snp e4-205 c/t in c-type lectin of portunus ...download.xuebalib.com/xuebalib.com.33457.pdf · with...
TRANSCRIPT

Aquaculture 442 (2015) 125–131
Contents lists available at ScienceDirect
Aquaculture
j ourna l homepage: www.e lsev ie r .com/ locate /aqua-on l ine
SNP E4-205 C/T in C-type lectin of Portunus trituberculatus is associationwith susceptibility/resistance to Vibrio alginolyticus challenge
Guijie Hao a,b, Feng Lin b, Changkao Mu a, Ronghua Li a, Jiayun Yao b, Xuemei Yuan b, Xiaoyi Pan b,Jinyu Shen b,⁎, Chunlin Wang a,⁎⁎a School of Marine Science, Ningbo University, Zhejiang, Ningbo 315211, PR Chinab Zhejiang Institute of Freshwater Fisheries, Key Laboratory of Fish Health and Nutrition of Zhejiang Province, Zhejiang, Huzhou 313001, PR China
⁎ Corresponding author. Tel./fax: +86 572 204 3927.⁎⁎ Corresponding author. Tel./fax: +86 574 876 00356.
E-mail addresses: [email protected] (J. Shen), wang
http://dx.doi.org/10.1016/j.aquaculture.2015.02.0070044-8486/© 2015 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 17 December 2014Received in revised form 4 February 2015Accepted 6 February 2015Available online 14 February 2015
Keywords:Portunus trituberculatusC-type lectinPolymorphismSusceptibility/resistanceHigh resolution melting (HRM) analysis
C-type lectins play an important role in innate immunity of invertebrates, especially in defending them againstbacterial pathogens. In this study, the full-length genomic DNA of C-type lectin (designated as CTL gene) was iso-lated from the swimming crab Portunus trituberculatus. The genomic DNA of CTL consists of 1473 bp, containingfour exons and three introns. Polymorphisms of the CTL gene were identified to explore their association withsusceptibility/resistance to Vibrio alginolyticus infection. Four sites of single nucleotide polymorphisms (SNPs)and one site of insertion/deletion (ins/del) polymorphismwere identified in CTL. Three of these SNPs were syn-onymous and one was non-synonymous. The distribution of these polymorphisms in the susceptible and resis-tant stocks was identified, according to the survival time after V. alginolyticus challenge. The non-synonymousSNP E4-205 C/T, a dimorphism caused from C to T transition that resulted in a Threonine to Isoleucine substitu-tion at position 152 in the peptide of CTL protein, showed significant difference between the two stocks accordingto Chi-squared test (P b 0.05). The SNP site has three genotypes that are C/T, C/C and T/T. The T/T genotypefrequency was 6.1% in susceptible stock, and 21.2% in resistant stock, with a significant difference betweenthem (P = 0.022). Therefore, the T/T genotype was suggested to be associated with increased resistance toV. alginolyticus (P b 0.05). The presumption was confirmed by another challenge experiment, in which high res-olution melting (HRM) analysis was used to analyze the three genotypes distribution. The cumulative mortalityof genotype T/T (57.1%) was significantly lower than C/C (82%) and T/C (81.4%) (P = 0.001). These resultssuggested that the E4-205 genotype T/T is a potential marker for selection of swimming crabs resistant toV. Alginolyticus challenge.
© 2015 Elsevier B.V. All rights reserved.
1. Introduction
Portunus trituberculatus is widely distributed along the coast ofChina, Japan, Korea, and other Southeast Asian countries. This speciesis one of the most common edible marine crabs found in China, andhas also been artificially cultured since the 1990s. However, the uncon-trolled expansion in the scale of farming, increase in stocking density,and the polluted breeding environment, all result in the frequent out-breaks of diseases. Someof themost recurrent of these are the infectionscaused by the Vibrio family, such as Vibrio metschnikovii (Wan et al.,2011), and Vibrio alginolyticus (Liu et al., 2007). The “emulsificationdisease” caused by V. alginolyticus has brought considerable economiclosses, and has seriously restricted the sustained and healthy develop-ment of crab aquaculture (Wang et al., 2006). Select breeding of new
[email protected] (C. Wang).
strains resistant to V. alginolyticus is a very important step towardscontrolling this disease.
Traditional breeding techniques are expensive, time-consuming,and easily influenced by the environment (Collard et al., 2005). One ofthe methods that could be used to improve breeding strategies ismarker assisted selection (MAS), a molecular method successfullyused in the improvement of agricultural population (Dekkers andHospital, 2002). It is a method where a marker is used for the indirectselection of the target trait. The marker used is closely linked to thetarget trait. Some studies have reported on the use of DNA markers inMAS of livestock in order to cultivate novel disease-resistant varieties(Maillard et al., 2003; Masoudi et al., 2007). However, the use of MASon P. trituberculatus is strategically difficult, as there are very fewmarkers associated with specific traits (Liu et al., 2012; Lee et al.,2013). Therefore, it is important to identify the marker associatedwith resistance toV. alginolyticus in the swimming crab to develop strat-egies for MAS.
Immune-related genes are considered as the optimal candidates forthe selection of markers associated with resistance to pathogens. Gene

126 G. Hao et al. / Aquaculture 442 (2015) 125–131
polymorphism could improve the genetic quality or quantity, enhanc-ing the ability of the immune system in protecting itself against infec-tion. In recent years, polymorphisms in some immune-related geneshave been reported to affect pathogen or disease resistance in aquaticanimals. Examples of such genes include the lysozyme gene in Zhikongscallop (Chlamys farreri) (Li et al., 2009), the superoxide dismutase genefamily in the bay scallop (Argopecten irradians) (Bao et al., 2010), theserine protease inhibitor gene in the eastern oyster (Crassostreavirginica Gmelin) (He et al., 2012; Yu et al., 2011), the MDA5 gene ingrass carp (Wang et al., 2012), and the mannose binding lectin (MBL)in Zebra fish (Danio rerio) (Jackson et al., 2007). Polymorphism inthese immune genes yielded different genotypes, which exhibitedimmunity phenotypes. Many genes involved in immune response ofswimming crab, such as the C-type lectin (Kong et al., 2008), C-type ly-sozyme (Pan et al., 2010), crustins (Yue et al., 2010), prophenoloxidase(Chen et al., 2010) and anti-lipopolysaccharide (Y. Liu et al., 2011,2012), were reported. However, polymorphisms in these genes, andthe correlation between these gene polymorphisms and diseasesusceptibility and resistance are not yet well understood (Li et al.,2013).
The C-type lectins play an important role in pattern recognition andinnate immune response in invertebrates (McGreal et al., 2004). Theygenerally contain one or more carbohydrate recognition domains. Inthe presence of Ca2+, they specifically recognize the oligosaccharidesexpressed on the cell surface of exogenousmicrobes, and initiate an im-mune reaction against the invasion of pathogenic microorganisms(Wang and wang, 2013). A number of C-type lectins have been identi-fied in crustaceans, most of which display anti-bacterial and anti-viralproperties (Huang et al., 2014; Wang et al., 2013; Wei et al., 2012). InP. trituberculatus, the cDNA of a C-type lectin-like domain (CTLD)-containing protein with one carbohydrate recognition domain (CRD)was cloned and designated as PtLP (CTLD-containing protein) (Konget al., 2008). In our previous study, a high level of PtLP transcripts wasfound in the hemocytes and hepatopancreas of P. trituberculatus follow-ing injection of V. alginolyticus. To elucidate the role of PtLP in suscepti-bility or resistance of P. trituberculatus to V. alginolyticus, the completeDNA sequence of PtLP was cloned, designated as CTL. Our objectivewas to identify polymorphisms in CTL and to investigate the possibleassociation of these polymorphisms with susceptibility/resistance toinfection by the V. alginolyticus.
Table 1Primers used in this study.
Name Sequence (5′-3′) PCR objective
M13-47 CGCCAGGGTTTTCCCAGTCACGAC SequencingRV-M GAGCGGATAACAATTTCACACAGG SequencingCTL-F1 TCCTGTTCTGACAACCACCAA Obtaining DNA sequenceCTL-R1 CTGTGCCCGAGTCAAGAAGTA Obtaining DNA sequenceCTL-F2 AGCAGCACCGATAAAAGAGGCACC Finding SNPsCTL-R2 AAGCTGGTTAGCACATTACAC Finding SNPsCTL –F3 TCAAATCATTTCCTTGTGAGGG Sequencing SNPs in two stocksCTL –R3 ACAAAAGGATGTCACCGTAGAA Sequencing SNPs in two stocksCTL –F4 GCAATTCTACGGTGACATCCTT Sequencing SNPs in two stocksCTL –R4 TTATCGCCATAGTTAGCCCAAA Sequencing SNPs in two stocksCTL –F5 ACTCCTTTTTGGGCTAACTATGGC Sequencing SNPs in two stocksCTL –R5 TCCAAAGTAAGCTGGTTAGCACAT Sequencing SNPs in two stocksCTL-HRM-F AAATGCCTACTGGAGGAGAGAAACA Genotyping amplificationCTL-HRM-R TTTCCTCGTTACTTCTCACAAATCG Genotyping amplification
2. Materials and methods
2.1. Crab breeding and V. alginolyticus challenge
One hundred and sixty swimming crabs with an average weightof 25 ± 3 g were collected from a commercial farm (Xiangshan,China) and acclimatized in aerated seawater at 25 °C for a weekbefore processing. During the entire period of the experiment, thecrabswere fed with clammeat once every night, and seawater changedevery day.
For the bacterial challenge experiment, the crabs were dividedinto four groups (40 crabs in each group). These were cultured in fourcement pools (2 m × 5 m × 0.9 m, width × length × depth). Crabsfrom the bacteria-challenged groups were injected of 200 μL liveV. alginolyticus suspended in 0.01 mol/L Phosphate Buffer Saline (PBS)(pH 7.2, 7.8 × 107 cfu/mL) and the injection site is the arthrodial mem-brane of the last walking leg. The control group crabs received an injec-tion of 200 μL PBS. All the crabs were observed every hour for the first24 h, followed by observation every 3 h after 120 h post-challenge.The crabs that died in the first 24 h were classified as the susceptiblestock, and the crabs that survived 120 h post-challenge were regardedas being relatively resistant to V. alginolyticus (resistant stock). Musclesin the last walking leg of all crabs from these two stocks were removedand stored at−80 °C for DNA extraction.
2.2. Acquiring of CTL DNA sequence
Genomic DNA was extracted from the muscle of P. trituberculatususing Genomic DNA Purification Kit (Axygen Biosciences, Corning, NY,USA). The primers CTL-F1 and CTL-R1 (Table 1) were designed, accord-ing the full length cDNA of PtLP in P. trituberculatus (GenBank accessionno. EU477491.1), to acquire the DNA sequence [21]. The reaction wascarried out in a 50 μL reaction mixture, containing 5 μL of 10× LA PCRbuffer (TaKaRa), 8 μL of dNTP (2.5 mM), 1 μL each of forward primerand reverse primer (10 μM), 0.5 μL (2.5 U) of LA Taq (TaKaRa), 3 μL oftarget DNA and 31.5 μL of double distilled water. The reaction wasperformed in a Tprofessional Thermocycler (Biometra, Goettingen,Germany) under the following conditions; 94 °C for 5 min, followedby 35 cycles of 94 °C for 30 s, 57 °C for 50 s, 72 °C for 2.5 min, and afinal extension at 72 °C for 10 min. The PCR products were purifiedby the manufacturer's protocol of PCR gel purification kit (Axygen)and cloned into pMD19-T simple vector (TaKaRa Bio Inc., Otsu, Japan).The vector was transformed into the Trans1-T1 phage resistantchemically competent cell (TransGen, Beijing, China), and the positiverecombinants identified through anti-Amp selection and PCR screeningusingM13-47 and RV-M primers (Table 1). Three of the positive cloneswere sequenced using an ABI3730 Automated Sequencer (AppliedBiosystems, Carlsbad, CA, USA).
2.3. Identification of polymorphic loci in the CTL
One pair of gene specific primer CTL-F2 and CTL-R2 (Table 1)was de-signed based on the obtained CTL DNA sequence to amplify the wholeCTL gene fragment. The reactionwas carried out in a 50 μL reactionmix-ture, containing 5 μL of 10× PCR buffer (TaKaRa), 6 μL of dNTP (2.5mM),1 μL each of forward primer and reverse primer (10 μM), 0.5 μL (1.25U) of PrimeSTAR HS DNA Polymerase (TaKaRa), 3 μL of target DNAand 33.5 μL of double distilled water. The reaction was performedfollowing conditions; one cycle of 94 °C for 4 min, followed by35 cycles of 94 °C for 50 s, 55 °C for 50 s, and 72 °C for 1.5 min, and afinal 10 min extension at 72 °C.
The PCR products from three susceptible and resistant crabs eachwere analyzed on 1% agarose gels and purified. The objective fragmentswere cloned into pMD19-T vector (TaKaRa) and at least three cloneswere sequenced from every sample. The objective nucleotide sequenceof CTL was aligned using the Vector NT1 Suite 9 (Life Technologies,Carlsbad, CA, USA). The polymorphisms in CTL gene were detectedfrom the sequence alignments of different crabs.
2.4. Analysis of SNPs in the CTL and association with V. alginolyticusresistance
Three pairs of gene-specific primers (CTL-F3/R3, CTL-F4/R4 andCTL-F5/R5) (Table 1) were designed based on the sites of polymorphism

127G. Hao et al. / Aquaculture 442 (2015) 125–131
in the CTL gene to amplify the genomic DNA obtained from the twostocks. Specific PCR products were purified and sequenced. The geno-type of every SNP from each sample was determined based on thesequencing chromatograms. Statistical analysis was carried out usingthe SPSS 11.0 software (IBM, Armonk, NY, USA). Significance analysisof genotype, and the allele frequencies between susceptible and resistantstocks were calculated by Chi-squared test. Results were considered asa significant difference when P b 0.05, and an extremely significantdifference when P b 0.01.
2.5. High resolution melting (HRM) for E4-205 locus genotyping
The significant SNP locus E4-205 was selected for genotyping withhigh resolution melting (HRM) assay. HRM analysis, which includedthe primer design target DNA template concentration, prediction ofmelting temperatures, and other parameters, was performed accordingProtocol of Rotor Gene Q (Qiagen, Venlo, Netherlands). This analysis isbased on the amplification of a short DNA fragment, for which the spe-cific primers CTL-HRM-F (R) were designed (Table 1). PCR mixtureconsisted of 1 μL genomic DNA (20 ng), 10 μL SsoFast EvaGreensupermix (Bio-Rad, Hercules, CA, USA), 0.8 μL each of the forward andreverse primer (10 μM), and 7.4 μL of double distilled water, makingup a volume of 20 μL. PCR cycling and HRM analysis were performedon the Rotor-Gene 6000 real-time PCR system (Qiagen) under the fol-lowing conditions: one cycle of 98 °C for 2 min, followed by 40 cyclesof 98 °C for 5 s, 58 °C for 20 s, 72 °C for 20 s, and a final 10min extensionat 72 °C. For melt curve analysis, the PCR products were heated from65 °C to 85 °C uniformly, and the fluorescent signal collected at every0.1 degree rise in temperature. Normalized and difference graphswere generated using the High Resolution Melting Analysis softwareprovided with the Rotor-Gene 6000.
2.6. Confirmation of significant SNP by challenge test of large sample
The association between the E4-205 locus and the susceptibility orresistance of swimming crab to V. alginolyticus was investigated by achallenge experiment. The bacterial challenge experiment was per-formed on 300 crabs. Muscle was collected from all the crabs (deadand surviving) and stored at −80 °C for DNA extraction. Genotype ofeach sample was determined by HRM assay (protocol as described inthe previous section). The mortality and survival rate for each genotypewere compared by Chi-squared analysis, and the statistical significancewas calculated using SPSS 11.0 (IBM).
3. Results
3.1. Sequence characterization of CTL
The full length cDNA of PtLP is 923 bp containing a 138 bp-long5ʹ-UTR and 290 bp 3 -UTR (Fig. 1A). The genomic DNA fragment be-tween positions 14 and 866 of PtLP cDNA was amplified. The obtained
Fig. 1. Schematic diagram of (A) full-length cDNA of PtLP obtained from P. tritu
amplicon was designated as CTL gene. The sequence and length(1473 bp) of CTL genomic DNA were deposited in GenBank under theaccession number JX195096.1. The exon–intron boundaries of CTLwere determined by alignmentwith the corresponding cDNA sequence.Three introns were found in the open reading frame region of CTL con-taining typical intron–exon junction structures, in which, donor and ac-ceptor (GT and AG) dinucleotide sequenceswere found (Figs. 1B and 2).
3.2. Analysis of crabs infected with live bacteria
In the challenge group, the first dead crabwas observed 6 h after in-fection. The deaths increased gradually and reached a peak at 60 h. Thecrabs that died in the first 24 h (n = 33) were classified as the suscep-tible stock, and the 33 crabs that survived the challenge experimentwere regarded as being relatively resistant to V. alginolyticus (resistantstock). No dead crabs were found in the control group.
3.3. CTL gene polymorphisms
A 1320 bp fragment was amplified from three susceptible and resis-tant crabs each using the CTL– F2 (R2) primer. This primer amplifies allthe exons and introns of CTL gene. 18 positive clones of each fragmentfrom the six crabs were sequenced. One insertion/deletion (ins/del)polymorphism and four SNPs were found in the amplified CTLfragments. Four polymorphisms were located in introns; I2-309 T-A,I3-89 A-C, I3-101A-G, and I3-104 CAA insertion/deletion. One non-synonymous substitution in the fourth exon, E4-205 C-T (Fig. 2), result-ed in the substitution of Threonine (ACT) to Isoleucine (ATT).
3.4. Association between gene polymorphisms and susceptibility/resistanceto V. alginolyticus
Genomic DNA from the 33 susceptible and resistant crabs each wasamplified via PCR. The SNP genotype isolated from each samplewas de-termined through sequencing chromatograms. Table 2 shows the alleleand genotype frequencies for CTL in the resistant and susceptible stocks.According to the results of the Chi-squared (x2) test, three SNPs (I2-309T-A, I3-89 A-C, I3-101A-G), and one ins/del polymorphism (I3-104 CAAins-del) exhibited significant (P b 0.05) difference in genotype frequen-cies, but nodifference in these allele frequencieswas found between theresistant and susceptible crabs. However, the SNP (E4-205) showed sig-nificant difference in both genotype and allele frequencies between thetwo stocks (P b 0.05). The T allele frequency was 33.3% in susceptiblestock, and 53% in resistant stock. In addition, the T/T genotype frequencywas 6.1% in susceptible stock, and 21.2% in resistant stock, with ademonstrated significant difference between them (P = 0.022).
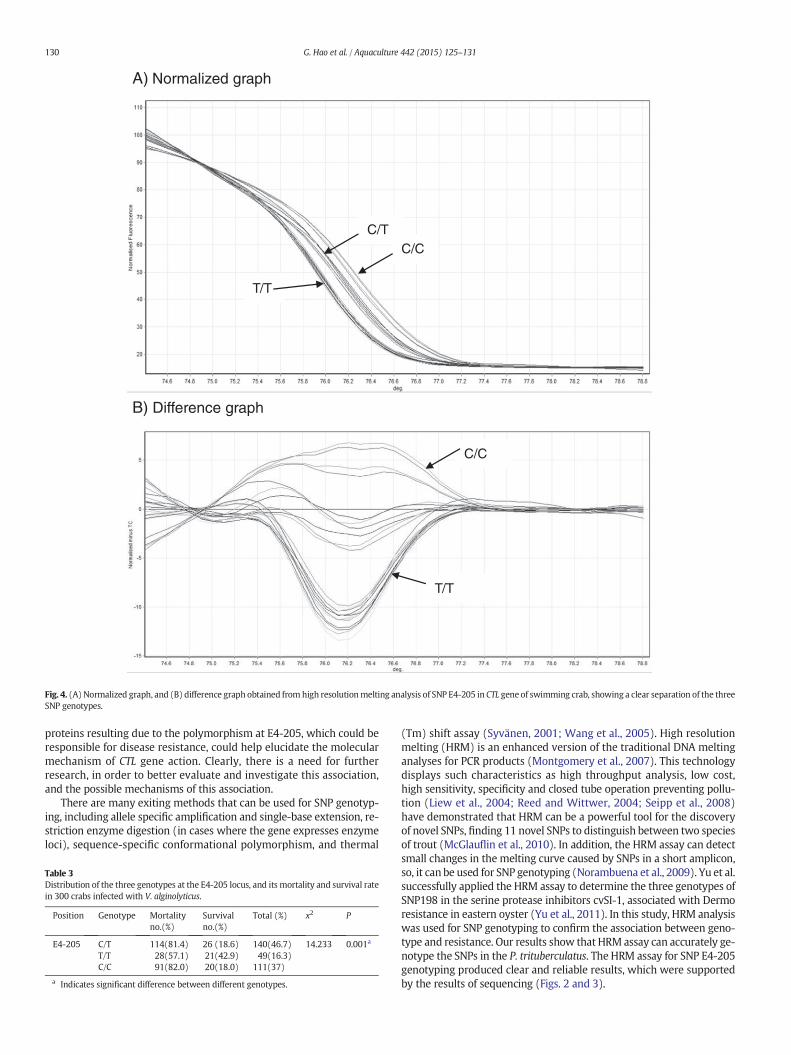
3.5. SNP E4-205 genotyping by HRM
The genotypes of SNP E4-205 were determined by the C/T, T/T andC/C sequencing chromatograms (Fig. 3). We have established the HRM
berculatus, and (B) partial genomic DNA of CTL obtained from PtLP cDNA.

Fig. 2. CTL genomic DNA nucleotide sequence and its polymorphisms obtained fromP. trituberculatus. Nucleotide numbering is shown at the left margin. The intron sequencesare highlighted. Intron dinucleotide acceptor and donor sites (GT/AG) for RNA splicing areunderlined. SNP sites are underlined and themutations are described below. The start andstop codons are displayed in boxes.
128 G. Hao et al. / Aquaculture 442 (2015) 125–131
method for the detection of the three genotypes, by screening ofprimers and optimizing other parameters. Normalized graph and differ-ence graph were generated with the aid of the High Resolution MeltingAnalysis software provided with the Rotor-Gene 6000. Genotypes ofeach sample were automatically indicated when the known genotypes(identified by sequencing) were included as positive controls. Fig. 4shows a clear separation of the three genotypes.
3.6. Relation of susceptibility/resistance phenotypes to genotypes atsignificant SNPs
To determine whether variation at CTL is associated withV. alginolyticus resistance, 300 crabs were infected of the bacteria, andthe SNP at E4-205 genotyped by HRM assay. Table 3 shows the distribu-tion of the three SNP genotypes in the 300 crabs. C/T genotype isobserved in 46.7%, T/T genotype in 16.3%, and C/C in 37% of the totalnumber of samples. The cumulative mortality of E4-205 T/T SNP was57.1%, which was extremely significantly lower than that of C/T(81.4%) and C/C (82%). However, no significant difference was detectedbetween samples expressing C/T and C/C polymorphism (P N 0.05)(Fig. 5).
4. Discussion
V. alginolyticus has been responsible for considerable economiclosses in the raising of swimming crabs over the past few years (Liuet al., 2007; Wang et al., 2006). The selective culture of swimmingcrabs by traditional methods in our laboratory has revealed thatsome crabs were disease resistant (Mu et al., 2012). However, little isknown about the molecular mechanism of disease resistance in crabs.Therefore, research conducted on the identification of genes, and itsSNPs related to disease resistance, could help improve our understand-ing of disease resistance in crabs, and may beneficial to cultivate anddevelop disease-resistant crabs.
Polymorphisms affecting gene expression, playing an important rolein resistance and susceptibility to diseases, is seen in humans(Mardan-Nik et al., 2014; Sun et al., 2007), scallops (Bao et al., 2010;Li et al., 2009), and oysters (Yu et al., 2011). In the present study, geno-mic DNA of C-type lectin is identified and one SNP is found to be relatedto susceptibility/resistance to V. alginolyticus in P. trituberculatus. To ourknowledge, this is the first study to identify an association between nu-cleotide polymorphisms of CTL genes and bacterial resistance. We haveanalyzed the temporal expression change of CTL gene in hemolymphand hepatopancreas after V. alginolyticus challenge in ordinary crabsand the disease-resistant stock cultured by traditional methods inour laboratory (unpublished data). The results demonstrated that theCTL gene was upregulated in both stocks, higher expression levels ofthe gene being observed in the disease-resistant stock, followingV. alginolyticus challenge. The combined results of these studies supportthe theory that polymorphisms in the CTL gene are the causative muta-tions affecting V. alginolyticus-resistance in swimming crab.
Among all the polymorphisms identified in the CTL gene, only theSNP at the E4-205 locus is significantly different in both genotype andallele frequencies between the two stocks of crabs (P b 0.05). TheE4-205 SNP is a non-synonymous variation, and its polymorphism re-sults in the substitution of Threonine (ACT) to Isoleucine (ATT). TheT/T SNP frequency was determined to be 6.1% in susceptible stock,and 21.2% in resistant stock, displaying a significant difference ofP=0.022. This indicated that E4-205 T/T was associated with the resis-tance of the swimming crabs to V. alginolyticus. Similar results havebeen reported in the P. trituberculatus anti-lipopolysaccharide factors(Li et al., 2013), and the SOD family of A. irradians (Bao et al., 2010).Non synonymous genetic variations, which improve crab's immune re-sistance, may cause considerable differences in the composition andstructure of the affected protein, thus changing its biological traits andfunctions. Therefore, the identification of the different functional

Table 2Distribution of CTL polymorphisms in susceptible and resistant stocks.
Serial no. Position Genotype Genotype no. (n = 33) x2 P Base type Allele no. x2 P
Susceptible Resistant Susceptible Resistant
1 I2-309 T/A 22(0.667) 29(0.879) 4.227 0.040a T 22(0.333) 29(0.439) 1.566 0.211A/A 11(0.333) 4(0.121) A 44(0.667) 37(0.561)
2 I3-89 A/C 22(0.667) 29(0.879) 4.227 0.040a A 44(0.667) 37(0.561) 1.566 0.211A/A 11(0.333) 4(0.121) C 22(0.333) 29(0.439)
3 I3-101 A/G 22(0.667) 29(0.879) 4.227 0.040a A 44(0.667) 37(0.561) 1.566 0.211A/A 11(0.333) 4(0.121) G 22(0.333) 29(0.439)
4 I3-104 CAA/CAA 11(0.333) 4(0.121) 4.227 0.040a ins 44(0.667) 37(0.439) 1.566 0.211I3-104 CAA/--- 22(0.667) 29(0.879) del 22(0.667) 29(0.439)
5 E4-205 C/T 18(0.545) 21 (0.636) 6.564 0.038a T 22(0.333) 35(0.530) 5.218 0.022a
T/T 2(0.061) 7(0.212) C 44(0.667) 31(0.470)C/C 13(0.394) 5(0.152)
I-intron, E-exon, and the number in position indicate number of base pairs. P values less than 0.05 are shown in bold.a Indicates significant difference between resistant and susceptible stocks.
C/TA
T/TB
C/CC
Fig. 3. The sequencing chromatograms of E4-205 SNP. The arrow shows that the genotype of E4-205 is (A) C/T, (B) T/T, and (C) C/C.
129G. Hao et al. / Aquaculture 442 (2015) 125–131
A) Normalized graph
B) Difference graph
T/T
C/TC/C
C/C
T/T
Fig. 4. (A) Normalized graph, and (B) difference graph obtained from high resolutionmelting analysis of SNP E4-205 in CTL gene of swimming crab, showing a clear separation of the threeSNP genotypes.
130 G. Hao et al. / Aquaculture 442 (2015) 125–131
proteins resulting due to the polymorphism at E4-205, which could beresponsible for disease resistance, could help elucidate the molecularmechanism of CTL gene action. Clearly, there is a need for furtherresearch, in order to better evaluate and investigate this association,and the possible mechanisms of this association.
There are many exiting methods that can be used for SNP genotyp-ing, including allele specific amplification and single-base extension, re-striction enzyme digestion (in cases where the gene expresses enzymeloci), sequence-specific conformational polymorphism, and thermal
Table 3Distribution of the three genotypes at the E4-205 locus, and its mortality and survival ratein 300 crabs infected with V. alginolyticus.
Position Genotype Mortalityno.(%)
Survivalno.(%)
Total (%) x2 P
E4-205 C/T 114(81.4) 26 (18.6) 140(46.7) 14.233 0.001a
T/T 28(57.1) 21(42.9) 49(16.3)C/C 91(82.0) 20(18.0) 111(37)
a Indicates significant difference between different genotypes.
(Tm) shift assay (Syvänen, 2001; Wang et al., 2005). High resolutionmelting (HRM) is an enhanced version of the traditional DNA meltinganalyses for PCR products (Montgomery et al., 2007). This technologydisplays such characteristics as high throughput analysis, low cost,high sensitivity, specificity and closed tube operation preventing pollu-tion (Liew et al., 2004; Reed and Wittwer, 2004; Seipp et al., 2008)have demonstrated that HRM can be a powerful tool for the discoveryof novel SNPs, finding 11 novel SNPs to distinguish between two speciesof trout (McGlauflin et al., 2010). In addition, the HRM assay can detectsmall changes in the melting curve caused by SNPs in a short amplicon,so, it can be used for SNP genotyping (Norambuena et al., 2009). Yu et al.successfully applied the HRM assay to determine the three genotypes ofSNP198 in the serine protease inhibitors cvSI-1, associated with Dermoresistance in eastern oyster (Yu et al., 2011). In this study, HRM analysiswas used for SNP genotyping to confirm the association between geno-type and resistance. Our results show that HRM assay can accurately ge-notype the SNPs in the P. trituberculatus. The HRM assay for SNP E4-205genotyping produced clear and reliable results, which were supportedby the results of sequencing (Figs. 2 and 3).

Fig. 5. Cumulative mortality of swimming crabs expressing SNP genotypes at the E4-205locus upon V. alginolyticus infection. ** indicates a highly significant statistical differenceof cumulative mortality between crabs expressing different SNP genotypes (P = 0.001).
131G. Hao et al. / Aquaculture 442 (2015) 125–131
In conclusion, polymorphisms in the CTL genewere investigated andsignificant association between the E4-205 SNP and V. alginolyticussusceptibility/resistance was established in P. trituberculatus. The devel-oped HRM assay provides a useful rapid method for SNP genotyping.The E4-205 T/T SNP variant might potentially be applied as the genemarker for selection of swimming crabs with enhanced resistance toV. alginolyticus.
Acknowledgements
The authors would like to thank Yanfei Wang, Chen Chen, andShaokun Lu for technical assistance in crab breeding and collection. Thisresearch was supported by the National High Technology Research andDevelopment Program of China (No. 2012AA10A409), the ZhejiangMajor Special Programof Breeding (2012C12907-3), the Zhejiang Provin-cial Natural Science Foundation of China under Grant No. LY12C19009,the National Natural Science Foundation of China (41106123), NingboNatural Science Foundation (2014A610183) and K.C. Wong MagnaFund by Ningbo University.
References
Bao, Y., Li, L., Zhang, G., 2010. Polymorphism of the superoxide dismutase gene family inthe bay scallop (Argopecten irradians) and its association with resistance/susceptibil-ity to Vibrio anguillarum. Dev. Comp. Immunol. 34 (5), 553–561.
Chen, P., Li, J., Liu, P., Gao, B., Wang, Q., 2010. Molecular cloning and characterization ofprophenoloxidase gene in swimming crab Portunus trituberculatus. Fish ShellfishImmunol. 28 (1), 106–112.
Collard, B., Jahufer, M., Brouwer, J., Pang, E., 2005. An introduction tomarkers, quantitativetrait loci (QTL) mapping and marker-assisted selection for crop improvement: thebasic concepts. Euphytica 142 (1–2), 169–196.
Dekkers, J.C., Hospital, F., 2002. The use of molecular genetics in the improvement of ag-ricultural populations. Nat. Rev. Genet. 3 (1), 22–32.
He, Y., Yu, H., Bao, Z., Zhang, Q., Guo, X., 2012.Mutation in promoter region of a serine pro-tease inhibitor confers Perkinsus marinus resistance in the eastern oyster (Crassostreavirginica). Fish Shellfish Immunol. 33 (2), 411–417.
Huang, Y., An, L., Hui, K.-M., Ren, Q., Wang, W., 2014. An LDLa domain-containing C-typelectin is involved in the innate immunity of Eriocheir sinensis. Dev. Comp. Immunol.42 (2), 333–344.
Jackson, A.N., McLure, C.A., Dawkins, R.L., Keating, P.J., 2007. Mannose binding lectin(MBL) copy number polymorphism in Zebrafish (D. rerio) and identification of hap-lotypes resistant to L. anguillarum. Immunogenetics 59 (11), 861–872.
Kong, H.J., Park, E.-M., Nam, B.-H., Kim, Y.-O., Kim, W.-J., Park, H.-J., Lee, C.H., Lee, S.-J.,2008. A C-type lectin like-domain (CTLD)-containing protein (PtLP) from the swim-ming crab Portunus trituberculatus. Fish Shellfish Immunol. 25 (3), 311–314.
Lee, H.J., Lee, D.-H., Yoon, S.-J., Kim, D.H., Kim, S.-G., Hyun, Y.S., Min, G.-S., Chung, K.W.,2013. Characterization of 20 microsatellite loci by multiplex PCR in swimming crab,Portunus trituberculatus. Genes Genom. 35 (1), 77–85.
Li, L., Zhao, J.,Wang, L., Qiu, L., Zhang, H., Dong, C., Cong,M., Song, L., 2009. The polymorphismof lysozyme gene in Zhikong scallop (Chlamys farreri) and its association with suscepti-bility/resistance to Listonella anguillarum. Fish Shellfish Immunol. 27 (2), 136–142.
Li, X., Cui, Z., Liu, Y., Song, C., Shi, G., Wang, C., 2013. Polymorphisms of anti-lipopolysaccharide factors in the swimming crab Portunus trituberculatus and theirassociation with resistance/susceptibility to Vibrio alginolyticus. Fish ShellfishImmunol. 34 (6), 1560–1568.
Liew, M., Pryor, R., Palais, R., Meadows, C., Erali, M., Lyon, E., Wittwer, C., 2004. Genotypingof single-nucleotide polymorphisms by high-resolution melting of small amplicons.Clin. Chem. 50 (7), 1156–1164.
Liu, Q., Wang, X., Dai, F., Liu, P., Li, J., 2007. Preliminary study on Vibrio alginolyticus dis-ease in Portunus trituberculatus. Shandong Fish. 24, 1–4.
Liu, Y., Cui, Z., Luan, W., Song, C., Nie, Q., Wang, S., Li, Q., 2011. Three isoforms of anti-lipopolysaccharide factor identified from eyestalk cDNA library of swimming crabPortunus trituberculatus. Fish Shellfish Immunol. 30 (2), 583–591.
Liu, L., Li, J., Liu, P., Zhao, F., Gao, B., Du, Y., Ma, C., 2012a. Correlation analysis of microsat-ellite DNA markers with growth related traits of swimming crab (Portunustrituberculatus). J. Fish. China 36 (7), 1034–1041.
Liu, Y., Cui, Z., Li, X., Song, C., Li, Q., Wang, S., 2012b. A new anti-lipopolysaccharide factorisoform (PtALF4) from the swimming crab Portunus trituberculatus exhibited struc-tural and functional diversity of ALFs. Fish Shellfish Immunol. 32 (5), 724–731.
Maillard, J.-C., Berthier, D., Chantal, I., Thevenon, S., Sidibé, I., Stachurski, F., Belemsaga, D.,Razafindraïbé, H., Elsen, J.-M., 2003. Selection assisted by a BoLA-DR/DQ haplotypeagainst susceptibility to bovine dermatophilosis. Genet. Sel. Evol. 35, S193–S200.
Mardan-Nik, M., Pasdar, A., Jamialahmadi, K., Biabangard-Zak, A., Mirhafez, S.R.,Ghalandari, M., Tajfard, M., Mohebati, M., Esmaily, H., Ferns, G.A., 2014. Associationof heat shock protein70-2 (HSP70-2) gene polymorphism with coronary artery dis-ease in an Iranian population. Gene 550 (2), 180–184.
Masoudi, A.A., Uchida, K., Yokouchi, K., Miyadera, K., Ogawa, H., Sugimoto, Y., Kunieda, T.,2007. Marker‐assisted selection for forelimb‐girdle muscular anomaly of JapaneseBlack cattle. Anim. Sci. J. 78 (6), 672–675.
McGlauflin, M.T., Smith, M.J., Wang, J.T., Young, S.F., Chen, N., Lee, Y.C., Pascal, C., Seeb,L.W., Stevens, J., Seeb, J.E., 2010. High-resolution melting analysis for the discoveryof novel single-nucleotide polymorphisms in rainbow and cutthroat trout for speciesidentification. Trans. Am. Fish. Soc. 139 (3), 676–684.
McGreal, E.P., Martinez-Pomares, L., Gordon, S., 2004. Divergent roles for C-type lectinsexpressed by cells of the innate immune system. Mol. Immunol. 41 (11), 1109–1121.
Montgomery, J., Wittwer, C.T., Palais, R., Zhou, L., 2007. Simultaneous mutation scanningand genotyping by high-resolution DNA melting analysis. Nat. Protoc. 2 (1), 59–66.
Mu, C., Liu, S., Song, W., Li, R., Wang, C., 2012. Enhanced resistance of Portunus trituberculatusto Vibrio alginolyticus by selective breeding. Chin. J. Oceanol. Limnol. 30, 638–643.
Norambuena, P.A., Copeland, J.A., Křenková, P., Štambergová, A., Macek Jr., M., 2009. Diag-nosticmethodvalidation:High resolutionmelting (HRM)of small amplicons genotypingfor the most common variants in theMTHFR gene. Clin. Biochem. 42 (12), 1308–1316.
Pan, L., Yue, F., Miao, J., Zhang, L., Li, J., 2010. Molecular cloning and characterization of anovel c-type lysozyme gene in swimming crab Portunus trituberculatus. Fish ShellfishImmunol. 29 (2), 286–292.
Reed, G.H., Wittwer, C.T., 2004. Sensitivity and specificity of single-nucleotide polymor-phism scanning by high-resolution melting analysis. Clin. Chem. 50 (10), 1748–1754.
Seipp, M.T., Pattison, D., Durtschi, J.D., Jama, M., Voelkerding, K.V., Wittwer, C.T., 2008.Quadruplex genotyping of F5, F2, and MTHFR variants in a single closed tube byhigh-resolution amplicon melting. Clin. Chem. 54 (1), 108–115.
Sun, T., Gao, Y., Tan, W., Ma, S., Shi, Y., Yao, J., Guo, Y., Yang, M., Zhang, X., Zhang, Q., 2007.A six-nucleotide insertion-deletion polymorphism in the CASP8 promoter is associat-ed with susceptibility to multiple cancers. Nat. Genet. 39 (5), 605–613.
Syvänen, A.-C., 2001. Accessing genetic variation: genotyping single nucleotide polymor-phisms. Nat. Rev. Genet. 2 (12), 930–942.
Wan, X., Shen, H., Wang, L., Cheng, Y., 2011. Isolation and characterization of Vibriometschnikovii causing infection in farmed Portunus trituberculatus in China. Aquac.Int. 19 (2), 351–359.
Wang, X.-W., Wang, J.-X., 2013. Diversity and multiple functions of lectins in shrimp im-munity. Dev. Comp. Immunol. 39 (1), 27–38.
Wang, J., Chuang, K., Ahluwalia, M., Patel, S., Umblas, N., Mirel, D., Higuchi, R., Germer, S.,2005. High-throughput SNP genotyping by single-tube PCR with T ~ m-shift primers.Biotechniques 39 (6), 885.
Wang, G., Shan, J., Chen, Y.-e., Li, Z., 2006. Study on pathogens and pathogenesis of emul-sification disease of Portunus trituberculatus. Adv. Mar. Sci. 4, 014.
Wang, L., Su, J., Yang, C., Wan, Q., Peng, L., 2012. Genomic organization, promoter activityof grass carp MDA5 and the association of its polymorphisms with susceptibility/re-sistance to grass carp reovirus. Mol. Immunol. 50 (4), 236–243.
Wang, L., Wang, L., Zhang, D., Li, F., Wang, M., Huang, M., Zhang, H., Song, L., 2013. A novelC-type lectin from crab Eriocheir sinensis functions as pattern recognition receptor en-hancing cellular encapsulation. Fish Shellfish Immunol. 34 (3), 832–842.
Wei, X., Liu, X., Yang, J., Fang, J., Qiao, H., Zhang, Y., Yang, J., 2012. Two C-type lectins fromshrimp Litopenaeus vannamei that might be involved in immune response againstbacteria and virus. Fish Shellfish Immunol. 32 (1), 132–140.
Yu, H., He, Y., Wang, X., Zhang, Q., Bao, Z., Guo, X., 2011. Polymorphism in a serine prote-ase inhibitor gene and its association with disease resistance in the eastern oyster(Crassostrea virginica Gmelin). Fish Shellfish Immunol. 30 (3), 757–762.
Yue, F., Pan, L., Miao, J., Zhang, L., Li, J., 2010. Molecular cloning, characterization andmRNA expression of two antibacterial peptides: crustin and anti-lipopolysaccharidefactor in swimming crab Portunus trituberculatus. Comp. Biochem. Physiol. B Biochem.Mol. Biol. 156 (2), 77–85.

本文献由“学霸图书馆-文献云下载”收集自网络,仅供学习交流使用。
学霸图书馆(www.xuebalib.com)是一个“整合众多图书馆数据库资源,
提供一站式文献检索和下载服务”的24 小时在线不限IP
图书馆。
图书馆致力于便利、促进学习与科研,提供最强文献下载服务。
图书馆导航:
图书馆首页 文献云下载 图书馆入口 外文数据库大全 疑难文献辅助工具