sex differences in ethanol-induced hypnosis and hypothermia in young long-evans rats
TRANSCRIPT

Sex Differences in Ethanol-Induced Hypnosis andHypothermia in Young Long-Evans Rats
Barbara Webb, Patricia W. Burnett, and Don W. Walker
Introduction: Females experience greater liver damage, have reduced brain size, and have greatermemory deficits than do males with a similar history of alcoholism. Females have higher peak alcohol levelsand faster elimination rates than males. Our goal was to study sex differences in the response of youngethanol-naïve outbred Long-Evans rats to acute ethanol exposure so that we may better understand whyfemales are more sensitive to alcohol toxicity than males.
Methods: Females aged 49 days and males aged 43 days, weighing 153.6 and 177.5 g, respectively, weretested for their initial response to ethanol. Fasted (12 hr) females (in diestrous) and males were given anintraperitoneal injection of 3.0 g/kg of ethanol (v/v in 0.9% sterile saline). Body temperature, loss of therighting reflex (LORR), return of the righting reflex, and tail blood alcohol concentration (BAC) weremonitored.
Results: LORR occurred at the same time in females and males. The return of the righting reflexoccurred later in males than in females. BACs were the same in the males and females except at LORR,when BAC was lower in the males. Acute ethanol tolerance developed in more males than females. Femalesdemonstrated a slower recovery from peak ethanol-induced hypothermia than males. The proportions oflean body mass, ethanol elimination, and ethanol metabolism were similar in the females and males.
Conclusions: Ethanol-naïve young male and female Long-Evans rats demonstrated sex differences intheir initial responses to ethanol. Males were more sensitive than females to the hypnotic effect of ethanol,whereas females were more sensitive than males to ethanol-induced hypothermia. In addition, more malesthan females developed acute ethanol tolerance. Investigating the mechanisms underlying these differencesmay help us to understand why females experience more of the adverse effects of alcohol consumption thanmales.
Key Words: Sex, Body Temperature, Acute Ethanol Tolerance, Ethanol Elimination, Rats.
RESEARCH HAS SHOWN that men and women differfrom each other in peak alcohol level, first-pass me-
tabolism of alcohol (Lieber, 2000), and the rate of alcoholelimination (Lancaster, 1994; Thomasson, 1995). These dif-ferences may account for the greater morbidity and mor-tality that women alcoholics show as compared with malealcoholics (Schmidt and Popham, 1980; Wilkinson, 1980).Reports in the literature show that sex differences in ro-dents parallel those in humans; e.g., females have higherblood alcohol levels (Rivier, 1993; Rivier et al., 1992) andfaster elimination rates (Collins et al., 1975; Erickson, 1984;Eriksson, 1973) than males. Still other studies report con-tradictory findings (Hutchins et al., 1981; Middaugh et al.,1992; Seitz et al., 1992). Many studies have reported sex
differences in behavioral effects of intoxication, such asduration of loss of righting reflex [LORR; for review, seeLancaster (1995)]. Drinking behaviors are regulated bymultiple neural systems. However, alcohol levels in thebrain are not different in female and male rats (Crippens etal., 1999).
The initial sensitivity of rats to ethanol varies accordingto rat strain and between females and males of a particularstrain. Blood alcohol concentration (BAC) at LORR isused as a measure of ethanol sensitivity. Average sleep time[time from LORR to the return of righting reflex(RORR)], acute ethanol tolerance, and body temperatureare all measurements that reflect how ethanol affects thebrain. Average sleep time is influenced by both the sensi-tivity of the brain to ethanol and the rate of ethanol me-tabolism. Blood ethanol correlates with metabolism andelimination in rat inbred strains; the higher the rate ofmetabolism, the higher the BAC at RORR (Spuhler andDeitrich, 1984). Most sex comparisons in rodent modelshave shown that females eliminate ethanol more quicklythan do males (Erickson, 1984; Eriksson, 1973; Marshall etal., 1983).
In this study we injected young ethanol-naïve rats withethanol and measured the initial ethanol sensitivity, the
From the Department of Neuroscience, McKnight Brain Institute (BW,PWB, DWW), and Research Service (DWW), Malcom Randall VeteransMedical Center, Gainesville, Florida.
