sharks and the origins of vertebrate immunity
TRANSCRIPT

Some 500 million years ago the an-cestor of all jawed vertebratesemerged in the warm waters of the
earth’s vast primordial sea. Although itsidentity is shrouded in mystery, somepaleontologists believe that this ances-tor resembled certain members of a lat-er group of fish known as placoderms,which are known, at least, from the fos-sils they left behind. These ungainly crea-tures, some of which apparently grew tolengths of about seven meters, had a
head and pectoral region encased in pro-tective bony plates.
A living placoderm, or one of the oth-er possible ancient vertebrate forerun-ners, would of course add immeasurablyto our understanding of evolution. Per-haps most significantly, we would beable to see the workings of one of themost complex of bodily constituents—
the immune system—that existed shortlyafter some vertebrates made the criticaltransition from jawless to jawed form.
The transition is a key one in evolutionbecause it is a link in the course leadingto more advanced animals, includingthose that eventually crawled onto landand evolved into humans. It is likely thatmulticomponent, adaptive immune sys-tems began with the first vertebrates.The immune systems of surviving inver-tebrates, which are probably similar tothose of ancient ones, do not have theremarkable adaptive capabilities of ver-tebrate immunity.
Although the placoderms and their an-cestors are long gone, we do have thenext best thing: several of their phyloge-netic relations, including sharks, skates,rays and ratfishes. These creatures—withimmune systems that have also proba-bly changed little if at all since their ear-liest appearance hundreds of millions ofyears ago—may provide a window ontothis distant and extraordinary period inevolution.
During the past several years, mycolleagues and I have studied theimmune systems of some of thesecreatures. As might be expected,immunity in these living fossilsis different from that in suchlater animals as frogs, mon-keys and humans. Yet intrigu-ingly, when it comes to protect-ing their hosts against disease,infection and other ills, theseancient immune systems appearto be every bit as effective—if notmore so—than their more mod-ern counterparts.
Perhaps this is not surprising; thesubclass of elasmobranchs, which
includes sharks, skates and rays, hasexisted for as many as 450 million
years (Homo sapiens has been aroundfor approximately half a million years),surviving several mass extinctions thateliminated countless species. It is hardto imagine how such evolutionary suc-cess could have occurred in creatureswith immune systems that were any-thing less than unusually effective. Ourefforts to identify the features that havemade elasmobranch immunity so suc-cessful have had a valuable side benefit:insights into human immunity.
The Two Parts of Immunity
The adaptive immune system has twobasic parts, called humoral and cel-
lular. The agents of humoral immunityare known as B lymphocytes, or B cells.B cells produce protein molecules, or an-tibodies, that bind to foreign substanc-
Sharks and the Origins of Vertebrate Immunity Scientific American November 1996 67
PLACODERMS, of which only fossils remain, are believed to have been among theearly beneficiaries of multipart, adaptive immune systems.
Sharks and the Origins ofVertebrate ImmunitySharks, which have existed for as many as 450 million years, offer glimpses of a distantperiod in the evolution of the immune system
by Gary W. Litman
Copyright 1996 Scientific American, Inc.
es, or antigens, on potentially harmfulbacteria and viruses in the bloodstream.This binding enables other bodily enti-ties to destroy the bacteria and virusesby various means. Antibodies are alsoknown as immunoglobulins; humanshave five major types of them.
All the antibodies on a single B cellare of the same type and bind to a spe-cific antigen. If this antibody encoun-ters and binds to its corresponding anti-gen, the B cell is stimulated to repro-duce and to secrete its antibody. Mostof the human body’s billions of B cellsmake antibodies that are different fromone another, because during the forma-tion of each B cell a genetic process thathas both random and inherited compo-nents programs the cell to produce alargely unique “receptor”—the part ofthe antibody that actually binds to theantigen. It is this incredible diversityamong antigen receptors that gives suchvast range to humoral immunity.
