starch synthesis in potato tubers is regulated by post ... · pdf filepost-translational redox...
TRANSCRIPT

The Plant Cell, Vol. 14, 2191–2213, September 2002, www.plantcell.org © 2002 American Society of Plant Biologists
Starch Synthesis in Potato Tubers Is Regulated byPost-Translational Redox Modification of ADP-Glucose Pyrophosphorylase: A Novel Regulatory MechanismLinking Starch Synthesis to the Sucrose Supply
Axel Tiessen, Janneke H. M. Hendriks, Mark Stitt, Anja Branscheid, Yves Gibon, Eva M. Farré,and Peter Geigenberger
1
Max Planck Institute of Molecular Plant Physiology, Am Mühlenberg 1, 14476 Golm, Germany
Transcriptional and allosteric regulation of ADP-Glc pyrophosphorylase (AGPase) plays a major role in the regulation ofstarch synthesis. Analysis of the response after detachment of growing potato tubers from the mother plant revealedthat this concept requires extension. Starch synthesis was inhibited within 24 h of tuber detachment, even though thecatalytic subunit of AGPase (AGPB) and overall AGPase activity remained high, the substrates ATP and Glc-1-P in-creased, and the glycerate-3-phosphate/inorganic orthophosphate (the allosteric activator and inhibitor, respectively)ratio increased. This inhibition was abolished in transformants in which a bacterial AGPase replaced the potato AGPase.Measurements of the subcellular levels of each metabolite between Suc and starch established AGPase as the onlystep whose substrates increase and mass action ratio decreases after detachment of wild-type tubers. Separation ofextracts on nonreducing SDS gels revealed that AGPB is present as a mixture of monomers and dimers in growing tu-bers and becomes dimerized completely in detached tubers. Dimerization led to inactivation of the enzyme as a resultof a marked decrease of the substrate affinity and sensitivity to allosteric effectors. Dimerization could be reversed andAGPase reactivated in vitro by incubating extracts with DTT. Incubation of tuber slices with DTT or high Suc levels re-duced dimerization, increased AGPase activation, and stimulated starch synthesis in vivo. In intact tubers, the Suc con-tent correlated strongly with AGPase activation across a range of treatments, including tuber detachment, aging of themother plant, heterologous overexpression of Suc phosphorylase, and antisense inhibition of endogenous AGPase ac-tivity. Furthermore, activation of AGPase resulted in a stimulation of starch synthesis and decreased levels of glycolyticintermediates.
INTRODUCTION
ADP-Glc pyrophosphorylase (AGPase) catalyzes the con-version of Glc-1-P and ATP to ADP-Glc and inorganic pyro-phosphate (PPi), which is the first committed step in thepathway of starch synthesis (Figure 1) (Preiss, 1988; Martinand Smith, 1995; Smith et al., 1997). The higher plant en-zyme is a heterotetramer, consisting of two “regulatory”subunits (AGPS; 51 kD) and two slightly smaller “catalytic”subunits (AGPB; 50 kD) (Okita et al., 1990). AGPase plays amajor role in the regulation of starch synthesis. Studies withan Arabidopsis
AGPS
mutant have demonstrated that AGPaseis a key site for the control of starch synthesis in leaves(Neuhaus and Stitt, 1990). In potato tubers expressing anantisense
AGPB
construct, starch synthesis is decreased
when activity is decreased to
�
50% of the wild-type level(Müller-Röber et al., 1992; Geigenberger et al., 1999a).
Two mechanisms are known to regulate AGPase activity.First, AGPase is subject to transcriptional regulation, with ex-pression being increased by sugars (Salanoubat and Belliard,1989; Müller-Röber et al., 1990; Sokolov et al., 1998) anddecreased by nitrate (Scheible et al., 1997) and phosphate(Nielsen et al., 1998). This may allow starch accumulation torespond to changes in the carbon and nutritional status(Scheible et al., 1997; Stitt and Krapp, 1999). Second, AGPaseis exquisitely sensitive to allosteric regulation, being acti-vated by glycerate-3-phosphate (3PGA) and inhibited by Pi(Sowokinos, 1981; Sowokinos and Preiss, 1982; Preiss, 1988).
Increasing levels of phosphorylated intermediates typi-cally lead to a marked increase of the 3PGA/Pi ratio. There-fore, activation of AGPase by an increasing 3PGA/Pi ratioallows the rate of starch synthesis to be adjusted in re-sponse to changes in the balance between photosynthesisand Suc synthesis in leaves (Heldt et al., 1977; Herold,
1
To whom correspondence should be addressed. E-mail [email protected]; fax 49-331-5678408.Article, publication date, and citation information can be found atwww.plantcell.org/cgi/doi/10.1105/tpc.003640.

2192 The Plant Cell
1980; Stitt et al., 1987) and to changes in the balance be-tween Suc breakdown and respiration in nonphotosyn-thetic tissues (Stark et al., 1992; Hajirezaei et al., 1994;Geigenberger et al., 1997, 1998a, 2000; Jenner et al., 2001).
Several situations have been reported in which starchsynthesis changes independently of overall AGPase activityand reciprocally to the levels of phosphorylated intermediates(Geigenberger et al., 1994; Geiger et al., 1998; Trethewey etal., 1998, 2001; Geigenberger and Stitt, 2000). This fact indi-cates that there may be important gaps in our understand-ing of the regulation of starch synthesis. One such situationformed the starting point for the experiments describedhere. Detachment of growing potato tubers from the motherplant leads within 24 h to a threefold decrease of ADP-Glcand a 50% inhibition of starch synthesis, even thoughAGPase activity remains unaltered and the overall levels ofhexose phosphates and 3PGA increase (Geigenberger etal., 1994).
Because only the overall metabolite levels were mea-sured, there are various explanations for these observa-tions. One is that AGPase is being inhibited by a novelmechanism. Another is that the substrate supply for AGP-ase may be changing as a result of regulation of the enve-lope hexose phosphate:phosphate transporter (Kammereret al., 1998), the plastidic phosphoglucomutase (PGM)(Tauberger et al., 2000), or the envelope adenylate translo-
cator (Tjaden et al., 1998; Geigenberger et al., 2001). Theseare required to transport the carbon substrates and ATPfrom the cytosol to the plastid, where AGPase is located(Figure 1). Another possibility is that AGPase activity may beresponding to changes in plastid 3PGA/Pi ratio caused bychanges in the activity of the envelope triose phos-phate:phosphate translocator. This transporter is requiredto transmit changes of the 3PGA/Pi ratio from one compart-ment to the other (Borchert et al., 1989; Schott et al., 1995)(Figure 1).
Here, we show that the inhibition of starch synthesis aftertuber detachment occurs via a mechanism that depends onthe properties of the plant AGPase; additionally, we usednonaqueous fractionation to investigate the response of cy-tosolic and plastidic metabolite levels and define a uniquecrossover point at AGPase. We also demonstrate that de-tachment does not lead to a decrease of the plastid 3PGA/Pi ratio and has no effect on AGPB protein or overall AGP-ase activity; instead, it leads to post-translational inactiva-tion of AGPase via a reversible mechanism that involves re-dox-dependent dimerization of the ABPB subunits. Finally,we present evidence that this novel mechanism contributesto the regulation of starch synthesis in response to a rangeof treatments that modify the Suc level in tubers.
RESULTS
Inhibition of Starch Synthesis in Response to Tuber Detachment Is Abolished in Transgenic Tubers That Express a Heterologous AGPase
We used two independent approaches to determine thestep or steps in the pathway of starch synthesis that are thetargets for the novel mechanism that inhibits starch synthe-sis when tubers are detached from the mother plant. Thefirst approach asked whether specific regulatory features ofpotato tuber AGPase are essential for the inhibition of starchsynthesis. To answer this question, the response was com-pared in wild-type tubers and in transformants in whichendogenous AGPase was replaced largely by a nonplantAGPase.
The introduced construct encodes a form of the mono-meric
Escherichia coli
enzyme (glgC16) that is different in itskinetic properties from the plant enzyme (Stark et al., 1992).It was introduced (Lloyd et al., 1999) into the AGPase anti-sense line AGP93, which has low activity of the higher plantAGPase (Müller-Röber et al., 1992). AGPase activity in wild-type tubers was strongly dependent on 3PGA (Figure 2A).AGPase activity in the antisense line AGP93 was reduced by
�
90% and remained dependent on 3PGA. AGPase activityin the double-transformed lines AF1-28 and AF1-20 wasthreefold higher than that in AGP93 and was independent of3PGA (Figure 2A) and Pi (data not shown) (Lloyd et al.,1999).
Figure 1. Pathway of Suc-to-Starch Conversion and Its SubcellularCompartmentation in Potato Tubers.
1, Suc synthase; 2, UDP-Glc pyrophosphorylase; 3, fructokinase; 4,cytosolic phosphoglucomutase; 5, phosphoglucoisomerase; 6, plas-tidic phosphoglucomutase; 7, ADP-Glc pyrophosphorylase; 8, alka-line pyrophosphatase; 9, granule-bound starch synthase; 10, solu-ble starch synthase; 11, branching enzyme; 12, hexose phosphatetranslocator; 13, triose phosphate translocator; 14, adenylate trans-locator. TCA, tricarboxylic acid.

Redox Regulation of Starch Synthesis 2193
To measure the rate of starch synthesis, low concentra-tions of
14
C-Glc were injected into a fine borehole in growingtubers that were attached to the mother plant or 1 and 3days after detaching them from the mother plant by sever-ing the stolon (for details, see Geigenberger et al., 1994). Af-ter 1 h, the area around the injected label was removed andanalyzed to determine how much of the injected label hadbeen converted to starch (Figure 2B). Metabolite levels weremeasured in the same material (Figures 2C and 2D). Similarresults were obtained in an independent experiment inwhich
14
C-Glc was supplied for 30 min to discs cut from at-tached or detached tubers (data not shown).
In wild-type tubers, 34% of the label was incorporatedinto starch in attached tubers, decreasing to 21 and 15% intubers that had been detached for 1 and 3 days, respec-tively (Figure 2B). The inhibition of starch synthesis was ac-companied by an increase of hexose phosphates (Figure2C) and a slight increase of 3PGA (data not shown). Suc de-creased gradually, declining by 24 and 70% after 1 and 3days, respectively (Figure 2D). This finding confirms the re-sults described previously (Geigenberger et al., 1994).
In the antisense line AGP93, 19% of the injected label wasincorporated into starch in attached tubers, decreasing to8% in detached tubers (Figure 2B). Hexose phosphateswere high in attached tubers and remained high after de-tachment (Figure 2C), and Suc decreased even more slowlythan in wild-type tubers (Figure 2D).
In the double transformants AF1-20 and AF1-28, 24 to26% of the label was incorporated into starch in attachedtubers (Figure 2B). Detachment did not lead to a rapid inhi-bition of starch synthesis in these lines (Figure 2B) but in-stead led to a marked decline of hexose phosphates (Figure2C) and a rapid 70 to 80% decrease of Suc during the firstday after detachment (Figure 2D). There were no substantialchanges in the specific activities of the internal hexose phos-phate pools in the various genotypes after detachment (datanot shown), demonstrating that the different response in the
Figure 2.
Inhibition of Starch Synthesis in Response to TuberDetachment Is Abolished in Transgenic Tubers Expressing the
glgC16
Gene in an AGPB Antisense Background.
(A)
AGPase activity in wild-type tubers (WT), the parental antisenseAGPB line (AGP93), and four independent transgenic AF1 lines ex-pressing the
glgC16
gene in an antisense AGPase (AGP93) back-ground. AGPase activity was assayed using a standard protocol(Müller-Röber et al., 1992) in the absence (black bars) or the pres-ence of 1 mM (dark-gray bars), 2 mM (medium-gray bars), or 3 mM(light-gray bars) 3PGA.
(B)
to
(D)
Changes in the rate of starch synthesis and metabolite lev-els after tuber detachment. Tubers from 8-week-old wild-type po-tato plants, AGPase antisense plants (AGP93), and supertrans-formed plants expressing
glgC16
(AF1-28 and AF1-20) wereanalyzed either directly (black bars) or 1 day (gray bars) or 3 days(white bars) after detachment from the plant. To measure the rate ofstarch synthesis
(B)
, U-
14
C-Glc of high specific activity (
�
40 to 50kBq per tuber) was injected into a fine borehole of an otherwise in-tact tuber. The tubers then were incubated for 1 h, and a concentriccore of material around the borehole was extracted and analyzed todetermine
14
C incorporation into starch. The data are expressed as apercentage of the total label injected. The same samples were usedto measure the levels of hexose phosphates (sum of Glc-6-P, Fru-6-P, and Glc-1-P)
(C)
and Suc
(D
) by enzymatic analysis. Results aremeans
�
SE
of four tubers from different plants.FW, fresh weight.