Received for publication May 1, 2001; accepted February 25, 2002.Supported by NIAAA Grants AA00200 and AA10480 and the Medical
Research Service Department of Veterans Affairs.Reprint requests: Barbara Webb, PhD, Department of Neuroscience, Uni-
versity of Florida, Box 100244, Gainesville, FL 32610; Fax: 352-392-8347;E-mail: [email protected]
Copyright © 2002 by the Research Society on Alcoholism.
0145-6008/02/2605-0695$03.00/0ALCOHOLISM: CLINICAL AND EXPERIMENTAL RESEARCH
Vol. 26, No. 5May 2002
Alcohol Clin Exp Res, Vol 26, No 5, 2002: pp 695–704 695

hypnotic response to ethanol, the effect of ethanol on bodytemperature, the development of acute ethanol tolerance,and ethanol elimination. We hypothesized that there aresex and individual differences in the response of Long-Evans rats to acute ethanol exposure.
METHODS
Ethanol-naïve 49-day-old female (n � 32) and 43-day-old male (n �32) Long-Evans rats (Charles River, Portage, Canada) were tested forinitial sensitivity to ethanol, ethanol elimination, and development ofacute ethanol tolerance. The females weighed 134 to 173 g, and the malesweighed 151 to 216 g when tested. All rats were housed individually inwire-bottomed cages that had holes in the back wall of the cage. Thisallowed for limited socialization between rats that were housed acrossfrom one another. The rats were given 1 week to adjust to their newsurroundings before testing. Estrous cycle was monitored in the females(vaginal smears) for the week before testing. The males were handled ina manner (removed from their cages) similar to the females. All femaleswere tested during diestrous.
Acute Ethanol Exposure Procedure
All the rats (32 females and 32 males) were fasted for 12 hr before andduring testing. Each rat was tested once for ethanol sensitivity. All maleand female rats were weighed, and rectal body temperature was measured.Next, the tip of the tail was clipped, and a control (preinjection) bloodsample (20 �l) was obtained. Rats were given an intraperitoneal (ip)injection of 3.0 g/kg of ethanol (95%) (20% v/v in sterile 0.9% saline) inthe right abdominal cavity. Blood samples were obtained for measuring(NAD-ADH Assay Kit, Sigma Chemical Co., St. Louis, MO) BACs im-mediately after the ip injection (time 0), at LORR, at RORR, and hourlyfor 4 to 5 hr after RORR. Body temperature was taken before blood wascollected at all time points (except at time 0). The length of time to LORRand to RORR was recorded. LORR was defined as the failure of the ratto right itself twice within 20 sec after being placed on its back. RORR wasdefined as the ability of the rat to right itself after being placed on its backtwice within 20 sec and support itself with all four legs under its body. Thedevelopment of acute tolerance occurred when the BAC at RORR washigher than the BAC at LORR. Ethanol sensitivity was determined byBACs at LORR. After RORR, the rats were returned to their cages, hadfree access to water, and were given food (rat chow) after 2 hr.
Ethanol Elimination and Metabolism
The term elimination is defined here as the apparent rate of disappear-ance of ethanol from the blood (mg/dl/hr) as calculated from the slope ofthe linear decreasing portion of the BAC curve (from 2 to 5 hr afterRORR) that was no longer significantly affected by the processes ofabsorption and distribution (Kalant, 1971; Ott et al., 1985; Sturtevant,1976; Widmark, 1933). Because the disappearance curve for each rat waslinear (r2 ranged from 0.954 to 0.997 in the females and from 0.97 to 0.997in the males) and the BAC values observed were all greater than 30.0mg/dl, the Widmark model (Sturtevant, 1976; Widmark, 1933) wasdeemed appropriate. The term metabolic rate is defined as milligrams ofethanol removed per hour per kilogram of body weight (mg/kg/hr) andcorrects for the effect of variations in the volume of distribution on therate of ethanol elimination from blood (Ott et al., 1985).
Statistics
BAC, body temperature, and time to LORR and RORR are reportedas mean � SD. Rats that did not show signs of LORR by 30 min wereeliminated from the statistical analyses. Linear regression was used todetermine the rate of ethanol elimination and metabolism. Treatmenteffects of ethanol and sex (and interactions) were determined by ANOVA
followed by Bonferroni/Dunn post hoc comparisons within significantvariables or, when appropriate, by repeated-measures ANOVA (Stat-View, SAS Institute, Inc., Cary, NC). The significance level was set at p �
0.05.
