somatosensory cortical activity in relation to arm posture
TRANSCRIPT

JOURNALOFNEUROPHYSIOLOGY
Vol. 76, No. 4, October 1996. Printed in U.S.A.
Somatosensory Cortical Activity in Relation to Arm Posture:
Nonuniform Spatial Tuning
S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
Graduate Program in Neuroscience, and Departments of Physiology and Neurosurgery, University of Minnesota, . Minneapolis, Minnesota 55455
SUMMARY AND CONCLUSIONS
1. Single unitary activity in primate somatosensory cortex (SI) was recorded while monkeys maintained a range of static arm postures. Unit discharge was related to parameters defining the posture of the arm by multiple linear regression techniques.
2. Two monkeys were trained to grasp a manipulandum pre- sented at locations distributed throughout their workspace. The discharge of single units in SI was recorded for 3 s while the monkeys maintained contact with the manipulandum and the mean discharge rate over this hold time was related to the location of the hand and to the shoulder and elbow joint angles of the arm.
3. Unitary activity of 17 1 neurons in the proximal arm region of areas 3, 1, and 2 was recorded during the task. Of the total, 78 neurons had activity that varied with the location of the hand in space. Neuronal discharge typically varied monotonically with the target location, reaching a maximum at the borders of the work- space. The discharge rate in most of these neurons varied with both shoulder and elbow angles.
4. Discharge rate was related to the hand’s location along three axes by means of a polynomial fit. In approximately half of the neurons, activity varied significantly only for displacements along a single axis in space. However, many neurons exhibited nonlinear relations between hand location along this preferred axis and dis- charge rate. Discharge rate did not vary for displacements of the hand in the plane perpendicular to this preferred axis (null plane).
5. In other neurons, discharge rate varied for hand displace- ments in a plane, i.e., along two perpendicular axes. Displacements of the hand along the axis perpendicular to this plane (null axis) did not affect the discharge rate. In only a small minority of neurons did discharge rate vary for hand displacements along all three axes in space.
6. The distribution of the sensitivity of the neural population to hand displacements along arbitrary directions in space was not uniform. On average, hand displacement along a vertical axis led to the smallest modulation of neural discharge, and displacement of the hand along the anteroposterior direction led to the largest modulation of activity.
INTRODUCTION
The aim of movement is to act on or interact with the environment. Controlling such an interaction, however, is extremely difficult in the absence of information about the relations between the movement and the environment. Whether tracing the edges of an object or simply trying to stand upright, one is heavily reliant on sensory inputs to monitor the ongoing status of action.
This is certainly true for the control of arm movements by the CNS. When the actions to be performed are either rudimentary or well rehearsed, then performance in the ab- sence of sensory information (either somatosensory or vi-
sual) can be quite adequate. If the task is changed or the starting arrangement is altered, however, performance de- grades quickly. For example, monkeys with bilateral deaffer- entation of the arms can perform conditioned movements about a single joint (Knapp et al. 1963 ) , planar pointing movements (Polit and Bizzi 1979)) and even three-dimen- sional pointing movements (Taub et al. 1975). If the starting position of the arm is changed without the animal’s knowl- edge, however, the animal is no longer able to reach to a visual target (Polit and Bizzi 1979). Even in well-rehearsed movements, careful analysis reveals performance flaws after deafferentation. Humans with large-fiber sensory neuropathy produce characteristic errors during both horizontal and ver- tical planar movements, even when the movements are well practiced (Ghez et al. 1990; Sainburg et al. 1993 ) .
These observations highlight the importance of informa- tion arising from somatosensors in the control of movement. Despite the intimate involvement of this information in the control of movement and the impact of somatosensory input on movement-related signals in the CNS, the question of how the information is processed and used remains unre- solved (Hasan 1992; Hasan and Stuart 1988). One use of somatosensory inputs may be to determine the static posture of the limb. For example, a representation of current arm posture is necessary to determine the direction and amplitude of an intended movement (Flanders et al. 1992; Helms Til- lery et al. 1991; Soechting and Flanders 1989). This repre- sentation could be provided by the somatosensory system.
Consistent with this viewpoint, physiological studies throughout the somatosensory system have revealed dis- charge related to static limb postures. Peripheral afferents (Edin 1992; Edin and Abbs 199 1; Hulliger et al. 1979; Knibestol and Vallbo 1970)) dorsal column nuclear neurons (Millar 1972; Surmeier and Towe 1987), thalamic relay neurons (Mountcastle et al. 1963; Poggio and Mountcastle 1963)) and somatosensory cortical (SI) neurons (Cohen et al. 1994; Gardner 1988; Gardner and Costanzo 198 1; Jen- nings et al. 1983; Mountcastle and Powell 1959) all show regular changes in activity related to the posture of the arm. Most of these studies involved changes in the posture of a single joint. Unit activity at all levels is monotonically re- lated to changes in joint angle, reaching a maximum at the anatomic limits of joint rotation.
Costanzo and 6ardner ( 198 1) showed that the discharge of some SI neurons is related to the position of more than one joint. Many muscles cross more than one joint, and muscle spindle afferents, are thought to contribute substan- tially to encoding limb posture (Goodwin et al. 1972; Mc-
0022-3077/96 $5.00 Copyright 0 1996 The American Physiological Society 2423

2424 S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
Closkey 1978). Because skin can also be expected to be deformed locally by rotation about more than one joint (Edin 1992; Edin and Abbs 1991; Edin and Johansson 1995)) one might expect that cutaneous afferents would also be sensitive to changes in the posture of the limb as a whole.
Neurons in cortical areas other than SI are also tuned to the position of the hand in space. Georgopoulos and colleagues (Georgopoulos and Massey 1985; Georgopoulos et al. 1984; Kettner et al. 1988) found that most cells in motor cortex and area 5 had a discharge that was linearly related to static hand position in space. Each cell’s activity was modulated maximally by a translation of the hand in one direction in space (the “best direction” ) . Thus static activity exhibited a tuning that was similar to that found in the same areas for active movements. Interestingly, although the static best directions in areas 4 and 5 appeared to be distributed uni- formly, Kalaska et al. ( 1983) found a nonuniform distribu- tion of best directions during active movements, with a pref- erence for movements directed anteriorly or medially. The interpretation of the cosine tuning of neural discharge has been controversial, because a variety of kinematic and ki- netic parameters may also exhibit cosine tuning under these experimental conditions (Georgopoulos et al. 1984; Helms Tillery et al. 1995; Kalaska 199 1; Lacquaniti et al. 1995).
The question thus arises: how does the spatial tuning of neurons in SI compare with the tuning that has been ascribed to motor cortex and area 5? This question has been addressed by Kalaska and colleagues (Cohen et al. 1994; Prud’homme and Kalaska 1994; Prud’homme et al. 1994) with the use of the same center-out task as the studies cited above. Those researchers found that discharge rate in SI also exhibits a cosine tuning to the location of the hand in a plane. However, the distribution of the cells’ preferred directions was nonuni- form, with a preponderance of the cells being tuned in the anteroposterior direction.
In this paper we extend the analysis of the spatial tuning of neurons in SI to three-dimensional displacements of the hand in space. We also used a larger range of hand locations than previous studies, because such an experimental design has the potential to provide a fuller description of a given cell’s discharge (Fu et al. 1993). In agreement with previous results, we found that SI discharge is related in a monotonic fashion to hand location, but that this relationship can be nonlinear. In particular, the discharge of each neuron only encodes passive hand displacement along a specific direction in space, or in a specific plane. In agreement with and ex- tending previous findings, we found that cell discharge was modulated maximally with hand displacement in the antero- posterior direction, and to lesser extent by displacement in the vertical or mediolateral directions.
Abstracts of a portion of these results have been presented previously (Helms Tillery et al. 1992, 1994; Soechting et al. 1992).
METHODS
Overview
Two young female rhesus monkeys (Macaca mulatta) were trained to grasp a manipulandum placed in arbitrary locations in a three-dimensional workspace by a robot arm (UMI RT-100 or Microbot Minimover- ) . Because the manipulandum provided postural support, this task requires little tonic activation of arm
muscles (Helms Tillery 1994). Once the monkey had been trained, the discharge of neurons in SI was recorded while the monkey maintained static postures. The recorded discharge was then related to the manipulandum location as well as the posture of the arm (shoulder and elbow angles) by means of linear regression tech- niques. Data were recorded from both left and right hemispheres, contralateral to the arm performing the task. For convenience of presentation, all data from the right hemisphere/left arm are re- flected across the midline.
Task and equipment
The task required monkeys to grasp a manipulandum consisting of either a small sphere or a 1-cm-diam bar that was oriented vertically. The locations, contained within a spherical wedge in front of the monkey’s shoulder, were presented in a random block design from two lists. The first list contained six targets: two lo- cated 5 cm in front of the monkey’s shoulder and 5 cm above or below the shoulder, and four at the corners of a square lying in the frontal plane at the limits of the animal’s reach. The second list contained targets at 3-cm intervals throughout a volume contained within 45’ to the left and right of the shoulder and 45’ above and below the shoulder. The nearest targets were located 5 cm from the monkey’s shoulder, and the farthest were at the limit of the monkey’s reach (25-30 cm). This volume is referred to as the monkey’s “workspace.”
Three colored light-emitting diodes (LEDs) were mounted on the manipulandum. A trial was initiated by lighting a red LED, cuing the monkey to press a 4-cm-diam button located near its lap. While the monkey pressed the “hold” button, the robot moved the manipulandum into position. Then a second cue lamp was lit. A yellow lamp cued the monkey to grasp the stationary manipulan- dum and maintain contact for 3 s ( “grasp” task). A green lamp cued the monkey to press a button on the manipulandum (Helms Tillery et al. 1995). I f the task was performed correctly, the mon- key was rewarded with a drop of juice.