2194 The Plant Cell
double transformants is not attributable to isotopic dilutionof the incoming label by internal pools. These results pro-vide strong genetic evidence that starch synthesis is inhib-ited after detachment by a regulatory mechanism that re-quires the presence of native potato tuber AGPase.
Separation of Tuber Material intoSubcellular Compartments
Regulated enzymes can be identified by perturbing the fluxthrough a metabolic pathway and measuring the resultingchanges in metabolite levels to identify the step or steps atwhich the substrate concentration(s) changes reciprocally tothe flux through the pathway (Rolleston, 1972). To allow theunbiased identification of the step(s) at which starch synthe-sis is inhibited after the detachment of wild-type tubers, weinvestigated changes in the subcellular levels of every me-tabolite from Suc to starch.
This was performed by nonaqueous fractionation, a tech-nique developed to analyze the subcellular compartmenta-tion of metabolites in leaves (Gerhardt and Heldt, 1984; Stittet al., 1989) and adapted recently for use with potato tubers(Farré et al., 2001). Tubers are frozen in liquid nitrogen toquench metabolism, homogenized in liquid nitrogen, ly-ophilized at low temperature, and resuspended in heptane.Enzymic reactions are blocked for the reminder of the frac-tionation procedure because water is absent. During lyoph-ilization, metabolites and proteins from a particular regionof the cell aggregate. The suspension then is ultrasoni-cated to generate particles that are partially enriched fordifferent material from subcellular compartments and thatcan be separated by nonaqueous density gradient centri-fugation.
Marker enzyme activities are measured to reveal how ma-terials from different cellular compartments distribute acrossthe gradient (Figure 3A). The vacuolar marker mannosidase(Boller and Kende, 1979) was highly enriched in the pellet,the cytosolic markers UDP-Glc pyrophosphorylase (UGP-ase; Kleczkowsky, 1994) and pyrophosphate:Fru-6-P1 phos-photransferase (MacDonald and Preiss, 1986) and the mito-chondrial marker citrate synthase were enriched in fractions0 and 1, and the plastidic marker AGPase (Kim et al., 1989)was enriched in the lightest fractions (fractions 2 and 3) (Fig-ure 3A) (Farré et al., 2001). This effect resembles the distri-bution in gradients of leaf material (Stitt et al., 1989); in thatearlier study, it also was shown that the distribution of en-zymes tracks the distribution of metabolites that are knownto be restricted to a particular compartment.
Metabolites are measured in each fraction, and theirsubcellular distribution are estimated by more-dimensionallinear regression. The estimated distribution can be com-pared with the overall content to estimate the metabolitecontent in each compartment (per gram of total dry weight)and with empirically determined values for the volume ofeach compartment (Farré et al., 2001) to estimate the sub-
cellular concentrations. This analysis yields estimates forthe vacuole, plastids, and cytosol. The estimated valuesfor the cytosol include the mitochondrial metabolites be-cause the cytosol and mitochondria are not separated (seeabove).
Changes in Subcellular Levels of Its Substrates Reveal That AGPase Is Involved in the Inhibition ofStarch Synthesis
The immediate substrates for AGPase are the pools of Glc-1-P and ATP in the plastid (Figure 1). Suc is degraded in thecytosol via Suc synthase (SuSy), UGPase, and PGM to formGlc-6-P, which is imported into the plastid via the Glc-6-P/Pi transporter and converted back to Glc-1-P by the plas-tidic PGM (Figure 1). Glc-6-P, Fru-6-P, and Glc-1-P weredistributed between the plastid and cytosol in attached tu-bers (Figures 3B to 3D) (Farré et al., 2001). The vacuole con-tained negligible hexose phosphates, except for traces ofGlc-1-P (Figure 3D) (Farré et al., 2001). Detachment led toan increase of Glc-6-P, Fru-6-P, and Glc-1-P in the cytosoland the plastid. The error bars are large for Glc-1-P becausethis metabolite is present at low levels, leading to analyticerrors that sum during the calculations. The estimated aver-age plastid Glc-1-P concentration increased fourfold from17 to 64
�
M (Table 1).ATP is imported from the cytosol via the envelope adenyl-
ate exchanger (Figure 1). A substantial proportion of the ad-enine nucleotides are located in the plastid (Figures 3E and3F), whereas uridine and guanidine nucleotides are locatedmainly in the cytosol of tubers (data not shown) (Farré et al.,2001), as is found for leaves (Dancer et al., 1990; Riens etal., 1991). Detachment led to a 50% increase of ATP in theplastid and a 50% decrease of ATP in the cytosol (Figure3E). ADP levels were not affected substantially after detach-ment (Figure 3F).
The ATP/ADP ratio was lower in the plastid than in thecytosol in attached tubers (1.2 compared with 3.3; datacalculated from Figures 3E and 3F), again resembling theratio in leaves (Stitt et al., 1982). The value attributed to thecytosol underestimates the actual cytosolic value, becausesignificant amounts of adenine nucleotides also are pres-ent in the mitochondria, and the ATP/ADP ratio in the mito-chondria is lower than that in the cytosol (Stitt et al., 1982).After detachment, the ATP/ADP ratio in the plastid in-creased to a value (1.9) similar to that estimated for the cy-tosol plus the mitochondria (2.0). The estimated plastidATP concentration increased after detachment from 179 to279
�
M (Table 1).The estimated plastidic concentrations of Glc-1-P and
ATP were in the range of the values determined in vitro forthe substrate affinity S
0.5
Glc-1-P (40 to 140
�
M) and S
0.5
ATP (120 to 190
�
M) of AGPase (Sowokinos and Preiss,1982; Ballicora et al., 1995). Detachment led to an increaseof the levels of both substrates, whereas the rate of starch

Redox Regulation of Starch Synthesis 2195
synthesis decreased. These results show that the inhibition ofstarch synthesis involves a mechanism that acts on AGPase.
Estimation of Mass-Action Ratios for EachEnzyme-Catalyzed Reaction and Transport Step between Suc and Starch Identifies AGPase as the Unique Site Involved in the Inhibition of Suc-to-Starch Interconversion after Tuber Detachment
The cytosolic concentrations of Suc, Fru, UDP-Glc, Glc-1-P,Glc-6-P, Fru-6-P, ATP, ADP, UTP, UDP, PPi, Pi, and 3PGA,as well as the plastidic concentrations of Glc-1-P, Glc-6-P,ADP-Glc, ATP, ADP, PPi, Pi, and 3PGA, are summarized inTable 1. The variation for some metabolites, including Glc-1-P and PPi in the plastid and ADP in the cytosol, was high.The variation for Glc-1-P was discussed above. The varia-tion in PPi was caused by the low level of PPi and by thefact that only a very small fraction of the total PPi was lo-cated in the plastid.
These results (Table 1) were used to calculate the ratiobetween the in vivo concentrations of the products and thesubstrates (termed the mass-action ratio) for every step be-tween Suc and ADP-Glc (Table 2). The theoretical equilib-rium constant (K
eq
; the ratio of product and substrate con-centrations at which the reaction is at its thermodynamicequilibrium and net flux is zero) is listed for comparison. K
eq
for the transport steps is set at unity. In attached tubers, themass-action ratios of the reactions catalyzed by SuSy,UGPase, and cytosolic PGM, the transport exchanges cata-lyzed by the Glc-6-P/Pi and ATP/ADP transporters, and thereaction catalyzed by plastidic PGM are close to their K
eq
.The mass-action ratios of fructokinase, AGPase, and inor-ganic pyrophosphatase are displaced from their K
eq
(Table2). These results resemble those reported previously for cy-tosol and plastids in leaves (Stitt et al., 1982, 1989) and forphloem sap (Geigenberger et al., 1993). Thus, our resultsprovide evidence for the reliability of the fractionation tech-nique and subsequent calculations.
The inhibition of starch synthesis after detachment wasaccompanied by a 40-fold decrease of the mass-action ra-tio for AGPase (Table 2). This was the result of an increase inthe concentration of the substrates Glc-1-P and ATP and adecrease in the concentration of the products ADP-Glc andPPi in the plastid (Table 1) (Geigenberger et al., 1994). Themass-action ratios for all of the other steps were unaltered
Figure 3.
Subcellular Analysis of Metabolite Levels in Attached and1-Day-Detached Wild-Type Tubers.
(A)
Marker enzyme distribution in a typical nonaqueous gradient oflyophilized wild-type tuber tissues. The values represent the enzymeactivity found in each fraction as a percentage of the total applied tothe gradient. The marker enzyme distribution in the fractions of eachgradient was used to calculate the distribution of metabolites in thesubcellular compartments. CS, citrate synthase; NAF, nonaqueousfractionation; PFP, pyrophosphate:Fru-6-P1 phosphotransferase.
(B)
to
(I)
Compartmentation of metabolites in potato tuber attachedto the plant (gray bars) or after 1 day of detachment (black bars).
Metabolites in the fractions of the nonaqueous gradients were ex-tracted with trichloroacetic acid before analysis of Glc-6-P
(B)
, Fru-6-P
(C)
, Glc-1-P
(D)
, ATP
(E)
, ADP
(F)
, 3PGA
(G)
, Pi
(H)
, and Suc
(I)
.The subcellular compartmentation of the metabolites was calculatedby more-dimensional regression analysis (best-fit method). The val-ues are standardized to tissue dry weight and give the mean of threeseparate gradients (means
�
SE
,
n
�
3).
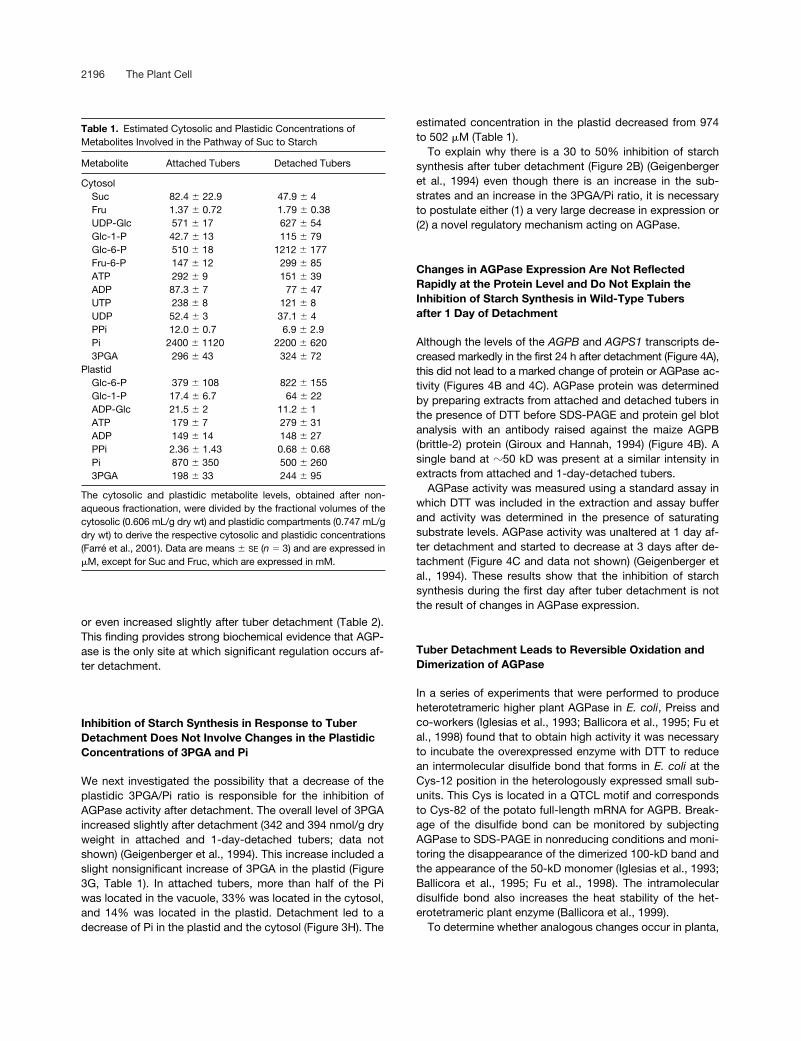
2196 The Plant Cell
or even increased slightly after tuber detachment (Table 2).This finding provides strong biochemical evidence that AGP-ase is the only site at which significant regulation occurs af-ter detachment.
Inhibition of Starch Synthesis in Response to Tuber Detachment Does Not Involve Changes in the Plastidic Concentrations of 3PGA and Pi
We next investigated the possibility that a decrease of theplastidic 3PGA/Pi ratio is responsible for the inhibition ofAGPase activity after detachment. The overall level of 3PGAincreased slightly after detachment (342 and 394 nmol/g dryweight in attached and 1-day-detached tubers; data notshown) (Geigenberger et al., 1994). This increase included aslight nonsignificant increase of 3PGA in the plastid (Figure3G, Table 1). In attached tubers, more than half of the Piwas located in the vacuole, 33% was located in the cytosol,and 14% was located in the plastid. Detachment led to adecrease of Pi in the plastid and the cytosol (Figure 3H). The
estimated concentration in the plastid decreased from 974to 502
�
M (Table 1).To explain why there is a 30 to 50% inhibition of starch
synthesis after tuber detachment (Figure 2B) (Geigenbergeret al., 1994) even though there is an increase in the sub-strates and an increase in the 3PGA/Pi ratio, it is necessaryto postulate either (1) a very large decrease in expression or(2) a novel regulatory mechanism acting on AGPase.
Changes in AGPase Expression Are Not Reflected Rapidly at the Protein Level and Do Not Explain the Inhibition of Starch Synthesis in Wild-Type Tubersafter 1 Day of Detachment
Although the levels of the
AGPB
and
AGPS1
transcripts de-creased markedly in the first 24 h after detachment (Figure 4A),this did not lead to a marked change of protein or AGPase ac-tivity (Figures 4B and 4C). AGPase protein was determinedby preparing extracts from attached and detached tubers inthe presence of DTT before SDS-PAGE and protein gel blotanalysis with an antibody raised against the maize AGPB(brittle-2) protein (Giroux and Hannah, 1994) (Figure 4B). Asingle band at
�
50 kD was present at a similar intensity inextracts from attached and 1-day-detached tubers.
AGPase activity was measured using a standard assay inwhich DTT was included in the extraction and assay bufferand activity was determined in the presence of saturatingsubstrate levels. AGPase activity was unaltered at 1 day af-ter detachment and started to decrease at 3 days after de-tachment (Figure 4C and data not shown) (Geigenberger etal., 1994). These results show that the inhibition of starchsynthesis during the first day after tuber detachment is notthe result of changes in AGPase expression.
Tuber Detachment Leads to Reversible Oxidation and Dimerization of AGPase
In a series of experiments that were performed to produceheterotetrameric higher plant AGPase in
E. coli
, Preiss andco-workers (Iglesias et al., 1993; Ballicora et al., 1995; Fu etal., 1998) found that to obtain high activity it was necessaryto incubate the overexpressed enzyme with DTT to reducean intermolecular disulfide bond that forms in
E. coli
at theCys-12 position in the heterologously expressed small sub-units. This Cys is located in a QTCL motif and correspondsto Cys-82 of the potato full-length mRNA for AGPB. Break-age of the disulfide bond can be monitored by subjectingAGPase to SDS-PAGE in nonreducing conditions and moni-toring the disappearance of the dimerized 100-kD band andthe appearance of the 50-kD monomer (Iglesias et al., 1993;Ballicora et al., 1995; Fu et al., 1998). The intramoleculardisulfide bond also increases the heat stability of the het-erotetrameric plant enzyme (Ballicora et al., 1999).
To determine whether analogous changes occur in planta,
Table 1.
Estimated Cytosolic and Plastidic Concentrations of Metabolites Involved in the Pathway of Suc to Starch
Metabolite Attached Tubers Detached Tubers
CytosolSuc 82.4
�
22.9 47.9
�
4Fru 1.37
�
0.72 1.79
�
0.38UDP-Glc 571
�
17 627
�
54Glc-1-P 42.7
�
13 115
�
79Glc-6-P 510
�
18 1212
�
177Fru-6-P 147
�
12 299
�
85ATP 292
�
9 151
�
39ADP 87.3
�
7 77
�
47UTP 238
�
8 121
�
8 UDP 52.4
�
3 37.1
�
4PPi 12.0
�
0.7 6.9
�
2.9Pi 2400
�
1120 2200
�
6203PGA 296
�
43 324
�
72Plastid
Glc-6-P 379
�
108 822
�
155Glc-1-P 17.4
�
6.7 64
�
22ADP-Glc 21.5
�
2 11.2
�
1ATP 179
�
7 279
�
31ADP 149
�
14 148
�
27PPi 2.36
�
1.43 0.68
�
0.68Pi 870
�
350 500
�
2603PGA 198
�
33 244
�
95
The cytosolic and plastidic metabolite levels, obtained after non-aqueous fractionation, were divided by the fractional volumes of thecytosolic (0.606 mL/g dry wt) and plastidic compartments (0.747 mL/gdry wt) to derive the respective cytosolic and plastidic concentrations(Farré et al., 2001). Data are means
�
SE
(
n
�
3) and are expressed in
�
M, except for Suc and Fruc, which are expressed in mM.

