simultaneous analysis the adhesion and release granule
TRANSCRIPT

Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 1071
_____________________________ _____________________________
Granulocyte Adhesion to MatrixProteins and the Effect on theRelease of Granule Proteins
Development of a Simple Method and its Application in
Experimental and Clinical Studies
BY
XIAOYAN XU
ACTA UNIVERSITATIS UPSALIENSISUPPSALA 2001

Dissertation for the Degree of Doctor of Philosophy (Faculty of Medicine) in Clinical Chemistry presented atUppsala University in 2001
ABSTRACT
Xu, X. 2001. Granulocyte Adhesion to Matrix Proteins and the Effect on the Release of Granule Proteins.Development of a Simple Method and its Application in Experimental and Clinical Studies. Acta UniversitatisUpsaliensis. Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 1071. 46 pp. Uppsala. ISBN 91-554-5112-8
Granulocyte adhesion and release of their granule proteins are key steps during selective accumulation of acertain cell to an inflammatory site. Eosinophils are specifically recruited to sites of allergic inflammation andparasitic infection, whereas neutrophil influx predominates in bacterial infection and rheumatoid arthritis.
A simple, reliable and convenient method was developed for the measurement of granulocyte adhesion andrelease of granule proteins by using the normal population of granulocytes. The design allows simultaneousquantitative assessment of eosinophil and neutrophil adhesion to proteins and degranulation.
Using this method, manganese ions (Mn2+) induced a higher level of eosinophil adhesion to fibronectin,fibrinogen and albumin as compared with neutrophils. PMA induced comparable levels of eosinophil andneutrophil adhesion. F-MLP stimulated a rapid, short-term adhesion of neutrophils to fibrinogen.
In the same conditions PMA alone stimulated a dose-dependent release of ECP from cells that adhered toboth fibronectin and fibrinogen. Meanwhile, Mn2+ amplified the release of ECP induced by PMA. Furthermore,release of ECP was shown to be associated with cell death.
PMA, in combination with Mn2+, induced a marked release of ∼ 80%of the intracellular content of lactoferrinand HNL in neutrophils. PMA or f-MLP alone induced 30-40% release of lactoferrin and HNL. A maximalrelease of MPO of 15-20% was obtained from neutrophils stimulated by PMA and Mn2+. Release of lactoferrinand HNL showed a significant negative relationship to the viability of cells.
Stimulated by PMA, eosinophils from pollen-atopic patients during early pollen season displayed a markedlyenhanced adhesion and release of ECP of eosinophils compared with eosinophils from the references. Primingwith IL-5 caused a significantly higher adhesion and release of ECP by eosinophils in response to PMA. GM-CSF priming enhanced eosinophil adhesion in response to PAF and PMA plus Mn2+, but did not enhance therelease of ECP.
In conclusion, the assay allows a simple quantification of eosinophil and neutrophil adhesion, as well asdegranulation by using the normal population of granulocytes. Cellular adhesion plays an important role in theregulation of both eosinophil and neutrophil degranulation, but adhesion and degranulation can be inducedseparately.
Key words: Eosinophils, neutrophils, adhesion, release of ECP, extracellular matrix proteins, granulocytes.
Xiaoyan Xu, Department of Medical Sciences, Clinical Chemistry, University Hospital, SE-751 85 Uppsala, Sweden
Xiaoyan Xu 2001
ISSN 0282-7476ISBN 91-554-5112-8
Printed in Sweden by Kopieringshuset AB, Uppsala 2001

To Zhurong, Yangzi and my parents

4
This thesis is based on the following papers, which are referred to by their
Roman numerals.
I. Xiaoyan Xu and Lena Håkansson
Simultaneous analysis of eosinophil and neutrophil adhesion to plasma and tissue
fibronectin, fibrinogen and albumin. J.Immunol.Methods 1999, 226, 93-104.
II. Xiaoyan Xu and Lena Håkansson
Regulation of the release of eosinophil cationic protein by eosinophil adhesion.
Clin.Exp.Allergy 2000, 30, 794-806
Ш. Xiaoyan Xu and Lena Håkansson
Degranulation of primary and secondary granules in adherent human neutrophils
Submitted for publication
IV. Xiaoyan Xu , L. Håkansson and G. Stålenheim
The capability of adhesion and release of ECP of eosinophils from pollen-atopic
patients and cytokine primed normal eosinophils
Manuscript
Reprints were made with the permission of the publisher.

5
CONTENTS
1. ABBREVIATIONS……………………………………………………………………..6
2. INTRODUCTION………………………………………………………………………7
General background…………………………………………………………………….8
Asthma and allergic diseases Eosinophil granulocytes Neutrophil granulocytes Extra cellular matrix proteins Adhesion Adhesion molecules
Degranulation Eosinophil priming Cytokines Cell death 3. AIMS OF THE STUDY………………………………………………………………20
4. MATERIAL AND METHODS………………………………………………………21
Isolation of granulocytes Purification of eosinophils Coating of wells Stimulation of cells Priming of eosinophils Adhesion assay Release assay Radioimmunoassays Flow cytometry Assessment of granulocyte death Inhibition of release and adhesion by monoclonal antibodies Statistical analysis
5. RESULTS AND DISCUSSION………………………………………………………..26
6. CONCLUSIONS………………………………………………………………………..34
7. ACKNOWLEDGEMENTS……………………………………………………………35
8. REFERENCES…………………………………………………………………………37
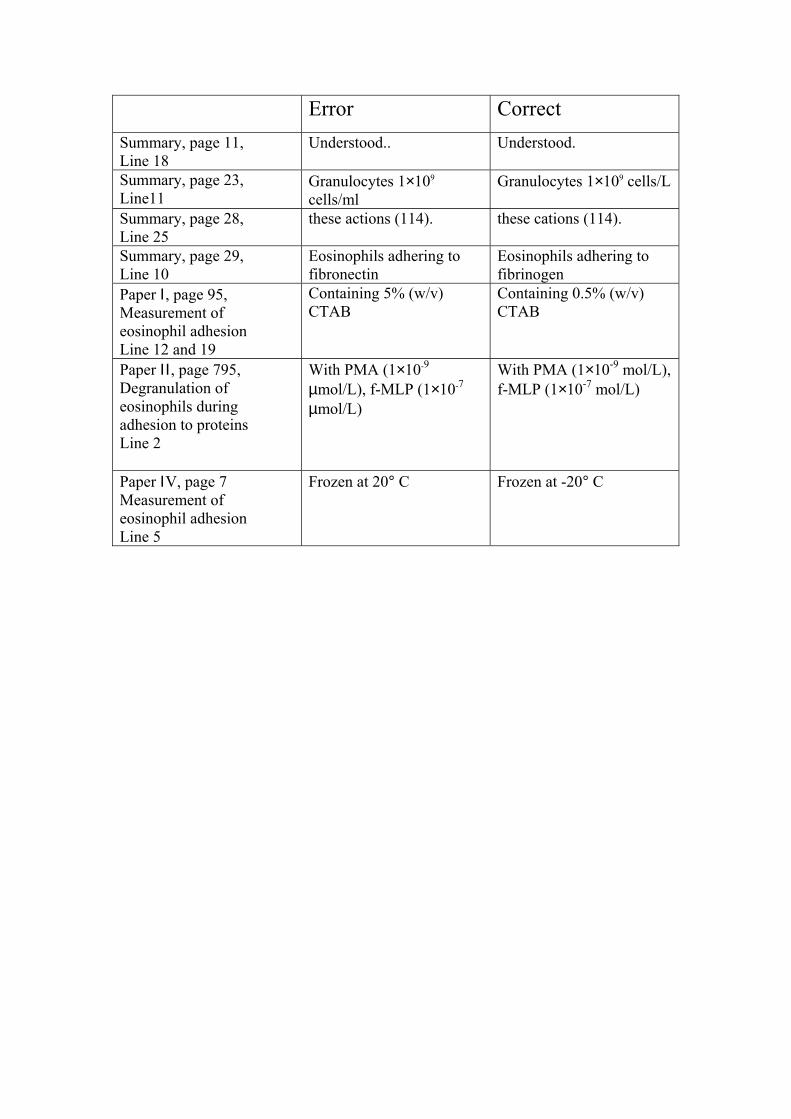
Error CorrectSummary, page 11, Line 18
Understood.. Understood.
Summary, page 23,Line11
Granulocytes 1×109
cells/mlGranulocytes 1×109 cells/L
Summary, page 28,Line 25
these actions (114). these cations (114).
Summary, page 29,Line 10
Eosinophils adhering tofibronectin
Eosinophils adhering tofibrinogen
Paper Ι, page 95,Measurement ofeosinophil adhesionLine 12 and 19
Containing 5% (w/v)CTAB
Containing 0.5% (w/v)CTAB
Paper ΙΙ , page 795,Degranulation ofeosinophils duringadhesion to proteinsLine 2
With PMA (1×10-9
µmol/L), f-MLP (1×10-7
µmol/L)
With PMA (1×10-9 mol/L),f-MLP (1×10-7 mol/L)
Paper ΙV, page 7Measurement ofeosinophil adhesionLine 5
Frozen at 20° C Frozen at -20° C

6
1. ABBREVIATIONS
BPI – bactericidal/permeability-increasing proteinCGRP – calcitonin gene-related peptideCTAB - N-cetyl-N,N,N-trimethyl-ammonium bromideECM – extracellular matrix proteinECP – eosinophil cationic proteinEDN – eosinophil-derived neurotoxinEPO – eosinophil peroxidaseERK – extracellular regulated kinase FDA – fluorescein diacetateFMCA – fluorometric microculture cytotoxicity assayf-MLP - formyl-methionyl-leucyl-phenylalanineGM-CSF – granulocyte-macrophage colony-stimulating factorHNL – human neutrophil lipocalinHSA - human serum albuminICAM – intercellular adhesion moleculeLTC – leukotriene CLTD – leukotriene DIL – interleukinmAb - monoclonal antibodyMadCAM – mucosal addressin cell adhesion moleculeMAPK – mitogen activated protein kinaseMBP – major basic proteinMEK – MAPK / ERK kinaseMFI - mean fluorescence intensityMPO – myeloperoxidaseNBS – new-born calf serumPAF – platelet-activating factorPI – propidium iodidePKC - protein kinase CPMA - phorbol-myristate-acetatePMN – polymorphonuclear leukocytesRIA - radioimmunoassaySTZ – serum-treated zymosanTNF – tumour necrosis factorVCAM-1 – vascular cell adhesion moleculeVLA – very late activation antigen

7
2. INTRODUCTION
Eosinophils are specifically recruited to sites of allergic inflammation and parasitic infection.
Neutrophil influx, on the other hand, predominates in bacterial infection. This biological
selectivity suggests that granulocytes may respond differentially to inflammatory mediators
during the recruitment to inflammatory foci (1). For instance, secretory IgA and secretory
component preferentially activate human eosinophils (2). The selective migration of different
granulocyte types from the circulation into the tissue might partly be due to the demonstrated
increased surface expression and activation of adhesion molecules, such as CD11/CD18 and
VLA-4 (3;4). Thus, adhesion may be regarded as one of the first functional events in the
activation cascade, whereas degranulation of the granule proteins most likely represents a
final functional event in this activation cascade (5).
In the activation cascade, leukocytes encounter the extracellular matrix protein (ECM) that
underlies endothelia and epithelia and surrounds connective tissue cells. It has been
demonstrated that adherence to the ECM is important for the localisation of leukocytes to a
site of inflammation (6) and that a number of key functions of leukocytes, including adhesion,
cytokine production (7;8), cell migration and activation (9) and release of granule contents
(10), are significantly affected. Examples of this are that adhesion of eosinophils to
fibronectin attenuated activation and release of EDN (eosinophil-derived neurotoxin) (11) and
prolonged eosinophil survival (12). The ability of the ECM to convey information to the cell
is thought to involve β1 and β2 integrins (13). Therefore, it seems reasonable that adhesion
molecules play an important role in normal immune surveillance and in the pathogenesis of
tissue injury, including cell recruitment at the site of allergic inflammation. Earlier studies
have shown increased expression of ICAM-1, E-selectin and VCAM-1 on endothelium in
allergic asthma (14). Meanwhile, secretion of granule proteins from eosinophils and
neutrophils was increased in asthma and cystic fibrosis (15;16).
It has become evident that specialised CD4+ type 2 T-helper cells are important in the
pathogenesis of allergic inflammation (17). The Th2 cells produce cytokines (e.g., IL-3 IL-5
and GM-CSF) (18). These cytokines are important modulators or primers of eosinophil
function (e.g., degranulation, adhesion, survival and superoxide production) (19-21). It has