Cellular immunity is carried out by adifferent group of immune cells, termedT lymphocytes, or T cells. In contrast toB cells, T cells do not produce antibodies;rather they recognize antigens bound toa type of molecule on the surface of adifferent kind of cell. For this purpose,they are equipped with a specializedclass of molecule, called a receptor. Typ-ical manifestations of T cells at workinclude such diverse phenomena as therejection of a foreign skin graft and thekilling of tumor cells.
Antibodies, or immunoglobulins, andT cell receptors are the primary meansby which the body can recognize spe-cific antigens. Although humoral andcellular immunity have basically differ-ent functions and purposes, they inter-act during an immune response. T cells,for example, help to regulate the func-tion of B cells.
In some ways, shark and skate immu-nity is similar to that of humans. Thesefish have a spleen, which, as in humans,is a rich source of B cells. When a sharkis immunized—that is to say, injectedwith an antigen—B cells respond by pro-ducing antibodies. The similarities ex-tend to cellular immunity. Like humans,sharks and skates have a thymus, inwhich T cells mature and from whichthey are released. Sharks also have T cellreceptors. Recent work by me and Jon-athan P. Rast, now at the California In-stitute of Technology, showed that, asin humans, diversity in these receptorsarises from the same kind of geneticmechanisms that give rise to antibody
diversity. Finally, skin grafted from oneshark to another ultimately results inrejection.
These similarities notwithstanding,there are some significant and fascinat-ing differences between the immune sys-tems of such cartilaginous fish as sharksand of humans. For example, cartilagi-nous fish have four different classes ofimmunoglobulin, only one of which isalso among the five major types in hu-mans. Furthermore, these shark anti-bodies lack the exquisite specificity thatpermits the recognition of, among oth-er things, the subtle differences betweentwo similar types of bacteria.
In addition, these antibodies lack thecapacity of human antibodies to bindmore and more strongly to an antigenduring the course of a prolonged im-mune response—a decided advantage infighting infection. A difference in cellu-lar immunity is implied by the fact thatsharks do not reject skin grafts vigor-ously and quickly, as humans do, butrather over a period of weeks.
Do these facts mean that the immunesystems of sharks and skates are lesssuited to the needs of the host in com-parison with those of humans and other
mammals? Not at all. Indeed, the idio-syncratic nature of this ancient immunesystem illustrates well the twists andturns that occurred during the evolu-tion of immunity. This sinuous course,moreover, suggests that evolution, atleast where the immune system is con-cerned, may not have always proceededin the inexorable, successive way inwhich it is often portrayed.
A Receptor for Every Antigen
Much of our work so far has beendevoted to elucidating the humor-
al immune system of the horned shark,a spotted creature that usually grows toabout a half meter in length. In this ani-mal, as in all vertebrates, the diversityin antigen receptors has a genetic basis.Specifically, each antibody’s antigen re-ceptor is formed through the interac-tions between two amino acid chains,which are protein molecules, character-ized as heavy and light. With few ex-ceptions, the basic antibody moleculehas two pairs of such chains and there-fore two antigen receptor sites. Exactlywhich antigen a receptor will bind todepends on the type and arrangement
Sharks and the Origins of Vertebrate Immunity68 Scientific American November 1996
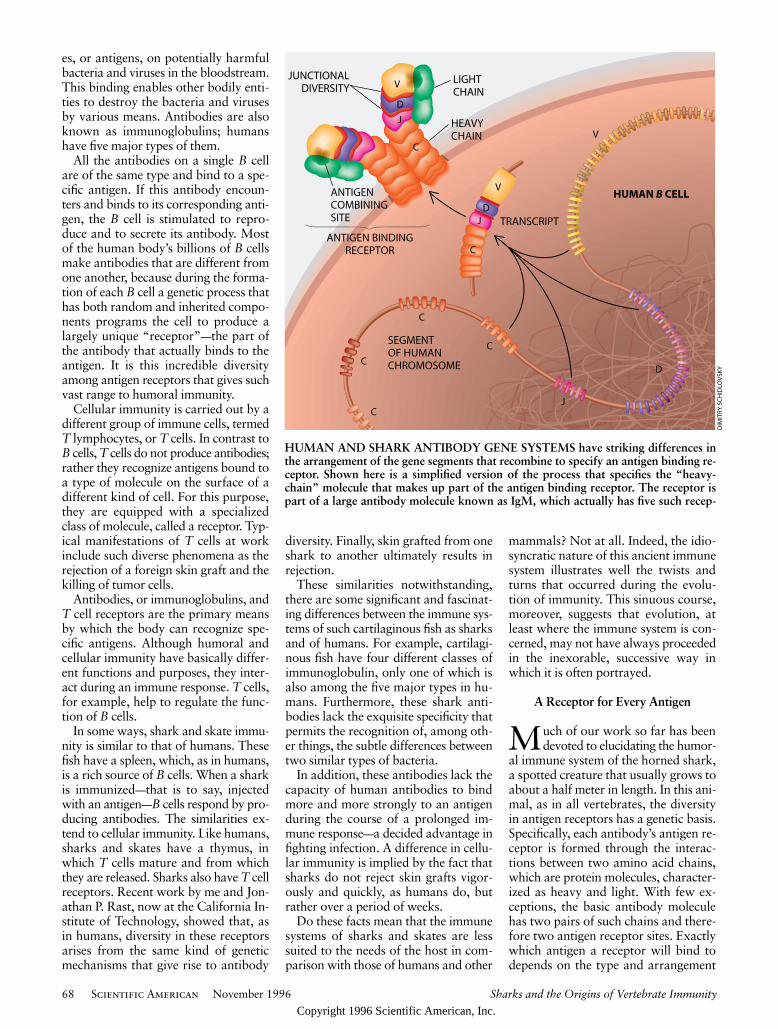
HUMAN AND SHARK ANTIBODY GENE SYSTEMS have striking differences inthe arrangement of the gene segments that recombine to specify an antigen binding re-ceptor. Shown here is a simplified version of the process that specifies the “heavy-chain” molecule that makes up part of the antigen binding receptor. The receptor ispart of a large antibody molecule known as IgM, which actually has five such recep-
V
D
J
C
C
C
C
V
D
J
C
JUNCTIONALDIVERSITY
V
DJ
C
ANTIGENCOMBININGSITE
ANTIGEN BINDINGRECEPTOR
SEGMENTOF HUMANCHROMOSOME
TRANSCRIPT
HUMAN B CELL
LIGHTCHAIN
HEAVYCHAIN
DIM
ITRY
SC
HID
LOV
SKY
Copyright 1996 Scientific American, Inc.
of the amino acids in the chains thatmake up the receptor.
Regardless of where they are producedin the body, amino acid chains are creat-ed in cells and specified by genes—whichact as a kind of blueprint—in the cell’snucleus. In the case of an antigen recep-tor, the amino acid chain is specified bygene segments, also known as antibodygenes, in the B cell’s nucleus. There arethree types of gene segments for thispurpose; they are designated V (“vari-able”), D (“diverse”) and J (“joining”).The amino acids in the heavy chain arespecified by all three types of gene seg-ments; the light chain is encoded by theV and J only. A fourth type of gene seg-ment, designated C (“constant”), deter-mines the class of antibody.
In humans the functional V, D, J andC segments are found on a single chro-mosome. As in most higher vertebrates,the segments are located in clusters,with, for example, some 50 functionalV, 30 D, six J and eight C elements in asingle location, occupying roughly amillion components, or “rungs,” of theDNA molecular “ladder.” (These rungsare the base pairs.) When a B cell’s gene-reading mechanisms produce an anti-
body, various cellular entities first re-combine single V, D and J segments ad-jacent to a C segment in a multistepprocess. This genetic material is then“read out” to the cell’s protein-makingsystems. The recombination of thesegene segments determines the antigen-binding characteristics of the antibody.In such higher vertebrates as humans,this joining of different V, D and J ele-ments, which is called combinatorial di-versity, is an important factor in antigenreceptor diversity.
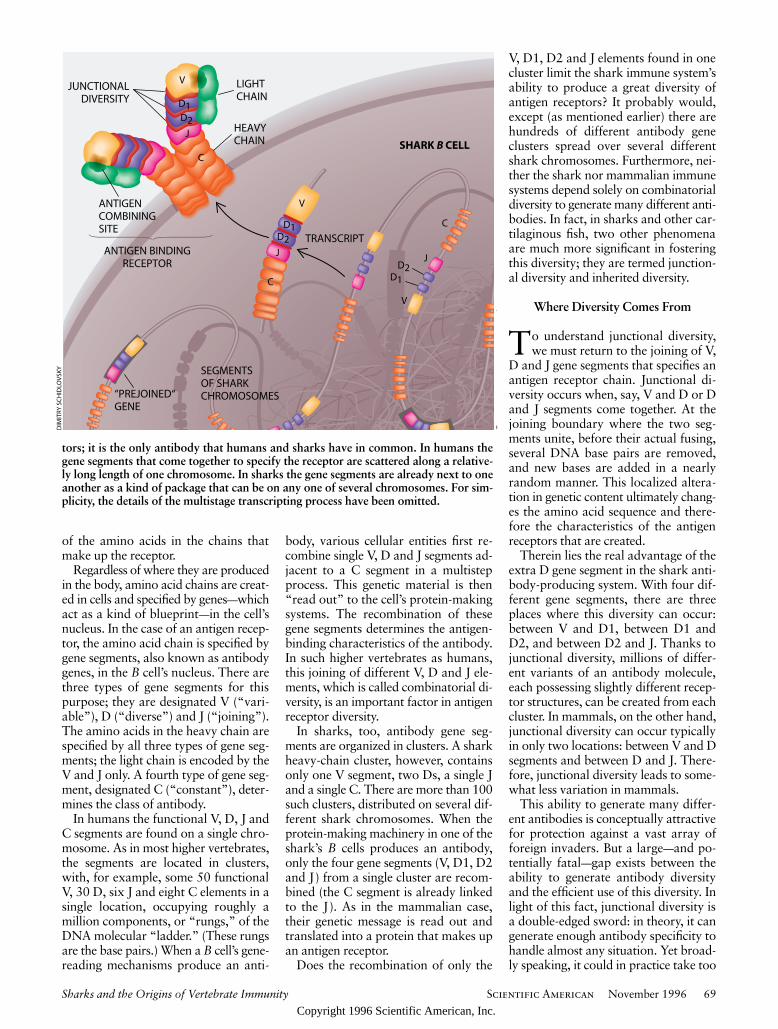
In sharks, too, antibody gene seg-ments are organized in clusters. A sharkheavy-chain cluster, however, containsonly one V segment, two Ds, a single Jand a single C. There are more than 100such clusters, distributed on several dif-ferent shark chromosomes. When theprotein-making machinery in one of theshark’s B cells produces an antibody,only the four gene segments (V, D1, D2and J) from a single cluster are recom-bined (the C segment is already linkedto the J). As in the mammalian case,their genetic message is read out andtranslated into a protein that makes upan antigen receptor.
Does the recombination of only the
V, D1, D2 and J elements found in onecluster limit the shark immune system’sability to produce a great diversity ofantigen receptors? It probably would,except (as mentioned earlier) there arehundreds of different antibody geneclusters spread over several differentshark chromosomes. Furthermore, nei-ther the shark nor mammalian immunesystems depend solely on combinatorialdiversity to generate many different anti-bodies. In fact, in sharks and other car-tilaginous fish, two other phenomenaare much more significant in fosteringthis diversity; they are termed junction-al diversity and inherited diversity.
Where Diversity Comes From
To understand junctional diversity,we must return to the joining of V,
D and J gene segments that specifies anantigen receptor chain. Junctional di-versity occurs when, say, V and D or Dand J segments come together. At thejoining boundary where the two seg-ments unite, before their actual fusing,several DNA base pairs are removed,and new bases are added in a nearlyrandom manner. This localized altera-tion in genetic content ultimately chang-es the amino acid sequence and there-fore the characteristics of the antigenreceptors that are created.
Therein lies the real advantage of theextra D gene segment in the shark anti-body-producing system. With four dif-ferent gene segments, there are threeplaces where this diversity can occur:between V and D1, between D1 andD2, and between D2 and J. Thanks tojunctional diversity, millions of differ-ent variants of an antibody molecule,each possessing slightly different recep-tor structures, can be created from eachcluster. In mammals, on the other hand,junctional diversity can occur typicallyin only two locations: between V and Dsegments and between D and J. There-fore, junctional diversity leads to some-what less variation in mammals.
This ability to generate many differ-ent antibodies is conceptually attractivefor protection against a vast array offoreign invaders. But a large—and po-tentially fatal—gap exists between theability to generate antibody diversityand the efficient use of this diversity. Inlight of this fact, junctional diversity isa double-edged sword: in theory, it cangenerate enough antibody specificity tohandle almost any situation. Yet broad-ly speaking, it could in practice take too
Sharks and the Origins of Vertebrate Immunity Scientific American November 1996 69
tors; it is the only antibody that humans and sharks have in common. In humans thegene segments that come together to specify the receptor are scattered along a relative-ly long length of one chromosome. In sharks the gene segments are already next to oneanother as a kind of package that can be on any one of several chromosomes. For sim-plicity, the details of the multistage transcripting process have been omitted.
V
J
C
D2
D2
D1
D1
V
D2D1
J
C
V
J
C
“PREJOINED“GENE
ANTIGENCOMBININGSITE
ANTIGEN BINDINGRECEPTOR
SEGMENTSOF SHARKCHROMOSOMES
TRANSCRIPT
SHARK B CELL
LIGHTCHAIN
HEAVYCHAIN
JUNCTIONALDIVERSITY
DIM
ITRY
SC
HID
LOV
SKY
Copyright 1996 Scientific American, Inc.
much time to generate enough anti-bodies, select the best ones, expand theirnumbers and then deal with the invad-ing pathogen; in other words, the hostcould lose a race with the infectiousagent.
To try to keep the host from losingthat race, the body relies on mechanismsthat rapidly select the “blueprint” of theimmediately needed antibody gene. Thisblueprint is first expressed by one B cellamong the body’s billions. In mammals,specialized cellular compartments andcomplex intercellular communi-cations mobilize and expand theimmune system for this purpose.
Sharks, on the other hand, relyheavily on a form of inherited di-versity. This form, the most dis-tinctive feature of the shark im-mune system, allows the animalsto avoid depending on a chanceoccurrence—for example, a fortu-itous combination of DNA basepairs attained through junctionaldiversity—to generate the rightantigen receptor at the right time.In a shark, a large percentage ofthe gene clusters in every cell areinherited with their V, D1, D2and J gene segments already en-tirely or partially “prejoined.”
In such clusters, there is limitedcapacity, or none at all, for junc-tional diversification. Analyses ofhundreds of these prejoined orpartially prejoined clusters haveshown their gene segments to beremarkably similar to those of or-dinary clusters, suggesting thatone type derived from the other atsome point in evolution.
But why? As in so many areas,our knowledge of genetic mecha-nisms has far surpassed an under-standing of their relation to func-tion. Still, it would be entirely reasonableto theorize that the humoral immunesystems of cartilaginous fish have evolvedto combine the best of two possibilities:a large number of genes that can recom-bine and thus provide immunologic flex-ibility, as well as some genes with fixedspecificities that can be mobilized quick-ly to make antibodies against pathogensthat these species encounter all the time.
Combinatorial, junctional and inherit-ed forms of diversity are not the extentof diversity-producing mechanisms. Inaddition, the two types of gene clustersundergo additional change through mu-tation, which occurs at a very high fre-quency in the antibody genes of higher
vertebrates. These mutations are direct-ed at altering the characteristics of theantigen receptor sites of antibodies.
One interesting conclusion from acomparison of human and shark hu-moral immunity is that some 450 mil-lion years of evolution did relatively lit-tle to change the molecules of antibodyimmunity; the protein structures of sharkand human antibodies are very similar.Moreover, the V, D and J sequences ofgene segments that specify the creationof antibodies are similar. What evolu-
tion did radically alter is the way thesegene segments that specify antibodiesare organized; it placed greater empha-sis on junctional and especially inheriteddiversity in sharks, for example. Thoughrelatively simple, the mechanisms of ge-netic diversification in the shark’s im-mune system seem in many ways moreefficient than those in such higher verte-brates as humans.
This finding confirms, not surprising-ly, that evolution has a way of uniquelyadapting systems to their hosts’ immedi-ate needs. In the case of immunity, evo-lution also has to provide for unexpectedchallenges as well. The surprise is thatin order to make that efficiency possi-
ble, enigmatic evolutionary leaps of un-characteristic magnitude apparentlysometimes occur, at least in antibodyimmunity, over relatively short periods.
Cellular Immunity
Many of the basic principles putforth in the discussion so far—the
rearrangement of widely spaced genesegments scattered along a stretch ofchromosome and the reading out andalteration of their genetic information
to specify the creation of antigenreceptors made up of amino acidchains—apply to cellular as wellas humoral immunity. After all, Tcells, just like the antibodies se-creted by B cells, must also recog-nize and bind to an almost limit-less assortment of antigens.
T cells and antibodies both havereceptors that are specified by sim-ilar gene segments. The basic mech-anisms of gene segment reassem-bly that produce antibody mole-cules also create T cell receptors.But a T cell receptor is found onlyon the cell’s surface and only rec-ognizes foreign material bound toa specialized molecule on a differ-ent cell. T cells’ affinities for for-eign materials are low in compar-ison to some antibodies, and theydo not undergo mutation in thesame manner as antibodies.
In the past, many immunolo-gists believed that cellular immu-nity predated humoral immunity.Yet the aforementioned chronicnature of skin graft rejection insharks suggests that, if anything,cellular immunity in the shark isnot robust and possibly lacks spec-ificity. This notion, in turn, impliedto some observers that sharks do
not have T cells.In order to test this hypothesis, my
colleagues and I set about determiningwhether the horned shark has T cells.Unequivocal proof of the existence of Tcells requires identification of their anti-gen receptors. For this purpose, theconventional approaches available untilrecently were inadequate. The break-through came with the development sev-eral years ago of a technique known asthe polymerase chain reaction (PCR),which can produce millions of copies ofa small section of DNA. We used a formof the PCR technique as part of a pro-cess that produced great numbers of Tcell receptor genes in order to character-
Sharks and the Origins of Vertebrate Immunity70 Scientific American November 1996
SHARKS AND HUMANS share a number of im-munologic features, including a thymus and a spleen.
ROB
ERTO
OST
I
THYMUS
SPLEEN
Copyright 1996 Scientific American, Inc.

ize them. Recently we found all fourclasses of mammalian T cell antigen re-ceptors in the skate and have evidencesuggesting their presence in the shark.
Extensive characterization of one ofthe classes of shark T cell receptorsshowed it to be about as complexly di-versified as its human equivalent. Thisfinding surprised us, indicating that incontrast to antibody gene organization,T cell receptor genes seem to have un-dergone no major changes since the timeof the divergence of the sharks from theevolutionary line leading to the mam-mals some 450 million years ago. Theantibody gene system and the T cell re-ceptor gene system may well have di-verged from a common ancestor thatmore closely resembled the latter, al-though the opposite can also be ar-gued—that it was an antibody-gene-likeancestor that gave rise to both catego-ries of immune gene systems.
As the genomes of sharks and theirrelatives continue to be characterized,we now recognize a variety of differentgene clusters. For example, a group ledby Martin F. Flajnik at the University ofMiami recently found gene clusters thatresemble those of both antibodies andT cell receptors. Intriguingly, the genesin these clusters undergo extraordinaryrates of mutation.