Redox Regulation of Starch Synthesis 2197
growing potato tubers were extracted in degassed SDS ex-traction buffer, separated immediately by nonreducing SDS-PAGE to maintain the in vivo redox status of the enzyme,and immunoblotted with antibodies raised against maize orpotato AGPB. To ensure the specificity of the antibodies,His-tagged potato AGPB and AGPS were overexpressed in
E. coli
and purified; their identities were confirmed by ma-trix-assisted laser-desorption ionization time-of-flight analy-sis. Then, they were applied in a concentration series, sepa-rated by SDS-PAGE, and immunoblotted. The antibodyraised against maize brittle-2 was highly specific for the po-tato AGPB subunit (Figure 5A).
Extracts from tubers that were attached to the plant gaveimmunopositive signals with the AGPB-specific maize anti-body at 50 and 100 kD (Figure 5B, lane 1). When 1-day-detached tubers were analyzed (Figure 5B, lane 2), the 50-kDband disappeared almost completely and the 100-kD bandbecame more intense. Incubation of the extracts in vitro with
DTT before SDS-PAGE completely converted the 100-kDband to the 50-kD band (Figure 5B, cf. lanes 3 and 4 withlanes 1 and 2). Similar results were obtained in repeated ex-periments over a period of months with separate batches oftubers. These results reveal that AGPB is present as a mix-ture of dimer and monomer in growing tubers and becomescompletely dimerized when the tubers are detached andSuc import from the mother plant is interrupted.
Establishment of a New Protocol to Extract and Measure AGPase Activity Reveals That the Changes in Redox State Are Accompanied by Changes in Activity and Kinetic Properties
We next investigated whether AGPase activity is altered whenAGPB becomes dimerized in planta. The standard protocol(Sowokinos, 1981; Müller-Röber et al., 1992; Ballicora et al.,
Table 2.
Estimated in Vivo Mass-Action Ratios of Enzyme and Transport Reactions Involved in Suc-to-Starch Conversion
Molar Mass-action Ratio
Reaction Formula Attached Tubers Detached Tubers Theoretical Equilibrium Constant (Keq)
SuSy(cytosol)
0.181 0.632 0.15 to 0.56
a
UGPase(cytosol)
1.48 3.21 3.2
b
Fructokinase(cytosol)
0.032 0.085 851
c
Phosphoglucoisomerase(cytosol)
3.47 4.05 2
c
PGM(cytosol)
11.9 10.5 19
c
Glc-6-P/Pitranslocator
2.04 2.97 1
d
3PGA/Pitranslocator
1.85 3.31 1
d
Adenylatetranslocator
0.359 0.961 1
d
PGM(plastid)
0.046 0.078 0.053
c
AGPase(plastid)
1.6
�
10
�
2
4
�
10
�
4
1
e
Inorganic pyrophosphatase(plastid)
0.323 0.373 1000
b
The data from Table 1 were used to calculate the in vivo molar mass-action ratios (ratio of product to substrate concentration) for each reaction.The theoretical equilibrium constant (ratio of product/substrate concentration) at which the net flux is zero) of each reaction is shown for compar-ison.a Geigenberger et al. (1993).b Weiner et al. (1987).c Stryer (1990).d Estimated value.e Kruger (1997).
Fru[ ] UDP-Glc[ ]×Suc[ ] UDP[ ]×
------------------------------------------------
Glc-1-P[ ] UTP[ ]×UDP-Glc[ ] PPi[ ]×
-------------------------------------------------
Fru-6-P[ ] ADP[ ]×Fru[ ] ATP[ ]×
-------------------------------------------------
Glc6P[ ]Fru6P[ ]
---------------------
Glc6P[ ]Glc1P[ ]
---------------------
Glc6P[ ]pl Pi[ ]cyt×Pi[ ]pl Glc6P[ ]cyt×
-----------------------------------------------
3PGA[ ]pl Pi[ ]cyt×Pi[ ]pl 3PGA[ ]cyt×
----------------------------------------------
ATP[ ]pl ADP[ ]cyt×ADP[ ]pl ATP[ ]cyt×
-------------------------------------------------
Glc1P[ ]Glc6P[ ]
---------------------
ADP-Glc[ ] PPi[ ]×Glc1P[ ] ATP[ ]×
------------------------------------------------
Pi[ ] Pi[ ]×PPi[ ]
--------------------------

2198 The Plant Cell
1995) for the extraction and assay of AGPase gave onlysmall and variable differences of activity between extractsfrom attached and detached tubers (data not shown). Thesolutions used in this protocol include DTT, and only the 50-kD band was found when aliquots from these extracts weresubjected to nonreducing SDS-PAGE (Figure 5C, lane 1).
To detect the large changes of activity of redox-modu-lated Calvin cycle enzymes that occur when chloroplasts orleaves are illuminated or darkened, it was necessary to de-velop very fast assay and extraction procedures (Laing etal., 1981; Wirtz et al., 1982). Therefore, we modified the pro-
tocol for the extraction and assay of AGPase by omitting allredox agents, presaturating all extraction and assay mediawith nitrogen, and decreasing the time between tissue dis-ruption and transfer into the assay to �1 min.
When an extract from growing tubers was prepared in thisway and subjected to nonreducing SDS-PAGE, a mixture ofbands at 50 and 100 kD (Figure 5C, lane 2) was found thatresembled a parallel sample that was extracted in SDS (Fig-ure 5C, lane 3). Large differences in AGPase activity werefound between extracts from attached tubers and tubersthat had been detached for 1 day (Figure 6). The differenceswere stable for at least 10 min in the assay conditions (datanot shown) but were lost rapidly if extract was incubated inthe absence of substrate (data not shown).
For routine determination of AGPase activity, the assaywas performed in the reverse direction (Figures 6A to 6C)using PGM and Glc-6-P dehydrogenase to couple Glc-1-Pformation in real time to NADP reduction. Detachment ledto a decrease of AGPase activity and the affinity for ADP-Glc when activity was assayed in the absence of allostericeffectors (Figure 6A). This inhibition was reversed com-pletely when 5 mM DTT was included during extraction andassay (Figure 6A).
Detachment also led to a decrease of activity and the af-finity for ADP-Glc when activity was assayed in the pres-ence of 300 �M 3PGA and 1200 �M Pi (Figure 6B), which liein the range of estimated concentrations in amyloplasts (Ta-ble 1). When assayed with limiting substrate concentrations,AGPase from detached tubers was strongly dependent onthe presence of 3PGA and was not activated until high con-centrations of 3PGA were present (Figure 6C). It also wasmore sensitive to inhibition by Pi than AGPase extractedfrom attached tubers (Figure 6C). Maximum activity in thepresence of high 3PGA and zero Pi was similar for attachedand detached tubers.
We also investigated whether AGPase activity was modi-fied when it was assayed in the ADP-Glc–producing direc-tion. A stopped assay was used, in which each individualsample was analyzed by HPLC to determine ADP-Glc (Fig-ures 6D to 6F). Control experiments showed that ADP-Glcwas not degraded during the incubation. Detachment led toa very marked inhibition of activity. The affinity for ATP wasdecreased when assayed in the presence of 200 �M 3PGA(Figure 3D) or 300 �M 3PGA plus 1200 �M Pi (Figure 6E).AGPase from detached tubers showed a strongly reducedsensitivity to 3PGA activation at physiological concentra-tions (Figure 6F).
To provide a routine microplate-compatible test forchanges in redox regulation in subsequent experiments,AGPase activity was measured in the ADP-Glc–cleaving di-rection in the presence of 600 �M ADP-Glc in the absence(Vsel) or presence of 5 mM DTT (Vred). Similar results wereobtained at a range of ADP-Glc concentrations between200 and 1000 �M (Figure 6A and data not shown). The ratiobetween the activities in these two assays (Vsel/Vred) istermed “activation.”
Figure 4. AGPB and AGPS Expression in Attached and 1-Day-Detached Wild-Type Tubers.
Attached (t0) and 1-day-detached (1 d) tubers were analyzed forsteady state mRNA levels of AGPB and AGPS1 (A), AGPase proteinafter reducing SDS-PAGE and immunoblot analysis with antiserumraised against the homologous maize brittle-2 protein (Giroux andHannah, 1994) (B), and overall activity of AGPase using a standardprotocol (Müller-Röber et al., 1992) including DTT in the extractionand assay buffers (C). Representative samples are shown in (A) and(B); in (C), results are means � SE (n � 4 extracts from separate tu-bers). FW, fresh weight.

Redox Regulation of Starch Synthesis 2199
Incubation of Tuber Discs with DTT to Redox Activate AGPase Leads to a Stimulation of Starch Synthesis and a Decrease of Phosphorylated Metabolites
To provide independent evidence that redox modulation ofAGPase can lead to large changes in the rate of starch syn-thesis, freshly cut discs from attached and 1-day-detachedtubers were incubated for 2 h in medium containing 0, 1, 5,or 10 mM DTT. To prevent postextraction modification ofAGPase, the discs were washed before extraction to pre-vent carryover of DTT into the extraction mixture. To mea-sure the rate of starch synthesis, 2 mM 14C-Glc was in-cluded in the incubation.
Increasing concentrations of DTT led to a progressive in-crease of AGPase activation (Figure 7A) and the proportionof AGPB that was present as a monomer (data not shown).There was a parallel increase of label incorporation intostarch (Figure 7B). AGPase activation (Figure 7A) and starchsynthesis (Figure 7B) were lower in discs from detached tu-bers than in discs from attached tubers in the absence ofDTT, and this difference was reversed partly by DTT. Whenthe results for all of the treatments were combined, a strongcorrelation was found between AGPase activation and labelincorporation into starch (Figure 7C).
The levels of hexose phosphates and label in the phos-phorylated intermediates were measured and used to calcu-late the specific activity of the hexose phosphate pool andthe absolute rate of starch synthesis. This calculation con-firmed that there is a strong correlation between AGPaseactivation and the rate of starch synthesis (data not shown).The stimulation of starch synthesis was accompanied by amarked decrease of the levels of hexose phosphates (Figure7D) and 3PGA (data not shown). This finding shows that thestimulation of starch synthesis by DTT is not caused by anadditional action besides redox activation of AGPase, such asa stimulation of Suc breakdown or a restriction of respiration.
Incubation of Tuber Discs Leads to a Decrease in AGPase Activation State, Which Is Prevented by Suc
Detachment of a tuber interrupts Suc import, leading to adecrease of Suc in the vacuole and the cytosol (Figure 3I). Itappeared plausible that changes in the import or level ofSuc might stimulate the post-translational inactivation ofAGPase. Therefore, experiments were performed to deter-mine whether redox modulation of potato tuber AGPase
Figure 5. Analysis of the Dimerization of AGPase.
(A) Demonstration of the specificity of the antiserum raised againstthe homologous maize brittle-2 protein (Giroux and Hannah, 1994)for the potato AGPB subunit. Similar amounts of heterologouslyoverexpressed His-tagged AGPB (B) and AGPS1 (S) proteins wereapplied in a concentration series (diluted 1 to 81 times), sepa-rated by SDS-PAGE in the presence of 4 mM DTT, and immuno-blotted.(B) Detachment leads to the dimerization of AGPase. Total proteinwas extracted from wild-type tubers, separated by SDS-PAGE, andexamined by immunoblot analysis using maize brittle-2 protein anti-body. Samples from attached tubers (t0; lanes 1 and 3) and 1-day-detached tubers (1 d; lanes 2 and 4) were prepared under anaerobicconditions in SDS and analyzed by SDS-PAGE in the absence ofDTT (lanes 1 and 2) or in the presence of 4 mM DTT (lanes 3 and 4).Each lane contained protein from �3 mg fresh weight of tubertissue.(C) Effect of different extraction methods on the dimerization ofAGPase after nonreducing SDS-PAGE and immunoblot analysiswith maize brittle-2 protein antibody. Lane 1, AGPase extracted us-ing a standard protocol as described by Müller-Röber et al. (1992);lane 2, AGPase extracted using a modified protocol omitting DTT(see Methods); lane 3, tissue extracted in SDS. Electrophoresis sam-ples of the different extracts were prepared in an argon atmosphereby mixing 100 �L of the respective extract (directly after its prepara-
tion) with 100 �L of argon-treated 2 � sample buffer lacking DTTand additional supplements. The sample was boiled immediately for5 min. Every lane contained the proteins originating from 1.5 mgfresh weight of tuber tissue. Representative sample blots using tu-bers from 8-week-old wild-type plants are shown.

2200 The Plant Cell
contributes to the regulation of starch synthesis in otherconditions in which the supply or level of Suc changes.
Geiger et al. (1998) reported that the addition of Suc to tu-ber discs stimulates starch synthesis and decreases the lev-els of glycolytic intermediate. In our experiments, when
freshly cut discs were incubated in the absence of Suc, theirinternal Suc content decreased by �30% during the first 90min and then stabilized (Figure 8A). AGPase activation de-creased between 30 and 120 min (Figure 8A). Inclusion of200 mM Suc in the medium prevented the decrease of the
Figure 6. Enzyme Kinetics of AGPase in Extracts from Attached or 1-Day-Detached Wild-Type Tubers Prepared Using the Modified Rapid Protocol.
(A) to (C) Reverse assay coupling Glc-1-P formation in real time to NADP reduction.(A) ADP-Glc saturation kinetics with or without DTT.(B) ADP-Glc saturation kinetics in the presence of 300 �M 3PGA and 1200 �M Pi.(C) 3PGA activation kinetics in the presence of 400 �M ADP-Glc with and without 1200 �M Pi.(D) to (F) Forward assay using a stopped assay and determination of ADP-Glc by HPLC.(D) ATP saturation kinetics in the presence of 200 �M 3PGA.(E) ATP saturation kinetics in the presence of 300 �M 3PGA and 1200 �M Pi.(F) 3PGA activation kinetics in the presence of 100 �M ATP.Results are from determinations with an extract from three pooled tubers for each condition. Similar results were obtained with extracts fromother pools of tubers. Closed circles, attached tubers; open circles, 1-day-detached tubers. FW, fresh weight.

Redox Regulation of Starch Synthesis 2201
internal Suc pool (Figure 8B) and the decrease of AGPaseactivation (Figure 8C). In a parallel experiment, 2 mM 14C-Glc was provided to tuber slices in the absence and pres-ence of 200 mM Suc. The increase in AGPase activation inthe presence of 200 mM Suc was accompanied by a 50%stimulation of label incorporation into starch and a decreaseof hexose phosphates (Table 3).
Inhibition of Starch Synthesis in Transgenic Tubers Overexpressing Suc Phosphorylase Involves the Inactivation of AGPase
In the second approach, heterologous expression of bacte-rial Suc phosphorylase in the cytosol was used as a tool todecrease Suc levels in planta (Trethewey et al., 2001). Sucphosphorylase converts Suc and Pi to Fru and Glc-1-P. Thisresembles the endogenous pathway for Suc breakdown viaSuSy and UGPase in not producing Glc but differs becauseit does not require PPi or produce UDP-Glc as an intermedi-ate. Critically, Suc phosphorylase has a far lower Km(Suc)(�1 mM; Silverstein et al., 1967) than SuSy (40 to 200 mM;Avigad, 1982).
Introduction of Suc phosphorylase led to a threefold tofourfold decrease of Suc (Figure 9A). Glc did not change(data not shown) (Trethewey et al., 2001), UDP-Glc de-creased slightly, PPi showed a slight and nonsignificant in-crease (data not shown), starch decreased (Figure 9B), andhexose phosphates (Figure 9C) and 3PGA (data not shown)(Trethewey et al., 2001) increased markedly. The inhibitionof starch synthesis in the transformants was accompaniedby a 30 to 50% decrease of overall AGPase activity (Figure9D). There also was a marked decrease of the AGPase acti-vation state (Figure 9E) and a marked decrease in the inten-sity of the 50-kD immunosignal relative to the 100-kD signalin nonreductive SDS-PAGE (Figure 9F).
Redox Inactivation of AGPase Correlates with the Decrease in Suc and the Inhibition of Starch Accumulation as Potato Plants Age
The experiments shown in Figures 2 to 9 used tubers from8-week-old plants, which contain high levels of Suc. A third
Figure 7. Redox Inactivation of AGPase and Inhibition of StarchSynthesis Both Can Be Reversed in Vivo by External Feeding of DTTto Wild-Type Tuber Discs for 2 h.
Discs were taken from attached tubers (black bars, black symbols)or 1-day-detached tubers (gray bars, gray symbols) and were incu-bated with 0 mM (circles), 1 mM (squares), 5 mM (triangles), or 10
mM (inverted triangles) DTT. Results are means � SE; n � 5 sepa-rate incubations with discs from separate tubers.(A) AGPase redox activation state (Vsel/Vred).(B) Labeling of starch after incubation of discs in 2 mM 14C-Glc (spe-cific activity of 18.5 kBq/�mol).(C) Starch labeling in relation to AGPase activation.(D) Hexose phosphate levels relative to AGPase activation. FW,fresh weight.

2202 The Plant Cell
approach investigated changes in wild-type tubers as themother plant ages. Tuber starch levels increased on a freshweight basis between 6 and 10 weeks but did not increasefurther between 10 and 12 weeks (Figure 10A). Suc levelsdecreased slightly between 6 and 8 weeks and decreasedmarkedly at 10 and 12 weeks (Figure 10B) (Mares andMarschner, 1980; Merlo et al., 1993). Hexose phosphate levelsincreased between 10 and 12 weeks, revealing that starchsynthesis is inhibited more strongly than Suc breakdown(Figure 10C). As shown previously (Mares and Marschner,1980; Merlo et al., 1993), SuSy activity decreased sharply(Figure 10D) and overall AGPase activity decreased mark-edly (assayed with DTT; Figure 10E) as the plant aged.There also was a marked decrease of AGPase activation(Figure 10F).
DISCUSSION
A Novel Mechanism Involving ReductivePost-Translational Regulation of AGPase InhibitsStarch Synthesis after Detachment of Tubers fromthe Mother Plant
The starting point for the experiments presented here wasthe observation (Geigenberger et al., 1994) that removal ofgrowing potato tubers from the mother plant leads within 1day to an inhibition of starch synthesis that cannot be ex-plained easily by allosteric regulation or by changes in theexpression of AGPase. This led us to suspect that theremight be a major gap in our understanding of the regulationof starch synthesis.
The first step in identifying the missing regulatory mecha-nism was to determine the step(s) at which flux was beinginhibited. Substitution of higher plant AGPase with a heter-ologous bacterial AGPase provided genetic evidence thatthe inhibitory mechanism required the presence of nativepotato AGPase (Figure 2). To provide biochemical evidencethat AGPase is the unique site at which flux is regulated, weperformed a systematic investigation of the subcellular lev-els of every metabolite in the pathway between Suc andstarch. This allowed the mass-action ratio (the product/sub-strate ratio) to be estimated for every enzyme and transport
Figure 8. Redox Inactivation of AGPase in Wild-Type Tuber Discs IsPrevented by External Feeding of Suc.
(A) Decrease in Suc content (gray circles) and AGPase activationstate (black circles) after cutting and incubating discs in buffer solu-tion in the absence of sugars for up to 4 h.(B) and (C) Suc levels (B) and AGPase redox activation (C) in discsincubated for 2 h with no sugars (gray bars) or 200 mM Suc (blackbars). After 2 h of incubation, subsamples of the discs were eitherwashed three times with Mes buffer before analyzing their Suc con-
tent or frozen directly in liquid nitrogen to measure AGPase activityunder selective (Vsel; �DTT) and reductive (Vred; DTT) assay con-ditions to calculate the activation state of the enzyme (Vsel/Vred).Suc levels and AGPase activation in tuber slices directly after cutting(white bars) are shown for comparison.Results are means � SE; n � 4 replicates from separate tubers. FW,fresh weight.

Redox Regulation of Starch Synthesis 2203
step between Suc in the cytosol and starch synthesis in theplastid. AGPase was the only step at which this ratio de-creased (by 2 orders of magnitude) after detachment of thetubers (Table 2). These genetic and biochemical experi-ments demonstrate (1) that the regulatory mechanism actsat AGPase and (2) that AGPase is the sole step involved inthe inhibition of starch synthesis after tuber detachment.
The second step was to exclude the possibility that knownmechanisms are responsible for the inhibition of AGPase ac-tivity. Measurements of 3PGA and Pi levels in the plastidshowed that the 3PGA/Pi ratio increased after detachment oftubers. Although there was a marked decrease in the tran-script levels for AGPB and AGPS1, there were no changes ofAGPB protein or AGPase activity. AGPase activity also changesmuch more slowly than transcript levels during the photo-period in Arabidopsis leaves (Sokolov et al., 1998) and po-tato tubers (Geigenberger and Stitt, 2000). Furthermore, stud-ies with antisense AGP transformants have shown that largechanges in the levels of AGP transcripts are required to pro-duce a significant change in AGPase activity (Müller-Röber etal., 1992) and that quite large changes of AGPase activity arerequired to produce a significant inhibition of starch synthesis(Müller-Röber et al., 1992; Geigenberger et al., 1999a).
The last step was to identify the missing mechanism.Since Sowokinos (1981) found that potato AGPase activityis stimulated by DTT, it has been included routinely in theextraction and assay media. An important clue was pro-vided by in vitro experiments by Preiss and co-workers inwhich they found that when the higher plant heterotet-rameric AGPase is overexpressed heterologously in E. coli,it is inactivated partly by an intermolecular disulfide bondthat forms between Cys residues in the N-terminal regionsof the AGPB subunits (Iglesias et al., 1993; Ballicora et al.,1995; Fu et al., 1998). Following up this clue, we investi-gated the biochemical and kinetic properties of AGPase ex-tracted from tubers in the absence of DTT using methodsadapted to allow rapid extraction in the absence of oxygen.
We found that AGPB exists as a mixture of monomersand dimers in growing tubers and is converted almost com-pletely to dimers in detached tubers. Furthermore, dimeriza-tion led to inactivation, as a result of a strong decrease ofthe substrate affinity, an increased requirement for and de-creased sensitivity to activation by 3PGA, and an increasedsensitivity to inhibition by Pi. The change in the kinetic prop-erties was especially dramatic when AGPase activity wasassayed in the forward reaction. Dimerization and inactiva-tion both were reversed by incubating extracts with DTT.
We interpret these results as evidence that the redoxchanges identified by Preiss and co-workers in heterolo-gously overexpressed AGPase operate as a regulatorymechanism in planta. However, biochemical analyses of theposition of the Cys bridges in AGPase holoenzyme isolatedfrom plants is needed to confirm this and to exclude thepossibility that other intermolecular or intramolecular Cysbridges also form in planta. Final genetic proof that thismechanism is responsible for the inhibition of starch synthe-
sis after detachment of tubers would require the productionof transformants in which the native AGPB is replaced by amodified form of AGPB in which Cys-82 is modified to pre-vent the formation of the intermolecular bridge. This experi-ment would require a mutant line in which transcription ofthe native AGPB is blocked to allow effective overexpres-sion without cosuppression of the modified construct. Atpresent, this is not possible in potato.
Redox Modulation of AGPase Allows Starch Synthesis to Be Increased, Whereas the Levels of Phosphorylated Metabolites Decrease
The dramatic changes in the kinetic properties of AGPaseproduced by redox modulation explain why starch synthesisis inhibited after tuber detachment even though there arechanges in the levels of substrates and allosteric effectorsthat would otherwise strongly stimulate starch synthesis. Toprovide independent biochemical evidence that redox acti-vation allows the rate of starch synthesis to be increased inplanta in the face of diametrically opposed changes in thelevels of substrates and metabolite effectors, discs wereprepared from tubers and incubated with DTT to achieve anartificial activation of AGPase. This led to a strong stimula-tion of starch synthesis and a marked decrease in the levelsof phosphorylated metabolites.
Post-Translational Redox Modulation of AGPase Also Makes an Important Contribution to the Regulation of Starch Synthesis in Tubers in Other Conditions
Other situations have been reported in which the rate ofstarch synthesis in potato tubers changes independently
Table 3. Suc Feeding Leads to Increased AGPase Redox Activation, Increased Starch Synthesis, and Decreased Hexose Phosphate Levels in Discs from Growing Tubers
Parameter Control200 mMSuc
Suc content (�mol/g fresh wt) 11.6 � 0.4 22.2 � 1.4AGPase activation [Vsel/Vred] (%) 33 � 2 49 � 3Starch synthesis (% of total 14C absorbed) 40 � 3 59 � 3Hexose phosphate level (nmol/g fresh wt) 225 � 20 148 � 11
Tuber discs were cut from an intact growing tuber and incubated inbuffer with and without 200 mM Suc for 2 h before they were eitherwashed three times to analyze their Suc content or frozen immedi-ately in liquid nitrogen to measure hexose phosphate levels and AGP-ase activity under selective (Vsel; �DTT) and reductive (Vred; DTT)assay conditions. In parallel samples, 2 mM 14C-Glc kBq/�mol spe-cific activity of 18.5) was provided to measure label incorporationinto starch. Results are means � SE (n � 3 replicate measurementson different discs from the same tuber).

2204 The Plant Cell
of overall AGPase activity and reciprocally to the changesin the levels of phosphorylated intermediates. When Sucis supplied to potato tuber slices (Geiger et al., 1998),starch synthesis is stimulated even though phosphory-lated intermediates, including 3PGA, decrease. When
Suc phosphorylase is overexpressed in growing tubers(Trethewey et al., 2001), starch synthesis is inhibited eventhough phosphorylated intermediates increase and AGPaseactivity decreases only slightly. The inhibition of starchsynthesis in tubers as the mother plant ages is not linked
Figure 9. Redox Activation State and Dimerization Degree of AGPase in Transgenic Tubers Expressing a Heterologous Suc Phosphorylase inthe Cytosol.
Tubers of 8-week-old wild-type plants and three independent transgenic lines were analyzed for Suc content (A), starch (B), hexose phos-phates (C), overall AGPase activity when DTT was included in the assay (Vred) (D), redox activation state of AGPase (Vsel/Vred) (E), anddimerization of AGPB protein in nonreductive SDS-PAGE using the maize brittle-2 antibody (F). Each lane contains the proteins originatingfrom 1 mg fresh weight. Results are means � SE; n � 3 tubers from different plants, except for (F), which documents a representative exam-ple. FW, fresh weight.

Redox Regulation of Starch Synthesis 2205
to a general decrease of phosphorylated metabolites(Merlo et al., 1993).
These results can be explained by post-translational re-dox modulation of AGPase. Feeding Suc to discs led to in-creased activation of AGPase and stimulated the rate ofstarch synthesis (Figure 8, Table 3), the heterologous ex-pression of Suc phosphorylase led to redox inactivation ofAGPase and a lower tuber starch content (Figure 9), and thegradual changes in the sink-source balance as the motherplant aged led to a decrease of AGPase activation. In thelatter two cases, there also was a 30 to 40% decrease ofoverall AGPase activity, but this alone (see above) is notlarge enough to explain the increase in hexose phosphatesand the decrease in the rate of starch synthesis.
Post-Translational Redox Modulation of AGPase Represents a Component in a Novel Regulatory Pathway That May Link the Rate of Starch Synthesis to Suc Supply
Although the evidence from the individual experiments iscircumstantial, together, our findings indicate that post-translational redox regulation of AGPase is part of a novelregulatory loop that links the rate of starch synthesis tochanges in the Suc supply. Detachment of tubers led to asudden interruption of phloem import, and aging of themother plant led to a gradual change in sink-source rela-tions. In both cases, the decreased rate of Suc import led toa decrease in the tuber Suc content (Figures 2D, 3I, and
Figure 10. Redox Activation State of AGPase in Tubers from Wild-Type Plants of Different Developmental Stages.
Tubers were harvested from 6-, 8-, 10-, and 12-week-old plants and analyzed for starch levels (A), Suc levels (B), hexose phosphate levels (C),SuSy activity (D), AGPase activity (Vred) (E), and redox activation state of AGPase (Vsel/Vred) (F). Results are means � SE; n � 3. FW, freshweight.

2206 The Plant Cell
10B) (Merlo et al., 1993), which correlates with the redox in-activation of AGPase. In these two treatments, of course,the import of other compounds as well as Suc was inter-rupted.
When tuber discs were incubated in the absence of Suc,the internal Suc level of the discs decreased and there wasa decrease of AGPase activation that could be reversed bythe addition of Suc (Figure 9, Table 3). Overexpression ofSuc phosphorylase to alter the Suc level and the preciseway in which Suc was metabolized in the tuber also led toredox inactivation of AGPase (Trethewey et al., 2001) (Figure10). Furthermore, in an antisense AGPB transformant line inwhich AGPase activity was reduced by 40 to 50%, therewas an increase of the tuber Suc level (Geigenberger et al.,2000) and a decrease in the dimerization and an increase ofthe activation state of AGPase (data not shown).
Figure 11 summarizes the data on tuber Suc content andAGPase activation from these different experiments, whichwere performed over a period of 2 years with many separatebatches of plants grown under different conditions. By com-paring the two parameters, a correlation coefficient of 0.85was found. These results provide very strong correlative ev-idence that post-translational redox modulation of AGPaseresponds to changes in Suc availability, although it is notpossible at this time to determine whether a specific influx, aparticular Suc pool, or changes in related metabolites pro-vide the immediate stimulus for the transduction pathway.
Candidate Components of Signal Transduction That Lead to the Redox Modulation of AGPase Have Yet toBe Identified and Confirmed
The light-dependent redox modulation of Calvin cycle en-zymes and enzymes involved in ATP synthesis and NADPHexport in chloroplasts is mediated by thioredoxin f andthioredoxin m, respectively (Schümann and Jacquot, 2000).Photosynthetic electron transport leads to a reduction offerredoxin, and reducing groups then are transferred byferredoxin:thioredoxin reductase to the thioredoxins, whichreact with their targets. Oxidation is thought to occur pri-marily by a reversal of this process. The reaction betweenthe thioredoxins and their individual targets is modulatedby changes in pH, Mg2, and the levels of their substratesor products, which alter the mid-redox potential of theCys groups on enzymes (Scheibe, 1991; Schümann andJacquot, 2000).
Ballicora et al. (2000) showed in in vitro experiments withheterologously overexpressed AGPase that DTT can be re-placed by thioredoxin f isolated from spinach. More studiesare needed to identify which thioredoxin(s) interacts withAGPase in planta and to determine how the flow of elec-trons to AGPase is regulated. Suc may lead to a large in-crease in the redox state of the plastid or, alternatively, mayact in some way to modulate the transfer of electrons fromNADPH over thioredoxin to AGPase.
Figure 11. Correlation between Tuber Suc Level and AGPase Redox Activation.
Data were taken from Figures 8B and 8C and Table 3 (feeding to tuber discs), Figures 9A and 9D (Suc phosphorylase–expressing tubers), andFigures 10B and 10F (aging of mother plant). Data from another tuber detachment experiment with wild-type plants and from transgenic tuberswith decreased expression of AGPase are included (means � SE; n � 4). FW, fresh weight.

Redox Regulation of Starch Synthesis 2207
Finally, our results raise the question of whether the post-translational regulation of AGPase contributes to the regula-tion of starch synthesis in other organs. Two lines of evi-dence indicate that this may be the case. First, anomaliessimilar to those that prompted our investigations on tubershave been reported for other tissues. Starch synthesischanged independently of overall AGPase activity and thelevels of phosphorylated intermediates after phloem trans-port was inhibited by detaching cotyledons of germinatingRicinus seedlings (Geigenberger and Stitt, 1991), whenspinach leaves were cold-girdled to decrease export (Krappand Stitt, 1995), when sugars were supplied to detachedspinach leaves (Krapp et al., 1991) or heterotrophic Che-nopodium rubrum suspension cells (Hatzfeld et al., 1990),and in leaves of transgenic tobacco plants when phloemtransport was inhibited by the phloem-specific expressionof E. coli pyrophosphatase (Geigenberger et al., 1996). In-triguingly, these all involve manipulations that were shownto or are likely to alter sugar export or sugar levels.
Second, the QTCL motif is conserved in the N-terminal re-gion of almost all sequenced AGPB genes of dicots and insome of the sequences available for monocots. Interest-ingly, the QTCL motif is absent in the major AGPB transcriptin cereal endosperm (Thorbjornsen et al., 1996; Hannah etal., 2001), which presumably encodes the cytosolic enzyme.This finding indicates that other mechanisms may be re-quired to link starch synthesis to the Suc supply during thelate stages of cereal grain filling.
The QTCL motif is absent in E. coli AGPase (glgC) and incyanobacterial AGPase and also is absent in the small sub-unit of the heterotetrameric Chlamydomonas reinhardtii AGP-ase (STA6) (Zabawinski et al., 2001), indicating that thispost-translational regulatory mechanism was developedduring the evolution of higher plants, possibly in parallel withthe evolution of the use of Suc as a transport metabolite andof thioredoxin function in photosynthetic eukaryotes.
This comparison also implies that it may be necessary toreevaluate the interpretation of experiments in which theoverexpression of glgC16 in higher plants was shown tolead to a strong stimulation of starch synthesis (Stark et al.,1992). This stimulation has been taken to date as evidencefor the crucial role of the allosteric properties of AGPase inthe regulation of starch synthesis. However, it is likely thatthis genetic switch also interrupts the post-translational re-dox inactivation of AGPase. The tuber detachment experi-ments with the double-transformed AF1 lines expressing anAGPase but lacking the QTCL motif and lacking redox mod-ulation suggest this possibility.
Regulation of AGPase at the Level of Expression,Post-Translational Regulation, and Allosteric Regulation
In conclusion, our results show that the redox modulation ofAGPase provides a powerful mechanism to regulate starchsynthesis. It was overlooked until now because it is reversed
rapidly under the standard methods used to extract and an-alyze AGPase protein and activity. Figure 12 presents amodel showing how this novel mechanism combines withknown mechanisms to coordinate AGPase activity, forminga network that allows starch synthesis in potato tubers torespond across a range of time scales to a variety of physio-logical and environmental stimuli.
Allosteric control by 3PGA and Pi operates in a time frame ofseconds to stimulate starch synthesis when phosphorylated
Figure 12. AGPase Is a Key Enzyme for the Regulation of StarchSynthesis in Potato Tubers.
Redox modulation of AGPase provides a novel mechanism thatcombines with known mechanisms to coordinate AGPase activity ina network that allows starch synthesis to respond across a range oftime scales to a variety of physiological and environmental stimuli.Allosteric control by 3PGA and Pi operates in a time frame of sec-onds to adjust the rate of starch synthesis to the balance betweenSuc breakdown and respiration. Post-translational redox modulationleads to changes in AGPase activity in a time frame of �30 to 60min. Activation occurs in response to factors directly or indirectly re-lated to increased Suc availability and leads to stimulation of starchsynthesis and decreased glycolytic metabolite levels. The signalingcomponents leading to redox modulation of AGPase are unknownand may involve thioredoxins as well as putative sugar sensors.Transcriptional regulation in response to changes of Suc allowsmore gradual changes in AGPase activity, which can require days todevelop. ABA, abscisic acid; ABI, abscisic acid insensitive; ISI, in-sensitive for Suc induction; SNF1, Suc non-fermenting1.

2208 The Plant Cell
intermediates increase and to inhibit it when they decrease.Existing physiological data indicate that allosteric controldoes not play a major role in coordinating the rate of starchsynthesis with the Suc supply. Instead, it adjusts the rate ofstarch synthesis to the balance between Suc breakdownand carbon use. Conditions in which changes in the 3PGA/Pi ratio correlate with starch synthesis include high temper-ature, during which an increasing rate of respiration leads toa depletion of phosphorylated intermediates and an inhibi-tion of starch synthesis (Geigenberger et al., 1998a), andmild water stress, during which post-transcriptional activa-tion of Suc phosphate synthase stimulates the resynthesisof Suc and leads to a decrease of phosphorylated interme-diates and an inhibition of starch synthesis (Geigenbergeret al., 1997, 1999b).
Post-translational redox modulation leads to changes ofactivity in a time frame of �30 to 60 min. Activation occursin responses to factors related directly or indirectly to an in-crease in Suc availability and leads to a stimulation of starchsynthesis and a decrease in the levels of phosphorylated in-termediates when Suc increases. This could have two im-portant consequences.
First, lower levels of phosphorylated intermediates will fa-vor Suc breakdown via the reversible reactions catalyzed bySuSy and UGPase (Geigenberger and Stitt, 1993), will re-lieve feedback inhibition of fructokinase by Fru-6-P (Renzand Stitt, 1993), and will decrease Suc resynthesis by inhib-iting Suc phosphate synthase (Geigenberger et al., 1999b).Second, stimulation of starch synthesis via a mechanismthat simultaneously decreases glycolytic intermediate levelsmay channel Suc toward starch without this leading to astimulation of glycolysis and respiration. This is importantbecause internal oxygen concentrations decrease to verylow levels during tuber bulking (Geigenberger et al., 2000),and it is important to increase the flow of carbon from Sucto starch without a large increase in oxygen consumption.
Transcriptional regulation in response to changes in Suc al-lows more gradual changes in AGPase activity that may re-quire days to develop. For example, during diurnal rhythms(Geigenberger and Stitt, 2000) and after detachment of tubers(Figure 4), the levels of the AGPS and AGPB transcriptschange within 12 to 24 h, but AGPase activity remains highfor several days (Geigenberger et al., 1994). Changes in tran-scription also may be responsible for the gradual decrease ofoverall AGPase activity on tubers as the mother plants age(Figure 10) and the slightly lower expression of AGPase in tu-bers that express Suc phosphorylase (Figure 9).
METHODS
Plant Material and Growth Conditions
Wild-type potato plants (Solanum tuberosum cv Desiree) were ob-tained originally from Saatzucht Fritz Lange (Bad Schwartau, Ger-
many). The generation and characterization of transgenic potatoplants expressing antisense ADP-Glc pyrophosphorylase (AGPase),a heterologous gene from a bacterial mutated AGPase enzyme(glgC16 from Escherichia coli), and a bacterial Suc phosphorylase inthe cytosol of their tubers were described by Müller-Röber et al.(1992), Lloyd et al. (1999), and Trethewey et al. (2001), respectively.
Potato plants were grown in a growth chamber (350 �mol·m�2·s�1
irradiance at 20C and 50% RH) under a 14-h-light/10-h-dark regimein 3-L plastic pots in soil supplemented with Hakaphos grün (100 gper 230 L of soil; BASF, Ludwigshafen, Germany) and were watereddaily from the top with tap water, or they were grown in a greenhouseduring the summer (16-h-light/8-h-dark regime at 20/18C day/nightand 60% RH) with supplementing light (62 400-W AgroSonT lampsper 100 m2) in 20-cm-diameter plastic pots with a 2:1 mixture of soilto sand and were irrigated daily with water (�200 mL per plant perday) containing nutrients (0.7 g/L Hakaphos rot [8% N, 12% P, 24% K,and 4% Mg]).
Reagents
Enzymes, reagents, and kits for molecular biology were purchasedfrom Roche (Mannheim, Germany), MBI Fermentas (St. Leon-Rot,Germany), Qiagen (Hilden, Germany), and Stratagene (Heidelberg,Germany). Biochemical enzymes were purchased from Roche andSigma (München, Germany), chemicals were purchased from Roche,Merck (Darmstadt, Germany), and Sigma, and reagents for SDS-PAGE were purchased from Bio-Rad (München). Radiochemicalsand x-ray films were purchased from Amersham-Buchler (Braun-schweig, Germany). If not stated otherwise, enzymes were grade IIand chemicals were of analytical purity.
Tuber Detachment Experiments
Tuber detachment experiments were performed as described byGeigenberger et al. (1994) and were started in the second half of thelight period (6 to 8 h into the photoperiod) using 8- to 9-week-oldplants. Tubers of these plants contained high Suc synthase activity,which is indicative of rapidly growing tubers.
Sampling of Potato Tuber Tissue
Immediately after harvesting the tuber, a cylinder was cut perpendic-ular to the stolon-apex axis. Tuber slices that were incubated subse-quently with radioactive Glc were cut (8 mm diameter, thickness of 2mm) and preincubated in 10 mM Mes-KOH, pH 6.5, for 10 to 20 min.Tuber tissues sampled for direct analysis or for analysis after non-aqueous fractionation were sliced immediately into liquid nitrogenand stored at �80C until use. Metabolic gradients have been re-ported for potato tubers along the stolon-apex axis (Merlo et al.,1993) and across the tuber (Geigenberger et al., 2000). Therefore, tu-ber slices were taken from the middle of the tuber, avoiding the outer3 mm and the tuber skin.
Labeling Experiments with Intact Tubers
Radioactive labeling experiments were performed essentially as de-scribed by Geigenberger et al. (1994). To measure carbon fluxes inwhole tubers, a 1-mm borehole was made (perpendicular to the sto-

Redox Regulation of Starch Synthesis 2209
lon-apex axis) into intact or detached tubers using a hypodermicneedle. Radioactive Glc (U-14C-Glc; specific activity of 11.5 GBq/mmol) was injected into the borehole, the end was sealed, and after1 h, a concentric cylinder of 8 mm diameter surrounding the injectionhole was cut and frozen in liquid nitrogen. The frozen tissue wasweighed before extraction.
Labeling Experiments with Tuber Slices
A cylinder was cut perpendicular to the stolon-apex axis to prepareslices (8-mm diameter, thickness of 2 mm) from intact or detachedtubers. Slices were washed briefly in 10 mM Mes-KOH, pH 6.5, blot-ted with tissue paper, and then incubated for 30 min or 2 h at 22C in4 mL of incubation buffer containing 2 mM radioactive U-14C-Glc(18.5 kBq/�mol) or U-14C-Suc (18.5 kBq/�mol) in Erlenmeyer flasksshaken at 90 rpm to maintain aerobic conditions. The incubationtimes, buffer compositions, and labeled compounds are specified inthe figure legends. For each treatment, 10 slices from four differenttubers were used (2 slices per replicate sample). After incubation, thediscs were washed (three times for 30 s each) in nonlabeled incuba-tion buffer, blotted briefly with paper, and frozen in liquid nitrogen.
Fractionation of 14C-Labeled Potato Tissue
Tuber material was extracted with 80% (v/v) ethanol at 80C (1 mL/0.1 g fresh weight) and reextracted in two subsequent steps with50% (v/v) ethanol (1 mL/0.1 g fresh weight at each step). The com-bined supernatants were dried under an air stream at 40C, taken upin 1 mL of water, and separated into ionic (phosphate esters, organicacids, and amino acids) and neutral (Suc, Glc, and Fru) componentsby ion exchange and thin layer chromatography as described byGeigenberger et al. (1997). The residue was analyzed for label instarch as described by Merlo et al. (1993).
Nonaqueous Fractionation of Tuber Tissue
Potato tuber tissue was fractionated as described by Farré et al.(2001), except that (1) five fractions (termed pellet, 0, 1, 2, and 3)were collected from the nonaqueous density gradients and (2) themass of the dried total aliquot was determined to allow the results tobe expressed per gram dry weight. The subcellular distribution ofeach metabolite was estimated from the distribution of marker en-zymes and the metabolite between the gradient fractions, based onequations given by Stitt et al. (1989). The equations were solved to abest fit by the method of least-squares values using an algorithmbased on the solver function incorporated into Excel version 5.0 (Mi-crosoft, Redmond, WA). This allowed direct and rapid calculationand evaluation on one spreadsheet using recovery values (percent-age of the value found in the total aliquot) of marker enzymes and ofmetabolites to make the calculations and regressions for subcellularcompartmentation. It also allowed more efficient detection of ana-lytic errors than earlier methods.
By using a higher number of fractions (five) than the number of an-alyzed compartments (three), the equation system is overdeter-mined, making the results for subcellular distribution more reliableand robust. Even when the data for one fraction is deleted, the re-gressions can predict a reliable value (data not shown). Estimates forthe subcellular volumes of growing potato tuber tissue derived fromplanimetry (Farré et al., 2001) were used to calculate the metabolite
concentrations in each compartment (vacuole, 3.384 mL/g dryweight; plastid, 0.747 mL/g dry weight; cytosol, 0.655 mL/g dryweight). There were no significant changes in subcellular volumes af-ter detachment (data not shown).
For each gradient, an aliquot of the initial homogenate was takento determine the recovery values of enzymes applied to the gradientand to determine the dry weight of the potato tissue. The recoveriesof enzymes were 72, 82, 89, 78, and 81% for mannosidase, AGPase,UDP-Glc pyrophosphorylase, pyrophosphate:Fru-6-P1 phospho-transferase, and citrate synthase, respectively. Analysis of starch lev-els in the different fractions revealed that starch is 100% located inthe plastids. As has been documented for leaves (Stitt et al., 1989),the distribution of enzymes tracks the distribution of metabolites thatare known to be restricted to a particular compartment.
Metabolite Analysis
Tuber material was extracted either directly or after subcellular frac-tionation (see above) in trichloroacetic acid as described by Weineret al. (1987) and Jellito et al. (1992); the reliability of the method hasbeen documented by Jellito et al. (1992). Nucleotides were mea-sured by HPLC according to Geigenberger et al. (1997) using a Par-tisil-10-SAX ion exchange column (4.6 mm � 250 mm; Whatman)and detection at 254 and 230 nm. The nucleotides were identified bycomparison of the elution times of authentic standards and the 254/230-nm ratio of absorbance. The metabolite amount was calculatedfrom the peak area using Datasystem 450 MT2 software (Kontron,Munich, Germany).
The levels of Glc-1-P, Glc-6-P, Fru-6-P, UDP-Glc, glycerate-3-phosphate (3PGA), Pi, Suc, and starch were measured as describedby Geigenberger et al. (1998b). Inorganic pyrophosphate (PPi) andADP-Glc were measured in the 96-well format by enzymatic cyclingassays (Gibon et al., 2002), except that arsenolysis was not used inthe case of PPi determination. Instead, aliquots of the samples werepreincubated for 30 min in the presence of glycerokinase, glycerol-3-phosphate oxidase, and glycerol and then heated at 95C for 20 minbefore determination of PPi, to decrease the amount of interferingmetabolites.
Analysis of Overall Enzyme Activities in Potato Tubers
The activities of Suc synthase and AGPase were measured in de-salted tuber extracts as described by Merlo et al. (1993). As detailedbelow, AGPase also was measured using a newly developed protocol.
Analysis of AGPase Activity and Kinetic Properties
A special extraction and assay protocol was developed to reveal thein planta AGPase activity and redox activation state. The extractionbuffer (50 mM Hepes-KOH, pH 7.8, and 5 mM MgCl2) was degassedby bubbling with nitrogen. Tuber discs were prepared rapidly (seeabove) and frozen in liquid nitrogen. The discs (�100 mg freshweight) were homogenized to a powder under liquid nitrogen, ex-tracted with 1 mL of extraction buffer, and centrifuged for 30 s at10,000g at 4C; 10 �L of the supernatant was used for the AGPaseassay. The entire procedure lasted �2 min. AGPase activity was fol-lowed on line and was linear for up to 30 min. The 96-well format al-lowed us to perform several comparative measurements of differentsamples at the same time.

2210 The Plant Cell
AGPase assay in the pyrophosphorolysis direction was performedat 30C in a total volume of 200 �L containing 50 mM Hepes-KOH,pH 7.8, 5 mM MgCl2, 10 �M Glc-1,6-bisP, 0.6 mM NADP, 2.5 mMNa-PPi, 1 unit/mL phosphoglucomutase (from rabbit muscle), 2.5units/mL Glc-6-P dehydrogenase (from yeast), and varying concen-trations of ADP-Glc (0 to 1 mM) with or without 5 mM DTT. The assaylacking DTT was termed Vsel, and the assay containing 5 mM DTTwas termed Vred.
The reaction was started by adding freshly prepared extract. Par-allel assays were performed lacking either Na-PPi or ADP-Glc. Blankactivity was negligible for at least 40 min and was subtracted whendetected. To analyze the kinetic properties of AGPase, the activator3PGA or the inhibitor Pi was included in the assay mixtures as indi-cated in the figure legends. To redox activate AGPase, extract waspreincubated for 10 min at 4C by adding 1 mM ADP-Glc and 5 mMDTT before transfer to the Vred assay. The ratio of Vsel to Vred of thephosphorolysis reaction at 600 �M ADP-Glc and without Pi and3PGA was termed the activation state of AGPase.
AGPase activity in the ADP-Glc–forming direction was analyzedin a stopped assay. The assay contained 50 mM Hepes-KOH, pH7.8, 5 mM MgCl2, 1.5 mM Glc-1-P, and 0 to 3 mM ATP in a total vol-ume of 200 �L. The assay was started by the addition of 10 �L ofextract (see above), incubated at 30C for 15 min with shaking at900 rpm, and stopped by incubating at 95C for 5 min. The allo-steric activator 3PGA or the inhibitor Pi was included in the assaysas indicated in the figure legends. The stopped assays were kept at4C until quantification. ADP-Glc was quantified via HPLC using ashort (25-min) program adapted to separate AMP, ADP-Glc, ADP,and ATP (Geigenberger et al., 1994).
RNA Gel Blot Analysis
Total RNA was isolated from different pools of tubers (two samplesper condition) according to Logemann et al. (1987), and RNA gel blotanalysis was performed according to Sambrook et al. (1989). Equalloading and transfer were confirmed by staining with ethidium bro-mide (data not shown). Radioactive hybridization probes were pre-pared with the random priming labeling kit (Boehringer Mannheim)using 32P-dCTP and following the manufacturer’s instructions. EcoRIrestriction fragments of plasmid B22-1 and plasmid S25-1 (Müller-Röber et al., 1990; La Cognata et al., 1995) were used as hybridiza-tion probes for AGPB and AGPS1, respectively.
Protein Gel Blot Analysis
Frozen plant material was extracted directly with 1 � sample buffer(62.5 mM Tris-Cl, pH 6.9, 2% SDS, 10% glycerol, and 0.02% brom-phenol blue, supplemented with 1 mM benzamidine, 1 mM �-aminocaproic acid, 1 mM EDTA, and 1 mM EGTA) that had been degassedeither by boiling the buffer or on an argon line. For every milligram ofplant material, 5 �L of buffer was added. After centrifugation of thesample for 30 s, the supernatant was boiled immediately for 5 to 10min. To a part of the sample, 4 mM DTT was added (DTT), whereasthe rest was used as prepared (�DTT). Equal amounts of both sampleswere loaded onto a 10% acrylamide gel containing SDS (Laemmli,1970) and separated. After transfer of the proteins to polyvinylidenedifluoride, the membrane was incubated with a rabbit antibodyraised against the brittle-2 protein (AGPB) from maize (Giroux andHannah, 1994) and a peroxidase-conjugated anti-rabbit antibody
(Bio-Rad). Peroxidase activity was detected on x-ray film by en-hanced chemiluminescence.
Heterologous Expression of Potato AGPB and AGPS1 in E. coli
The potato sequences of the presumed mature proteins of AGPB(complete SWISS-PROT entry P23509) and AGPS1 (starting atamino acid 41 of SWISS-PROT entry Q00081) were cloned into thevector pQE (Qiagen), causing the encoding of the N-terminal exten-sion MRGSHHHHHHGSVD. The proteins were expressed in E. coliand purified under denaturing conditions on a nickel–nitrilotriaceticacid agarose column. The resulting preparations were �90 and�50% pure for AGPB and AGPS1, respectively. The identities of thepurified preparations were confirmed by matrix-assisted laser-desorp-tion ionization time-of-flight analysis of peptides, generated by in-geldigestion with trypsin (Walz et al., 2002), before they were used todetermine the specificity of the maize brittle-2 and potato AGPB an-tibodies.
Upon request, all novel materials described in this article will bemade available in a timely manner for noncommercial research pur-poses. No restrictions or conditions will be placed on the use of anymaterials described in this article that would limit their use for non-commercial research purposes.
ACKNOWLEDGMENTS
We are grateful to L. Curtis Hannah (University of Florida) for the giftof the maize brittle-2 antibody, to Lothar Willmitzer, Jens Kossmann,and Bernd Müller-Röber for providing transgenic plants, and to UrsulaLa Cognata for providing the potato AGPB and AGPS cDNAs. Weare grateful to Tobias Haas, Katrin Prescha, and Melanie Höhne forexcellent technical help and to Joost van Dongen for help in prepar-ing the figures. This work was supported by the Deutsche For-schungsgemeinschaft, Grant Ge 878/1-1 (to A.T., A.B., and P.G.),and the Max Planck Society (support to J.H.M.H., M.S., Y.G., andE.M.F.). The article is dedicated to Hans W. Heldt on the occasion ofhis retirement.
Received April 3, 2002; accepted June 10, 2002.
REFERENCES
Avigad, G. (1982). Sucrose and other disaccharides. In Encyclope-dia of Plant Physiology, T.A. Loewus and W. Tanner, eds (Heidel-berg, Germany: Springer), pp. 217–347.
Ballicora, M.A., Frueauf, J.B., Fu, Y., Schürmann, P., and Preiss,J. (2000). Activation of the potato tuber ADP glucose pyrophos-phorylase by thioredoxin. J. Biol. Chem. 275, 1315–1320.
Ballicora, M.A., Fu, Y., Frueauf, J.B., and Preiss, J. (1999). Heatstability of the potato ADP-glucose pyrophosphorylase: Role ofCys subunit 12 in the small subunit. Biochem. Biophys. Res.Commun. 257, 782–786.
Ballicora, M.A., Laughlin, M.J., Fu, Y., Okita, T.W., Barry, G.F.,and Preiss, J. (1995). Adenosine 5�-diphosphate-glucose pyro-phosphorylase from potato tuber: Significance of the N terminus

Redox Regulation of Starch Synthesis 2211
of the small subunit for catalytic properties and heat stability.Plant Physiol. 109, 245–251.
Boller, T., and Kende, H. (1979). Hydrolytic enzymes in the centralvacuole of plant cells. Plant Physiol. 63, 1123–1132.
Borchert, S., Grosse, H., and Heldt, H.W. (1989). Specific transportof inorganic phosphate, glucose-6-phosphate, dihydroxyacetone-phosphate and 3-phosphoglycerate into amyloplast from pearoots. FEBS Lett. 253, 183–186.
Dancer, J.E., Neuhaus, H.E., and Stitt, M. (1990). The subcellu-lar compartmentation of uridine nucleotides and nucleoside-5-diphosphate kinase in leaves. Plant Physiol. 92, 637–641.
Farré, E.M., Tiessen, A., Roessner, U., Geigenberger, P., Trethewey,R.N., and Willmitzer, L. (2001). Analysis of the compartmentationof glycolytic intermediates, nucleotides, sugars, amino acids andsugar alcohols in potato tubers using a non-aqueous fractionationmethod. Plant Physiol. 127, 685–700.
Fu, Y., Ballicora, M.A., Leykam, J.F., and Preiss, J. (1998). Mecha-nism of reductive activation of potato tuber ADP-glucose pyro-phosphorylase. J. Biol. Chem. 273, 25045–25052.
Geigenberger, P., Fernie, A.R., Gibon, Y., Christ, M., and Stitt, M.(2000). Metabolic activity decreases as an adaptive response tolow internal oxygen in growing potato tubers. Biol. Chem. 381,723–740.
Geigenberger, P., Geiger, M., and Stitt, M. (1998a). High-tempera-ture perturbation of starch synthesis is attributable to inhibition ofADP-glucose pyrophosphorylase by decreased levels of glycer-ate-3-phosphate in growing potato tubers. Plant Physiol. 117,1307–1316.
Geigenberger, P., Hajirezaei, M., Geiger, M., Deiting, U., Sonnewald,U., and Stitt, M. (1998b). Overexpression of pyrophosphataseleads to increased sucrose degradation and starch synthesis,increased activities of enzymes for sucrose-starch interconversions,and increased levels of nucleotides in growing potato tubers.Planta 205, 428–437.
Geigenberger, P., Langenberger, S., Wilke, I., Heineke, D., Heldt,H.W., and Stitt, M. (1993). Sucrose is metabolised by sucrosesynthase and glycolysis within the phloem complex of Ricinuscommunis L. seedlings. Planta 190, 446–453.
Geigenberger, P., Lerchl, J., Stitt, M., and Sonnewald, U. (1996).Phloem-specific expression of pyrophosphatase inhibits long dis-tance transport of carbohydrates and amino acids in tobaccoplants. Plant Cell Environ. 19, 43–55.
Geigenberger, P., Merlo, L., Reimholz, R., and Stitt, M. (1994).When growing potato tubers are detached from their mother plantthere is a rapid inhibition of starch synthesis, involving inhibition ofADP-glucose pyrophosphorylase. Planta 193, 486–493.
Geigenberger, P., Müller-Röber, B., and Stitt, M. (1999a). Contri-bution of adenosine 5�-diphosphoglucose pyrophosphorylase tothe control of starch synthesis is decreased by water stress ingrowing potato tubers. Planta 209, 338–345.
Geigenberger, P., Reimholz, R., Deiting, U., Sonnewald, U., andStitt, M. (1999b). Decreased expression of sucrose phosphatesynthase strongly inhibits the water stress-induced synthesis ofsucrose in growing potato tubers. Plant J. 19, 119–129.
Geigenberger, P., Reimholz, R., Geiger, M., Merlo, L., Canale, V.,and Stitt, M. (1997). Regulation of sucrose and starch metabolismin potato tubers in response to short-term water deficit. Planta201, 502–518.
Geigenberger, P., Stamme, C., Tjaden, J., Schulz, A., Quick,P.W., Betsche, T., Kersting, H.J., and Neuhaus, H.E. (2001).Tuber physiology and properties of starch from tubers of trans-
genic potato plants with altered plastidic adenylate transporteractivity. Plant Physiol. 125, 1667–1678.
Geigenberger, P., and Stitt, M. (1991). A “futile” cycle of sucrosesynthesis and degradation is involved in regulating partitioningbetween sucrose, starch and respiration in cotyledons of germi-nating Ricinus communis L. seedlings when phloem transport isinhibited. Planta 185, 81–90.
Geigenberger, P., and Stitt, M. (1993). Sucrose synthase catalysesa readily reversible reaction in vivo in developing potato tubersand other plant tissues. Planta 189, 329–339.
Geigenberger, P., and Stitt, M. (2000). Diurnal changes in sucrose,nucleotides, starch synthesis, and AGPS transcript in growingpotato tubers that are suppressed by decreased expression ofsucrose phosphate synthase. Plant J. 23, 795–806.
Geiger, M., Stitt, M., and Geigenberger, P. (1998). Metabolism inpotato tuber slices responds differently after addition of sucroseand glucose. Planta 206, 245–252.
Gerhardt, R., and Heldt, H.W. (1984). Measurement of subcellularmetabolite levels in leaves by fractionation of freeze-stoppedmaterial in non-aqueous media. Plant Physiol. 75, 542–547.
Gibon, Y., Vigeolas, H., Tiessen, A., Geigenberger, P., and Stitt,M. (2002). Sensitive and high throughput metabolite assays usinga novel enzymic cycling system: Utilisation to investigate theimpact of decreased external oxygen concentration on energyand storage metabolism in developing Arabidopsis thalianaseeds. Plant J., 30, 221–235.
Giroux, M.J., and Hannah, L.C. (1994). ADP-glucose pyrophosphor-ylase in shrunken2 and brittle2 mutants of maize. Mol. Gen.Genet. 243, 400–408.
Hajirezaei, M., Sonnewald, U., Viola, R., Carlisle, S., Dennis, D.,and Stitt, M. (1994). Transgenic potato plants with stronglydecreased expression of pyrophosphate:fructose-6-phosphatephosphotransferase show no visible phenotype and only minorchanges in metabolic fluxes in their tubers. Planta 192, 16–30.
Hannah, L.C., Shaw, J.R., Giroux, M.J., Reyss, A., Prioul, J.-L.,Rae, J.-M., and Lee, J.-Y. (2001). Maize genes encoding thesmall subunit of ADP-glucose pyrophosphorylase. Plant Physiol.127, 173–183.
Hatzfeld, W.D., Dancer, J.E., and Stitt, M. (1990). Fructose 2,6-bis-phosphate, metabolites and “coarse” control of pyrophos-phate:fructose-6-phosphate phosphotransferase during triosephosphate cycling in heterotrophic cell suspension cultures ofChenopodium rubrum. Planta 180, 205–211.
Heldt, H.W., Chon, C.J., Maronde, D., Herold, A., Stankovic, Z.S.,Walker, D.A., Kraminer, A., Kirk, M.R., and Heber, U. (1977).Role of orthophosphate and other factors in the regulation ofstarch formation in leaves and isolated chloroplast. Plant Physiol.59, 1146–1155.
Herold, A. (1980). Regulation of photosynthesis by sink activity: Themissing link. New Phytol. 86, 131–144.
Iglesias, A.A., Barry, G.F., Meyer, C., Bloksberg, L., Nakata, P.A.,Greene, T., Laughlin, M.J., Okita, T.W., Kishore, G.M., and Preiss,J. (1993). Expression of the potato tuber ADP-glucose pyrophos-phorylase in Escherichia coli. J. Biol. Chem. 268, 1081–1086.
Jellito, T., Sonnewald, U., Willmitzer, L., Hajirezaei, M.R., andStitt, M. (1992). Inorganic pyrophosphate content and metabo-lites in leaves and tubers of potato and tobacco plantsexpressing E. coli pyrophosphatase in their cytosol. Planta 188,238–244.
Jenner, H.L., Winning, B.M., Millar, A.H., Tomlinson, K.L., Leaver,C.J., and Hill, S.A. (2001). NAD malic enzyme and the control of

