shellfish aquaculture and the environment (shumway/shellfish aquaculture and the environment) || the...
TRANSCRIPT

Chapter 17
The i mplications of g lobal c limate c hange for m olluscan a quaculture Edward H. Allison , Marie - Caroline Badjeck , and Kathrin Meinhold
Introduction
The overwhelming majority of the world ’ s climate scientists agree that human activity is changing the global climate (Anderegg et al. 2010 ). Rising CO 2 and other greenhouse gas emissions from industry, domestic use and agriculture, and the degradation of carbon sinks through deforestation and other land use and coastal ecosystem conversions are together leading to rapid warming of the earth ’ s atmo-sphere (IPCC 2007a ). Public awareness of the changing climate is increasing, although people in industrialized countries remain divided on its cause and distracted by more immediate concerns (Lorenzoni and Pidgeon 2006 ; Dunlap and McCright 2008 ). Commitment by
individuals, civil society, businesses, and states to reduce emissions and invest in adapting to change is diffuse or limited, hence the continu-ing diffi culties in reaching a global accord on emissions targets (Lazarus 2009 ).
While humanity debates the issues and how to deal with them, global warming is leading to changes in the basic biophysical processes that determine the structure and function of the oceans (Harley et al. 2006 ). This, in turn, has impacts on the coastal and marine ecosys-tems within which molluscan shellfi sh are farmed. Climate change should therefore be an issue of concern to shellfi sh farmers, proces-sors, and traders, as well as to researchers and regulators and to all of us who enjoy eating shellfi sh. All these activities will be affected by
Shellfi sh Aquaculture and the Environment, First Edition. Edited by Sandra E. Shumway.© 2011 John Wiley & Sons, Inc. Published 2011 by John Wiley & Sons, Inc.
461

462 Shellfi sh Aquaculture and the Environment
the changing climate, and some are already being affected. In this chapter, we review the relevant elements of climate change and their likely impact on shellfi sh farming operations and value chains, and we propose measures to respond to them, so that we may retain pro-ductive industries and access to a highly valued source of food.
Molluscs are cultured in both temperate and tropical environments, in marine and brackish water, in a variety of farming systems, ranging from extensive seabed cultivation of oysters and clams, through semi - intensive line and raft cultivation of mussels and scallops, through to intensive recirculation systems for abalone (Chapter 1 in this book). They also contribute to economy and society in various ways — from providing low - input food pro-duction systems for small - scale farmers and fi sherfolk, by being part of a diversifi ed coastal economy sustaining small communities, to being major commercial concerns (Chapter 2 in this book). As we will show, even the effects of a single climate change - related variable — ocean acidifi cation due to increasing concen-tration of CO 2 in the atmosphere — cannot be accurately predicted. When we consider the multiple pathways through which climate change (and societal responses to it) will take place, prediction becomes diffi cult on the basis of current knowledge. In this chapter, rather than attempting to make ill - founded model predictions of future harvests, and their distri-bution and value, we limit ourselves to identi-fying the pathways of potential impact of climate change on molluscan shellfi sh aquacul-ture, reviewing the evidence for impact so far, and giving some examples of the nature and scale of predicted impacts from recent case studies. We also review possible adaptive responses, mindful that investment in such adaptation will have to contend with continu-ing uncertainty about the nature and extent of impacts. Finally, we consider how shellfi sh farms can become part of the solution to a
lower carbon future for our societies, by exploring their role in producing food that has a small “ carbon footprint ” (Bunting and Pretty 2007 ; Hickey 2008 ), and by evaluating the proposition that shellfi sh aquaculture, through the production of calcium carbonate shells, can act as a “ carbon sink. ”
Climate c hange in the o ceans and c oastal z ones
As recently reviewed by the Intergovernmental Panel on Climate Change (IPCC) (IPCC 2007a ), global warming leads to changes in physical processes in aquatic systems, such as alternations in the major ocean currents and in the local circulation patterns in coastal systems; changes in the frequency and severity of the El Ni ñ o Southern Oscillation ( ENSO ; driven by the Humboldt Current in the Pacifi c); and thermal expansion and addition of ice melt to the oceans, leading to sea level rise (Fig. 17.1 ). In freshwater, rainfall and river-fl ow patterns and rates of evapotranspiration are changing. Storms, fl oods, and droughts are changing in frequency and intensity. Chemical changes brought about by rising levels of atmospheric CO 2 are also leading to an acidi-fi cation of the oceans, with consequences for ocean food chains and for coral reefs.
Drawing on recent reviews by the IPCC (2007a) and by Barange and Perry (2009) , supplemented by reference to other specifi c studies, the main chemical and physical ele-ments of global change affecting the oceans can be summarized as follows.
Heat c ontent and t emperature
The oceans are warming, but there are geo-graphical differences in the extent of that warming, and there continue to be decadal

Implications of global climate change 463
Ocean s alinity, d ensity, and s tratifi cation
Overall indications are that, globally, oceans are becoming less saline due to greater precipi-tation, higher runoff, ice melting, and advec-tion, but that there are large regional differences. Salinity is increasing in the surface of the subtropical North Atlantic Ocean (15 – 42 ° N), while further north and in the Southern Ocean waters are becoming less saline. Lower salinity is also reported in the Pacifi c, except in the upper 300 m and in the subtropical gyre, where salinity is increasing, and the Indian Ocean is generally showing increases in salin-ity in the upper layers. Salinity affects species distribution, and the growth and reproductive success of individual shellfi sh species, and will therefore affect the composition and produc-tivity of both natural and farmed shellfi sh populations.
cycles which can mean short - term cooling in some areas, despite the overall warming trend. Warming is not exclusive to surface waters, with deep warming being seen particularly clearly in the Atlantic Ocean. While coastal large marine ecosystems (LMEs) on the West Coast of the Americas have shown slight cooling, most other areas have warmed, with enclosed shelf seas (e.g., Mediterranean, Black Sea) showing particularly large increases.
Increases in temperature (and salinity; see below) from changes in precipitation, evapora-tion, river runoff, and ice melt are likely to lead to increased vertical stratifi cation and water column stability in oceans and lakes, reducing nutrient availability to the euphotic zone and thus primary and secondary produc-tion. Ecosystem productivity is likely to be reduced in most tropical and subtropical oceans, seas, and lakes, and increased in high latitudes.
Figure 17.1 The main physical changes in the oceans attributable to anthropogenic global warming. (Reproduced from Harley et al. 2006 .)
Human activities
Increased airtemperature
Increased stormfrequency
IncreasedCO2
IncreasedUV
Increased watertemperature
Sea levelrise
Decreased pH
Increased greenhousegas concentrations
Intensified atmosphericpressure gradients
Intensifiedupwelling (?)

464 Shellfi sh Aquaculture and the Environment
pended sediment loads. In contrast, damming and channelization have greatly reduced the supply of sediments to the coast from other rivers through retention of sediment by dams (many of which are themselves now being built as adaptations to climate change — e.g., to increase water storage, regulate fl oods, or reduce emissions from fossil fuel - based power generation). Levels of sedimentation have the potential to have signifi cant impacts on shell-fi sh farming through changes in turbidity, salinity, stratifi cation, and nutrient availability, all of which affect estuarine and coastal eco-systems and the productivity of the phyto-plankton community.
Changes in l ow - f requency c limate v ariability p atterns
Natural climate variability takes place through intermittent 1 – 2 - year duration events (e.g., ENSO) and intrinsic variability operating at decadal and longer timescales. ENSO events are associated with many atmospheric and oceanic patterns, including abnormal patterns of rainfall over the tropics, Australia, southern Africa and India, and parts of the Americas, easterly winds across the entire tropical Pacifi c, air pressure patterns throughout the tropics, and sea surface temperatures. Coincident eco-logical changes are both vast and global through their impacts in coastal upwelling systems: They increase coastal temperatures, reduce plankton production by lowering the thermo-cline (which inhibits upwelling of nutrients), and change trophodynamic relationships; they also have knock - on effects on the hydrological cycle and therefore on agriculture and even on forest fi re severity, as in Indonesia following the 1997 ENSO event. Examples of aquaculture impacts of El Ni ñ o are given in the next section.
Some studies expect stronger and more fre-quent ENSO events as a result of global warming, while others suggest that the evi-dence is still lacking because ENSO is not well -
Ocean c irculation and c oastal u pwelling
Observed and predicted changes in the ocean ’ s heat content and salinity are and will continue to affect circulation patterns. These are expected to decline, but at present there is confl icting and unclear evidence as to whether climate change will increase or decrease coastal upwelling. This is important in terms of its implications for biological production in the major fi sheries and shellfi sh farms of upwelling areas.
Sea l evel r ise
Global average sea level has been rising at an average rate of 1.8 mm per year since 1961. The rate has accelerated since 1993 to about 3.1 mm per year. Higher rates in coming decades are likely, and recent research (e.g., Rahmstorf et al. 2007 ) indicates that they may be higher than even the most pessimistic (IPCC 2007a ) scenario.
Sea level change is not geographically uniform, however, because it is controlled by regional ocean circulation processes. The largest losses of land and impact on popula-tions and economies are likely to be in East and Southeast Asia, although proportional impacts may be even higher on low - lying small - island developing states in the Pacifi c and Indian Oceans in particular. Rises in sea levels may affect coastal bays, lagoons, and wetlands, which provide sites for shellfi sh cul-tivation, and coastal onshore infrastructure used by fi sherfolk (e.g., housing, harbors).
Land – o cean e xchanges
Land use change contributing to and resulting from climate change, particularly deforesta-tion and hydrological modifi cations, has downstream impacts, especially in terms of erosion in catchment areas and increased sus-

Implications of global climate change 465
expected that pH reduction will change the depth below which calcium carbonate dis-solves, increasing the volume of ocean that is undersaturated with respect to aragonite and calcite, which are used by marine organisms to build their shells and by corals to build their skeletons. The impacts of these changes will be greater for some regions and ecosystems and may be most severe for shell - bearing organ-isms, tropical coral reefs, and cold - water corals in the Southern Ocean. Ocean acidifi cation has been portrayed in apocalyptic terms. The con-clusions of two recent reviews (Doney et al. 2009 ; Hendriks et al. 2010 ) are more mea-sured but still point to the need for urgent action on curbing emissions to prevent rapid change. The direct implications for shellfi sh have galvanized much recent research, which is reviewed in the next section.
Impacts of the above physical and chemical changes in coastal and oceanic waters have potentially profound ecological impacts. Drawing on Barange and Perry (2009) and Brierley and Kingsford (2009) , these impacts include as follows.
Timing and s uccess of p hysiological, s pawning, and r ecruitment p rocesses
All living organisms have specifi c ranges of environmental conditions to which they are adapted and within which they perform opti-mally. Physiological performance may degrade and cause stress at temperatures outside the normal limits. Likely impacts of climate change include changes in timing and success of fertil-ization, survival and growth of shellfi sh, timing and extent of migrations, and structure of food webs.
Primary p roduction
Satellite observations suggest a 6% reduction in global oceanic primary production between
enough simulated in climate models to have full confi dence in these projected changes (Cane 2005 ).
Other naturally occurring climatic variabil-ity being impacted by climate change relates to atmospheric associations or teleconnections; changes in the position and intensity of atmo-spheric convection in one area result in adjust-ments in pressure cells in adjacent areas and can lead to altered wind and ocean current patterns on a global scale. Understanding these connec-tions and how climate change will affect them is a research frontier for climate scientists.
Increased f requency and s everity of e xtreme c limate e vents
Most major tropical storms are generated around latitudes 8 ° and 35 ° north and south of the equator. There are ongoing scientifi c controversies around the oft - stated perception that storm frequency and severity are increas-ing due to climate change, and that there may be changes in seasonality and typical track of storm events. An emerging consensus around recent trends and predictions for hurricane energy (a measure of storm severity) indicate no evidence for increased frequency of storms, but strong evidence for a trend for increasing sever-ity and therefore destructiveness of storms over the last 20 – 40 years (i.e., exceeding the period-icity of decadal cycles in the ocean climate system and attributable to anthropogenic warming); this trend is particularly evident in the western Atlantic (Emanuel et al. 2008 ).
Ocean a cidifi cation and c hanges in c hemical p roperties
Continued uptake of atmospheric CO 2 by the oceans has decreased the pH of surface seawa-ter by 0.1 units in the last 200 years. Model estimates predict further reduction of 0.3 to 0.5 pH units over the next 100 years. It is