RESULTS
Females
Sensitivity and Tolerance to Acute Ethanol Exposure. Thefasted females (n � 32) were all cycling (determined byvaginal cytology) and were tested for their initial responseto ethanol during diestrous. LORR occurred in 4.6 � 4.9min after ip injection of 3.0 g/kg ethanol in the females (n �28) that showed definite signs of LORR before 30 min(Table 1). The BAC at LORR was 248 � 90 mg/dl (Table2). The control body temperature was 37.66 � 0.7°C, andbody temperature was 37.2 � 0.7°C at LORR (Table 3).
RORR occurred 71.0 � 30.0 min after ip injection ofethanol in the females (Table 1). BAC at RORR was 259 �23 mg/dl, and body temperature had decreased to 35.7 �0.72°C. The females demonstrated individual differences inthe development of acute ethanol tolerance. Acute ethanoltolerance developed in only 10 of the females (Table 2). Asa result of the low number of females to develop acutetolerance, the BACs in the females (n � 28) were the sameat LORR and RORR.
Body Temperature, Ethanol Elimination, and Ethanol Me-tabolism. BACs were measured at LORR and RORR and
Table 1. Loss of Righting Reflex (LORR) and Return of Righting Reflex (RORR)in Female Long-Evans Rats
Tolerance No. LORR (min) RORR (min) Time L-R
1 1 47 462 3 99 963 2 79 774 5 68 636 3 48 457 2 88 868 3 63 609 3 37 34
10 2 44 4211 2 68 6612 2 50 4814 4 21 1715 2 78 7616 3 22 1917 6 76 7018 7 120 11320 4 84 8021 2 95 9322 3 67 6423 18 105 8724 3 56 5325 3 25 2226 2 121 11927 22 120 9828 4 41 3729 12 120 10830 3 81 7831 2 65 63
Tolerance No. is the number of the individual. LORR and RORR are expressedas minutes after ip injection of 3 g/kg of ethanol. Females that did not show LORRby 30 min (n � 4) were excluded. Time L-R is the duration of LORR in minutes.
696 WEBB ET AL.
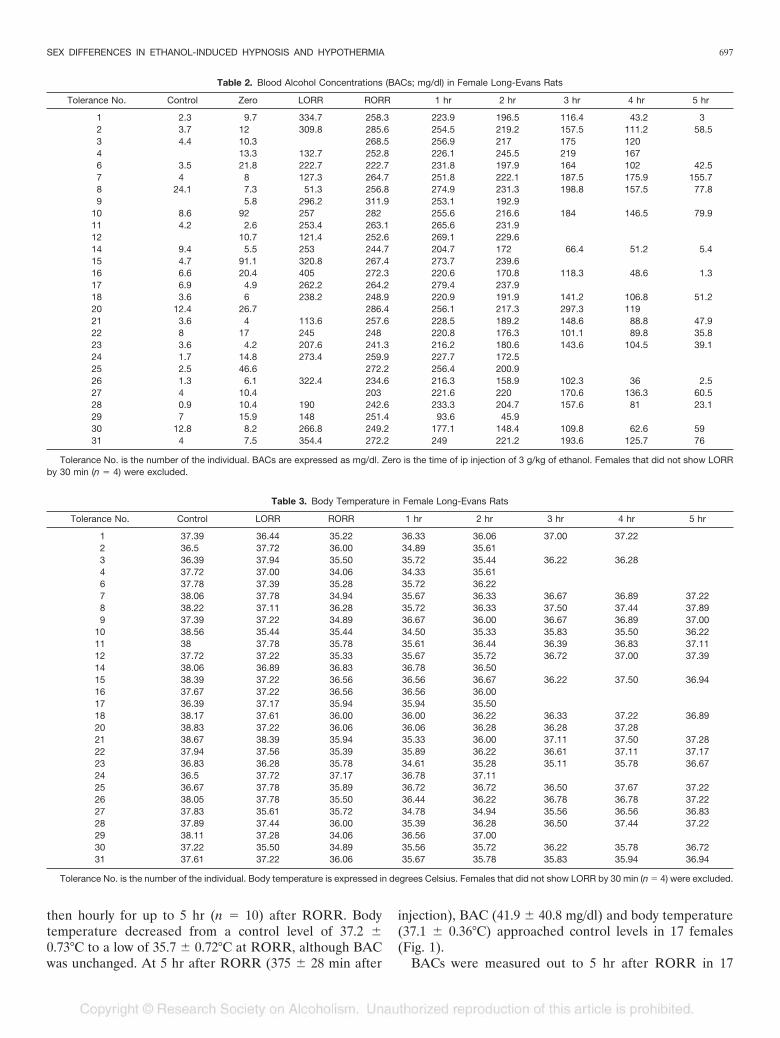
then hourly for up to 5 hr (n � 10) after RORR. Bodytemperature decreased from a control level of 37.2 �0.73°C to a low of 35.7 � 0.72°C at RORR, although BACwas unchanged. At 5 hr after RORR (375 � 28 min after
injection), BAC (41.9 � 40.8 mg/dl) and body temperature(37.1 � 0.36°C) approached control levels in 17 females(Fig. 1).