In this paper we describe the discharge observed in the grasp task, where the monkey was able to receive support for the weight of the arm by grasping the manipulandum. Interspersed among the trials in which the monkeys grasped the manipulandum were other trials in which they were required to press a button on the face of the manipulandum (press) or grasp the handle after it had been rotated into an oblique orientation (grasp oblique). Results from these paradigms are described elsewhere (Helms Tillery 1994).
Behavioral analysis
The posture of the arm during these tasks was analyzed by a video based system (Motion Analysis), with markers placed at the shoulder, elbow, wrist, and the manipulandum. Arm posture was defined in terms of segment orientation angles (Soechting and Ross 1984), namely the yaw and elevation of the upper and forearm segments. One angle (7 for the upper arm, a for the lower arm) defined the yaw angle, measured in the horizontal plane, and the other (0 for the upper arm, 0 for the forearm) gave the elevation, measured in the vertical plane. The video derived data were used to analyze the relations between target location, task, and arm posture as described by the four orientation angles. The results of this analysis are described in detail elsewhere (Helms Tillery et al. 1995 ) . In the task described in this paper, the monkeys tended to hold the manipulandum with the arm held within 10’ of a vertical plane (cf. the natural posture of Scott and Malaska 1995 ) .
Surgical preparation
Once trained, the monkeys were outfitted with chambers for chronic recording with the use of techniques previously described (Fu et al. 1993). The monkeys were deeply anesthetized with

POSTURE-RELATED ACTIVITY IN SI 2425
ketamine (20 mg/kg ) and xylazine ( 1 mg/kg ) and placed in a stereotactic frame. The skull was exposed by midline incision and an 18-mm craniotomy was made at 7 mm anterior, 16 mm lateral (Snider and Lee 196 1) . A metal cylinder was then affixed to the skull with cranial screws and sealed with cranioplasty. During postoperative recovery, the animals received analgesics (Nubane or Buprenex) as needed and were kept on oral Amoxycillin for a period of 2-4 wk.
The monkeys were maintained on a water-restricted diet throughout the training and recording periods. The animals typi- cally received between 150 and 300 ml of water while working, and supplements of fruit. The animals did not receive water while in their home cages on any day that they worked, although food was available ad libitum. The animals were cared for in strict accordance with National Institutes of Health guidelines for the care and use of laboratory animals.
Physiological methods
During recording sessions, the animals’ heads were fixed and the discharge of cortical neurons was recorded with the use of tungsten electrodes (impedance 5- 10 MO). Once a candidate neu- ron was isolated, several samples of the action potential were over- lain on a computer screen. If the waveform was stable, and the neuron showed no evidence of injury discharge, its chamber coordi- nates and the waveform of a single action potential were recorded, along with the cell’s receptive field characteristics.
An experiment began with repeated presentation of the small block of six target locations (see Fig. 2). The neural activity was recorded during the 3-s period in which the animal held the manipu- landum. As the experiment progressed, a cumulative average of the cell’s discharge rate within fixed portions of the workspace was displayed as a color-coded map on a computer monitor. This permitted us to determine 1) whether the cell’s discharge was modulated in relation to changes in arm position and 2) whether these changes were consistent with repeated trials. I f the cell’s discharge showed both of these characteristics after two or three repetitions of each location, the larger block of target positions was presented. These targets were presented in a randomized block design, and each cell’s discharge was recorded for 200-300 trials or until isolation was lost. Activity was recorded as spike times at 3-kHz resolution throughout the 3-s hold period, and stored on a trial-by-trial basis for later analysis. In each of the hemispheres, recording was continued 5 or 6 days/wk for 3-6 mo.
Receptive Jield characterization
As each neuron was isolated, its receptive field properties were determined and noted. Response type was classified either as “cu- taneous” (brisk response to touch or stroke of a light brush or the handle of the brush) or “deep” (no cutaneous responsiveness, but response to rotation of a joint or palpation of a joint or muscle). Note that the cutaneous classification was not exclusive: a neuron classified as cutaneous could also have had a deep responsiveness. All neurons described here had receptive fields on the arm.
Anatomic methods
Two methods were used to identify the relationship between the coordinates of the recording chamber and locations within the cor- tex. In two hemispheres, several small lesions were produced by passing current through the recording electrodes. In the remaining hemisphere, five pins were placed into the cortex in the shape of a cross centered in the chamber. In each case, the locations of the marks provided both the X and Y directions of the chamber on the cortex, and the vertical direction of travel for electrodes in the chamber. After recording was completed, each monkey was deeply anesthetized with an intramuscular injection of 0.5 ml xylazine and
0.5 ml ketamine. The animal was perfused via the heart with normal Ringer solution followed by Zamboni’s fixative. The brain was removed, photographed, blocked, and placed in Zamboni’s fixative for several days. The blocks were then infiltrated with sucrose and cut into 40-pm frontal sections on a freezing microtome. Each 10th section was mounted and stained for Nissl substance with the use of either cresyl violet or thionin. The slides were carefully checked for lesion sites, identifying as many as possible. The identified lesions were localized by cross-checking the chamber map with the photographs of the cortical surface.
Data analysis
The first step in the analysis of each cell’s discharge was to class the cell as either modulated or nonmodulated with changes in arm posture. This determination was made by an analysis of variance. Because most of the recording sessions did not involve repeated measures, we used multiple linear regression techniques to determine whether each cell was modulated with hand location. To this end the firing rate for each trial was averaged over the 3- s sampling period and fitted to cubic equations in Cartesian coordi- nates of the hand
f = w, y, 2) (0
where fis the discharge rate of the neuron, P is a cubic polynomial having 20 terms, x is the anterior direction, y is medial, and z is up. The 95% confidence intervals for each term were computed and the terms for which this interval bracketed zero were discarded. The remaining terms comprised the “best-fit” equation for that neuron. The regressions were performed with the use of a singular value decomposition-based method (Press et al. 1988). For each neuron, an r2 value was computed as
r 2 = 1 - [X(X -jg”/C(f; -$>2] (2)
where 5 was the firing rate on the ith trial, 7 was the mean firing rate over all the trials, and j was the firing rate predicted for the ith trial by the regression polynomial. Next, an F value was com- puted as
F = (r2/k)l[(l - r2)lm] 0
where k was the number of coefficients with nonzero values, m was y2 - (k + l), and y2 was the number of trials. Cells for which the F value was significant to the 0.05 level were defined as modulated in the task.
Monotonicity
To provide a measure of the extent to which firing rate was monotonically related to hand location, the target location giving the highest discharge rate was identified. Target locations were first binned according to their height. The trials were then divided into three subsets, one for each bin, and the polygon that defined the perimeter of the sampled workspace in each bin was computed. The perpendicular distance from the point of maximal discharge to each of the edges of the polygon was then computed. The small- est such value gave the distance from the point of highest discharge to the edge of the sampled workspace.
Fitting to multiple axes
In some cases, a cell’s spatial tuning was restricted to a modula- tion along one axis in space. Displacements in a plane perpendicu- lar to this axis did not affect neural discharge (null plane). In other cases, neural activity was modulated by displacement of the hand along two perpendicular axes (i.e., in a plane), and displacements

2426 S. I. HELMS TILLER-Y, J. F. SOECHTING, AND T. J. EBNER
of the hand in a direction perpendicular to this plane (null direc- tion) did not affect neural activity.
The analytical procedure to define the best-fit axis or axes began with a linear model of the discharge rate
f = CC) + Cl x + c,y + c3z (4)
This fit provided the axis x’, which best described the cell’s dis- charge
x’ = (c,, c2, c&c:. + c; + cg) (5)
Thus x’ is a unit vector pointing in a direction given by the X, y, z coefficients from Eq. 4. Discharge rates were then fitted to a cubic polynomial along this axis
f = ho + b,x’ + b2xt2 -I- b3xr3 (6)
To determine whether the discharge rate was modulated in direc- tions perpendicular to this best axis, we used the following proce- dure. We first defined 18 planes containing the vector x’, each separated by 10’ increments, and fitted the data with the use of a cubic polynomial on two variables, X’ as defined above, and y I) which was always perpendicular to x ‘. For each of these 18 planes an r* value was computed. The y ’ giving the highest value for Y* was chosen to be the second axis defining the modulation plane. Finally, the data were fitted to a cubic polynomial along all three axes
f= M-C y’>
f= i(x’, y’, z’) (7)
This procedure resulted in three equations (Eq. 6 and 7) relating the discharge rate of the neuron to the location of the hand. In each instance, terms whose coefficients did not differ significantly from zero were discarded. In order to classify each cell as requiring one, two, or three axes to account for the observed changes in discharge rate, the r* values for the three fits were compared. We used the criterion that for cells described as being tuned along a best direction or in a modulation plane, the reduced fits should account for 290% of the variance accounted for by the complete fit.
Joint angles
We can also express the discharge directly in terms of joint angles, because there is a unique relation between the location of targets in space and the angles defining the posture of the arm (Helms Tillery et al. 1995). One likely starting point is to classify each neuron as being related to either the shoulder or the elbow. To make this assessment, the data were fitted to a cubic polynomial in elbow flexion (4)
f = a0 + a,+ i- a242 + a3+3 (8)
The Y* obtained from this fit was then divided by the y2 that resulted from the fit to the full cubic equation (Eq. 7). To fit the discharge rate to single axes in joint space, we regressed the discharge rates of the neurons to linear functions of the joint angles. This analysis provided a single axis a’ that captured the greatest component of variance in the same way as the first axis in Cartesian coordinates (Eq. 5), except this axis had four terms, one for each of the joint angles
a’ = (c,, ~0, c,, “~V~(C;; + c; -I- c: -I- c$) (9)
Distribution of tuning directions
As we will show, different neurons are tuned along different numbers of spatial axes. Given this result, displaying the distribu- tion of the cells’ best axes does not fully capture the spatial proper- ties of the population. The difficulty is to combine the results for
TABLE 1. Enumeration of cells according to cortical location
and receptive Jield characterization
No. of No. of Cells Cutaneous Deep Receptive Area Cells Modulated Receptive Field Field
3a 8 5 2 (0) 5 (4) 3b 42 19 16 (8) 25 (11) 1 55 24 24 (10) 27 (14) 2 38 13 20 (10) 12 (2) 215 28 17 25 (17) 2 (0) Total 171 78 87 (45) 71 (31)
Numbers in parentheses refer to cells whose activity was modulated during the task.
those cells modulated along single axes with those modulated in two spatial directions. We have addressed this issue by reframing the question: for a hand displacement along a given direction, what proportion of the neurons is significantly modulated? We made the assumption that, for cells modulated along a single axis, the responsiveness to displacements along any other axis is propor- tional to the cosine of the angle between the two. For neurons with null axes, we assumed that the cells’ responsiveness would be proportional to the projection of the displacement onto the cells’ modulation planes. This is equal to the absolute value of the sin of the angle between the displacement and the null axis.
To present these data, we plotted the results on an equal-area projection of an upper hemisphere (Watson 1983; see also Schwartz et al. 1988). In this type of projection, equal areas on the hemisphere are accorded equal areas on the projection. The pole is represented as the center of the circle, and the equator is at the circumference. This provides for a representation with minimal distortion near the pole and maximum distortion near the equator.
RESULTS
Data base
Table 1 gives a breakdown of the cells recorded from the arm area of SI into cortical area, receptive field type, and whether they were modulated with the location of the hand. Of the 17 1 cells, 78 (45%) were significantly modulated by hand position in space. The cells that were significantly modulated came from pools with both cutaneous and deep sensory modalities and were distributed primarily across areas 3b, 1, and 2 and the transition zone between areas 2 and 5. As noted in METHODS, the criteria used to determine a cell’s response modality favored a designation of cuta- neous: a determination of cutaneous modality did not ex- clude the possibility that a cell also had deep responsiveness, whereas a determination of deep response was exclusive. Therefore the slight preponderance of cutaneous respon- siveness in the sample does not necessarily imply a prepon- derance of recording sites in areas of cortex that are largely cutaneous.
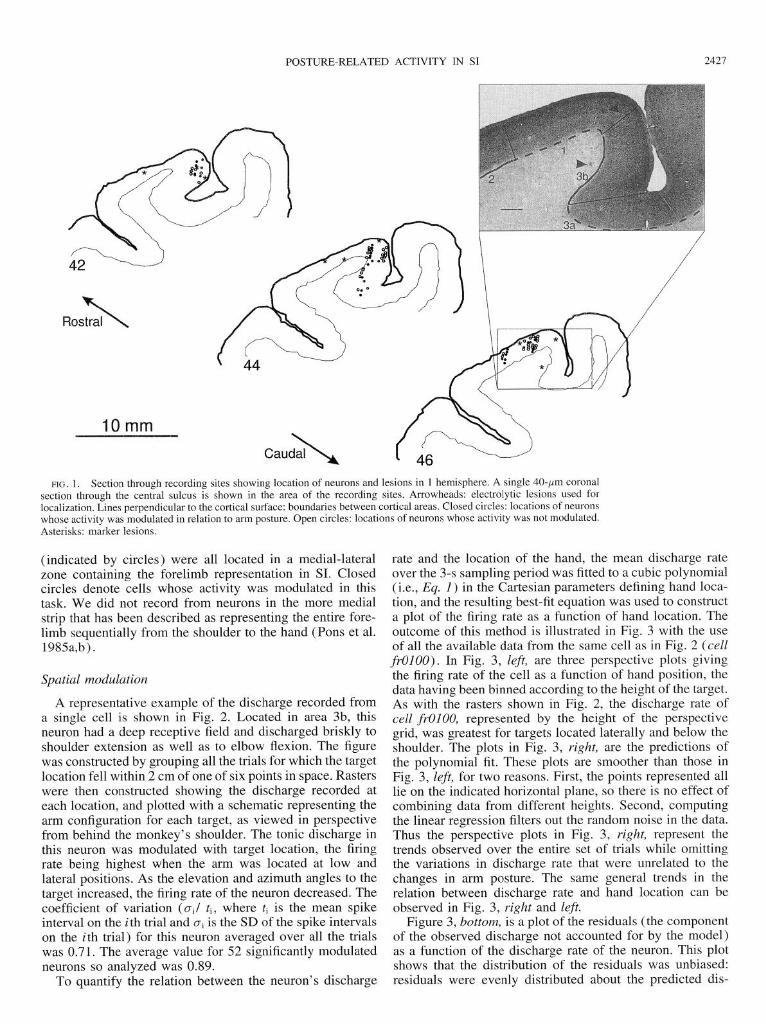
Figure 1 shows a coronal section through the central sul- cus in the region of the recordings. Each cortical area is indicated by a line at the gray-white border, and the lines perpendicular to the cortical surface denote the demarcations between areas. These were determined with the use of stan- dard criteria (Jones et al. 1978; Powell and Mountcastle 1959; Prud’homme et al. 1994). The arrowheads in Fig. 1 denote two marker lesions that are indicated as asterisks in the camera lucida drawing of this section (46) showing a reconstruction of cell locations for this hemisphere. The cells
POSTURE-RELATED ACTIVITY IN SI 2427
IOmm
FIG. 1. Section through recording sites showing location of neurons and lesions in 1 hemisphere. A single 40-pm coronal section through the central sulcus is shown in the area of the recording sites. Arrowheads: electrolytic lesions used for localization. Lines perpendicular to the cortical surface: boundaries between cortical areas. Closed circles: locations of neurons whose activity was modulated in relation to arm posture. Open circles: locations of neurons whose activity was not modulated. Asterisks: marker lesions.
(indicated by circles) were all located in a medial-lateral zone containing the forelimb representation in SI. Closed circles denote cells whose activity was modulated in this task. We did not record from neurons in the more medial strip that has been described as representing the entire fore- limb sequentially from the shoulder to the hand (Pons et al. 1985a,b).
Spatial modulation
A representative example of the discharge recorded from a single cell is shown in Fig. 2. Located in area 3b, this neuron had a deep receptive field and discharged briskly to shoulder extension as well as to elbow flexion. The figure was constructed by grouping all the trials for which the target location fell within 2 cm of one of six points in space. Rasters were then constructed showing the discharge recorded at each location, and plotted with a schematic representing the arm configuration for each target, as viewed in perspective from behind the monkey’s shoulder. The tonic discharge in this neuron was modulated with target location, the firing rate being highest when the arm was located at low and lateral positions. As the elevation and azimuth angles to the target increased, the firing rate of the neuron decreased. The coefficient of variation (ai/ ti, where ti is the mean spike interval on the ith trial and cri is the SD of the spike intervals on the ith trial) for this neuron averaged over all the trials was 0.71. The average value for 52 significantly modulated neurons so analyzed was 0.89.
To quantify the relation between the neuron’s discharge
rate and the location of the hand, the mean discharge rate over the 3-s sampling period was fitted to a cubic polynomial (i.e., Eq. 1) in the Cartesian parameters defining hand loca- tion, and the resulting best-fit equation was used to construct a plot of the firing rate as a function of hand location. The outcome of this method is illustrated in Fig. 3 with the use of all the available data from the same cell as in Fig. 2 (ceZZ frOlO0). In Fig. 3, left, are three perspective plots giving the firing rate of the cell as a function of hand position, the data having been binned according to the height of the target. As with the rasters shown in Fig. 2, the discharge rate of cell fro1 00, represented by the height of the perspective grid, was greatest for targets located laterally and below the shoulder. The plots in Fig. 3, right, are the predictions of the polynomial fit. These plots are smoother than those in Fig. 3, left, for two reasons. First, the points represented all lie on the indicated horizontal plane, so there is no effect of combining data from different heights. Second, computing the linear regression filters out the random noise in the data. Thus the perspective plots in Fig. 3, right, represent the trends observed over the entire set of trials while omitting the variations in discharge rate that were unrelated to the changes in arm posture. The same general trends in the relation between discharge rate and hand location can be observed in Fig. 3, right and left.
Figure 3, bottom, is a plot of the residuals (the component of the observed discharge not accounted for by the model) as a function of the discharge rate of the neuron. This plot shows that the distribution of the residuals was unbiased: residuals were evenly distributed about the predicted dis-

2428 S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
Time (sets)
FIG. 2. Activity of a representative neuron at different arm postures. Rasters denote the activity of 1 neuron in the somatosensory cortex (ST) at 6 different arm postures. Postures of the right arm and limb segments (S, shoulder; E, elbow; H, hand) corresponding to each hand location are indicated schematically as they would appear viewed from behind the monkey’s shoulder. In this neuron, the tonic discharge was highest for hand locations below and lateral to the shoulder (bottom right). Each raster was constructed by combining all trials for which the hand location fell within 2 cm of the indicated location.
charge rate, and did not vary as a function of the firing rate. This measure of the goodness of the fit indicates that the cubic polynomial provided a reasonable description of the discharge rate. In addition, this fit gave r2 = 0.84 [P( 6,94) = 83.2, P < O.OOl] ; thus the fit accounted for >80% of the variance present in the data.
Two features of these plots are representative of results from all of the modulated cells. First, the discharge rate changed monotonically as the hand was displaced along any given line. For example, as the target locations changed from left to right at a given height, the firing rate of the neuron increased continuously. Second, the major component of the change in discharge rate occurred with displacements along a horizontal axis. The magnitude of the change in discharge rate for vertical displacements was less than that occurring along the horizontal axis. In addition, the fact that the sur- faces of the plots were nearly planar is consistent with this neuron’s discharge rate being linearly related to hand posi- tion. In terms of the multiaxis analysis discussed later, this neuron’s discharge could be specified by knowing the loca- tion of the hand along the single axis <0.06, -0.91, -0.41>
(Es* 5). Figure 4 shows representative plots of the spatial tuning
for four neurons. As in Fig. 3, the firing rate is plotted on three horizontal planes located above, at, and below the
monkey’s shoulder. In Fig. 4 the firing rate is represented by shading and the three planes are drawn in the correct spatial relation to each other. Each plane is bounded by target elevation and azimuth angles of 545” and distances of 6 and 20 cm from the shoulder. The monkey’s shoulder is located at the intersection of the center plane with the vertical grid line.
The features observed on these plots capture the spectrum of spatial modulation observed in SI during the grasp task. For most of the neurons, discharge changed monotonically with hand location. Examples include the plots for the cells shown at the top Zeft, bottom Left, and top right. In the plot shown at the top left (same cell as in Figs. 2 and 3)) the contour lines are parallel to each other along the anterior- posterior direction. The maximal changes in discharge rate occurred as the target location was displaced in a direction perpendicular to these lines, along a left-right axis. The evenly spaced contour lines imply that this neuron’s dis- charge is related linearly to hand location in space.
The two plots shown at the top right (labeled fro 111) and bottom left (labeled liOO52) have the characteristics most representative of this pool of neurons. Cell $41 II, also lo- cated in area 3b, had a small receptive field with deep respon- siveness located on the posterior aspect of the axilla, whereas ceZ2 ZiOO52 was located in area 3a and also had a receptive

POSTURE-RELATED ACTIVITY IN SI 2429
Data Representation Data Cubic Fit
Residuals vs. Predicted Discharge
s ., ,. . . .: _ ‘, I ;;;. ?.‘“. i ., ,. ? .:” .,. .._. I . ...,. . . . . _I . .,‘., , :. .: ,”
2 0 10 20 30
PK. 3. Modulatton of neural acttvtty wrth hand locatton. Left. data from the same neuron as shown m Fig 2; height of perpecttve plot denotes
mean tirmg rate. To construct thts figure. the workspace war spin into 3 horrzontal bms, and all the trrals for whrch the target fell wrthm a particular
bm were pooled. The mean herght of each bin relative to the shoulder 1s mdtcated to the right of the plots. The monkey’s shoulder is approxrmately centered m the medial-lateral dnection on the near-rrght edge of the middle <lab Each slab extends 25 cm antertorly and 1-15 cm to each side of the caglttal plane. As m Frg. 2, thus neuron’s dtscharge rate Increased as the hand was located more laterally or downward. Riglrt: perspective plot\ constructed from the tit to a cubic polynomral \how the same general trends. Bol~m. resrduali fat thts fit. Residuals are normally drstrrbutcd about the predicted dtscharge rate, and do not show any regular change as a functton of the discharge rate. Both measures rndrcate that the model order was approprtate.
field on the arm that was classified as deep. In the plots for both cells fro111 and liO0.52, firing rate increases monotoni- cally as the workspace is traversed, but the contour lines are neither straight nor evenly spaced, indicating that firing rate cannot be characterized by a planar model, as was the case for cell,frOlOO. The data plotted at the bottom left are closer to linear than those shown at the top right, but neither is adequately described by a linear function. As described be- low, the data at the top right require two axes to capture all the variance, whereas the data at the bottom left were well described with the use of only one spatial axis.
The final plot, shown at the bottom right (cell frOO75, located in area 1 with a large cutaneous receptive field on the ulnar side of the arm, extending over the upper arm and forearm), illustrates a neuron whose relation between
discharge rate and hand location was more complex. The discharge of this neuron was not monotonically related to hand location. Instead, there was a saddle point near the center of the workspace, the rate of discharge increasing with hand locations both posterior as well as anterior to this saddle point. Of the 78 neurons with significant modulation in the grasp task, -20% had a complex relation to hand location like that shown at the bottom right. The remainder appeared monotonic.
Monotonicity
One measure of whether a neuron’s tiring rate varies monotonically with the location of the hand in space is the distance from the edge of the sampled workspace to the target location giving the maximum tiring rate (see METH-
ODS). For cells satisfying this criterion, this distance will be zero: the target location giving maximal discharge rate should be located directly on the edge of the sampled work- space. Figure 5 shows that in >50% of the cells maximum firing rate occurred at the borders of the workspace. and in all cases maximum firing was within 3 cm of the boundary. However, as seen for cell,fr-0075 in Fig. 4, there is a limita- tion to this measure. It is possible for a cell to have maximal
0 ‘0
Spikes/sex
0 10 0 50
FIG. 4. Spatial discharge profiles for 4 representative neurons. The dis- charge rate as a function of hand position is shown for 4 neurons. These
figures were constructed with the use of a cubic polynomial fit to the experimental data. Each panel shows activity in 3 horizontal planes: at the shoulder and 8 cm above and below the shoulder. Dashed grid lines: azimuth angles of +45”, 0”. and -45” and a distance of 20 cm. Except for the example shown at the hottom right, neural activity changed monotonically as a function of hand location. The spatial tuning of nelrron ,JrOrOO (the same neuron depicted in Figs. 2 and 3) is planar, indicating a linear relation between hand position and discharge rate. The profiles for IIWTOIZS ,fiOI I I and liOO52 are monotonic, but not linear. Ne~on ,JrOO75, shown at the bottom right, has a nonmonotonic spatial profile, but such neurons were encountered less frequently.

S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
Distribution of Peak-Edge Distances
0
0.0 0.5 1.0 1.5 2.0
Distance (cm)
2.5 3.0
FIG. 5. Distribution of distances from the hand location at which neural activity is maximal to the edge of the workspace. For each neuron, the trial for which the neuron fired maximally was identified. The distance between that location and the edge of the sampled workspace was then computed, and the distribution of all such distances is shown in the figure. The mode of this distribution is at 0 cm, as would be expected if most of the neurons had monotonic spatial discharge profiles.
discharge for targets at the edge of the workspace without having a monotonic discharge profile. As mentioned before, -20% of the cells had a spatial tuning that exhibited a saddle point.
Spatial tuning along a single axis or in a plane
Many of the cells could be characterized by their re- sponses to a displacement of the hand along a single axis. In these instances, neural discharge could be predicted by taking the projection of the hand location onto that axis. In other instances, neural discharge depended on the projection of hand location onto a plane. The analytical procedures used to characterize response along a single axis or in a plane have been described in METHODS, the first step in the procedure being to define the best direction. Figure 6 shows the firing rates of the four cells illustrated in Fig. 4 as a function of hand location along the best direction. The two cells at the left (labeled fro100 and liO052) show a simple relation between discharge rate and hand location along this axis. The small amount of scatter of the data points indicates that one axis could account for the modulation of neural activity for these two cells. For cell frOIO0 (top left), the fit along a single axis accounted for 98% of the variance that was accounted for by the full three-dimensional fit. Fur- thermore, except for a threshold nonlinearity, the fit was
close to linear. The larger amount of scatter of the data points at the bottom left (cell liOO52) indicates that a second axis contributed to the cell’s modulation: the first axis ac- counted for 87% of the variance, the second axis accounted for 11%. For the two cells shown at the right, the plots exhibit wide scatter for any given point along the best direc- tion. This indicates that the single axis has not captured the variance present in the data. For cell fro1 11 (top right), the first axis accounted for 67% of the variance, with the second axis accounting for most of the remainder (32%). A similar result holds true for cellfrOO7.5, the first axis accounting for 60% of the variance and the second axis for 39%.
The spatial tuning of the four cells we have examined in detail (Figs. 4 and 6) can be better appreciated in Fig. 7, in which we show the predicted firing rate plotted in the modulation plane. The axes extending to the right are the best direction (cf. Fig. 6) and the axis extending back and to the left is the axis, perpendicular to the best direction, that provided the greatest reduction in the variance unac- counted for by the single-axis model ( see METHODS). In agreement with the qualitative observations obtained from Fig. 6, there is little modulation in firing rate along the second axis for the two cells shown at the left. The reason why the activity of the two cells at the right cannot be described by modulation along a single axis also becomes apparent. Along a direction close to the best direction, the activity of these cells appears to be increase monotonically, but the tuning along the second axis appears closer to bell shaped.
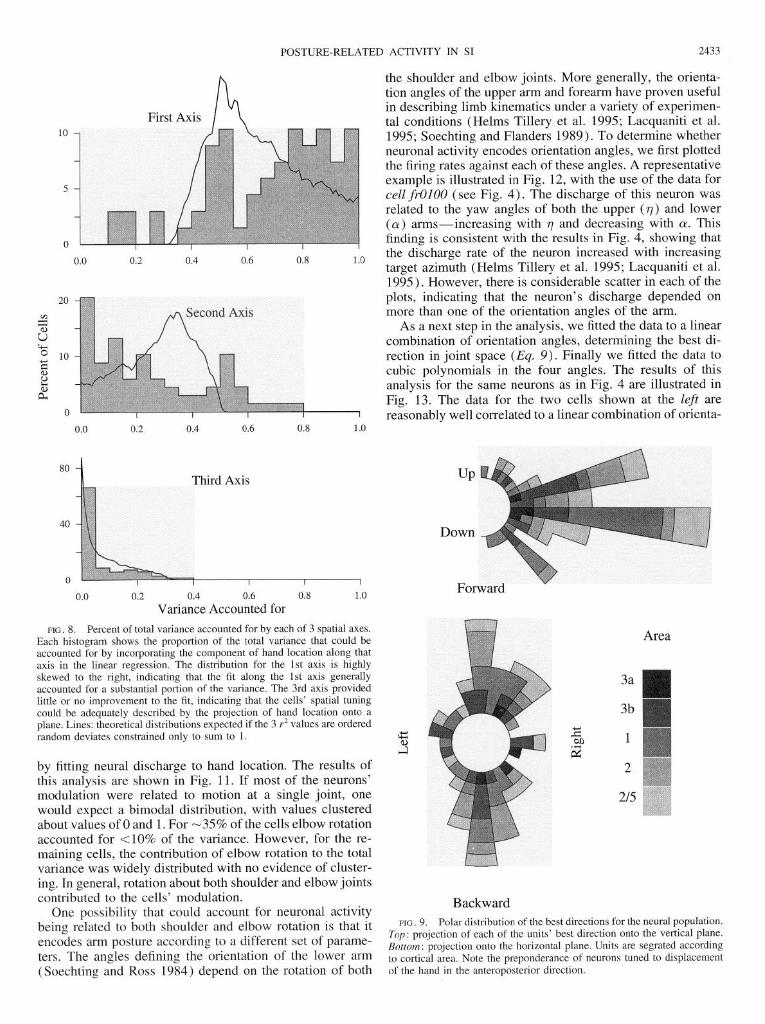
Figure 8 shows the distribution of the fraction of the total variance accounted for due to the cells’ modulation along each of the three axes. For -40% of the cells, the first axis accounted for X30% of the variance accounted for by the full three-dimensional fit. When cell discharge was fitted to the projection of hand location onto a plane, the model could account for >80% of the variance in 88% of the cells, and for >90% of the variance in 75% of the cells. Therefore the third axis contributed little to the modulation of neural discharge. It is also clear that some cells can be adequately characterized by their tuning along a single direction in space, but that others cannot. In fact, for 22% of the cells the first axis accounted for <50% of the variance.
The extent to which tuning along a best direction in space could account for the modulation in the activity of SI neurons did not appear to be uniform across areas. In areas 3b and 1, a single axis accounted for 80% of the total variance in 44 and 45% of the cells, respectively, whereas this propor- tion was much less in area 2 (25%) and in the transition zone between areas 2 and 5 (27%). (For area 3a, in 2 of 5 cells, a single axis accounted for >80% of the variance.)
The analytical procedure that we used to fit the data has an implicit rank ordering. One would expect the first axis to contribute most of the variance, the second axis a lesser amount, and the third axis even less. (Under ideal conditions, the propor- tion of the variance accounted for by the 1st axis should be > l/3, and that for the 2nd axis <0.5. As can be seen in Fig. 8, this condition was not always satisfied; sometimes a single axis did not give an adequate fit to the data.) The question arises: to what extent can the distributions shown in Fig. 8 arise from a random process of tuning in three dimensions, assuming a strict rank ordering? In other words, is the finding that the tuning of many cells can be characterized by a best

POSTURE-RELATED ACTIVITY IN SI 243 1
-15 -10 -5 0 5 10 -25 -20 -15 -10 -5 0
.06x -.91y -.41z -.72x - .05y - .7Oz
liO052 . l
. . . . 8 l
l . .*
* . . . . .
. . . 0. .
. l a. l l . l
. . . . . .
. . . . .
. .
25 -
20 -
15 -
10 -
5 -
0 -
-.33x -.94y -.08z -.64x + .76y - .09z
FIG. 6. Discharge rate as plotted against hand location along the neuron’s “best direction.” The same data shown in Fig. 4 are plotted as a function of hand location along the best-fit axis for that neuron. The extent of scatter of data points indicates the extent to which each neuron’s activity was modulated by hand displacement perpendicular to this axis. Activity in neurons frO100 and ZiOO.52 depended principally on hand displacement along a single axis, whereas activity in the other 2 neurons (fro1 11 and frO07.5) exhibited considerable variation unrelated to hand location along the best-fit axis. Discharge rate is stated in spikes/s.
direction and that of virtually all cells by a “modulation plane’ ’ a result of a chance process, or does it reflect neural processing to organize information about limb posture in a particular manner?
To answer this question, we performed a simple simula- tion. Fifteen hundred random numbers uniformly distributed over the interval (0,l ) were generated in groups of three. Each triplet of numbers was then normalized so that its sum of squares was equal to 1, and the three resulting numbers were ordered from largest to smallest. This gave three distri- butions of 500 numbers each. The distributions of the squares of these numbers are plotted as lines in Fig. 8. These are the distributions one would expect for the variance accounted for by each of the three axes if those distributions were generated by a completely random process. According to this simulation, one would expect a single axis to account for 90% of the variance in only 9% of the cells, and 80% of the variance in 20% of the cells. Two axes would suffice in 65% of the cases to account for 90% of the variance, and in 86% of the cases they would account for 80% of the variance. Thus the finding that the third axis generally con- tributed <20% of the variance is to be expected by chance. However, this simulation indicates that many more cells were tuned to hand displacement along a single axis than expected by chance. Compared against the Monte Carlo re-
sults, the first axis in the experimental data accounted for more variance than predicted by a random process (Kolmo- gorov-Smirnov test for the difference between the distribu- tions shown in Fig. 8, top, D = 0.228, IV = 68, P < 0.05). According to the same criterion, the second and third axes contributed less to the overall variance of the data.
Spatial distribution of best directions and modulation planes
The tuning of the population of cells we recorded from was not uniformly distributed in space. More cells had activ- ity that was modulated according to the location of the hand along the anteroposterior axis than along either the vertical or mediolateral axes. Figure 9 shows the distribution of the best directions for the cells, projected onto the vertical plane at the top and onto the horizontal plane at the bottom. There is a preponderance of cells whose preferred direction was close to horizontal (top) and in the anteroposterior direction (bottom). The number of cells that had maximal discharge rates at distal hand locations and at proximal hand locations was about equal. No clear distinctions in the tuning of cells in different cortical areas are obvious from inspection of Fig. 9.
Because the modulation of only a fraction of the cells

2432 S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
_s-- _*_.--
. . . - l .
- .
z - . - - . l . - . l . - .
- . l . si - .
s __-- . .
__-- - . . . .
_ . - -
: - . .
_ . - - ( --. ’
liOO52 -.-- _--- _--- ’ --._ _.-- : --.-
_a--- --._
FIG. 7. Modulation planes. Data from the same 4 neurons are now plotted along the best-fit plane. The best-fit axis is plotted going to the right, and the 2nd axis extends back and to the left. Neurons frOlO0 and ZiOO.52 (left) show very little dependence on location along the 2nd axis, whereas neurons fro111 and fvO07.5 (right) exhibit significant modulation along the 2nd axis. Discharge rate is stated in spikes/s.
could be characterized by their tuning along a single spatial axis ( see Fig. 8 ) , we also took into account the fact that some cells could best be described as being modulated in a plane. The equal-area projection in Fig. 10 takes this fact into account. In this plot we illustrate the percentage of cells whose discharge would be modulated as the hand is
’ laced (quasistatically) in a particular direction in space. rice again, it is clear that there is a maximum for forwardly
for backwardly directed displacements of the hand (79%)) and minima for displacements of the hand in the mediolateral (56% ) and vertical (57% ) directions. To deter- rnine the statistical significance of this difference, we per- formed a bootstrap simulation, sampling at random from our data set with substitution. For the anterior direction, the 95% confidence limits were 86% and 72% and did not overlap those for the other two directions (65% and 64% at the upper limit).
So far, we have shown that the firing rate of SI neurons can be rel ated to the locati on of the hand in space. This does
more plausible, that these neurons actually encode parame- ters more closely related to the rotation of the elbow and shoulder joints, i.e., parameters that are more closely related to the sensitivity of the mechanoreceptors. Joint rotation angles are uniquely and monotonically related to the location of the hand in space (Flanders et al. 1992; Helms Tillery et al. 1995; Lacquaniti et al. 1995; Soechting et al. 1995) at least for a particular task. It is possible that the neuronal discharge could be described in a simpler manner by relating it directly to joint angles rather than hand location. This would constitute evidence that these neurons encode arm posture rather than hand location.
Therefore we also computed the relation between neural discharge and joint angles for each of the modulated neurons, but we did not find that this description was any simpler than the description we have provided above. In some neurons, a simple relation can be seen between one or another joint angle and static discharge rate, but this observation did not generalize to all the neurons. As a first step in the analysis, we tested whether some neurons were preferentially tuned to shoulder rotation and others to rotation at the elbow joint. To this end, we fitted the data to a cubic polynomial in
not imply, however, that hand location in space is actually elbow flexion (Es. 8) and computed the variance accounted encoded by these neurons. It is equally possible, and perhaps for by this model as a fraction of the variance accounted for
POSTURE-RELATED ACTIVITY IN SI 2433
0.0 0.2 0.4 0.6 0.8 1.0
0.0 0.2 0.4 0.6 0.8 1.0
80
Third Axis
40
0
0.0 0.2 0.4 0.6 0.8 1.0
Variance Accounted for
FIG. 8. Percent of total variance accounted for by each of 3 spatial axes. Each histogram shows the proportion of the total variance that could be accounted for by incorporating the component of hand location along that axis in the linear regression. The distribution for the 1st axis is highly skewed to the right, indicating that the fit along the 1st axis generally accounted for a substantial portion of the variance. The 3rd axis provided little or no improvement to the fit, indicating that the cells’ spatial tuning could be adequately described by the projection of hand location onto a plane. Lines: theoretical distributions expected if the 3 T* values are ordered random deviates constrained only to sum to 1.
by fitting neural discharge to hand location. The results of this analysis are shown in Fig. 11. If most of the neurons’ modulation were related to motion at a single joint, one would expect a bimodal distribution, with values clustered about values of 0 and 1. For -35% of the cells elbow rotation accounted for <lo% of the variance. However, for the re- maining cells, the contribution of elbow rotation to the total variance was widely distributed with no evidence of cluster- ing. In general, rotation about both shoulder and elbow joints contributed to the cells’ modulation.
One possibility that could account for neuronal activity being related to both shoulder and elbow rotation is that it encodes arm posture according to a different set of parame- ters. The angles defining the orientation of the lower arm (Soechling and Ross 1984) depend on the rotation of both
the shoulder and elbow joints. More generally, the orienta- tion angles of the upper arm and forearm have proven useful in describing limb kinematics under a variety of experimen- tal conditions (Helms Tillery et al. 1995; Lacquaniti et al. 1995; Soechting and Flanders 1989). To determine whether neuronal activity encodes orientation angles, we first plotted the firing rates against each of these angles. A representative example is illustrated in Fig. 12, with the use of the data for cell frOl00 (see Fig. 4). The discharge of this neuron was related to the yaw angles of both the upper (q) and lower ((Y) arms-increasing with q and decreasing with (Y. This finding is consistent with the results in Fig. 4, showing that the discharge rate of the neuron increased with increasing target azimuth (Helms Tillery et al. 1995; Lacquaniti et al. 1995). However, there is considerable scatter in each of the plots, indicating that the neuron’s discharge depended on more than one of the orientation angles of the arm.
As a next step in the analysis, we fitted the data to a linear combination of orientation angles, determining the best di- rection in joint space (Eq. 9). Finally we fitted the data to cubic polynomials in the four angles. The results of this analysis for the same neurons as in Fig. 4 are illustrated in Fig. 13. The data for the two cells shown at the left are reasonably well correlated to a linear combination of oricnta-
Area
Backward
FIG. 9. Polar distribution of the best directions for the neural population. Top: projection of each of the units’ best direction onto the vertical plane. Bottom: projection onto the horizontal plane. Units are segrated according
to cortical area. Note the preponderance of neurons tuned to displacement of the hand in the anteroposterior direction.

2434 S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
Modulation Distribution
Forw arcl
Backward
q 56%.60%
61% 65%
71% - 75%
H 76%-X0%
FIG. 10. Spatial tuning of neural responses. Sensitivity of the population of neurons to displacements of the hand along different directions is shown on an equal-area projection of the upper hemisphere. This plot takes into account that the discharge of some neurons was modulated by hand dis- placement in a plane. The relative sensitivity of the population to displace- ments along the anteroposterior direction (79%) was greater than the sensi- tivity to displacements along the other 2 cardinal axes (56%).
tion angles. The activity of cell liOO.52 was most strongly related to the yaw angles of the upper arm (coefficient for 17 = 0.41) and forearm (coefficient for (Y = -0.89). This is consistent with the observation that this cell’s activity increased as the hand was displaced laterally (cf. Fig. 4), corresponding to an increase in n and a decrease in c~. The activity for cell fro100 (top Zeft) was mostly related to the yaw angle of the upper arm (q), and to a lesser extent to the elevation angles of both the upper arm (0) and forearm (/3). Note that for both the cells at the left, there is consider- able scatter of the data points, indicating that there was also modulation of neuronal activity about joint axes perpendicu- lar to the ones indicated in the plots. The data points for the two cells at the right show a much greater amount of scatter. In fact, a comparison of Fig. 13 with Fig. 6 (illustrating results for the same 4 cells) shows that there is much less scatter of the data points when activity is plotted along a best direction in the Cartesian space of hand displacement than when it is plotted along a best direction in the joint space of the orientation angles.
The activity of our sample of neurons can clearly be de- scribed in terms of joint angles. However, this description is no simpler than when the activity is described in terms of hand location in Cartesian coordinates. Only a small por- tion of the neurons (6 of 78) was related to only one of the orientation angles. In fact, because there are four orientation
angles and only three parameters describing hand location, the description of neural activity in terms of hand location appears to be simpler. However, this should not be taken to imply that the activity of these neurons encodes hand location.
DISCUSSION
Activity of neurons in SI is related monotonically to the static location of the hand in space, generally reaching a maximum at the extremes of the workspace. In some neu- rons, activity was defined by the projection of the hand’s location on a single axis in space (best direction), akin to the code of movement direction that has been described for neurons in other cortical areas (cf. Caminiti et al. 1991; Georgopoulos 1995; Georgopoulos et al. 1988; Kalaska et al. 1990). However, the activity of the majority of the cells in SI was defined by the projection of the hand’s location on a plane (modulation plane). Considered as a population, the sensitivity of these neurons to a quasistatic displacement of the hand in space is not uniform: more neurons are tuned to a displacement of the hand in the anteroposterior direction than to hand displacements in the other two directions. This last finding confirms and extends the findings of Cohen et al. (1994) and of Prud’homme and Kalaska (1994). In the following, we take up the implications of these findings and the similarities and differences in the spatial tuning of neu- rons in SI compared with those in areas 4 and 5.
0.2 0.4 0.6 0.8 1.0
Elbow Fit r2/ Complete Fit r2
FIG. 11. Dependence of neural discharge rate on changes in elbow joint angle. The data for each neuron were fitted to a cubic polynomial in elbow angle, and the Y* for that fit was normalized against the Y’ for a cubic fit to the hand location. Those neurons that were related only to rotations about the elbow would give a value of 1.0 for this measure. Those neurons that were independent of the elbow and responsive only to rotations about the
shoulder would give a value of 0.0. The distribution is skewed toward neurons related to the shoulder (see text), but there are a substantial number
of neurons giving values (>0.2 and <0.8) consistent with sensitivity to rotation about both joints.

Yaw .
POSTURE-RELATED ACTIVITY IN SI 2435
Elevation
25 1 E 2o . 2 15 -
8 10 -
&5-
0 ’ . .
0” 50” 100”
0. l . .
. . . . .
. l .
. l l .
.
:
. . 0. .s* . .
. . r & : : - . . e
. . . ! * .
. . . . * . . . * . . I I 1 I
. . l
l . l
: . *
. . l
. l * l .
a l . :* . . .* . . . l . . . l . l l
8 . . : .A
.
. * l .
-20° O0 20" 4o" 20' 40" 60" 80' loo0 FIG. 12. Discharge rate as a function of segment orientation angles. The discharge rate of neuron frM00 is plotted as a
function of the segment orientation angles. This neuron exhibited a monotonic relation between discharge rate and the yaw angles of both the upper and lower arm segments, but the scatter in the plots indicates this neuron’s discharge depended on the elevation angles as well. Discharge rate is stated in spikes/s.
Spatial tuning
Our observation that the relations between discharge rate and target location tended to be monotonic agrees with previ- ous findings in both somatosensory thalamus (Mountcastle et al. 1963) and SI (Cohen et al. 1994; Gardner and Costanzo 198 1; Mountcastle and Powell 1959) as well as peripheral afferents (Edin and Abbs 199 1; Hulliger et al. 1979). In each of these reports, the discharge of tonically active neurons modulated monotonically with increasing joint angles. This held true whether the neurons had receptive fields on the finger, wrist, elbow, or shoulder joints. The joint angles of the shoulder and elbow are monotonically related to hand location (Helms Tillery et al. 1995; Lacquaniti et al. 1995; Soechting and Flanders 1989). Therefore one could expect the discharge of SI neurons to vary monotonically with hand position.
For the majority of cells, activity could not be character- ized in terms of a best direction, either in the Cartesian space of hand location or in the four-dimensional space of joint angles. In this respect our conclusions differ from those of Cohen et al. (1994) and of Prud’homme and Kalaska ( 1994), who found that a cosine tuning of cells in SI pro- vided an adequate description of their results both during active movements and during the maintenance of a static posture. However, the difference between their results and ours is more apparent than real. They examined movements to only eight targets in a plane, with the use of repeated measures, whereas we examined posture at locations distrib- uted randomly throughout the workspace. Furthermore, they restricted their analysis only to that portion of cells (-60- 70%) that was fitted adequately by a sinusoid. One might expect the characterization of a cell’s spatial tuning to un- cover a more complex pattern if one were to examine a
larger range of target locations (Caminiti et al. 199 1; Fu et al. 1993, 1995).
Activity in a number of other cortical areas such as precen- tral area 4 (Georgopoulos et al. 1986; Schwartz et al. 1988) and postcentral area 5 (Kalaska et al. 1990) as well as the cerebellum (Fortier et al. 1989) has been described as being cosine tuned. Before and during movement, neurons in these areas discharge at a rate that is related to the angle between the direction in which the hand is about to move and the neuron’s best direction. A similar result has been shown to hold in these same areas for discharge occurring while the hand is held at various locations in a two-dimensional work- space (Fortier et al. 1989; Georgopoulos and Massey 1985; Kettner et al. 1988).
The question of such a vector code of movement direction in area 5 has recently been reexamined by Lacquaniti et al. ( 1995). Those researchers examined neural activity during movements from a central starting position to eight targets located at the vertices of a cube and found that a vector code could adequately characterize neural activity. However, when the starting location was shifted medially or laterally, the best direction of most neurons was found to depend on the starting position. Testing a variety of other coding schemes for these neurons, these investigators found that one based on distance and direction (elevation and azimuth) from the shoulder (Flanders et al. 1992) provided the most parsimonious description. (There is a unique relation be- tween elbow angle and distance from the shoulder, and Lac- quaniti et al. used the elbow angle rather than distance in their model.) Their study reveals similarities and differences between the spatial tuning of neurons in SI and in area 5. They found a tendency for clustering of neural tuning along one of the three coordinate axes (elbow angle, elevation,

2436 S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
30 -
20 -
10 -
0 d*
fro100 . . . -t . l *. . .
. .
40 60 80 100 120 -80 -60 -40 -20
.81q + .408 - .06a + 0.4lf3 -.Olq - .13e + .80a + .59p
20 - . . . liO052 l
. l
. . . 15 -
. . * . : . . . . . . . l * . l .
lo- l
t l . .
. .
:
. .
. .
. .
5 8 ii l : l , ; . : : - ’ : l
.
25 -
20 - 15 -
10 -
5 -
0 *
fro075
. .
-20 0 20 40 60 -90 -80 -70 -60
.41q - ,030 -.89a + .21f3 -.48q -.638 - .46a - .4lC3
FIG. 13. Best-fit axes in joint space. Data for the same neurons as shown in Fig. 4 are plotted along the best-fit axis in joint space. Scatter of the data points indicates that neural activity was modulated by hand displacements along axes that were perpendicular to the one indicated in each panel.
and azimuth), whereas we were unable to find evidence for such clustering in SI. More neurons were tuned along the elbow angle axis than the other two axes, both under static and dynamic conditions. This finding reflects another differ- ence between the two neural populations: we found few if any neurons that were tuned predominately to the elbow angle (Fig. 11). However, there is one fundamental similar- ity between the conclusions of Lacquaniti et al. and the results we have presented here: neurons in both cortical areas appear to encode the location of the hand in space, although it is undetermined whether that is in terms of the spherangu- lar coordinates used by Lacquaniti et al. ( 1995), the Cartesian coordinates used here, or shoulder and elbow joint angles. (There is a unique relationship between each of the three sets of parameters, at least for any one particular task.)
Because SI provides information to both areas 4 and 5, it is relevant to consider the differences and similarities in the kinematic spatial tuning of cells in these areas compared with those in SI. Most of the studies that have ascribed cosine tuning to motor cortical neurons have also used only a limited number of movement directions, and it is possible that if one were to use a greater number of target locations, a single best direction would inadequately characterize the activity of motor cortical neurons. However, the studies of Schwartz ( 1992, 1994) suggest that a vector code of move- ment direction may in fact characterize motor cortical neu- ronal activity, because he was able to predict the hand’s trajectory during sinusoidal drawing movements (i.e., a task comprising an infinite number of movement directions) from neuronal activity on the basis of a population vector code (Georgopoulos et al. 1983). Nevertheless, it may be useful to reexamine this issue by using the same experimental para- digms and analysis techniques in the study of pre- and post- central cortical areas.
Nonuniform spatial tuning
We found that neurons in SI were preferentially sensitive to changes in the location of the hand in the anteroposterior direction. This asymmetry in tuning was statistically signifi- cant. It is also consonant with the observations of Cohen et al. (1994) and Prud’homme and Kalaska (1994). Here we take up two issues that are pertinent to this topic: I) the possible origin of this asymmetry and 2) a comparison of the distribution of spatial tuning of neurons in SI and in areas 4 and 5.
Sampling bias is one possible source of the spatial asym- metries in tuning that we found. We recorded from neurons in a large portion of the proximal arm region of areas 3b, 1, and 2 (Fig. 1). Nevertheless, it is possible that areas contiguous to those we recorded from could contain neurons that are preferentially tuned to displacements of the hand in the mediolateral and vertical directions. This would imply that neurons tuned to different movement directions are dis- tributed inhomogeneously in SI. At present there is no evi- dence in support of this possibility, either in SI or in other precentral or postcentral areas. As noted in RESULTS, we did not record from the more medial strip described by Pons et al. ( 1985a,b), and it is possible that neurons in that region would exhibit spatial tuning different from what we de- scribed. We also recorded from only a few cells in area 3a, and it is possible that our conclusions do not hold true for this region of SI.
Another explanation is anatomic: peripheral somatosen- sors may be more sensitive to displacements of the hand in the fore-aft direction than to displacements in other direc- tions. Thus, if a displacement of the hand by 1 cm in the anteroposterior direction led to a larger change in shoulder and elbow angles than did a l-cm displacement of the hand

POSTURE-RELATED ACTIVITY IN SI 2437
in other directions, then one might expect preferential tuning, EDIN, B. B. Quantitative analysis of static strain sensitivity in human mecha-
expressed in Cartesian coordinates, even if the population noreceptors from hairy skin. J. Neurophysiol. 67: 1105 - 1113, 1992.
of neurons were equally sensitive to joint excursions in all EDIN, B. B. AND ABBS, J. H. Finger movement responses of cutaneous
directions. This explanation appears unlikely, because there mechanoreceptors in the dorsal skin of the human hand. J. Neurophysiol. 65: 657-670, 1991.
is no bias in the sensitivity of the joint angles toward the EDIN, B. B. AND JOHANSSON, N. Skin strain patterns provide kinaesthetic
anterior direction. In fact, the two yaw angles (7 and a) are information to the human central nervous system. J. Physiol. Lund. 487:
biased toward the mediolateral direction, changing maxi- 243-251, 1995.
mally for movements directed -45’ and 65” away from FLANDERS, M., HELMS TILL~ERY, S. I., AND SOECHTING, J. F. Early stages
in a sensorimotor transformation. Behav. Brain Sci. 15: 309-362. 1992. the anterior direction ( see also Helms Tillery et al. 1995; FORTIER, P. A., KALASKA, J. F., AND SMITH, A. M. Cerebellar neuronal
Lacquaniti et al. 1995 ) . In the horizontal plane, the two activity related to whole-arm reaching movements in the monkey. J.
elevation angles (7 and p) are tuned within a few degrees Neurophysiol. 62: 198-211, 1989.
of the anterior direction, but their maximum sensitivity in FROMM, C. AND EVARTS, E. V. Pyramidal tract neurons in somatosensory
the vertical plane is to hand displacements 40-45” from cortex: central and peripheral inputs during voluntary movement. Brain Res. 238: 186-191, 1982.
horizontal. Fu, Q.-C., FLAMENT, D., COLTZ, J. D., AND EBNER, T. J. Temporal encoding
We have already mentioned that our data are in agreement with observations of Cohen et al. ( 1994) and of Prud’homme and Kalaska ( 1994), who also found that the preferred direc- tions of SI neurons were biased in the anterolateral direction both during active movements and the maintenance of static postures. There appears to be a similar bias for the spatial tuning in area 5. Kalaska et al. ( 1983) found a preference for movements that were directed either anteriorly or toward the left, although, interestingly, the same group did not find such an asymmetry under static conditions (Georgopoulos et al. 1984). By contrast, the distribution of best directions in area 4 appears to be uniform (Schwartz et al. 1988).
In summary, one can suggest that neuronal activity in areas 3, 1, and 2 resembles more closely that found in area 5 than in area 4 in two ways: 1) there is a positional code of hand location rather than a vector code of movement direction and 2) the sensitivity of the neural activity to changes in hand location does not appear to be uniform for all directions in space. There is one more important distinc- tion between neurons in area 5 and area 4 that this study has not addressed: neurons in area 5 appear to be related primarily to kinematic parameters, whereas neurons in motor cortical areas appear to be influenced as well by parameters related to movement kinetics (Georgopoulos et al. 1992; Kalaska et al. 1990). In this respect as well, it appears the properties of SI neurons more closely resemble those in area 5 (Helms Tillery 1994; see also Fromm and Evarts 1982; Prud’homme and Kalaska 1994; Soso and Fetz 1980).
This work was supported by National Institute of Neurological Disorders
and Stroke Grants NS-15018 and NS-31530, a Human Frontiers in Science Program grant, and a National Science Foundation Graduate Fellowship to S. I. Helms Tillery.
Present address of S. I. Helms Tillery: Research Services ( 15 1) , VA Medical Ctr., Syracuse, NY 13210.
Address for reprint requests: J. F. Soechting, Dept. of Physiology, 6-255 Millard Hall, University of Minnesota, Minneapolis, MN 55455.
Received 9 January 1995; accepted in final form 24 May 1996.
of movement kinematics in the discharge of primate primary motor and premotor neurons. J. Neurophysiol. 73: 836-854, 1995.
Fu, Q.-C., SUAREZ, J. I., AND EBNER, T. J. Neuronal specification of direc- tion and distance during reaching movements in the superior precentral premotor area and primary motor cortex of monkeys. J. Neurophysiol. 70: 2097-2116, 1993.
GARDNER, E. P. Somatosensory cortical mechanisms of feature detection in tactile and kinesthetic discrimination. Can. J. Physiol. Pharmacol. 66: 439-454, 1988.
GARDNER, E. P. AND COSTANZO, R. M. Properties of kinesthetic neurons in somatosensory cortex of awake monkeys. Brain Res. 214: 301-3 19, 1981.
GEORGOPOULOS, A. P. Current issues in directional motor control. Trends Neurosci. 18: 506-510, 1995.
GEORGOPOULOS, A. P., ASHE, J., SMYRNID, N., AND TAIRA, M. Motor cortex
and the coding of force. Science Wash. DC 256: 1692-1695, 1992. GEORGOPOULOS, A. P., CAMINITI, R., AND KALASKA, J. F. Static spatial
effects on motor cortex and area 5: quantitative relations in a two-dimen- sional space. Exp. Brain Res. 54: 446--454, 1984.
GEORGOPOULOS, A. P., CAMINITI, R., KALASKA, J. F., AND MASSEY,
J. T. Spatial coding of movement: a hypothesis concerning the coding
of movement direction by motor cortical populations. Exp. Brain Res. Suppl. 7: 327-336, 1983.
GEORGOPOULOS, A. P., KETTNER, R. E., AND SCHWARTZ, A. B. Primate motor cortex and free arm movements to visual targets in three-dimen- sional space. II. Coding of the direction of movement by a neuronal population. J. Neurosci. 8: 2928-2937, 1988.
GEORGOPOULOS, A. P. AND MASSEY, J. T. Static versus dynamic effects in motor cortex and area 5: comparison during movement time. Behav.
Brain Res. 18: 159-166, 1985. GEORGOPOULOS, A. P., SCHWARTZ, A. B., AND KETTNER, R. E. Neuronal
population coding of movement direction. Science Wash. DC 233: 1416- 1419, 1986.
GHEZ, C., GORDON, J., GHILARDI, M. F., CHRISTAKOS, C. N., AND COOPER,
S. E. Roles of proprioceptive input in the programming of arm trajector- ies. Cold Spring Harbor Syrnp. Quant. Biol. 55: 837-847, 1990.
GOODWIN, G. M., MCCLOSKEY, D. I., AND MATTHEWS, P. B. C. The contri-
bution of muscle afferents to kinesthesia shown by vibration induced illusions of movement and by the effects of paralysing joint afferents. Brain 95: 705-748, 1972.
HASAN, 2. Role of proprioceptors in neural control. Curr. Opin. Neurobiol. 2: 824-829, 1992.
HASAN, Z. AND STUART, D. G. Animal solutions to problems of movement control: the role of proprioceptors. Annu. Rev. Neurosci. 11: 199-223, 1988.
REFERENCES
HELMS TILLERY, S. I. Encoding of Arm Posture in Sornatosensory Cortex (PhD thesis). Minneapolis, MN: Univ. of Minnesota, 1994.
HELMS TILLERY, S. I., EBNER, T. J., AND SOECHTING, J. F. Differential
CAMINITI, R., JOHNSON, P. B., GALLI, C., FERRAINA, S., AND BURNOD, Y. processing of vertical and horizontal components of arm posture in SI.
Making arm movements within different parts of space: the premotor Sot. Neurosci. Abstr. 20: 755, 1994.
and motor cortical representations of a coordinate system for reaching HELMS TILLERY, S. I., EBNER, T. J., AND SOECHTING, J. F. Task dependence
to visual targets. J. Neurosci. 11: 1182- 1197, 199 1. of primate arm postures. Exp. Brain Res. 104: 1 - 11, 1995.
COHEN, D. A. D., PRUD’HOMME, M. J. L., AND KALASKA, J. F. Tactile HELMS TILLERY, S. I., FLANDERS, M., AND SOECHTING, J. F. A coordinate
activity in primate somatosensory cortex during active arm movements: system for the synthesis of visual and kinesthetic information. J. Neu-
correlation with receptive field properties. J. Neurophysiol. 7 1: 16 1 - 172, rosci. 11: 770-780, 1991.
1994. HELMS TILLERY, S. I., SOECHTING, J. F., AND EBNER, T. J. Discharge of COSTANZO, R. M. AND GARDNER, E. P. Multiple-joint neurons in somatosen- parietal neurons during static postures. I. Force dependence. Sot. Neu-
rosci. Abstr. 18: 503, 1992. sory cortex of awake monkeys. Brain Res. 214: 321-333, 1981.

2438 S. I. HELMS TILLERY, J. F. SOECHTING, AND T. J. EBNER
HULLIGER, M., NORDH, E., THELIN, A.-E., AND VALLBO, A. The responses of afferent fibres from the glabrous skin of the hand during voluntary finger movements in man. J. Physiol. Lond. 291: 233-249, 1979.
JENNINGS, V. A., LAMOUR, Y., SOLIS, H., AND FROMM, C. Somatosensory cortex activity related to position and force. J. Neurophysiol. 49: 1216- 1229, 1983.
JONES, E. G., COULTER, J. D., AND HENDRY, S. H. C. Intracortical connectiv- ity of architectonic fields in the somatic sensory, motor and parietal cortex of monkeys. J. Comp. Neurol. 181: 291-348, 1978.
KALASKA, J. F. What parameters of reaching are encoded by discharges of cortical cells? In: Motor Control: Concepts and Issues, edited by D. R. Humphrey and H.-J. Freund. Chichester, UK: Wiley-Interscience, 1991, p. 307-330.
KALASKA, J. F., CAMINITI, R., AND GEORGOPOULOS, A. P. Cortical mecha- nisms related to the direction of two-dimensional arm movements: rela- tions in parietal area 5 and comparison with motor cortex. Exp. Brain Res. 51: 247-260, 1983.
KALASKA, J. F., COHEN, D. A. D., PRUD’HOMME, M., AND HYDE, M. L. Parietal area 5 neuronal activity encodes movement kinematics, not movement dynamics. Exp. Brain Res. 80: 351-364, 1990.
KETTNER, R. E., SCHWARTZ, A. B., AND GEORGOPOULOS, A. P. Primate motor cortex and free arm movements to visual targets in three-dimen- sional space. III. Positional gradients and population coding of movement direction from various movement origins. J. Neurosci. 8: 2938-2947, 1988.
KNAPP, H. D., TAUB, E., AND BERMAN, J. Movements in monkeys with deafferented forelimbs. Exp. Neural. 7: 305 -3 15, 1963.
KNIBESTOL, M. AND VALLBO, A. B. Single unit analysis of mechanoreceptor activity from the human glabrous skin. Acta Physiol. Stand. 79: 17% 195, 1970.
LACQUANITI, F., GUIGNON, E., BIANCHI, L., FERRAINA, S., AND CAMINITI,
R. Representing spatial information for limb movement: role of area 5 in the monkey. Cereb. Cortex 5: 391-409, 1995.
MCCLOSKEY, D. I. Kinesthetic sensibility. Physiol. Rev. 58: 763-820, 1978.
MILLAR, J. Loci of joint cells in the cuneate and external cuneate nuclei of the cat. Brain Res. 167: 385-390, 1972.
MOUNTCASTLE, V. B., POGGIO, G. F., AND WERNER, G. The relation of thalamic cell response to peripheral stimuli varied over an intensive con- tinuum. J. Neurophysiol. 26: 807-834, 1963.
MOUNTCASTLE, V. B. AND POWELL, T. P. S. Central nervous mechanisms subserving position sense and kinesthesis. Bull. Johns Hopkins Hosp.
105: 173-200, 1959. POGGIO, G. F. AND MOUNTCASTLE, V. B. The functional properties of ven-
trobasal thalamic neurons studied in unanesthetized monkeys. J. Neuro- physiol. 26: 775 - 806, 1963.
POLIT, A. AND BIZZI, E. Characteristics of motor programs underlying arm movements in monkeys. J. Neurophysiol. 42: 183- 194, 1979.
PONS, T. P., GARRAGHTY, P. E., CUSICK, C. G., AND KAAS, J. H. A sequential representation of the occiput, arm, forearm and hand across the rostrocau- da1 dimension of areas 1, 2 and 5 in macaque monkeys. Brain Res. 335: 350-353, 1985a.
PONS, T. P., GARRRAGHTY, P. E., CUSICK, C. G., AND KAAS, J. H. The somatotopic organization of area 2 in macaque monkeys. J. Comp. Neu- rol. 241: 445-466, 1985b.
POWELL, T. P. S. AND MOUNTCASTLE, V. B. The cytoarchitecture of the postcentral gyrus of the monkey Macaca mulatta. Bull. Johns Hopkins Hosp. 105: 108-131, 1959.
PRESS, W. H., FLANNERY, B. P., TEUKOLSKY, S. A., AND VETTERLING,
W. T. Numerical Recipes in C: The Art of Scientific Computing. Cam- bridge, MA: Cambridge Press, 1988.
PRUD’HOMME, M. J. L., COHEN, D. A. D., AND KALASKA, J. F. Tactile activity in primary somatosensory cortex during active arm movements: cytoarchitectonic distribution. J. Neurophysiol. 7 1: 173 - 18 1, 1994.
PRUD’HOMME, M. J. L. AND KALASKA, J. F. Proprioceptive activity in primate primary somatosensory cortex during active arm reaching move- ments. J. Neurophysiol. 72: 2280-2301, 1994.
SAINBURG, R. L., POIZNER, H., AND GHEZ, C. Loss of proprioception pro- duces deficits in interjoint coordination. J. Neurophysiol. 70: 2 136-2147, 1993.
SCHWARTZ, A. B. Motor cortical activity during drawing movements: single-
unit activity during sinusoid tracing. J. Neurophysiol. 68: 52%-541, 1992. SCHWARTZ, A. B. Direct cortical representation of drawing. Science Wash.
DC 265: 540-542, 1994. SCHWARTZ, A. B., KETTNER, R. E., AND GEORGOPOULOS, A. P. Primate
motor cortex and free arm movements to visual targets in three-dimen- sional space. I. Relations between single cell discharge and direction of movement. J. Neurosci. 8: 2913-2927, 1988.
SCOTT, S. H. AND KALASKA, J. F. Changes in motor cortex activity during reaching movements with similar hand paths but different postures. J. Neurophysiol. 73: 2563-2567, 1995.
SNIDER, R. S. AND LEE, J. C. A Stereotaxic Atlas of the Monkey Brain (Macaca mulatta). Chicago, IL: Univ. of Chicago Press, 1961.
SOECHTING, J. F., BUNEO, C. A., HERRMANN, U., AND FLANDERS, M. Moving effortlessly in three dimensions: Does Donders’ law apply to arm move- ment? J. Neurosci. 15: 6271-6280, 1995.
SOECHTING, J. F. AND FLANDERS, M. Errors in pointing are due to approxi- mations in sensorimotor transformations. J. Neurophysiol. 62: 595-608, 1989.
SOECHTING, J. F., HELMS TILLERY, S. I., AND EBNER, T. J. Discharge of parietal neurons during static postures. II. Relation to postural parameters. Sot. Neurosci. Abstr. 18: 503, 1992.
SOECHTING, J. F. AND Ross, B. Psychophysical determination of coordinate representation of human arm orientation. Neuroscience 13: 595 -604, 1984.
Soso, M. J. AND FETZ, E. E. Responses of identified cells in postcentral cortex of awake monkeys during comparable active and passive joint movements. J. Neurophysiol. 43: 1090- 1110, 1980.
SURMEIER, D. J. AND TOWE, A. L. Properties of proprioceptive neurons in the cuneate nucleus of the cat. J. Neurophysiol. 57: 938-961, 1987.
TAUB, E., GOLDBERG, I. A., AND TAUB, P. Deafferentation in monkeys: pointing at a target without visual feedback. Exp. Neural. 46: 178- 186, 1975.
WATSON, G. S. Statistics on Spheres. New York: Wiley-Interscience, 1983.