8
been reported that patients with asthma show altered surface expression of the adhesion
molecules on airway granulocytes compared with blood granulocytes (22).
Though the eosinophil is a central effector cell in allergic diseases, neutrophils are also found
in bronchial lavage from patients undergoing acute exacerbations that were not associated
with infection (23).
The aim of the present work was to develop a simple, reliable and convenient assay for the
simultaneous measurement of eosinophil and neutrophil adhesion and release. Most of the
studies in this field are based on separate experiments performed with either purified
eosinophils or neutrophils whereas these two types of cells coexist in the blood. The present
method may offer a useful tool for comparative studies on eosinophils and neutrophils from
different patient groups. The method should also be helpful in characterising the effect of
inflammatory mediators or pharmacological agents on the selective adhesion and release of
granule contents in different types of granulocyte. As an important priority, we wanted to
assay the function of eosinophils within the normal population of granulocytes, making it
possible to perform a large number of experiments during states approaching physiological
conditions. As a comparison, neutrophil adhesion and degranulation of granule proteins were
measured. To investigate whether priming in vivo affects adhesion and degranulation
eosinophils from atopic patients exposed to allergens were compared with granulocytes from
healthy individuals. Adhesion to extracellular matrix proteins (ECM) and release of ECP were
investigated. In an attempt to mimic the situation in vivo, IL-5 and GM-CSF were used to
prime normal eosinophils in vitro. We then investigated the effect of this procedure on
eosinophil adhesion and release after further stimulation by PMA, PAF and Mn2+.
GENERAL BACKGROUND
1. Asthma and allergic diseases
Asthma is a common disease in the industriliazed world, for instance affecting 4 to 10% of
the population in the United States (24). It is an inflammatory disease involving mast cells,
antigen-presenting cells, macrophages, eosinophils, neutrophils, Th2-lymphocytes and
epithelial cells.

9
Blood and local tissue eosinophilia are outstanding features of allergic diseases such as
asthma. The evidence for the contribution of the eosinophil and eosinophil-elaborated
mediators to the pathophysiology of these disorders is increasing (25). This observation is one
of the cornerstones of the current hypothesis suggesting a central role for eosinophil-derived
mediators in causing asthma and related allergic diseases. Accordingly, selective
accumulation of eosinophils in the airways in allergic diseases has attracted significant
attention as an important mechanism underlying the diseases. However, the exact role of
eosinophils in the pathophysiology of chronic allergic disease remains obscure. The
recruitment of eosinophils to inflamed tissues is a multi-stage process that entails eosinophil
priming (26), adhesion, migration, generated cytokines, degranulation and survival (4;27;28).
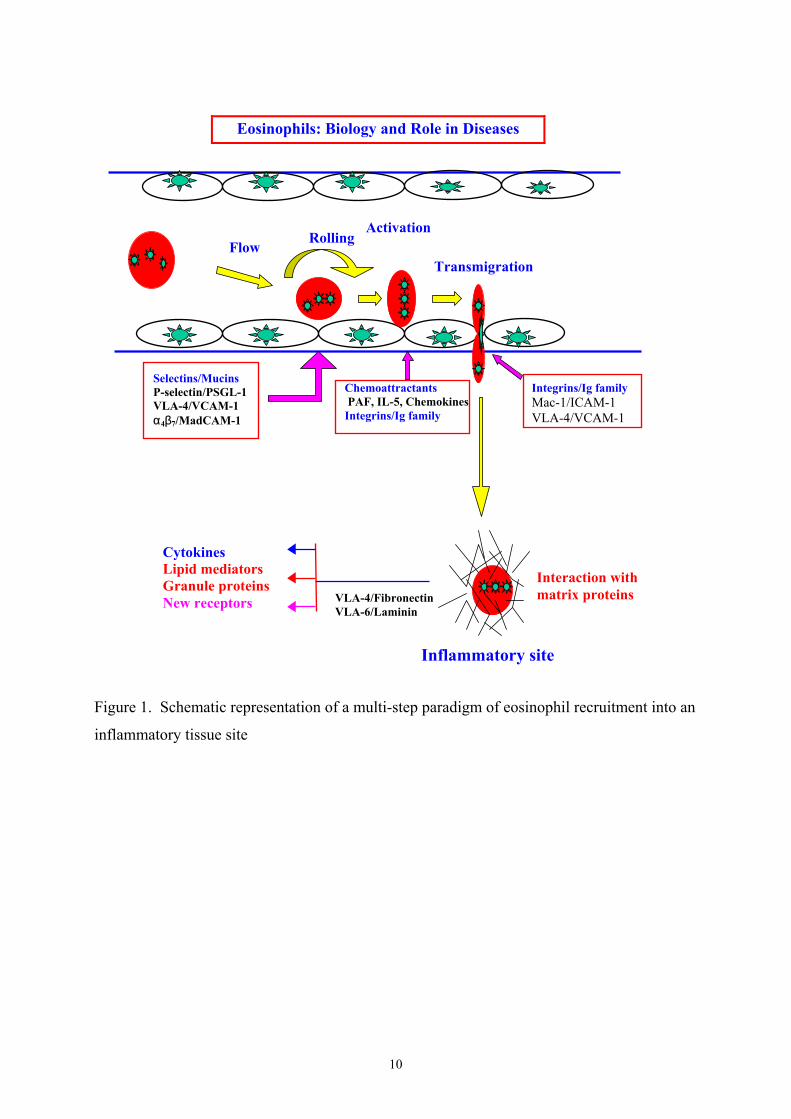
10
Figure 1. Schematic representation of a multi-step paradigm of eosinophil recruitment into an
inflammatory tissue site
Eosinophils: Biology and Role in Diseases
FlowRolling
Activation
Transmigration
Selectins/MucinsP-selectin/PSGL-1VLA-4/VCAM-1α4β7/MadCAM-1
Chemoattractants PAF, IL-5, ChemokinesIntegrins/Ig family
Integrins/Ig familyMac-1/ICAM-1VLA-4/VCAM-1
Interaction withmatrix proteins
CytokinesLipid mediatorsGranule proteinsNew receptors VLA-4/Fibronectin
VLA-6/Laminin
Inflammatory site

11
2. Eosinophil granulocytes
Since Paul Ehrhich’s discovery of eosinophils in 1879, eosinophils have been found to be
associated with helminth infection, bronchial asthma and other allergic diseases (29).
Eosinophils play a principal role during the late-phase reaction in allergic inflammation,
accounting for about 30% of the accumulated cells (30); in the blood; however, only 1-4% of
all leukocytes are eosinophils. The influx of eosinophils in the late-phase response has been
shown to contribute to the chronic inflammation of the bronchial mucosa that characterises
persistent asthma (31-33). On the other hand, eosinophils are normally not present in lung
tissue. However, eosinophils are primarily tissue-dwelling cells in the lamina propria of the
normal gut, where they appear as intact cells. In helminth and allergic diseases eosinophils
often undergo massive degranulation. Preformed and newly formed eosinophil-derived
inflammatory mediators including cationic proteins like ECP, EDN, MBP and EPO (34), lipid
mediators such as LTC4, LTD4 and PAF, and oxygen metabolites (35-37) are thought to cause
many of the symptoms observed in allergic disease (25). The fact that eosinophils accumulate
in large numbers despite being present in peripheral blood at much lower levels suggests that
a pathway exists that selectively permits their local influx and survival. The mechanisms
underlying the selective eosinophil accumulation at the sites of inflammation and the
activation and release of mediators are, however, still poorly understood.. A multi-step
process is thought to be involved (see below).
a. Increased eosinophilopoiesis and release from the bone marrow (38;39).
b. Adhesion to endothelium and matrix proteins via the adhesion receptor VLA-4 and
selective priming of eosinophil adhesion by cytokine IL-5 (4;40).
c. Regulation of eosinophil migration into the tissue by eosinophil chemoattractants,
particularly chemokines (41;42).
d. Maintenance of eosinophil numbers in the tissue by prolonged survival, i.e. inhibition of
apoptosis (43).
The differentiation and proliferation of eosinophils are controlled by IL-3, GM-CSF and IL-5.
All these growth factors are also capable of priming eosinophils for enhanced biological
functions, such as increased mediator release and prolonged survival (3;20). The list of
stimuli that can elicit rapid activation of eosinophils includes antibodies, complement-derived
opsonins and anaphylatoxins, PAF, leukotrienes, formylated methionyl peptides and other
chemoattractants.

12
Table 1. Activation and Regulation of Eosinophil FunctionActivators & regulators Degranu
lation
Production
of lipid
mediators
Priming Chemotaxis/chemokinesis
Adhesion Survival Cytokineproduction
IgGIgEIgA
↑↑↑
↑↑
↑ ↑↑↑
C3aC5aZymosan
↑↑↑ ↑
↑↑ ↑
PAFLTB4
↑ ↑ ↑ ↑↑
↑ ↑
IL-3IL-4IL-5GM-CSFTNFα
↑↓↑↑↑
↑
↑↑
↑
↑↑
↑
↑↑
↑
↑↑↑
↑↑
IL-8RANTES ↑
↑↑ ↑
β2-integrinligandsFibronectinVCAM-1
↑ ↑
↑↑
↑
↑↑
↑
FMLPPMA
↑↑
↑ ↑ ↑↑
↑
3. Neutrophil granulocytes
Neutrophils are known to be involved in the defence of the body against bacteria, fungi and
virus. They are the first cell type to enter the lung following allergen challenge (44-46).
Unlike eosinophils, the influx of neutrophils is transient after allergen challenge. Furthermore,
high neutrophil counts are not only observed in bronchial lavage from patients undergoing
acute exacerbations that were not associated with infection, but are also present in high
numbers in the airways of patients with chronic severe asthma (47). Neutrophils are also

13
found to be another major participant in late-phase reactions. One of the mechanisms
underlying the attraction of neutrophils to the area of a type Ι reaction is probably by a
neutrophil chemotactic factor that is released from degranulating mast cells (30).
Neutrophils can produce a wide range of products, including lipids (LTB4,PAF), cytokines
(IL-8, TNFα), proteases (elastase, collagenase) and microbicidal products (lactoferrin, MPO)
(47). The granule proteins secreted by neutrophils are compartmentalised into at least three
distinct granule populations: primary, specific and tertiary granules. Primary granules contain
MPO (myeloperoxidase), elastase and BPI (bactericidal/permeability-increasing protein);
specific granules comprise lysozyme, lactoferrin and HNL (human neutrophil lipocalin); and
tertiary granules include gelatinase. Specific granules have been characterised as secretory
granules that are known to play an important role in the initiation of the inflammatory
response (48).
Specific granules and tertiary granules are associated with diapedesis and phagocytosis via
several mechanisms listed below (49):
a. Primary contact between endothelium and circulating neutrophils is established via
selectins and their ligands. The contact causes neutrophil to roll along the activated
endothelium.
b. Integration of the membrane of secretory vesicles and its associated CD11b/CD18
molecules into the cell membrane enhances the potential of the neutrophils for firm
adhesion to endothelium.
c. Exocytosis of gelatinase from tertiary granules may facilitate degradation of type IV
collagen in basement membranes.
d. Mobilisation of specific granules to the surface membrane may enhance the phagocytic
potential of the neutrophils by providing CD11b/CD18, i.e. the C3b-receptor.
4. Extracellular matrix proteins
The ECM is a relatively stable structural matrix that underlies epithelia and endothelia and
surrounds connective tissue cells. During adhesion to endothelium and subsequent
transendothelial migration, leukocytes encounter matrices and bind to components of the
ECM via integrins and other receptors on their surface (6). The ECM has been shown to
significantly affect a number of different leukocyte responses to inflammatory stimuli. For
instance, eosinophil degranulation and activation induced by PAF, C5a and IL-5 was