BACs were measured out to 5 hr after RORR in 17
Table 2. Blood Alcohol Concentrations (BACs; mg/dl) in Female Long-Evans Rats
Tolerance No. Control Zero LORR RORR 1 hr 2 hr 3 hr 4 hr 5 hr
1 2.3 9.7 334.7 258.3 223.9 196.5 116.4 43.2 32 3.7 12 309.8 285.6 254.5 219.2 157.5 111.2 58.53 4.4 10.3 268.5 256.9 217 175 1204 13.3 132.7 252.8 226.1 245.5 219 1676 3.5 21.8 222.7 222.7 231.8 197.9 164 102 42.57 4 8 127.3 264.7 251.8 222.1 187.5 175.9 155.78 24.1 7.3 51.3 256.8 274.9 231.3 198.8 157.5 77.89 5.8 296.2 311.9 253.1 192.9
10 8.6 92 257 282 255.6 216.6 184 146.5 79.911 4.2 2.6 253.4 263.1 265.6 231.912 10.7 121.4 252.6 269.1 229.614 9.4 5.5 253 244.7 204.7 172 66.4 51.2 5.415 4.7 91.1 320.8 267.4 273.7 239.616 6.6 20.4 405 272.3 220.6 170.8 118.3 48.6 1.317 6.9 4.9 262.2 264.2 279.4 237.918 3.6 6 238.2 248.9 220.9 191.9 141.2 106.8 51.220 12.4 26.7 286.4 256.1 217.3 297.3 11921 3.6 4 113.6 257.6 228.5 189.2 148.6 88.8 47.922 8 17 245 248 220.8 176.3 101.1 89.8 35.823 3.6 4.2 207.6 241.3 216.2 180.6 143.6 104.5 39.124 1.7 14.8 273.4 259.9 227.7 172.525 2.5 46.6 272.2 256.4 200.926 1.3 6.1 322.4 234.6 216.3 158.9 102.3 36 2.527 4 10.4 203 221.6 220 170.6 136.3 60.528 0.9 10.4 190 242.6 233.3 204.7 157.6 81 23.129 7 15.9 148 251.4 93.6 45.930 12.8 8.2 266.8 249.2 177.1 148.4 109.8 62.6 5931 4 7.5 354.4 272.2 249 221.2 193.6 125.7 76
Tolerance No. is the number of the individual. BACs are expressed as mg/dl. Zero is the time of ip injection of 3 g/kg of ethanol. Females that did not show LORRby 30 min (n � 4) were excluded.
