stable carbon isotope fractionation by acetotrophic sulfur-reducing bacteria
TRANSCRIPT

R E S E A R C H A R T I C L E
Stable carbon isotope fractionation byacetotrophicsulfur-reducing bacteriaDennis Goevert & Ralf Conrad
Max Planck Institute for Terrestrial Microbiology, Marburg, Germany
Correspondence: Ralf Conrad, Max Planck
Institute for Terrestrial Microbiology, Karl-von-
Frisch-Str., 35043 Marburg, Germany. Tel.:
149 6421 178801; fax: 149 6421 178809;
e-mail: [email protected]
Received 18 August 2009; revised 26 October
2009; accepted 30 October 2009.
Final version published online 27 November
2009.
DOI:10.1111/j.1574-6941.2009.00811.x
Editor: Gary King
Keywords
acetate turnover; Desulfuromonas;
Desulfurella; Hippea; acetyl-CoA pathway;
tricarboxylic acid cycle.
Abstract
Acetate is the most important intermediate in anaerobic degradation of organic
matter. The carbon isotope effects associated with the oxidation of acetate (eac)
were examined for four acetotrophic sulfur reducers, Desulfuromonas acetoxidans,
Desulfuromonas thiophila, Desulfurella acetivorans, and Hippea maritima. During
the consumption of acetate and sulfur, acetate was enriched in 13C by 11.5 and
11.2% in Desulfuromonas acetoxidans and Desulfuromonas thiophila, respectively.
By contrast, isotope fractionation in D. acetivorans and H. maritima resulted in
isotope enrichment factors of eac =� 6.3% and � 8.4%, respectively. These
sulfur-reducing bacteria all metabolize acetate via the tricarboxylic acid cycle, but
have different mechanisms for the initial activation of acetate. In Desulfuromonas
acetoxidans, acetyl-CoA is formed by succinyl-CoA : acetate-CoA-transferase, and
in D. acetivorans by acetate kinase and phosphate acetyltransferase. Hence, values
of eac seem to be characteristic for the type of activation of acetate to acetyl-CoA in
acetotrophic sulfur reducers. Summarizing eac-values in anaerobic acetotrophic
microorganisms, it appears that isotope fractionation depends on the mechanism
of acetate activation to acetyl-CoA, on the key enzyme of the acetate dissimilation
pathway, and on the bioavailability of acetate, which all have to be considered
when using d13C of acetate in environmental samples for diagnosis of the involved
microbial populations.
Introduction
Acetate is a key intermediate during the anaerobic degrada-
tion of organic matter. It can be used by different groups of
anaerobic microorganisms, such as those reducing nitrate,
iron, sulfate, protons, or producing methane. For example,
4 70% of biological methanogenesis in flooded soils (Con-
rad, 1999) and about 50% of sulfate reduction in marine
sediments (Soerensen et al., 1981) result from acetate
consumption. Besides several sulfate-reducing bacteria, a
few sulfur-reducing bacteria are also capable of oxidizing
acetate by a coupled reduction of elemental sulfur to sulfide.
However, little is known about carbon isotope fractionation
in acetate-dissimilating microorganisms, although such
knowledge may be helpful to diagnose the involvement of
particular groups of microorganisms in acetate turnover in
natural environments (Penning et al., 2006; Goevert &
Conrad, 2009).
Two main pathways for the oxidation of acetate by
anaerobic organisms are known: the acetyl-CoA/carbon
monoxide dehydrogenase pathway and the tricarboxylic
acid (TCA) cycle. Recently, it was observed that sulfate
reducers show differences in the fractionation of stable
carbon when they oxidize acetate via these different path-
ways (Goevert & Conrad, 2008). However, until now, to our
knowledge, all isolated sulfur reducers that were found to be
capable of utilizing acetate use the TCA cycle (Pfennig &
Biebl, 1976; Bonch-Osmolovskaya et al., 1990; Galushko &
Schink, 2000) for the following reaction:
CH3COO� þHþ þ 4S0 þ 2H2O! 2CO2 þ 4H2S ð1Þ
However, it was found that sulfur-reducing bacteria differ
in their biochemical activation of acetate. Desulfuromonas
acetoxidans activates acetate to acetyl-CoA via CoA transfer
FEMS Microbiol Ecol 71 (2010) 218–225c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
MIC
ROBI
OLO
GY
EC
OLO
GY

from succinyl-CoA (Gebhardt et al., 1985):
acetateþ succinyl-CoA! acetyl-CoAþ succinate ð2Þcatalyzed by a succinyl-CoA : acetate-CoA-transferase. In
contrast, for Desulfurella acetivorans the following reaction
was reported (Schmitz et al., 1990):
acetateþ ATP! acetyl-phosphateþ ADP ð3Þ
acetyl-phosphateþ CoASH! acetyl-CoAþ phosphate ð4Þwhere acetate is activated by two enzymes, acetate kinase
and phosphate acetyltransferase.
Therefore, we determined carbon isotope ratios for
Desulfuromonas acetoxidans and D. acetivorans to find out
whether sulfur reducers show different carbon isotope
fractionation when they activate acetate via different me-
chanisms. By studying fractionation in Desulfuromonas
thiophila (level of similarity to Desulfuromonas acetoxidans:
96.6%; Finster et al., 1997) and Hippea maritima (closely
related to D. acetivorans by approximately 88%; Miroshni-
chenko et al., 1999) our aim was to investigate whether
sulfur reducers with high 16S rRNA gene sequence similarity
express similar isotope fractionation. We hypothesize that if
this is true, the determination of carbon isotope fractiona-
tion may help to elucidate the initial activation mechanism
in acetotrophic, sulfur-reducing bacteria. Furthermore, it
might help to identify microbial groups of acetate utilizers
in natural environments by analyzing the isotopic signature
of acetate.