Ongoing studies have also suggestedthat immune system genes from differ-ent clusters have “mixed and matched”with one another during evolution. Withhundreds of clusters and plenty of genet-ic backup, exchange between clustersmay have been a very efficient means ofgenerating novel gene clusters. It is quitepossible, too, that our continuing stud-ies will identify even more receptors inthe shark immune system.
With respect to this exchange amongdifferent clusters, the peculiar redundan-cy of different immune receptor gene
clusters in the shark—the groupings ofessentially identical V, D1, D2 and J seg-ments repeated over and over on variouschromosomes—can be seen in an en-tirely new light. In short, this recombi-nation, along with other unique fea-tures of the shark’s genetic mechanisms,affords a means for rapidly evolvingnew families of receptor molecules. Inmammals the gene segments are isolat-ed to single chromosomes, and littlestructural redundancy is evident; thesefacts mean that the opportunity for thistype of recombination is remote.
Furthermore, duplication of gene seg-ments—the existence of multiple Vs, Dsand Js, a hallmark of the mammalianimmune system—appears to come atthe price of introduction and retentionof significant numbers of nonfunctionalgenetic elements. In sharks and skates,on the other hand, nonfunctional ele-ments are uncommon and probably arelost quickly from the genome.
As surviving representatives of a very
ancient line, sharks, skates and their re-lations may be our only remaining linkto the distant origins of T and B cell im-munity. These fish offer a unique glimpseof a pivotal moment in the course ofevolution. Through this window wemay someday begin to see the elementsthat drove the evolution of a systemthat in different ways is as protective, ifnot more so, as the armor plates of theancient placoderm.
If we are correctly reading the evolu-tionary record, several questions cometo mind. Was it the relentless nature ofthe challenge from pathogens that ledto relatively sudden, radical changes inthe way that antibody genes are orga-nized? Do these lessons from the pre-historic vertebrates and the profounddifferences seen in contemporary mam-mals suggest that the immune system ispoised for quick change? This scenariomay well be the case, forcing us to re-think our notions of evolutionary selec-tion and adaptation.
Sharks and the Origins of Vertebrate Immunity Scientific American November 1996 71
The Author
GARY W. LITMAN is Hines Professor of Pediatrics andprofessor of biochemistry and molecular biology in the Col-lege of Medicine at the University of South Florida. He is alsohead of the division of molecular genetics at All Children’sHospital in St. Petersburg. The author of 187 publicationsand reviews (and counting), he recently received a MERITAward from the Allergy and Infectious Disease Branch of theNational Institutes of Health. He spends his free time withhis family, his orchids and a collection of rare palm trees. Theauthor wishes to acknowledge the assistance of colleaguesChris Amemiya, Michele K. Anderson, Martin F. Flajnik,Carl Luer, Jonathan P. Rast and Michael J. Shamblott.
Further Reading
Evolution of the Immune System. Louis DuPasquier in FundamentalImmunology. Third edition. Edited by William E. Paul. Raven Press, 1993.
How the Immune System Recognizes Invaders. Charles A. Janeway, Jr.,in Scientific American, Vol. 269, No. 3, pages 72–79; September 1993.
Phylogenetic Diversification of Immunoglobulin Genes and theAntibody Repertoire. Gary W. Litman, Jonathan P. Rast, Michael J.Shamblott et al. in Molecular Biology and Evolution, Vol. 10, No. 1, pages60–72; January 1993.
Molecular Evolution of the Vertebrate Immune System. SimonaBartl, David Baltimore and Irving L. Weissman in Proceedings of the Na-tional Academy of Sciences, Vol. 91, No. 23, pages 10769–10770;November 8, 1994.
SA
MIC
HA
EL S
EXTO
N A
ll C
hild
ren’
s H
ospi
tal
HORNED SHARKS are among the most ancient creatures in which T cells, the agentsof cellular immunity, have been conclusively identified.
Copyright 1996 Scientific American, Inc.