Table 3. Body Temperature in Female Long-Evans Rats
Tolerance No. Control LORR RORR 1 hr 2 hr 3 hr 4 hr 5 hr
1 37.39 36.44 35.22 36.33 36.06 37.00 37.222 36.5 37.72 36.00 34.89 35.613 36.39 37.94 35.50 35.72 35.44 36.22 36.284 37.72 37.00 34.06 34.33 35.616 37.78 37.39 35.28 35.72 36.227 38.06 37.78 34.94 35.67 36.33 36.67 36.89 37.228 38.22 37.11 36.28 35.72 36.33 37.50 37.44 37.899 37.39 37.22 34.89 36.67 36.00 36.67 36.89 37.00
10 38.56 35.44 35.44 34.50 35.33 35.83 35.50 36.2211 38 37.78 35.78 35.61 36.44 36.39 36.83 37.1112 37.72 37.22 35.33 35.67 35.72 36.72 37.00 37.3914 38.06 36.89 36.83 36.78 36.5015 38.39 37.22 36.56 36.56 36.67 36.22 37.50 36.9416 37.67 37.22 36.56 36.56 36.0017 36.39 37.17 35.94 35.94 35.5018 38.17 37.61 36.00 36.00 36.22 36.33 37.22 36.8920 38.83 37.22 36.06 36.06 36.28 36.28 37.2821 38.67 38.39 35.94 35.33 36.00 37.11 37.50 37.2822 37.94 37.56 35.39 35.89 36.22 36.61 37.11 37.1723 36.83 36.28 35.78 34.61 35.28 35.11 35.78 36.6724 36.5 37.72 37.17 36.78 37.1125 36.67 37.78 35.89 36.72 36.72 36.50 37.67 37.2226 38.05 37.78 35.50 36.44 36.22 36.78 36.78 37.2227 37.83 35.61 35.72 34.78 34.94 35.56 36.56 36.8328 37.89 37.44 36.00 35.39 36.28 36.50 37.44 37.2229 38.11 37.28 34.06 36.56 37.0030 37.22 35.50 34.89 35.56 35.72 36.22 35.78 36.7231 37.61 37.22 36.06 35.67 35.78 35.83 35.94 36.94
Tolerance No. is the number of the individual. Body temperature is expressed in degrees Celsius. Females that did not show LORR by 30 min (n � 4) were excluded.
SEX DIFFERENCES IN ETHANOL-INDUCED HYPNOSIS AND HYPOTHERMIA 697
females. The BAC curves had a good linear fit (r2 rangingfrom 0.954 to 0.997) between 2 and 5 hr after RORR (asdetermined by linear regression), and BAC was greaterthan 30 mg/dl (the criterion for using Widmark model) in10 of the 17 females. In those 10 females, the ethanolelimination rate was 47.05 � 9.5 mg/dl/hr.
We first calculated the theoretical time 0 BAC (C0) andthe theoretical volume of distribution, r, in the 10 femalesto calculate the rate of ethanol metabolism (Ott et al.,1985). For each rat, C0 was taken as the time 0 intercept ofthe linear regression equation fitting the individual datapoints from 2 to 5 hr after RORR. The Widmark r valuesfor each rat were calculated from the equation r � dose(g/kg)/C0 (g/liter) and represent the theoretical volume ofdistribution of ethanol in the body, indicating the propor-tion of lean body mass of the animal (Ott et al., 1985). C0in the females was 361.44 � 32.5 mg/dl, r was 0.837 � 0.082,and the metabolic rate [elimination rate � r (Widmark,1933)] was 38.78 � 6.6 mg/dl/hr.
Males
Sensitivity and Tolerance to Acute Ethanol Exposure.LORR occurred in 6.3 � 6.6 min after ip injection of 3.0g/kg of ethanol in the males (n � 29) that showed definitesigns of LORR before 30 min (Table 4). BAC at LORRwas 161 � 91 mg/dl (Table 5). Control body temperaturewas 36.8 � 0.58°C and was unchanged (36.7 � 0.81°C) atLORR (Table 6).
RORR occurred in 97.6 � 29 min after ip injection ofethanol in the males (Table 4). BAC at RORR was 260 �26 mg/dl when peak hypothermia occurred (34.7 � 0.97°C).The males demonstrated individual differences in the de-velopment of acute ethanol tolerance. Acute ethanol toler-ance developed in 22 of the 28 males (Table 2). The highnumber of males with low BACs at LORR that developedacute ethanol tolerance caused BAC to be significantly
higher (df � 27, paired t test; p � 0.0001) at RORR than atLORR.
Body Temperature, Ethanol Elimination, and Ethanol Me-tabolism. BACs were measured at LORR and RORR andthen hourly for 5 hr after RORR. At 5 hr after RORR (398� 33 min after injection), BAC approached control levels(35.6 � 27.2 mg/dl) and body temperature increased tocontrol levels (36.8 � 0.49°C; Fig. 2) in the males (n � 10).
BACs were measured out to 5 hr after RORR in 19males. In 10 of the males, the BAC curves had a good linearfit (r2 ranging from 0.97 to 0.997) between 2 and 5 hr afterRORR (as determined by linear regression), and BAC wasgreater than 30 mg/dl (criterion for using the Widmarkmodel). Ethanol elimination rate in the 10 males was 53.2� 6.6 mg/dl/hr (see previous section). The calculated C0 inthose males was 395.89 � 38.9 mg/dl, r was 0.764 � 0.073,and the metabolic rate was 40.4 � 4.2 mg/dl/hr.