2212 The Plant Cell
carbohydrate metabolism in potato tubers. Plant Physiol. 126,1139–1149.
Kammerer, B., Fischer, K., Hilpert, B., Schubert, S., Gutensohn,M., Weber, A., and Flugge, U.I. (1998). Molecular characteriza-tion of a carbon transporter in plastids from heterotrophic tissues:The glucose 6-phosphate/phosphate antiporter. Plant Cell 10,105–117.
Kim, W.T., Franceschi, V.R., Okita, T.W., Robinson, N.L., Morell,M., and Preiss, J. (1989). Immunocytochemical localization ofADPglucose pyrophosphorylase in developing potato tuber cells.Plant Physiol. 91, 217–220.
Kleczkowsky, L. (1994). Glucose activation and metabolismthrough UDP-glucose pyrophosphorylase in plants. Phytochemis-try 37, 1507–1515.
Krapp, A., Quick, W.P., and Stitt, M. (1991). Rubisco, other Calvincycle enzymes and chlorophyll decrease when glucose is sup-plied to mature spinach leaves via the transpiration stream. Planta186, 58–69.
Krapp, A., and Stitt, M. (1995). An evaluation of direct and indirectmechanisms for the “sink regulation” of photosynthesis of spin-ach: Changes in gas exchange, carbohydrates, metabolites,enzyme activities and steady-state transcript levels after cold gir-dling of source leaves. Planta 195, 313–323.
Kruger, N.J. (1997). Carbohydrate synthesis and degradation. InPlant Metabolism, D.T. Dennis, D.H. Turpin, D.D. Lefebvre, andD.B. Layzell, eds (Harlow, UK: Longman), pp. 83–104.
La Cognata, U., Willmitzer, L., and Müller-Rober, B. (1995).Molecular cloning and characterization of novel isoforms of potatoADP-glucose pyrophosphorylase. Mol. Gen. Genet. 246, 538–548.
Laemmli, U.K. (1970). Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680–685.
Laing, W.A., Stitt, M., and Heldt, H.W. (1981). Control of CO2 fixa-tion: Changes in the activity of ribulose 5-phosphate kinase andfructose- and sedoheptulose-bisphosphatase in chloroplasts.Biochim. Biophys. Acta 637, 348–359.
Lloyd, J.R., Springer, F., Buleon, A., Müller-Röber, B., Willmitzer,L., and Kossmann, J. (1999). The influence of alterations in ADP-glucose pyrophosphorylase activities on starch structure andcomposition in potato tubers. Planta 209, 230–238.
Logemann, J., Schell, J., and Willmitzer, L. (1987). Improvedmethod for the isolation of RNA from plant tissues. Anal. Bio-chem. 163, 21–26.
MacDonald, F.D., and Preiss, J. (1986). The subcellular locationand characteristics of pyrophosphate-fructose-6-phosphate phos-photransferase from suspension-cultured cells of soybean. Planta167, 240–245.
Mares, D.J., and Marschner, H. (1980). Assimilate conversion inpotato tubers in relation to starch deposition and cell growth. Ber.Dtsch. Bot. Ges. 95, 299–313.
Martin, C., and Smith, A.M. (1995). Starch biosynthesis. Plant Cell7, 971–985.
Merlo, L., Geigenberger, P., Hajirezaei, M.R., and Stitt, M. (1993).Changes of carbohydrates, metabolites and enzyme activities inpotato tubers during development, and within a single tuber alonga stolon-apex gradient. J. Plant Physiol. 142, 392–402.
Müller-Röber, B.T., Kossmann, J., Hannah, L.C., Willmitzer, L.,and Sonnewald, U. (1990). One of two different ADP-glucosepyrophosphorylase genes from potato responds strongly to ele-vated levels of sucrose. Mol. Gen. Genet. 224, 136–146.
Müller-Röber, B.T., Sonnewald, U., and Willmitzer, L. (1992). Inhi-bition of ADP-glucose pyrophosphorylase leads to sugar storing
tubers and influences tuber formation and expression of tuberstorage protein genes. EMBO J. 11, 1229–1238.
Neuhaus, H., and Stitt, M. (1990). Control analysis of photosyn-thate partitioning: Impact of reduced activity of ADP-glucosepyrophosphorylase or plastid phosphoglucomutase on the fluxesto starch and sucrose in Arabidopsis thaliana L. Heynh. Planta182, 445–454.
Nielsen, T.H., Krapp, A., Röper-Schwarz, U., and Stitt, M. (1998).The sugar-mediated regulation of genes encoding the small sub-unit of Rubisco and the regulatory subunit of ADP glucose pyro-phosphorylase is modified by nitrogen and phosphate. Plant CellEnviron. 21, 443–455.
Okita, T.W., Nakata, P.A., Anderson, J.M., Sowokinos, J., Morell,M., and Preiss, J. (1990). The subunit structure of potato tuberADPglucose pyrophosphorylase. Plant Physiol. 93, 785–790.
Preiss, J. (1988). Biosynthesis of starch and its regulation. In TheBiochemistry of Plants, Vol. 14, J. Preiss, ed (San Diego, CA: Aca-demic Press), pp. 181–254.
Renz, A., and Stitt, M. (1993). Substrate specificity and productinhibition of different forms of fructokinase and hexokinases indeveloping potato tubers. Planta 190, 166–175.
Riens, B., Lohaus, G., Heineke, D., and Heldt, H.W. (1991). Aminoacid and sucrose content determined in the cytosolic, chloroplas-tic, and vacuolar compartments and in the phloem sap of spinachleaves. Plant Physiol. 97, 227–233.
Rolleston, F.S. (1972). A theoretical background to the use of mea-sured intermediates in the study of the control of intermediarymetabolism. Curr. Top. Cell. Regul. 5, 47–75.
Salanoubat, M., and Belliard, G. (1989). The steady-state level ofpotato sucrose synthase mRNA is dependent on wounding,anaerobiosis and sucrose. Gene 84, 181–185.
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989). MolecularCloning: A Laboratory Manual. (Cold Spring Harbor, NY: ColdSpring Harbor Laboratory Press).
Scheibe, R. (1991). Redox-modulation of chloroplast enzymes: Acommon principle for individual control. Plant Physiol. 96, 1–3.
Scheible, W.R., González-Fontes, A., Lauerer, M., Müller-Röber,B., Caboche, M., and Stitt, M. (1997). Nitrate acts as a signal toinduce organic acid metabolism and repress starch metabolism intobacco. Plant Cell 9, 783–798.
Schott, K., Borchert, S., Müller-Röber, B., and Heldt, H.W. (1995).Transport of inorganic phosphate, and C3- and C6-sugar phos-phates across the envelope membranes of potato tuber amylo-plast. Planta 196, 647–652.
Schümann, P., and Jacquot, J.-P. (2000). Plant thioredoxin systemsrevisited. Annu. Rev. Plant Physiol. Plant Mol. Biol. 51, 371–400.
Silverstein, R., Voet, J., Reed, D., and Abeles, R.H. (1967). Purifi-cation and mechanism of action of sucrose phosphorylase. J.Biol. Chem. 242, 1338–1345.
Smith, A.M., Denyer, K., and Martin, C. (1997). The synthesis of thestarch granule. Annu. Rev. Plant Physiol. Plant Mol. Biol. 48, 67–87.
Sokolov, L.N., Dejardin, A., and Kleczkowski, L.A. (1998). Sugarsand light/dark exposure trigger differential regulation of ADP-glu-cose pyrophosphorylase genes in Arabidopsis thaliana (thalecress). Biochem. J. 336, 681–687.
Sowokinos, J.R. (1981). Pyrophosphorylases in Solanum tubero-sum. II. Catalytic properties and regulation of ADP-glucose andUDP-glucose pyrophosphorylase activities in potatoes. PlantPhysiol. 68, 924–929.
Sowokinos, J.R., and Preiss, J. (1982). Phosphorylases in Solanumtuberosum. III. Purification, physical and catalytical properties of

Redox Regulation of Starch Synthesis 2213
ADP-glucose pyrophosphorylase in potatoes. Plant Physiol. 69,1459–1466.
Stark, D.M., Timmerman, K.P., Barry, G.F., Preiss, J., andKishore, G.M. (1992). Regulation of the amount of starch in plant tis-sues by ADP-glucose pyrophosphorylase. Science 258, 287–292.
Stitt, M., Huber, S., and Kerr, P. (1987). Control of photosyntheticsucrose synthesis. In The Biochemistry of Plants, Vol. 10, M.D. Hatchand N.K. Boardman, eds (New York: Academic Press), pp. 327–409.
Stitt, M., Lilley, R.M., Gerhardt, R., and Heldt, H.W. (1989). Metab-olite levels in specific cells and subcellular compartments of plantleaves. Methods Enzymol. 174, 518–550.
Stitt, M., Lilley, R.M., and Heldt, H.W. (1982). Adenine nucleotidelevels in the cytosol, chloroplast, and mitochondria of wheat leafprotoplasts. Plant Physiol. 70, 971–977.
Stitt, M., and Krapp, A. (1999). The interaction between elevatedcarbon dioxide and nitrogen nutrition: The physiological andmolecular background. Plant Cell Environ. 22, 583–621.
Stryer, L.M. (1990). Biochemistry. (Weinheim, Germany: VerlagChemie).
Tauberger, E., Fernie, A.R., Emmermann, M., Renz, A., Kossman,J., Willmitzer, L., and Trethewey, R.N. (2000). Antisense inhibi-tion of plastidial phosphoglucomutase provides compelling evi-dence that potato tuber amyloplasts import carbon from the cyto-sol in the form of glucose-6-phosphate. Plant J. 23, 43–53.
Thorbjornsen, T., Villand, P., Kleczkowski, L.A., and Olsen, O.-A.(1996). A single gene encodes two different transcripts for theADP-glucose pyrophosphorylase small subunit from barley (Hor-deum vulgare). Biochem. J. 313, 149–154.
Tjaden, J., Möhlmann, T., Kampfenkel, K., Henrichs, G., andNeuhaus, H.E. (1998). Altered plastidic ATP/ADP-transporteractivity influences potato (Solanum tuberosum L.) tuber morphol-ogy, yield and composition of starch. Plant J. 16, 531–540.
Trethewey, R.N., Fernie, A.R., Bachmann, A., Fleischer-Notter,H., Geigenberger, P., and Willmitzer, L. (2001). Expression of abacterial sucrose phosphorylase in potato tubers results in a glu-cose-independent induction of glycolysis. Plant Cell Environ. 24,357–365.
Trethewey, R.N., Geigenberger, P., Hajirezaei, M., Sonnewald,U., Stitt, M., Riesmeier, J., and Willmitzer, L. (1998). Combinedexpression of glucokinase and invertase in potato tubers leads toa dramatic reduction in starch accumulation and a stimulation ofglycolysis. Plant J. 15, 109–118.
Walz, C., Juenger, M., Schad, M., and Kehr, J. (2002). Evidencefor the presence and activity of a complete antioxidant defencesystem in mature sieve tubes. Plant J. 31, 189–197.
Weiner, H., Stitt, M., and Heldt, H.W. (1987). Subcellular compart-mentation of pyrophosphate and pyrophosphatase in leaves. Bio-chim. Biophys. Acta 893, 13–21.
Wirtz, W., Stitt, M., and Heldt, H.W. (1982). Light activation ofCalvin cycle enzymes as measured in pea leaves. FEBS Lett. 142,223–226.
Zabawinski, C., van den Koornhuyse, N., D’Hulst, C., Schlichting,R., Giersch, C., Delrue, B., Lacroix, J.-M., Preiss, P., and Ball,S. (2001). Starchless mutants of Chlamydomonas reinhardtii lackthe small subunit of a heterotetrameric ADP-glucose pyrophos-phorylase. J. Bacteriol. 183, 1069–1077.

DOI 10.1105/tpc.003640 2002;14;2191-2213Plant Cell
Peter GeigenbergerAxel Tiessen, Janneke H. M. Hendriks, Mark Stitt, Anja Branscheid, Yves Gibon, Eva M. Farré and
Sucrose SupplytheADP-Glucose Pyrophosphorylase: A Novel Regulatory Mechanism Linking Starch Synthesis to
Starch Synthesis in Potato Tubers Is Regulated by Post-Translational Redox Modification of
This information is current as of May 21, 2018
References /content/14/9/2191.full.html#ref-list-1
This article cites 78 articles, 28 of which can be accessed free at:
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists