fact that the 1 3C glycosylation shift displacements relative to the free sugar C-2-C-6 are all < 0.5 ppm. The residue with the anomeric carbon at δC 100.2 ppm must be a branch-point. The 1 3C glycosylation shifts of about 6 ppm for both C-2 and C-3 and β-effects for C-1 and C-4 show that this residue is →2,3)-β-D-Glcp-(1→ .
NOESY spectra with short mixing times (10, 25 and 50 ms) give information about both inter- and intra-residual correlations. In all of the NOESY spectra performed, the correlation between H-1’ and H-3’ was the most intense (FIGURE 11). In the case of the terminal sugar residue, a H-1 to H-2’ correlation was always present. In the HSQC-NOESY spectrum correlations between H-1’ and C-3’ and C-1 and H-2’ were observed, which indicates that the backbone of the EPS consists of β-(1→3)-linkages and that the terminal residue is β-(1→2)-linked to the backbone.
HMBC spectrum showed both the intra-residual correlation between C-1 and H-2 and between C-1’ to H-2’, 2JC , H ≈ 7 Hz. Inter-residual correlations observed in the HMBC confirmed the connectivity determined above with correlations of C-1’ to H-3’, C-1 to H-2’ and H-1 to C-2’.
The EPS contains two glucosyl residues in the repeating unit , and has the structure:
→3)[β-D-Glcp-(1→2)]-β-D-Glcp-(1→
48 C . Gran t and S . Sa lminen , In Lac t i c Ac id Bac ter ia – Microb io logy and Func t iona l Aspec t s ( 2 n d edn ) ; S . Sa lminen , A . von Wr igh t Eds , M. Decker , New York , 1998 , 589-601 .
30

5.005.15
3.5
3.6
3.7
3.8
3.9
4.0
4.1
4.2
3.4
H5'
H4'
H2'
H3'
H2'
H3
H5H2
1H /ppm
1H
/p
pm
F I G U R E 11. Par t of the NOESY NMR spectrum with a mixing t ime of 50 ms of the EPS from Propionibacterium freudenreichi i ssp. shermanii JS, with annoted correlat ions f rom anomeric protons. Atoms in the backbone sugar res idue are pr imed, whereas those in the terminal sugar res idue are unprimed. The chemical shif ts for H-1 and H-1’ are δH 4.99 and δH 5.15, respect ively.
The CPS from Streptococcus pneumonia type 3749 is composed
of a disaccharide repeating unit identical to the repeating unit of the EPS investigated above. Homopolysaccharides produced by lactic acid bacteria may have health promoting effects, for example some gluco- and fructo-oligosaccharides function as prebiotics.50 β-(1→3)-D-Glucans51 have been reported to have immuno stimulatory effects, which depend on branching pattern, higher-order structure and molecular weight. The EPS produced by Propionibacterium freudenreichii ssp. shermanii JS may have similar effects but this needs to be further studied.
49 A . Adeyeye , P . -E . J ansson , B . L indberg and J . Henr ichsen , Carbohydr . Res . , 180 (1988) 295-299 . 50 P . Monsan , S . Bozonne t , C . Albenne , G . Jou la , R . -M. Wi l l emo t and M. Remaud-S iméon , In t . Da i ry J . , 11 (2001) 675-685 . 51 B .H . Fa lch , T . Espev ik , L . Ryan and B .T . S tokke , Carbohydr . Res . , 329 (2000) 587 -596 .
31

P a p e r I I I : S t r u c t u r e o f a n A c i d i c P o l y s a c c h a r i d e f r o m t h e M a r i n e B a c t e r i u m P s e u d o a l t e r o m o n a s a l i e n a T y p e S t r a i n K M M 3 5 6 2 T C o n t a i n i n g t w o R e s i d u e s o f L - S e r i n e i n t h e R e p e a t i n g U n i t SEVEN BACTERIAL STRAINS isolated from sea-water samples collected from different locations in the Amursky Bay (Sea of Japan, Pacific Ocean) constitute a novel species, Pseudoalteromonas aliena . The isolates showed a cytotoxic effect on tumour cells.
Component analysis of the polysaccharide revealed 2-amino-2-deoxyglucose (GlcN), glucuronic acid (GlcA), 4-amino-4,6-dideoxyglucose (Qui4N) and an amino acid, serine (Ser). GlcN, Qui4N and GlcA have the D-configuration and Ser has L-configuration. 2-amino-2-deoxymannuronic acid (ManNA) was identified with the aid of NMR spectroscopy instead of component analysis.
In the 1H NMR spectrum, eight signals were present in the anomeric region but the HSQC NMR spectrum showed five signals in the anomeric region (left-hand bottom, FIGURE 12). This indicates a pentasaccharide repeating unit .
3.64.24.85.4
60
65
70
75
80
85
90
95
100
55
1H /ppm
13C
/ppm
F I G U R E 12. HSQC NMR spectrum of the polysaccharide f rom Pseudoal teromonas al iena type s t ra in KMM 3562T.
32

The five residues were denoted A-E with respect to decreasing δH of the anomeric signals at δH 5.57, 5.40, 5.02, 4.95 and 4.54. The signal at δH 4.51 originates from H-2 in the ManpNAcA residue and the two signals at δH 4.46 and 4.45 from two α-H in the serines. The corresponding 1 3C shifts are 56.8 and 56.9 ppm, respectively.
In residue A i ts C-4 resonance at δC 57.2 indicates an aminosugar. Residues B-D have C-2 resonances at δC 53.9, 56.7 and 54.3 which also indicate aminosugars. The 1H NMR spectrum showed the presence of four methyl signals from N-acetyl groups at δH 2.02-2.07.
JH - 1 , C - 1 values of 179 Hz for one of the GlcpNAc residues (B) and 177 Hz for the Quip4NAc residue (A) revealed that they are α-linked. The JH - 1 , C - 1 values of about 167 Hz showed that the second GlcpNAc residue (C), the ManpNAcA residue (D) and the GlcpA residue (E) are β-linked.
The sequence of the sugar residues in the repeating unit was determined by HMBC and NOESY experiments. Through-bond heteronuclear connectivities from an HMBC spectrum and inter-residue NOE connectivities from the NOESY defined all five glycosidic linkages. A BS-CT-HMBC experiment was also carried out to unravel the carbonyl region.
3.53.63.73.83.94.04.1
4.5
4.7
4.9
5.1
5.3
5.5
E1-B4
B1-D4
D1-C3
C1-E3
A1-E4
Ser-α,β
1H
/p
pm
1H /ppm
F I G U R E 13. Par t of the NOESY NMR spectrum with a mixing t ime of 40 ms of the polysaccharide from Pseudoal teromonas al iena
type s t ra in KMM 3562T.
33

The NOE correlations mentioned below are shown in FIGURE 13. From the anomeric proton H-1 in residue A , an NOE was observed to H-4 in residue E . This correlation and inter-residual HMBC connectivities that were found between both C-1 in residue A and H-4 in residue E and between H-1 in residue A and C-4 in residue E and showed that the Quip4NAc residue is l inked to position 4 in the GlcpA residue. The latter is further l inked to position 4 in residue B , the α-linked GlcpNAc, as indicated by an NOE between H-1 in residue E and H-4 in residue B . This is further corroborated by the following HMBC connectiveties: between H-1 in residue E and C-4 in residue B and between C-1 in residue E and H-4 in residue B .
Residue B shows an NOE from H-1 to H-4 in residue D . Thus, the GlcpNAc residue is l inked to position 4 in the ManpNAcA residue. An HMBC connectivity between C-1 in residue B and H-4 in residue D supports this. The ManpNAcA residue D shows an NOE from its anomeric proton H-1 to H-3 in residue C and an HMBC connectivity to C-3 in residue C . The ManpNAcA residue is thus linked to position 3 in the β-linked GlcpNAc residue. The sequence is t ied together by HMBC connectivities between the anomeric proton H-1 in residue C and C-3 in residue E and C-1 in residue C and H-3 in residue E . Thus the β-linked GlcpNAc is l inked to the position 3 in GlcpA which is consequently 3,4-substituted. This means that the Quip4NAc residue is a terminal residue on the polysaccharide repeating unit .
The uronic acid residues D and E are substituted via an amide linkage, by the serine residues, indicated by the chemical shifts of C-6 resonances. The chemical shifts of the C-6 resonances are 170.5 and 170.7 ppm and correlations between C-6 and H-4 as well as Ser-αH are observed in the BS-CT-HMBC spectrum (FIGURE 14) and confirm the substitution pattern of the serines.
34

3.94.04.14.24.34.44.5
170.2
170.4
170.6
170.8
171.0
13C
/p
pm
1H /ppm
Ser-αE
D
F I G U R E 14. Par t of the BS-CT-HMBC NMR spectrum of the polysaccharide from Pseudoal teromonas al iena
type s t ra in KMM 3562T.
The structure of the repeating unit of the polysaccharide produced by Pseudoalteromonas aliena type strain KMM 3562T is defined below as a pentasaccharide with two serine residues.
(6-N)-L-Ser (6-N)-L-Ser ⏐ E B ⏐ D C
→3)-β-D-GlcpA-(1→4)-α-D-GlcpNAc-(1→4)-β-D-ManpNAcA-(1→3)-β-D-GlcpNAc-(1→ 4 ↑ 1 α-D-Quip4NAc A
35

P a p e r I V : S t r u c t u r a l E l u c i d a t i o n o f t h e O - a n t i g e n i c P o l y s a c c h a r i d e f r o m t h e E n t e r o a g g r e g a t i v e E s c h e r i c h i a c o l i S t r a i n 1 8 0 / C 3 a n d i t s I m m u n o c h e m i c a l R e l a t i o n s h i p w i t h E s c h e r i c h i a c o l i O 5 a n d O 6 5 ESCHERICHIA COLI STRAIN 180/C3 is an enteroaggregative strain and was isolated in 1991 from an infant with persistent diarrhoea in León, Nicaragua. It could not be serotyped by conventional methods; the strain was non-typable in slide agglutination.
Component analysis revealed that the polysaccharide contains four major components: ribose, 3-amino-3,6-dideoxyglucose (Qui3N), galactose and 2-amino-2-deoxygalactose (GalN). All sugars had the D-configuration. Glucose and a heptose were also detected but these were attributed to the core. Methylation analysis showed that the monosaccharides were linked as follows: the ribose was 3-substituted, the Quip3NAc 2-substituted, the galactose 4-substituted as a pyranoside or 5-substituted as furanoside, which was determined by NMR and the GalpNAc 3-substituted. The ribose was a furanoside; the ring form was determined by the mass fragmentation pattern. The galactose was a pyranoside as determined by NMR.
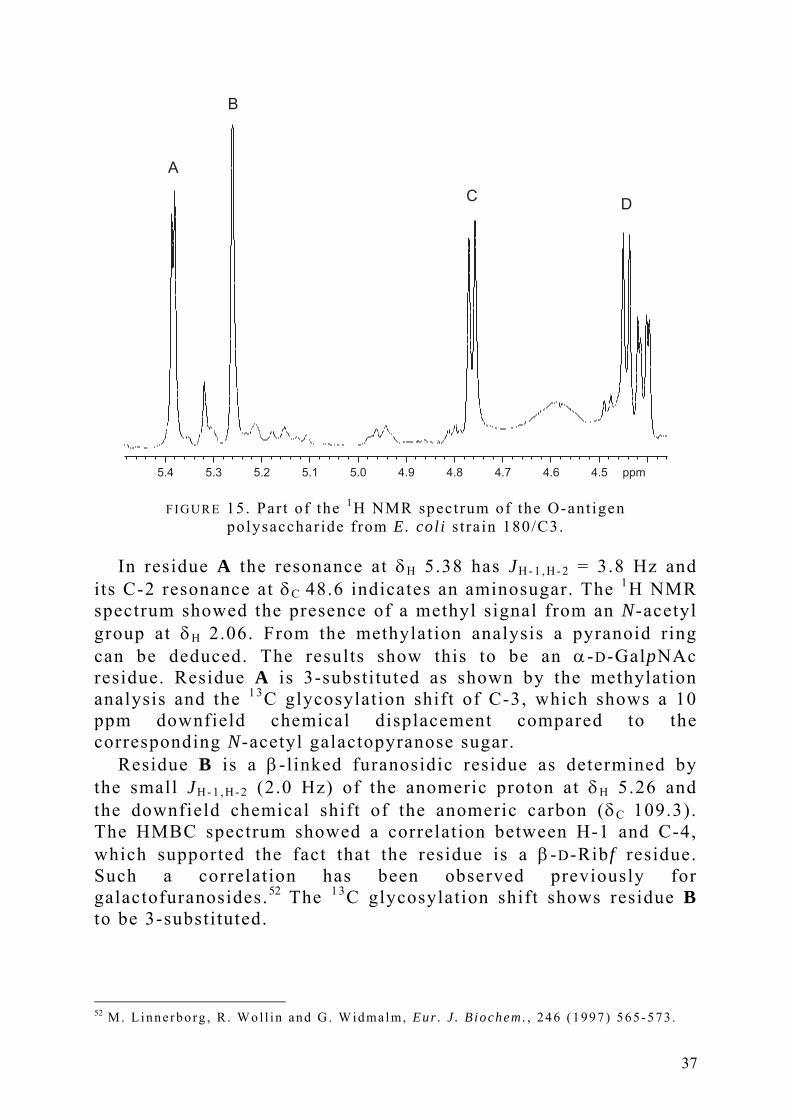
In the 1H NMR spectrum five signals were present in the anomeric region. Four of them originate from anomeric protons. The 1 3C NMR showed four signals in the anomeric region, 109.3, 105.9, 103.5 and 96.3 ppm. This indicates that the polysaccharide consists of tetrasaccharide repeating units. The four residues are denoted A-D with respect to decreasing δH of the anomeric signals at δH 5.38, 5.26, 4.76 and 4.44. The signal at δH 4.41 originates from the H-2 in the GalpNAc residue. These proton resonances are shown in FIGURE 15.
36
4.54.64.74.84.95.05.15.25.35.4 ppm
A
B
CD
F I G U R E 15. Par t of the 1H NMR spectrum of the O-ant igen polysaccharide f rom E. col i s t ra in 180/C3.
In residue A the resonance at δH 5.38 has JH - 1 , H - 2 = 3.8 Hz and
its C-2 resonance at δC 48.6 indicates an aminosugar. The 1H NMR spectrum showed the presence of a methyl signal from an N-acetyl group at δH 2.06. From the methylation analysis a pyranoid ring can be deduced. The results show this to be an α-D-GalpNAc residue. Residue A is 3-substituted as shown by the methylation analysis and the 1 3C glycosylation shift of C-3, which shows a 10 ppm downfield chemical displacement compared to the corresponding N-acetyl galactopyranose sugar.
Residue B is a β-linked furanosidic residue as determined by the small JH - 1 , H - 2 (2.0 Hz) of the anomeric proton at δH 5.26 and the downfield chemical shift of the anomeric carbon (δC 109.3). The HMBC spectrum showed a correlation between H-1 and C-4, which supported the fact that the residue is a β-D-Rib f residue. Such a correlation has been observed previously for galactofuranosides.52 The 1 3C glycosylation shift shows residue B to be 3-substituted.
52 M. L innerbo rg , R . Wol l in and G . W idmalm, Eur . J . B iochem. , 246 (1997) 565-573 .
37

The anomeric proton resonance of residue C at δH 4.76 has JH - 1 , H - 2 = 7.6 Hz. The resonance of C-3 at δC 56.3 showed the presence of an amino group. In the 1H NMR spectrum a methyl signal from an N-acetyl group was found at δH 2.07. This residue is β-D-Quip3NAc, which is 2-substituted.
Finally, residue D could be assigned to a 4-substituted β-D-Galp residue. Its anomeric proton resonance is present at δH 4.44 and has JH - 1 , H - 2 = 7.6 Hz.
The sequence of sugar residues in the repeating unit was determined by HMBC and NOESY experiments. Through-bond heteronuclear connectivities from an HMBC spectrum and inter-residue NOE connectivities from the NOESY defined all four glycosidic linkages.
From the anomeric proton H-1 in residue A , an NOE was observed to H-2 in residue C and also an inter-residue HMBC connectivity was found between the anomeric carbon C-1 in residue A and H-2 in residue C . The GalpNAc residue is thus linked to position 2 in the Quip3NAc residue. The latter is l inked to position 3 in residue B , the furanoid ribose, as indicated by an NOE between H-1 in residue C and H-3 in residue B . HMBC connectivities between both H-1 in residue C and C-3 in residue B and between C-1 in residue C and H-3 in residue B add further support. The sequence of the fragment -A-C-B- could be established from the these results.
Residue B shows an NOE from its anomeric proton to H-4 in residue D which indicates that residue B is l inked to position 4 in residue D . This is corroborated by the following HMBC connectivities: between H-1 in residue B and C-4 in residue D and between C-1 in residue B and H-4 in residue D . The sequence is t ied together by HMBC connectivities between the anomeric proton H-1 in residue D and C-3 in residue A and between the anomeric carbon C-1 in residue D and H-3 in residue A . Thus, the Galp residue is l inked to position 3 in the GalpNAc residue.
The sequence -C-B-D-A- can be deduced from the results above. Thus, the structure of the O-antigen polysaccharide from E. coli strain 180/C3 is as shown below. →2)-β-D-Quip3NAc-(1→3)-β-D-Ribf-(1→4)-β-D-Galp-(1→3)-α-D-GalpNAc-(1→
C B D A
38

In the 1H NMR spectrum minor peaks were found with an appearance similar to those in the repeating unit determined above. Upon a closer inspection they could be originating from protons on the C’ , B’ and D’ residues. The HSQC NMR spectrum identified signals (not shown at the contour level presented in FIGURE 16) indicating that the minor peaks come from the terminal repeating unit .
An HMBC correlation between C-1 and H-2 in residue C’ indicated a terminal β-D-Quip3NAc residue with respect to the chemical shift for H-2. The H-2 of the terminal β-D-Quip3NAc residue has 0.2 ppm lower chemical shift , compared to the H-2 of the β-D-Quip3NAc residue in the repeating unit . Furthermore correlations in HMBC and NOESY NMR spectra added further support to this assumption. In accordance with these findings and biosynthetic considerations, an α-linked N-acetylgalactosamine is presumed to be present at the reducing end of the biological repeating unit . Nevertheless signals at δH 4.80 and δC 102.5 probably originate from a β-linked N-acetylgalactosamine at the reducing end, closest to the core. This hypothesis is consistent with the case of the LPS from E. coli O6 which has a β-linkage between the O-chain and the core.53
Serological cross-reactions based on the O-antigens occur frequently between different bacterial isolates within the same species but may also occur between different species. The cross-reactivities are based on structural similarities in the cell wall associated antigens.
The structures of the O-antigenic polysaccharides from the enteroaggregative E. coli strain 180/C3 and the enterohemorrhagic E. coli O51 6 are identical but for the substitution pattern of the Quip3NAc. In E. coli strain 180/C3 the Quip3NAc is 2-substituted whereas in E. coli O5 the same residue is 4-substituted. Thus, these O-antigenic polysaccharides are positional isomers. Parts of their 1 3C-decoupled HSQC NMR spectra are shown in FIGURE 16.
53 L . Grozdanov , U . Zähr inger , G . B lum-Oeh le r , L . Brade , A . Henne , Y .A . Kn i re l , U . Schombe l , J . Schu lze , U . Sonnenborn , G . Go t t scha lk , J . Hacker , E .T . R ie t sche l and U. Dobr ind t , J . Bac te r io l . , 184 (2002) 5912-5925 .
39

98
100
102
104
106
108
110
98
100
102
104
106
108
110
96
96
4.85.0 4.6 4.45.25.4
a
b
1H /ppm
13C
/ppm
F I G U R E 16. The anomeric region of the HSQC NMR spectrum of the O-ant igen polysaccharide f rom
E. col i s t ra in 180/C3 (a) and E. col i O5 (b) .
In addition, the O-antigen from E. coli strain 180/C3 has some similarities with the O-antigen from E. coli O65.54 The E. coli O65 O-antigen contains as part of i ts l inear pentasaccharide repeating unit a similar structural element, thereby indicating that a common epitope could be present for the two polysaccharides.
O5 →4)-β-D-Quip3NAc-(1→3)-β-D-Ribf-(1→4)-β-D-Galp-(1→3)-α-D-GalpNAc-(1→ 180/C3 →2)-β-D-Quip3NAc-(1→3)-β-D-Ribf-(1→4)-β-D-Galp-(1→3)-α-D-GalpNAc-(1→ Part of O65 →2)-β-D-Quip3NAc⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅⋅→4)-β-D-GalpA-(1→3)-α-D-GlcpNAc-(1→
In order to further investigate similarities between the three O-antigens an ELISA was used. The curve in FIGURE 17a shows that the LPS from strain 180/C3 reacts nearly identical with the anti-180/C3-serum and anti-O5-serum, but the anti-O65-antibodies show a slightly weaker response. In FIGURE 17b the difference 54 M.B . Per ry and L .L . MacLean , Carbohydr . Res . , 322 (1999) 57 -66 .
40

between O65 and the two others becomes somewhat larger. FIGURE 17c shows that LPS O65 has less similarit ies to 180/C3 and O5 than 180/C3 and O5 have in comparison with each other.
1
0.1
0.01
10
A4
05
a b c
Dilution
10-3 10-4 10-610-5 10-7 10-8
1
0.1
0.01
10
1
0.1
0.01
10
10-3 10-4 10-610-5 10-7 10-8 10-3 10-4 10-610-5 10-7 10-8
F I G U R E 17. Enzyme immunoassay with rabbi t – ant i -E. col i 180/C3 (■ ) , O5 (● ) and O65 (□ ) sera t i t ra ted against LPS isolated from E.
col i s t ra in 180/C3 (a) , O5 (b) and O65 (c) .
Thus, the ELISA curves show that the anti-180/C3, as well as the anti-O5-sera, react similarly but differ from the anti-O65-sera. The cross-reactivity between E. coli O5 and O65 was observed before. The O-antigens show a structural relationship that makes it most logical to type the O-antigen of E. coli 180/C3 as a variant of E. coli O5 and not as a new type. The O5 group should be divided into subtypes O5ab and O5ac.
41

P a p e r V : I n t e r a c t i o n S t u d i e s o f a S a l m o n e l l a e n t e r i t i d i s O - a n t i g e n o c t a s a c c h a r i d e t o P h a g e P 2 2 T a i l S p i k e P r o t e i n b y N M R S p e c t r o s c o p y a n d M o l e c u l a r M o d e l i n g THE BACTERIOPHAGE P2255 belongs to a large group of phages that use surface carbohydrates as receptors. The homotrimeric (having three identical binding sites) tail spike protein recognizes (in this case) the O-antigen repeating units of the cell surface lipopolysaccharides from Salmonella typhimurium2 0 , 56 and Salmonella enteritidis .57, 58
P22 has a receptor destroying endorhamnosidase activity, cleaving the α-(1→3)-glycosidic linkage between Rha and Gal (in FIGURE 18 between Rha1 and Gal2). The cleavage procedure produces an octasaccharide and to lesser extent a tetrasaccharide. This receptor destroying enzymatic activity is well known for viruses that use carbohydrates as receptor targets.5 5
O
OO
OO
O
OH
HO
HO
OH
HO
HO
HO
HO
O
O
HO
Gal2
Man2Tyv2
Rha2
O
OO
O
O
HO
HO
OH
HO
HO
HO
HO
O
O
HO
Gal1
Man1
Tyv1
Rha1
OH
OH
OH
F I G U R E 18. Two repeat ing uni ts ( the invest igated octasaccharide) of the O-ant igen of the LPS from Salmonel la enteri t idis .
55 S . S te inbache r , S . Mi l l e r , U Baxa , N . Bud i sa , A . Wein t r aub , R . Seck le r and R. Huber , J . Mo l . B io l . , 267 (1997) 865-880 . 56 K . Bock , M. Melda l , D . Bund le , T . Iver sen , M.B . P in to , P . J . Garegg , I . Kvans t röm, T . Norberg , A . A . L indbe rg and S .B . Svenson , Carbohydr . Res . , 130 (1984) 35-53 . 57 U . Baxa , A . Cooper , A . W ein t raub , W. Pfe i l and R . Seck le r , Biochemis t ry , 40 (2001) 5144-5150 . 58 D .R . Bund le , H . Baumann , J . -R . Br i s son , S .M. Gagné , A . Zdanov and M. Cyg le r , Biochemis t ry , 33 (1994) 5183-5192 .
42

The pocket-like binding site of the phage protein is in the central part of the β-helix of the protein. The binding cleft is 21Å long and accommodates two repeating units (an octasaccharide). In this region, there is a stacking of aromatic side chains with pyranose rings in a hydrophobic manner and H-bonds to polar and ionic side chains. When the oligosaccharide binds, about 40% of its surface is accessible to the solvent relative to the free oligosaccharide. FIGURE 19 shows the crystal structure of bacteriophage P22 and the octasaccharide.
F I G U R E 19. The crystal s t ructure5 5 of bacter iophage P22 and the octasaccharide f rom Salmonel la enteri t idis .
43

In this study tr-NOESY and T-ROESY experiments were performed to compare the conformation of the ligand in its bound and free state. The ligand is the octasaccharide (two repeating units) derived from the Salmonella enteritidis O-antigen. When bound to the protein, the ligand showed a similar cross peak pattern in the tr-NOESY spectrum as it does in the T-ROESY spectrum of the free ligand. The conformation of the free and bound ligand do not seem to differ very much, which indicates that the ligand does not undergo any large changes when binding to the protein. Thus the conformation of the bound ligand and the major conformation of the free ligand in solution resemble each other.
When investigating the interaction between a ligand and a protein, mapping the binding epitope is important and the STD experiment is very useful. The protons in the ligand closest to the protein give the strongest STD signals. The narrow signals in the 1H NMR reference spectrum in FIGURE 20a originate from the ligand and the broad signals from the protein. The STD NMR spectrum is shown in FIGURE 20b. Both of the galactose residues show about the same STD intensity at their anomeric protons, at 5.20 and 5.22 ppm. The anomeric protons of the two mannose residues have the same chemical shift and show an intensive STD signal at 5.26 ppm. One of the rhamnoses seems to be closer to the protein than the other, Rha1 showing a STD signal at the resonance for i ts α-configurated anomeric proton, 5.10 ppm. Also, the H-4s of both the rhamnoses are involved in binding to the active site, showing STD peaks at 3.52 and 3.55 ppm. Both of the tyvelose residues are involved in the binding to the active site of the protein, indicated by the intensive STD signals at their methylene protons at 1.81 and 2.05 ppm. The methyl groups of the tyveloses are to some extent also involved, showing a STD signal at 1.26 ppm. The methyl groups of the rhamnoses also show strong STD, but are hard to distinguish. This agrees with results in previous studies on the Salmonella serogroups, where the 3,6-dideoxyhexoses were found to function as antigenic determinants.2 1
44

10 9 8 7 6 3 2 1 0 -15 4
~HDO
Chemical shift /ppm
b
a
10 9 8 7 6 3 2 1 0 -15 4
*
F I G U R E 20. (a) 1H NMR reference spectrum of a mixture of the P22 ta i l spike protein and the octasaccharide from of the O-ant igen of the LPS of Salmonel la enteri t idis in a molar ra t io of 1:30. (b) STD NMR spectrum of the same sample. The aster isk indicates an ar tefact .
45

46

C O N C L U S I O N S NMR SPECTROSCOPY is an invaluable tool in order to structurally investigate polysaccharides. A combination of one- and two-dimensional NMR experiments together with component analysis are useful in a structural elucidation. The sequence of the sugars in the repeating unit of the polysaccharides was determined by NOESY and HMBC experiments. Interaction studies were performed by trNOE and STD NMR experiments.
In the structural studies of the EPS produced by Streptococcus thermophilus THS, the NMR experiments were run at a 1H frequency of 800 MHz. The signals were stil l difficult to assign due to severe overlaps and the fact that this EPS is viscous. The resulting structure was shown to consist of a pentasaccharide repeating unit of only D-glucose and D-galactose, which has not been reported earlier for a Streptococcus thermophilus species.
The structural elucidation of the EPS produced by Propionibacterium freudenreichii ssp. shermanii JS revealed that i t is a homopolysaccharide, containing two glucose residues in the repeating unit . β-(1→3)-D-glucans are reported to have immuno stimulatory effects and therefore the EPS produced by Propionibacterium freudenreichii ssp. shermanii JS needs to be further studied to see if i t has some beneficial effects to human health.
The acidic polysaccharide from the marine bacterium Pseudoalteromonas aliena type strain KMM 3562T contains, in addition to a glucuronic acid and a mannuronic acid, two serine residues in the repeating unit . To unravel the carbonyl region a BS-CT-HMBC experiment was carried out.
The diarrhoeagenic Escherichia coli strain 180/C3 was investigated and is shown to be a variant of Escherichia coli O5, and therefore the O5 group should be divided into subtypes O5ab and O5ac.
Interaction studies on the octasaccharide derived from the Salmonella enteritidis O-antigen and the P22 tail spike protein showed similar conformations of the bound ligand and the free ligand in solution.
47

A C K N O W L E D G E M E N T S I WOULD LIKE TO THANK.. . GÖRAN WIDMALM for very good and inspiring supervision during these years and for the discussions about a.o. chemistry, l i terature and music TINA - grrr lpower, JENNIE - , ULRIKA - Mallorcabaren, TARA - l i l l schmü, JENS - Skuggan med klös, CLAS - Kung-Fu-Fight ing, ROBBAN - s tabi l , ANDREAS - pest , MAGNUS - kolera, HANNA, MICKE, CHRISTER, GLEN, KRISTINA, PIA and ROLAND for amusing years with work, Åre-trips and conferences with “THE WIDMALMS” JAN ERLING BÄCKVALL, BENGT LINDBERG and PER J. GAREGG for kind interest in our work MAGNUS JÄDER, JOHAN NILSSON, ANDREAS LARSSON and JENNIE JANSSON for important computer help everybody at the department of organic chemistry TORBJÖRN ASTLIND for always having time to help me with all the messy NMRs GLIBS for very interesting and funny annual meetings and money to conferences and projects ROBERT SECKLER, EINE HUTTUNEN, ANDREJ WEINTRAUB, ANDREI PEREPELOV and MAJA ROŠČ IĆ for fruitful collaborations GLEN ROCKWELL for l inguistic help of the thesis TINA LYCKNERT for many giving proof readings and strengthening afternoon talks in the blue chairs HANNA LIND for “my” sofa ☺ at inorganic chemistry and a lot of fun during the years all my friends, cousins, sysslings, aunts and uncles in both Sweden and Germany my lovely sister PÄLS, my gorgeous mother SMIRRE and my marvellous father PJ for löv and support and everything DANIEL ♥
48

”Jag v i l l a ldr ig b l i s tor , sa han bes tämt . ”Inte jag he l ler” , sa
Annika . ”Nä, de t är inge t a t t s tå e f ter” , sa Pippi . ”Stora människor har a ldr ig nånt ing ro l ig t . Dom har
bara en hoper t råkig t arbe te och fåniga k läder och l ik tornar och
kumminalskat t .”
A S T R I D L I N D G R E N
49

50