Sex Differences
There were sex differences in the response of theethanol-naïve Long-Evans rats to acute ethanol exposure.Although LORR occurred at the same time in females andmales, the females were less sensitive to the hypnotic effectof ethanol. BACs were significantly lower [F(1) � 13.877; p� 0.0005] in the males at LORR than in the females,reflecting an increased sensitivity of the males to ethanol.As a result of their increased sensitivity to the hypnotic
Fig. 1. Female Long-Evans rats’ hypothermic response to ethanol injection.Females were fasted for 12 hr and then given an ip injection of 3.0 g/kg of ethanol.Body temperature (°C) was measured before injection, at loss of righting reflex, atreturn of righting reflex (RORR), and then hourly after RORR for up to 5 hr. Peakhypothermia (35.7 � 0.72°C) occurred at RORR (71.0 � 30.0 min after ethanolinjection).
Table 4. Loss of Righting Reflex (LORR) and Return of Righting Reflex (RORR)in Male Long-Evans Rats
Tolerance No. LORR (min) RORR (min) Time L-R
1 2 120 1183 4 68 644 28 103 755 17 85 686 3 56 538 3 71 68
10 9 120 11111 3 120 11712 23 120 9713 5 92 8714 3 55 5215 9 120 11116 3 40 3717 3 120 11718 3 39 3619 16 109 9320 3 120 11721 3 120 11722 2 120 11823 3 52 4924 12 120 10825 3 120 11726 4 117 11327 2 120 11828 3 120 11729 3 84 8130 3 59 5631 6 120 11432 3 120 117
Tolerance No. is the number of the individual. LORR and RORR are expressedas minutes after ip injection of 3 g/kg of ethanol. Males that did not show LORRby 30 min (n � 3) were excluded. Time L-R is the duration of LORR in minutes.
698 WEBB ET AL.

response to ethanol, LORR was significantly longer [F(1)� 10.879; p � 0.0017] in the males (97.6 � 29 min) com-pared with the females (71.0 � 30.0 min).
We used change from baseline body temperature valuesfor sex comparisons because body temperature was signif-icantly higher [F(1) � 38.464; p � 0.0001] in the femalesthan in the males. Repeated-measures ANOVA (sex �time) of body temperature change over the entire timeperiod resulted in a significant effect of sex [F(1) � 5.169;p � 0.0301] and of time [F(6) � 60.680; p � 0.0001]. Theinteraction between sex and time, a reflection of the effectof sex on the rate of temperature change, was also signifi-cant [F(1,6) � 2.322; p � 0.0347], with females demonstrat-ing a slower recovery from peak hypothermia than themales (Figs. 1 and 2). Although the magnitude ofthe change in body temperature at RORR was the same inthe males and females, the decrease in body temperaturein the females occurred without any significant change inBAC, whereas BAC was significantly higher in the males.
Repeated-measures ANOVA (sex � time) of BACs overthe entire time period resulted in a significant effect of time[F(8) � 244.002; p � 0.0001], but not sex. There was asignificant interaction between sex and time [F(1,8) �15.465; p � 0.0001], a reflection of the effect of sex on therate of change in BAC. The proportion of lean body mass,the rate of ethanol elimination, and the rate of ethanolmetabolism were the same in the females and males.
However, BAC curves of the data from all of the females(n � 28) and males (n � 29) showed that there was morevariability in ethanol elimination among the females thanthe males (Figs. 3 and 4). The BAC curves also demon-strated that males took significantly longer [F(1) � 5.675;p � 0.0411] than females to eliminate ethanol. Because therate of ethanol elimination and the BACs were the same inthe females and males during the linear portion of thecurve (between 2 and 5 hr after RORR) and LORR waslonger in the males than in the females, the longer elimi-nation time in the males probably represents sex differ-ences in the absorption and distribution of ethanol duringthe early, nonlinear portion of the BAC curve.
DISCUSSION
The overall objective of this study was to investigate sexdifferences in the response of rats to acute ethanol expo-sure. There were sex differences in initial ethanol sensitiv-ity. Males showed greater sensitivity than females to thehypnotic effect of ethanol. The greater sensitivity of malesto ethanol was reflected by LORR occurring at a lowerBAC and by LORR lasting longer than in the females. Sexdifferences in ethanol sensitivity in the Long-Evans ratsseemed to be the result of differences in the absorption anddistribution of ethanol during the early, nonlinear portionof the BAC curve. Sex also affected the rate of recovery
Table 5. Blood Alcohol Concentrations (BACs) in Male Long-Evans Rats
Tolerance No. Control Zero LORR RORR 1 hr 2 hr 3 hr 4 hr 5 hr
1 4.9 13.6 97.7 252.2 219 178.5 133 91.5 30.22 1.6 3.1 251.1 265.8 247.8 225 201.8 156.8 88.83 8 13.2 244.9 275.4 246.2 201.1 153.1 85.9 6.74 6.7 8.3 222.1 275.2 249.7 197.65 10.9 5.4 254.7 246.4 237 196.26 1.6 2.9 64.2 275.2 256.7 212.1 184.6 107.4 59.97 3.4 8.3 236.9 253.5 230.3 198.1 145.9 82.4 23.18 3.4 10.2 160.3 271.4 286.7 185.79 3.4 12.8 223.4 235 231.8 202.9 152.8 89.7 21.8
10 8.7 6 218.6 253.3 215.1 185.5 142.1 111.5 53.511 3.4 19.2 111.3 250.3 202.9 182.8 122.9 79.2 14.512 3.4 5.1 197 218.3 217 174.213 6.9 8.5 323.5 284.1 227.5 211.914 4.9 4.7 138 304.7 265.3 226.8 188.4 109.8 53.515 2.2 10.5 235.1 255.3 230.2 181.7 151.9 76 4.216 6.9 7.6 242.3 297.1 272.3 229.5 181.9 112.8 38.117 6.9 3.1 72.9 267.3 222.1 184 117.3 67.8 8.518 4.9 13.4 65.3 256.2 271.8 234.7 194.9 156.5 88.119 1.8 4.7 238 240 228.2 181.720 0.9 5.1 319 261.8 117.3 200.6 147.1 116.721 0 10.9 55.5 183.6 207.3 202.9 152.922 0.9 18 253.8 244.4 196.3 157.3 102.723 2.6 8.5 25.2 243.3 249.7 198.2 133.5 91.4 36.924 1.3 1.1 203.9 262.1 228.7 188.2 122.4 57.3 5.625 1.3 10.5 86.7 238.7 193.9 173.1 104.1 57.3 426 3.9 6.3 108.3 263.2 269.5 233.3 194.5 129.3 77.627 2 4.3 66 258.9 233.3 204.5 157.9 116.4 53.328 1.1 5.4 40.4 298.4 256.5 215.9 173 115.7 3529 2.4 4.6 281.8 282.5 241.1 185.830 5.9 5.3 22.5 294.1 262.4 237.431 4.8 5.9 192.3 240.9 217.9 19532 5 8.3 137.2 236.5 232.6 196.5
Tolerance No. is the number of the individual. BACs are expressed as mg/dl. Zero is the time of ip injection of 3 g/kg of ethanol. Males that did not show LORRby 30 min (n � 3) were excluded.
SEX DIFFERENCES IN ETHANOL-INDUCED HYPNOSIS AND HYPOTHERMIA 699

from peak hypothermia, with females having a slower rateof recovery than males.
Our results demonstrated that female Long-Evans ratshad higher BACs at LORR, higher variability in ethanolelimination, less sensitivity to the hypnotic effect of etha-nol, and a slower rate of recovery from peak hypothermiathan male Long-Evans rats. The fact that BACs werehigher at LORR in the females than in the males for thesame amount of alcohol is consistent with reports in hu-
mans. In humans, women drinking the same amount ofalcohol as men have higher BACs, mostly because womenhave less body water than men of similar body weight(Frezza et al., 1990; Taylor et al., 1996) and lower gastric(first-pass) alcohol metabolism (Frezza et al., 1990; Lieber,2000).