466 Shellfi sh Aquaculture and the Environment
larval survival rates, disease immunity, etc.) are not negatively affected by climate change. Thus, it cannot be expected that warming will bring more and faster - growing shellfi sh.
Phenomenological c hanges
More than half of all terrestrial, freshwater, or marine species studies have exhibited measur-able changes in their phonologies (timing of life cycle events) over the past 20 – 140 years, in line with climate changes. In the oceans, this includes changes in the timing and extent of seasonal phytoplankton blooms which have not been tracked by zooplankton, suggesting an emerging mismatch between food supply and predator life cycles.
Species i nvasions and d iseases
Pathogens are spreading to higher latitudes and harmful algal blooms (HABs) are becom-ing more common (although these may also be due to localized nutrient enrichment). Ecosystems may become more liable to inva-sive species as they change conditions from those that may have prevented warmer water species from breeding, for example. Thermally stressed shellfi sh in aquaculture systems may become more liable to viral, fungal, and bacte-rial infections.
Regime s hifts and e xtreme e vents
Gradual and variable climate changes can provoke sudden and perhaps irreversible bio-logical responses as ecosystems shift from one state to another (de Young et al. 2008 ). The altered state may provide an alternative fi shery, or it may simply be less productive. Examples of regime shifts are the alternations between sardine and anchovy - dominated pelagic fi sher-ies in some upwelling areas, and arguably, the
the early 1980s and late 1990s, but with sub-stantial regional differences. The climate – plankton link in the ocean is found most strongly in the tropics and mid latitudes, where the typically low levels of surface nutrients limit phytoplankton growth. Climate warming further inhibits mixing, reducing the upward nutrient supply and lowering productivity (Doney 2006 ). Reduced primary production is likely to mean reduced fi sh and shellfi sh pro-duction in many aquatic ecosystems, although the relationship may not be straightforward as much productivity is derived from the “ micro-bial loop ” and dependent on infl ux and cycling of organic material from terrestrial areas for continental shelf seas.
Changes in d istributions
Climate change is expected to drive most ter-restrial and marine species ranges toward the poles, expanding the range of warmer - water species and contracting that of colder - water species. Such changes have already been docu-mented in the North Sea and North Atlantic for fi nfi sh. A recent study modeling “ climate envelopes ” based on existing species distribu-tions of both fi sh and exploited invertebrates suggests that there will be substantive changes in distribution of fi sh and invertebrate com-munities, and large numbers of extinctions of species with restricted habitat ranges (Cheung et al. 2009 ).
Abundance c hanges
At the simplest level, biological production processes occur faster at higher temperatures. However, increased growth rates of shellfi sh (and hence increased biomass and production) will only occur in response to higher tempera-tures when food supply is adequate to these increased demands, and when other life cycle processes (spawning migrations, fertilization,

Implications of global climate change 467
development, and protected areas; and changes in coastal land use and associated marine habi-tats which affect key life cycle stages of shell-fi sh populations or water quality. Markets change, too: People no longer eat the quantity of oysters that New Yorkers did in the eigh-teenth and nineteenth centuries (Kurlansky 2006 ) but expectations on quality and variety are likely to increase. Although farming tech-niques for many shellfi sh remain simple, tech-nology also evolves, notably in the development of hatcheries, in the selective breeding that this makes possible, and in the development of closed or recirculation systems. The legislative and policy context for shellfi sh farming, pro-cessing, and trade also adapts and responds to new threats and opportunities. Climate change is thus an additional driver of change in a sector in which environmental management and food safety already have a strong infl u-ence. Both of these drivers will themselves be affected by climate change, as we will discuss in the next section.
Human activities are generating increased emissions of greenhouse gasses — principally methane and carbon dioxide. The changing chemical composition of the earth ’ s atmo-sphere is leading to a global change in the climate. There is an overall warming trend, accompanied by changing weather patterns (which may include localized or seasonal cooling). Increased CO 2 emissions are also responsible for the “ other CO 2 problem ” — ocean acidifi cation, which has far - reaching implications for all calcifying marine organ-isms, of obvious concern to shellfi sh farming (hence the large number of experimental studies, reviewed below) and with particularly alarming prognoses for the future of coral reefs (Doney et al. 2009 ). Warming and chang-ing weather are leading to rising sea surface temperatures and changes in the fl ow and variability of ocean currents (the two are linked). These changes lead to linked sets of physical, chemical, and biological effects, which include reduced oxygenation in waters
collapse of the Newfoundland cod fi shery signaled a regime shift driven by the com-bined stressors of overfi shing and climate variability.
The links between global warming, these physical and chemical changes in hydrology and oceanography, and the ecological changes in aquatic food webs are thus complex, and when we then add the operations and econom-ics of the shellfi sh farming sector, the complex-ity increases further. In order to identify issues that can be evaluated and acted upon in the context of mollusc culture, the next section is structured around a simplifi ed schema that draws on the mass of observation and predic-tion that underlies the above brief review, and illustrates some the main potential pathways of impact from climate change to shellfi sh specifi cally.
The e ffects of c limate c hange on s hellfi sh a quaculture s ystems
Drawing on a review of the extensive literature on existing and potential climate change impacts on the oceans, fi sheries, and on aqua-culture systems (e.g., Handisyde et al. 2006 ; Brander 2007 ; Brierley and Kingsford 2009 ; De Silva and Soto 2009 ; Badjeck et al. 2010 ), the main pathways through which climate variability and change are hypothesized to impact shellfi sh are illustrated in Figure 17.2 . After a brief explanation of the diagram, we review the available evidence for each of the illustrated pathways.
First, it is important to remind ourselves that climate change is not the only “ driver ” affecting the operation of shellfi sh farms and may not be the most infl uential one. Nonclimate drivers of change could include pollution that affects the location and operating costs of shellfi sh farms; competing uses for the fore-shore and coastal waters that constrain the expansion of the sector — for example, indus-try and urbanization, tourism and residential

468 Shellfi sh Aquaculture and the Environment
The increasing heat energy in the oceans and atmosphere and change in currents also affect the formation of storm events. The warming sea and melting polar ice caps are leading to a rise in sea level, which, when taken together with potentially increased storm intensity, are likely to increase the sever-ity of coastal fl ooding, extreme rainfall events, and stormy weather. This combination of changes threatens to bring increased losses of stock and direct damage to aquaculture instal-lations and service facilities, to coastal infra-structure, and indeed to people ’ s homes. In countries without effective storm early warning and evacuation systems, and with large, vul-nerable coastal populations, hundreds or even thousands of human lives are lost with every major storm, as was the case with the major
where circulation is limited, changes in primary production and food web structure and func-tion, and in life history processes such as spat-fall of shellfi sh. Warming and changes in currents are also associated with increased fre-quency and poleward range expansion of HABs, as well as facilitating the spread of pests and alien species. These are issues that already plague the shellfi sh farming sector.
In combination, the changes outlined above are likely to lead to l physiological stresses that decrease growth and production, although warming could improve growth rates by speed-ing up metabolism, if primary production is not limiting. There will certainly be changes in where particular species can be successfully farmed, or in which species are farmed in areas traditionally used for shellfi sh cultivation.
Figure 17.2 Multiple pathways by which climate change potentially impacts molluscan shellfi sh aquaculture. GHGs, greenhouse gasses.
Humanactivity
Nonclimate drivers of change: e.g., pollution, competing uses for
coastal waters, fishing pressure, land use and habitat change, changing
Increased CO2
Increased Increased CO2 concentration in
Ocean acidification
patterns of demand, technology development, legislation and policy
emissionsemissions:other GHGS oceans
(reduced pH)
Reduced rates of
calcificationin shellfish Atmospheric warming
and changing weather patterns
Reduced oxygenation of
heated, enclosed watersDecreased growth and productionIncreased mortality,
Increased costs of
production
Changes in
Changes in primary
production and food web
Rising sea surface
temperature
Increased frequency of
Changes in natural spatfall
altered recruitment patterns
production volume
and value
Needs and Shifting sites of
structure and function
Changingocean currents
pathogenic infections
Changes in distribution of
pests and alien species
opportunities for
adaptation in
culture locations,
species grown,technology,
production
Changes in species
cultured
Sea level rise
Increased frequency
Increased losses and direct damage to aquaculture installations
system
management,
transportation and
marketingCoastal flooding
Extreme rainfall events
and severity of
extreme weather
events
and coastal
infrastructure
High winds and wave energy

Implications of global climate change 469
tions in Chesapeake Bay in a warm future (Paolisso et al. 2010 ) might look like present - day conditions in the Gulf of Mexico, so the shellfi sh culture conditions on the Gulf coast may be a spatial analogue for those that can be expected in Chesapeake Bay at the end of the twenty - fi rst century.
Literature that makes reference to the potential impacts of climate change on shell-fi sh aquaculture is concentrated on looking at two of the pathways illustrated in Figure 17.2 : ocean acidifi cation and its potential impacts on shell formation and growth rate; and the spread and impact of HABs and its effects (Tables 17.1 and 17.2 ). Studies of the direct and indirect effects of warming, and of mul-tiple, interacting pathways are rarer (Table 17.3 ). While there are many reports of storm and fl ood damage to shellfi sh farming opera-tions, there are none in the peer - reviewed lit-erature which demonstrate or claim increased frequency or severity of such events, linked with global warming.
Studies of the impacts of ocean acidifi cation of shellfi sh have burgeoned in the last 5 years, since ocean acidifi cation became a global concern. While some early studies used a range of pH outside values even the gloomiest prog-nosis for the future, the studies reviewed in Table 17.1 have confi ned experimental manip-ulation of pH to levels corresponding to values predicted by 2050 or 2100 under IPCC emis-sions scenarios. In general, the 0.5 pH unit decrease in seawater forecast by 2100 is asso-ciated with decreased growth and calcifi cation rates in juvenile and adult oysters, mussels, and scallops, although the results for mussels are ambiguous. Waldbusser et al. (2011) point to the complexity of the relationship, which is mediated by salinity and temperature changes that will occur concurrently with acidifi cation. Talmage and Gobler (2010) include preindus-trial CO 2 levels and associated seawater pH in their experiments, which demonstrate that bivalve development may already have been impacted by the changes that have taken place
cyclones in Bangladesh and Myanmar in the last 3 years (ESCAP and UNISDR 2010 ).
Through these pathways of impact, shellfi sh farming systems and the value chains associ-ated with them are likely to face increased costs of production and changes in the volume, species composition and value of their produc-tion. These costs will be passed on to consum-ers, affecting demand. There will be a need — or, more optimistically, an opportunity — to adapt. Adaptation options are discussed in Section 4.
What, then, do we know of the likelihood and potential magnitude of the impacts described above? In Tables 17.1 – 17.3 , we summarize research on the relationship between climate variables and impacts on farmed shellfi sh.
First, a caveat: Our review is not compre-hensive as it is confi ned to studies in which the authors explicitly make reference to climate change. For this reason, it is largely recent. There is of course an extensive literature, dating back more than a century, linking shell-fi sh population dynamics to climate variability but as it predates concern for anthropogenic warming it makes no reference to climate change and no prediction of likely impacts of such change. A more comprehensive review — beyond the scope of this one — is required to make use of this data set to search for relation-ships that can help predict future change. The use of “ climate analogues ” is standard in climate change research applied to other agri-cultural systems. Using temporal analogues involves investigating the effects of past cli-matic events, including climate variability, to predict future changes and inform adaptation (Glantz 1988 ; Easterling et al. 1992 ). Spatial analogues refer to conducting research in one region and identifying parallels to how another might be affected (Ford et al. 2010 ). In agri-cultural research, this involves looking at what species and farming systems will be suitable for the anticipated climate, based on studying areas of the world where such conditions are the current norm. To take an example, condi-

