spata2 promotes cyld activity and regulates tnf...
TRANSCRIPT

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 1
Manuscript EMBO-2016-42592 SPATA2 promotes CYLD activity and regulates TNF-induced NF- B signaling and cell death Lisa Schlicher, Manuela Wissler, Florian Preiss, Prisca Brauns-Schubert, Celia Jakob, Veronica Dumit, Christoph Borner, Jörn Dengjel, and Ulrich Maurer Corresponding author: Ulrich Maurer, Albert Ludwigs University Freiburg Review timeline: Submission date: 21 April 2016 Editorial Decision: 13 May 2016 Revision received: 27 June 2016 Editorial Decision: 02 July 2016 Revision received: 04 July 2016 Accepted: 05 July 2016 Editor: Achim Breiling Transaction Report: (Note: With the exception of the correction of typographical or spelling errors that could be a source of ambiguity, letters and reports are not edited. The original formatting of letters and referee reports may not be reflected in this compilation.)
1st Editorial Decision 13 May 2016
Thank you for the submission of your research manuscript to EMBO reports. We have now received reports from the three referees that were asked to evaluate your study, which can be found at the end of this email. As you will see, all three referees acknowledge the high interest of the findings and the quality of the data and support publication after revision. All referees have raised a number of suggestions to improve the manuscript or to strengthen the data and the conclusions drawn. As the reports are below, I will not detail them here, but in particular all three referees suggest to analyze also K63 chains in the DUB assays. Given these constructive comments, we would like to invite you to revise your manuscript with the understanding that all referee concerns (as detailed in their reports) must be fully addressed in a complete point-by-point response. Acceptance of the manuscript will depend on a positive outcome of a second round of review. It is EMBO reports policy to allow a single round of revision only and acceptance or rejection of the manuscript will therefore depend on the completeness of your responses included in the next, final version of the manuscript. Revised manuscripts should be submitted within three months of a request for revision; they will otherwise be treated as new submissions. Please contact us if a 3-months time frame is not sufficient for the revisions so that we can discuss the revisions further. Supplementary/additional data: The Expanded View format, which will be displayed in the main

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 2
HTML of the paper in a collapsible format, has replaced the Supplementary information. You can submit up to 5 images as Expanded View. Please follow the nomenclature Figure EV1, Figure EV2 etc. The figure legend for these should be included in the main manuscript document file in a section called Expanded View Figure Legends after the main Figure Legends section. Additional Supplementary material should be supplied as a single pdf labeled Appendix. The Appendix includes a table of content on the first page, all figures and their legends. Please follow the nomenclature Appendix Figure Sx throughout the text and also label the figures according to this nomenclature. For more details please refer to our guide to authors. Important: All materials and methods should be included in the main manuscript file. Regarding data quantification and statistics, can you please specify the number "n" for how many experiments were performed, the bars and error bars (e.g. SEM, SD) and the test used to calculate p-values in the respective figure legends? This information must be provided in the figure legends. Please also include scale bars in all microscopy images. We now strongly encourage the publication of original source data with the aim of making primary data more accessible and transparent to the reader. The source data will be published in a separate source data file online along with the accepted manuscript and will be linked to the relevant figure. If you would like to use this opportunity, please submit the source data (for example scans of entire gels or blots, data points of graphs in an excel sheet, additional images, etc.) of your key experiments together with the revised manuscript. Please include size markers for scans of entire gels, label the scans with figure and panel number, and send one PDF file per figure or per figure panel. I look forward to seeing a revised version of your manuscript when it is ready. Please let me know if you have questions or comments regarding the revision. REFEREE REPORTS Referee #1: In this manuscript, Schlicher and colleagues report the identification and characterisation of SPATA2. They find that SPATA2 functions as critical adaptor linking the deubiquitylase CYLD to LUBAC. Previous work demonstrated that the E3 ligase LUBAC recruits CYLD into the TNFR1 signalling complex-I, where it restrains Ub-mediated activation of NFkB and enhances TNF-induced cell death. However, previous work failed to explain how CYLD was recruited to LUBAC. Here the authors demonstrate that SPATA2 binds to both LUBAC and CYLD and that SPATA2 is a novel component of the TNFR signalling complex-I. They demonstrate that SPATA2 is essential for the recruitment of CYLD into this complex. Knockout of SPATA2 abolishes CYLD recruitment to TNFR1 complex-I and alters the kinetics of NFkB signalling. The authors further show that SPATA2 interacts with HOIP and that SPATA2 competes with OTULIN for HOIP binding, suggesting that the interaction is mediated via the respective PIM and PUB domains. Consistent with its role to recruit CYLD to LUBAC, they find that knockout of SPATA2 increases M1-linked ubiquitylation of components of complex-I. Their data are consistent with a model whereby SPATA2 modulates the DUB activity of CYLD. Moreover, they find that knockout of SPATA2 sensitises cells to TNF-induced cell death in a RIPK1-dependent manner. All in all this is an interesting and timely ms, which reports an important finding. The ms is well written and the experiments clearly controlled. What is lacking is a more in depth analysis of complex-I and complex-II in the presence and absence of SPATA2. Below are a few comments that should help to improve the ms. Figure 1 • A better clarification of the identification of SPATA2 should be included. It is not clear how the mass spec experiment was performed, was SPATA2 identified upon TNF treatment and does the protein only bind CYLD in a TNF-dependent manner? In addition, the whole interactome of CYLD binding partners should be shown with respective spectral counts.

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 3
• In Figure 1C, the authors show that overexpressed CYLD is able to co-purify SPATA2. However, even though SPATA2 is clearly visible in the IP , there is no signal in the lysate. Its important to show the expression level of SPATA2 in the lysate. • In figure 1E, the authors IP CYLD upon SPATA2 overexpression, is the IP blot mislabelled as FLAG CYLD, should this be FLAG SPATA2? In addition, the authors show that the N-terminus of SPATA2 is able to IP CYLD. To support the conclusion that SPATA2 interacts with CYLD through its N-terminus the authors should show that the C-terminus of the protein doesn't co-purify CYLD. Figure 2 • The authors carry out a competition assay to investigate whether OTULIN and SPATA2 compete for the binding to HOIP. Here the authors overexpress SPATA2 but not Otulin and show that SPATA2 is able to outcompete Otulin for HOIP binding. To show a true competition, both SPATA2 and Otulin should be titrated against one another, and both molecules should be overexpressed. • Figure 2C and E. The authors show that SPATA2 is recruited to complex 1 in a TNF dependent manner. The authors only show CYLD as an established complex 1 component. The authors should carry out a proper complex 1 characterisation by showing that other established components such as RIPK1, Sharpin, cIAP 1/2, HOIP and HOIL are also recruited to the complex in a TNF dependent manner. Please refer to T.L. Haas et al. Mol. cell, 36 (2009), pp. 831-844 for the protocol on how to carry out a comprehensive complex 1 IP. Figure 3 • Fig 3A. Here the authors knockout SPATA2 and show that there is an increase in M1 linear chains in the TNFR signalling complex. Again, the authors need to show the TNF time dependent recruitment of established complex-I components such as HOIP, HOIL and TRADD and RIPK1 ubiquitylation upon TNF treatment. In addition, by showing other complex 1 components and the levels of TNFR1 itself, the authors could rule out that the effect seen on M1 ubiquitylation is not just due to a different stoichiometry of the complex 1 components in the WT and SPATA2 knockout MEFs. • Figure 3C. The authors show by a DUB activity assay that in the presence of SPATA2, there is increased monoubiquitin. In order to make the conclusion that SPATA2 enhances the M1 DUB activity of CYLD, the authors need to boil the beads and show that the amount of CYLD is equal between the samples. • In addition, CYLD has an established role in digesting K63 chains, the authors don't comment on the effect that SPATA2 has on this linkage type. It would be interesting to know whether loss of SPATA2 also impacts on the ability of CYLD to influence K63 chains in complex 1. Figure 4 • Fig 4A and 4B. Here the authors look at the effect of SPATA2 knockout on NFkB signalling. IkB degradation upon TNF treatment is a hallmark of NFkB signalling but it is also necessary that the authors show that this coincides with IkB phosphorylation and p65 phosphorylation. In addition, the authors should carry out cyotkine ELISA to investigate whether SPATA2 knockout also increases the levels of cytokines such as IL6, IL8 and TNF. • Fig 4C. Here the authors investigate the role of SPATA2 in cell death upon TNF treatment. It is recommended that the authors carry out complex-II isolation assays as outlined by Feoktistova et al. Mol Cell 2011; 43: 449-463 in wild type and SPATA2 knockout MEFS to support their conclusion that SPATA2 is required for TNF induced cell death. Minor comments: General note: Blots should include molecular weight of proteins Referee #2: This study identified SPATA2 as a binding partner of CYLD by Mass Spectrometry (Fig. 1A&B). The authors confirm and define the CYLD interaction in a number of assays (Figure 1&2) and then explore the consequences of SPATA2 loss on TNF signalling.

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 4
Identification of a Linear Ubiquitin Assembly Complex (LUBAC) / CYLD binding factor is highly topical and will be of interest to EMBOR readers. The affects of SPATA2 expression on TNF signalling also make this of general interest. Conceptually there are no problems and I believe the manuscript mostly satisfies the published criteria for EMBOR, namely: 2. Physiological/functional relevance demonstrated (detailed insight into the mechanism is not always necessary) - Some relevance data with knock-out cells 3. Strong evidence for the conclusions that are drawn - reasonable 4. Novelty - high, new player 5. Broad biological significance - yes LUBAC and M1 ubiquitylation 6. Importance to the specific field - yes I have made a number of technical comments that would improve the quality of the manuscript. Specific Remarks Fig. 2B one presumes that Spata2 doesn't bind to Otulin and that in Fig. 2B that CYLD binding increases. It would be important to demonstrate this to support the authors' contention that "it is the competition of SPATA2 and OTULIN, which underlies the mutual exclusive binding of CYLD and OTULIN to the LUBAC complex". Fig. 2C yes there is an increase in Spata2 and cyld at 5 minutes post TNF but this data as it stands is not convincing. When others do these assays (eg Draber et al) there is really no CYLD at t=0 and it is hard to understand why there is here. Fig. 2E unfortunately in Fig. 2E there is the same problem of binding of cyld at t=0 and also there is so little signal in the wild type that it is hard to interpret the lack of cyld binding in spata2 knock-out cells. Also need to see that HOIP is recruited. Fig. 3A same comments as to 2E would like to see HOIP, HOIL or Sharpin binding. page 8: "whether SPATA2 dependent recruitment of CYLD to the TNF-RSC results in increased M1 ubiquitylation", don't they mean decreased? The lack of the protein causes increased M1 not its presence. Fig. 3C There is a hint that SPATA CYLD has more activity but it is not a huge difference and I think it would be important to repeat this a few times to be sure. It would be good to also examine K63 diubiquitin chains, perhaps the effect is more pronounced on this chain type. And most impressive to have a control for a chain type that it doesn't cleave to show that specificity of CYLD doesn't change when bound to SPATA2. Fig. 4A NF-κB is good, MAPK signalling might be more convincing if the authors included p38 and ERK. Fig. 4B The lack of a JNK effect in Sertoli cell line makes one wonder how robust the effect in MEFs is. I recommend looking at other MAPKs in Sertoli cells. Fig. 4C perhaps the most difficult (and interesting) result of the whole manuscript. This is really hard to reconcile with increased complex 1 formation in Spata2 knock-out cells because TAK1 is not present to drive activation of IKK2 and NF-κB. The most obvious explanation is that increased M1 ubiquitylation directly prevents/reduces the formation of complex 2. Caspase-8 pull downs are a relatively easy and quick way to test this hypothesis. An experiment of this nature is required because otherwise there is no good explanation for why loss of Spata2 reduces cell death. Referee #3: Schlicher et al. identified SPATA2 as new interaction partner of CYLD and therefore, as a new component of the TNF-RSC by using quantitative mass spectrometry. They found that SPATA2 interacts with CYLD and that it is required for the recruitment of CYLD

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 5
to the TNF-RSC. Moreover, depletion of SPATA2 prevents the recruitment of CYLD to the TNF-RSC and most strikingly results in increased ubiquitination of RIPK1, a well-known LUBAC target. In addition, authors showed that SPATA2 is involved in NFκB signaling and TNF-α -induced apoptosis. Taken together, the study from Schlicher et al. provides exciting new insights into the composition and assembly of the TNF-RSC. It was established, that CYLD is a part of the TNF-RSC. However, it was not known how CYLD is recruited to the receptor complex. Draber et al. (Cell Rep. 2015) demonstrated for the first time, that it is CYLD's interaction with LUBAC via HOIP that is responsible for it´s recruitment. In the present manuscript authors propose, that SPATA2 acts as a critical adaptor for CYLD and LUBAC and mediates their recruitment to the TNF-RSC. This is a novel and important finding that can be further improved with the following: Specific comments/suggestions: 1. Authors should try to map the interaction domain of SPATA2 and HOIP. Coulf authors map the potential PUB interacting motif (PIM) of SPATA2 that is similar to the motif defined in OTULIN (Scheefer et al. Mol Cell; Eliot et all Mol Cell) 2. Draber et al. (Cell Reports) showed, that two pools of LUBAC exist in the cytosol: one is associated with CYLD, whilst another one is associated with OTULIN. In that study authors showed, that the interaction of HOIP with OTULIN is abrogated upon high SPATA2 concentrations. The authors should check, if under these conditions CYLD is preferentially recruited to HOIP and if CYLD´s binding to HOIP correlate with the level of SPATA2 expression. 3. The authors claim that SPATA2 function as an allosteric activator for CYLD´s DUB-activity. This is a very important part of the manuscript and should be further supported by the following experiments a. For Fig. 3c authors need to provide a loading control of CYLD input. b. It is known that CYLD cleaves M1-, as well as K-63 -linkages. Both linkage types are of critical importance for TNF-RSC assembly and for the signaling output. Therefore, authors should also include K-63 Di-Ub chains for the DUB assay. It might be that activation is specific for only M1 chains or for both linear and K63 chains Minor comments: 1. Fig. 1E is labeled wrong. Flag-SPATA2 was immunoprecipitated.. 2. Fig 2E & Fig 3A: Authors should provide blots with longer exposure times to prove, that depletion of SPATA2 really abolishes the recruitment of CYLD to the TNF-RSC. 3. Fig. 3A: Why is in the wt settings no M1-ubiquitination detected after 20 min TNF-stimulation? Other groups showed also in MEF cells strongly increased M1- ubiquitination signals after 20 min TNF stimulation at the TNF-RSC. Is it cell type specific difference? 4. Authors should include anti-SPATA2 staining in experiments, when using SPATA2 KO cells, to prove that SPATA2 was indeed depleted in the particular setting (Fig. 2E, 3A, 4A-C). 1st Revision - authors' response 27 June 2016
We would like to thank the reviewers for their highly valuable, thoughtful and constructive input. Based on the suggestions of all three reviewers, we think we could significantly improve the paper. By having added a number of new and improved experiments, we think the study provides more profound insight into the mechanism, as we hope the reviewers will agree. Referee #1: In this manuscript, Schlicher and colleagues report the identification and characterisation of SPATA2. They find that SPATA2 functions as critical adaptor linking the deubiquitylase CYLD to

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 6
LUBAC. Previous work demonstrated that the E3 ligase LUBAC recruits CYLD into the TNFR1 signalling complex-I, where it restrains Ub-mediated activation of NFkB and enhances TNF-induced cell death. However, previous work failed to explain how CYLD was recruited to LUBAC. Here the authors demonstrate that SPATA2 binds to both LUBAC and CYLD and that SPATA2 is a novel component of the TNFR signalling complex-I. They demonstrate that SPATA2 is essential for the recruitment of CYLD into this complex. Knockout of SPATA2 abolishes CYLD recruitment to TNFR1 complex-I and alters the kinetics of NFkB signalling. The authors further show that SPATA2 interacts with HOIP and that SPATA2 competes with OTULIN for HOIP binding, suggesting that the interaction is mediated via the respective PIM and PUB domains. Consistent with its role to recruit CYLD to LUBAC, they find that knockout of SPATA2 increases M1-linked ubiquitylation of components of complex-I. Their data are consistent with a model whereby SPATA2 modulates the DUB activity of CYLD. Moreover, they find that knockout of SPATA2 sensitises cells to TNF-induced cell death in a RIPK1-dependent manner. All in all this is an interesting and timely ms, which reports an important finding. The ms is well written and the experiments clearly controlled. What is lacking is a more in depth analysis of complex-I and complex-II in the presence and absence of SPATA2. Below are a few comments that should help to improve the ms. Figure 1 • A better clarification of the identification of SPATA2 should be included. It is not clear how the mass spec experiment was performed, was SPATA2 identified upon TNF treatment and does the protein only bind CYLD in a TNF-dependent manner? In addition, the whole interactome of CYLD binding partners should be shown with respective spectral counts. This mass spec experiment was originally performed with a different hypothesis. In an ongoing study on CYLD phosphorylation by TAK1, we wanted to identify CYLD phosphorylation-dependent interaction partners. Thus, the SILAC interaction screen was done with three conditions a.) CYLD ko cells, b.) CYLD ko cells expressing FLAG-CYLD and c.) CYLD ko cells expressing FLAG-CYLD in which TAK1 was knocked down by stable expression of an shRNA. In all three conditions, a TNF stimulation of 20 min was done (in order to activate TAK1). In this experiment, we found SPATA2 being in complex with FLAG-CYLD independently from TAK1 knockdown or not. For this reason, we did not describe the TAK1 part of the experiment, as it is irrelevant for our finding. Also, experiments shown in new Figures 1C-E, and Figures 2A,B, D and E, along with data from others (Draber et al. 2015) indicate that the interaction of SPATA2 with CYLD and HOIP does not depend TNFR complex 1 formation. This is now discussed in the discussion section. We also attached to this rebuttal, for the reviewers’ information, the complete dataset of the MS experiment, including all three SILAC conditions, with the corresponding intensities measured, that are calculated from the original spectra (this includes columns with the 'ratio count '). Please note, that for a better understanding in the manuscript, we relabelled the original “medium” condition (CYLD ko cells expressing FLAG-CYLD) to “heavy” condition, as the original “heavy” condition (CYLD ko cell expressing FLAG-CYLD in which TAK1 was knocked down by stable expression of an shRNA) is not part of the manuscript. In the revised manuscript, we now also provide a list of the other proteins we have found to be associated with CYLD, while SPATA2 clearly stood out with an eight-fold enrichment as compared to a two-fold enrichment (at best) for other proteins. This is shown in a new Fig. 1B in the manuscript. • In Figure 1C, the authors show that overexpressed CYLD is able to co-purify SPATA2. However, even though SPATA2 is clearly visible in the IP, there is no signal in the lysate. Its important to show the expression level of SPATA2 in the lysate. The commercial anti-SPATA2 antibody we have used in this experiment was not very sensitive and did hardly pick up endogenous SPATA2. We have now redone the experiment using a different antibody and we were able to detect endogenous SPATA2 expression in the lysate. This is shown in new Fig 1C.
EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 7
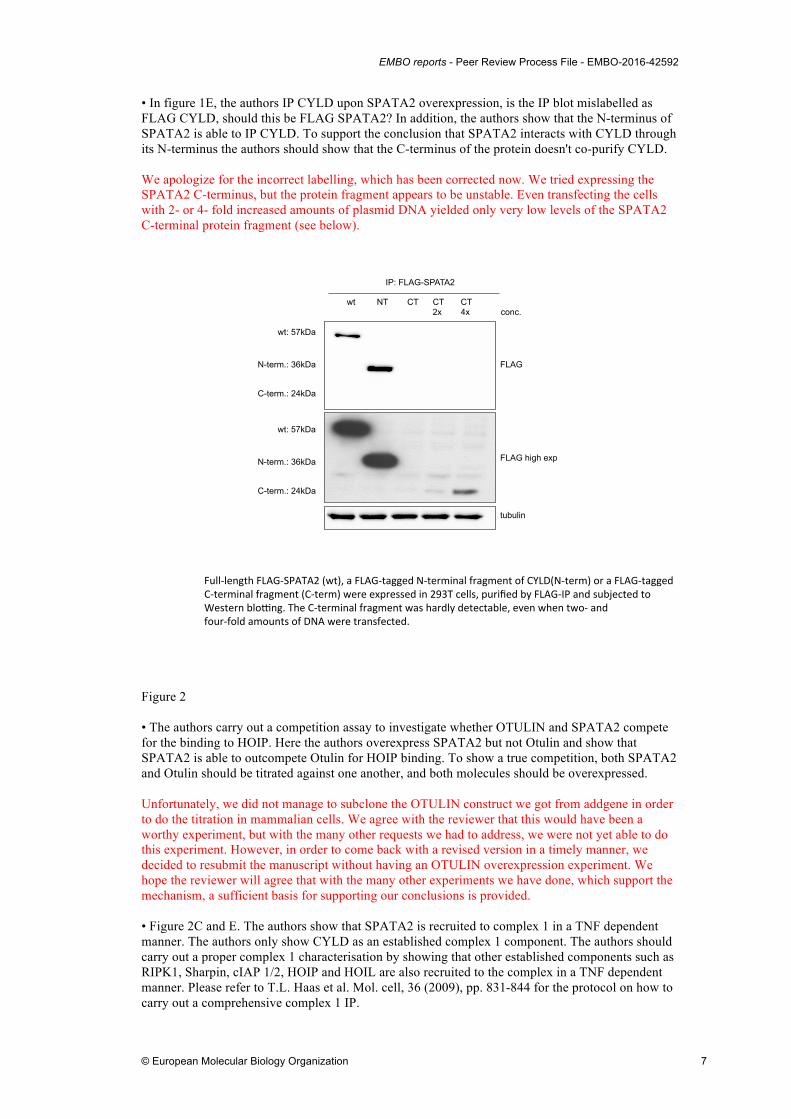
• In figure 1E, the authors IP CYLD upon SPATA2 overexpression, is the IP blot mislabelled as FLAG CYLD, should this be FLAG SPATA2? In addition, the authors show that the N-terminus of SPATA2 is able to IP CYLD. To support the conclusion that SPATA2 interacts with CYLD through its N-terminus the authors should show that the C-terminus of the protein doesn't co-purify CYLD. We apologize for the incorrect labelling, which has been corrected now. We tried expressing the SPATA2 C-terminus, but the protein fragment appears to be unstable. Even transfecting the cells with 2- or 4- fold increased amounts of plasmid DNA yielded only very low levels of the SPATA2 C-terminal protein fragment (see below).
Figure 2 • The authors carry out a competition assay to investigate whether OTULIN and SPATA2 compete for the binding to HOIP. Here the authors overexpress SPATA2 but not Otulin and show that SPATA2 is able to outcompete Otulin for HOIP binding. To show a true competition, both SPATA2 and Otulin should be titrated against one another, and both molecules should be overexpressed. Unfortunately, we did not manage to subclone the OTULIN construct we got from addgene in order to do the titration in mammalian cells. We agree with the reviewer that this would have been a worthy experiment, but with the many other requests we had to address, we were not yet able to do this experiment. However, in order to come back with a revised version in a timely manner, we decided to resubmit the manuscript without having an OTULIN overexpression experiment. We hope the reviewer will agree that with the many other experiments we have done, which support the mechanism, a sufficient basis for supporting our conclusions is provided. • Figure 2C and E. The authors show that SPATA2 is recruited to complex 1 in a TNF dependent manner. The authors only show CYLD as an established complex 1 component. The authors should carry out a proper complex 1 characterisation by showing that other established components such as RIPK1, Sharpin, cIAP 1/2, HOIP and HOIL are also recruited to the complex in a TNF dependent manner. Please refer to T.L. Haas et al. Mol. cell, 36 (2009), pp. 831-844 for the protocol on how to carry out a comprehensive complex 1 IP.
FLAG
FLAG high exp
tubulin
wt NT CT CT CT 2x 4x conc.
IP: FLAG-SPATA2
wt: 57kDa
N-term.: 36kDa
C-term.: 24kDa
wt: 57kDa
N-term.: 36kDa
C-term.: 24kDa
Full$length*FLAG$SPATA2*(wt),*a*FLAG$tagged*N$terminal*fragment*of*CYLD(N$term)*or*a*FLAG$tagged**C$terminal*fragment*(C$term)*were*expressed*in*293T*cells,*purified*by*FLAG$IP*and*subjected*to**Western*bloMng.*The*C$terminal*fragment*was*hardly*detectable,*even*when*two$*and**four$fold*amounts*of*DNA*were*transfected.*

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 8
We would like to thank the reviewer for this comment as the requested experiments have made our data more convincing, as we believe, and we hope the reviewer will agree. Unfortunately, the antibodies we could obtain for the detection of SHARPIN, HOIL and HOIP did not work with the murine proteins. Therefore, we have now done the TNFR pull-down with human A549 cells, which are an established model system for TNFR complex formation (Draber et al. 2015). Using this system, we show that endogenous SPATA2, CYLD, SHARPIN, RIPK1 and TRADD are recruited after 5 and 15 min (but not at time zero) to the TNFR complex. This is now shown in new Fig 2F. Figure 3 • Fig 3A. Here the authors knockout SPATA2 and show that there is an increase in M1 linear chains in the TNFR signalling complex. Again, the authors need to show the TNF time dependent recruitment of established complex-I components such as HOIP, HOIL and TRADD and RIPK1 ubiquitylation upon TNF treatment. In addition, by showing other complex 1 components and the levels of TNFR1 itself, the authors could rule out that the effect seen on M1 ubiquitylation is not just due to a different stoichiometry of the complex 1 components in the WT and SPATA2 knockout MEFs. Again, this was a very helpful suggestion. As mentioned above, we were not able to show recruitment of HOIP or SHARPIN to the TNF receptor complex I as the antibodies did not work well with MEF. To show the dependence of CYLD recruitment on the presence of SPATA2, we now created SPATA2 knockout A549 cells by CRISPR/Cas9 and show that in cells lacking SPATA2, reduced amounts of SPATA2 and CYLD were recruited to the TNFR complex (note that during the revision of the manuscript we could only generate mixed cell CRIPR/Cas9 populations but not select single cell clones, therefore the loss of SPATA2 was not 100%. Nevertheless the effects are, as we hope the reviewer will agree, quite clear). In contrast, the recruitment of SHARPIN and HOIP was not affected by the absence of SPATA2. • Figure 3C. The authors show by a DUB activity assay that in the presence of SPATA2, there is increased monoubiquitin. In order to make the conclusion that SPATA2 enhances the M1 DUB activity of CYLD, the authors need to boil the beads and show that the amount of CYLD is equal between the samples. We apologise, as the experiment was not well described. Actually the Western blot of the FLAG-IP in Figure 3B represents 50 % of the very same CYLD isolates which were subjected to the assay shown in Figure 3C. We have repeated this experiment, with an improved way to normalize for CYLD input (see below). • In addition, CYLD has an established role in digesting K63 chains, the authors don't comment on the effect that SPATA2 has on this linkage type. It would be interesting to know whether loss of SPATA2 also impacts on the ability of CYLD to influence K63 chains in complex 1. Again this was a very helpful suggestion resulting in an improved experiment, as we think, and we highly appreciate this input. We have now used as substrates not only M1-linked di-ubiquitin, but also K63-linked and K48-linked di-ubiquitin. We again found that the presence of SPATA2 increased CYLD DUB activity with the M1-, but also with the K63-linked di-ubiquitin. Consistent with the findings by others (Komander et al. 2009) the K48-linked di-ubiquitin was not cleaved by CYLD, and this was not changed by the presence of SPATA2. We believe that low levels of mono-ubiquitin are resulting from a contaminating K48-specific DUB, as it was present also with the inactive CYLDC598A. For improved normalization, we have cut the gel after electrophoresis. The lower part was silver stained for detection of di-and mono-ubiquitin, and the upper part was transferred to a membrane and probed with antibodies for FLAG (CYLD) and V5 (SPATA2). Thus the amounts of FLAG-CYLD represent the very same protein resulting in di-ubiquitin cleavage. This is now shown in new Fig. 4A-C.

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 9
Figure 4 • Fig 4A and 4B. Here the authors look at the effect of SPATA2 knockout on NFkB signalling. IkB degradation upon TNF treatment is a hallmark of NFkB signalling but it is also necessary that the authors show that this coincides with IkB phosphorylation and p65 phosphorylation. In addition, the authors should carry out cyotkine ELISA to investigate whether SPATA2 knockout also increases the levels of cytokines such as IL6, IL8 and TNF. We have now repeated this experiment with both MEF and 15P-1 Sertoli cells (from which we now have generated single cell clones) and observed, in the absence of SPATA2, increased p65 and IkBa phosphorylation, and increased phosphorylation of the MAP Kinases JNK, ERK and p38. These data are now shown in new Fig. 5A and B (note that the lower signal for phospho-IkBa in Fig. 5A with SPATA ko MEF after 5 min is certainly due to low levels of IkBa due to degradation at this time point, as evident from the probing for IkBa below). Unfortunately, we were not able to optimize the qPCR for the detection of various cytokines. While we observed elevated mRNA levels with SPATA2 ko cells for, TNFa and IL1a, but we were not confident enough in the quality of the PCR and the generality of the result but could to date not repeat the experiment more often. In the interest of a timely resubmission, and with the data now shown in Fig. 5A and B, we hope that the reviewer will agree that there is solid support for our conclusions. • Fig 4C. Here the authors investigate the role of SPATA2 in cell death upon TNF treatment. It is recommended that the authors carry out complex-II isolation assays as outlined by Feoktistova et al. Mol Cell 2011; 43: 449-463 in wild type and SPATA2 knockout MEFS to support their conclusion that SPATA2 is required for TNF induced cell death. Again we would like to thank the reviewer for a very useful suggestion. We have performed immuno-precipitations with an anti-FADD antibody after stimulation of TAK1-/- MEF, lacking or retaining SPATA2. We found a strongly reduced association of RIPK1 with FADD in the absence of SPATA2, while RIPK1 was associated with FADD, indicating proper complex II formation, in SPATA wild-type cells. Unfortunately, despite trying a number of different antibodies for detection of FLIP and caspase-8, we did not find antibodies which sufficiently in the mouse system. Nevertheless, we think the experiment is convincing, as we hope the reviewer agrees. This is now shown in new Fig. 5F. Minor comments: General note: Blots should include molecular weight of proteins The molecular weight is now indicated on all Figures. Again, we would like to thank the reviewer for his/her time, effort and valuable input. Referee #2: This study identified SPATA2 as a binding partner of CYLD by Mass Spectrometry (Fig. 1A&B). The authors confirm and define the CYLD interaction in a number of assays (Figure 1&2) and then explore the consequences of SPATA2 loss on TNF signalling. Identification of a Linear Ubiquitin Assembly Complex (LUBAC) / CYLD binding factor is highly topical and will be of interest to EMBOR readers. The affects of SPATA2 expression on TNF signalling also make this of general interest. Conceptually there are no problems and I believe the manuscript mostly satisfies the published criteria for EMBOR, namely: 2. Physiological/functional relevance demonstrated (detailed insight into the mechanism is not always necessary) - Some relevance data with knock-out cells 3. Strong evidence for the conclusions that are drawn - reasonable 4. Novelty - high, new player

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 10
5. Broad biological significance - yes LUBAC and M1 ubiquitylation 6. Importance to the specific field - yes I have made a number of technical comments that would improve the quality of the manuscript. Specific Remarks Fig. 2B one presumes that Spata2 doesn't bind to Otulin and that in Fig. 2B that CYLD binding increases. It would be important to demonstrate this to support the authors' contention that "it is the competition of SPATA2 and OTULIN, which underlies the mutual exclusive binding of CYLD and OTULIN to the LUBAC complex". We have addressed this important request by expressing FLAG-HOIP or FLAG-SPATA2, followed by FLAG-IP. While FLAG-HOIP pulled down SPATA2 and OTULIN, FLAG-SPATA co-immunoprecipated with CYLD, but not with OTULIN. This shows the absence of a SPATA2-OTULIN interaction and is now included in the manuscript in Fig. 2D. We tried to reprobe the blot with an anti-CYLD antibody, but unfortunately we could not activate the blot anymore. We planned to redo the experiment, but in the meantime we obtained a result, which shows that CYLD binds to the TNFR complex exactly to the same extent as does SPATA2. This is now shown in new Fig. 3A. By obtaining these data we decided to focus on the other experiments requested by the reviewer and we hope the reviewer will agree that the data sufficiently support our conclusions. Fig. 2C yes there is an increase in Spata2 and cyld at 5 minutes post TNF but this data as it stands is not convincing. When others do these assays (eg Draber et al) there is really no CYLD at t=0 and it is hard to understand why there is here. Again this was an important and helpful request as we think that the resulting experiment helped to improve the manuscript. We used A549 cells, as in the Draber et al. (2015) paper and found SPATA2, CYLD, SHARPIN, RIPK1 and TRADD are recruited after 5 and 15 min (but not at time zero) to the TNFR complex. This result is now shown in new Fig. 2F. Fig. 2E unfortunately in Fig. 2E there is the same problem of binding of cyld at t=0 and also there is so little signal in the wild type that it is hard to interpret the lack of cyld binding in spata2 knock-out cells. Also need to see that HOIP is recruited. This experiment was redone as well with A549, infected with CRISPR/Cas9, targeting the spata2 gene (a mixed cell culture was generated). Using these cells, exhibiting highly reduced SPATA2 expression, we can show that recruitment of CYLD is abrogated, while the recruitment of SHARPIN and HOIP is not affected by the presence or absence of SPATA2. The data are now shown in new Fig. 3A. Fig. 3A same comments as to 2E would like to see HOIP, HOIL or Sharpin binding. Unfortunately we could not obtain antibodies for SHARPIN and HOIP, which detect the mouse proteins. We hope that the reviewer will agree that experiment shown in the new Fig. 3A, showing SPATA2 independent recruitment of SHARPIN and HOIP in human cells, sufficiently supports the mechanism we postulate. page 8: "whether SPATA2 dependent recruitment of CYLD to the TNF-RSC results in increased M1 ubiquitylation", don't they mean decreased? The lack of the protein causes increased M1 not its presence. We apologize for this error and thank the reviewer for pointing this out. We have corrected this in the revised version. Fig. 3C There is a hint that SPATA CYLD has more activity but it is not a huge difference and I think it would be important to repeat this a few times to be sure. It would be good to also examine K63 diubiquitin chains, perhaps the effect is more pronounced on this chain type. And most impressive to have a control for a chain type that it doesn't cleave to show that specificity of CYLD

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 11
doesn't change when bound to SPATA2. We thank the reviewer for this valuable suggestion. We have repeated this experiment, and can show now that the interaction of CYLD with SPATA2 enhances not only its M1-linkage-specific, but also the K63 linkage-specific DUB activity of CYLD. In contrast the absence of cleavage of K48-linked di-ubiquitin substrates remained unchanged. This is now shown in new Fig. 4 A-C. Fig. 4A NF-κB is good, MAPK signalling might be more convincing if the authors included p38 and ERK. We have redone the experiments with MEF lacking SPATA2 and probed for p65 and IkBa phosphorylation and phospho-p38, -JNK and -ERK, all of which exhibit stronger phosphorylation in the absence of SPATA2. This is now shown in new Fig. 5A (note that the lower signal for phospho-IkBa in Fig. 5A with SPATA ko MEF after 5 min is certainly due to IkBa degradation at this time point, as evident from the probing for IkBa below). Fig. 4B The lack of a JNK effect in Sertoli cell line makes one wonder how robust the effect in MEFs is. I recommend looking at other MAPKs in Sertoli cells. We have now repeated the TNF stimulation of 15P-1 Sertoli cells lacking or retaining SPATA2, having now generated single cell clones after CRISPR/Cas9 mediated knock-out of SPATA2. We found that the absence of SPATA2 indeed affects the phosphorylation/activation of MAP Kinases and p65. In the absence of SPATA2, we observed increased p65 and IkBa phosphorylation, and increased phosphorylation of IkBa as well as MAPKinases JNK, ERK and p38. We think that in the original manuscript, the use of a mixed 15P-1 cell population generated by CRSPR/Cas9 targeteing the spata2 gene, and therefore containing a fraction of cells retaining SPATA2 expression, has masked the effects on JNK phosphorylation. The new result is now shown in new Fig. 5B. We would like to thank the reviewer for this valuable suggestion. Fig. 4C perhaps the most difficult (and interesting) result of the whole manuscript. This is really hard to reconcile with increased complex 1 formation in Spata2 knock-out cells because TAK1 is not present to drive activation of IKK2 and NF-κB. The most obvious explanation is that increased M1 ubiquitylation directly prevents/reduces the formation of complex 2. Caspase-8 pull downs are a relatively easy and quick way to test this hypothesis. An experiment of this nature is required because otherwise there is no good explanation for why loss of Spata2 reduces cell death. Again, we thank the reviewer for this valuable suggestion. We have done a complex II IP using an anti-FADD antibody. We found that, while in the presence of SPATA2, TNF stimulation results in pulldown of RIPK1, indicating formation of complex II, while this was not observed in absence of SPATA2, indicating the absence of complex II formation. We intended to show recruitment of FLIP and caspase-8 to FADD as well but among a number of different antibodies we tested, not a single one appeared to work with mouse protein to detect FLIP or caspase-8. We nevertheless think the result is convincing, and we hope the reviewer will agree. This result is now shown in Fig. 5. As with reviewer #1, we thank the reviewer for his/her input, which, as we think helped substantially to improve the paper. Referee #3: Schlicher et al. identified SPATA2 as new interaction partner of CYLD and therefore, as a new component of the TNF-RSC by using quantitative mass spectrometry. They found that SPATA2 interacts with CYLD and that it is required for the recruitment of CYLD to the TNF-RSC. Moreover, depletion of SPATA2 prevents the recruitment of CYLD to the TNF-RSC and most strikingly results in increased ubiquitination of RIPK1, a well-known LUBAC target. In addition, authors showed that SPATA2 is involved in NFκB signaling and TNF-α -induced apoptosis.

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 12
Taken together, the study from Schlicher et al. provides exciting new insights into the composition and assembly of the TNF-RSC. It was established, that CYLD is a part of the TNF-RSC. However, it was not known how CYLD is recruited to the receptor complex. Draber et al. (Cell Rep. 2015) demonstrated for the first time, that it is CYLD's interaction with LUBAC via HOIP that is responsible for it´s recruitment. In the present manuscript authors propose, that SPATA2 acts as a critical adaptor for CYLD and LUBAC and mediates their recruitment to the TNF-RSC. This is a novel and important finding that can be further improved with the following: Specific comments/suggestions: 1. Authors should try to map the interaction domain of SPATA2 and HOIP. Coulf authors map the potential PUB interacting motif (PIM) of SPATA2 that is similar to the motif defined in OTULIN (Scheefer et al. Mol Cell; Eliot et all Mol Cell) We would like to thank the reviewer for this highly valuable suggestion. We have generated a PIM mutant (Y338A) of SPATA2 (in analogy to the OTULIN PIM mutant Y56A as published (Elliott et al. 2014, Schaeffer et al. 2014) and also a PUB domain mutant (F108A) of SPATA2. By Co-IP of V5-tagged SPATA2 carrying these mutations, we observed that the PUB domain mutation of SPATA2 abolished the binding to CYLD, whereas the PIM domain mutant of SPATA2 did not interact with HOIP, but retained the ability to bind CYLD. We think that this experiment suggested by the reviewer provides a much more detailed understanding of the interaction of those three proteins. These results are now shown in Fig. 2 B and E. 2. Draber et al. (Cell Reports) showed, that two pools of LUBAC exist in the cytosol: one is associated with CYLD, whilst another one is associated with OTULIN. In that study authors showed, that the interaction of HOIP with OTULIN is abrogated upon high SPATA2 concentrations. The authors should check, if under these conditions CYLD is preferentially recruited to HOIP and if CYLD´s binding to HOIP correlate with the level of SPATA2 expression. We tried to reprobe the blot as shown in Fig. 2B of the original manuscript with an anti-CYLD antibody, but unfortunately the detection did not work anymore with the membrane having dried for some weeks. We then intended to do the experiment again, but in the meantime we obtained the result now shown in new Fig. 3A. This clearly shows that the binding of CYLD to the TNFR complex is reduced exactly to the extent of SPATA2 reduction at the TNFR complex. Thus we think the correlation of CYLD binding to HOIP, depending on SPATA2 expression can be deduced from this experiment. We hope the reviewer will agree. 3. The authors claim that SPATA2 function as an allosteric activator for CYLD´s DUB-activity. This is a very important part of the manuscript and should be further supported by the following experiments a. For Fig. 3c authors need to provide a loading control of CYLD input. b. It is known that CYLD cleaves M1-, as well as K-63 -linkages. Both linkage types are of critical importance for TNF-RSC assembly and for the signaling output. Therefore, authors should also include K-63 Di-Ub chains for the DUB assay. It might be that activation is specific for only M1 chains or for both linear and K63 chains As with reviewer #1 and #2, we thank this reviewer for a very valuable suggestion and we think that the result has made the paper considerably stronger, as we hope the reviewer will agree. We apologize for not having clearly indicated that Figure 3C represented the loading control for the assay shown if Figure 4C, as 50% of the material used for the assay shown in Figure 3C was loaded on the gel shown in Fig 3C. We have now repeated the experiment with M1-, K63- and K48-linked di-ubiquitin substrates. We now show that that presence of SPATA2 augments CYLD enzymatic activity towards M1-linked and K63-linked di-ubiquitin substrates, while CYLD does not cleave K48-linked di-ubiquitin, and this was independent of presence or absence of SPATA2. To directly control for the amounts of CYLD resulting in di-ubquitin cleavage, the gel was now cut in two pieces after electrophoresis, and the lower part was silver stained for the detection of di-and mono-ubiquitin, while the upper part was transferred to a membrane and probed for FLAG (CYLD)

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 13
and V5 (SPATA). Thus the anti-FLAG signal represents exactly the amount of CYLD resulting in the cleavage (or not) in the respective condition. These data are now shown in new Fig. 4 A-C. Minor comments: 1. Fig. 1E is labeled wrong. Flag-SPATA2 was immunoprecipitated.. We apologize for this mistake and thank the reviewer for having eyed this. This has been corrected. 2. Fig 2E & Fig 3A: Authors should provide blots with longer exposure times to prove, that depletion of SPATA2 really abolishes the recruitment of CYLD to the TNF-RSC. Using human A549 cells we were able to obtain results, which clearly show that in the SPATA2 CRISPR/Cas9 cells, the recruitment of CYLD was reduced corresponding to the reduction of SPATA2. This is now shown in Figure 3A. 3. Fig. 3A: Why is in the wt settings no M1-ubiquitination detected after 20 min TNF-stimulation? Other groups showed also in MEF cells strongly increased M1- ubiquitination signals after 20 min TNF stimulation at the TNF-RSC. Is it cell type specific difference? We think this discrepancy may be due to cell type-specific effects, or possibly due TNF used by others from different manufacturers (or different lots) with slightly different activity. 4. Authors should include anti-SPATA2 staining in experiments, when using SPATA2 KO cells, to prove that SPATA2 was indeed depleted in the particular setting (Fig. 2E, 3A, 4A-C). Unfortunately the anti SPATA2 antibodies we could obtain did not pick up mouse SPATA2 very well. However, using human A549 cells, we can clearly show the reduction of SPATA2 in mixed cell cultures expressing CRISPR/Cas9 targeting the spata2 gene (however containing a fraction of cells retaining SPATA2), and the strong reduction of SPATA2 in the respective TNFR complex I pull-downs from these cells. This is shown in new Fig. 3A. We were able to pick up SPATA2 with a SPATA2 antibody with the MEF we describe in Fig. 3B and Fig. 5A. This is now shown in Fig. EV1. For reasons we cannot explain we could not pick up a signal for endogenous SPATA2 with 15P-1 Sertoli cells. However we have generated single clones now, and by sequencing, we could pick clones with a frameshift on both alleles, resulting in an immature STOP-codon and therefore cannot express SPATA2 protein. One of these clones is shown in Fig. 5B. The sequencing of the respective clone is shown in Fig. EV3. As with reviewer #1 and #2, we thank the reviewer for his/her efforts in evaluating our manuscript, and we feel that the additional experiments we performed and the changes we have made according to his/her input have substantially improved the work. We hope you will agree. 2nd Editorial Decision 02 July 2016
Thank you for the submission of your revised manuscript to our editorial offices. We have now received the enclosed reports on it. As you will see, all three referees find the revised manuscript suitable for publication in EMBO reports. Before we can proceed with the formal acceptance of your manuscript we will require that you upload as soon as possible: - a Microsoft Word file (.doc) of the final manuscript text - editable TIFF or EPS-formatted figure files in high resolution (that need then to be uploaded as single files) - single files for the tables in the methods section and call outs for these in the text (Table 1, Table 2). - a conflict of interest statement in the manuscript text after the author contribution part. Further, for a short report we would require that the results part and discussion part are combined in

EMBO reports - Peer Review Process File - EMBO-2016-42592
© European Molecular Biology Organization 14
one section (called Results and Discussion). We also strongly encourage the publication of original source data with the aim of making primary data more accessible and transparent to the reader. The source data will be published in a separate source data file online along with the accepted manuscript and will be linked to the relevant figure. If you would like to use this opportunity, please submit the source data (for example scans of entire gels or blots, data points of graphs in an excel sheet, additional images, etc.) of your key experiments together with the revised manuscript. Please include size markers for scans of entire gels, label the scans with figure and panel number, and send one PDF file per figure or per figure panel. Please contact me any time if you have any questions. REFEREE REPORTS Referee #1: The authors have done a good job in addressing all my concerns, and the ms is much improved. In light of the timely nature of this story I recommend publication of the revised ms without any further delay. Referee #2: The authors have done a really excellent job in addressing the reviewers comments, perhaps one of the strongest I have seen in a short time. They have improved the analysis of complex I signalling substantially and they have provided an answer for why tak1 ko cells are protected by loss of SPATA2. Referee #3: The authors have address major questions successfully. The manuscript is now acceptable for publication. 2nd Revision - authors' response 04 July 2016
Author made the necessary changes. 3rd Editorial Decision 05 July 2016
I am very pleased to accept your manuscript for publication in the next available issue of EMBO reports. Thank you for your contribution to our journal.

USEFUL LINKS FOR COMPLETING THIS FORM
http://www.antibodypedia.comhttp://1degreebio.orghttp://www.equator-‐network.org/reporting-‐guidelines/improving-‐bioscience-‐research-‐reporting-‐the-‐arrive-‐guidelines-‐for-‐reporting-‐animal-‐research/
http://grants.nih.gov/grants/olaw/olaw.htmhttp://www.mrc.ac.uk/Ourresearch/Ethicsresearchguidance/Useofanimals/index.htmhttp://ClinicalTrials.govhttp://www.consort-‐statement.orghttp://www.consort-‐statement.org/checklists/view/32-‐consort/66-‐title
è
http://www.equator-‐network.org/reporting-‐guidelines/reporting-‐recommendations-‐for-‐tumour-‐marker-‐prognostic-‐studies-‐remark/è
http://datadryad.orgè
http://figshare.comè
http://www.ncbi.nlm.nih.gov/gapè
http://www.ebi.ac.uk/ega
http://biomodels.net/
http://biomodels.net/miriam/è http://jjj.biochem.sun.ac.zaè http://oba.od.nih.gov/biosecurity/biosecurity_documents.htmlè http://www.selectagents.gov/è
èè
èè
� common tests, such as t-‐test (please specify whether paired vs. unpaired), simple χ2 tests, Wilcoxon and Mann-‐Whitney tests, can be unambiguously identified by name only, but more complex techniques should be described in the methods section;
� are tests one-‐sided or two-‐sided?� are there adjustments for multiple comparisons?� exact statistical test results, e.g., P values = x but not P values < x;� definition of ‘center values’ as median or average;� definition of error bars as s.d. or s.e.m.
1.a. How was the sample size chosen to ensure adequate power to detect a pre-‐specified effect size?
1.b. For animal studies, include a statement about sample size estimate even if no statistical methods were used.
2. Describe inclusion/exclusion criteria if samples or animals were excluded from the analysis. Were the criteria pre-‐established?
3. Were any steps taken to minimize the effects of subjective bias when allocating animals/samples to treatment (e.g. randomization procedure)? If yes, please describe.
For animal studies, include a statement about randomization even if no randomization was used.
4.a. Were any steps taken to minimize the effects of subjective bias during group allocation or/and when assessing results (e.g. blinding of the investigator)? If yes please describe.
4.b. For animal studies, include a statement about blinding even if no blinding was done
5. For every figure, are statistical tests justified as appropriate?
Do the data meet the assumptions of the tests (e.g., normal distribution)? Describe any methods used to assess it.
Is there an estimate of variation within each group of data?
Is the variance similar between the groups that are being statistically compared?
no statistical testing was performed
does not apply
does not apply
does not apply
YOU MUST COMPLETE ALL CELLS WITH A PINK BACKGROUND ê
does not apply
no animal studies were performed
no exclusion from analysis
no
no animal studies were performed
no exclusion from analysis
no animal studies were performed
definitions of statistical methods and measures:
1. Data
the data were obtained and processed according to the field’s best practice and are presented to reflect the results of the experiments in an accurate and unbiased manner.figure panels include only data points, measurements or observations that can be compared to each other in a scientifically meaningful way.graphs include clearly labeled error bars for independent experiments and sample sizes. Unless justified, error bars should not be shown for technical replicates.if n< 5, the individual data points from each experiment should be plotted and any statistical test employed should be justified
Please fill out these boxes ê (Do not worry if you cannot see all your text once you press return)
a specification of the experimental system investigated (eg cell line, species name).
C-‐ Reagents
B-‐ Statistics and general methods
the assay(s) and method(s) used to carry out the reported observations and measurements an explicit mention of the biological and chemical entity(ies) that are being measured.an explicit mention of the biological and chemical entity(ies) that are altered/varied/perturbed in a controlled manner.
the exact sample size (n) for each experimental group/condition, given as a number, not a range;a description of the sample collection allowing the reader to understand whether the samples represent technical or biological replicates (including how many animals, litters, cultures, etc.).
Each figure caption should contain the following information, for each panel where they are relevant:
2. Captions
The data shown in figures should satisfy the following conditions:
Source Data should be included to report the data underlying graphs. Please follow the guidelines set out in the author ship guidelines on Data Presentation.
a statement of how many times the experiment shown was independently replicated in the laboratory.
Any descriptions too long for the figure legend should be included in the methods section and/or with the source data.
Please ensure that the answers to the following questions are reported in the manuscript itself. We encourage you to include a specific subsection in the methods section for statistics, reagents, animal models and human subjects.
In the pink boxes below, provide the page number(s) of the manuscript draft or figure legend(s) where the information can be located. Every question should be answered. If the question is not relevant to your research, please write NA (non applicable).
Manuscript Number: EMBOR-‐2016-‐42592V1
EMBO PRESS
A-‐ Figures
Reporting Checklist For Life Sciences Articles (Rev. July 2015)
This checklist is used to ensure good reporting standards and to improve the reproducibility of published results. These guidelines are consistent with the Principles and Guidelines for Reporting Preclinical Research issued by the NIH in 2014. Please follow the journal’s authorship guidelines in preparing your manuscript.
PLEASE NOTE THAT THIS CHECKLIST WILL BE PUBLISHED ALONGSIDE YOUR PAPER
Journal Submitted to: EMBO ReportsCorresponding Author Name: Ulrich Maurer

6. To show that antibodies were profiled for use in the system under study (assay and species), provide a citation, catalog number and/or clone number, supplementary information or reference to an antibody validation profile. e.g., Antibodypedia (see link list at top right), 1DegreeBio (see link list at top right).
7. Identify the source of cell lines and report if they were recently authenticated (e.g., by STR profiling) and tested for mycoplasma contamination.
* for all hyperlinks, please see the table at the top right of the document
8. Report species, strain, gender, age of animals and genetic modification status where applicable. Please detail housing and husbandry conditions and the source of animals.
9. For experiments involving live vertebrates, include a statement of compliance with ethical regulations and identify the committee(s) approving the experiments.
10. We recommend consulting the ARRIVE guidelines (see link list at top right) (PLoS Biol. 8(6), e1000412, 2010) to ensure that other relevant aspects of animal studies are adequately reported. See author guidelines, under ‘Reporting Guidelines’. See also: NIH (see link list at top right) and MRC (see link list at top right) recommendations. Please confirm compliance.
11. Identify the committee(s) approving the study protocol.
12. Include a statement confirming that informed consent was obtained from all subjects and that the experiments conformed to the principles set out in the WMA Declaration of Helsinki and the Department of Health and Human Services Belmont Report.
13. For publication of patient photos, include a statement confirming that consent to publish was obtained.
14. Report any restrictions on the availability (and/or on the use) of human data or samples.
15. Report the clinical trial registration number (at ClinicalTrials.gov or equivalent), where applicable.
16. For phase II and III randomized controlled trials, please refer to the CONSORT flow diagram (see link list at top right) and submit the CONSORT checklist (see link list at top right) with your submission. See author guidelines, under ‘Reporting Guidelines’. Please confirm you have submitted this list.
17. For tumor marker prognostic studies, we recommend that you follow the REMARK reporting guidelines (see link list at top right). See author guidelines, under ‘Reporting Guidelines’. Please confirm you have followed these guidelines.
18. Provide accession codes for deposited data. See author guidelines, under ‘Data Deposition’.
Data deposition in a public repository is mandatory for:a. Protein, DNA and RNA sequencesb. Macromolecular structuresc. Crystallographic data for small moleculesd. Functional genomics data e. Proteomics and molecular interactions19. Deposition is strongly recommended for any datasets that are central and integral to the study; please consider the journal’s data policy. If no structured public repository exists for a given data type, we encourage the provision of datasets in the manuscript as a Supplementary Document (see author guidelines under ‘Expanded View’ or in unstructured repositories such as Dryad (see link list at top right) or Figshare (see link list at top right).20. Access to human clinical and genomic datasets should be provided with as few restrictions as possible while respecting ethical obligations to the patients and relevant medical and legal issues. If practically possible and compatible with the individual consent agreement used in the study, such data should be deposited in one of the major public access-‐controlled repositories such as dbGAP (see link list at top right) or EGA (see link list at top right).21. As far as possible, primary and referenced data should be formally cited in a Data Availability section. Please state whether you have included this section.
Examples:Primary DataWetmore KM, Deutschbauer AM, Price MN, Arkin AP (2012). Comparison of gene expression and mutant fitness in Shewanella oneidensis MR-‐1. Gene Expression Omnibus GSE39462Referenced DataHuang J, Brown AF, Lei M (2012). Crystal structure of the TRBD domain of TERT and the CR4/5 of TR. Protein Data Bank 4O26AP-‐MS analysis of human histone deacetylase interactions in CEM-‐T cells (2013). PRIDE PXD00020822. Computational models that are central and integral to a study should be shared without restrictions and provided in a machine-‐readable form. The relevant accession numbers or links should be provided. When possible, standardized format (SBML, CellML) should be used instead of scripts (e.g. MATLAB). Authors are strongly encouraged to follow the MIRIAM guidelines (see link list at top right) and deposit their model in a public database such as Biomodels (see link list at top right) or JWS Online (see link list at top right). If computer source code is provided with the paper, it should be deposited in a public repository or included in supplementary information.
23. Could your study fall under dual use research restrictions? Please check biosecurity documents (see link list at top right) and list of select agents and toxins (APHIS/CDC) (see link list at top right). According to our biosecurity guidelines, provide a statement only if it could.
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
does not apply
see table in experimental procedures pages 17, 18
see in experimental procedure, page 15
does not apply
does not apply
does not apply
G-‐ Dual use research of concern
F-‐ Data Accessibility
D-‐ Animal Models
E-‐ Human Subjects