sowing date and the relationships between plant and apex development in winter cereals
TRANSCRIPT

Field Crops Research, 14 (1986) 321--337 321 Elsevier Science Publishers B.V., Amsterdam -- Printed in The Netherlands
SOWING DATE AND THE RELATIONSHIPS BETWEEN PLANT AND APEX DEVELOPMENT IN WINTER CEREALS
R.K.M. HAY
Botany Department, West o f Scotland Agricultural College, Auchincruive, Ayr, KA6 5HW (Great Britain)
(Accepted 22 April 1986)
ABSTRACT
Hay, R.K.M., 1986. Sowing date and the relationships between plant and apex develop- ment in winter cereals. Field Crops Res., 14: 321--337.
The plant and mainstem apex development of a range of sowings of winter wheat, barley and rye (early September to late April) was monitored at weekly intervals at a single site in the West o f Scotland throughout the growing season 1982--1983. For each species there was no relationship between mainstem leaf number and the stage of apex development. However, in each case, three developmental events -- the terminal spikelet (or awn primordium) stage, the beginning of mainstem extension and the death of later- formed tillers -- were clearly associated. In wheat and rye, stem internode extension be- gan shortly before the terminal spikelet or awn primordium stage, but the interval be- tween the two events was progressively reduced by delay in sowing such that they were virtually synchronous for the spring sowings. In contrast, the awn primordium stage in barley occurred before the onset of stem extension. The start of tiller death, which is difficult to record objectively, appeared to occur at approximately the same time as stem internode extension in each species.
For each sowing of the three species the length of the sheath o f malnstem leaves in- creased exponentially with leaf number; furthermore, the sheath length of a given leaf increased progressively with delay in sowing so that successive leaves had further to ex- tend before they reached the ligule of the previous leaf and were classed as having appear- ed. The implications of this effect for the rate of leaf appearance are discussed. The crop measurements also provide further evidence for a relationship between pseudostem height and the achievement of the double ridge stage not only in wheat but also in rye and barley.
Overall, the findings for each species confirm the usefulness of field measurements of pseudostem height and "ear at 1 cm" as reliable guides to the development of the main- stem apex in wheat, but much more field evidence is required for winter barley.
INTRODUCTION
Over the last decade, considerable advances have been made in the under- standing of cereal crop growth and yield by field studies which have com- bined physiological measurements with detailed monitoring of plant devel- opment. This approach has led to the formulation of simulation models
0378-4290/86/$03.50 © 1986 Elsevier Science Publishers B.V.

322
which can be used to predict plant development and crop phenology with some degree of accuracy from a relatively simple set o f environmental data, principally temperature and daylength {e.g., Porter, 1984; Weir et al., 1984). It has also led to an appreciation o f the uncertainties in our knowledge.
For example, three important events in crop development -- (1) achieve- ment of terminal spikelet stage (wheat) or maximum number of primordia stage (barley, rye) on the main shoot (Kirby and Appleyard, 1984a), (2) the beginning of the rapid upward movement of the mainstem apex (stem inter- node extension), and (3) the rapid decline in number of living tillers -- ap- pear to be closely linked (e.g., in wheat, Baker and Gallagher, 1983), but there is a lack of information as to how the timing o f these interrelated developmental events varies amongst species and sowing dates. This question is not only of scientific interest but also of considerable practical impor- tance; recent work (Darwinkel, 1983) has shown that precise timing of ni- trogen fertilization at the onset of stem elongation can give increases in the number of fertile tillers per plant. Before such a practice can be considered for general adoption (Biscoe and Willington, 1984; Appleton and Haggar, 1985) it is necessary to know more about tiller and stem apex development under a range o f field conditions.
There is a second important uncertainty involving the development of leaf sheaths. Very close linear relationships have been established between leaf appearance (based on counts of the number of leaves per mainstem whose tip or ligule has become visible) and accumulated soil or air temperature, at least during the period before stem extension (Baker et al., 1980;Del~colle and Gurnade, 1980; Hay and Tunnicliffe Wilson, 1982). However, because the distance from the soil surface to the ligule of the youngest fully-expand- ed leaf (pseudostem length) increases with leaf number (Tottman, 1977), successive leaves must extend further before they can be seen and classed as "appeared". Two important questions arise out of this observation. Firstly, does this mean that successive leaves extend more rapidly in order to emerge on time according to a linear accumulated temperature/ leaf appearance re- lationship, and, secondly, does the sheath length of a given leaf vary signifi- cantly between sowing dates?
This paper, which is a continuation of work reported by Hay (1978), examines these areas of uncertainty by studying the plant and stem apex development of winter wheat, rye and barley sown over a wide range of dates from early September to late April.
MATERIALS AND METHODS
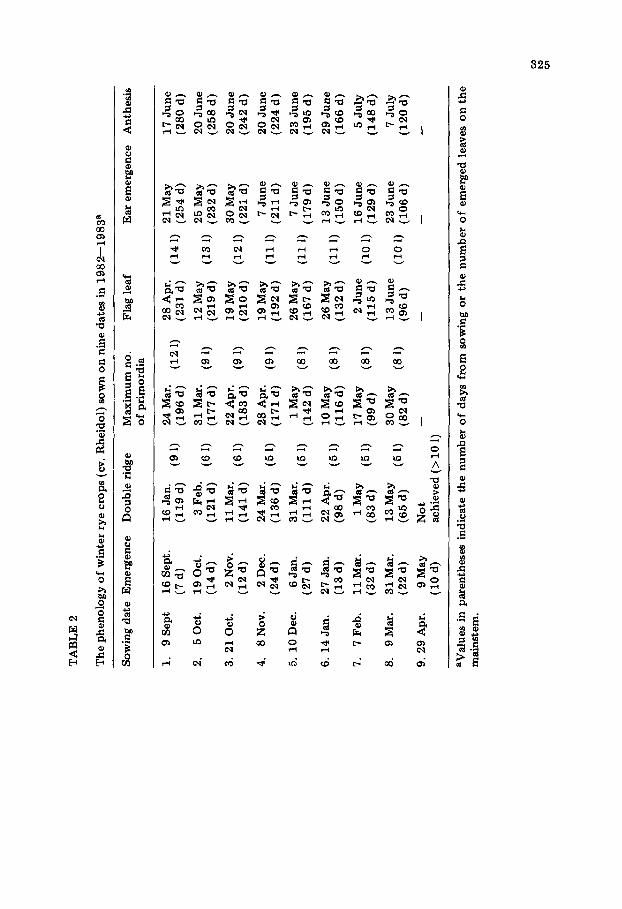
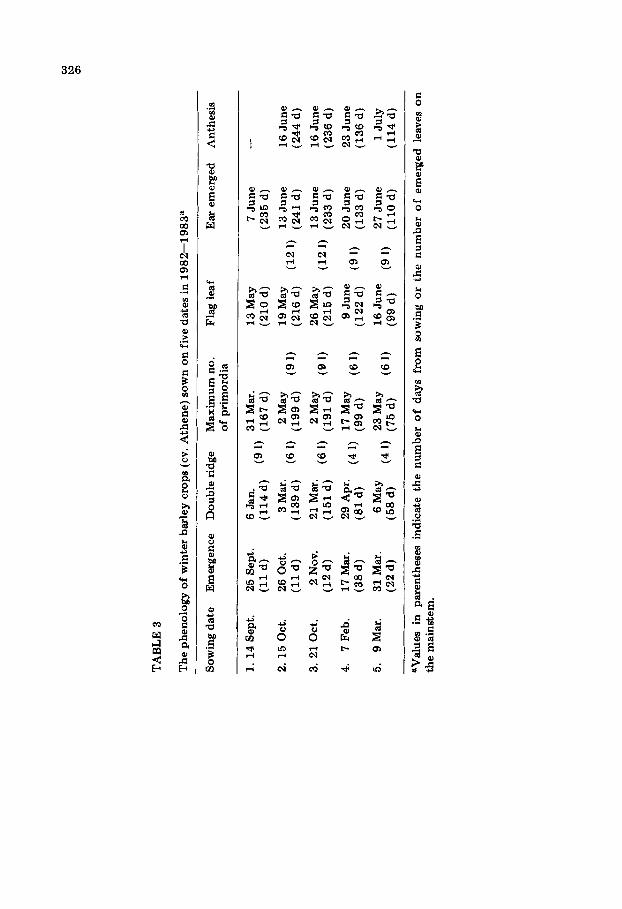
The measurements presented in this paper were obtained from a series of 1-m 2 hand-sown plots (15 cm between rows) of winter wheat (cv. Maris Hustler) and winter rye (cv. Rheidol) sown at Auchincruive on nine sowing dates during the season 1982--1983 (Tables 1 and 2). These results are sup- plemented by measurements of an early-sown commercial c rop and four

323
hand sowings of winter barley (cv. Athene) (Table 3). Each plot was given 20 kg P, 40 kg K and 20 kg N/ha in the seedbed and, on 17 March, the first seven wheat and rye sowings, and the first three barley sowings were given a further 50 kg N/ha. A single dressing of 70 kg N/ha was given to the re- maining plots at sowing. The timing of these nitrogen treatments was design- ed to reduce the risk of nitrogen deficiency at terminal spikelet/maximum number of primordia and stem internode extension.
At emergence, 25 uniform plants from each sowing, each plant separated from adjacent plants in the row by 2--3 cm, were marked using 1-cm diam- eter loops of coloured wire. These spacings correspond to plant population densities of 225--350 plants/m 2. Thereafter, mainstem leaf appearance and emerged, living, tiller numbers were recorded at 7-day intervals as described in Hay and Tunnicliffe Wilson (1982). As in the majority of previous studies, a tiller was classed as dead when the youngest leaf began to yellow. From December onwards, destructive harvests were made (five plants/plot) and the plants were dissected following the procedures of Kirby and Appleyard (1984a) to determine the developmental stage, length, and position relative to the soil surface, of the mainstem apex, as well as the sheath length of the youngest fully-extended leaf (soil surface to ligule). These measurements were continued up to ear emergence and ceased at anthesis.
The successive sowings of each species were made under deteriorating soil and weather conditions with the result that plant establishment was general- ly poorer for sowings 5--7, in spite of increased seed rates. This made selec- tion of suitably-spaced plants more difficult for these sowings and, in the case of sowing 6, where there was also predation by mice, the measurements are of widely-spaced plants. This may have had a significant influence on the rate of development of the plants of this sowing (e.g., Kirby and Faris, 1972) and it presumably accounts for the higher number of surviving tillers of the wheat crop (Fig. 2). However, in the present paper it is assumed that sowing date is the predominant factor influencing plant development, at least up to canopy closure. Disease and weed incidences were negligible, and although many of the stands suffered considerable leaf damage due to frost in Janu- ary, there was little evidence of winterkill amongst the marked plants.
RESULTS
Crop phenology and stem apex development
The development of successive sowings of each of the three species was found to converge so that, for example, the eight wheat crops sown between 9 September and 9 March all reached anthesis within a period of only 17 days (Table 1); similar effects were observed for rye (Table 2) and barley (Table 3). This phenomenon is caused mainly by the progressive acceleration of the development of later-sown crops (Stern and Kirby, 1979; Baker et al., 1980; Hay and Tunnicliffe Wilson, 1982) mediated by photoperiod and by

TA
BL
E 1
The
phe
nolo
gy o
f w
inte
r w
heat
cro
ps (
cv. M
aris
Hus
tler
) so
wn
on n
ine
date
s in
198
2--1
983
a
O~
tO
Sow
ing
date
E
mer
genc
e D
oubl
e ri
dge
Ter
min
al s
pike
let
Fla
g le
af
Ear
em
erge
nce
Ant
hesi
s
1.
9 S
ept.
21
Sep
t.
17 F
eb.
22 A
pr.
30 M
ay
20 J
une
29 J
une
(12
d)
(161
d)
(9 1
) (2
25 d
) (1
1 1)
(2
63 d
) (1
4 l)
(2
84 d
) (2
93 d
)
2.
5 O
ct.
21 O
ct.
14 A
pr.
13 M
ay
30 M
ay
23 J
une
29 J
une
(16
d)
(191
d)
(9 1
) (2
20 d
) (1
1 1)
(2
37 d
) (1
3 1)
(2
61 d
) (2
67 d
)
3. 2
1 O
ct.
8 N
ov.
2 M
ay
21 M
ay
7 Ju
ne
27 J
une
5 Ju
ly
(18
d)
(193
d)
(9 l
) (2
12 d
) (1
1 1)
(2
29 d
) (1
3 1)
(2
49 d
) (2
57 d
)
4.
8 N
ov.
9 D
ec.
6 M
ay
21 M
ay
7 Ju
ne
27 J
une
5 Ju
ly
(31
d)
(179
d)
(8 1
) (1
94 d
) (9
1)
(211
d)
(11
1)
(231
d)
(239
d)
5.10
Dec
. 14
Jan
. 6
May
21
May
7
June
29
Jun
e 7
July
(3
5 d)
(1
47 d
) (7
1)
(162
d)
(8 1
) (1
79 d
) (1
0 1)
(2
01 d
) (2
09 d
)
6. 1
4 Ja
n.
3 M
ar.
6 M
ay
25 M
ay
17 J
une
3 Ju
ly
11 J
uly
(48
d)
(112
d)
(6 l
) (1
31 d
) (8
l)
(154
d)
(10
1)
(170
d)
(178
d)
7.
7 F
eb.
17 M
ar.
13 M
ay
25 M
ay
23 J
une
5 Ju
ly
13 J
uly
(38
d)
(95
d)
(6 1
) (1
07 d
) (8
1)
(136
d)
(10
1)
(148
d)
(156
d)
8.
9 M
ar.
7 A
pr.
23 M
ay
7 Ju
ne
1 Ju
ly
9 Ju
ly
15 J
uly
(29
d)
(75
d)
(6 1
) (9
0 d)
(8
1)
(114
d)
(10
1)
(122
d)
(128
d)
9. 2
9 A
pr.
12 M
ay
5 Ju
ly b
15 J
uly
b 21
Jul
y b
2 A
ug. b
16
Aug
. b
(13
d)
(37
d)
(10
1)
(47
d)
(12
1)
aVal
ues
in p
aren
thes
es i
ndic
ate
the
num
ber
of d
ays
from
sow
ing
or t
he n
umbe
r o
f em
erge
d le
aves
on
the
mai
nste
m.
bVer
y va
riab
le r
epro
duct
ive
deve
lopm
ent
(see
tex
t).
TA
BL
E 2
The
phe
nolo
gy o
f w
inte
r ry
e cr
ops
(cv.
Rhe
idol
) so
wn
on n
ine
date
s in
198
2--1
983
a
Sow
ing
date
E
mer
genc
e D
oubl
e ri
dge
Max
imum
no.
F
lag
leaf
E
ar e
mer
genc
e A
nthe
sis
of p
rim
ordi
a
1.
9 S
ept
16 S
ept.
16
Jan
. 24
Mar
. 28
Apr
. 21
May
17
Jun
e (7
d)
(119
d)
(9 I
) (1
96 d
) (1
2 I)
(2
31 d
) (1
4 l)
(2
54 d
) (2
80 d
)
2.
5 O
ct.
19 O
ct.
3 F
eb.
31 M
ar.
12 M
ay
25 M
ay
20 J
une
(14
d)
(121
d)
(6 1
) (1
77 d
) (9
l)
(219
d)
(13
l)
(232
d)
(258
d)
3.21
Oct
. 2
Nov
. 11
Mar
. 22
Apr
. 19
May
30
May
20
Jun
e (1
2 d)
(1
41 d
) (6
l)
(183
d)
(9 1
) (2
10 d
) (1
2 1)
(2
21 d
) (2
42 d
)
4.
8 N
ov.
2 D
ec.
24 M
ar.
28 A
pr.
19 M
ay
7 Ju
ne
20 J
une
(24
d)
(136
d)
(5 l
) (1
71 d
) (9
l)
(192
d)
(11
l)
(211
d)
(224
d)
5. 1
0 D
ec.
6 Ja
n.
31 M
ar.
1 M
ay
26 M
ay
7 Ju
ne
23 J
une
(27
d)
(lll
d)
(51)
(1
42
d)
(81)
(1
67
d)
(lll
) (1
79
d)
(19
5d
)
6. 1
4 Ja
n.
27 J
an.
22 A
pr.
10 M
ay
26 M
ay
13 J
une
29 J
une
(13
d)
(98
d)
(5 1
) (1
16 d
) (8
l)
(132
d)
(11
l)
(150
d)
(166
d)
7.
7 F
eb.
11 M
ar.
1 M
ay
17 M
ay
2 Ju
ne
16 J
une
5 Ju
ly
(32
d)
(83
d)
(5 1
) (9
9 d)
(8
1)
(115
d)
(10
1)
(129
d)
(148
d)
8.
9 M
ar.
31 M
ar.
13 M
ay
30 M
ay
13 J
une
23 J
une
7 Ju
ly
(22
d)
(65
d)
(5 1
) (8
2 d)
(8
l)
(96
d)
(10
l)
(106
d)
(120
d)
9.2
9 A
pr.
9 M
ay
Not
.
..
.
(10
d)
achi
eved
(>
10 l
)
aVal
ues
in p
aren
thes
es i
ndic
ate
the
num
ber
of d
ays
from
sow
ing
or t
he n
umbe
r o
f em
erge
d le
aves
on
the
mai
nste
m.
C~
¢j1
CO
t~
TA
BL
E 3
The
phe
nolo
gy o
f w
inte
r ba
rley
cro
ps (
cv.
Ath
ene)
sow
n on
fiv
e da
tes
in 1
982-
-198
3 a
Sow
ing
date
E
mer
genc
e D
oubl
e ri
dge
Max
imum
no.
F
lag
leaf
E
ar e
mer
ged
Ant
hesi
s o
f pr
imor
dia
1.1
4 S
ept.
25
Sep
t.
6 Ja
n.
31 M
ar.
13 M
ay
7 Ju
ne
--
(11
d)
(114
d)
(9 1
) (1
67 d
) (2
10 d
) (2
35 d
)
2. 1
5 O
ct.
26 O
ct.
3 M
ar.
2 M
ay
19 M
ay
13 J
un
e 16
Ju
ne
(11
d)
(139
d)
(6 1
) (1
99 d
) (9
1)
(216
d)
(12
l)
(241
d)
(244
d)
3. 2
1 O
ct.
2 N
ov.
21 M
ar.
2 M
ay
26 M
ay
13 J
un
e 16
Jun
e (1
2 d)
(1
51 d
) (6
1)
(191
d)
(9 l
) (2
15 d
) (1
2 1)
(2
33 d
) (2
36 d
)
4.
7 F
eb.
17 M
ar.
29 A
pr.
17 M
ay
9 Ju
ne
20 J
un
e 23
Jun
e (3
8 d)
(8
1 d)
(4
l)
(99
d)
(6 l
) (1
22 d
) (9
l)
(133
d)
(136
d)
5.
9 M
ar.
31 M
ar.
6 M
ay
23 M
ay
16 J
une
27 J
une
1 Ju
ly
(22
d)
(58
d)
(4 1
) (7
5 d)
(6
1)
(99
d)
(9 l
) (1
10 d
) (1
14 d
)
aVal
ues
in parentheses
indicate the
number of da
ys from sowing or
the
number of e
merged leaves on
the ma
inst
em.

327
low-temperature vernalization. Thus, for wheat sowings 3 (21 October) to 6 (14 January), vernalization was complete in time for the plants of each sowing to respond similarly to photoperiod, achieving double ridge within a period of 5 days of each other (Table 1).
Tables 1--3 reveal several other features of the development of winter cereals. Firstly, there were clear differences between the cultivars of wheat and rye used in this study in their requirement for low-temperature vernal- ization (Tables 1 and 2; Purvis, 1961). Thus, all of the rye plants sown in late April remained vegetative, whereas in the corresponding wheat sowing the vegetative phase of development was longer than for the previous sowing, but 90% of the plants did eventually produce normal ears about a month later than the previous sowings. Secondly, although the acceleration of the development of the later-sown plants was, to a certain extent, at the expense of leaf production, there was evidence of a minimum leaf number for each species (10 for wheat and rye, nine for barley) under normal combinations of daylength and temperature at the experimental site. Further reduction of final leaf number could have been achieved by other combinations of tem- peratures and daylengths (Kirby et al., 1985a). Thirdly, in no case was there evidence of a simple relationship between mainstem leaf number and apex development; for example, the mainstems of rye plants of different sowing dates carried between five and nine leaves at the double ridge stage (Table 2).
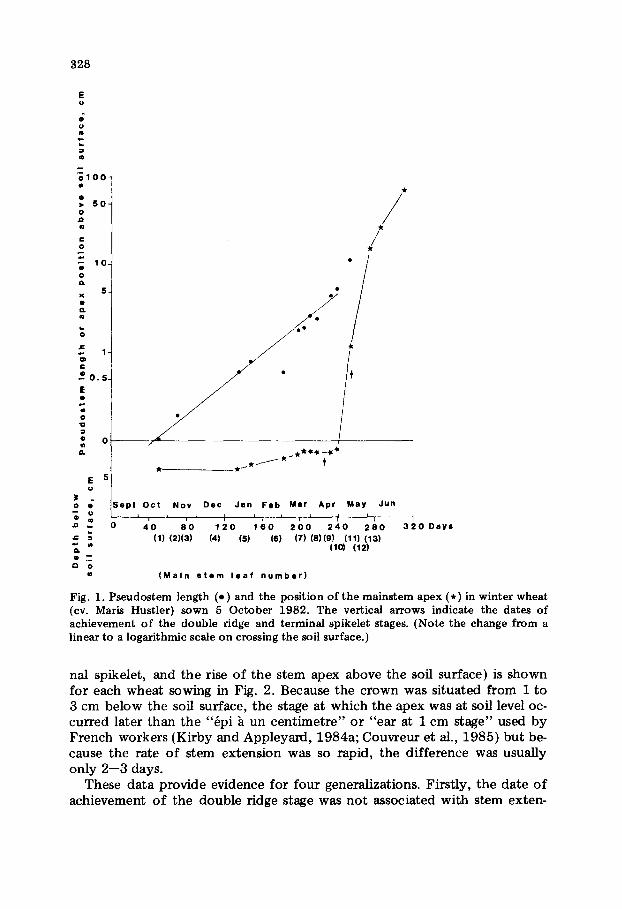
The time course of plant development varied significantly among species and among sowing dates, but the general features of mainstem development can be illustrated by the measurements from the second wheat sowing (Fig. 1). The mainstem apex remained below the soil surface until the beginning of rapid stem internode extension, although, earlier, there was an apparent upward movement of several mm, presumably caused by soil consolidation and/or the accumulation of stem nodes (see also Hay, 1978). The double ridge stage was achieved while the apex was still below the soil surface but, for this sowing at least, terminal spikelet did not occur until the apex was ap- proximately 1 cm above the soil, and the major phase of ear growth (i.e., a 25-fold increase in length) took place between this time and ear emergence. Sheath length increased continuously between successive leaves, from leaf I onwards, and exponentially with time (Fig. 1). Consequently, the pseudo- stem length was less than 1 cm long for leaves up to the appearance of leaf 5 but by the time that leaf 8 was fully expanded, it had increased to 5 cm (Zadoks code 30, Tottman et al., 1979). Thus leaves appearing before leaf 6 had to extend between 0.3 and 1 cm above the soil surface before the tip be- came visible, whereas the corresponding distances for leaves 6--10 were be- tween 1 and 6 cm.
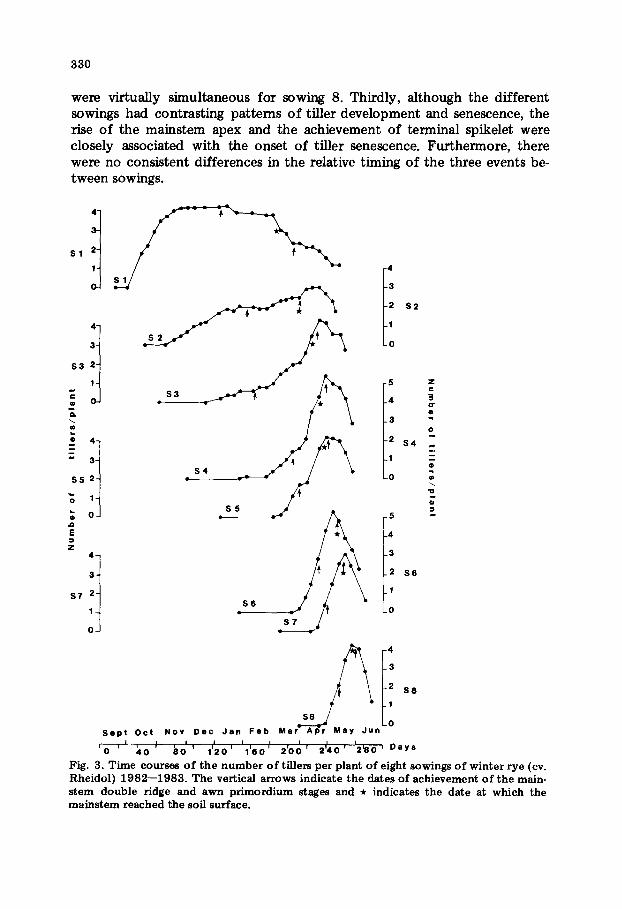
Mainstem apex development and the time course o f tiller numbers
The relationship between the number of emerged living tillers per plant and the timing of three mainstem development events (double ridge, termi-
328
E u
@
m
o l 0 0 - w
e > 5 0 - o .o
" 1 0 - o 0 Q.
5 - x @ D.
c
_°o.s_
o
e o "o
• 0 w 0.
/ /
e •
/, /
~ - ~
E 5
o o" S e p t O c t Nov Dec Jan Feb Mar Apr May Jun
® ~ ' 8 ' 0 I ~ ' ~ ' I - . ~ 0 4 0 1 2 0 1 6 0 2 0 0 2 4 0 2 8 0 3 0 D S y 8
J= = (1 ) ( 2 ) ( 3 ) (4) (5 ) (6 ) ( 7 ) ( 8 ) ( 9 ) (11) ( 1 3 ) ~ (10) (12 )
• - -
o ( M a i n s t e m l e a f n u m b e r )
Fig. 1. Pseudostem length (e) and the position of the mainstem apex (*) in winter wheat (cv. Marls Hustler) sown 5 October 1982. The vertical arrows indicate the dates o f achievement o f the double ridge and terminal spikelet stages. (Note the change from a linear to a logarithmic scale on crossing the soil surface.)
nal spikelet, and the rise of the stem apex above the soil surface) is shown for each wheat sowing in Fig. 2. Because the crown was situated from 1 to 3 cm below the soil surface, the stage at which the apex was at soft level oc- curred later than the "~pi h u n cent imetre" or "ear at 1 cm stage" used by French workers (Kirby and Appleyard, 1984a; Couvreur et al., 1985) but be- cause the rate of stem extension was so rapid, the difference was usually only 2--3 days.
These data provide evidence for four generalizations. Firstly, the date of achievement of the double ridge stage was not associated with stem exten-

329
s31 c ¢1 o. 5
9)
-- 3 S5 2
1- o 0 .Q
E
"1 S7
$2
S3
S4
$5
°
L
-$6 Y
$2
4
3
2 S 4
1
0
6
5
4
3
2 S6
1
0
$7 .~
3
2 S8 1
_ 0 S e p t O c t N o v D e c J a n F e b M a r A p r M a y J u n
i i / J i I r ~ r I i I I , iJ i I r f
0 4 0 8 0 1 2 0 1 6 0 2 fO0 2 4 0 2 8 0 D a y s
Z ¢
3 o
o
e
t¢
fl}
Fig. 2. T ime courses o f the number o f tillers per plant o f eight sowings o f winter w h e a t (cv. Maria Hustler) 1 9 8 2 - - 1 9 8 3 . the vertical arrows indicate the dates o f achievement o f the mains tem double ridge and terminal spikelet stages and * indicates the date at which the mains tem reached the soil surface.
sion or tiller development, and it occurred at a range of mainstem leaf num- bers (Table 1). Secondly, in all cases, the rapid extension of stem internodes began before terminal spikelet but, because o f the tendency towards syn- chronous development in the later sowings, the interval between the rise of the apex above the soft surface and the achievement of terminal spikelet de- creased from 15 days (sowing 1) to a day (sowing 7), and the two events
330
were virtually simultaneous for sowing 8. Thirdly, although the different sowings had contrasting patterns o f tiller development and senescence, the :rise of the mainstem apex and the achievement of terminal spikelet were closely associated with the onset of tiller senescence. Furthermore, there were no consistent differences in the relative timing of the three events be- tween sowings.
: 3 S I
D,
5
0..1
O . J .
$2
0
• 5 Z
o" e
o
.2 S4
.1 "- e
- O t~
"O
0~
.4
_3
.2 $6
.1
.0
A L ~ L0 Dec Jan Feb Mar- Ap-r May Jun S e p t O c t Nov
' ~ ~ o '~ ' ~ ' ' ' 2 ~ o ' O ° , . ' 0 ~ 1 0 i I r O ' 1~60 2 0 0 2 ' 4 0 '
Fig. 3. T ime courses o f t h e n u m b e r o f t i l lers pe r p lan t o f e ight sowings o f w i n t e r rye (cv. Rhe ido l ) 1 9 8 2 - - 1 9 8 3 . T he ver t ical a r rows ind ica te t h e da tes of a ch i evemen t o f t he main- s t em d o u b l e r idge and awn p r i m o r d i u m stages and * ind ica tes t h e da te at wh ich the m a i n s t e m r eached t he soil surface.

331
Finally, it is commonly assumed that a given tiller will appear when its subtending leaf and two subsequent leaves are fully expanded (Kirby and Appleyard, 1984b). Thus it would be expected that the first tiller forming within the sheath of leaf I would appear simultaneously with the tip of leaf 4. This was the case for sowings 1, 2 and 6--9, but under the low tempera- tures of January and February, the rate of tiller emergence was depressed more than the rate of leaf appearance, with the result that, for sowing 3, the first tiller to emerge did not do so until leaf 7 was expanding. However, it is not known whether this was the result of a delay in emergence, or the complete inhibition, o f tillers forming at the first two or three nodes. It is possible that the first tiller to emerge was developing within the sheath of leaf 3 or leaf 4.
Although the development of the rye plants was more rapid than that of the corresponding wheat sowings (Fig. 3; Hay and Abbas A1-Ani, 1983), the pattern o f response of the two species was very similar, such that the pre- ceding generalizations apply equally well to rye. However, for the barley sowings {Fig. 4), there was one very important difference: the maximum number of primordia (awn primordium stage; Kirby and Appleyard, 1984a)
==
w 2
@
; ' S 4 1
o
s3 y 3 e
e s4 "--
m 2 =
f" ~ 1 $5
0
S e p t h O c t N o v D e c J a n F e b M a r A p r M a y J u n e , i i I t I , l i i r i ,
( ) 4 0 8 0 1 2 0 1 6 0 2 0 0 2 4 0 2 8 0 3 2 0
D a y s
Fig. 4. Time courses o f the number o f tillers per plant o f three sowings o f winter barley (cv. Athene) 1982--1983. The vertical arrows indicate the dates o f achievement o f the mainstem double ridge and awn primordium stages and * indicates the date at which the mainstem reached the soil surface.

332
was achieved 15 (sowing 3), 4 (sowing 4) or 2 (sowing 5) days before rather than after the first appearance o f the apex above the soil surface (Fig. 4). Thus although the close relationship between the three events was maintain- ed in winter barley, the "ear at 1 cm" stage was an index of a later stage o f apex development than for wheat or rye.
Mainstem apex development and the development of leaf sheaths
Plotting the length o f the sheath o f the youngest fully-emerged leaf against mainstem leaf number revealed that sheath length increased exponen- tially with leaf number, but that the exponent differed between sowings. The difference in sheath length between sowings, therefore, increased with leaf number (Fig. 5). Thus the ligule o f leaf 4 was approximately 1 cm above the soil surface for wheat sowings 1--4 and 2.5 cm for sowing 7, whereas the corresponding heights for leaf 7 were 2 cm {sowing 2), 5 cm (4) and 9.5 cm (7). Very similar trends were observed in rye and barley (Fig. 5). These find- ings confirm the observation that successive leaves must extend further be-
16 W h e a t Bar ley
14
cJ
12
w
o 1 0 " o
0 ,a 8 "
• 6
E o w 0 4 "o
o
~ 2 "
1 2 3 4 5 6 7 8 9 10 11 1 2 3 4
Leaf number
5 6 7 8 9 10 11
Fig. 5. The influence o f leaf number and sowing date on pseudostem length in winter wheat (cv. Maris Hustler, sowings 2, 4 and 7) and winter barley (cv. Athene, sowings 3 and 5), 1982--1983. The curves were fitted by linear regression using log e (pseudoatem length); r = 0.992 (wheat 2), 0 .999 (wheat 4), 0.981 (wheat 7), 0.975 (barley 3), 0.979 (barley 5).

333
fore they can be classed as "appeared" but also show that the distance which the tip of a given leaf must travel to the ligule of the previous leaf increases with delay in sowing.
As a result of this interrelationship between leaf sheath extension and sowing date, growth stage 30 (pseudostem = 5 cm) occurred at progressively lower mainstem leaf numbers with delay in sowing. However, as shown in Tables 1--3, later sowing was also associated with acceleration of stem apex development such that, for example, the double ridge stage also occurred at progressively lower leaf numbers. Consequently, there exists a close relation- ship, between leaf sheath length and the achievement of double ridge, which appears to be independent of sowing date within the range of dates studied in this experiment (Table 4). Thus the double ridge stage occurred when the pseudostem length, averaged over all sowings, was 4.0 + 0.38 for wheat, 3.1 + 0.29 for rye, and 2.0 + 0.31 for barley.
TABLE 4
Pseudostem length (cm) at the mainstem double ridge stage for a range o f sowings of winter cereals
Sowing Wheat Rye Barley
1 4.1 5.2 2 3.4 3.1 3 3.0 2.7 4 4.9 2.5 5 <6.0 2.8 6 3.0 2.5 7 2.7 2.8 8 4.7 2.9
2.3 1.0 2.0 1.5
<3.0
Very similar values for wheat, falling within the range 3.2--4.0 cm, have been recorded elsewhere for October/November sowings (Tottman, 1977; Baker and GaUagher, 1983), leading to the proposal that sheath length be used as a convenient index of the achievement of the double ridge stage, for example in the timing of herbicide applications. However, Kirby et al. (1985b) have recently questioned the usefulness of this approach by demon- strating that wheat plants sown during the first 2 weeks of September in southern England did not reach the double ridge stage until their pseudo- stems (8.1 cm) were more than twice the corresponding values for October- sown crops (3.3--3.8 cm). This effect was not repeated in the present work, presumably owing to different combinations of temperature and photo- period, but there is an indication of a similar trend in winter rye (Table 4). In spite of the commercial importance of the crop, there is a lack of compar- able data for winter barley.

334
DISCUSSION
The remarkably synchronous anthesis of mainstem and tiller ears o f dif- ferent ages, and of the florets of different ages within each ear, is the out- come of a long and complex series o f developmental processes, the rate and duration o f which are largely controlled by environmental factors, principal- ly temperature and photoperiod. This ordered outcome of a range of diverse processes (e.g., the initiation, development and senescence of leaves, spike lets and tillers; extension of leaf sheaths and stem internodes, etc.) depends upon the coordination in t ime and space of some, but not all, of these pro- cesses. Two of these interrelationships are explored in the present paper.
Firstly, Figs. 1--3 support the idea o f a close temporal and spatial relation- ship between the terminal spikelet or awn primordium stage, stem extension and tiller mortal i ty in crops of all three species sown at widely differing sow- ing dates, and, in particular, Fig. 2 supports the observation of Biscoe and Willington {1984) that the achievement o f the terminal spikelet in mainstems of Avalon wheat immediately precedes rapid tiller death. Important features of these relationships include the progressive shortening of the period be- tween the start o f stem extension and the terminal spikelet or awn primordia stage in wheat and rye, and the distinctly different pattern of response for winter barley. Secondly, although the usefulness of pseudostem length as a guide to the achievement of the double ridge stage in wheat (Tottman, 1977) is confirmed over a wider range of sowing dates by the present work, there is an urgent need to understand why the relationship does not appear to hold for very early crops in the South of England (Kirby et al., 1985b) and to confirm that a similar relationship holds for barley (Table 4). The present work was not designed to explore the mechanism of these interrelationships but, from other studies of the influence o f environmental factors on induc- tion of flowering and cell expansion (e.g., Hay and Heide, 1983), it seems likely that the synchrony of apical development and sheath or stem exten- sion is the result of concerted responses to photoperiod, possibly mediated by gibberellins. In contrast, the association between the terminal spikelet stage and tiller mortali ty is, presumably, the consequence of intraplant com- petition between the extending mainstem and tillers, especially for nitrogen.
Figs. 1 and 5 show that, up to the start of rapid stem extension, each suc- cessive leaf sheath is longer than its predecessor, whereas the position of the apex from which the leaves are growing remains essentially stationary a few mm above the crown. Consequently, the distance which each successive leaf must extend before its tip or ligule is visible increases, and the effect is more pronounced with delay in sowing. In spite of this, each successive leaf ap- pears at regular intervals of accumulated temperature (Baker et al., 1980; Del6colle and Gurnade, 1980).
Hay and Tunnicliffe Wilson (1982) found that the rate of wheat leaf ex- pansion in the field, per unit o f accumulated temperature, did not vary with leaf number, but this investigation was concerned only with extension of

335
leaves after appearance; although this phase of extension can account for some 80% of the final leaf length, it is clear from the dissection studies of Gallagher (1979) that the rate of leaf extension has already fallen markedly by the time of leaf appearance. There have been few investigations of the growth within the leaf sheaths of pre-emergent cereal leaves. However, the detailed measurements of Kemp {1980) indicate that the rate of extension of emerging leaves should be resolved into two components, the relative rate of extension of the extension zone and the length of the extension zone. Since the latter varies with stage of development of the leaf and with the length of the encircling leaf sheath, it may not prove necessary to invoke dif- ferences in relative rates of extension to explain the temperature relations of leaf appearance. This is currently being examined in a simpler system, the leek, which is large, uniculm and remains vegetative throughout the first growing season. The mechanism of the control of leaf extension by sheath length is not yet understood, although Kemp {1980) suggests the involve- ment of light penetrating through (or down) the enclosing sheaths.
Recently, there has also been renewed interest in the part played by leaf sheaths in controlling the growth of grass plants. It has been known for some time that the pattern of regrowth of a grass sward can be strongly dependent upon the length of the stubble left at harvest (Davies et al., 1983) but this is only partly the consequence of differences in photosynthetic area. By the use of simulated sheath tubes of varying lengths, Wilson and Laidlaw (1985) have shown that the sheath or sheath tube length has a pronounced morpho- genic influence, affecting the final dimensions of the emergent leaf of perennial ryegrass tillers by its effect on the expansion of the cells in the younger areas of the leaf. The authors suggest that this is also a light-medi- ated effect, by which the growth of the younger tissues is affected by the emergence of the tip into an environment in which the spectral composition of the incident radiation is changed. Further work will no doubt prove that these two phenomena are closely related.
In conclusion, in view of the fact that pratical guides to the timing of nitrogen applications for wheat crops are now being devised (e.g., Appleton and Haggar, 1985) which rely upon the recognition of the double ridge stage and the approach of the terminal spikelet stage, it is essential for farmers and advisors to have reliable field indices of apical development which do not depend upon detailed dissection. The present work supports the usefulness of two readily-measured indices of apical development in wheat (pseudostem length and the height of the ear above the crown) but it is clear from the ob- servations of Kirby et al. (1985b) that pseudostem length may not be a reli- able index of double ridge for early-sown wheat crops in southern England and, presumably, in southern Europe. Finally, the demonstration that the timing of stem extension in relation to apical development is different for winter barley indicates that much more information is required before such a practical guide to the fertilization of this crop can be drawn up.

336
ACKNOWLEDGEMENTS
The a u t h o r is grateful to J o a n n a Asbeek-Brusse, Jacquel ine Davidson and Janice H a m p s o n fo r technical assistance, to Dr. E.J.M. Kirby fo r his thor- ough reading o f this pape r in draft , and to Dr. D.R. K e m p for useful discus- sions on leaf extension.
REFERENCES
Appleton, R.J. and Haggar, P.H.F., 1985. Apical-timed N could add precision to ADAS recommendations. Arable Farming, Feb., pp. 26--30.
Baker, C.K. and Gallagher, J.N., 1983. The development of winter wheat in the field. 1. Relation between apical development and plant morphology within and between sea- sons. J. Agric. Sci., Camb., 101: 327--335.
Baker, C.K., Gallagher, J.N. and Monteith, J.L., 1980. Daylength change and leaf appear- ance in winter wheat. Plant Cell Env., 3 : 285--287.
Biscoe, P.V. and Willington, V.B.A., 1984. Cereal crop physiology -- a key to accurate nitrogen timing. In: Marketable Yield of Cereals, Course Papers 1984. Arable Unit, National Agricultural Centre, Coventry, pp. 67--74.
Couvreur, F., Gate, P., Ingoat, G. and Mass6. J., 1985. Les Stades de B16. Institut Tech- nique des C~r~ales et des Fourrages, Paris, 12 pp.
Darwinkel, A., 1983. Ear formation and grain yield of winter wheat as affected by time of nitrogen supply. Neth. J. Agric. Sci., 31: 211--225.
Davies, A., Evans, M.E. and Exley, J.K., 1983. Regrowth of perennial ryegrass as affected by simulated leaf sheaths. J. Agric. Sci., Camb., 101: 131--137.
Del~coUe, R. and Gurnade, J.C., 1980. Liaisons entre le d~veloppement et la morphologie du bl~ tendre d'hiver. 1. Stades de d~veloppement de l'apex, apparition des feuilles et croissance de la tige. Ann. Am~lior Plant., 30: 479--498.
Gallagher, J.N., 1979. Field studies of cereal leaf growth. 1. Initiation and expansion in relation to temperature and ontogeny. J. Exp. Bot., 30: 625--636.
Hay, R.K.M., 1978. Seasonal changes in the position of the shoot apex of winter wheat and spring barley in relation to the soil surface. J. Agric. Sci., Camb., 91: 245--248.
Hay, R.K.M. and Abbas AI-Ani, M.K., 1983. The physiology of forage rye (Secale cere- ale). J. Agric. Sci., Camb., 101 : 63--70.
Hay, R.K.M. and Heide, O.M., 1983. Specific photoperiodic stimulation of dry matter production in a high-latitude cultivar of Poa pratensis. Physiol. Plant., 57 : 135--142.
Hay, R.K.M. and Tunnicliffe Wilson, G., 1982. Leaf appearance and extension in field- grown winter wheat plants: the importance of soil temperature during vegetative growth. J. Agric. Sci., Camb., 99: 403--410.
Kemp, D.R., 1980. The location and size of the extension zone of emerging wheat leaves. New Phytol., 84: 729--737.
Kirby, E.J.M. and Appleyard, M., 1984a. Cereal Development Guide, 2nd Edn. Arable Unit, National Agricultural Centre, Coventry, 95 pp.
Kirby, E.J.M. and Appleyard, M., 1984b. Cereal plant development -- assessment and use. In: The Nitrogen Requirements of Cereals. Reference Book 385. Ministry of Agricul- ture, Fisheries and Food, H.M.S.O., London, pp. 21--38.
Kirby, E.J.M. and Faris, D.G., 1972. The effect of plant density on tiller growth and morphology in barley. J. Agric. Sci., Camb., 78: 281--288.
Kirby, E.J.M., Appleyard, M. and Fellowes, G., 1985a. Effect of sowing date and variety on main shoot leaf emergence and number of leaves of barley and wheat. Agronomie, 5: 117--126.

337
Kirby, E.J.M., Appleyard, M. and Fellowes, G., 1985b. Variation in development of wheat and barley in response to sowing date and variety. J. Agric. Sci., Camb., 104: 383--396.
Porter, J.R., 1984. A model of crop canopy development in winter wheat. J. Agric. Sci., Camb., 102: 383--392.
Purvis, O.N., 1961. The physiological analysis of vernalization. Encycl. Plant Physiol., 16: 76--122.
Stern, W.R. and Kirby, E.J.M., 1979. Primordium initiation at the shoot apex in four con- trasting varieties of spring wheat in response to sowing date. J. Agric. Sci., Camb., 93 : 203--215.
Tottman, D.R., 1977. The identification of growth stages in winter wheat with reference to the application o f growth-regulator herbicides. Ann. Appl. Biol., 87: 213--224.
Tottman, D.R., Makepeace, R.J. and Broad, H., 1979. An explanation of the decimal code for the growth of cereals, with illustrations. Ann. Appl. Biol., 93: 221--234.
Weir, A.H., Bragg, P.L., Porter, J.R. and Raynor, J.H., 1984. A winter wheat crop simula- tion model without water or nutrient limitations. J. Agric. Sci., Camb., 102: 371-- 382.
Wilson, R.E. and Laidlaw, A.S., 1985. The role of the sheath tube in the development of expanding leaves in perennial ryegrass. Ann. Appl. Biol., 106: 385--391.