Table 17.1 Evidence for impact of increased CO 2 concentration and ocean acidifi cation on shellfi sh species and culture operations.
Species and location Impact Nature of evidence Reference
Oyster ( Crassostrea virginica ); Chesapeake Bay, USA, laboratory studies
Estuarine waters are more susceptible to acidifi cation because they are subject to multiple acid sources and are less buffered than marine waters. Biocalcifi cation declined signifi cantly with a reduction of 0.5 pH units and higher temperature, but salinity mitigated the decrease in biocalcifi cation. Complex relationships between water chemistry, eutrophication, pH, and biocalcifi ation do not allow simple prediction of reduced biocalcifi cation with increasing CO 2 concentration in the atmosphere.
Field water quality monitoring in Chesapeake Bay, and experimental studies on oyster larvae
Waldbusser et al. (2011)
Hard - shell clams and scallops: Mercenaria mercenaria and Argopecten irradians ; laboratory studies, USA
Compared with present CO 2 conditions (390 ppm), larvae grown under near preindustrial CO 2 concentrations (250 ppm) had signifi cantly faster growth and metamorphosis, higher survival and lipid accumulation rates, as well as having thicker, more robust shells. Bivalves exposed to CO 2 levels expected later this century (750 ppm) had shells that were malformed and eroded.
Experimental studies on larvae, with link to past, current, and projected oceanic CO 2 concentrations
Talmage and Gobler (2010)
Eastern oyster ( Crassostrea virginica ) and the Suminoe oyster ( Crassostrea ariakensis ); laboratory studies, USA
Oyster larvae grown in estuarine water under four pCO 2 regimes, representing preindustrial, present, and projected concentrations. Crassostrea virginica experienced a 16% decrease in shell area and a 42% reduction in calcium content when preindustrial and end of twenty - fi rst - century pCO 2 treatments were compared. Crassostrea ariakensis showed no change to either growth or calcifi cation. Both species demonstrated net calcifi cation and growth, even when aragonite was undersaturated.
Experimental studies on larvae, with link to past, current, and projected oceanic CO 2 concentrations
Miller et al. (2009)
Mussels ( Mytilus edulis ); laboratory studies, Norway
Virtually no growth at pH 6.7 and reduced growth at pH 7.1, but no signifi cant difference between growth at pH 8.1 (current) and pH 7.4 and 7.6
Short - term experimental studies on juveniles and adults
Berge et al. (2006)
Mussels ( Mytilus edulis ) and oysters ( Crassostrea gigas ); laboratory studies, the Netherlands
The calcifi cation rates of the edible mussel and Pacifi c oyster decline linearly with increasing pCO 2 . Mussel and oyster calcifi cation may decrease by 25% and 10%, respectively, by the end of the century, following one of the IPCC emissions scenarios (740 ppm in 2100).
Experimental studies with juvenile and adult organisms
Gazeau et al. (2007)
470
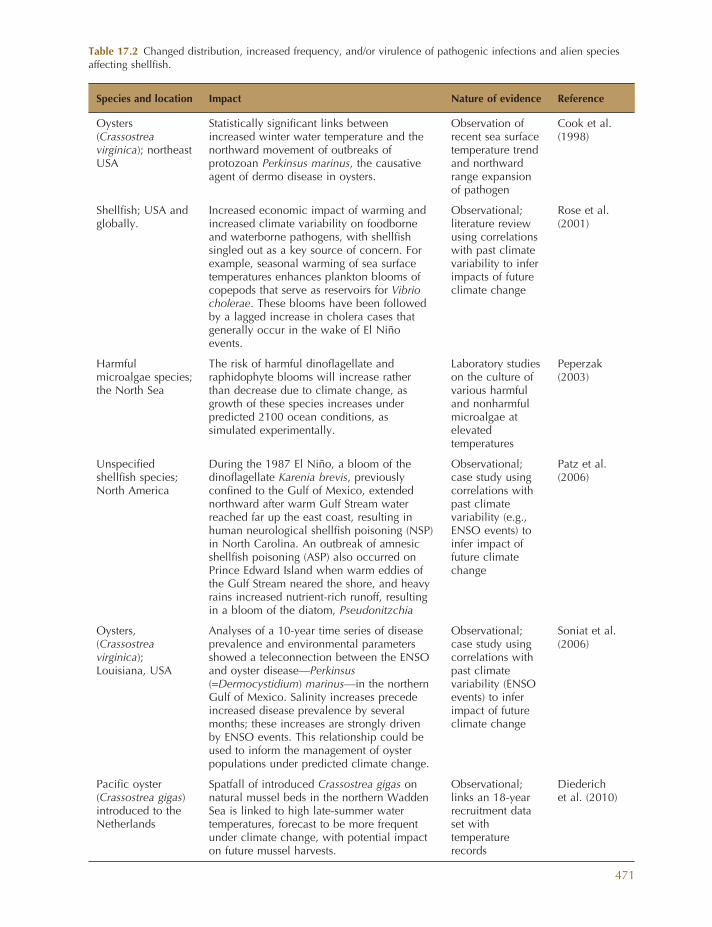
Table 17.2 Changed distribution, increased frequency, and/or virulence of pathogenic infections and alien species affecting shellfi sh.
Species and location Impact Nature of evidence Reference
Oysters ( Crassostrea virginica ); northeast USA
Statistically signifi cant links between increased winter water temperature and the northward movement of outbreaks of protozoan Perkinsus marinus , the causative agent of dermo disease in oysters.
Observation of recent sea surface temperature trend and northward range expansion of pathogen
Cook et al. (1998)
Shellfi sh; USA and globally.
Increased economic impact of warming and increased climate variability on foodborne and waterborne pathogens, with shellfi sh singled out as a key source of concern. For example, seasonal warming of sea surface temperatures enhances plankton blooms of copepods that serve as reservoirs for Vibrio cholerae . These blooms have been followed by a lagged increase in cholera cases that generally occur in the wake of El Ni ñ o events.
Observational; literature review using correlations with past climate variability to infer impacts of future climate change
Rose et al. (2001)
Harmful microalgae species; the North Sea
The risk of harmful dinofl agellate and raphidophyte blooms will increase rather than decrease due to climate change, as growth of these species increases under predicted 2100 ocean conditions, as simulated experimentally.
Laboratory studies on the culture of various harmful and nonharmful microalgae at elevated temperatures
Peperzak (2003)
Unspecifi ed shellfi sh species; North America
During the 1987 El Ni ñ o, a bloom of the dinofl agellate Karenia brevis , previously confi ned to the Gulf of Mexico, extended northward after warm Gulf Stream water reached far up the east coast, resulting in human neurological shellfi sh poisoning ( NSP ) in North Carolina. An outbreak of amnesic shellfi sh poisoning ( ASP ) also occurred on Prince Edward Island when warm eddies of the Gulf Stream neared the shore, and heavy rains increased nutrient - rich runoff, resulting in a bloom of the diatom, Pseudonitzchia
Observational; case study using correlations with past climate variability (e.g., ENSO events) to infer impact of future climate change
Patz et al. (2006)
Oysters, ( Crassostrea virginica ); Louisiana, USA
Analyses of a 10 - year time series of disease prevalence and environmental parameters showed a teleconnection between the ENSO and oyster disease — Perkinsus ( = Dermocystidium ) marinus — in the northern Gulf of Mexico. Salinity increases precede increased disease prevalence by several months; these increases are strongly driven by ENSO events. This relationship could be used to inform the management of oyster populations under predicted climate change.
Observational; case study using correlations with past climate variability (ENSO events) to infer impact of future climate change
Soniat et al. (2006)
Pacifi c oyster ( Crassostrea gigas ) introduced to the Netherlands
Spatfall of introduced Crassostrea gigas on natural mussel beds in the northern Wadden Sea is linked to high late - summer water temperatures, forecast to be more frequent under climate change, with potential impact on future mussel harvests.
Observational; links an 18 - year recruitment data set with temperature records
Diederich et al. (2010)
471

Table 17.3 Evidence for other direct and indirect impacts of climate change on coastal waters that affect shellfi sh species and culture operations.
Climate variable Species and location Impact Nature of evidence Reference
ENSO cycles Oysters ( Crassostrea virginica ); Gulf of Mexico, USA
In a National Oceanic and Atmospheric Administration study of chemical contaminant burdens in oysters (trace metal, polynuclear aromatic hydrocarbon , and pesticides), ENSO cycles were hypothesized to be important in establishing the interannual variability in contaminant body burden. The implications are that climate variability and change may affect levels of contaminants in shellfi sh.
Observational; correlations between interannual patterns of contaminant loads and large - scale climate cycles, 1986 – 1992
Kim et al. (1999)
Spatfall and/or recruitment to natural populations
Macoma baltica ; the Wadden Sea, the Netherlands
Rising seawater temperatures affect recruitment by a decrease in reproductive output and by spring advancement of bivalve spawning. Apparently, global warming upsets the evolved reproductive strategy of this marine bivalve to tune its reproduction to the most optimal environmental conditions for the fi rst vulnerable life stages, most importantly the match/mismatch of time of spawning with that of the phytoplankton bloom and the settlement of juvenile shrimps on the tidal fl ats
Field observation, 1973 – 2001
Philippart et al. (2003)
Ocean warming
Red abalone, Haliotis rufescens , and green abalone, Haliotis fulgens ; California
For red abalone, warm temperatures increased the onset of withering syndrome, a fatal abalone disease, and halted growth and reproduction. In contrast, green abalone survivorship, growth, and reproduction were relatively robust irrespective of temperature, while their growth and reproduction were most strongly infl uenced by food quantity. Cool - water red abalone suffer stronger consequences in warm water than do green abalone.
Experimental study of synergistic effects of temperature and food quantity and quality on survivorship, growth, and reproduction
Vilchis et al. (2005)
472

Climate variable Species and location Impact Nature of evidence Reference
Changes in primary production
Phytoplankton; global
Reduced primary production in the post - 1999 warming period (likely to have impact on shellfi sh production).
Emergent relationships between observed warming and productivity, and future predicted warming
Behrenfeld et al. (2006)
Ocean warming
Bivalve species; global
Greater tolerance for temperature variability in temperate - zone bivalves means that warming will have proportionately greater impact on survival, range, and productivity of tropical species than temperate ones.
Literature review of upper thermal tolerance limits
Compton et al. (2007)
Increased frequency of extreme rainfall events and river fl ows
Mussels ( Mytilus edulis ); Conwy Estuary, Wales, UK
Climate change will bring more frequent extreme rainfall and high fl ow events in rivers, which can impact shellfi sh populations and farms in estuaries through decreased salinity and/or increased transport of sediments, organic matter, and nutrients. One extreme event impacted condition index, while the other reduced total hemocyte count, suggesting impact on health and resilience of estuarine mussel populations.
Evidence from sampling over 18 months at control and impacted sites, covering two extreme fl ow events, with a hypothesized link to climate change
Oliver et al. (2008)
Coastal hypoxia, driven by eutrophication and sea temperature rise
Review of 206 species of benthic metazoans; globally
Molluscs are the benthic organisms most tolerant to hypoxia, crustaceans the least. This is therefore not likely to be an important way in which global warming affects molluscan aquaculture.
Review of 872 experimental studies of hypoxia tolerance
Vaquer - Sunyer and Duarte (2008)
Table 17.3 (Continued)
473

474 Shellfi sh Aquaculture and the Environment
Climate variable Species and location Impact Nature of evidence Reference
Reduced upwelling in coastal waters and increased water retention times in bays
Mussels ( Mytilus edulis ); Galicia, northwest Spain
The extent and intensity of the upwelling season has decreased by 30% and 45% over the last 40 years, respectively. This has led to a 240% increase in water retention time in the four large coastal inlets where 15% of the world ’ s harvest of blue mussels come from, which is causing increasing occurrence of harmful microalgae, with dramatic negative effects on raft cultivation of mussels, confi ning harvests to progressively fewer number of days per year.
Hydrographic, biological oceanography and aquaculture production studies over a 40 - year period
Á lvarez - Salgado et al. (2008)
Ocean warming and acidifi cation (synergistic effects)
Sydney rock oyster, Saccostrea glomerata ; southeast Australia
Fertilization rate, embryonic development rate, and frequency of abnormal veligers increased with temperature and CO 2 increase. Predicted changes in ocean acidifi cation and temperature over the next century may have severe implications for the distribution and abundance of this commercially - important species.
Laboratory studies, both short term and prolonged
Parker et al. (2009)
Table 17.3 (Continued)
to date. The species and life stage specifi city of acidifi cation impacts makes easy generaliza-tions impossible. Recent meta - analysis of the experimental studies of potential impacts of acidifi cation on marine organisms remains dis-puted on whether the oceans face a crisis or not (Dupont et al. 2010 ; Hendriks and Duarte 2010 ; Hendriks et al. 2010 ).
The predicted impacts of warming on pathogens and parasites of shellfi sh (Chapter 13 in this book) and of invasive species (Chapter 14 in this book) are apparently more clear - cut than those of acidifi cation (Table 17.2 ). Correlating past variability and warming
with the spread of diseases and alien species leads to a prediction that such problems will be more frequent in the future. This claim is strengthened by being rooted in observation on change that has already happened, particu-larly concerning HABs.
The most important harmful algae and their poisoning syndromes (in brackets) relevant to shellfi sh are diatoms from the genus Pseudo - nitzschia (amnesic shellfi sh poisoning), and species of dinofl agellates from the genera Alexandrium , Pyrodinium , and Gymnodinium (paralytic shellfi sh poisoning), Karenia (neuro-toxic shellfi sh poisoning and aerosolized

Implications of global climate change 475
adapt genetically and phenotypically to the pace of current climate change. This makes prediction of the future distribution “ fraught with diffi culties ” (Hallegraeff 2010 ). Of course, this should not stop anyone from trying ,and the presentation of future scenar-ios, even if of unknown probability, can be useful to galvanize responsive action. For example, Moore et al. (2008) predict that future warming will lengthen the season in which oysters from Puget Sound will be dan-gerous to consume due to possible phytotoxin contamination (Fig. 17.3 ). This communicates clearly to growers, traders, and consumers the likely practical implications of climate change.
So far, the news is all bad. However, climate change can bring positive benefi ts in certain places and to some shellfi sh fi sheries and farming systems. When we consider the range of potential pathways of impact of variables
Florida red tide respiratory syndrome), Dinophysis and Prorocentrum (diarrhetic shellfi sh poisoning). In a global review of health issues relating to climate change and oceans, Moore et al. (2008) state that, “ . . . evi-dence that climate change has infl uenced the frequency, duration, and geographical range of HABs is emerging as monitoring data . . . accu-mulate. ” Similarly, Marcogliese (2008) links past climate variability with the dynamics of pathogens and parasites to hypothesize that “ transmission rates of parasites and pathogens are expected to increase with increasing tem-perature. Evidence suggests that the virulence of some pathogens and parasites may also increase with global warming. ” Despite this apparent weight of evidence, we have limited understanding of marine ecosystem responses to the multiple, interacting pathways of poten-tial impact of climate change, and lack of any knowledge of whether the marine biota can
Figure 17.3 Potential climate change impacts on Puget Sound shellfi sh toxicity. Climatological monthly means of reconstructed sea surface temperature ( SST ) in Sequim Bay, Puget Sound, from 1921 to 2007 indicate an average 68 - day duration of the 13 ° C threshold for accelerated growth of the paralytic shellfi sh poisoning agent Alexandrium catenella (shaded period). Scenarios for warmer SST conditions by 2, 4, and 6 ° C are shown in gray with the associated widening of the window of increased opportunity for Alexandrium catenella growth. (Reproduced from Moore et al. 2008 .)
Sea surface temperature
13°C threshold
Month
J F M M J J A S O N D
Historical windowof increased opportunity
68 days
A
Tem
pera
ture
(°C
)
20
15
10
+2°C 69 more days
+4°C 127 more days
+6°C 191 more days

476 Shellfi sh Aquaculture and the Environment
exploited scallop populations in the middle of their current species range show an increase in recruitment — associated with warm years in a variable climate in the case of Peruvian scal-lops and with both warm years and an overall warming trend in the case of European scal-lops (Fig. 17.4 ).
associated with ocean warming and changes in currents (Table 17.3 ), the outlook becomes more mixed. While some fi eld observations and experimental studies show negative population - level impacts of warming on clam and abalone populations (Philippart et al. 2003 ; Vilchis et al. 2005 ), studies of wild,
Figure 17.4 Warmer waters may favor shellfi sh reproduction: evidence from recruitment to scallop fi sheries. (A) Time series of mean spring sea temperature (a principal component derived from four time series of water temperature) in the North Irish Sea, and scallop ( Pecten maximus ) recruitment (R) 2 years later (reproduced from Shephard et al. 2010 ). (B) Annual catch of the Peruvian scallop ( Argopecten purpuratus ) and sea surface temperature changes in Pisco, Peru, 1982 – 2003 (Badjeck et al. 2009 ). During the 1982 – 1983 El Ni ñ o, a 2 - year time lag is observed between temperature peaks and increases in landings as fi shing effort peaked in 1984 – 1985 when the 1982 – 1983 cohort reached its market-able size. In the 1997 – 1998 El Ni ñ o, the scallop stock again increased but instead of waiting for the cohort biomass to build up, small juvenile scallops of low market value were extracted immediately, leading to both a shorter lag (1 year) and to lower landings. Temp PCA, temperature principle component analysis.
R
R (
# a
ge–2 s
callo
ps tow
–1)
Tem
p P
CA
140
120
100
80
60
40
20
0
2.5
2
1.5
1
0.5
0
–0.5
–1
–1.5
–2
Temp PCA
(A)
Year
1990 1995 2000 2005 2010
0
500
1000
1500
2000
2500
3000
3500
4000
4500
(B)
82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 00 01 02 03
Landin
gs (
10 o
f to
ns)
10
11
12
13
14
15
16
17
18
19
20
Sea S
urf
ace T
em
pera
ture
s (
°C)
Landings Sea Surface Temperatures

Implications of global climate change 477
stressors have been made, although usually these are based on inductive reasoning rather than empirical evidence or formal prediction. This is not a criticism: Plausible scenarios of climate change ’ s potential impacts are required to inform policy decisions, and expert judge-ment based on a broad knowledge of path-ways and relationships can be useful. For example, according to Bell et al. (2010) , pearl farming in the tropical Pacifi c faces risks from increased acidifi cation of the ocean as the shells of black - lipped pearl oysters will be weaker. This is likely to lead to higher rates of predation of juveniles and lower rates of col-lection of wild spat. Large - scale farms may be forced to rely more heavily on hatcheries to produce spat, increasing production costs. Acidifi cation may also impair the ability of pearl oysters to form nacre. If so, pearl quality may decline progressively, reducing the value of pearls produced in the future. More severe cyclones can also be expected to increase the risk of damage to the infrastructure of pearl farms in subtropical Pacifi c Island countries.
Major shellfi sh culture sites also merit a holistic view of the potential impacts of climate change. Paolisso et al. (2010) have conducted an overall diagnosis of the potential impact of climate change on Chesapeake Bay that serves as a good example. Scenarios for CO 2 emis-sions indicate that by the end of the twenty - fi rst century the bay region will experience increases in CO 2 concentrations, sea level, and water temperature of the order 50 – 160%, 0.7 – 1.6 m, and 2 – 6 ° C, respectively. Also likely are increases in precipitation amount (very likely in the winter and spring) and intensity, and the intensity of cyclones (though their fre-quency may decrease), and in sea level vari-ability. The greatest uncertainty is associated with changes in annual streamfl ow, though it is likely that winter and spring fl ows will increase. Climate change alone will cause the bay to function very differently in the future. The authors conclude that likely changes include increases in coastal fl ooding and sub-mergence of estuarine wetlands, increases in
Elevated temperatures of coastal waters could also lead to increased production of aquaculture species by expanding their range. These species could be cultivated in higher lati-tudes as well as in existing farms as a result of a longer warm season during which water tem-perature will be near optimal. A decrease in sea ice cover could widen the geographical boundaries, allowing cultivation of commer-cially valuable species in areas hitherto unsuit-able (McCarthy et al. 2001 ). However, tropical species are often already near their thermal limits and a slight temperature change might have signifi cant negative effects on their physi-ology. In the case of bivalves, tropical species live closer to their upper thermal lethal limits than temperate species, which are better adapted to a wider temperature range fl uctua-tion (Compton et al. 2007 ). This has poten-tially serious consequences for tropical shellfi sh farmers, especially as overall vulnerability of aquaculture systems to climate change is higher in those developing countries in the tropics that have a high risk of exposure to multiple climate stressors; a relatively greater economic and dietary dependence on fi sh and shellfi sh resources than wealthier countries with larger, more diverse economies; and limited government and personal resources (such as savings and insurance) with which to adapt to change (Handisyde et al. 2006 ; Allison et al. 2009 ).
Complex, multifactorial impacts have also been demonstrated from review of recent change, monitoring of the impacts of extreme events, and from laboratory studies that manipulate more than one variable (Table 17.3 ). Again, the prognosis is mixed. Reduced oxygen concentration in warmer, more strati-fi ed waters will impact molluscs less than other coastal shellfi sh, such as crabs (Vaquer - Sunyer and Duarte 2008 ). Resilience of mussel popu-lations in estuaries may decrease if extreme fl ow events become more frequent or more intense (Oliver et al. 2008 ).
For particular species and culture systems, predictions that integrate a range of possible

478 Shellfi sh Aquaculture and the Environment
ence and act as one among a range of disincen-tives for them to return.
In aquaculture, problems with disease man-agement, limited availability of suitable sites, and a lack of stable and equitable access to input and output markets mean that it can be a high - risk venture in many parts of the world. The addition of climate - associated risks could slow the growth of aquaculture in developing countries and reduce recent gains in closing the “ supply gap ” through provision of lower - cost food to lower - income consumers, and provid-ing millions of small - holder farmers in devel-oping countries with a new cash crop for export and a nutritious supplement to staple crops (De Silva and Soto 2009 ).
Adapting s hellfi sh f arming to c limate c hange i mpacts
Projected impacts of climate change tend to sound apocalyptic when they do not address the possibility of adaptation. In this case, we are considering adaptation as the actions of people and their institutions, rather than in the Darwinian sense of genetic change by organ-isms in response to changing environments, although that, too, may occur under the selective pressure of rapid climate change. Humans — even conservative ones — are highly adaptable, and adaptation is possible at mul-tiple scales and in multiple domains. Figure 17.5 provides a framework through which to summarize the possibilities for adaptation in the shellfi sh farming sector.
Modifying slightly the ideas of Preston and Stafford - Smith (2009) and Grafton (2010) , we consider: Who can adapt? What elements of the system can be adapted? How can resources and capacity required to adapt be mobilized, and what processes support these adaptation options?
Adaptive actions can be taken by individual shellfi sh farmers, processors, and traders, or even individual consumers ( “ Who ” ?). An
salinity variability, in HABs, and the extent and duration of hypoxia. Eelgrass, the domi-nant submerged aquatic vegetation in the bay, will decrease in abundance and extent. There are also likely to be alterations in interactions among trophic levels, with subtropical fi sh and shellfi sh species ultimately being favored in the bay. The magnitude of these changes is sensi-tive to the CO 2 emission trajectory, so that actions taken now to reduce CO 2 emissions will reduce longer - term climate impacts on the bay. More studies of this type will be useful to the shellfi sh farming sector in other major growing areas.
Almost all the studies in the literature focus on the biological impacts of climate change on production systems, but climate change will impact all links in the value chain linking pro-ducers and consumers. For example, any increases in the intensity and frequency of extreme climatic events such as storms, fl oods, and droughts will negatively impact aquacul-ture production and may also result in signifi -cant damage mainly related to decreased farming capacity (loss of infrastructure) or decreased access to markets (damaged roads). This may translate into economic losses that small - scale farmers are unable to cope with. In Indian River Lagoon, FL, the Florida Department of Agriculture estimated Hurricanes Frances and Charley (2004) caused $8.7 million in losses for clam and oyster farmers (Bierschenk 2004 ). This estimate per-tained only to the value of the stock and to lost incomes and to traded value; it did not include infrastructure losses such as to build-ings, docks, vessels, and nursery and hatchery facilities. Climate change may also act through indirect pathways which may be diffi cult to foresee. For example, Yeoman and McMahon - Beattie (2006) , in a study of the potential impacts of climate change on tourism to Scotland, included a concern that removing local shellfi sh from menus, as a result of increased risk of disease associated with warming, could detract from visitor experi-

Implications of global climate change 479
time periods, such as rezoning a coastal area to refl ect habitat changes. In conjunction with the private sector, governments can help shell-fi sh farmers cope with climate change impacts through risk sharing and transfer mechanisms such as insurance schemes and emergency funds. For instance in the United States, clam growers can be insured for losses that occur because of decrease in salinity, disease, freeze, hurricane, ice fl oe, oxygen depletion, storm surge, and tidal wave under the Cultivated Clam Pilot Crop Insurance Program, part of the Federal Crop Insurance Program (USDA 2010 ). However, state - sponsored crop and income insurance is only a partial solution; it does not prevent the loss of lives or assets, and has been criticized as being a new mechanism for transferring subsidies (van Anrooy et al. 2006 ).
Governments have also an important role in creating an enabling environment for adaptation to take place, and a regulatory
example of autonomous adaptation is described by Nell (2001) in New South Wales, Australia, where summer temperatures on exposed leases can rise to 30 – 40 ° C and kill both Sydney rock and Pacifi c oysters. The problem is most acute when low tides coincide with the warmest period on a hot summer ’ s day. Some farmers have installed irrigation sprinkler systems on their leases and spray saltwater over the oysters to keep them cool. Other farmers use shade cloth over trays to keep oysters cool and predators out, but this practice restricts water fl ow over the trays and increases the risk of mudworm infestations. Spraying oysters with seawater also keeps the oysters cooler than the shade cloth does.
Individuals can be adaptive in adjusting to short - run changes of the type described above and to short - lived “ shocks, ” while govern-ments and communities have a comparative advantage in large - scale adaptation that requires substantial investment over longer
Figure 17.5 The adaptation process. (Adapted from Preston and Stafford - Smith 2009 and Grafton 2010 .)
What to Adapt?
How to Adapt?Who Adapts?
Biophysical System
Individuals Groups Drawing on Capital
Approaches
Responses
Government
Social and Economic system
Scale
Habitat
Human
Autonomous
Social Economic Technical
Palnned Flexible Mandated
Social Natural Physical Financial
Employment
Local Regional National Global
Market Governance
Water quality Species
Farmers
Traders
Processors
Retailers
Consumers
Firms
Communities
Producer associations
Regulatory bodies
Advisory services

480 Shellfi sh Aquaculture and the Environment
require new markets to be developed, or exist-ing consumer preferences to be adjusted. In today ’ s globalized seafood markets, such adjustments are relatively minor and take place quickly. Governance arrangements may also have to adapt to new conditions — this can mean anything from extending the coverage of food safety inspections to changing regulations to allow species translocations, changing legal designation of shellfi sh growing areas, or rene-gotiating leases to the seabed to allow for changing distribution of cultivated shellfi sh.
Adaptive measures can be implemented ( “ How to adapt ” ?) by deploying available capital, whether that comprises knowledge and skills (human capital), or involves mobiliz-ing farmers ’ groups to lobby for changing policy or legislation, or simply getting support from friends and neighbors to deal with loss of assets during a storm or fl ood (social capital). Land or water rights (natural capital) can be sold, leased, or deployed for different uses (e.g., coastal leisure tourism). Processing plants can be adjusted to new product lines and small boats switched from tending fi sh farms to inshore fi shing (physical capital). Loans, savings, and insurance (fi nancial capital) can all be mobilized to invest in new ventures, ease temporary production turn-downs, or recover from shocks such as fl ood damage of closure due to shellfi sh poisoning outbreaks. Improved depuration technologies can be brought into use to deal with increased microbial or HAB concerns.
Adaptation can be autonomous (bottom - up), planned (top - down), negotiated between stakeholders (fl exible), or mandated in law, depending on the nature of required response. Autonomous decisions usually take place at farm or household level, while planned responses, such as investment in disaster pre-paredness and response strategies, require multilevel planning and coordination. An example of autonomous adaptation to fre-quent storms is found among the oyster farmers of the western coast of Taiwan (Chen
environment that prevents reactive adaptation that has negative environmental, social, and economic consequences.
At national level, climate change adaptation should be mainstreamed in aquaculture devel-opment plans. In Peru, where scallops repre-sented 43% of the total value aquaculture products exported in 2009(Produce 2010a ), the new aquaculture development plan 2010 – 2021 (Produce 2010b ) considers climate changes and natural disasters but no explicit risk management and adaptation plans are put forward for scallops, a resource highly sensi-tive to climate fl uctuations (Wolff and Mendo 2000 ). Similarly, national adaptation pro-grams of action ( NAPA s) prepared by least developing countries should include the aqua-culture sector since they are a basis for chan-neling the adaptation funds promised in recent global climate change agreements (UNFCCC 2009 ). Some NAPAs contain provision for adaptation of fi shing and fi sh farming - dependent communities, but in light of their high vulnerability, more needs to be done to support adaptation through capacity building, fi nance, technology, and innovation.
In terms of what actions can be taken ( “ What to adapt ” ?), adaptations to ensure that farming systems are suited to the changing biophysical conditions may include choice of species cultivated or location chosen, or, at a larger scale, water quality or habitat manage-ment to maintain good growing conditions in the face of change. As an example of adapta-tion through species change, due to increased sea surface temperatures attributed to global warming, abalone ( Haliotis ), are now present in England ’ s West Country and the local gov-ernment has been promoting this new industry to rejuvenate the ailing fi shing sector (Brown and Sutton 2002 ).
Maintaining a livelihood or a community may entail adjustments in livelihood strategy and employment, including diversifi cation out of shellfi sh farming if conditions become unsuitable. Changes in species grown may

Implications of global climate change 481
likely to have to increase their investments in developing coherent “ climate - proof ” sectoral policy and legislation, management, and devel-opment, including some of the elements described above. The following costs are likely to arise to address adaptation, or in response to fi sheries and aquaculture decline due to climate change:
• Investments in “ climate - proofi ng ” infra-structure (e.g., coastal defenses, design of harbors);
• Social and economic costs of redundancies in fi shing and aquaculture sector;
• Weather - linked unemployment insurance for fi sherfolk (mostly used in developed countries; few developing countries have this kind of formal social security system);
• Costs of ecosystem maintenance and repair — for example, marine protected areas, restoration of salt marshes and man-groves, artifi cial reef construction, beach replenishment, maintaining “ environmental fl ows ” in rivers; and
• There will also be a need to consider the costs to fi sheries and aquaculture of adapta-tion in other sectors; for instance, the use of “ soft ” coastal defenses with fi sh and shell-fi sh habitat value (mangroves, artifi cial reefs, protecting existing reefs) where fi sher-ies and aquaculture are important to coastal livelihoods, rather than “ hard ” defenses such as sea walls, which prioritize land values over coastal habitat values.
Many of the adaptation options and processes described above are not specifi c to the shellfi sh sector, so, to conclude this section, it seems appropriate to consider the sorts of sector - specifi c technical adaptations we foresee. First, we think there will be widespread changes in species cultured in existing shellfi sh - growing areas. Second, the increased frequency of HABs could close off fi sheries and farming areas more frequently, and there will be a need for improved surveillance systems to prevent
2008 ): Information available on the size and timing of typhoons (the provision of which is a planned adaptive response) is used by farmers to make decisions on dates of setting and har-vesting oysters. Farmers also adapt to the increased frequency and severity of storms by selling small - size or medium - size oysters that can be harvested between typhoon seasons, rather than growing to a larger size and risking loss in the typhoon season.
Responses to climate change can be techni-cal (e.g., early warning systems, “ climate - proofi ng ” of infrastructure, improved means of testing for shellfi sh toxicity and depura-tion), economic (e.g., weather - linked insur-ance, planned economic restructuring to reduce dependence on climate - sensitive indus-tries), or social/institutional (e.g., community self - help programs, participation in grassroots adaptation knowledge networks).
Adaptation is also a political process. Climate change justice is a term usually heard with reference to the moral obligation of high - emissions wealthy nations to assist adaptation by low - emission poor nations that are vulner-able to climate change, but it may apply on a microscale as well. Shellfi sh farmers are likely to be in the vanguard of climate change as they often experience the impacts of changes on both land (and freshwater) and sea. Issues such as investment in sea defenses versus managed retreat in the face of sea level rise are not purely economic ones, as they involve preser-vation or loss of culturally valued landscapes and of people ’ s homes and livelihoods. For example, in Norfolk, England, the principle of social justice has been invoked by local resi-dents and politicians to challenge a policy deci-sion to discontinue maintenance of coastal defenses (McKenna et al. 2009 ).
Costs of a daptation
To maintain the fl ow of benefi ts to society and the economy from fi sheries, governments are

482 Shellfi sh Aquaculture and the Environment
sumption constitutes the most obvious and widely assessed source of carbon emissions from aquatic farming (Bunting and Pretty 2007 ). They are generated through a range of activities, for example, the collection/production of juveniles, general system opera-tions, harvesting, processing, and distribution of the product. The dependence of an aquacul-ture system on industrial energy varies with the means of production, the intensity of the oper-ation, the degree of mechanization, and the quality and quantity of feed used (Troell et al. 2004 ). Indirect or embodied energy inputs nec-essary for aquaculture production include, for example, the energy required to sustain human labor and to build and maintain fi xed capital assets such as farm infrastructure, processing facilities, harvesting machinery, and transpor-tation equipment. Depending on the aquacul-ture system, the scale and form of these inputs will vary widely (Tyedmers and Pelletier 2007 ). In different studies, the differentiation between direct and indirect energy use may vary. Most importantly, the energetic costs of producing, harvesting, processing, and transporting feed components are sometimes counted as direct, and in other cases as indirect, energy consump-tion. This needs to be kept in mind when com-paring the energy effi ciencies and carbon footprints of different food production systems estimated by different studies. In Table 17.4 , the global warming potential ( GWP ) and cumulative energy demand of different aqua-culture systems is presented, highlighting the relatively low GWP and energy inputs for shell-fi sh aquaculture.
Various possibilities exist to reduce green-house gas emissions in aquaculture still further, at both farm level and higher levels.
At the farm level, these include the reduc-tion of energy and fuel use, the generation and application of renewable energies, as well as the adoption of more resource - effi cient culture practices. Direct and indirect fossil fuel use on farms can be reduced by use of energy effi cient machinery, minimizing waste, using energy -
health incidents to consumers and economic losses to growers. Third, given the unpredict-able nature of climate change impacts, we can foresee a shift toward adoption of more “ climate - proof ” closed or recirculation systems in aquaculture — akin to intensive poultry production — where the environment is under more direct human control. Aquaculture dependent on the collection from the wild of climate - sensitive natural larvae, spat, or “ seed ” could begin to disappear in their current loca-tions. An obvious adaptive measure is to close the life cycle of these species by developing hatcheries, but this can be both technologically demanding and expensive. With strong markets providing an incentive for continued innovation, we are confi dent that the sector will adapt and persist.
Shellfi sh a quaculture and c limate c hange m itigation
Aquaculture is not a large contributor to agri-cultural greenhouse gasses, and neither, there-fore, does it have enormous global potential for halting or reversing climate change, but, nevertheless, it is responsible for a small (and currently unquantifi ed) proportion of the greenhouse gas emissions from agriculture, which formed around 13.5% of total anthro-pogenic emissions in 2004 (IPCC 2007b ). There is also potential for making aquaculture a lower - carbon food production system. There have even been discussions on the role that aquaculture could play in carbon capture and storage. Molluscan shellfi sh aquaculture fea-tures in both the debate on low - carbon food futures and on aquaculture ’ s potential for mitigation.
Aquaculture systems are responsible for greenhouse gas emissions, caused by direct energy consumption, indirect or embodied energy consumption, land conversion, and as a result of soil, water, and waste management (Bunting and Pretty 2007 ). Direct energy con-

Table 17.4 Global warming potential ( GWP ) and cumulated energy demand ( CED ) for aquatic products derived from aquaculture, shellfi sh in boldface.
Species System GWP (kg CO 2 e t − 1 ) CED (MJ t − 1 ) Source
Turbot Inland recirculating system; France
6017 290,986 Aubin et al. (2009)
Arctic char Land - based freshwater recirculating system; Canada
10,300 233,000 Ayer and Tyedmers (2009)
Atlantic salmon
Land - based saltwater fl ow - through system; Canada
5410 132,000
Rainbow trout
Freshwater raceways; France 2753 78,229 Aubin et al. (2009)
Prawns Extensive polyculture system (prawn, tilapia, milkfi sh and crab) Philippines
5108 67,000 Baruthio et al. (2009)
Trout Experimental recirculating system; France
1602 57,659 d ’ Orbcastel et al. (2009)
Sea bass Sea cages; Greece 3601 54,656 Aubin et al. (2009)
Pangasius Pump feed; Vietnam ca. 1320 54,411 Henriksson (2009)
Salmon United Kingdom 3270 47,900 Pelletier et al. (2009)
Polyculture Extensive polyculture system (prawn, tilapia, milkfi sh, and crab); Philippines
3553 46,000 Baruthio et al. (2009)
Atlantic salmon
Marine fl oating bag system; Canada
2250 37,300 Ayer and Tyedmers (2009)
Trout Flow - through system; France 2015 34,869 d ’ Orbcastel et al. (2009)
Salmon Chile 2300 33,200 Pelletier et al. (2009)
Salmon Canada 2370 31,200
Atlantic salmon
Conventional marine net pen system; Canada
2073 26,900 Ayer and Tyedmers (2009)
Tilapia Pond based; Indonesia 2100 26,500 Pelletier and Tyedmers (2010)
Salmon Norway 1790 26,200 Pelletier et al. (2009)
Tilapia Lake based; Indonesia 1520 18,200 Pelletier and Tyedmers (2010)
Milkfi sh Semi - intensive; Philippines ca. 1050 14,879 Henriksson (2009)
Pangasius Pond based, intensive; Vietnam
8,930 13,200 Bosma et al. (2009)
Mussel — 12,000 Troell et al. (2004)
Milkfi sh Intensive; Philippines ca. 920 11,547 Henriksson (2009)
Milkfi sh Extensive; Philippines ca. 500 10,799
Pangasius Tidal feed; Vietnam ca. 1030 9861
Mussel Longline system — 4000 Tlusty and Lagueux (2009)
Oysters Low - maintenance aquaculture system; Thailand
ca. 80 580 Henriksson (2009)
483

484 Shellfi sh Aquaculture and the Environment
Ca HCO CaCO CO H O23 3 2 22+ −+ → + +
(17.1)
The calcifi cation process thus actually releases CO 2 into the water, rather than contributing to its net removal from the atmosphere (Chauvaud et al. 2003 ; Laffoley and Grimsditch 2009 ). Therefore, shellfi sh farming does not seem an appropriate means for effective carbon sequestration. However, the carbon cycles in marine systems are very complex involving tightly coupled processes such as photosynthe-sis, respiration, calcifi cation, and calcite dis-solution (Chauvaud et al. 2003 ), and further research may lead to different understanding of carbon cycling in coastal waters and the potential role of shellfi sh. We do not, however, expect piles of oyster shells to be eligible for carbon payments anytime soon. The most useful contributions of shellfi sh farming to climate change mitigation is the compatibility of such farming with the maintenance of coastal environments that do act as carbon sinks, and the energy effi ciency with which they can produce high - quality food for people.
Conclusion
Molluscan shellfi sh fi shing and farming is a small and climate - sensitive sector of the world economy, but one that is both locally impor-tant to sustain livelihoods and coastal cultures and environments, and globally valued for its contribution to many distinctive food cultures — from New Orleans ’ oysters Rockefeller, to the cockles that fl avor Penang ’ s renowned street food noodle dish char kuay teow , and from the quotidian moules - frites of Europe ’ s pays - bas to the upmarket, ice - sculpted displays of fruits de mer outside Brussels ’ Rue des Bouchers restaurants.
Shellfi sh aquaculture is also a dynamic and responsive sector which already adapts to rapidly changing environments and markets. The recent reorientation of environmental
effi cient lighting, using low - carbon and/or recycled building materials, sourcing inputs locally (feed, seed, fertilizers, etc.) and selling to local markets (Bunting and Pretty 2007 ).
As well as emissions reductions, the coastal environment is also receiving attention as a potential site of carbon sequestration, defi ned as “ the process of increasing the carbon content of a reservoir other than the atmo-sphere ” (UNEP 2006 ). Much of this interest relates to maintaining vegetation - dominated coastal ecosystems that are net carbon sinks, such as salt marshes, mangroves, and eelgrass beds (Laffoley and Grimsditch 2009 ). There is also growing interest in using microalgae and seaweed (macroalgae) cultivation as a means to sequester carbon, as they can do so more effi ciently then terrestrial plants (Kaladharan et al. 2009 ; Kumar et al. 2010 ). They could potentially be grown with shellfi sh in inte-grated farming systems, to make such systems “ carbon neutral. ”
The big unresolved question is whether shellfi sh farms can sequester carbon — a pro-posal made due to the fact that the shell con-sists to a large extent of calcium carbonate — CaCO 3 (Bunting and Pretty 2007 ; Hickey 2008 ). Hickey (2008) demonstrated that the potential of oysters to sequester carbon could even be competitive with some plant species.
Based on production data relating to the culture of mussels ( Mytilus edulis ) on rafts in Killary Harbour, Ireland, Rodhouse and Roden (1987) estimated that 10.8 t C year − 1 would be assimilated in mussel production and that the removal rate of carbon during harvest was 0.008 t C m − 2 year − 1 , equating to 80 t C ha − 1 year − 1 .
However, the results of this study are based, most importantly, on the shell carbon content without considering factors such as respiration or the calcifi cation process. Calcifi cation induces shifts in the seawater carbonate equi-librium to generate dissolved CO 2 (Chauvaud et al. 2003 ):

Implications of global climate change 485
monitoring is already vital to the sector so the potential increases in harmful algae popula-tions, and on other pests and pathogens, are likely to be rapidly detected, which will help with timely response and adaptation. Shellfi sh farming associations also have an opportunity to market their products as “ low - carbon ” food sources. Finally, maintaining clean coastal environments for growing shellfi sh may also contribute to the maintenance of other coastal ecosystem services, including carbon seques-tration. Adaptable growers, buoyant market demand, effective monitoring and regulatory systems, and synergies between shellfi sh farming, coastal environmental protection, and low - carbon food production systems will all help ensure a future for shellfi sh farming in a changing climate. It would all be so much simpler, though, if we could agree to cut emis-sions to slow and reverse the warming trend.
Acknowledgments
Thanks to Malcolm Beveridge for early guid-ance on the structure and scope of this chapter, and for helpful comments and suggestions on a later draft. This review was made possible by funding from the U.K. Natural Environment Research Council QUEST thematic program (QUEST_Fish) and from the German Academic Exchange Services (DAAD)
Literature cited
Allison , E.H. , Perry , A.L. , Badjeck , M - C. , Adger , W.N. , Andrew , N.A. , Brown , K. , Conway , D. , Halls , A. , Pilling , G.M. , Reynolds , J.D. , and Dulvy , N.K. 2009 . Vulnerability of national economies to potential impacts of climate change on fi sheries . Fish and Fisheries 10 : 173 – 196 .
Á lvarez - Salgado , X.A. , Labarta , U. , Fern á ndez - Reiriz , M.J. , Figueiras , F.G. , Ros ó n , G. , Piedracoba , S. , Filgueira , R. , and Cabanas , J.M. 2008 . Renewal time and the impact of harmful algal blooms on the extensive mussel raft culture
research on shellfi sh toward understanding the impacts of climate change is building the knowledge base required for planned adapta-tion but we still face a daunting complexity of multiple interacting pathways of impact linking climate with the operation and eco-nomics of shellfi sh culture and the marketing of shellfi sh. The three main areas of adaptive response — social, economic, and technical — will all have a part to play. Their relative importance and ultimate success overall in terms of resilient shellfi sh value chains will be very much dependent on context: location, species, technologies, vulnerabilities of pro-ducers, and fl exibility of markets.
Apart from some scattered anecdotes, at present there is little documented experience of the adaptive strategies pursued by shellfi sh farmers and other actors in the shellfi sh value chain, but this is unsurprising as climate change adaptation is a relatively recent fi eld of applied research. What several authors have noted is that individual small - scale producers and traditional production environments are vulnerable, while major growers and retailers are resilient as they have the capital and national or global reach to invest in sourcing from new areas, developing new markets, and switching to new products. Understanding processes of adaptation at multiple scales should be prioritized as an area of research on which future adaptive responses can be developed.
There are also surprising gaps in our knowl-edge of carbon cycling in coastal waters and oceans — an omission being addressed by the United Nations Environment Programme ’ s “ Blue Carbon ” initiative (Nellemann et al. 2009 ).
Climate change will not destroy the shellfi sh farming sector, but adapting will impose further costs on it. In some places, however, there may be gains rather than losses as warming increases spatfall and growth rates and potentially extends the range of farming toward the poles. Close environmental health

486 Shellfi sh Aquaculture and the Environment
of the Iberian coastal upwelling system (SW Europe) . Harmful Algae 7 ( 6 ): 849 – 855 .
Anderegg , W.R.L. , Prall , J.W. , Harold , J. , and Schneider , S.H. 2010 . Expert credibility in climate change . Proceedings of the National Academy of Sciences of the United States of America 107 ( 27 ): 12107 – 12109 .
Aubin , J. , Papatryphon , E. , van der Werf , H.M.G. , and Chatzifotis , S. 2009 . Assessment of the envi-ronmental impact of carnivorous fi nfi sh produc-tion systems using life cycle assessment. The sustainability of seafood production and con-sumption . Journal of Cleaner Production 17 : 354 – 361 .
Ayer , N.W. , and Tyedmers , P.H. 2009 . Assessing alternative aquaculture technologies: life cycle assessment of salmonid culture systems in Canada. The sustainability of seafood produc-tion and consumption . Journal of Cleaner Production 17 : 362 – 373 .
Badjeck , M. - C. , Mendo , J. , Wolff , M. , and Lange , H. 2009 . Climate variability and the Peruvian scallop fi shery: the role of formal institutions in resilience building . Climatic Change 94 ( 1 – 2 ): 211 – 232 .
Badjeck , M. - C. , Allison , E.H. , Halls , A.S. , and Dulvy , N.K. 2010 . Impacts of climate variability and change on fi shing - based livelihoods . Marine Policy 34 ( 3 ): 375 – 383 .
Barange , M. , and Perry , I. 2009 . Physical and eco-logical impacts of climate change relevant to marine and inland capture fi sheries and aquacul-ture . In: Cochrane , K. , De Young , C. , Soto , D. , and Bahri , T. (eds.), Climate Change Implications for Fisheries and Aquaculture. Overview of Current Scientifi c Knowledge . Food and Agriculture Organization of the United Nations , Rome , pp. 7 – 106 .
Baruthio , A. , Aubin , J. , Mungkung , R.L.J. , and van der Werf , H.M. 2009 . Environmental assess-ment of Filipino fi sh/prawn polyculture using Life Cycle Assessment . In: Nemecek , T. , and Gaillard , G. (eds.), Proceedings of the 6th International Conference on LCA in the Agri - Food Sector — Towards A Sustainable Management of the Food Chain. November 12 – 14, 2008, Z ü rich, Switzerland , pp. 242 – 247 .
Behrenfeld , M.J. , O ’ Malley , R.T. , Siegel , D.A. , McClain , C.R. , Sarmiento , J.L. , Feldman , G.C. ,
Milligan , A.J. , Falkowski , P.G. , Letelier , R.M. , and Boss , E.S. 2006 . Climate - driven trends in contemporary ocean productivity . Nature 444 : 752 – 755 . PG(2046; Bell, Batty, Ganachaud, Gehrke, Hobday, Hoegh - Guldberg, Johnson, Le Borgne, Lehodey, Lough, Pickering, Pratchett, Sheaves, Waycott; 2010) Bell , J. , Batty , M. , Ganachaud , A. , Gehrke , P. , Hobday , A. , Hoegh - Guldberg , O. , Johnson , J. , Le Borgne , R. , Lehodey , P. , Lough , J. , Pickering , T. , Pratchett , M. , Sheaves , M. , and Waycott , M. 2010 . Preliminary assessment of the effects of climate change on fi sheries and aquaculture in the Pacifi c . In: The Contribution of Fisheries to the Economies of Pacifi c Island Countries and Territories . Pacifi c Studies Series, Asian Development Bank , Manila, Philippines .
Berge , J.A. , Bjerkeng , B. , Pettersen , O. , Schaanning , M.T. , and Oxnevad , S. 2006 . Effects of increased seawater concentrations of CO 2 on growth of the bivalve Mytilus edulis L . Chemosphere 62 ( 4 ): 681 – 687 .
Biershenck , E. 2004 . Storm damages shock local shellfi sh industry . Victoria, Australia, GROWfi sh, Gippsland Aquaculture Industry Network - GAIN (17/09/2004).
Bosma , R.H. , Hanh , C.T.T. , and Potting , J. 2009 . Environmental impact assessment of the panga-sius sector in the Mekong Delta . Wageningen University.
Brander , K.M. 2007 . Global fi sh production and climate change . Proceedings of the National Academy of Sciences of the United States of America 104 ( 50 ): 19709 – 19714 .
Brierley , A.S. , and Kingsford , M.J. 2009 . Impacts of climate change on marine organisms and eco-systems . Current Biology 19 : R602 – R614 .
Brown , P. , and Sutton , T. 2002 . Global warming brings new cash crop for fi shermen . The Guardian (10 December 2002), London.
Bunting , S.W. , and Pretty , J. 2007 . Global carbon budgets and aquaculture — emissions, sequestra-tion and management options . Centre for Environment and Society Occasional Paper 2007 - 1. University of Essex, UK.
Cane , M.A. 2005 . The evolution of El Nino, past and future . Earth and Planetary Science Letters 230 : 227 – 240 .
Chauvaud , L. , Thompson , J.K. , Cloern , J.E. , and Thouzeau , G. 2003 . Clams as CO 2 generators:

Implications of global climate change 487
the Potamocorbula amurensis example in San Francisco Bay . Limnology and Oceanography 48 : 2086 – 2092 .
Chen , Y.L. 2008 . The adaptive strategies of oyster farmers to climate variation: typhoon in Tainan, Taiwan . Masters thesis, Institute of Marine Aquaculture, University of Taiwan, 121 pp (in Chinese — English abstract accessed via etd.lib.nsysu.edu.tw/ETD - db/ETD - search/view_etd?URN=etd - 0907109 - 173345
Cheung , W.W.L. , Lam , V.W.Y. , Sarmiento , J.L. , Kearney , K. , Watson , R. , and Pauly , D. 2009 . Projecting global marine biodiversity impacts under climate change scenarios . Fish and Fisheries 10 ( 3 ): 235 – 251 .
Compton , T.J. , Rijkenberg , M.J.A. , Drent , J. , and Piersma , T. 2007 . Thermal tolerance ranges and climate variability: a comparison between bivalves from differing climates . Journal of Experimental Marine Biology and Ecology. 352 : 200 – 211 .
Cook , T. , Folli , M. , Klinck , J. , Ford , S. , and Miller , J. 1998 . The relationship between increasing sea - surface temperature and the northward spread of perkinsus marinus (dermo) disease epi-zootics in oysters . Estuarine, Coastal and Shelf Science 46 : 587 – 597 .
De Silva , S.S. , and Soto , D. 2009 . Climate change and aquaculture: potential impacts, adaptation and mitigation . In: Cochrane , K. , De Young , C. , Soto , D. , and Bahri , T. (eds.), Climate Change Implications for Fisheries and Aquaculture. Overview of Current Scientifi c Knowledge . Food and Agriculture Organization of the United Nations , Rome , pp. 151 – 212 .
de Young , B. , Barange , M. , Beaugrand , G. , Harris , R. , Perry , R.I. , Scheffer , M. , and Werner , F. 2008 . Regime shifts in marine ecosystems: detection, prediction and management . Trends in Ecology and Evolution 23 ( 7 ): 402 – 409 .
Diederich , S. , Nehls , G. , van Beusekom , J.E.E. , and Reise , K. 2010 . Introduced Pacifi c oysters ( Crassostrea gigas ) in the northern Wadden Sea: invasion accelerated by warm summers? Helgoland Marine Research 59 ( 2 ): 97 – 106 .
Doney , S.C. 2006 . Plankton in a warmer world . Nature 444 : 695 – 696 .
Doney , S.C. , Fabry , V.J. , Feely , R.A. , and Kleypas , J.A. 2009 . Ocean acidifi cation: the other CO 2
problem . Annual Review of Marine Science 1 : 169 – 192 .
d ’ Orbcastel , E.R. , Blancheton , J. - P. , and Aubin , J. 2009 . Towards environmentally sustainable aquaculture: comparison between two trout farming systems using life cycle assessment . Aquacultural Engineering 40 : 113 – 119 .
Dunlap , R.E. , and McCright , A.M. 2008 . A widening gap: republican and democratic views on climate change . Environment 50 : 26 – 35 .
Dupont , S. , Dorey , N. , and Thorndyke , M. 2010 . What can meta - analysis tell us about vulnerabil-ity of marine biodiversity to ocean acidifi cation? Estuarine, Coastal and Shelf Science 89 ( 2 ): 182 – 185 .
Easterling , W.E. , Rosenberg , N.J. , McKenney , M.S. , and Allan Jones , C. 1992 . An introduc-tion to the methodology, the region of study, and a historical analog of climate change . Agricultural and Forest Meteorology 59 ( 1 – 2 ): 3 – 15 .
Emanuel , K. , Sundararajan , R. , and Williams , J. 2008 . Hurricanes and global warming; results from downscaling IPCC AR4 simulations . Bulletin of the American Metrological Society 89 ( 3 ): 347 – 367 .
ESCAP and UNISDR . 2010 . Protecting develop-ment gains, reducing disaster vulnerability and building resilience in Asia and the Pacifi c . The Asia - Pacifi c Disaster Report, 2010. Economic and Social Commission for Asia and the Pacifi c, United Nations International Strategy for Disaster Reduction, Bangkok, Thailand. 129 pp.
Ford , J.D. , Keskitalo , E.C.H. , Smith , T. , Pearce , T. , Berrang - Ford , L. , Duerden , F. , and Smit , B. 2010 . Case study and analogue methodologies in climate change vulnerability research . Wiley Interdisciplinary Reviews: Climate Change 1 ( 3 ): 374 – 392 .
Gazeau , F. , Quiblier , C. , Jansen , J.M. , Gattuso , J. - P. , Middelburg , J.J. , and Heip , C.H.R. 2007 . Impact of elevated CO 2 on shellfi sh calcifi cation . Geophysical Research Letters 34:L07603.
Glantz , M. 1988 . Societal Responses to Regional Climatic Change: Forecasting by Analogy . Westview Press, Boulder , CO .
Grafton , R.Q. 2010 . Adaptation to climate change in marine capture fi sheries . Marine Policy 34 : 606 – 615 .

488 Shellfi sh Aquaculture and the Environment
Hallegraeff , G.M. 2010 . Ocean climate change, phytoplankton community responses and harmful algal blooms; a formidable predictive challenge . Journal of Phycology 46 ( 2 ): 220 – 235 .
Handisyde , N.T. , Ross , L.G. , Badjeck , M. - C. , and Allison , E.H. 2006 . The effects of climate change on world aquaculture: a global perspective . Final Technical Report. DFID Aquaculture and Fish Genetics Research Programme, Stirling Institute of Aquaculture, Stirling, U.K., 151 pp. www.aqua.stir.ac.uk/GISAP/climate/index.htm
Harley , C.D.G. , Hughes , A.R. , Hultgren , K.M. , Miner , B.G. , Sorte , C.J.B. , Thornber , C.S. , Rodriguez , L.F. , Tomanek , L. , and Williams , S.L. 2006 . The impacts of climate change in coastal marine systems . Ecology Letters 9 : 228 – 241 .
Hendriks , I.E. , and Duarte , C.M. 2010 . Ocean acidifi cation: seperating evidence from judgment—— A reply to Dupont et al. Estuarine, Coastal and Shelf Science 89 ( 2 ): 186 – 190 .
Hendriks , I.E. , Duarte , C.M. , and Á lvarez , M. 2010 . Vulnerability of marine biodiversity to ocean acidifi cation: a meta - analysis . Estuarine, Coastal and Shelf Science 86 ( 2 ): 157 – 164 .
Henriksson , P. 2009 . Energy intensity in tropical aquaculture. Cumulative energy demand of milkfi sh, pangasius and oyster production in SE Asia . Master ’ s Thesis, Department of Systems Ecology, Stockholm University,Sweden.
Hickey , J.P. 2008 . Carbon sequestration potential of shellfi sh. S eminars on Sustainability. University of South Australia. Accessed via: www.oysterssa.com.au/media/fi les/755.pdf
IPCC . 2007a . Climate Change 2007: The Physical Science Basis. Summary for Policymakers . Intergovernmental Panel on Climate Change, UNEP , Geneva .
IPCC . 2007b . Climate Change 2007: Mitigation of Climate Change . Working Group III Contribution to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. UNEP , Geneva .
Kaladharan , P. , Veena , S. , and Vivekanandan , E. 2009 . Carbon sequestration by a few marine algae: observation and projection . Journal of the Marine Biological Association of India 51 : 107 – 110 .
Kim , Y. , Powell , E.N. , Wade , T.L. , Presley , B.J. , and Brooks , J.M. 1999 . Infl uence of climate change on interannual variation in contaminant body burden in Gulf of Mexico oysters . Marine Environmental Research 48 ( 4 – 5 ): 459 – 488 .
Kumar , A. , Ergas , S. , Yuan , X. , Sahu , A. , Zhang , Q. , Dewulf , J. , Malcata , F.X. , and van Langenhove , H. 2010 . Enhanced CO 2 fi xation and biofuel production via microalgae: recent developments and future directions . Trends in Biotechnology 28 ( 7 ): 371 – 380 .
Kurlansky , M. 2006 . The Big Oyster: History on the Half Shell . Random House , New York .
Laffoley , D.d ’ A. , and Grimsditch , G. 2009 . The Management of Natural Coastal Carbon Sinks . IUCN , Gland, Switzerland .
Lazarus , R.J. 2009 . Super wicked problems and climate change: restraining the present to liber-ate the future . Cornell Law Review 94 : 1153 – 1234 .
Lorenzoni , I. , and Pidgeon , N. 2006 . Public views on climate change: european and USA perspec-tives . Climatic Change 77 : 73 – 95 .
Marcogliese , D.J. 2008 . The impact of climate change on the parasites and infectious diseases of aquatic animals . Revue scientifi que et tech-nique 27 ( 2 ): 467 – 484 .
McCarthy , J. , Canziani , O.S. , Learly , N. , Dokken , D. , and White , K. (eds.). 2001 . Climate Change 2001: Impacts, Adaptation, and Vulnerability . Cambridge University Press , Cambridge .
McKenna , J. , Cooper , J.A.G. , and O ’ Hagan , A.M. 2009 . Coastal erosion management and the European principles of ICZM: local versus stra-tegic perspectives . Journal of Coastal Conservation 13 ( 2 – 3 ): 165 – 173 .
Miller , A.W. , Reynolds , A.C. , Sobrino , C. , and Riedel , G.F. 2009 . Shellfi sh face uncertain future in high CO 2 world: infl uence of acidifi cation on oyster larvae calcifi cation and growth in estuar-ies . Public Library of Science One 4 ( 5 ): e5661 . doi:10.1371/journal.pone.0005661
Moore , S.K. , Trainer , V.L. , Mantua , N.J. , Parker , M.S. , Laws , E.A. , Backer , L.C. , and Fleming , L.E. 2008 . Impacts of climate variability and future climate change on harmful algal blooms and human health . Environmental Health 7 ( Suppl. 2 ): S4 .
Nell , J.A. 2001 . The history of oyster farming in Australia . Marine Fisheries Review 3 : 14 – 25 .

Implications of global climate change 489
Nellemann , C. , Corcoran , E. , Duarte , C.M. , Valdes , L. , DeYoung , C. , Fonseca , L. , and Grimsditch , G. (eds.). 2009 . Blue Carbon — The Role of Healthy Oceans in Binding Carbon — A Rapid Response Assessment . UNEP , Nairobi, Kenya .
Oliver , L.R. , Seed , R. , and Reynolds , B. 2008 . The effect of high fl ow events on mussels ( Mytilus edulis ) in the Conway estuary, North Wales, UK . Hydrobiologia 597 ( 1 ): 117 – 127 .
Paolisso , M. , Secor , D. , Sellner , K. , Wardrop , D. , and Wood , R. 2010 . Potential climate - change impacts on the Chesapeake Bay . Estuarine, Coastal and Shelf Science 86 ( 1 ): 1 – 20 .
Parker , L.M. , Ross , P.M. , and O ’ Connor , W.A. 2009 . The effect of ocean acidifi cation and tem-perature on the fertilization and embryonic development of the Sydney rock oyster Saccostrea glomerata (Gould 1850) . Global Change Biology 15 : 2123 – 2136 .
Patz , J.A. , Olson , S.H. , and Gray , A.L. 2006 . Climate change, oceans, and human health . Oceanography 19 ( 2 ): 52 – 59 .
Pelletier , N. , and Tyedmers , P. 2010 . Life cycle assessment of frozen tilapia fi llets from Indonesian lake - based and pond - based intensive aquaculture systems . Journal of Industrial Ecology. 14 ( 3 ): 467 – 481 .
Pelletier , N. , Tyedmers , P. , Sonesson , U. , Scholz , A. , Ziegler , F. , Flysjo , A. , Kruse , S. , Cancino , B. , and Silverman , H. 2009 . Not all salmon are created equal: life cycle assessment (LCA) of global salmon farming systems . Environmental Science & Technology 43 : 8730 – 8736 .
Peperzak , L. 2003 . C limate change and harmful algal blooms in the North Sea . Acta Oecologica 24 ( S1 ): S139 – S144 .
Philippart , C.J.M. , van Aken , H.M. , Beukema , J.J. , Bos , O.G. , Cad é e , G.C. , and Dekker , R. 2003 . Climate - related changes in recruitment of the bivalve Macoma balthica . Limnology & Oceanography 48 ( 6 ): 2171 – 2185 .
Preston , B. , and Stafford - Smith , M. 2009 . Framing vulnerability and adaptive capacity assessment: discussion paper . CSIRO Climate Adaptation Flagship Working Group Paper No 2, CSIRO, Canberra. www.csiro.au/org/climateadaptation-fl agship.html
Produce . 2010a . Peru: Valor de la exportacion de los productos hidrobiologicos procedentes de la
actividad de acuicultura por especie segun pais de destino 2000 – 2009 . www.produce.gob.pe/portal/portal/apsportalproduce/internapesqueria?ARE=3 & JER=460 (accessed on September 12, 2010). PRODUCE, Lima, Peru.
Produce . 2010b . National plan for aquaculture development — PNDA . General Aquaculture Directorate Deputy Minister ’ s Offi ce for Fisheries Ministry of Production, Lima, Peru.
Rahmstorf , S. , Cazenave , A. , Church , J.A. , Hansen , J.E. , Keeling , R.F. , Parker , D.E. , and Sommervile , R.C.J. 2007 . Recent climate observations com-pared to projections . Science 316 ( 5825 ): 709 .
Rodhouse , P.G. , and Roden , C.M. 1987 . Carbon budget for a coastal inlet in relation to intensive cultivation of suspension - feeding bivalve mol-luscs . Marine Ecology Progress Series 36 : 225 – 236 .
Rose , J.B. , Epstein , P.R. , Lipp , E.K. , Sherman , B.H. , Bernard , S.M. , and Patz , J.M. 2001 . Climate vari-ability and change in the United States: potential impacts on water - and foodborne diseases caused by microbiologic agents . Environmental Health Perspectives 109 ( 2 ): 211 – 220 .
Shephard , S. , Beukers - Stewart , B. , Hiddink , J. , Brand , A. , and Kaiser , M. 2010 . Strengthening recruitment of exploited scallops Pecten maximus with ocean warming . Marine Biology 157 ( 1 ): 91 – 97 .
Soniat , T.M. , Klinck , J.M. , Powell , E.N. , and Hofmann , E.E. 2006 . Understanding the success and failure of oyster populations: climatic cycles and Perkinsus marinus . Journal of Shellfi sh Research 25 ( 1 ): 83 – 93 .
Talmage , S.C. , and Gobler , C.J. 2010 . Effects of past, present, and future ocean carbon dioxide concentrations on the growth and survival of larval shellfi sh . Proceedings of the National Academy of Sciences of the United States of America 107 ( 40 ): 17246 – 17251 .
Tlusty , M.F. , and Lagueux , K. 2009 . Isolines as a new tool to assess the energy costs of the produc-tion and distribution of multiple sources of seafood. The sustainability of seafood produc-tion and consumption . Journal of Cleaner Production 17 : 408 – 415 .
Troell , M. , Tyedmers , P. , Kautsky , N. , and R ö nnb ä ck , P. 2004 . Aquaculture and energy use . In: Cleveland , C.J. (ed.), Encyclopedia of Energy . Elsevier , New York , pp. 97 – 108 .

490 Shellfi sh Aquaculture and the Environment
Tyedmers , P. , and Pelletier , N. 2007 . Biophysical accounting in aquaculture: insights from current practice and the need for methodological devel-opment . In: Bartley , D.M. , Brug è re , C. , Soto , D. , Gerber , P. , and Harvey , B. (eds.), Comparative Assessment of the Environmental Costs of Aquaculture and Other Food Production Sectors. Methods for Meaningful Comparisons . FAO/WFT Expert Workshop, 24 – 28 April 2006, Vancouver, Canada. Food and Agriculture Organization of the United Nations , Rome .
UNEP . 2006 . Marine and coastal ecosystems and human well - being . A synthesis report based on the fi ndings of the Millennium Ecosystem Assessment. United National Environment Programme, Nairobi, Kenya.
UNFCCC . 2009 . Copenhagen accord . Advance unedited version. Decision - /CP.15 18 December 2009. unfccc.int/fi les/meetings/cop_15/applica-tion/pdf/cop15_cph_auv.pdf . United Nations Framework Convention on Climate Change. (accessed on Octorber 31, 2010).
USDA . 2010 . Cultivated clam pilot crop insurance underwriting guide . 2010 and succeeding crop years FCIC - 24100 (08 - 2010). United States Department of Agriculture, Federal Crop Insurance Corporation, Product Development Division, Kansas City. 36 pp.
van Anrooy , R. , Secretan , P.A.D. , Lou , Y. , Roberts , R. , and Upare , M. 2006 . Review of the current state of world aquaculture insurance . FAO
Fisheries Technical Paper. No. 493. FAO, Rome. 92 pp.
Vaquer - Sunyer , R. , and Duarte , C.M. 2008 . Thresholds of hypoxia for marine biodiversity . Proceedings of the National Academy of Sciences of the United States of America 105 ( 40 ): 15452 – 15457 .
Vilchis , L.I. , Tegner , M.J. , Moore , J.D. , Friedman , C.S. , Riser , K.L. , Robbins , T.T. , and Dayton , P.K. 2005 . Ocean warming effects on growth, repro-duction, and survivorship of southern California abalone . Ecological Applications 15 ( 2 ): 469 – 480 .
Waldbusser , G.G. , Voigt , E.P. , Bergschneider , H. , Green , M.A. , and Newell , R.I.E. 2011 . Biocalcifi cation in the Eastern Oyster ( Crassostrea virginica ) in relation to long - term trends in Chesapeake Bay pH . Estuaries and Coasts 34 ( 2 ): 221 – 231 .
Wolff , M. , and Mendo , J. 2000 . Management of the Peruvian bay scallop metapopulation with regard to environmental change . Aquatic Conservation Marine Freshwater Ecosystems 10 : 117 – 126 .
Yeoman , I. , and McMahon - Beattie , U. 2006 . Understanding the impact of climate change on Scottish tourism . Journal of Vacation Marketing 12 ( 4 ): 371 – 379 .