In this study we used tail blood as our source of bloodused to measure BAC. Levitt et al. (1994) demonstrated
Table 6. Body Temperature in Male Long-Evans Rats
Tolerance No. Control LORR RORR 1 hr 2 hr 3 hr 4 hr 5 hr
1 37.44 37.22 34.00 34.39 35.39 35.33 36.50 36.223 37.06 36.78 36.22 36.00 35.39 35.94 36.22 36.784 36.22 35.22 35.22 34.22 34.895 36.78 35.89 35.22 34.56 36.116 36.33 37.22 34.22 34.94 35.06 35.56 35.72 37.448 37.78 38.22 34.78 34.94 36.00
10 36.89 36.33 34.89 34.72 34.94 35.72 36.28 36.5611 36.00 37.39 34.11 36.00 35.50 35.78 37.67 37.1712 36.39 34.56 34.39 34.22 34.5613 37.11 36.61 35.33 34.44 35.1114 36.78 37.78 34.11 34.89 34.39 36.00 36.67 36.7215 36.89 35.72 32.89 35.22 33.94 35.78 36.78 36.8316 36.67 35.89 32.83 34.61 34.50 35.17 36.28 36.2217 36.11 36.56 33.89 35.39 35.28 35.83 37.39 37.3918 37.06 36.94 34.56 34.50 34.11 34.78 35.28 35.8319 37.11 36.61 33.61 34.50 34.1720 36.78 36.56 34.94 35.17 34.28 35.44 35.3921 37.39 37.56 33.00 34.00 34.50 34.7822 37.11 37.11 36.06 35.94 35.11 36.2223 37.61 37.28 36.67 35.78 35.72 36.11 35.72 36.6724 37.11 35.72 35.39 35.33 36.00 35.94 35.72 37.1125 36.06 36.78 35.11 35.50 35.22 36.22 35.72 37.2226 36.28 36.67 34.83 34.50 34.39 35.22 36.50 36.5027 36.11 36.39 34.00 35.33 35.33 35.28 35.94 36.7228 36.67 36.78 35.11 34.50 35.28 36.11 36.72 37.5629 38.50 37.39 35.39 36.22 36.7230 36.83 35.72 35.22 35.22 36.2231 36.28 37.06 35.61 35.61 35.5632 37.00 37.72 35.78 35.67 35.78
Tolerance No. is the number of the individual. Body temperature is expressed in degrees Celsius. Males that did not show LORR by 30 min (n � 3) were excluded.
Fig. 2. Male Long-Evans rats’ hypothermic response to ethanol injection.Males were fasted for 12 hr and then given an ip injection of 3.0 g/kg of ethanol.Body temperature (°C) was measured before injection, at loss of righting reflex, atreturn of righting reflex (RORR), and then hourly after RORR for up to 5 hr. Peakhypothermia (34.7 � 0.97°C) occurred at RORR (97.6 � 29 min after ethanolinjection).
Fig. 3. Combined BAC curve for all female Long-Evans rats that showed signsof LORR by 30 min (n � 28). Females were fasted for 12 hr and then given an ipinjection of 3.0 g/kg of ethanol. BAC (mg/dl) was measured before injection,immediately after injection, at LORR, at RORR, and up to 5 hr after RORR. Timeis measured in minutes after injection. BAC values were plotted against the timesfor RORR and 1 to 5 hr after RORR by regression. The combined BAC curvedemonstrates that there was considerable variability in ethanol elimination amongthe females (r2 � 0.779).
700 WEBB ET AL.
that BACs in tail vein blood lags behind that of arterial,jugular, or femoral vein blood. They reported that this wasthe result of a low blood perfusion:tissue water ratio in thetail and that it disappeared when the tail was heated to37°C. Because normal body temperature was lower in themales than in the females in our study, the males in ourstudy may have also had lower tail temperatures. If this wasthe case, we could have false low BACs for the males atLORR and, therefore, a false interpretation of their sensi-tivity to ethanol. To rule out this possibility, we examinedthe relationship between temperature at LORR and BACin females and males having low BACs at LORR (less than200 mg/dl). We found that body temperature and change inbody temperature were the same at LORR in all males andfemales with BACs less than 200 mg/dl. Although we can-not rule out the possibility that the temperature of the tailcaused lower BACs in some of the rats, it does not seemlikely that only male BACs were affected.
Women also seem to eliminate alcohol from the bloodfaster than men. This finding may be explained by the factthat alcohol is metabolized almost entirely in the liver(Levitt et al., 1997), and women have a higher liver volumeper unit lean body mass than do men (Kwo et al., 1998; