shellfish aquaculture and the environment (shumway/shellfish aquaculture and the environment) ||...
TRANSCRIPT

Chapter 10
Bivalves a s b ioturbators and b ioirrigators Joanna Norkko and Sandra E. Shumway
Bivalves a re k ey s pecies in s oft - s ediment h abitats
Both natural and cultured populations of bivalve molluscs provide a range of important ecosystem functions and services (see Chapters 1 , 4 , 8 , and 9 in this book), some of which stem from the activities of different bivalve species living buried in the sediments. Bioturbation and bioirrigation are the mixing and fl ushing of sediments that are the result of the burrowing, feeding, and other activities of the animals in the sediment. Thus, bioturba-tion and bioirrigation are integral to a healthy soft - sediment ecosystem and, in general, infau-nal bivalves such as clams have a positive, desirable infl uence on the sand or mud in
which they live, just as the earthworms in a vegetable patch or garden compost.
Soft sediments are the most widespread habitats in the sea, extending from coastal estuaries and sandy shores, to continental shelves and the open ocean abyssal plains. Bivalve molluscs are often dominant in terms of biomass and/or abundance in estuarine and coastal soft - sediment habitats. Due to their habitat engineering and their infl uence on benthic - pelagic coupling and nutrient cycling, bivalve molluscs are key species in many habi-tats (Chapters 4 and 9 in this book; Jones et al. 1994 ; Dame et al. 2002 ). Consequently, changes in the condition, abundance, or distri-bution of different bivalve species may have cascading effects on both benthic and pelagic
Shellfi sh Aquaculture and the Environment, First Edition. Edited by Sandra E. Shumway.© 2011 John Wiley & Sons, Inc. Published 2011 by John Wiley & Sons, Inc.
297

298 Shellfi sh Aquaculture and the Environment
ecosystems (Vaughn and Hakenkamp 2001 ; Dahlhoff et al. 2002 ; Newell 2004 ; Norkko et al. 2006 ). In addition, bivalves are relatively long - lived and are therefore often used as indi-cator species when monitoring environmental change.
Bivalves are linked to their environment through a multitude of processes that operate over different spatial and temporal scales. Large - scale processes, such as oceanographic conditions, and hydrodynamic and nutrient regimes, defi ne the more general settings and the environmental framework that defi nes, for example, species distribution ranges and the level of primary production. Small - scale pro-cesses, such as individual physiology or behav-ior, are easier to study and amenable to experimental manipulation, but the extrapola-tion and scaling up of the results to the scale of an estuary or ecosystem are much more dif-fi cult. Therefore, generalizations based on small - scale studies are often not accurate at the larger scales of relevance to environmental problems and management practices. The way a pattern or a process is perceived, and there-fore its accuracy, depends to a large extent on the scale at which the phenomenon is studied (Hewitt et al. 2007 ). This highlights the impor-tance of explicitly incorporating scale in all environmental issues, for example, in the way the role of bivalves is assessed. This includes considering which processes operate at which scales, how they are linked, and which ones are most relevant for a particular issue, such as the most effi cient and sustainable siting of aquaculture installations in different types of estuaries (Dumbauld et al. 2009 ).
This chapter focuses on the mechanisms, dynamics, and effects of the bivalve bioturba-tion and bioirrigation processes and how these infl uence the health and stability of marine ecosystems. Particular attention is given to aspects of bioturbation/irrigation and other related sediment processes that are relevant for sustainable management of bivalve aquacul-ture operations.
What a re b ioturbation and b ioirrigation?
Soft - sediment habitats are greatly infl uenced by the activities of different taxa living within the sediment, including bivalves, crustaceans, and polychaete worms. The combination of their movements, feeding, ventilation, burrow-ing, tube construction, and biodeposit produc-tion results in structuring, mixing, and fl ushing of the sediment (Fig. 10.1 , Table 10.1 ). The term “ bioturbation ” refers to the reworking of aquatic sediments by the organisms in the sedi-ments and, in its broadest sense, includes the structuring activities of both burrowing animals and rooting plants, as well as microbes. In addition, animals living on the sediment surface (epifauna) and animals just visiting the sediment in search of food, such as rays and dugongs, may have bioturbating effects. The corresponding processes also occur in terres-trial soils, where earthworms are the most commonly used example. “ Bioirrigation, ” on the other hand, refers to the enhanced trans-port of solutes between the sediment and the overlaying water. This is the fl ushing of burrows that stems from the suspension feeding of the animals and their ventilation activities to facilitate transport of oxygen and excretory products. Bivalves are major players in the modifi cation of sediments, and the effects we observe are a combination of both bioturbation and bioirrigation. In the follow-ing sections, “ bioturbation ” will be used to describe both phenomena, that is, to include both particle and solute transport.
Bioturbation fundamentally affects the physical and chemical characteristics of soft - sediment habitats (Rhoads 1974 ; Aller 1988 ; Kristensen 1988 ; Levinton 1995 ; Aller 2001 ; Reise 2002 ; Meysman et al. 2006b ) and can transform a homogenous sandy or muddy bottom into a heterogeneous landscape with physical structures such as burrows, pits, and mounds. Also, the mere physical presence of, for example, shells of both live and dead

Figure 10.1 Schematic diagram of the processes around a bivalve buried in the sediment.
Excretion of nutrients and CO2
biodeposition of feces and pseudofecesSuspension and/or deposit feedingconsumption of O2
Sediment mixing, flushing,and oxygenation
Added structure
Water column
Oxidized sediment
Reduced sediment
RPD layer
Nut
rient
s
Table 10.1 Combined effects of bivalve physical structure, bioturbation, bioirrigation, and feeding activities in natural bivalve populations in soft - sediment habitats. In addition, the relevance of these processes to aquaculture is summarized.
Process Effects on local environment Relevance to aquaculture
Physical structure Added heterogeneity; burrows, pits, mounds, shell material
Racks, bags, and longlines also provide structure; at high stocking densities, the habitat is homogenized and the diversity reduced
Individual bivalves provide substrate and habitat
Promotes overall higher diversity
Bioturbation; sediment reworking
Oxygenation of the sediment; oxygen penetrates deeper into the sediment
Only applies if bivalves are able to bury into the sediment
Sediment mixing and sorting affects grain size Racks, bags, and longlines may increase siltation
Sediment destabilization, resuspension, and erosion
Prevents siltation
Bioirrigation; ventilation/feeding
Flushing of sediments with overlaying water Only applies if bivalves are able to bury into the sediment
Transport of oxygen into the sediment and excretory products out of the sediment
Biodeposition of feces and pseudofeces on the sediment surface
Benthic - pelagic coupling Potential negative effects are highly dependent on fl ow, depth, and stocking density Increased organic content of the sediment; may
provide food for other benthic species
- Higher microbial activity and oxygen demand in the sediment; potential for hypoxia
Altered nutrient fl uxes at the sediment - water interface
299

300 Shellfi sh Aquaculture and the Environment
tion to the underlying darker reduced sediment layer (Fig. 10.1 ). Depending on sediment type (oxygen penetrates deeper into porous sandy sediments compared with cohesive mud; Volkenborn et al. 2007 ) and the species com-position and abundance of the benthic com-munity, this lighter layer may range from just a millimeter to several centimeters thick. The border between oxidative and reducing reac-tions is called the redox potential discontinuity layer ( RPD ). Bioturbators may substantially extend the depth of this layer, and the depth of the RPD layer is often used as an indicator of environmental status, with deeper oxidized sediment layers indicating the presence of more animals and thus better conditions. This can roughly be measured in sediment cores taken with a see - through corer or quantifi ed, for example, using sediment profi le imagery ( SPI ), where vertical images are taken into the sediment with a prism mounted on a digital camera and subsequently analyzed using image analysis software (Grizzle and Penniman 1991 ). On a fi ner scale, the depth profi les of pH, oxygen, sulfi des, and other compounds can be measured with millimeter accuracy using microsensors penetrating the sediment (Glud 2008 ).
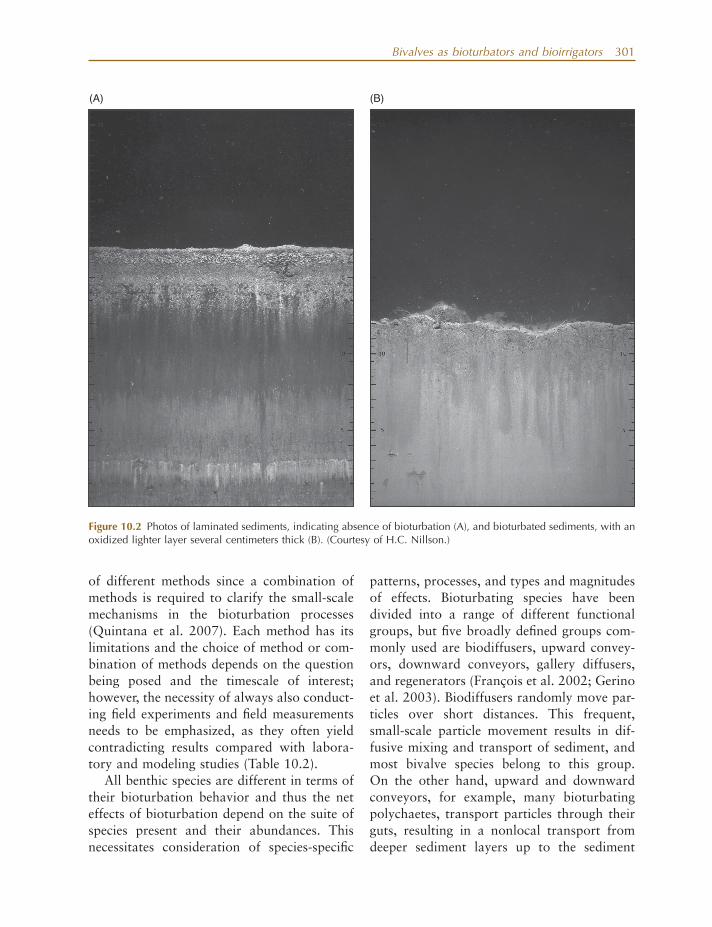
Present - day bioturbation generally does not affect more than about the top 30 cm of the sediment, that is, the sediment layers where most of the infaunal species live (although there are exceptions, such as deep - burrowing geoducks). Where sediment accumulates over time, however, some traces of bioturbation activities are preserved and can be seen in thick sediment layers that are thousands of years old (Erwin 2008 ). Similarly, absence of fauna is also recorded in the sedimentary record in the form of layers upon layers of undisturbed, laminated sediments, which are formed during periods of either natural or eutrophication - induced anoxic conditions (Schaffner et al. 1992 ; Karlson et al. 2002 ; Fig. 10.2 ).
The mechanisms of bioturbation and bioir-rigation have been studied through a number
bivalves in the sediment creates added struc-ture and heterogeneity. Reworking of the sedi-ment affects the sorting of the sediment, its grain size distribution, porosity, and the verti-cal profi les of both solids and solutes. It changes the rates and pathways of reaction processes within the sediment, and between the sediment and the overlaying water (Table 10.1 ). This subsequently affects biodiversity and ecosystem function in soft - sediment habi-tats. In general, bioturbation increases all fl uxes at the sediment – water interface, includ-ing both particulates and solutes. Effi cient fl uxes are a prerequisite for any well - functioning ecosystem and are at the base of all ecosystem services society needs from the environment. Understanding these fl uxes will help manage our coastal ecosystems effi ciently and sustainably.
Bioturbation increases the depth of oxygen penetration into the sediment from the over-laying water. The presence or absence of oxygen governs a multitude of both chemical and biological processes in the sediment, and therefore bioturbation affects the composition of both the invertebrate and microbial com-munities in the sediment (Glud 2008 ). The burrowing, mixing, and tube construction of different invertebrate species substantially increases the area of the sediment – water inter-face, by extending this layer deeper into the sediment as an irregular surface. In combina-tion with active suspension feeding and burrow ventilation, this allows oxygen - rich water to penetrate deeper into the sediment, thereby oxygenating otherwise reduced and anoxic sediments. Bioturbation thus affects the area available for aerobic microbial activity, which affects the functioning of the whole system, since the sediment microbial community per-forms essential functions such as decomposi-tion of deposited organic material and mineralization of nutrients.
The depth of oxygen penetration may be seen as an upper, lighter oxidized sediment layer, with a sometimes relatively sharp transi-
Bivalves as bioturbators and bioirrigators 301
patterns, processes, and types and magnitudes of effects. Bioturbating species have been divided into a range of different functional groups, but fi ve broadly defi ned groups com-monly used are biodiffusers, upward convey-ors, downward conveyors, gallery diffusers, and regenerators (Fran ç ois et al. 2002 ; Gerino et al. 2003 ). Biodiffusers randomly move par-ticles over short distances. This frequent, small - scale particle movement results in dif-fusive mixing and transport of sediment, and most bivalve species belong to this group. On the other hand, upward and downward conveyors, for example, many bioturbating polychaetes, transport particles through their guts, resulting in a nonlocal transport from deeper sediment layers up to the sediment
of different methods since a combination of methods is required to clarify the small - scale mechanisms in the bioturbation processes (Quintana et al. 2007 ). Each method has its limitations and the choice of method or com-bination of methods depends on the question being posed and the timescale of interest; however, the necessity of always also conduct-ing fi eld experiments and fi eld measurements needs to be emphasized, as they often yield contradicting results compared with labora-tory and modeling studies (Table 10.2 ).
All benthic species are different in terms of their bioturbation behavior and thus the net effects of bioturbation depend on the suite of species present and their abundances. This necessitates consideration of species - specifi c
Figure 10.2 Photos of laminated sediments, indicating absence of bioturbation (A), and bioturbated sediments, with an oxidized lighter layer several centimeters thick (B). (Courtesy of H.C. Nillson.)
(A) (B)

302 Shellfi sh Aquaculture and the Environment
bottom of the burrow after it has been deserted. Common to all of these groups is that they facilitate fl uxes of particulate matter and solutes between the sediment and the overlay-ing water, but models of bioturbation need to consider the different types of bioturbation (Michaud et al. 2006 ).
surface or from the sediment surface down, respectively. Gallery diffusers create tube systems and mix sediment in the process. Regenerators also dig tube systems, with addi-tional release of large amounts of sediment into the overlaying water during digging and transport of sediment from the surface to the
Table 10.2 Examples of methodological approaches used for studying bioturbation and bioirrigation (not restricted to only bivalves), the spatial scale these methods apply to (individual organism, laboratory studies, fi eld experiments, fi eld surveys and monitoring, ecosystem level effects), and selected references to studies utilizing these methods.
Method Bioturbation - related effect/process studied Spatial scale Example reference
Visual assessment, photographs, video
Tracks, pits, burrows, mounds Laboratory, fi eld experiments, surveys
Rhoads and Young (1971)
Visual assessment of RPD layer depth
Net effects of bioturbation; oxygen penetration in the sediment
Laboratory, fi eld experiments, surveys
Sediment analysis (including slicing)
Depth distribution of sediment grain size, water content, pelletization, organic content
Laboratory, fi eld experiments, surveys
Maire et al. (2008)
Inert particle addition (e.g., luminophores, glass beads) or natural tracers (radionuclides)
Particle mixing, sediment reworking
Laboratory, fi eld experiments
Gerino et al. (1998) ; Montserrat et al. (2009)
Flume studies Sediment resuspension and erosion
Laboratory Willows et al. (1998)
Inert solute addition (e.g., bromide)
Pore water irrigation Laboratory, fi eld experiments
Martin and Banta (1992)
Microelectrodes, planar optodes
Depth profi les of nutrients, O 2 , S, H 2 S
Very fi ne - scaled; laboratory, fi eld experiments
Glud (2008)
Sediment incubations Fluxes of nutrients, solutes and gasses across the water – sediment interface
Laboratory, fi eld experiments
Asmus et al. (1998) ; Michaud et al. (2006)
Sediment profi le imaging
Two - dimensional structures and oxygen penetration in the sediment
Surveys Grizzle and Penniman (1991) ; Solan et al. (2004)
Computer - aided tomography
Three - dimensional structures in the sediment
Individual cores (slow/expensive)
Perez et al. (1999) ; Bouchet et al. (2009)
Models Nutrient profi les and fl uxes, sediment reworking and transport
Individual bivalves, cores, ecosystems
Willows et al. (1998) ; Meysman et al. (2006a)
See Maire et al. (2008) for a review of methods for quantifying sediment reworking rates.

Bivalves as bioturbators and bioirrigators 303
variable and dependent on species, and the density and size structure of the population. The basic increased fl uxes of particulates and solutes that bioturbation induces are rather straightforward, but they have a range of complex and context - dependent effects. In the following sections these different mechanisms by which soft - sediment bivalves infl uence their surroundings are described. In particular the aim is to illustrate the numerous multiway interactions between bioturbation, benthic biodiversity, and ecosystem function.
Provide s tructure, h eterogeneity in s ediments, s tabilize/ d estabilize the s ediment
Bivalve beds physically infl uence the diversity and functioning of surrounding communities in a number of ways (see Chapter 9 in this book for an in - depth discussion on reef - forming bivalves as ecosystem engineers). Although infaunal species such as clams do not provide the same type of three - dimensional habitat as do mussels and oysters on hard substrates, the presence of bivalves neverthe-less provides increased structural heterogene-ity within the sediments, with potential for a larger number of different favorable micro-habitats in the interstitial spaces and thus increased diversity (Guti é rrez et al. 2003 ). Even the very small - scale movements around individual bioturbating bivalves are important and add heterogeneity to the soft - sediment system. Further, infaunal species have either siphons, feeding shafts, or parts of their shells reaching the sediment surface, which create additional bed roughness and heterogeneity. Bivalves protruding above the sediment surface will also affect fl ow in the benthic boundary layer, both due to their physical structure and to their feeding currents (Green et al. 1998 ). Shells provide hard substrate for recruitment in otherwise soft, unconsolidated sediments, which increases the diversity of epifaunal
How d o h ealthy s oft - s ediment b ivalve p opulations a ffect t heir s urroundings?
Bivalves have profound infl uences on sur-rounding macrobenthic communities. In com-parison with many other bioturbating species such as highly active crabs, shrimp, and poly-chaete worms, bivalves may seem rather sta-tionary and, at fi rst glance, not seem to be doing much, at least not in terms of bioturba-tion. Due to their frequently dominating abun-dances and biomasses, however, and active fi ltration of large volumes of water, the net impact of their bioturbation and bioirrigation activities on the system are considerable (Jackson et al. 2001 ; Vaughn and Hakenkamp 2001 ; Jaramillo et al. 2007 ; Petersen et al. 2008 ). Although most soft - sediment bivalves can be classifi ed as biodiffusers in terms of their bioturbation mode, different species have different burrowing depths (Table 10.3 ), which in turn infl uences their impact on sediment mixing, irrigation, and nutrient fl uxes, and their effects on the surrounding communities. Moreover, the juveniles of most soft - sediment bivalves live at or close to the sediment surface, and move successively deeper into the sedi-ment as they grow. Burrowing depth depends on siphon length (Zwarts and Wanink 1989 ); bivalves with short siphons live just under-neath the sediment surfaces (e.g., hard clams or northern quahog, Mercenaria mercenaria ), while species with longer siphons may be buried up to 40 cm down in the sediment (e.g., soft - shelled clams, Mya arenaria ) or even 80 cm (geoducks, Panopea abrupta ); however, shallow - burying bivalves (such as clams) tend to move more, thereby bioturbating the upper-most sediment layers more than the deep bur-rowers. Sediment reworking rates may further differ with season and food availability, even within a bivalve species (Stead and Thompson 2006 ; Maire et al. 2007 ). Thus, the patterns of bivalve bioturbation and facilitation effects on surrounding communities are temporally

304 Shellfi sh Aquaculture and the Environment
important and diversity no longer increases (Whitlatch et al. 1997 ).
Bivalve bioturbation has the potential to modify signifi cantly the coastal sea and land-scape. The prevalence of different types of soft - sediment habitats in coastal areas depends on the stability of the sediments, that is, the balance between rates of sediment deposition and sediment erosion. These processes are ulti-mately governed by hydrodynamic forcing (currents, waves) transporting the sediment
species in the system (sponges, bryozoans, tunicates, algae, etc.) (Guti é rrez et al. 2003 ). The bivalves affect the sedimentary habitat even after they die, as the shell debris contin-ues to provide structure in the habitat (Hewitt et al. 2005 ; Erwin 2008 ) with positive infl u-ences on biodiversity. The effect of bivalves on the physical heterogeneity of the habitat is naturally dependent on bivalve density and aggregation. For example, at very high bivalve densities, competition for space becomes
Table 10.3 Adult burrowing depth in natural populations of selected bivalve species and the type of shellfi sh aquaculture they are used in. In addition, their potential to actively bioturbate sediments in different types of culture is indicated with ( + ) or ( − ).
Species Burrowing depth Type of culture Potential for bioturbation in culture
Mercenaria mercenaria (northern quahog, hard clam)
1 – 3 cm (up to 20 cm) Bottom ( “ free ” ) +
On - bottom bags, cover nets
+
Mya arenaria (soft - shelled clam)
10 – 20 cm (up to 40 cm) Bottom +
Panopea abrupta (geoduck clam)
60 – 80 cm Bottom +
Crassostrea gigas (Pacifi c oyster)
0 Bottom −
Longlines, off - bottom bags and racks
−
Crassostrea virginica (eastern oyster)
0 Bottom −
Suspended −
Mytilus edulis (blue mussel) 0 Bottom −
Suspended −
Mytilus galloprovincialis (Mediterranean mussel)
0 Bottom −
Suspended −
Ruditapes philippinarum (Manila clam)
0 – 5 cm Bottom +
For many species, the burrowing depth differs between summer and winter (Zwarts and Wanink 1989 ). In cold climates, bivalves move deeper into the sediment in winter to avoid freezing, whereas in hot climates, bivalves may move deeper in summer to avoid overheating.

Bivalves as bioturbators and bioirrigators 305
philippinarum ), and Baltic clams ( Macoma balthica ) (Widdows and Brinsley 2002 ; Sgro et al. 2005 ; Ciutat et al. 2007 ). Although natural clam populations may act to destabilize sediments, enhance erosion rates, and prevent siltation, the nets used to keep clams in place in aquaculture installations increase the rates of sedimentation and increase the sediment silt content (Kaiser et al. 1996 ; Chapter 9 in this book). Problems with sedimentation can, however, be avoided with development of better culture methods, such as the bags used for clam culture in Cedar Key, Florida (http://shellfi sh.ifas.ufl .edu). These bags also provide habitat for other species and enhance diversity, provided the siting and layout of the bags is appropriate and coordinated. To predict sedi-ment transport patterns, the critical thresholds for sediment erosion need to be identifi ed (Le Hir et al. 2007 ). Faunal effects on sediment erosion, transport, and deposition have mainly been studied in intertidal areas, and our knowl-edge about these effects in deeper areas is much more limited.
The interactions between sediment stabiliz-ing and destabilizing processes are complex, dynamic, and continually at work. While bio-turbating bivalves may destabilize the sedi-ment, their bioturbation may simultaneously enhance nutrient fl uxes from the sediment, which promotes the growth of sediment stabi-lizing microphytobenthos (Sandwell et al. 2009 ). Within a bivalve bed the continuous reworking of the sediment, which is character-istic for biodiffusers such as most infaunal bivalves, homogenizes the sediment and pre-vents formation of permanent structures and may prevent establishment of, for example, invasive salt marsh species. Conversely, spread-ing salt marshes reduce the habitat available for shellfi sh (Neira et al. 2006 ). Sediment sta-bilizers and destabilizers have spatially and temporally variable effects on the community, usually favoring their own type, which facili-tates the persistence of patches (Volkenborn
and affecting its sorting and grain size, but biota signifi cantly infl uences the formation and persistence of different habitat types by affecting the stability of the sediment (Widdows et al. 2004 ; Le Hir et al. 2007 ).
In aquatic ecosystems some species act as sediment stabilizers (enhance sediment deposi-tion, inhibit erosion), while other species act as sediment destabilizers (enhance erosion and sediment transport) (Widdows et al. 2000 ; Bouma et al. 2009 ). By providing physical pro-tective structures, for example, mussel beds, oyster reefs, seagrass meadows, salt marsh plants, and tube - building polychaetes stabilize the sediment, modify the local hydrodynamics, and enhance sediment deposition. Also, micro-phytobenthos (i.e., microalgae/microorganisms living on the sediment surface) enhance sedi-ment cohesiveness through the extracellular polymeric substances they secrete, which form a protective biofi lm on the sediment surface.
Other species act as sediment destabilizers by disrupting the cohesiveness of the sediment, which decreases the sediment mud content and increases erosion rates. The sediment is desta-bilized, for example, by clams, crustaceans, and lugworms that bioturbate or mix the sedi-ment and facilitate resuspension (Widdows and Brinsley 2002 ). The sediment is also desta-bilized by grazers removing the protective microphytobenthic biofi lm. Further, increased bed roughness and scouring around patches of animals or plants can destabilize sediments and enhance erosion locally. Bioturbating bivalves affect resuspension rates, both directly and indirectly, actively and passively (Graf and Rosenberg 1997 ). Experiments in nonaquacul-ture settings have demonstrated a positive rela-tionship between bivalve density and sediment erosion, which is likely a combination of the bivalve - induced bioturbation and increased bed roughness, both of which promote sedi-ment erosion. These patterns have been found for a number of species, including cockles ( Cerastoderma edule ), Manila clams ( Ruditapes

306 Shellfi sh Aquaculture and the Environment
impact benthic ecosystems also during seed collection and harvesting. These other aspects, however, concern management practices and are dealt with in Chapters 3 and 11 in this book.
Bivalves have a dominating organizational role in ecosystems as they shunt a lot of the production from the pelagic to the benthic system in the form of biodeposits (Dame 1993 ; Newell 2004 ). Both feces and pseudofeces are ejected as mucus - bound aggregates, which sink faster than nonaggregated particles, thus aiding the vertical downward fl ux and increas-ing the deposition locally above background levels of sedimentation (Chapter 4 in this book; Ward and Shumway 2004 ). Rates of biodeposit production, and the ratio of feces to pseudofeces depends on the concentrations of food particles (phytoplankton, some detri-tus) and suspended sediment (inorganic matter), all of which vary seasonally, and also on the bivalve species, as different species have different clearance rates and abilities to select particles preferentially (Kautsky and Evans 1987 ; Jaramillo et al. 1992 ; Newell 2004 ; Ward and Shumway 2004 ; Giles et al. 2006 ). High loads of suspended sediments in the water (e.g., in sheltered, turbid estuaries) lead to larger amounts of pseudofeces being pro-duced, which also increases the downward fl ux of particulate matter to the sediment (Iglesias et al. 1996 ; Norkko et al. 2001 ).
Effects of w ater fl ow
Patterns of biodeposit accumulation or erosion in bivalve beds or underneath suspended bivalve cultures are mainly driven by the water fl ow at the site, with higher current velocities promoting erosion and effi cient dispersal of the biodeposits (Chamberlain et al. 2001 ; Hartstein and Rowden 2004 ; Weise et al. 2009 ; Chapters 4 , 6 , and 9 in this book). With greater tidal currents and wave action, resuspension of biodeposits is more likely in intertidal or shallow bivalve beds, compared
et al. 2009 ). These processes thus act both to modify and to maintain specifi c habitats. The antagonistic interactions between bioturbators and habitat - forming ecosystem engineers such as seagrasses, salt marshes, or oyster reefs create patchiness and habitat diversity, thereby favoring an overall higher biodiversity at the estuary scale (Jones et al. 1994 ; Bouma et al. 2009 ).
To minimize disruption of sediment trans-port patterns in an estuary and thus maintain the current extent of the existing habitat types in the estuary, the complex interactions between sediment stabilizers and destabilizers need to be considered. This has implications for where bivalve aquaculture plots should be located in an estuary and what the spatial scale, type of culture, and layout of the instal-lations should be, for example, continuous cover or several separate sections (Dumbauld et al. 2009 ).
Biodeposit p roduction and d egradation: e ffects on s ediment b iogeochemistry
Bivalves affect fl uxes of particles to and from the seafl oor, both by directly intercepting and ejecting particles as part of their feeding, and by physically altering the hydrodynamic conditions that govern sedimentation and erosion rates (Graf and Rosenberg 1997 ; Widdows et al. 1998 ) (Chapter 9 in this book). Through their feeding, bivalves produce feces and pseudofeces, collectively termed biodeposits when they reach the seafl oor. Chapter 4 (in this book) on “ Bivalves as bio-fi lters ” provides details on regulation of feeding and production of feces and pseudofe-ces. In this section the focus is on biodeposits after they have been deposited on the seafl oor and their context - dependent effects on benthic nutrient fl uxes and surrounding benthic habitats. In addition to the ongrowing phase considered here, bivalve aquaculture may

Bivalves as bioturbators and bioirrigators 307
subsidy to benthic systems (Wotton and Malmqvist 2001 ). As the biodeposits may be rich in carbon and nitrogen, surrounding deposit - feeding macro - , meio - , and micro-fauna can benefi t from increased food quality (Kautsky and Evans 1987 ; Norkko et al. 2001 ; Giles et al. 2006 ). The effects of increasing biodeposition follow the general hump - shaped benthic responses to organic enrichment, with increasing benthic diversity, abundance, and biomass at early stages of enrichment, and increasing sediment oxygen demand and anoxic conditions with benthic community disruption after severe enrichment (Pearson and Rosenberg 1978 ) (Fig. 10.3 ). The effects are directly related to the amounts of organic matter deposited and remaining on the sea-fl oor, and as noted earlier, these amounts are dependent on bivalve species and density, the quality and quantity of their food supply, and the hydrodynamic setting, water fl ow, and depth at the site (Fig. 10.3 ).
with bivalve beds or suspended aquaculture systems in deeper waters. In enclosed embay-ments, however, the tidal fl ushing may not be suffi cient and biodeposits accumulate under-neath shellfi sh farms (Cranford et al. 2009 ; Chapters 4 and 6 in this book) (Fig. 10.3 ). In deeper waters the biodeposits from suspended culture may be dispersed over a larger area before reaching the seafl oor and thus have no detectable impact on the benthic habitat. The rates of biodeposit accumulation or erosion are further affected by the density of biotur-bating infaunal bivalves and other fauna as they enhance resuspension and dispersal of biodeposits by acting as sediment destabilizers (see previous section).
Organic e nrichment
Bivalve biodeposition represents an organic enrichment of the sediment and this additional organic matter provides an important food
Figure 10.3 Schematic diagram of the context and density dependence of the effects of bivalve populations and their biodeposits on surrounding benthic communities. Panels A – C represent increasing densities of bivalves in either natural or bottom - cultured populations, while panel D represents a high - density suspended culture, where the bivalves are not able to bioturbate the sediments. Computational model by Alf Norkko.

308 Shellfi sh Aquaculture and the Environment
may also develop in areas of high biodeposi-tion and high sediment oxygen demand. Since Beggiatoa requires both oxygen and H 2 S for metabolism, its presence indicates that the reducing conditions in the sediment have reached the sediment surface.
Effects of b ioturbation on b enthic n utrient fl uxes and the p rocessing of b iodeposits
The biogeochemical effects of biodeposition from shellfi sh aquaculture are strongly depen-dent on the quantity and quality of the mate-rial deposited. Effi cient bivalve bioturbation does, however, ameliorate the potentially neg-ative effects of excessive biodeposition since particle and solute transport by bioturbating fauna signifi cantly infl uences rates and path-ways of organic matter mineralization, par-ticularly by increasing the oxygen penetration depth in the sediments. For example, Mya arenaria burrows are sites of enhanced micro-bial activity and high rates of sulphate reduc-tion (Hansen et al. 1996 ) and burrowing Mercenaria mercenaria increase benthic fl uxes of oxygen and nutrients (Doering et al. 1987 ). Bioturbation enhances coupled nitrifi cation - denitrifi cation, and under fully oxygenated conditions any remaining phosphorus in the biodeposits is also permanently buried with the accumulating sediments.
Infaunal bivalves both produce biodeposits and facilitate the remineralization of them in the sediment (Figs. 10.1 and 10.3 ). Thus, the combination of biodeposition and bioturba-tion by clams maintains a balanced benthic metabolism (Nizzoli et al. 2006 ), with the benthic nutrient fl uxes being dependent on bivalve density (Sandwell et al. 2009 ). In natural infaunal bivalve beds and bottom cul-tures of clams and mussels, rates of biodeposi-tion are not likely to reach high - enough levels to induce anoxic conditions and high levels of sulfi de in sediments due to the negative feed-back excessive biodeposition would have on
Effects of b iodeposition on s ediment b iogeochemistry
Biodeposits affect the biogeochemistry in the sediment and at the sediment – water interface. Although bivalves also directly excrete inor-ganic nutrients, the remineralization processes of the biodeposits in the sediment are usually more important for nutrient regeneration and fl uxes (Prins et al. 1998 ; Newell 2004 ). Accumulation of biodeposits increases sedi-ment organic content, promotes high micro-bial activity and increases the sediment oxygen demand (Rodhouse and Roden 1987 ; Wotton and Malmqvist 2001 ; Nizzoli et al. 2006 ; Hargrave et al. 2008 ; Cranford et al. 2009 ), but the effects of biodeposition are site specifi c (Chamberlain et al. 2001 ; Hartstein and Rowden 2004 ).
The decomposition of organic material con-sumes oxygen and increased sediment oxygen demand is commonly recorded, for example, under mussel culture longlines. High rates of biodeposition may adversely affect the sedi-ment microbial community by shifting it from an aerobic to an anaerobic metabolism. The changing oxygen conditions subsequently affect nutrient fl uxes in and out of the sedi-ment. Under anoxic conditions phosphorus is released from iron compounds in the sediment. Biodeposition also affects the cycling, reten-tion, and loss of nitrogen from the sediment (Cranford et al. 2007 ). Coupled nitrifi cation - denitrifi cation removes nitrogen from the system in the form of N 2 gas, but requires adjacent oxygenated sediments with nitrifying microbes and anoxic sediments with denitrify-ing microbes (Newell et al. 2002 ). Thus, the process is inhibited if the overlying water turns anoxic. Sediments underlying intensive shell-fi sh aquaculture release ammonium since the coupled nitrifi cation - denitrifi cation has been disrupted (Hatcher et al. 1994 ; Newell et al. 2002 ). If the sediments turn anoxic, concentra-tions of H 2 S may build up to toxic levels. White sulfur bacterial mats ( Beggiatoa spp.)

Bivalves as bioturbators and bioirrigators 309
and added physical structure from either bivalve shells or farming installations. The effects of organic enrichment from biodeposi-tion, however, have proven to be highly site specifi c and context dependent, and range from signifi cant positive effects (Inglis and Gust 2003 ; D ’ Amours et al. 2008 ), to no or minimal effects (Hatcher et al. 1994 ; Grant et al. 1995 ; Kaiser et al. 1996 ; Chamberlain et al. 2001 ; Crawford et al. 2003 ), to signifi -cant negative effects with impoverished benthic communities under/near the farms (Smith and Shackley 2004 ; Hargrave et al. 2008 ; Cranford et al. 2009 ). Inside bivalve beds and under lines of suspended bivalves, the benthic infau-nal species composition generally shifts toward species smaller, opportunistic detritivores, more tolerant to higher organic content, such as Oligochaeta and capitellid polychaetes (Beadman et al. 2004 ; Commito et al. 2005 ; Norling and Kautsky 2007 ; Ysebaert et al. 2009 ). Shells falling from mussel lines may also provide additional habitat and heteroge-neity, with positive effects on the surrounding fauna (Grant et al. 1995 ). At very high rates of biodeposit accumulation, however, the sedi-ment becomes anoxic and the macrofauna dis-appears. The net effects will depend both on the scale, stocking densities, and intensities of farming, as well as the hydrodynamic regime at the site (water depth, currents, sedimenta-tion rates), which will interact to determine the amount of waste products accumulating on the seabed (Fig. 10.3 ). It has, however, repeat-edly been reported that effects of, for example, mussel beds are relatively local and depend on bivalve density (Beadman et al. 2004 ; Hartstein and Rowden 2004 ; Ysebaert et al. 2009 ). Therefore, using a somewhat lower stocking density in combination with suffi cient fl ow can reduce the potential negative effects on the benthic environment (Beadman et al. 2004 ).
Since shellfi sh aquaculture is based on natural food sources, no additional nutrients or organic matter are added to the system. Therefore, the amount of biodeposition, and
bivalve densities. Nizzoli et al. (2006) com-pared mussel and clam farming and found that overall impacts of suspended mussel farming on oxygen and nutrient dynamics were greater than those of infaunal clam farming, and attributed this to the fact that infaunal, biotur-bating bivalves stimulate transfer of both organic matter and oxygen in to the sediment, whereas suspended bivalves only enhance transfer of organic matter to the sediment via biodeposition. This also affected the pathways of nitrogen cycling.
The feedback mechanisms between sedi-ment organic content, oxygen conditions, and bioturbation are complex. For example, hard clams do not burrow as deep under hypoxic conditions, with a potential for cascading negative effects on sediment oxygenation (Weissberger et al. 2009 ). It is therefore impor-tant to limit rates of biodeposit accumulation to a level at which the bioturbators can still process effectively. Model calculations indicate that even though the harvest represents a net removal of phosphorus and nitrogen from the ecosystem, the deposition of feces and pseu-dofeces from a mussel farm increases the reten-tion time of nutrients in some coastal areas (Cranford et al. 2007 ; Brigolin et al. 2009 ). With careful scaling and placement of farms in areas with suffi cient fl ow, and by maintain-ing suffi cient levels of bioturbation under the farms, mussel aquaculture can be used to remove nutrients from the system and thus combat eutrophication (Newell 2004 ; Lindahl et al. 2005 ; Gren et al. 2009 ; Chapter 8 in this book).
Effects of b iodeposits on b enthic c ommunity c omposition
Shellfi sh aquaculture infl uences the surround-ing benthic habitat through three main mecha-nisms: increased input of organic matter through biodeposition; increasing hypoxic and sulfi dic sediments due to increasing oxygen demand from decomposition of biodeposits;

310 Shellfi sh Aquaculture and the Environment
Heck, 2001b ). Bioturbating bivalves inside seagrass meadows also renew the water at the sediment surface and thereby reduce the risk of oxygen depletion. Nevertheless, particularly in eutrophic areas, high levels of biodeposition may lead to toxic sulfi de concentrations in the sediment and increase the ammonium effl ux from the sediment, which facilitates epiphyte growth, resulting in impaired growth of sea-grass (Vinther and Holmer 2008 ; Vinther et al. 2008 ). Also, rack and stake oyster culture negatively infl uence seagrasses through physi-cal disturbance and shading (Everett et al. 1995 ). If densities of aquaculture gear are suf-fi ciently low, however, there are only minimal effects on seagrasses of, for example, oyster depuration gear (Vaudrey et al. 2009 ). Therefore, the multiple interactions between seagrass habitats and shellfi sh aquaculture need to be considered and managed at appro-priate scales (Dumbauld et al. 2009 ).
Effects of b ioturbation on c ontaminants and r esting s tages in the s ediments
Bivalve bioturbation also affects the mixing and depth distribution of contaminants and has the potential to either bury these elements deeper or to bring them back into circulation. For example, Ciutat et al. (2006) found a posi-tive correlation between the density of cockles ( Cerastoderma edule ) and the resuspension of both sediment and polycyclic aromatic hydro-carbon s ( PAH s). Similarly, bioturbation by the bivalve Tellina deltoidalis in metal contami-nated sediments caused metal release from the pore waters and higher concentrations of iron and manganese in overlying waters (Atkinson et al. 2007 ). On the other hand, bioturbation by Baltic clams increased the retention of cadmium and a hydrophobic organic pollutant in the sediment (Hedman et al. 2008 ). Thus, the fate of contaminant is dependent both on
thus organic enrichment and potential subse-quent negative effects, is signifi cantly less under shellfi sh aquaculture installations com-pared with fi nfi sh aquaculture (Folke and Kautsky 1989 ; Naylor et al. 2000 ; Newell 2004 ). In fi nfi sh farming stocking densities are higher and the fi sh are fed artifi cial diets, resulting in a net addition of nutrients and organic matter to the site, which may result in waste accumulation and anoxic bottoms under the fi sh cages, disturbed benthic communities, and eutrophication of the surrounding ecosys-tem (Gowen and Bradbury 1987 ; Wu 1995 ; Kalantzi and Karakassis 2006 ). Due to this fundamental difference, it is important and essential to consider fi nfi sh and shellfi sh aqua-culture as two separate industries.
Interactions between b ivalve b ioturbation and s eagrasses
In coastal areas, seagrass meadows are impor-tant structured habitats, providing nursery areas for fi sh and shellfi sh (Heck et al. 2003 ). Suspension - feeding bivalves reduce water tur-bidity, which enhances light penetration and thereby potentially promotes benthic primary producers, such as seagrasses and microphyto-benthos. For example, successful recruitment of Mya arenaria was instrumental in turning a Danish lagoon from a turbid state to a clear - water state (Petersen et al. 2008 ). In some areas, improved water clarity resulting from large beds of cultured bivalves has even led to the reestablishment of seagrasses (Shumway and Kraeuter 2004 ). The effects are, however, species dependent and, for example, eastern oysters ( Crassostrea virginica ) have higher clearance rates than hard clams and therefore promote seagrass production more (Newell and Koch 2004 ). Sediment nutrient enrich-ment by biodeposits from natural bivalve pop-ulations can also elevate seagrass productivity (Peterson and Heck 2001a ; Peterson and

Bivalves as bioturbators and bioirrigators 311
organic content, hydrodynamic conditions, and environmental stress. Nevertheless, we are lacking in our understanding of the generality and ecosystem - scale consequences of these fl uxes. Since the positive or negative effects of bivalve biodeposits are quality, quantity, and context dependent, and modifi ed by bivalve bioturbation, studies of the effects of organic enrichment from shellfi sh aquaculture on sedi-ment biogeochemistry, benthic communities, and seagrass beds have yielded site - specifi c and sometimes nonconclusive results. The associ-ated complex biogeochemical processes are diffi cult to measure over larger scales in the fi eld, but at the same time results gained from laboratory experiments are diffi cult to general-ize to, for example, sites with different hydro-dynamic regimes (Porter et al. 2004 ). Therefore, an integrated approach consisting of well - designed laboratory experiments, fi eld studies across different sites and along gradients within sites, and modeling is required to understand these processes and to apply them at the appropriate scale to sustainable manage-ment of shellfi sh aquaculture (Dumbauld et al. 2009 ).
The benthic effects of bivalve aquaculture differ signifi cantly between cultured species and also between different types of culture of the same species (Nizzoli et al. 2006 ; Ysebaert et al. 2009 ). Suspended culture generally only affects benthic environments through their biodeposition, while on - bottom culture affects benthic habitats through both biodeposition, altered physical structure at the sediment surface, and bioturbation of the sediments. Therefore, for example, clam farming results in a more balanced benthic metabolism, although it may be limited by available space. As the effects of biodeposition on the benthic environment are relatively local and dependent on the hydrodynamic regime, in order to mini-mize negative impacts on the benthic commu-nities, farms should be sited in areas with relatively high current velocities and suffi cient
the type of compound and the rates of biotur-bation in the habitat.
Many planktonic species form dormant cysts or resting stages under adverse environ-mental conditions. The bioturbation - induced mixing processes apply to these cysts as well and the burial and reemergence of these is part of the natural cycle for many species. For example, Baltic clams extend the distribution of cladoceran resting eggs deeper into the sedi-ment (Viitasalo 2007 ) and also differentially affect the emergence of several zooplankton species from the sediment (Viitasalo et al. 2007 ).
Summary
Bioturbation and bioirrigation are very small - scale activities by individual bivalves, but the net effects of a whole bivalve population may have consequences for the long - term develop-ment of an entire coastal system, both in terms of nutrient dynamics and sediment stability. This highlights the importance of considering the appropriate spatial and temporal scales for both studying and managing any particular process, installation, or environmental effect, including shellfi sh aquaculture operations.
Bivalve bioturbation facilitates transport of particulate and dissolved material between the sediment and the overlying water, oxygenates the sediment, and produces complex and favorable microhabitats, which facilitate benthic communities with higher overall diver-sity. It is commonly assumed that there is a general positive relationship between high diversity and well - functioning, healthy ecosys-tems. One way to assess ecosystem function is to estimate rates of nutrient fl uxes in and out of the sediments (Lohrer et al. 2004 ). There are numerous studies on nutrient fl uxes and bivalves, and we know that they are infl u-enced, for example, by the interactions between bivalve density and species, sediment type,

312 Shellfi sh Aquaculture and the Environment
Journal of Experimental Marine Biology and Ecology 371 : 20 – 33 .
Bouma , T.J. , Olenin , S. , Reise , K. , and Ysebaert , T. 2009 . Ecosystem engineering and biodiversity in coastal sediments: posing hypotheses . Helgoland Marine Research 63 : 95 – 106 .
Brigolin , D. , Dal Maschio , G. , Rampazzo , F. , Giani , M. , and Pastres , R. 2009 . An individual - based population dynamic model for estimating biomass yield and nutrient fl uxes through an off - shore mussel ( Mytilus galloprovincialis ) farm . Estuarine, Coastal and Shelf Science 82 : 365 – 376 .
Chamberlain , J. , Fernandes , T.F. , Read , P. , Nickell , T.D. , and Davies , I.M. 2001 . Impacts of biode-posits from suspended mussel ( Mytilus edulis L.) culture on the surrounding surfi cial sediments . ICES Journal of Marine Science 58 : 411 – 416 .
Ciutat , A. , Widdows , J. , and Readman , J.W. 2006 . Infl uence of cockle Cerastoderma edule biotur-bation and tidal - current cycles on resuspension of sediment and polycyclic aromatic hydrocar-bons . Marine Ecology Progress Series 328 : 51 – 64 .
Ciutat , A. , Widdows , J. , and Pope , N.D. 2007 . Effect of Cerastoderma edule density on near - bed hydrodynamics and stability of cohesive muddy sediments . Journal of Experimental Marine Biology and Ecology 346 : 114 – 126 .
Commito , J.A. , Celano , E.A. , Celico , H.J. , Como , S. , and Johnson , C.P. 2005 . Mussels matter: postlarval dispersal dynamics altered by a spa-tially complex ecosystem engineer . Journal of Experimental Marine Biology and Ecology 316 : 133 – 147 .
Crawford , C.M. , Macleod , C.K.A. , and Mitchell , I.M. 2003 . Effects of shellfi sh farming on the benthic environment . Aquaculture 224 : 117 – 140 .
Cranford , P.J. , Strain , P.M. , Dowd , M. , Hargrave , B.T. , Grant , J. , and Archambault , M. - C. 2007 . Infl uence of mussel aquaculture on nitrogen dynamics in a nutrient enriched coastal embay-ment . Marine Ecology Progress Series 347 : 61 – 78 .
Cranford , P.J. , Hargrave , B.T. , and Doucette , L.I. 2009 . Benthic organic enrichment from sus-pended mussel ( Mytilus edulis ) culture in Prince Edward Island, Canada . Aquaculture 292 : 189 – 196 .
water exchange (Hartstein and Rowden 2004 ; Dumbauld et al. 2009 ) since in enclosed embayments the dispersive capacity may be surpassed (Cranford et al. 2009 ).
Finally, homogenization of habitats removes the small - scale heterogeneity that sustains diversity, and loss of biodiversity will ulti-mately lower the ocean ’ s capacity to provide food for human consumption (Worm et al. 2006 ). Thus, shellfi sh aquaculture should be managed in the light of minimizing negative impacts on diversity.
Literature cited
Aller , R.C. 1988 . Benthic fauna and biogeochemical processes in marine sediments: the role of burrow structures . In: Blackburn , T.H. , and S ø rensen , J. (eds.), Nitrogen Cycling in Coastal Marine Environments . Wiley , New York , pp. 301 – 338 .
Aller , R.C. 2001 . Transport and reactions in the bioirrigated zone . In: Boudreau , B. , and J ø rgensen , B.B. (eds.), The Benthic Boundary Layer: Transport Processes and Biogeochemistry . Oxford University Press , Oxford , pp. 269 – 301 .
Asmus , R.M. , Jensen , M.H. , Jensen , K.M. , Kristensen , E. , Asmus , H. , and Wille , A. 1998 . The role of water movement and spatial scaling for measurement of dissolved inorganic nitrogen fl uxes in intertidal sediments . Estuarine, Coastal and Shelf Science 46 : 221 – 232 .
Atkinson , C.A. , Jolley , D.F. , and Simpson , S.L. 2007 . Effect of overlying water pH, dissolved oxygen, salinity and sediment disturbances on metal release and sequestration from metal con-taminated marine sediments . Chemosphere 69 : 1428 – 1437 .
Beadman , H.A. , Kaiser , M.J. , Galanidi , M. , Shucksmith , R. , and Willows , R.I. 2004 . Changes in species richness with stocking density of marine bivalves . Journal of Applied Ecology 41 : 464 – 475 .
Bouchet , V.M.P. , Sauriau , P. - G. , Debenay , J. - P. , Mermillod - Blondin , F. , Schmidt , S. , Amiard , J. - C. , and Dupas , B. 2009 . Infl uence of the mode of macrofauna - mediated bioturbation on the vertical distribution of living benthic foramin-ifera: fi rst insight from axial tomodensitometry .

Bivalves as bioturbators and bioirrigators 313
Dahlhoff , E.P. , Stillman , J.H. , and Menge , B.A. 2002 . Physiological community ecology: varia-tion in metabolic activity of ecologically impor-tant rocky intertidal invertebrates along environmental gradients . Integrative and Comparative Biology 42 : 862 – 871 .
Dame , R.F. 1993 . Bivalve Filter Feeders and Coastal and Estuarine Ecosystem Processes . Springer Verlag , Heidelberg .
Dame , R. , Bushek , D. , Allen , D. , Lewitus , A. , Edwards , D. , Koepfl er , E. , and Gregory , L. 2002 . Ecosystem response to bivalve density reduction: management implications . Aquatic Ecology 36 : 51 – 65 .
D ’ Amours , O. , Archambault , P. , McKindsey , C.W. , and Johnson , L.E. 2008 . Local enhancement of epibenthic macrofauna by aquaculture activities . Marine Ecology Progress Series 371 : 73 – 84 .
Doering , P.H. , Kelly , J.R. , Oviatt , C.A. , and Sowers , T. 1987 . Effect of the hard clam Mercenaria mercenaria on benthic fl uxes of inorganic nutri-ents and gases . Marine Biology 94 : 377 – 383 .
Dumbauld , B.R. , Ruesink , J.L. , and Rumrill , S.S. 2009 . The ecological role of bivalve shellfi sh aquaculture in the estuarine environment: a review with application to oyster and clam culture in West Coast (USA) estuaries . Aquaculture 290 : 196 – 223 .
Erwin , D.H. 2008 . Macroevolution of ecosystem engineering, niche construction and diversity . Trends in Ecology and Evolution 23 : 304 – 310 .
Everett , R.A. , Ruiz , G.M. , and Carlton , J.T. 1995 . Effect of oyster mariculture on submerged aquatic vegetation: an experimental test in a Pacifi c Northwest estuary . Marine Ecology Progress Series 125 : 205 – 217 .
Folke , C. , and Kautsky , N. 1989 . The role of eco-systems for a sustainable development of aqua-culture . Ambio 18 : 234 – 243 .
Fran ç ois , F. , Gerino , M. , Stora , G. , Durbec , J. - P. , and Poggiale , J. - C. 2002 . Functional approach to sediment reworking by gallery - forming macrobenthic organisms: modeling and applica-tion with the polychaete Nereis diversicolor . Marine Ecology Progress Series 229 : 127 – 136 .
Gerino , M. , Aller , R.C. , Lee , C. , Cochran , J.K. , Aller , J.Y. , Green , M.A. , and Hirschberg , D. 1998 . Comparison of different tracers and
methods used to quantify bioturbation during a spring bloom: 234 - thorium, luminophores and chlorophyll a . Estuarine, Coastal and Shelf Science 46 : 531 – 547 .
Gerino , M. , Stora , G. , Fran ç ois , F. , Gilbert , J.C. , Poggiale , J. - C. , Mermillod - Blondin , F. , Desrosiers , G. , and Vervier , P. 2003 . Macro - invertebrate functional groups in freshwater and marine sediments: a common mechanistic clas-sifi cation . Vie Milieu 53 : 222 – 231 .
Giles , H. , Pilditch , C.A. , and Bell , D.G. 2006 . Sedimentation from mussel ( Perna canaliculus ) culture in the Firth of Thames, New Zealand: impacts on sediment oxygen and nutrient fl uxes . Aquaculture 261 : 125 – 140 .
Glud , R.N. 2008 . Oxygen dynamics of marine sedi-ments . Marine Biology Research 4 : 243 – 289 .
Gowen , R.J. , and Bradbury , N.B. 1987 . The eco-logical impact of salmonid farming in coastal waters: a review . Oceanography and Marine Biology: An Annual Review 25 : 563 – 575 .
Graf , G. , and Rosenberg , R. 1997 . Bioresuspension and biodeposition: a review . Journal of Marine Systems 11 : 269 – 278 .
Grant , J. , Hatcher , A. , Scott , D.B. , Pocklington , P. , Schafer , C.T. , and Winters , G.V. 1995 . A multi-disciplinary approach to evaluating impacts of shellfi sh aquaculture on benthic communities . Estuaries 18 : 124 – 144 .
Green , M.O. , Hewitt , J.E. , and Thrush , S.F. 1998 . Seabed drag coeffi cient over natural beds of horse mussels ( Atrina zelandica ) . Journal of Marine Research 56 : 613 – 637 .
Gren , I. - M. , Lindahl , O. , and Lindqvist , M. 2009 . Values of mussel farming for combating eutro-phication: an application to the Baltic Sea . Ecological Engineering 35 : 935 – 945 .
Grizzle , R.E. , and Penniman , C.A. 1991 . Effects of organic enrichment on estuarine macrofaunal benthos: a comparison of sediment profi le imaging and traditional methods . Marine Ecology Progress Series 74 : 249 – 262 .
Guti é rrez , J.L. , Jones , C.G. , Strayer , D.L. , and Iribarne , O.O. 2003 . Molluscs as ecosystem engineers: the role of shell production in aquatic habitats . Oikos 101 : 79 – 90 .
Hansen , K. , King , G.M. , and Kristensen , E. 1996 . Impact of the soft - shell clam Mya arenaria on sulfate reduction in an intertidal sediment . Aquatic Microbial Ecology 10 : 181 – 194 .

314 Shellfi sh Aquaculture and the Environment
Hargrave , B.T. , Doucette , L.I. , Cranford , P.J. , Law , B.A. , and Milligan , T.G. 2008 . Infl uence of mussel aquaculture on sediment organic enrich-ment in a nutrient - rich coastal embayment . Marine Ecology Progress Series 365 : 137 – 149 .
Hartstein , N.D. , and Rowden , A.A. 2004 . Effect of biodeposits from mussel culture on macroinver-tebrate assemblages at sites of different hydro-dynamic regime . Marine Environmental Research 57 : 339 – 357 .
Hatcher , A. , Grant , J. , and Schofi eld , B. 1994 . Effects of suspended mussel culture ( Mytilus spp.) on sedimentation, benthic respiration and sediment nutrient dynamics in a coastal bay . Marine Ecology Progress Series 115 : 219 – 235 .
Heck , K.L. , Jr. , Hays , G. , and Orth , R.J. 2003 . Critical evaluation of the nursery role hypothesis for seagrass meadows . Marine Ecology Progress Series 253 : 123 – 136 .
Hedman , J.E. , Bradshaw , C. , Thorsson , M.H. , Gilek , M. , and Gunnarsson , J.S. 2008 . Fate of contaminants in Baltic Sea sediments: role of bioturbation and settling organic matter . Marine Ecology Progress Series 356 : 25 – 38 .
Hewitt , J.E. , Thrush , S.F. , Halliday , J. , and Duffy , C. 2005 . The importance of small - scale habitat structure for maintaining beta diversity . Ecology 86 : 1619 – 1626 .
Hewitt , J.E. , Thrush , S.F. , Dayton , P.K. , and Bonsdorff , E. 2007 . The effect of spatial and temporal heterogeneity on the design and analy-sis of empirical studies of scale - dependent systems . The American Naturalist 169 : 398 – 408 .
Iglesias , J.I.P. , Urrutia , M.B. , Navarro , E. , Alvarez - Jorna , P. , Larretxea , X. , Bougrier , S. , and Heral , M. 1996 . Variability of feeding processes in the cockle Cerastoderma edule (L.) in response to changes in seston concentration and composi-tion . Journal of Experimental Marine Biology and Ecology 197 : 121 – 143 .
Inglis , G.J. , and Gust , N. 2003 . Potential indirect effects of shellfi sh culture on the reproductive success of benthic predators . Journal of Applied Ecology 40 : 1077 – 1089 .
Jackson , J.B.C. , Kirby , M.X. , Berger , W.H. , Bjorndal , K.A. , Botsford , L.W. , Bourque , B.J. , Bradbury , R.H. , Cooke , R. , Erlandson , J. , Estes , J.A. , Hughes , T.P. , Kidwell , S. , Lange , C.B. ,
Lenihan , H.S. , Pandolfi , J.M. , Peterson , C.H. , Steneck , R.S. , Tegner , M.J. , and Warner , R.R. 2001 . Historical overfi shing and the recent col-lapse of coastal ecosystems . Science 293 : 629 – 638 .
Jaramillo , E. , Bertr á n , C. , and Bravo , A. 1992 . Mussel biodeposition in an estuary in southern Chile . Marine Ecology Progress Series 82 : 85 – 94 .
Jaramillo , E. , Contreras , H. , and Duarte , C. 2007 . Community structure of the macroinfauna inhabiting tidal fl ats characterized by the pres-ence of different species of burrowing bivalves in Southern Chile . Hydrobiologia 580 : 85 – 96 .
Jones , C.G. , Lawton , J.H. , and Shachak , M. 1994 . Organisms as ecosystem engineers . Oikos 69 : 373 – 386 .
Kaiser , M.J. , Edwards , D.B. , and Spencer , B.E. 1996 . Infaunal community changes as a result of commercial clam cultivation and harvesting . Aquatic Living Resources 9 : 57 – 63 .
Kalantzi , I. , and Karakassis , I. 2006 . Benthic impacts of fi sh farming: meta - analysis of com-munity and geochemical data . Marine Pollution Bulletin 52 : 484 – 493 .
Karlson , K. , Rosenberg , R. , and Bonsdorff , E. 2002 . Temporal and spatial large - scale effects of eutro-phication and oxygen defi ciency on benthic fauna in Scandinavian and Baltic waters — a review . Oceanography and Marine Biology: An Annual Review 40 : 427 – 489 .
Kautsky , N. , and Evans , S. 1987 . Role of biodeposi-tion by Mytilus edulis in the circulation of matter and nutrients in a Baltic coastal ecosystem . Marine Ecology Progress Series 38 : 201 – 212 .
Kristensen , E. 1988 . Benthic fauna and biogeo-chemical processes in marine sediments: micro-bial activities and fl uxes . In: Blackburn , T.H. , and S ø rensen , J. (eds.), Nitrogen Cycling in Coastal Marine Environments . Wiley , New York , pp. 275 – 299 .
Le Hir , P. , Monbet , Y. , and Orvain , F. 2007 . Sediment erodability in sediment transport mod-elling: can we account for biota effects? Continental Shelf Research 27 : 1116 – 1142 .
Levinton , J. 1995 . Bioturbators as ecosystem engi-neers: control of the sediment fabric, inter - indi-vidual interactions, and material fl uxes . In: Jones , C.G. , and Lawton , J.H. (eds.), Linking

Bivalves as bioturbators and bioirrigators 315
Species and Ecosystems . Chapman & Hall , New York , pp. 29 – 38 .
Lindahl , O. , Hart , R. , Hernroth , B. , Kollberg , S. , Loo , L. - O. , Olrog , L. , Rehnstam - Holm , A. - S. , Svensson , J. , Svensson , S. , and Syversen , U. 2005 . Improving marine water quality by mussel farming: a profi table solution for Swedish society . Ambio 34 : 131 – 138 .
Lohrer , A.M. , Thrush , S.F. , and Gibbs , M.M. 2004 . Bioturbators enhance ecosystem function through complex biogeochemical interactions . Nature 431 : 1092 – 1095 .
Maire , O. , Duch ê ne , J.C. , Gr é mare , A. , Malyuga , V.S. , and Meysman , F.J.R. 2007 . A comparison of sediment reworking rates by the surface deposit - feeding bivalve Abra ovata during sum-mertime and wintertime, with a comparison between two models of sediment reworking . Journal of Experimental Marine Biology and Ecology 343 : 21 – 36 .
Maire , O. , Lecroart , P. , Meysman , F. , Rosenberg , R. , Duch ê ne , J. - C. , and Gr é mare , A. 2008 . Quantifi cation of sediment reworking rates in bioturbation research: a review . Aquatic Biology 2 : 219 – 238 .
Martin , W.R. , and Banta , G.T. 1992 . The measure-ment of sediment irrigation rates: a comparison of the BR − tracer and 222 RN/ 226 RA disequilibrium techniques . Journal of Marine Research 50 : 125 – 154 .
Meysman , F.J.R. , Galaktionov , O.S. , Gribsholt , B. , and Middelburg , J.J. 2006a . Bio - irrigation in permeable sediments: an assessment of model complexity . Journal of Marine Research 64 : 589 – 627 .
Meysman , F.J.R. , Middelburg , J.J. , and Heip , C.H.R. 2006b . Bioturbation: a fresh look at Darwin ’ s last idea . Trends in Ecology and Evolution 21 : 688 – 695 .
Michaud , E. , Desrosiers , G. , Mermillod - Blondin , F. , Sundby , B. , and Stora , G. 2006 . The functional group approach to bioturbation: II. The effects of the Macoma balthica community on fl uxes of nutrients and dissolved organic carbon across the sediment – water interface . Journal of Experimental Marine Biology and Ecology 337 : 178 – 189 .
Montserrat , F. , Van Colen , C. , Provoost , P. , Milla , M. , Ponti , M. , Van den Meersche , K. , Ysebaert , T. , and Herman , P.M.J. 2009 . Sediment segrega-
tion by biodiffusing bivalves . Estuarine, Coastal and Shelf Science 83 : 379 – 391 .
Naylor , R.L. , Goldburg , R.J. , Primavera , J.H. , Kautsky , N. , Beveridge , M.C.M. , Clay , J. , Folke , C. , Lubchenco , J. , Mooney , H. , and Troell , M. 2000 . Effect of aquaculture on world fi sh sup-plies . Nature 405 : 1017 – 1024 .
Neira , C. , Grosholz , E.D. , Levin , L.A. , and Blake , R. 2006 . Mechanisms generating modifi cation of benthos following tidal fl at invasion by a Spartina hybrid . Ecological Applications 16 : 1391 – 1404 .
Newell , R.I.E. 2004 . Ecosystem infl uences of natural and cultivated populations of suspen-sion - feeding bivalve molluscs: a review . Journal of Shellfi sh Research 23 : 51 – 61 .
Newell , R.I.E. , Cornwell , J.C. , and Owens , M.S. 2002 . Infl uence of simulated bivalve biodeposi-tion and microphytobenthos on sediment nitro-gen dynamics: a laboratory study . Limnology and Oceanography 47 : 1367 – 1379 .
Newell , R.I.E. , and Koch , E.W. 2004 . Modeling seagrass density and distribution in response to changes in turbidity stemming from bivalve fi l-tration and seagrass sediment stabilization . Estuaries 27 : 793 – 806 .
Nizzoli , D. , Welsh , D.T. , Fano , E.A. , and Viaroli , P. 2006 . Impact of clam and mussel farming on benthic metabolism and nitrogen cycling, with emphasis on nitrate reduction pathways . Marine Ecology Progress Series 315 : 151 – 165 .
Norkko , A. , Hewitt , J.E. , Thrush , S.F. , and Funnell , G.A. 2001 . Benthic - pelagic coupling and suspen-sion - feeding bivalves: linking site - specifi c sedi-ment fl ux and biodeposition to benthic community structure . Limnology and Oceanography 46 : 2067 – 2072 .
Norkko , A. , Hewitt , J.E. , Thrush , S.F. , and Funnell , G.A. 2006 . Conditional outcomes of facilitation by a habitat - modifying subtidal bivalve . Ecology 87 : 226 – 234 .
Norling , P. , and Kautsky , N. 2007 . Structural and functional effects of Mytilus edulis on diversity of associated species and ecosystem functioning . Marine Ecology Progress Series 351 : 163 – 175 .
Pearson , T.H. , and Rosenberg , R. 1978 . Macrobenthic succession in relation to organic enrichment and pollution of the marine environ-ment . Oceanography and Marine Biology: An Annual Review 16 : 229 – 311 .

316 Shellfi sh Aquaculture and the Environment
Perez , K.T. , Davey , E.W. , Moore , R.H. , Burn , P.R. , Rosol , M.S. , Cardin , J.A. , Johnson , R.L. , and Kopans , D.N. 1999 . Application of computer - aided tomography (CT) to the study of estuarine benthic communities . Ecological Applications 9 : 1050 – 1058 .
Petersen , J.K. , Hansen , J.W. , Laursen , M.B. , Clausen , P. , Carstensen , J. , and Conley , D.J. 2008 . Regime shift in a coastal marine ecosys-tem . Ecological Applications 18 : 497 – 510 .
Peterson , B.J. , and Heck , K.L. , Jr . 2001a . An experi-mental test of the mechanism by which suspen-sion feeding bivalves elevate seagrass productivity . Marine Ecology Progress Series 218 : 115 – 125 .
Peterson , B.J. , and Heck , K.L. , Jr . 2001b . Positive interactions between suspension - feeding bivalves and seagrass — a facultative mutualism . Marine Ecology Progress Series 213 : 143 – 155 .
Porter , E.T. , Cornwell , J.C. , Sanford , L.P. , and Newell , R.I.E. 2004 . Effect of oysters Crassostrea virginica and bottom shear velocity on benthic – pelagic coupling and estuarine water quality . Marine Ecology Progress Series 271 : 61 – 75 .
Prins , T.C. , Smaal , A.C. , and Dame , R.F. 1998 . A review of the feedbacks between bivalve grazing and ecosystem processes . Aquatic Ecology 31 : 349 – 359 .
Quintana , C.O. , Tang , M. , and Kristensen , E. 2007 . Simultaneous study of particle reworking, irriga-tion transport and reaction rates in sediment bioturbated by the polychaetes Heteromastus and Marenzelleria . Journal of Experimental Marine Biology and Ecology 352 : 392 – 406 .
Reise , K. 2002 . Sediment mediated species interac-tions in coastal waters . Journal of Sea Research 48 : 127 – 141 .
Rhoads , D.C. 1974 . Organism - sediment relations on the muddy sea fl oor . Oceanography and Marine Biology: An Annual Review 12 : 223 – 300 .
Rhoads , D.C. , and Young , D.K. 1971 . Animal - sediment relations in Cape Cod Bay, Massachusetts II. Reworking by Molpadia oolit-ica (Holothuroidea) . Marine Biology 11 : 255 – 261 .
Rodhouse , P.G. , and Roden , C.M. 1987 . Carbon budget for a coastal inlet in relation to intensive cultivation of suspension - feeding bivalve mol-luscs . Marine Ecology Progress Series 36 : 225 – 236 .
Sandwell , D.R. , Pilditch , C.A. , and Lohrer , A.M. 2009 . Density dependent effects of an infaunal suspension - feeding bivalve ( Austrovenus stutchburyi ) on sandfl at nutrient fl uxes and microphytobenthic productivity . Journal of Experimental Marine Biology and Ecology 373 : 16 – 25 .
Schaffner , L.C. , Jonsson , P. , Diaz , R.J. , Rosenberg , R. , and Gapcynski , P. 1992 . Benthic communi-ties and bioturbation history of estuarine and coastal systems: effects of hypoxia and anoxia . In: Vollenweider , R.A. , Marchetti , R. , and Viviani , R. (eds.), Marine Coastal Eutrophication . Elsevier , Amsterdam , pp. 1001 – 1016 .
Sgro , L. , Mistri , M. , and Widdows , J. 2005 . Impact of infaunal Manila clam, Ruditapes philippina-rum , on sediment stability . Hydrobiologia 550 : 175 – 182 .
Shumway , S.E. , and Kraeuter , J.N. (eds.). 2004 . Molluscan shellfi sh research and management: charting a course for the future. Final Proceedings from the Cooperative Research and Information Institute (CRII) Workshop, Charleston, SC, January 2000.
Smith , J. , and Shackley , S.E. 2004 . Effects of a com-mercial mussel Mytilus edulis lay on a sublitto-ral, soft sediment benthic community . Marine Ecology Progress Series 282 : 185 – 191 .
Solan , M. , Wigham , B.D. , Hudson , I.R. , Kennedy , R. , Coulon , C.H. , Norling , K. , Nilsson , H.C. , and Rosenberg , R. 2004 . In situ quantifi cation of bioturbation using time - lapse fl uorescent sedi-ment profi le imaging (f - SPI), luminophore tracers and model simulation . Marine Ecology Progress Series 271 : 1 – 12 .
Stead , R.A. , and Thompson , R.J. 2006 . The infl u-ence of an intermittent food supply on the feeding behaviour of Yoldia hyperborea (Bivalvia: Nuculanidae) . Journal of Experi-mental Marine Biology and Ecology 332 : 37 – 48 .
Vaudrey , J.M.P. , Getchis , T. , Shaw , K. , Markow , J. , Britton , R. , and Kremer , J.N. 2009 . Effects of oyster depuration gear on eelgrass (Zostera marina L.) in a low density aquaculture site in Long Island Sound . Journal of Shellfi sh Research 28 : 243 – 250 .
Vaughn , C.C. , and Hakenkamp , C.C. 2001 . The functional role of burrowing bivalves in fresh-water ecosystems . Freshwater Biology 46 : 1431 – 1446 .

Bivalves as bioturbators and bioirrigators 317
Viitasalo , S. 2007 . Effects of bioturbation by three macrozoobenthic species and predation by necto - benthic mysids on cladoceran benthic eggs . Marine Ecology Progress Series 336 : 131 – 140 .
Viitasalo , S. , Katajisto , T. , and Viitasalo , M. 2007 . Bioturbation changes the patterns of benthic emergence in zooplankton . Limnology and Oceanography 52 : 2325 – 2339 .
Vinther , H.F. , and Holmer , M. 2008 . Experimental test of biodeposition and ammonium excretion from blue mussels ( Mytilus edulis ) on eelgrass ( Zostera marina ) performance . Journal of Experimental Marine Biology and Ecology 364 : 72 – 79 .
Vinther , H.F. , Laursen , J.S. , and Holmer , M. 2008 . Negative effects of blue mussel (Mytilus edulis) presence in eelgrass ( Zostera marina ) beds in Flensborg fjord, Denmark . Estuarine, Coastal and Shelf Science 77 : 91 – 103 .
Volkenborn , N. , Polerecky , L. , Hedtkamp , S.I.C. , van Beusekom , J.E.E. , and de Beer , D. 2007 . Bioturbation and bioirrigation extend the open exchange regions in permeable sediments . Limnology and Oceanography 52 : 1898 – 1909 .
Volkenborn , N. , Robertson , D.M. , and Reise , K. 2009 . Sediment destabilizing and stabilizing bio - engineers on tidal fl ats: cascading effects of experimental exclusion . Helgoland Marine Research 63 : 27 – 35 .
Ward , J.E. , and Shumway , S.E. 2004 . Separating the grain from the chaff: particle selection in suspen-sion - and deposit - feeding bivalves . Journal of Experimental Marine Biology and Ecology 300 : 83 – 130 .
Weise , A.M. , Cromey , C.J. , Callier , M.D. , Archambault , P. , Chamberlain , J. , and McKindsey , C.W. 2009 . Shellfi sh - DEPOMOD: modelling the biodeposition from suspended shellfi sh aquaculture and assessing benthic effects . Aquaculture 288 : 239 – 253 .
Weissberger , E.J. , Coiro , L.L. , and Davey , E.W. 2009 . Effects of hypoxia on animal burrow con-struction and consequent effects on sediment redox profi les . Journal of Experimental Marine Biology and Ecology 371 : 60 – 67 .
Whitlatch , R.B. , Hines , A.H. , Thrush , S.F. , Hewitt , J.E. , and Cummings , V. 1997 . Benthic faunal responses to variations in patch density and patch size of a suspension - feeding bivalve .
Journal of Experimental Marine Biology and Ecology 216 : 171 – 189 .
Widdows , J. , and Brinsley , M. 2002 . Impact of biotic and abiotic processes on sediment dynam-ics and the consequences to the structure and functioning of the intertidal zone . Journal of Sea Research 48 : 143 – 156 .
Widdows , J. , Brinsley , M.D. , Salkeld , P.N. , and Elliott , M. 1998 . Use of annular fl umes to deter-mine the infl uence of current velocity and bivalves on material fl ux at the sediment - water interface . Estuaries 21 : 552 – 559 .
Willows , R.I. , Widdows , J. , and Wood , R.G. 1998 . Infl uence of an infaunal bivalve on the erosion of an intertidal cohesive sediment: a fl ume and modeling study . Limnology and Oceanography 43 : 1332 – 1343 .
Widdows , J. , Brown , S. , Brinsley , M.D. , Salkeld , P.N. , and Elliott , M. 2000 . Temporal changes in intertidal sediment erodability: infl uence of bio-logical and climatic factors . Continental Shelf Research 20 : 1275 – 1289 .
Widdows , J. , Blauw , A. , Heip , C.H.R. , Herman , P.M.J. , Lucas , C.H. , Middelburg , J.J. , Schmidt , S. , Brinsley , M.D. , Twisk , F. , and Verbeek , H. 2004 . Role of physical and biological processes in sediment dynamics of a tidal fl at in Westerschelde Estuary, SW Netherlands . Marine Ecology Progress Series 274 : 41 – 56 .
Worm , B. , Barbier , E.B. , Beaumont , N. , Duffy , J.E. , Folke , C. , Halpern , B.S. , Jackson , J.B.C. , Lotze , H.K. , Micheli , F. , Palumbi , S.R. , Sala , E. , Selkoe , K.A. , Stachowicz , J.J. , and Watson , R. 2006 . Impacts of biodiversity loss on ocean ecosystem services . Science 314 : 787 – 790 .
Wotton , R.S. , and Malmqvist , B. 2001 . Feces in aquatic ecosystems . BioScience 51 : 537 – 544 .
Wu , R.S.S. 1995 . The environmental impact of marine fi sh culture: towards a sustainable future . Marine Pollution Bulletin 31 : 159 – 166 .
Ysebaert , T. , Hart , M. , and Herman , P.M.J. 2009 . Impacts of bottom and suspended cultures of mussels Mytilus spp. on the surrounding sedimentary environment and macrobenthic biodiversity . Helgoland Marine Research 63 : 59 – 74 .
Zwarts , L. , and Wanink , J. 1989 . Siphon size and burying depth in deposit - and suspension - feed-ing benthic bivalves . Marine Biology 100 : 227 – 240 .