Materials and methods
Cultures and growth conditions
The following pure cultures were used in this study:
Desulfuromonas acetoxidans (DSM 684), Desulfuromonas
thiophila (DSM 8987), D. acetivorans (DSM 5264), and
H. maritima (DSM 10411) were obtained from the Deutsche
Sammlung von Mikroorganismen und Zellkulturen
(Braunschweig, Germany). All cultures were grown under
N2/CO2 (80 : 20) in 500-mL glass bottles (Ochs, Bovenden-
Lenglern, Germany) in bicarbonate-buffered mineral med-
ium. Pure cultures of Desulfuromonas acetoxidans were
incubated with shaking (120 r.p.m.) at 30 1C using the
original basal medium (Pfennig & Biebl, 1976). Desulfur-
omonas thiophila was also grown at 30 1C, using a modified
medium with the following composition (in g L�1 unless
otherwise noted): KH2PO4, 1.0; NH4Cl, 0.5; MgSO4 � 7H2O,
0.4; CaCl2 � 2H2O, 0.1; NaHCO3, 2.0; Na2S � 9H2O, 0.4; trace
element solution SL-10, 1.0 mL (Chin et al., 1998); vitamin
solution, 1 mL (Wolin et al., 1963); and resazurine at
0.1% w/v, 1 mL. Desulfurella acetivorans and H. maritima
were incubated without shaking at 55 1C. The medium used
for cultivation of D. acetivorans contained the following (in
g L�1 unless otherwise noted): KH2PO4, 0.33; NH4Cl, 0.33;
MgCl2 � 6H2O, 0.33; CaCl2 � 2H2O, 0.33; KCl, 0.33; NaHCO3,
2.0; Na2S � 9H2O, 0.5; trace element solution SL-10, 1.0 mL
(Chin et al., 1998); vitamin solution, 10 mL (Wolin et al.,
1963); and resazurine at 0.1% w/v, 1 mL. For growth of
H. maritima a similar medium was used, which additionally
contained 0.1 g L�1 yeast extract and 25 g L�1 NaCl and the
pH was adjusted to 6.0 with 5 M H2SO4 (Miroshnichenko
et al., 1999). All bacteria were grown with 3.8–5.3 mM
acetate as electron donor and 6.2 mM sulfur (subl., purum,
purchased from Fluka, Buchs, Switzerland) as electron
acceptor. For experiments, 10% of bacterial suspensions in
the late exponential phase were inoculated (resulting in a
final volume of 250 mL) and several samples from the
headspace and the liquid phase were removed to determine
pH, concentration of acetate, sulfide, carbon dioxide, and
carbon isotope composition of acetate and carbon dioxide.
All experiments were performed in triplicate.
Chemical and isotopic analyses
Chemical and isotopic analyses were performed as described
previously (Goevert & Conrad, 2008). Sulfide was deter-
mined photometrically after reaction with CuSO4 to form
CuS as described by Cord-Ruwisch (1985). CO2 was ana-
lyzed by GC using a flame ionization detector (Shimadzu,
Kyoto, Japan) after conversion to CH4 with a methanizer
(Ni-catalyst at 350 1C, Chrompack, Middelburg, the Nether-
lands). Stable isotope analysis of 13C/12C in gas samples was
performed using a gas chromatograph combustion isotope
ratio mass spectrometer system (Thermo Fisher Scientific,
Bremen, Germany). Isotopic measurements and quantifica-
tion of acetate were performed on an HPLC system coupled
to Finnigan LC IsoLink (Thermo Fisher Scientific).
Calculations
Fractionation factors for a reaction A ! B are defined after
Hayes (1993) as:
aA=B ¼ ðdA þ 1000Þ=ðdB þ 1000Þ ð5Þ
also expressed as e � 103 (1�a). The carbon isotope
enrichment factor associated with acetate oxidation (eac)
was calculated from the temporal change of d13C of acetate
as described by Mariotti et al. (1981) from the residual
reactant
dr ¼ dri þ e½lnð1� f Þ� ð6Þwhere dri and dr are the isotope compositions of the reactant
(acetate) at the beginning and of the residual acetate,
respectively, and f is the fractional yield of the product based
on the consumption of acetate (0o fo 1). Linear
FEMS Microbiol Ecol 71 (2010) 218–225 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
219Acetotrophic sulfur-reducing bacteria

regression of dr against ln(1� f) yields e as the slope of best-
fit lines.