14
attenuated by fibronectin and laminin compared with fibrinogen (11). Furthermore, when
granulocytes become adherent to ECM proteins or endothelial cells, they are thought to be
primed to respond to additive stimuli (e.g., cytokines or chemoattractants) (50;51). Although
the function of cell-matrix interaction in inflammation remains ambiguous, some observations
indicate that fibronectin and collagen are found in increased amounts in the bronchoalveolar
lavage fluid or lung tissue of children with obstructive bronchitis (52). Recent evidence
suggests that diseases characterised by airway inflammation (e.g., asthma and chronic
obstructive lung disease) are also associated with increased ECM deposition (53).
The ECM participates in the following areas:
A. Attachment of cells
B. Tissue growth and repair
C. Proliferation and differentiation
D. Cell migration and activation
E. Cell survival/delay of apoptosis
F. Chemotaxis
G. Degranulation
5. Adhesion
Granulocyte adhesion during an inflammatory process involves a series of interactions as the
cells migrate into the tissue. First, there is adhesion to vascular endothelium followed by
interaction with an extracellular matrix and resident cells. Each of these steps involves a
distinct set of cell surface receptors. The first step, in which leukocytes tether and roll along
the vascular endothelium, is now recognised to be selectin-dependent, including E-selectin, L-
selectin and P-selectin (54). Following this, β2 and β1 integrin expressions on the surface of
eosinophils and neutrophils are rapidly increased (55;56), whereas ICAM-1 and VCAM-1 on
the surface of the endothelium are activated and increase their expression (14;57;58). This, in
turn, leads to firm adhesion between the leukocytes and endothelium. Finally, the cells reach
the site of inflammation and come in contact with ECM (e.g., fibronectin and laminin) via
integrins, which are cell surface receptors. Eosinophils and neutrophils express β2 integrins:
CD11a/CD18, CD11b/CD18 and CD11c/CD18. Although β2 integrins, especially
CD11b/CD18, play an essential role in the adherence of both cell types to albumin (59;60)
and fibrinogen (61), eosinophils also express several β1 integrins, particularly CD49d/CD29

15
(VLA-4), which neutrophils do not express (62-64). VLA-4, the α4β1 integrin and α4β7,
which mediate eosinophil adhesion to VCAM-1 and fibronectin (57;65), may account for the
selective accumulation of eosinophils to the site of allergic inflammation. However, VLA-4 is
also expressed by monocytes and subgroups of lymphocytes, implying that several pathways
are involved in the selective accumulation of eosinophils (66;67).
6. Adhesion molecules
Multiple receptor systems mediate a variety of leukocyte adhesion capabilities. The cell
adhesion receptors are usually transmembrane glycoproteins that mediate binding interactions
at the extracellular surface and determine the specificity of cell-cell and cell-ECM
recognition. They include members of the integrin, cadherin, immunoglobulin, selectin and
proteoglycan superfamilies (68).
Since their initial discovery by Springer and others in the past decade (69), adhesion
molecules have been shown to have pivotal roles in normal immune surveillance and
function, as well as in the pathogenesis of tissue injury (70). The interaction of granulocytes
with ECM is mainly mediated by receptors of the integrin family (71). For example,
neutrophils are thought to bind to collagen via the β2-integrin CD11b/ CD18 and the β1-
integrins VLA-1 and VLA-2 and to laminin via β2-integrins and the β1 integrins VLA-5 and
VLA-6. Eosinophil adhesion to fibrinogen has been found to be mediated by β1 and β2
integrins and neutrophil adhesion to fibrinogen by the β2 integrin CD11b/CD18 and the β1
integrin VLA-5. Neutrophil adhesion to fibronectin is mediated by β2 integrins and by the β1
integrin VLA-5, whereas eosinophil adhesion to fibronectin is mediated by VLA-4 and α4β7
(12;72-75). In addition, recent evidence suggests that integrins, particularly the β2 integrin,
are capable of modulating the eosinophil degranulation response stimulated with various
physiologic secretagogues (76).
In contrast to the principal mechanism of modulation of the selectins and immunoglobulin
family adhesion receptors, which is accomplished through changing the number of expressed
receptors, integrin function is regulated by the conformational state (77).
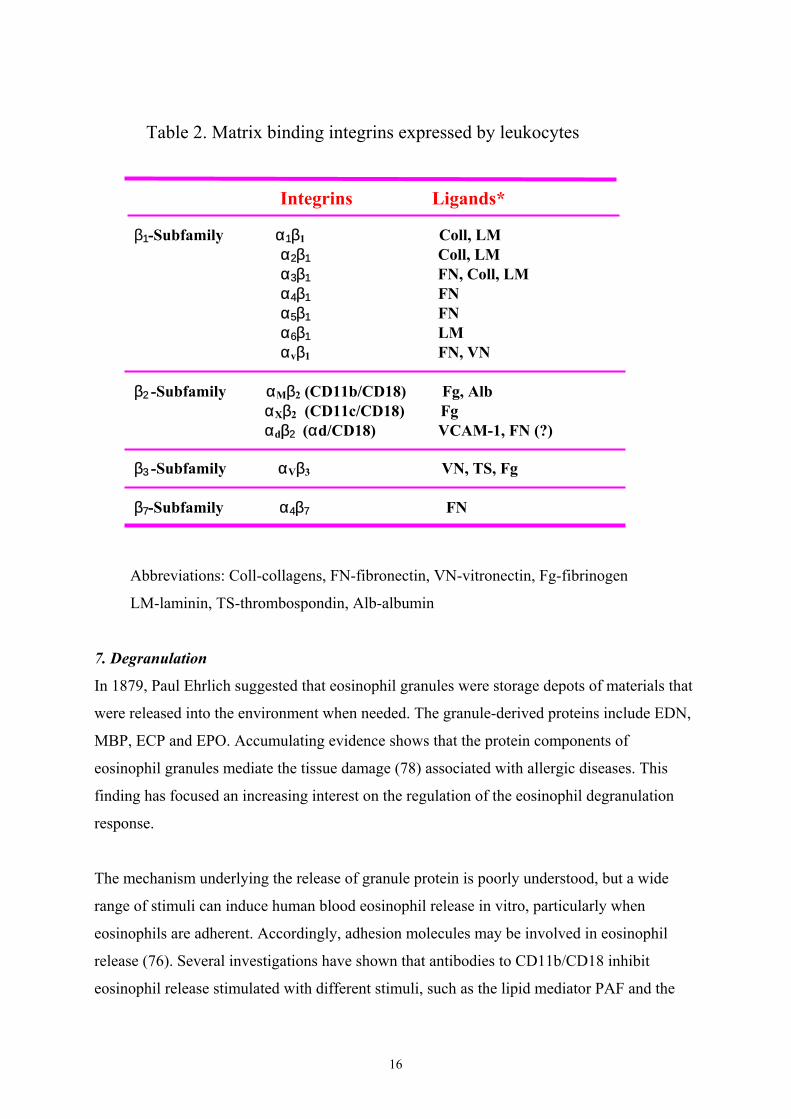
16
Table 2. Matrix binding integrins expressed by leukocytes
Abbreviations: Coll-collagens, FN-fibronectin, VN-vitronectin, Fg-fibrinogen
LM-laminin, TS-thrombospondin, Alb-albumin
7. Degranulation
In 1879, Paul Ehrlich suggested that eosinophil granules were storage depots of materials that
were released into the environment when needed. The granule-derived proteins include EDN,
MBP, ECP and EPO. Accumulating evidence shows that the protein components of
eosinophil granules mediate the tissue damage (78) associated with allergic diseases. This
finding has focused an increasing interest on the regulation of the eosinophil degranulation
response.
The mechanism underlying the release of granule protein is poorly understood, but a wide
range of stimuli can induce human blood eosinophil release in vitro, particularly when
eosinophils are adherent. Accordingly, adhesion molecules may be involved in eosinophil
release (76). Several investigations have shown that antibodies to CD11b/CD18 inhibit
eosinophil release stimulated with different stimuli, such as the lipid mediator PAF and the
β 1-Subfamily α1β1 Coll, LM α2β1 Coll, LM α3β1 FN, Coll, LM α4β1 FN α5β1 FN α6β1 LM αvβ1 FN, VN
β2 -Subfamily αMβ2 (CD11b/CD18) Fg, Alb αXβ2 (CD11c/CD18) Fg αdβ2 (αd/CD18) VCAM-1, FN (?)
β3 -Subfamily αVβ3 VN, TS, Fg
β7-Subfamily α4β7 FN
Integrins Ligands*

17
cytokine GM-CSF (60). The finding of eosinophil granule proteins in diseased tissue or on the
surface of the parasite, where few or no intact eosinophils were seen, has suggested eosinophil
degranulation to be associated with cell death (79). From the results described above, it is
evident that the degranulation process can be affected by adhesion. Furthermore, in resting
neutrophils, 75% of Mac-1 is localised in gelatinase granules (tertiary granules) and specific
granules, 20% in secretory vesicles and 5% on the plasma membrane (80). It is also
biologically important that exocytosis of those compartments is regulated by different signals.
8. Eosinophil Priming
Priming of an eosinophil response is defined as an increased reaction to a particular stimulus
by an agent that, in itself, induces little or no response. It has previously been demonstrated
that eosinophils in the peripheral blood of patients with allergic disorders have a primed
phenotype that resembles eosinophils from normal donors primed in vitro with cytokines (18).
Priming factors for eosinophils include neuropeptides (CGRP, substance P) and PAF,
chemokines like RANTES, IL-8 and eotaxin, but the most essential proteins are the
eosinophilopoietic cytokines, of which GM-CSF is the most potent and IL-5 the most
eosinophil-specific (21;18; 26).
Despite the knowledge of the importance of priming of eosinophils, little is known about the
signals that mediate this priming response. Furthermore, over the years the distinction
between primed and activated eosinophils has been somewhat unclear. The shift in density of
eosinophils toward light density or hypodense eosinophils is a well known phenomenon,
which is related to such eosinophilic diseases as allergic asthma, allergic rhinitis and parasitic
diseases (81-83). The low density eosinophils are presumed to represent primed eosinophils,
but the literature is far from conclusive (84).
9. Cytokines
Cytokines are low-molecular-weight glycoproteins that, when released at sites of
inflammation, induce their biological effect by binding to cytokine cell surface receptors.
They regulate a number of cellular functions important to allergic inflammation. Some of
these functions include the immunoglobulin E (IgE) synthesis, expression of adhesion
molecules by endothelium and inflammatory cell function (85). Eosinophils have functional

18
receptors for IL-3, GM-CSF and IL-5 that can regulate cellular function (degranulation,
adhesion, survival, superoxide production, etc.) (19;20).
Recent kinetic data from patients undergoing antigen challenge showed that IL-5 levels
increased between days 2 and 7 post challenge, whereas GM-CSF peaked at day 2 and
remained elevated through to day 16 (86). Furthermore, recruitment of eosinophils is
associated with elevated IL-5 in the airways of asthma patients (87). Blood eosinophils from
allergic subjects have an enhanced ability to transmigrate across endothelial cells in vitro, a
process that involves in vivo priming by IL-5 and related cytokines (21).
IL-5, IL-3 and GM-CSF act on eosinophils at three levels (88):
• Stimulation of the production from bone marrow
• Stimulation of functional activity
• Prolongation of survival of eosinophils at the inflammatory site
10. Cell death
Cell death is the process that culminates with cessation of biological activity. In contrast to
cell quiescence or dormancy, which is also characterised by decreased rates of many
biological activities, the changes that accompany cell death are irreversible. It is generally
accepted that apoptosis and necrosis are two distinct, mutually exclusive, modes of cell death.
Apoptosis is an active and physiological mode of cell death, in which the cell itself designs
and executes the program of its own demise and subsequent body disposal (89). This evidence
points to the importance of apoptosis as a component of processes designed to limit and
resolve inflammation.
Eosinophil apoptosis, like neutrophil apoptosis, occurs constitutively in vivo and the cells are
recognised by macrophages by a specific process without discharge of their histotoxic
contents. The regulation of eosinophil survival and apoptosis seems to play a major role in
diseases, demonstrating increased numbers of circulating and tissue eosinophils such as
allergic reactions (43). The presence of eosinophils or their granule proteins, or both, was
observed in diseased tissues or on the parasite’s surface. Frequently, few or no intact
eosinophils are seen despite extensive deposition of the eosinophil granule major basic protein
(MBP) and ECP in diseased tissues (90). This observation suggests that either the eosinophils
degranulate and leave the tissue or degranulate and die. However, some studies show that cell

19
death is not always accompanied by the classical features of either apoptosis or necrosis but
had features of both (91-93). It has been reported that PMA-treated neutrophils show
morphological changes quite different from those of typical apoptosis or necrosis (94).

20
AIMS OF THE STUDY
There were two aims of this investigation. The first was to develop a reliable, convenient and
specific method for the simultaneous analysis of eosinophil adhesion and release of ECP by
granulocytes that adhered to ECM. The second aim was to apply the method in experimental
and clinical studies. This would extend our knowledge of selective accumulation of different
types of granulocyte into the site of various inflammatory reactions. The specific aims of this
study were the following:
1. To compare the adhesion of eosinophils and neutrophils to fibronectin, fibrinogen and
albumin.
2. To study the relationship between eosinophil adhesion and release of ECP from
granulocytes that adhered to ECM.
3. To investigate the relationship between the release of ECP and eosinophil death.
4. To compare eosinophil adhesion and ECP release from granulocytes and purified
eosinophils that adhered to ECM.
5. To compare degranulation from primary and secondary granules of neutrophils that
adhered to ECM.
6. To compare the eosinophil priming in vivo and in vitro on eosinophil adhesion and
simultaneous release of ECP.
21
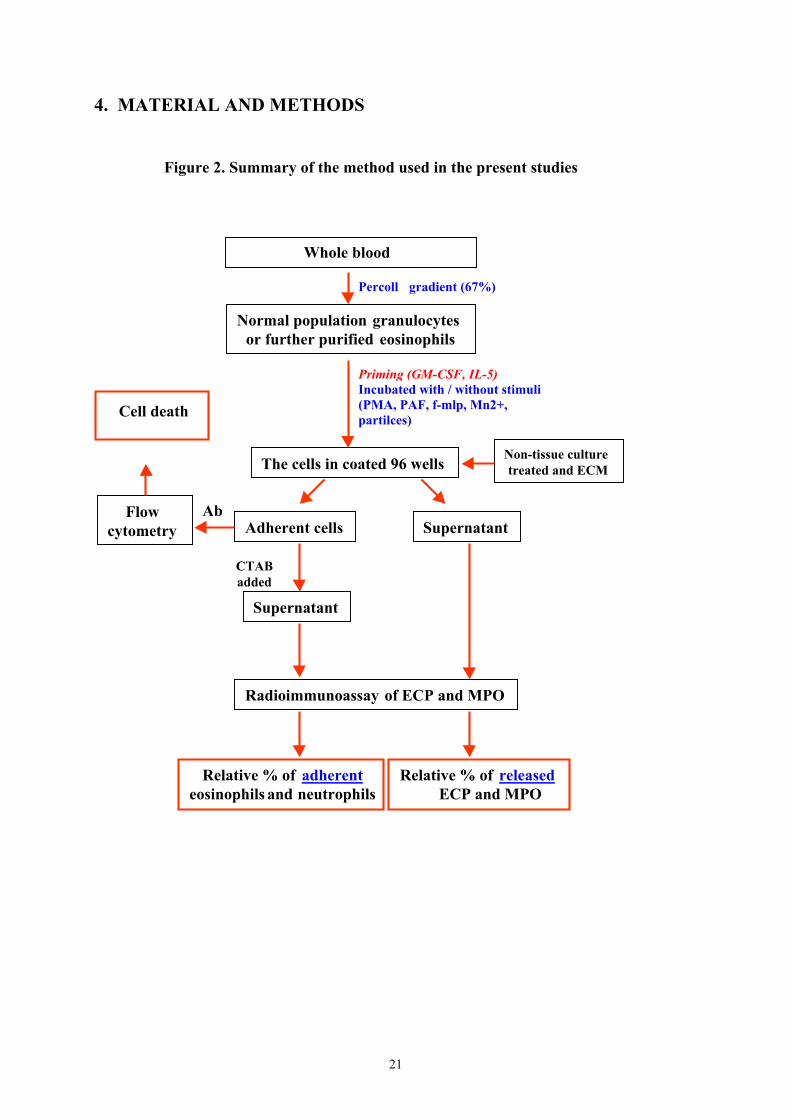
4. MATERIAL AND METHODS
Figure 2. Summary of the method used in the present studies
Whole blood
Normal population granulocytes or further purified eosinophils
The cells in coated 96 wells
Adherent cells
Radioimmunoassay of ECP and MPO
Relative % of adherenteosinophils and neutrophils
Percoll gradient (67%)
Priming (GM-CSF, IL-5)Incubated with / without stimuli(PMA, PAF, f-mlp, Mn2+,partilces)
Non-tissue culturetreated and ECM
Flowcytometry
Ab
Cell death
Supernatant
CTABadded
Supernatant
Relative % of released ECP and MPO

22
Isolation of granulocytes (papers I- IV)
Granulocytes were isolated from peripheral blood of non-allergic, normal volunteers or atopic
patients by Percoll (Pharmacia & Upjohn Diagnostic AB, Uppsala, Sweden) gradient
centrifugation as previously described (95;96). In a 50 mL albumin-coated tube ~18 mL
heparinized blood was mixed with an equal volume of PBS, and 15 mL Percoll (67%) was
applied at the bottom of the tube. After centrifugation at 1000 x g for 30 minutes at room
temperature, the mononuclear leukocytes on top of the Percoll gradient and the Percoll
gradient were removed. The remaining cell suspension in the bottom of the tube contained
granulocytes and erythrocytes. The erythrocytes were lysed by the addition of 40 mL of
NH4Cl (155 mmol/L) and incubated on ice for 15 minutes. The purified granulocytes (≥99%)
were washed twice with the assay buffer plus 1% (w/v) HSA and then suspended in the same
buffer. The cell concentration was 1 × 109 cells/L. The eosinophil content in the purified
granulocytes was 6% ± 2% (SD)
Purification of eosinophils (papers I, II)
Granulocytes, isolated as above, were incubated with anti-CD16-labelled magnetic beads in
PBS plus 2% NBS for one hour at 4° C (10 × 106 cells: 15 µl particles: 30 µl PBS + 2% NBS).
Resuspended cells (PBS + 2% NBS) were loaded onto the separation column positioned in a
strong magnetic field. The neutrophils bound to the anti-CD16 labelled magnetic beads
remained in the column while the eosinophils were eluted. The pure eosinophils (≥99%) were
diluted to a concentration corresponding to that in the granulocyte suspension from which
they were isolated.
Coating of wells (papers I-IV)
Ninety-six well, flat-bottom, non-tissue culture treated, high protein binding plates (Greiner
Labortechnik, Germany) were coated overnight at 37°C with 50 µg/ml fibronectin (both
plasma and tissue form) or albumin, or with 25 µg/ml fibrinogen (all matrix proteins were
from Sigma Chemical Co. St. Louis, Mo) in coating buffer, 100 µl/well. Before use, the wells
were washed twice in the assay buffer supplemented with 1% HSA. Finally, one more
washing was done with the same buffer in the absence or presence of Mn2+ (5 mmol/L).

23
Stimulation of cells (papers I-IV)
The stimulating agents tested were PMA, PAF, f-MLP, Mn2+ and serum-opsonized Sephadex
G-15 particles. The serum-opsonized Sephadex G-15 particles were prepared as described
previously (15). Suspensions containing 1 × 109 granulocytes/L and PMA, PAF, f-MLP or
Mn2+ at the concentrations of 10-9 mol/L, 10-7 mol/L, 10-7 mol/L and 5 mmol/L, respectively,
were added to the wells. For the particle stimulation, one volume of washed serum-opsonized
Sephadex G-15 particles was mixed with two volumes of granulocytes, with or without Mn2+
(5 mmol/L). The final concentration of cells was 1 × 109cells/L and the final concentration of
the serum-opsonized Sephadex G-15 particles was 27.8 g/L.
Priming of granulocytes (papers IV)
Granulocytes 1×109 cells/ml from normal donors were primed with IL-5 or GM-CSF for 20
minutes at 37°C in 5% CO2 in a humidified atmosphere. IL-5 was used at a concentration of
10-12 mol/L. GM-CSF was used at a concentration of 10-14 mol/L. The selected concentration
were found to be optimal for priming in preliminary experiments.
Adhesion assays (papers I- IV)
After incubation at 37° C in a humidified atmosphere containing 5% CO2 for the tested time,
the non-adherent cells were removed by three repeated washes with a room temperature assay
buffer. Adhering cells were then solubilized by the addition of a 100µl CTAB detergent
solution (5 g CTAB/L, 9 g NaCl/L). The supernatants of the extracts were harvested and
centrifuged at 600 × g for 10 minutes. The supernatants were collected again and frozen at
-20°C until assayed for the content of ECP (eosinophil marker) and MPO (neutrophil marker).
The total intracellular ECP and MPO contents extracted from granulocytes or from purified
eosinophils, solubilized with CTAB detergent solution, were measured in parallel. Results
from the adhesion experiments were expressed as the percentage of the total amount (100%)
of ECP and MPO in the respective population of eosinophils and neutrophils, which
corresponds to the relative amount of eosinophils and neutrophils.

24
Release assay (paper II- IV)
The samples for determination of the release of granule contents were collected from adhesion
experiments. After incubation, the supernatants were transferred to tubes before washing
away the non-adherent cells. These tubes were centrifuged at 600 × g for 10 minutes at room
temperature. The supernatants were carefully collected and mixed with an equal volume of
CTAB detergent solution and frozen at -20°C until assayed for ECP and MPO by
radioimmunoassay. The release of ECP was calculated according to the same principle as the
calculation of the relative amount of adherent eosinophils.
Radioimmunoassay (papers I- IV)
The ECP and MPO concentrations of the supernatants were analysed by a double antibody
competition radioimmunoassay(RIA) as described earlier (Pharmacia & Upjohn Diagnostics
AB, Uppsala, Sweden) (97;98).
Flow cytometry (paper II)
To observe the relationship between the release of granule contents and cell death
granulocytes were stimulated with different stimuli in plasma fibronectin or fibrinogen-coated
wells, as above. After incubation, optimally titrated FITC-labelled mAbs CD9 and propidium
iodide were added and afterwards plates were incubated for another 30 minutes at 4° C. The
EDTA-solution was added to detach the adherent cells (details are described in paper II). An
EPICS XL flow cytometer (Coulter Corporation, Miami, FL) was employed to analyse the
cells. Eosinophils were identified as CD9-positive cells (99). The relative amount of
eosinophils and neutrophils stained with propidium iodide was measured.
Assessment of granulocyte death (papers II- Ш)
The trypan blue exclusion assay was applied to assess the amount of dead granulocytes. Fifty
µl PMN suspension with different stimuli were incubated in plasma fibronectin- or
fibrinogen-coated wells for 90 minutes. After aspiration of the supernatant, 50 µl trypan blue
solution was added for 5 minutes and then aspirated. Next, 100 µl of PBS was added to each

25
well and the relative number of cells stained with trypan blue was evaluated blindly with an
inverted microscope.
Inhibition of release and adhesion by monoclonal antibodies (paper II)
The inhibitory effect of mAbs against CD18 and CD49d was tested by co-incubation of
granulocytes or purified eosinophils with antibodies (final concentration 10 µl/ml) and
activating agents for 90 minutes.
Patients (paper IV)
Eleven birch pollen-atopic patients with asthma (seven men and four women), with a mean
age of 39 years (range, 20 to 51 years), were selected for the study. The patients were
considered atopic because of positive skin tests to birch allergen. None of the patients were
smokers and all patients had a positive methacholine test. Blood was collected during early
pollen season from March to April. The range of eosinophil counts was 60 - 520 x 106/L
(median 210 x 106/L).
References (paper IV)
The reference group consisted of 9 healthy, non-allergic, laboratory employees and normal
blood donors matched for age and sex with the patients. The range of eosinophil counts was
60 – 160 x 106/L (median 135 x 106/L).
Statistical analysis (papers I- IV)
Statistical analyses were performed using Student’s paired t test, one-way ANOVA, Mann-
Whitney U test, and Wilcoxon’s paired test. Statistically significant differences were assumed
when p< 0.05. All calculations were done using CSS Statistica (Statsoft Inc., Tulsa, OK).

26
5. RESULTS AND DISCUSSION
Development of a method for the simultaneous analysis of eosinophil adhesion and ECP
release (I,II)
The measurement of ECP and MPO has been used in earlier research to quantitatively
estimate eosinophil and neutrophil involvement (97;98). In the present study, the number of
adherent eosinophils and neutrophils was quantified indirectly by expressing the amount of
eosinophil cationic protein (ECP) and myeloperoxidase (MPO) extracted from the adherent
fraction of cells in percentage of the total amount of ECP and MPO extracted from the
granulocytes added to the wells. Degranulation was quantified in the same way.
The cellular content of ECP has been estimated to be 10 µg/106 eosinophils (100). The normal
level of eosinophils in peripheral blood is about 1-4% of all leukocytes. The RIA measures
ECP within the range of 2-200 µg/L. In all experiments, the concentration of 1 x
109granulocytes/L was used. Thus, although there was only 1% of eosinophil adhesion or
release, the concentration of ECP in supernatants was enough for measurement by the RIA.
The adhesion of purified eosinophils was compared with that of eosinophils in the normal
granulocyte population. At a short incubation time (15 minutes), eosinophils in the
granulocyte population showed a significantly higher basal adhesion to both fibronectin and
fibrinogen. The results indicate that the dominating presence of neutrophils might induce a
short time enhancement of the spontaneous adhesion of eosinophils. After a prolonged
incubation period (90 minutes), purified eosinophils evidenced a higher adhesion to
fibronectin induced by Mn2+ and to fibrinogen induced by PMA as compared with eosinophils
in the granulocyte population. Comparison of the release of ECP from granulocytes and
purified eosinophils adhered to fibronectin and fibrinogen showed a higher release of ECP
from purified eosinophils than that from eosinophils when induced by serum-treated
Sephadex G-15 particles. The overall difference between isolated eosinophils and eosinophils
in the normal granulocyte population was minor. The results indicate that the pattern of
responses of isolated eosinophils versus eosinophils in the granulocyte population to the
different stimuli was similar. In conclusion, the method allows the measurement of eosinophil
adhesion and degranulation by using granulocytes.

27
The effects of PMA, f-MLP and Mn2+ on eosinophil and neutrophil adhesion (I)
Depending on both the stimulus and the protein that the cells reacted to, induction of adhesion
by the activation of integrins revealed different patterns of eosinophil and neutrophil adhesion
to tissue and plasma fibronectin, albumin and fibrinogen. When the incubation time was 105
minutes, Mn2+ preferentially enhanced eosinophil adhesion compared with neutrophil
adhesion in the presence of these four proteins. Furthermore, background adhesion of
eosinophils to albumin was higher as compared with the other three proteins. These data are
compatible with the previous finding that approximately 50% of the eosinophils adhered to
albumin-Sepharose or albumin-coated plastic surfaces, as compared with only 25-30% of the
isolated neutrophils (101). The level of eosinophil adhesion, however, is lower in our study
than the level reported in the above study. The difference might be due to the use of
eosinophils purified to varying degrees and by slightly different procedures. Furthermore,
tissue fibronectin supported a greater eosinophil and neutrophil adhesion than plasma
fibronectin. This may be explained by the fact that tissue fibronectin, in comparison with
plasma fibronectin, contains approximately 50% more of the CS-1 cell binding region
recognised by the integrin α4β1 (102;103). The number of RGD cell binding domains
recognised by the integrin α5β1 (104) in tissue fibronectin is not higher than in plasma
fibronectin, however (27). This observation means that there are probably other structural
differences between plasma and tissue fibronectin to explain this phenomenon.
It has been demonstrated that PKC is important for eosinophil activation. For example, the
PKC activator PMA induces a variety of eosinophil functions, such as adhesion (105),
respiratory burst (106) and degranulation (107). In the present studies, PMA induced a similar
pattern of adhesion of both eosinophils and neutrophils to the four tested proteins. The results
are in agreement with studies that have shown that human neutrophils and eosinophils from
the peripheral circulation had similar average levels of activable PKC (108) and that PMA
removed the environmental restriction on eosinophil respiratory burst (109). This is probably
because PMA induces a direct stimulation of PKC of the cells, which would allow that the
receptor mechanism is bypassed (110).

28
It has been reported that chemoattractants differentially regulate the avidity of β1 and β2
integrins in eosinophils. For example, f-MLP stimulated the expression of an activation-
specific Mac-1 epitope defined by the CBRM1/5 mAB on the surface of eosinophils, but did
not upregulate the 9EG7 neoepitope, which is related to the β1 subunit (111). On the other
hand, neutrophils mobilise Mac-1 to a greater extent when responding to receptor-dependent
factors related to bacterial infections, although eosinophils had a larger total pool of Mac-1
than neutrophils (112). In accordance with these results, the present study showed that f-MLP
induced a very early and short-term increment of the adhesion of neutrophils to fibrinogen
and albumin but not to fibronectin. A lower f-MLP induced adhesion of eosinophils to
fibrinogen was also observed. Moreover, PMA amplified the effect of Mn2+ on the adhesion of
both eosinophils and neutrophils to these four proteins suggesting that Mn2+ and PMA exert
synergistic effects on granulocyte adhesion. This event is in contrast to Mn2+ in combination
with f-MLP which had no synergetic effect on granulocyte adhesion.
The effect of eosinophil adhesion on the release of ECP (II)
Evidence suggests that ligation of the β2 integrins triggers activation and degranulation of
human eosinophils (76) and that ECM can attenuate activation and degranulation of
stimulated eosinophils (11). These findings clearly indicate that cellular adhesion is important
for cellular degranulation. Our findings extend the knowledge that adhesion molecules are not
simply involved in cell-cell or cell-matrix interactions but may also modulate selective cell
functions (72). Manganese ions (0.5-15mmol/L), which are known to activate integrins
directly (113), did not induce any ECP release, regardless of the degree of adhesion and to
which protein the cells adhered. The results could be explained by the results of the study
performed in cell suspensions. That study demonstrated that Mn2+ had dual effects: it
stimulated the ECP release in the absence of Mg2+ and Ca2+ and it inhibited the release in the
presence of these actions (114). In our system, the buffer contains Mg2+ and Ca2+. PMA alone
induced a dose-dependent eosinophil degranulation, independent of whether the eosinophils
were adhering to fibronectin or fibrinogen. The implication of these results is that PMA
removes the influence of ECM on eosinophil degranulation, which has been demonstrated to
attenuate activation and degranulation of eosinophils stimulated by PAF (11). An intriguing
finding was that PMA at the concentration of 10-9 mol/L did not induce any significant release
of ECP, but this concentration of PMA in combination with Mn2+ (5 mmol/L) dramatically

29
enhanced the release of ECP. Furthermore, antibody inhibition studies showed that the release
of ECP induced by PMA and Mn2+ in combination was dependent on β2 integrins on
granulocytes that adhered to fibronectin and fibrinogen, whereas the adhesion of eosinophils
to fibronectin could be inhibited by both anti-β1 (α4) and anti-β2 (CD18) monoclonal
antibodies. These results indicate that adhesion and degranulation can be regulated by
different signalling pathways. The anti-CD18 inhibited adhesion of eosinophils to fibronectin,
previously considered β1 dependent, may be explained by the newly discovered αdβ2 integrin
(115). This integrin was recently described to function as an alternative ligand for VCAM-1
and, in analogy with VLA-4, might also function as a ligand to fibronectin (57).
A small but significant release of ECP was found when eosinophils adhering to fibronectin
and HSA, but not to fibronectin, were stimulated by f-MLP. Earlier investigations have
demonstrated that eosinophils adhere to fibronectin via β1 and β7 integrins, to fibrinogen via
β1 and β2 integrins and to HSA via β2 integrins (61;75;111;116-119). Consequently, the
difference in f-MLP induced release of ECP could plausibly be explained by a differential
regulation of β1 and β2 integrins (111;112). In another study it was found that when adherent
to laminin- and fibronectin-coated wells, degranulation of eosinophils (induced by PAF and
C5a) was reduced by about 50% compared with cells adherent to non-coated, HSA-blocked
wells (11). Furthermore, Mn2+ suppressed f-MLP induced eosinophil degranulation.
When induced by serum-opsonized Sephadex particles, extracellular release of approximately
12-14% of ECP from eosinophils occurred in the presence of both plasma fibronectin and
fibrinogen. In the presence of the serum-opsonized particles adhesion to protein-coated wells
was decreased. This preferential adherence to the particles could be explained by sterical
hindrance in the sense that the particles prevent contact between eosinophils and
fibronectin/fibrinogen. Another probable mechanism is the competition between C3b particles
and fibronectin/fibrinogen in the binding to eosinophils. In a study on the release induced by
serum-opsonized Sephadex particles, almost all eosinophils were found to adhere to the
particles (101). Furthermore, ECP release appears to be associated with cell death when it is
induced by PMA plus Mn2+ or serum-opsonized Sephadex particles. One earlier study has
demonstrated that if stimulated by protein-coated beads, eosinophil degranulation is
accompanied by cellular injury and necrosis (79). Further investigations are needed to
confirm the kind of cell death in our study, i.e. apoptosis or necrosis.

30
Comparison of purified eosinophils and eosinophils in the normal population of granulocytes
regarding the release of ECP in general demonstrated the same pattern of responses. The only
difference was that serum-opsonized Sephadex particles stimulated a slightly, but
significantly, higher release of ECP from purified eosinophils than eosinophils in the normal
population of granulocytes.
The effect of neutrophil adhesion on degranulation of primary and secondary granules
(Ш)
Neutrophils are the first cell type to enter the lung after allergen challenge and another major
participant in late phase reactions (30). It has also been shown that neutrophils help eosinophil
migration across lung epithelium (120). Furthermore, neutrophil release granule constituents,
lipid-derived mediators, and reactive oxygen intermediates (ROI) which are involved in
killing of pathogens and tissue damage (121).
In the present study, we found that PMA, in combination with Mn2+, induced a marked release
of ∼ 80% of the intracellular content of lactoferrin and HNL after 60-75 minutes of incubation,
regardless to which protein the cells were associated. Under the same conditions, a maximal
release of MPO of 15-20% was obtained after a 90-minute stimulation of neutrophils by PMA
and Mn2+. This finding is agreement with a report describing the hierarchy of the mobilisation
of granule populations. This hierarchy leads to a higher release of lactoferrin and HNL from
the secondary granules as compared with MPO from the primary granules (48). The
biological relevance of the observed lag-phase between primary and specific granule
degranulation could be to facilitate cell adhesion and migration to inflammatory sites.
Adhesion molecules are considered as stores in specific granules (80) and exocytosis of
gelatinase from tertiary granules may facilitate degradation of type IV collagen in basement
membranes. Depending on the protein that the cells were plated on, PMA or f-MLP alone
induced 30-40% release of lactoferrin and HNL after 60 minutes of incubation; fibrinogen
and HSA were observed to support a higher release of lactoferrin than fibronectin. The
difference can likely be explained by differences in signalling from β2 and β1 integrins.
According to recent findings, adhesion to albumin is mediated by CD11b/CD18 (122),
whereas adhesion to fibronectin is mediated by VLA-5 and possibly αdβ2 (115;123). Lastly,

31
adhesion to fibrinogen is mediated by both CD11b/CD18 and VLA-5 (61;74;75). It is also
known that neutrophils from patients with leukocyte adhesion deficiency (LAD) lacking
CD18, are unable to spread on thrombospondin, and also fail to release lactoferrin in response
to f-MLP (124).
Adhesion and release of lactoferrin and HNL were quantitatively related when induced by
PMA and PMA plus Mn2+, but not when they were induced by f-MLP. This pattern may result
because f-MLP is more of a chemoattractant and preferentially induces migration, whereas
PMA directly activates proteinkinase C, which would by-pass the early steps in physiological
ligand-receptor activation of PMN (125). Earlier study has reported that concanavalin A (Con
A) at low doses is a potent stimulus for adhesion and does not induce any O2- production,
whereas at high doses it stimulate both adhesion and O2- production (126). The mean release
of lactoferrin and HNL induced by different stimuli showed a significant negative relationship
to the viability of cells. Thus, adhesion is important for the regulation of degranulation, but
adhesion and degranulation are not necessarily quantitatively related. The results also indicate
that neutrophil responses have various transductional pathways that may depend on the type
of stimulus.
The effect of priming in vivo or in vitro on eosinophil adhesion and release of ECP (IV)
Granulocytes obtained from pollen-atopic patients during early pollen season were stimulated
by PMA, Mn2+ and serum-opsonized particles. Afterwards, adhesion and release of ECP from
eosinophils were measured. In an attempt to mimic the situation in vivo, IL-5 and GM-CSF
were used to prime normal eosinophils in vitro and the effect on eosinophil adhesion and
release further stimulated by PMA, PAF and Mn2+ was investigated.
When stimulated by PMA, eosinophils from pollen-atopic patients during early pollen season
exhibited greater adhesion to fibronectin and fibrinogen as compared with eosinophils from
the reference group. In addition, eosinophils from pollen-atopic patients during early pollen
season displayed greater release of ECP than eosinophils from the reference group. The other
stimuli did not induce this kind of differential response. These findings indicate that
eosinophils from atopic patients during early pollen season already possess a higher PKC
activity. This view concurs with the finding that low-density eosinophils from allergic
asthmatic patients possess higher PKC activity than high-density eosinophils (108). The

32
finding of a higher proportion of low-density blood eosinophils was among the earliest
evidence suggesting a primed eosinophil phenotype in allergic disease was (21). It has also
been shown that there is an increase in the expression of the PKC ζ isoform in eosinophils
after allergen challenge (127). The evidence from all these studies suggests that protein kinase
C (PKC) is involved in eosinophil priming in vivo.
The priming is likely to be caused by IL-3, IL-5 and GM-CSF in the peripheral blood, where
IL-5 is especially important in the recruitment and activation of eosinophils. It has been
demonstrated that IL-5 mRNA and protein levels were elevated in lung tissue and
bronchoalveolar lavage fluid from symptomatic asthma patients (18;128). Priming with IL-5
caused a significantly higher adhesion to fibrinogen and simultaneous release of ECP in
response to PMA at 90 minutes. In the same conditions simulation with PMA plus Mn2+ for
90 minutes enhanced the adhesion to fibrinogen. These differences between eosinophils
primed in vivo versus in vitro can be explained by varying effects of short- versus long-term
priming (129) and that eosinophils from the patients have been primed in vivo by
combinations of cytokines. It has been found that allergen challenge induces the late-phase
nasal responses and the local expression of Th2-type cytokines, IL-4 and IL-5 (130;131), as
well as proinflammatory cytokines such as IL-6 (132).
GM-CSF, which shares a common β chain of the receptor with IL-5, was observed to prime
eosinophil adhesion to fibronectin and fibrinogen in response to PAF or PMA plus Mn2+,
whereas IL-5 did not prime this kind of response. The results indicate that IL-5 and GM-CSF
can signal through cytokine-specific pathways not dependent on the common β chain.
Furthermore, priming with GM-CSF only slightly affected the release of ECP. IL-5, but not
GM-CSF, was found to prime eosinophil generation of O2- and intracellular calcium
movement (133); however, an increased release of ECP by eosinophils upon short-term (20
minutes) priming with GM-CSF and stimulation with PAF has also been reported (134) . It is
likely that different experimental conditions are responsible for these apparently conflicting
results.
In conclusion, eosinophils from pollen-atopic patients primed in vivo and normal eosinophils
primed in vitro displayed similar, although not identical, patterns of upregulation of

33
eosinophil adhesion and release. The fact that the responses to PMA were primed both in vivo
and in vitro suggests that PKC activity is involved in priming of eosinophils.

34
6. CONCLUSIONS1. Normal population granulocytes can be used to measure eosinophil adhesion and
degranulation.
2. Mn2+ preferentially enhanced eosinophil adhesion compared with neutrophil adhesion,
though it did not induce any release of ECP.
3. PMA stimulated a dose-dependent eosinophil adhesion and degranulation, regardless to
which protein the eosinophils adhered. Mn2+ amplified the effects of PMA on eosinophil
adhesion and degranulation.
4. When stimulated by f-MLP, eosinophil adhesion and degranulation were affected by the
matrix proteins to which eosinophils adhered. Co-stimulation with f-MLP and Mn2+
exhibited different effects on eosinophil adhesion and degranulation.
5. When induced by PMA and Mn2+, eosinophil degranulation was β2 integrin-dependent,
regardless of whether the eosinophils adhered to fibronectin or fibrinogen; adhesion to
fibrinogen was β2 integrin-dependent and adhesion to fibronectin was β1 and β2 integrin-
dependent.
6. Adhesion modulates neutrophil degranulation, but it is not always quantitatively related or
related in time.
7. PKC is involved in the priming of eosinophils.

35
7. ACKNOWLEDGEMENTSI would like to express my sincere gratitude to everyone who contributed to this work and in
particular to the following persons:
Associate Professor Lena Håkansson, for being my tutor and supervisor, for introducing me to
the field of granulocyte adhesion, for giving me the privilege of sharing her plentiful, unique
knowledge in cellular adhesion science and flow cytometer, for her patience in reading and
revising my manuscripts and for her concern about my daily life.
Professor Per Venge, head of the Department of Clinical Chemistry, my supervisor, for
sharing his vast knowledge of science and for giving me a chance to work and study in a
friendly and inspiring atmosphere which he has worked hard to create.
Gunilla Strömstedt and Barbro Bjurhäll, for offering superb secretarial assistance.
Agneta Trulson, my former roommate, for friendship and kindness.
Kerstin Lindblad, Ing-Britt Persson, Lena Moberg and Ulla-Britt Jönsson, for their excellent
work in interpreting Swedish into English and narrating Swedish culture and customs.
Charlotte Woschnagg, Jonas Byström, Kristina Sandström, Malgorzata Karawajczyk and
Maria Lampinen, for skilful technical assistance and friendship.
Lixin Liu, Linshu Zhao and Shengyuan Xu, my Chinese colleagues, for their friendship and
reminding me of my homeland.
All the staff at the Department of Clinical Chemistry for their warm friendship.
Yangzi Liu, my dear daughter, for creating a rich and colourful life for me outside the
laboratory during these years in Sweden.
Zhurong Liu, my husband, for his love, understanding and continuous support.

36
My parents, sisters and brother, for their expectations, encouragement and taking care of my
daughter in times of need.
This work was supported by grants from the following sources:
The Medical Faculty of the University of Uppsala
The National Association against Asthma and Allergy
The Swedish Foundation for Health Care and Allergy Research

37
REFERENCES
1. Nagata,M., Sedgwick,J.B., and Busse,W.W. 1995. Differential effects of granulocyte-macrophage colony- stimulating factor on eosinophil and neutrophil superoxide aniongeneration. J.Immunol. 155:4948-4954.
2. Motegi,Y. and Kita,H. 1998. Interaction with secretory component stimulates effectorfunctions of human eosinophils but not of neutrophils. J.Immunol. 161:4340-6.
3. Resnick,M.B. and Weller,P.F. 1993. Mechanisms of eosinophil recruitment.Am.J.Respir.Cell Mol.Biol. 8:349-355.
4. Wardlaw,A.J., Symon,F.S., and Walsh,G.M. 1994. Eosinophil adhesion in allergicinflammation. J.Allergy Clin.Immunol. 94:1163-1171.
5. Weller,P.F. 1997. Human eosinophils. J.Allergy Clin.Immunol. 100:283-287.
6. Pakianathan,D.R. 1995. Extracellular matrix proteins and leukocyte function.J.Leukoc.Biol. 57:699-702.
7. Kasahara,K., Strieter,R.M., Chensue,S.W., Standiford,T.J., and Kunkel,S.L. 1991.Mononuclear cell adherence induces neutrophil chemotactic factor/interleukin-8 geneexpression. J.Leukoc.Biol. 50:287-295.
8. Dubucquoi,S. 1994. Interleukin 5 synthesis by eosinophils: association with granulesand immunoglobulin-dependent secretion. J.Exp.Med. 179:703-8.
9. Cara,D.C., Negrao-Correa,D., and Teixeira,M.M. 2000. Mechanisms underlyingeosinophil trafficking and their relevance in vivo. Histol.Histopathol. 15:899-920.
10. Reimert,C.M., Skov,P.S., and Poulsen,L.K. 1998. A microtiter assay for activation ofeosinophils. Simultaneous monitoring of eosinophil adhesion and degranulation. Allergy53:129-138.
11. Kita,H., Horie,S., and Gleich,G.J. 1996. Extracellular matrix proteins attenuateactivation and degranulation of stimulated eosinophils. J.Immunol. 156:1174-1181.
12. Anwar,A.R., Moqbel,R., Walsh,G.M., Kay,A.B., and Wardlaw,A.J. 1993. Adhesion tofibronectin prolongs eosinophil survival. J.Exp.Med. 177:839-843.
13. Goldring,K. and Warner,J.A. 1997. Cell matrix interactions in asthma. Clin.Exp.Allergy27:22-27.
14. Gosset,P. 1995. Expression of E-selectin, ICAM-1 and VCAM-1 on bronchial biopsiesfrom allergic and non-allergic asthmatic patients. Int.Arch.Allergy Immunol. 106:69-77.
15. Carlson,M., Håkansson,L., Peterson,C., Stalenheim,G., and Venge,P. 1991. Secretion ofgranule proteins from eosinophils and neutrophils is increased in asthma. J.AllergyClin.Immunol. 87:27-33.

38
16. Koller,D.Y., Urbanek,R., and Gotz,M. 1995. Increased degranulation of eosinophil andneutrophil granulocytes in cystic fibrosis. Am.J.Respir.Crit.Care Med. 152:629-633.
17. Romagnani,S. 1997. The Th1/Th2 paradigm. Immunol.Today 18:263-266.
18. Koenderman,L., van der Bruggen,T., Schweizer,R.C., Warringa,R.A., Coffer,P.,Caldenhoven,E., Lammers,J.W., and Raaijmakers,J.A. 1996. Eosinophil priming bycytokines: from cellular signal to in vivo modulation. Eur.Respir.J.Suppl. 22:119s-125s.Review.
19. Sung,K.L., Li,Y., Elices,M., Gang,J., Sriramarao,P., and Broide,D.H. 1997.Granulocyte-macrophage colony-stimulating factor regulates the functional adhesivestate of very late antigen-4 expressed by eosinophils. J.Immunol. 158:919-927.
20. Horie,S. 1996. Cytokines directly induce degranulation and superoxide production fromhuman eosinophils. J.Allergy Clin.Immunol. 98:371-81.
21. Sampson,A.P. 2001. IL-5 priming of eosinophil function in asthma. Clin.Exp.Allergy31:513-7.
22. in 't Veen,J.C. 1998. CD11b and L-selectin expression on eosinophils and neutrophils inblood and induced sputum of patients with asthma compared with normal subjects.Clin.Exp.Allergy. 28:606-15.
23. Lamblin,C., Gosset,P., Tillie-Leblond,I., Saulnier,F., Marquette,C.H., Wallaert,B., andTonnel,A.B. 1998. Bronchial neutrophilia in patients with noninfectious statusasthmaticus. Am.J.Respir.Crit.Care Med. 157:394-402.
24. Lazarus,S.C. 1998. Inflammation, inflammatory mediators, and mediator antagonists inasthma. J.Clin.Pharmacol. 38:577-82.
25. Gleich,G.J. 1990. The eosinophil and bronchial asthma: current understanding. J.AllergyClin.Immunol. 85:422-36.
26. Sedgwick,J.B., Quan,S.F., Calhoun,W.J., and Busse,W.W. 1995. Effect of interleukin-5and granulocyte-macrophage colony stimulating factor on in vitro eosinophil function:comparison with airway eosinophils. J.Allergy Clin.Immunol. 96:375-85.
27. Walsh,G.M., Symon,F.A., and Wardlaw,A.J. 1995. Human eosinophils preferentiallysurvive on tissue fibronectin compared with plasma fibronectin. Clin.Exp.Allergy25:1128-1136.
28. Wardlaw,A.J., Brightling,C., Green,R., Woltmann,G., and Pavord,I. 2000. Eosinophilsin asthma and other allergic diseases. Br.Med.Bull. 56:985-1003.
29. Silberstein,D.S. 1995. Eosinophil function in health and disease.Crit.Rev.Oncol.Hematol. 19:47-77.
30. Kuby,J. 1997. Hypersentive reactions. In Immunology. W. H. Freeman and Company,New York. 413-439.

39
31. Calhoun,W.J. 1991. The role of eosinophils in the pathophysiology of asthma.Ann.N.Y.Acad.Sci. 629:62-72.
32. Broide,D.H. 2001. Molecular and cellular mechanisms of allergic disease. J.AllergyClin.Immunol. 108:S65-S71.
33. Schatz,M. 1982. The eosinophil and the lung. Arch.Intern.Med. 142:1515-9.
34. Young,J.D., Peterson,C.G., Venge,P., and Cohn,Z.A. 1986. Mechanism of membranedamage mediated by human eosinophil cationic protein. Nature 321:613-616.
35. Ayars,G.H., Altman,L.C., McManus,M.M., Agosti,J.M., Baker,C., Luchtel,D.L.,Loegering,D.A., and Gleich,G.J. 1989. Injurious effect of the eosinophil peroxide-hydrogen peroxide-halide system and major basic protein on human nasal epithelium invitro. Am.Rev.Respir.Dis. 140:125-131.
36. Lee,T.H. and Austen,K.F. 1986. Arachidonic acid metabolism by the 5-lipoxygenasepathway, and the effects of alternative dietary fatty acids. Adv.Immunol. 39:145-75.:145-175.
37. Shaw,R.J., Walsh,G.M., Cromwell,O., Moqbel,R., Spry,C.J., and Kay,A.B. 1985.Activated human eosinophils generate SRS-A leukotrienes following IgG-dependentstimulation. Nature 316:150-152.
38. Sehmi,R. and Denburg,J.A. 2000. Differentiation of human eosinophils. Role in allergicinflammation. Chem.Immunol. 76:29-44.:
39. Denburg,J.A. 1999. Bone marrow in atopy and asthma: hematopoietic mechanisms inallergic inflammation. Immunol.Today 20:111-113.
40. Bochner,B.S. 1994. The role of adhesion molecules in human eosinophil and basophilrecruitment. J.Allergy Clin.Immunol. 94:427-38; quiz 439.
41. Baggiolini,M. and Loetscher,P. 2000. Chemokines in inflammation and immunity.Immunol.Today 21:418-420.
42. Alam,R. 1997. Chemokines in allergic inflammation. J.Allergy Clin.Immunol. 99:273-277.
43. Simon,H.U. 1998. Eosinophil apoptosis in allergic diseases--an emerging new issue.Clin.Exp.Allergy 28:1321-1324.
44. Smith,H.R., Larsen,G.L., Cherniack,R.M., Wenzel,S.E., Voelkel,N.F., Westcott,J.Y.,and Bethel,R.A. 1992. Inflammatory cells and eicosanoid mediators in subjects with lateasthmatic responses and increases in airway responsiveness. J.Allergy Clin.Immunol.89:1076-84.
45. Teran,L.M., Carroll,M., Frew,A.J., Montefort,S., Lau,L.C., Davies,D.E., Lindley,I.,Howarth,P.H., Church,M.K., and Holgate,S.T. 1995. Neutrophil influx and interleukin-8release after segmental allergen or saline challenge in asthmatics. Int.Arch.AllergyImmunol. 107:374-5.

40
46. Casale,T.B., Costa,J.J., and Galli,S.J. 1996. TNF alpha is important in human lungallergic reactions. Am.J.Respir.Cell Mol.Biol. 15:35-44.
47. Sampson,A.P. 2001. The role of eosinophils and neutrophils in inflammation.Clin.Exp.Allergy 30 Suppl 1:22-7.:22-7.
48. Borregaard,N. and Cowland,J.B. 1997. Granules of the human neutrophilicpolymorphonuclear leukocyte. Blood. 89:3503-21.
49. Borregaard,N. 1996. Current opinion about neutrophil granule physiology. Curr OpinHematol 3:11.
50. Tourkin,A., Anderson,T., LeRoy,E.C., and Hoffman,S. 1993. Eosinophil adhesion andmaturation is modulated by laminin. Cell Adhes.Commun. 1:161-176.
51. Suchard,S.J. 1993. Interaction of human neutrophils and HL-60 cells with theextracellular matrix. Blood Cells. 19:197-221.
52. Nagy,B., Katona,E., Erdei,J., Szekely,E., Marialigeti,T., Karmazsin,L., and Fachet,J.1988. Fibronectin in bronchoalveolar lavage fluid and plasma from children withchronic inflammation of lungs. Acta Paediatr.Scand. 77:727-733.
53. Roman,J. 1996. Extracellular matrix and lung inflammation. Immunol.Res. 15:163-178.
54. Bevilacqua,M.P. 1993. Selectins. J.Clin.Invest. 91:379-87.
55. Gahmberg,C.G., Valmu,L., Kotovuori,A., Kotovuori,P., Hilden,T.J., Fagerholm,S.,Kantor,C., Nurminen,T., Ihanus,E., and Tian,L. 1999. Leukocyte adhesion--anintegrated molecular process at the leukocyte plasma membrane. Biosci.Rep. 19:273-281.
56. Seminario,M.C. and Bochner,B.S. 1997. Expression and function of beta 1 integrins onhuman eosinophils. Mem.Inst.Oswaldo Cruz 92 Suppl 2:157-64.:157-164.
57. Matsumoto,K., Sterbinsky,S.A., Bickel,C.A., Zhou,D.F., Kovach,N.L., andBochner,B.S. 1997. Regulation of alpha 4 integrin-mediated adhesion of humaneosinophils to fibronectin and vascular cell adhesion molecule-1. J.AllergyClin.Immunol. 99:648-656.
58. Wegner,C.D., Rothlein,R., and Gundel,R.H. 1991. Adhesion molecules in thepathogenesis of asthma. Agents Actions Suppl. 34:529-544.
59. Freyer,D.R., Boxer,L.A., Axtell,R.A., and Todd,R.F.3. 1988. Stimulation of humanneutrophil adhesive properties by adenine nucleotides. J.Immunol. 141:580-586.
60. Horie,S. and Kita,H. 1994. CD11b/CD18 (Mac-1) is required for degranulation ofhuman eosinophils induced by human recombinant granulocyte-macrophage colony-stimulating factor and platelet-activating factor. J.Immunol. 152:5457-5467.

41
61. Altieri,D.C., Agbanyo,F.R., Plescia,J., Ginsberg,M.H., Edgington,T.S., and Plow,E.F.1990. A unique recognition site mediates the interaction of fibrinogen with theleukocyte integrin Mac-1 (CD11b/CD18). J.Biol.Chem. 265:12119-12122.
62. Hemler,M.E. 1990. VLA proteins in the integrin family: structures, functions, and theirrole on leukocytes. Annu.Rev.Immunol. 8:365-400.
63. Elices,M.J. 1990. VCAM-1 on activated endothelium interacts with the leukocyteintegrin VLA-4 at a site distinct from the VLA-4/fibronectin binding site. Cell. 60:577-84.
64. Tachimoto,H. and Bochner,B.S. 2000. The surface phenotype of human eosinophils.Chem.Immunol. 76:45-62.:45-62.
65. Wakugawa,M., Nakagawa,H., Yamada,N., and Tamaki,K. 1996. Chronologic analysisof eosinophil granule protein deposition and cell adhesion molecule expression in miteallergen-induced dermatitis in atopic subjects. Int.Arch.Allergy Immunol. 111 Suppl 1:5-11.
66. Teixeira,M.M., Williams,T.J., and Hellewell,P.G. 1995. Mechanisms andpharmacological manipulation of eosinophil accumulation in vivo.Trends.Pharmacol.Sci. 16:418-423.
67. Wein,M. and Bochner,B.S. 1993. Adhesion molecule antagonists: future therapies forallergic diseases? [editorial]. Eur.Respir.J. 6:1239-1242.
68. Gumbiner,B.M. 1996. Cell adhesion: the molecular basis of tissue architecture andmorphogenesis. Cell 84:345-57.
69. Springer,T.A. 1990. Adhesion receptors of the immune system. Nature 346:425-34.
70. Hickey,P.R. and McGowan,F.X. 1995. Adhesion molecules and inflammation: the nexttargets for perioperative organ protection? Anesth.Analg. 81:1123-4.
71. Wei,J., Shaw,L.M., and Mercurio,A.M. 1997. Integrin signaling in leukocytes: lessonsfrom the alpha6beta1 integrin. J.Leukoc.Biol. 61:397-407.
72. Hynes,R.O. 1992. Integrins: versatility, modulation, and signaling in cell adhesion. Cell69:11-25.
73. Monboisse,J.C., Garnotel,R., Randoux,A., Dufer,J., and Borel,J.P. 1991. Adhesion ofhuman neutrophils to and activation by type-I collagen involving a beta 2 integrin.J.Leukoc.Biol. 50:373-380.
74. Diamond,M.S. and Springer,T.A. 1993. A subpopulation of Mac-1 (CD11b/CD18)molecules mediates neutrophil adhesion to ICAM-1 and fibrinogen. J.Cell Biol.120:545-556.
75. Wright,S.D., Weitz,J.I., Huang,A.J., Levin,S.M., Silverstein,S.C., and Loike,J.D. 1988.Complement receptor type three (CD11b/CD18) of human polymorphonuclearleukocytes recognizes fibrinogen. Proc.Natl.Acad.Sci.U.S.A. 85:7734-7738.

42
76. Kato,M., Abraham,R.T., Okada,S., and Kita,H. 1998. Ligation of the beta2 integrintriggers activation and degranulation of human eosinophils. Am.J.Respir.Cell Mol.Biol.18:675-86.
77. Newton,R.A., Thiel,M., and Hogg,N. 1997. Signaling mechanisms and the activation ofleukocyte integrins. J.Leukoc.Biol. 61:422-426.
78. Weller,P.F. 1991. The immunobiology of eosinophils. N.Engl.J.Med. 324:1110-8.
79. Weiler,C.R., Kita,H., Hukee,M., and Gleich,G.J. 1996. Eosinophil viability duringimmunoglobulin-induced degranulation. J.Leukoc.Biol. 60:493-501.
80. Sengelov,H., Kjeldsen,L., Diamond,M.S., Springer,T.A., and Borregaard,N. 1993.Subcellular localization and dynamics of Mac-1 (αmβ2) in human neutrophils.J.Clin.Invest. 92:1467-1476.
81. Fukuda,T., Dunnette,S.L., Reed,C.E., Ackerman,S.J., Peters,M.S., and Gleich,G.J. 1985.Increased numbers of hypodense eosinophils in the blood of patients with bronchialasthma. Am.Rev.Respir.Dis. 132:981-985.
82. Frick,W.E., Sedgwick,J.B., and Busse,W.W. 1988. Hypodense eosinophils in allergicrhinitis. J.Allergy Clin.Immunol. 82:119-125.
83. De Simone,C., Donneli,G., Meli,D., Rosati,F., and Sorice,F. 1982. Human eosinophilsand parasitic diseases. II. Characterization of two cell fractions isolated at differentdensities. Clin.Exp.Immunol. 48:249-255.
84. Wardlaw,A. 1995. Eosinophil density: what does it mean? Clin.Exp.Allergy 25:1145-1149.
85. Broide,D.H. 1995. Cytokines and adhesion molecules in asthma. In Curr Allergy ClinImmunol. 19-22.
86. Shaver,J.R., Zangrilli,J.G., Cho,S.K., Cirelli,R.A., Pollice,M., Hastie,A.T., Fish,J.E.,and Peters,S.P. 1997. Kinetics of the development and recovery of the lung from IgE-mediated inflammation: dissociation of pulmonary eosinophilia, lung injury, andeosinophil-active cytokines. Am.J.Respir.Crit Care Med. 155:442-448.
87. Sur,S., Gleich,G.J., Swanson,M.C., Bartemes,K.R., and Broide,D.H. 1995. Eosinophilicinflammation is associated with elevation of interleukin-5 in the airways of patients withspontaneous symptomatic asthma. J.Allergy Clin.Immunol. 96:661-8.
88. Ramshaw,H.S., Woodcock,J.M., Bagley,C.J., Mcclure,B.J., Hercus,T.R., andLopez,A.F. 2001. New approaches in the treatment of asthma. Immuology and CellBiology 79:154-159.
89. Darzynkiewicz,Z., Juan,G., Li,X., Gorczyca,W., Murakami,T., and Traganos,F. 1997.Cytometry in cell necrobiology: analysis of apoptosis and accidental cell death(necrosis). Cytometry 27:1-20.
90. Frigas,E. and Gleich,G.J. 1986. The eosinophil and the pathophysiology of asthma.J.Allergy Clin.Immunol. 77:527-537.

43
91. Catchpoole,D.R. and Stewart,B.W. 1993. Etoposide-induced cytotoxicity in two humanT-cell leukemic lines: delayed loss of membrane permeability rather than DNAfragmentation as an indicator of programmed cell death. Cancer Res. 53:4287-96.
92. Cohen,G.M., Sun,X.M., Snowden,R.T., Dinsdale,D., and Skilleter,D.N. 1992. Keymorphological features of apoptosis may occur in the absence of internucleosomal DNAfragmentation. Biochem.J. 286:331-4.
93. Zamai,L., Falcieri,E., Marhefka,G., and Vitale,M. 1996. Supravital exposure topropidium iodide identifies apoptotic cells in the absence of nucleosomal DNAfragmentation. Cytometry 23:303-11.
94. Takei,H., Araki,A., Watanabe,H., Ichinose,A., and Sendo,F. 1996. Rapid killing ofhuman neutrophils by the potent activator phorbol 12- myristate 13-acetate (PMA)accompanied by changes different from typical apoptosis or necrosis. J.Leukoc.Biol.59:229-40.
95. Hansel,T.T., de Vries,I.J.M., Iff,T., Rihs,S., Wandzilak,M., Betz,S., Blaser,K., andWalker,C. 1991. An improved immunomagnetic procedure for the isolation of highlypurified human blood eosinophils. J.Immunol.Methods. 145:105-110.
96. Håkansson,L., Nielsen,L.S., and Teder,P. 1994. Measurement of neutrophil andeosinophil adhesion to E-selectin, VCAM-1, and ICAM-1 by the use of transfectedfibroblast cell lines. J.Immunol.Methods 176:53-66.
97. Olofsson,T., Olsson,I., Venge,P., and Elgefors,B. 1977. Serum myeloperoxidase andlactoferrin in neutropenia. Scand.J.Haematol. 18:73-80.
98. Peterson,C.G., Enander,I., Nystrand,J., Anderson,A.S., Nilsson,L., and Venge,P. 1991.Radioimmunoassay of human eosinophil cationic protein (ECP) by an improvedmethod. Establishment of normal levels in serum and turnover in vivo. Clin.Exp.Allergy21:561-567.
99. Saito,H., Yamada,K., Breard,J., Yoshie,O., and Mathe,G. 1986. A monoclonal antibodyreactive with human eosinophils. Blood 67:50-55.
100. Venge,P., Bystrom,J., Carlson,M., Hakansson,L., Karawacjzyk,M., Peterson,C.,Seveus,L., and Trulson,A. 1999. Eosinophil cationic protein (ECP): molecular andbiological properties and the use of ECP as a marker of eosinophil activation in disease.Clin.Exp.Allergy 29:1172-86.
101. Winqvist,I., Olofsson,T., and Olsson,I. 1984. Mechanisms for eosinophil degranulation;release of the eosinophil cationic protein. Immunology 51:1-8.
102. Wayner,E.A. 1989. Identification and characterization of the T lymphocyte adhesionreceptor for an alternative cell attachment domain (CS-1) in plasma fibronectin. J.CellBiol. 109:1321-30.
103. Wayner,E.A. 1992. Activation-dependent recognition by hematopoietic cells of theLDV sequence in the V region of fibronectin. J.Cell Biol. 116:489-97.

44
104. Bohnsack,J.F. and Zhou,X.N. 1992. Divalent cation substitution reveals CD18- and verylate antigen- dependent pathways that mediate human neutrophil adherence tofibronectin. J.Immunol. 149:1340-1347.
105. Dobrina,A., Menegazzi,R., Carlos,T.M., Nardon,E., Cramer,R., Zacchi,T., Harlan,J.M.,and Patriarca,P. 1991. Mechanisms of eosinophil adherence to cultured vascularendothelial cells. Eosinophils bind to the cytokine-induced ligand vascular cell adhesionmolecule-1 via the very late activation antigen-4 integrin receptor. J.Clin.Invest. 88:20-6.
106. Sedgwick,K.B., Geiger,K.M., and Busse,W.W. 1990. Superoxide generation byhypodense eosinophils from patients with asthma. Am.Rev.Respir.Dis. 142:120-5.
107. Kita,H., Abu Ghazaleh,R.I., Gleich,G.J., and Abraham,R.T. 1991. Role of pertussistoxin-sensitive G proteins in stimulus- dependent human eosinophil degranulation.J.Immunol. 147:3466-3473.
108. Bates,M.E., Bertics,P.J., Calhoun,W.J., and Busse,W.W. 1993. Increased protein kinaseC activity in low density eosinophils. J.Immunol. 150:4486-93.
109. Dri,P., Cramer,R., Spessotto,P., Romano,M., and Patriarca,P. 1991. Eosinophilactivation on biologic surfaces. Production of O-
2 in response to physiologic solublestimuli is differentially modulated by extracellular matrix components and endothelialcells. J.Immunol. 147:613-620.
110. Nishizuka,Y. 1984. The role of protein kinase C in cell surface signal transduction andtumour promotion. Nature. 308:693-8.
111. Weber,C., Katayama,J., and Springer,T.A. 1996. Differential regulation of beta 1 andbeta 2 integrin avidity by chemoattractants in eosinophils. Proc.Natl.Acad.Sci.U.S.A.93:10939-10944.
112. Lundahl,J., Hallden,G., and Hed,J. 1993. Differences in intracellular pool and receptor-dependent mobilization of the adhesion-promoting glycoprotein Mac-1 betweeneosinophils and neutrophils. J.Leukoc.Biol. 53:336-341.
113. Altieri,D.C. 1991. Occupancy of CD11b/CD18 (Mac-1) divalent ion binding site(s)induces leukocyte adhesion. J.Immunol. 147:1891-1898.
114. Winqvist,I., Olofsson,T., and Persson,E. 1985. Effect of zinc and other cations on therelease of the eosinophil cationic protein. Scand.J.Clin.Lab.Invest. 45:671-7.
115. Grayson,M.H., van der Vieren,M., Sterbinsky,S.A., Gallatin,W.M., Hoffman,P.A.,Staunton,D.E., and Bochner,B.S. 1998. αdβ2 integrin is expressed on humaneosinophils and functions as an alternative ligand for vascular cell adhesion molecule 1(VCAM-1). J.Exp.Med. 188:2187-91.
116. Anwar,A.R., Walsh,G.M., Cromwell,O., Kay,A.B., and Wardlaw,A.J. 1994. Adhesionto fibronectin primes eosinophils via α4β1 (VLA-4). Immunology 82:222-228.

45
117. Diamond,M.S., Garcia Aguilar,J., Bickford,J.K., Corbi,A.L., and Springer,T.A. 1993.The I domain is a major recognition site on the leukocyte integrin Mac-1(CD11b/CD18) for four distinct adhesion ligands. J.Cell Biol. 120:1031-1043.
118. Walsh,G.M., Symon,F.A., Lazarovils,A.L., and Wardlaw,A.J. 1996. Integrin α4β7mediates human eosinophil interaction with MAdCAM-1, VCAM-1 and fibronectin.Immunology 89:112-119.
119. Davis,G.E. 1992. The Mac-1 and p150,95 beta 2 integrins bind denatured proteins tomediate leukocyte cell-substrate adhesion. Exp.Cell Res. 200:242-252.
120. Zuurbier,A.E., Liu,L., Mul,F.P., Verhoeven,A.J., Knol,E.F., and Roos,D. 2001.Neutrophils enhance eosinophil migration across monolayers of lung epithelial cells.Clin.Exp.Allergy 31:444-52.
121. Berton,G., Yan,S.R., Fumagalli,L., and Lowell,C.A. 1996. Neutrophil activation byadhesion: mechanisms and pathophysiological implications. Int.J.Clin.Lab.Res. 26:160-177.
122. Anderson,D.C., Miller,L.J., Schmalstieg,F.C., Rothlein,R., and Springer,T.A. 1986.Contributions of the Mac-1 glycoprotein family to adherence- dependent granulocytefunctions: structure-function assessments employing subunit- specific monoclonalantibodies. J.Immunol. 137:15-27.
123. Mould,A.P., Akiyama,S.K., and Humphries,M.J. 1995. Regulation of integrin alpha 5beta 1-fibronectin interactions by divalent cations. Evidence for distinct classes ofbinding sites for Mn2+, Mg2+, and Ca2+. J.Biol.Chem. 270:26270-26277.
124. Suchard,S.J. and Boxer,L.A. 1994. Exocytosis of a subpopulation of specific granulescoincides with H2O2 production in adherent human neutrophils. J.Immunol. 152:290-300.
125. Schleiffenbaum,B., Moser,R., Patarroyo,M., and Fehr,J. 1989. The cell surfaceglycoprotein Mac-1 (CD11b/CD18) mediates neutrophil adhesion and modulatesdegranulation independently of its quantitative cell surface expression. J.Immunol.142:3537-3545.
126. Bellavite,P., Chirumbolo,S., Mansoldo,C., Gandini,G., and Dri,P. 1992. Simultaneousassay for oxidative metabolism and adhesion of human neutrophils: evidence forcorrelations and dissociations of the two responses. J.Leukoc.Biol. 51:329-35.
127. Evans,D.J., Lindsay,M.A., Webb,B.L., Kankaanranta,H., Giembycz,M.A.,O'Connor,B.J., and Barnes,P.J. 1999. Expression and activation of protein kinase C-zetain eosinophils after allergen challenge. Am.J.Physiol. 277:L233-9.
128. Humbert,M., Corrigan,C.J., Kimmitt,P. 1997. Relationship between IL-4 and IL-5mRNA expression and disease severity in atopic asthma. Am.J.Respir.Crit Care Med.156:704-8.
129. Heath,H., Qin,S., Rao,P., Wu,L., LaRosa,G., Kassam,N.,Ponath,P.D., andMackay,C.R.1997. Chemokine receptor usage by human eosinophils. The importance of

46
CCR3 demonstrated using an antagonistic monoclonal antibody. J.Clin.Invest. 99:178-84.
130. Durham,S.R., Ying,S., Varney,V.A., Jacobson,M.R., Sudderick,R.M., Mackay,I.S.,Kay,A.B., and Hamid,Q.A. 1992. Cytokine messenger RNA expression for IL-3, IL-4,IL-5, and granulocyte/macrophage-colony-stimulating factor in the nasal mucosa afterlocal allergen provocation: relationship to tissue eosinophilia. J.Immunol. 148:2390-4.
131. Ghaffar,O., Laberge,S., Jacobson,M.R., Lowhagen,O., Rak,S., Durham,S.R., andHamid,Q. 1997. IL-13 mRNA and immunoreactivity in allergen-induced rhinitis:comparison with IL-4 expression and modulation by topical glucocorticoid therapy.Am.J.Respir.Cell Mol.Biol. 17:17-24.
132. Sim,T.C., Grant,J.A., Hilsmeier,K.A., Fukuda,Y., and Alam,R. 1994. Proinflammatorycytokines in nasal secretions of allergic subjects after antigen challenge.Am.J.Respir.Crit.Care Med. 149:339-44.
133. Coeffier,E., Joseph,D., and Vargaftig,B.B. 1991. Activation of guinea pig eosinophilsby human recombinant IL-5. Selective priming to platelet-activating factor-acether andinterference of its antagonists. J.Immunol. 147:2595-602.
134. Simon,H.U., Weber,M., Becker,E., Zilberman,Y., Blaser,K., and Levi-Schaffer,F. 2001.Eosinophils maintain their capacity to signal and release eosinophil cationic proteinupon repetitive stimulation with the same agonist. J.Immunol. 165:4069-75.