As total oxidized carbon was distributed among different
carbon species, d13C of total inorganic carbon (dTIC) was
calculated by the following mass-balance equation:
dTIC ¼ Xgdg þ Xddd þ Xbdb þ Xcdc ð7Þwhere X is the mole fraction and d is the isotopic composi-
tion of the C of gaseous CO2 (g), dissolved CO2 (d), HCO3�
(b), and CO32� (c). The distribution of carbon among these
species was calculated using solubility and equilibrium
constants (Stumm & Morgan, 1995). dg was measured
directly, the remaining isotopic compositions were calcu-
lated from the relevant equilibrium isotope fractionation
factors (Deines & Langmuir, 1974; Mook et al., 1974):
dd ¼ ad=g þ dg þ ðad=g�1Þ1000 ð8Þ
db ¼ ab=g þ dg þ ðab=g � 1Þ1000 ð9Þ
dc ¼ ac=g þ dg þ ðac=g � 1Þ1000 ð10Þ
Results
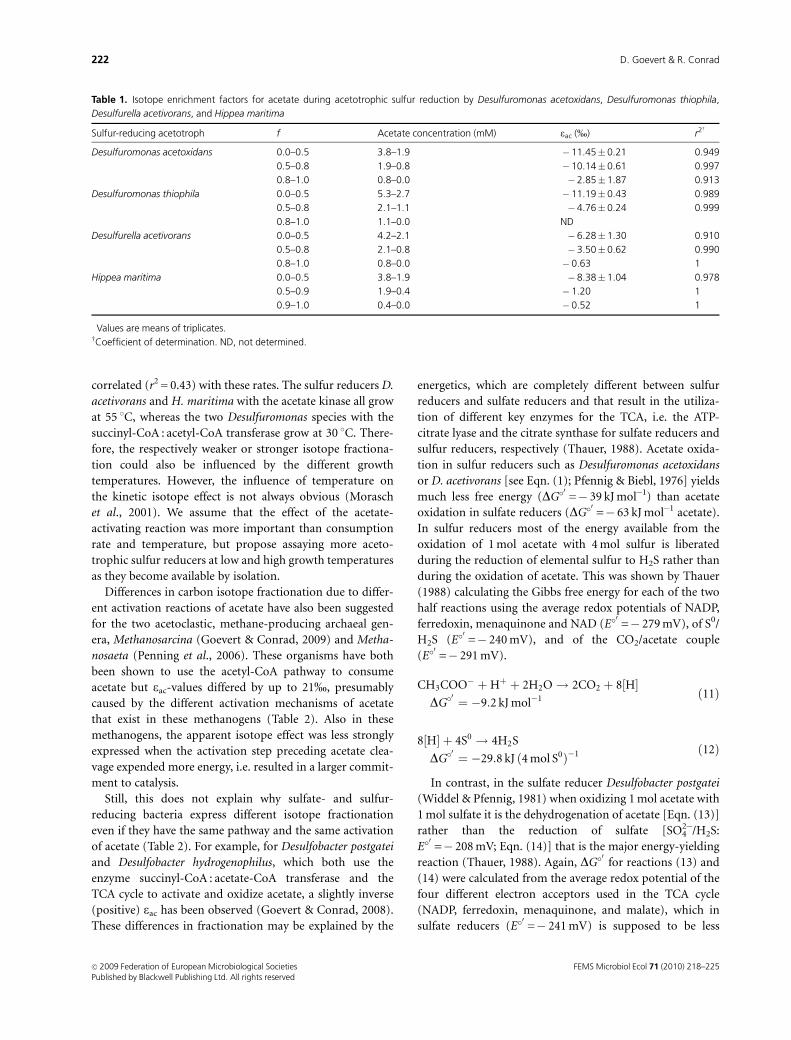
Desulfuromonas acetoxidans and Desulfuromonas thiophila
were grown to study isotope fractionation of stable carbon
during acetate oxidation via the TCA cycle and activation of
acetate to acetyl-CoA via CoA transfer from succinyl-CoA.
This reaction was shown to drive acetate activation in
Desulfuromonas acetoxidans (Gebhardt et al., 1985) and is
assumed to be also active in Desulfuromonas thiophila (see
Discussion). During oxidation of acetate, both Desulfuro-
monas species converted acetate and sulfur into CO2 and
H2S, respectively, following the known stoichiometry [see
Eqn. (1); Fig. 1a and d]. Concentrations of CO2 are not
shown because the high concentrations of bicarbonate in the
medium made it impractical to measure yields of this
product. During sulfur reduction the preferred consump-
tion of [12C]acetate caused an enrichment of the heavier
isotope 13C in the remaining acetate (Fig. 1b and e). d13CCO2
[illustrated in Fig. 1 as TIC] became slightly depleted in 13C
with time but was not used for determination of isotope
fractionation because, as mentioned above, the high back-
ground of bicarbonate hindered a precise quantification of
the d13C of the newly formed TIC. Carbon isotope fractio-
nation of acetate during sulfur reduction was determined
using Eqn. (6), based on Rayleigh distillation (Fig. 1c and f
and Table 1). The carbon isotope enrichment factor for
acetate (eac) was calculated for three different phases of
acetate consumption to respect substrate limitation (Goe-
vert & Conrad, 2009): until 50% of acetate was consumed
(f-values between 0.0 and 0.5), between 50% and 80%
consumption (0.5o fo 0.8), and from 80% to maximum
consumption of acetate (0.8o fo 1.0). As carbon isotope
fractionation decreased after the first phase, e-values for the
range 0o fo 0.5 were finally used to determine and
compare isotope fractionation. Toward the end of experi-
ments and thus at lower substrate concentrations, acetate
might have become limiting and no longer fractionated
during consumption, as observed before (Kinnaman et al.,
2007; Goevert & Conrad, 2008, 2009). Such an effect of
bioavailability of substrates on stable isotope fractionation
has recently been explained theoretically for pollutant
biodegradation (Kampara et al., 2008; Thullner et al., 2008)
and shown to occur during acetoclastic methanogenesis
(Goevert & Conrad, 2009). It seems also to occur during
acetate oxidation under sulfur-reducing conditions.
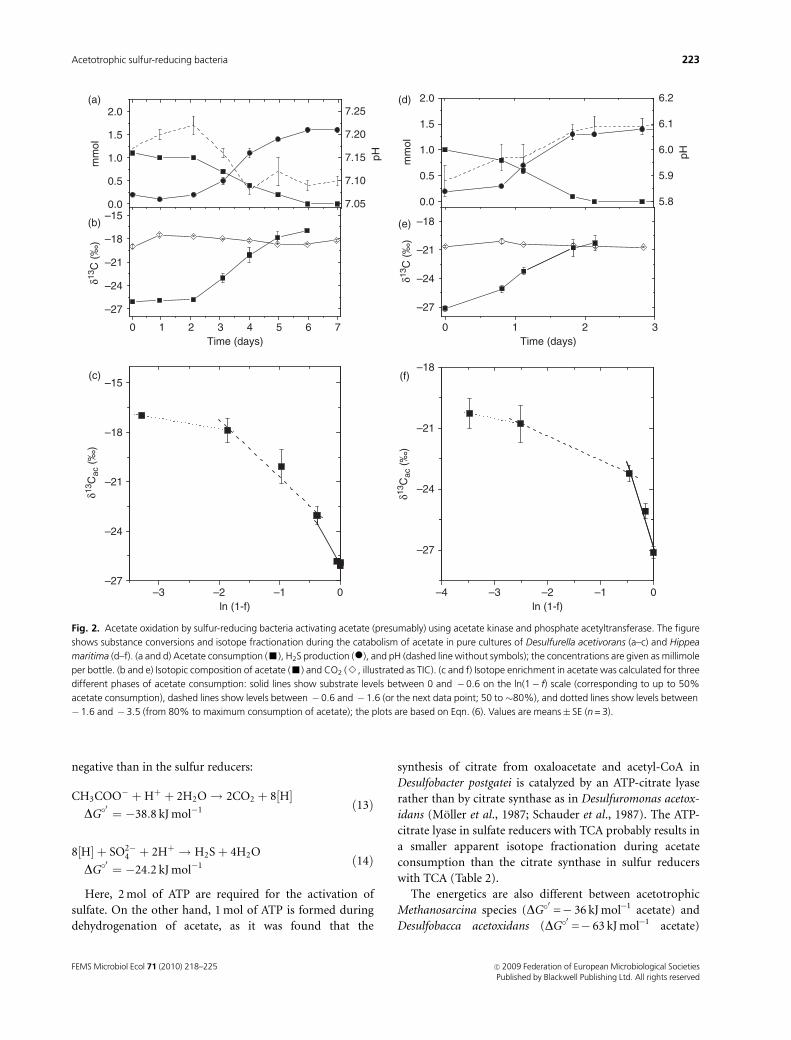
In contrast to the two Desulfuromonas spp., D. acetivorans
(Schmitz et al., 1990) and apparently also H. maritima (see
Discussion) both activate acetate directly to acetyl-CoA,
catalyzed by the two enzymes acetate kinase and phosphate
acetyltransferase. During growth of D. acetivorans and H.
maritima, acetate was consumed below the detection limit
(approximately 20 mM) and was converted to CO2 (Fig. 2a
and d). When acetate was oxidized, again the preferred
consumption of light 12C-acetate caused an enrichment of
heavy 13C in the residual acetate (Fig. 2b and e), as expected
for a closed system approach. Similar to the Desulfuromonas
species, eac-values decreased with decreasing substrate con-
centrations (Fig. 2c and f and Table 1). But, interestingly,
isotope enrichment factors of acetate in D. acetivorans and
H. maritima differed from eac in Desulfuromonas spp. and
were significantly weaker (Table 1).
Discussion
Acetotrophic sulfate reducers apparently express different
carbon isotope fractionation when they use different bio-
chemical pathways for the oxidation of acetate, either the
TCA cycle or the acetyl-CoA pathway (Table 2). In the
present study, we found that carbon isotope fractionation of
acetate differs among acetotrophic sulfur-reducing bacteria
by up to 5%, even if they are using the same biochemical
pathway (TCA) for the oxidation of acetate. Thus, we
conclude that in these bacteria it is not the oxidation
pathway that determines carbon isotope fractionation as
suggested recently for sulfate-reducing bacteria (Goevert &
Conrad, 2008). Instead we propose that the initial activation
of acetate was important for the fractionation of acetate.
Whereas in D. acetivorans, acetate is activated by acetate
kinase and phosphate acetyltransferase at the expense of one
high-energy phosphate bond, in Desulfuromonas acetoxi-
dans, acetate is activated without expense of additional
energy. Acetate activation, by the latter reaction, exhibited
the stronger isotope effect during the subsequent oxidation
FEMS Microbiol Ecol 71 (2010) 218–225c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
220 D. Goevert & R. Conrad

of acetyl-CoA in the TCA. The apparent isotope effect is
mainly dependent on the mechanism of the first irreversible
step (Northrop, 1981). In the acetotrophic sulfur reducers
this is the reaction of the citrate synthase, the key enzyme of
the TCA (Thauer, 1988). However, other reactions preced-
ing the citrate synthase may also influence the apparent
isotope effect. This has been termed ‘commitment to
catalysis’ (Northrop, 1981; Elsner et al., 2005). We may
assume that the energy-utilizing acetate kinase creates a
larger commitment to catalysis, so that the apparent isotope
effect by the citrate synthase is not as strongly expressed as
with the succinyl-CoA : acetate-CoA-transferase.
If our assumption is true, then the determination of eac
may help to identify the biochemical mechanism of acetate
activation in acetotrophic sulfur reducers. Hence, by
comparison of isotope enrichment in acetate among
different sulfur reducers, we assume that Desulfuromonas
thiophila (eac =� 11.2%) activates acetate directly via succi-
nyl-CoA : acetate-CoA transferase as in Desulfuromonas acet-
oxidans (eac =� 11.5%), and that in H. maritima (eac =
� 8.4%) this reaction is catalyzed via acetate kinase and
phosphate acetyltransferase as in D. acetivorans (eac =
� 6.3%). However, enzyme or genomic data are not avail-
able for these species.
The rates of acetate consumption were different in
the different cultures, increasing from H. maritima4D.
acetivorans4Desulfuromonas thiophila4Desulfuromonas
acetoxidans (Figs 1 and 2). However, eac was only weakly
1.5
2.0
7.10
7.151.5
2.0
7.00
7.05(d)(a)
0.0
0.5
1.0
6.95
7.00
7.05mm
ol
pH
0.0
0.5
1.0
6.85
6.90
6.95
mm
ol
pH
(e)(b)
–15
–10
–5
–18
–15
–5
0 –12(c) (f)
0 4 8 12 16 20
–25
–20
δ13C
(‰
)
Time (days)0 2 4 6 8 10
–24
–21δ13C
(‰
)
Time (days)
–20
–15
–10
δ13C
ac (
‰)
–21
–18
–15δ13
Cac
(‰
)
–3 –2 –1 0–30
–25
ln (1-f)–2.0 –1.5 –1.0 –0.5 0.0
–24
ln (1-f)
Fig. 1. Acetate oxidation by sulfur-reducing bacteria activating acetate (presumably) via succinyl-CoA : acetate-CoA-transferase. The figure shows
substance conversions and isotope fractionation during the catabolism of acetate in pure cultures of Desulfuromonas acetoxidans (a–c) and
Desulfuromonas thiophila (d–f). (a and d) Acetate consumption (’), H2S production (�), and pH (dashed line without symbols); the concentrations
are given as millimole per bottle. (b and e) Isotopic composition of acetate (’) and CO2 (B, illustrated as TIC). (c and f) Isotope enrichment in acetate
was calculated for three different phases of acetate consumption: solid lines show substrate levels between 0 and � 0.6 on the ln(1� f) scale
(corresponding to up to 50% acetate consumption), dashed lines show levels between � 0.6 and � 1.6 (50–80%), and dotted lines show levels
between � 1.6 and � 3.0 (from 80% to maximum consumption of acetate); the plots are based on Eqn. (6). Values are means� SE (n = 3).
FEMS Microbiol Ecol 71 (2010) 218–225 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
221Acetotrophic sulfur-reducing bacteria
correlated (r2 = 0.43) with these rates. The sulfur reducers D.
acetivorans and H. maritima with the acetate kinase all grow
at 55 1C, whereas the two Desulfuromonas species with the
succinyl-CoA : acetyl-CoA transferase grow at 30 1C. There-
fore, the respectively weaker or stronger isotope fractiona-
tion could also be influenced by the different growth
temperatures. However, the influence of temperature on
the kinetic isotope effect is not always obvious (Morasch
et al., 2001). We assume that the effect of the acetate-
activating reaction was more important than consumption
rate and temperature, but propose assaying more aceto-
trophic sulfur reducers at low and high growth temperatures
as they become available by isolation.
Differences in carbon isotope fractionation due to differ-
ent activation reactions of acetate have also been suggested
for the two acetoclastic, methane-producing archaeal gen-
era, Methanosarcina (Goevert & Conrad, 2009) and Metha-
nosaeta (Penning et al., 2006). These organisms have both
been shown to use the acetyl-CoA pathway to consume
acetate but eac-values differed by up to 21%, presumably
caused by the different activation mechanisms of acetate
that exist in these methanogens (Table 2). Also in these
methanogens, the apparent isotope effect was less strongly
expressed when the activation step preceding acetate clea-
vage expended more energy, i.e. resulted in a larger commit-
ment to catalysis.
Still, this does not explain why sulfate- and sulfur-
reducing bacteria express different isotope fractionation
even if they have the same pathway and the same activation
of acetate (Table 2). For example, for Desulfobacter postgatei
and Desulfobacter hydrogenophilus, which both use the
enzyme succinyl-CoA : acetate-CoA transferase and the
TCA cycle to activate and oxidize acetate, a slightly inverse
(positive) eac has been observed (Goevert & Conrad, 2008).
These differences in fractionation may be explained by the
energetics, which are completely different between sulfur
reducers and sulfate reducers and that result in the utiliza-
tion of different key enzymes for the TCA, i.e. the ATP-
citrate lyase and the citrate synthase for sulfate reducers and
sulfur reducers, respectively (Thauer, 1988). Acetate oxida-
tion in sulfur reducers such as Desulfuromonas acetoxidans
or D. acetivorans [see Eqn. (1); Pfennig & Biebl, 1976] yields
much less free energy (DG0=� 39 kJ mol�1) than acetate
oxidation in sulfate reducers (DG0=� 63 kJ mol�1 acetate).
In sulfur reducers most of the energy available from the
oxidation of 1 mol acetate with 4 mol sulfur is liberated
during the reduction of elemental sulfur to H2S rather than
during the oxidation of acetate. This was shown by Thauer
(1988) calculating the Gibbs free energy for each of the two
half reactions using the average redox potentials of NADP,
ferredoxin, menaquinone and NAD (E0=� 279 mV), of S0/
H2S (E0=� 240 mV), and of the CO2/acetate couple
(E0=� 291 mV).
CH3COO� þHþ þ 2H2O! 2CO2 þ 8½H�DG
0 ¼ �9:2 kJ mol�1 ð11Þ
8½H� þ 4S0 ! 4H2S
DG0 ¼ �29:8 kJ ð4 mol S0Þ�1 ð12Þ
In contrast, in the sulfate reducer Desulfobacter postgatei
(Widdel & Pfennig, 1981) when oxidizing 1 mol acetate with
1 mol sulfate it is the dehydrogenation of acetate [Eqn. (13)]
rather than the reduction of sulfate [SO42�/H2S:
E0=� 208 mV; Eqn. (14)] that is the major energy-yielding
reaction (Thauer, 1988). Again, DG0
for reactions (13) and
(14) were calculated from the average redox potential of the
four different electron acceptors used in the TCA cycle
(NADP, ferredoxin, menaquinone, and malate), which in
sulfate reducers (E0=� 241 mV) is supposed to be less
Table 1. Isotope enrichment factors for acetate during acetotrophic sulfur reduction by Desulfuromonas acetoxidans, Desulfuromonas thiophila,
Desulfurella acetivorans, and Hippea maritima�
Sulfur-reducing acetotroph f Acetate concentration (mM) eac (%) r2w
Desulfuromonas acetoxidans 0.0–0.5 3.8–1.9 � 11.45� 0.21 0.949
0.5–0.8 1.9–0.8 � 10.14� 0.61 0.997
0.8–1.0 0.8–0.0 � 2.85� 1.87 0.913
Desulfuromonas thiophila 0.0–0.5 5.3–2.7 � 11.19� 0.43 0.989
0.5–0.8 2.1–1.1 � 4.76� 0.24 0.999
0.8–1.0 1.1–0.0 ND
Desulfurella acetivorans 0.0–0.5 4.2–2.1 � 6.28� 1.30 0.910
0.5–0.8 2.1–0.8 � 3.50� 0.62 0.990
0.8–1.0 0.8–0.0 � 0.63 1
Hippea maritima 0.0–0.5 3.8–1.9 � 8.38� 1.04 0.978
0.5–0.9 1.9–0.4 � 1.20 1
0.9–1.0 0.4–0.0 � 0.52 1
�Values are means of triplicates.wCoefficient of determination. ND, not determined.
FEMS Microbiol Ecol 71 (2010) 218–225c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
222 D. Goevert & R. Conrad
negative than in the sulfur reducers:
CH3COO� þHþ þ 2H2O! 2CO2 þ 8½H�DG
0 ¼ �38:8 kJ mol�1 ð13Þ
8½H� þ SO2�4 þ 2Hþ ! H2Sþ 4H2O
DG0 ¼ �24:2 kJ mol�1 ð14Þ
Here, 2 mol of ATP are required for the activation of
sulfate. On the other hand, 1 mol of ATP is formed during
dehydrogenation of acetate, as it was found that the
synthesis of citrate from oxaloacetate and acetyl-CoA in
Desulfobacter postgatei is catalyzed by an ATP-citrate lyase
rather than by citrate synthase as in Desulfuromonas acetox-
idans (Moller et al., 1987; Schauder et al., 1987). The ATP-
citrate lyase in sulfate reducers with TCA probably results in
a smaller apparent isotope fractionation during acetate
consumption than the citrate synthase in sulfur reducers
with TCA (Table 2).
The energetics are also different between acetotrophic
Methanosarcina species (DG0=� 36 kJ mol�1 acetate) and
Desulfobacca acetoxidans (DG0=� 63 kJ mol�1 acetate)
(a) (d)
1.5
2.0
6.1
6.22.0 7.25
(b) (e)–18
–150.0
0.5
1.0
5.8
5.9
6.0
mm
ol
pH
–21
–18
0.0
0.5
1.0
1.5
7.05
7.10
7.15
7.20
mm
ol
pH(c)
–15(f)
–18
0 1 2 3 4 5 6 7
–27
–24
–21
δ13C
(‰
)
Time (days)0 1 2 3
–27
–24δ13C
(‰
)
Time (days)
–21
–18
δ13C
ac (
‰)
–24
–21
δ13C
ac (
‰)
–3 –2 –1 0–27
–24
ln (1-f)–4 –3 –2 –1 0
–27
ln (1-f)
Fig. 2. Acetate oxidation by sulfur-reducing bacteria activating acetate (presumably) using acetate kinase and phosphate acetyltransferase. The figure
shows substance conversions and isotope fractionation during the catabolism of acetate in pure cultures of Desulfurella acetivorans (a–c) and Hippea
maritima (d–f). (a and d) Acetate consumption (’), H2S production (�), and pH (dashed line without symbols); the concentrations are given as millimole
per bottle. (b and e) Isotopic composition of acetate (’) and CO2 (B, illustrated as TIC). (c and f) Isotope enrichment in acetate was calculated for three
different phases of acetate consumption: solid lines show substrate levels between 0 and �0.6 on the ln(1� f) scale (corresponding to up to 50%
acetate consumption), dashed lines show levels between � 0.6 and � 1.6 (or the next data point; 50 to 80%), and dotted lines show levels between
� 1.6 and � 3.5 (from 80% to maximum consumption of acetate); the plots are based on Eqn. (6). Values are means� SE (n = 3).
FEMS Microbiol Ecol 71 (2010) 218–225 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
223Acetotrophic sulfur-reducing bacteria

despite use of the same mechanism of acetate activation
(acetate kinase) and the same dissimilatory pathway (acetyl-
CoA pathway) (Table 2). The Methanosarcina species with
less energy available than Desulfobacca acetoxidans showed a
stronger fractionation. The catalytic basis of the different
isotope fractionation is unclear at present. However, we
hypothesize that the commitment for catalysis was larger in
Desulfobacca than in Methanosarcina spp.
In conclusion, we propose that closely related aceto-
trophic sulfur reducers (based on high 16S rRNA gene
sequence similarities) use the same mechanism for the
activation of acetate to acetyl-CoA and thus express similar
isotope fractionation of stable carbon. In general, the
apparent isotope fractionation in anaerobic acetotrophs is
probably affected not only by the acetate dissimilation
pathway and its key enzymes (citrate synthase, ATP-citrate
lyase, acetyl-CoA synthase complex) but also by the preced-
ing acetate activating enzymes (acetate kinase, acetyl-CoA
synthetase, succinyl-CoA : acetyl-CoA transferase) creating a
particular commitment of catalysis. These effects are fully
expressed only if acetate is not limiting. However, if the
concentration of acetate is so low that it is not fully available
to the microorganisms, the apparent stable isotope fractio-
nation is decreased (Thullner et al., 2008; Goevert & Con-
rad, 2009). Hence, quite a number of factors have to be
considered when acetate fractionation data collected in
nature are to be used for diagnosis of the operation of a
particular type of acetate dissimilation pathway or acetate-
dissimilating microbial community.
Acknowledgements
This work was supported by the Fonds der Chemischen
Industrie, Germany. We thank Peter Claus for excellent
technical assistance and Rudolf Thauer for helpful discus-
sions. We also thank two anonymous reviewers for helpful
comments.
References
Bonch-Osmolovskaya EA, Sokolova TG, Kostrikina NA &
Zavarzin GA (1990) Desulfurella acetivorans gen. nov. and sp.
nov. – a new thermophilic sulfur-reducing eubacterium. Arch
Microbiol 153: 151–155.
Chin KJ, Rainey FA, Janssen PH & Conrad R (1998)
Methanogenic degradation of polysaccharides and the
characterization of polysaccharolytic clostridia from anoxic
rice field soil. Syst Appl Microbiol 21: 185–200.
Conrad R (1999) Contribution of hydrogen to methane
production and control of hydrogen concentrations in
methanogenic soils and sediments (review). FEMS Microbiol
Ecol 28: 193–202.
Cord-Ruwisch R (1985) A quick method for the determination of
dissolved and precipitated sulfides in cultures of sulfate-
reducing bacteria. J Microbiol Meth 4: 33–36.
Deines P & Langmuir D (1974) Stable carbon isotope ratios and
the existence of a gas phase in the evolution of carbonate
ground waters. Geochim Cosmochim Ac 38: 1147–1164.
Elsner M, Zwank L, Hunkeler D & Schwarzenbach RP (2005) A
new concept linking observable stable isotope fractionation to
Table 2. Comparison of isotope enrichment factors for acetate during sulfur reduction, sulfate reduction, and methane production using different
pathways and activation mechanisms for acetate utilization
Microorganism
Electron
acceptor
Growth
temperature
( 1C) eac (%)
Acetate
dissimilation
pathway Acetate activation Reference
Desulfuromonas acetoxidans S0 reduction 30 � 11.5 TCA SuccCoA : acCoA transferase This paper
Desulfuromonas thiophila S0 reduction 30 � 11.2 TCA SuccCoA : acCoA transferase � This paper
Desulfurella acetivorans S0 reduction 55 �6.3 TCA Acetate kinase This paper
Hippea maritima S0 reduction 55 �8.4 TCA Acetate kinase � This paper
Desulfobacter postgatei SO42� reduction 30 11.8 TCA1citrate
lyase
SuccCoA : acCoA transferase Goevert & Conrad
(2008)
Desulfobacter hydrogenophilus SO42� reduction 30 11.5 TCA1citrate
lyase
SuccCoA : acCoA transferase Goevert & Conrad
(2008)
Desulfobacca acetoxidans SO42� reduction 37 � 19.1 acCoA Acetate kinase Goevert & Conrad
(2008)
Methanosarcina barkeri CH4 production 30 � 30.5 acCoA Acetate kinase Goevert & Conrad
(2009)
Methanosarcina acetivorans CH4 production 37 � 35.3 acCoA Acetate kinase Goevert & Conrad
(2009)
Methanosaeta concilii CH4 production 37 � 13.6 acCoA AcCoA synthetase Penning et al. (2006)
�Apparent mechanism of acetate activation based on carbon isotope fractionation.
acCoA, acetyl-CoA; succCoA, succinyl-CoA.
FEMS Microbiol Ecol 71 (2010) 218–225c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
224 D. Goevert & R. Conrad

transformation pathways of organic pollutants (review).
Environ Sci Technol 39: 6896–6916.
Finster K, Coates JD, Liesack W & Pfennig N (1997)
Desulfuromonas thiophila sp. nov., a new obligately sulfur-
reducing bacterium from anoxic freshwater sediment. Int J Syst
Bacteriol 47: 754–758.
Galushko AS & Schink B (2000) Oxidation of acetate through
reactions of the citric acid cycle by Geobacter sulfurreducens in
pure culture and in syntrophic coculture. Arch Microbiol 174:
314–321.
Gebhardt NA, Thauer RK, Linder D, Kaulfers PM & Pfennig N
(1985) Mechanism of acetate oxidation to CO2 with elemental
sulfur in Desulfuromonas acetoxidans. Arch Microbiol 141:
392–398.
Goevert D & Conrad R (2008) Carbon isotope fractionation by
sulfate-reducing bacteria using different pathways for the
oxidation of acetate. Environ Sci Technol 42: 7813–7817.
Goevert D & Conrad R (2009) Effect of substrate concentration
on carbon isotope fractionation during acetoclastic
methanogenesis by Methanosarcina barkeri and M. acetivorans
and in rice field soil. Appl Environ Microb 75: 2605–2612.
Hayes JM (1993) Factors controlling 13C contents of sedimentary
organic compounds: principles and evidence. Mar Geol 113:
111–125.
Kampara M, Thullner M, Richnow HH, Harms H & Wick LY
(2008) Impact of bioavailability restrictions on microbially
induced stable isotope fractionation. 2. Experimental
evidence. Environ Sci Technol 42: 6552–6558.
Kinnaman FS, Valentine DL & Tyler SC (2007) Carbon and
hydrogen isotope fractionation associated with the aerobic
microbial oxidation of methane, ethane, propane and butane.
Geochim Cosmochim Ac 71: 271–283.
Mariotti A, Germon JC, Hubert P, Kaiser P, Letolle R, Tardieux A
& Tardieux P (1981) Experimental determination of nitrogen
kinetic isotope fractionation: some principles; illustration for
the denitrification and nitrification processes. Plant Soil 62:
413–430.
Miroshnichenko ML, Rainey FA, Rhode M & Bonch-
Osmolovskaya EA (1999) Hippea maritima gen. nov., sp. nov.,
a new genus of thermophilic, sulfur-reducing bacterium from
submarine hot vents. Int J Syst Bacteriol 49: 1033–1038.
Moller D, Schauder R, Fuchs G & Thauer RK (1987) Acetate
oxidation to CO2 via a citric acid cycle involving an ATP-
citrate lyase: a mechanism for the synthesis of via substrate
level phosphorylation in Desulfobacter postgatei growing on
acetate and sulfate. Arch Microbiol 148: 202–207.
Mook WG, Bommerso JC & Staverma WH (1974) Carbon
isotope fractionation between dissolved bicarbonate and
gaseous carbon dioxide. Earth Planet Sc Lett 22: 169–176.
Morasch B, Richnow HH, Schink B & Meckenstock RU (2001)
Stable hydrogen and carbon isotope fractionation during
microbial toluene degradation: mechanistic and
environmental aspects. Appl Environ Microb 67: 4842–4849.
Northrop DB (1981) The expression of isotope effects on
enzyme-catalyzed reactions. Annu Rev Biochem 50: 103–131.
Penning H, Claus P, Casper P & Conrad R (2006) Carbon isotope
fractionation during acetoclastic methanogenesis by
Methanosaeta concilii in culture and a lake sediment. Appl
Environ Microb 72: 5648–5652.
Pfennig N & Biebl H (1976) Desulfuromonas acetoxidans gen. nov
and sp. nov., a new anaerobic, sulfur-reducing, acetate-
oxidizing bacterium. Arch Microbiol 110: 3–12.
Schauder R, Widdel F & Fuchs G (1987) Carbon assimilation
pathways in sulfate reducing bacteria: 2. Enzymes of a
reductive citric acid cycle in the autotrophic Desulfobacter
hydrogenophilus. Arch. Microbiology 148: 218–225.
Schmitz RA, Bonch-Osmolovskaya EA & Thauer RK (1990)
Different mechanisms of acetate activation in Desulfurella
acetivorans and Desulforomonas acetoxidans. Arch Microbiol
154: 274–279.
Soerensen J, Christensen D & Joergensen BB (1981) Volatile fatty
acids and hydrogen as substrates for sulfate-reducing bacteria
in anaerobic marine sediment. Appl Environ Microb 42: 5–11.
Stumm W & Morgan JJ (1995) Aquatic Chemistry. John Wiley
and Sons, New York.
Thauer RK (1988) Citric-acid cycle, 50 years on. Eur J Biochem
176: 497–508.
Thullner M, Kampara M, Ricnow HH, Harms H & Wick LY
(2008) Impact of bioavailability restrictions on microbially
induced stable isotope fractionation. 1. Theoretical
calculation. Environ Sci Technol 42: 6544–6551.
Widdel F & Pfennig N (1981) Studies on dissimilatory sulfate-
reducing bacteria that decompose fatty acids. 1. Isolation of
new sulfate-reducing bacteria enriched with acetate from
saline environments – description of Desulfobacter postgatei
gen. nov., sp. nov. Arch Microbiol 129: 395–400.
Wolin EA, Wolin MJ & Wolfe RS (1963) Formation of methane
by bacterial extracts. J Biol Chem 238: 2882–2886.
FEMS Microbiol Ecol 71 (2010) 218–225 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
225Acetotrophic sulfur-reducing bacteria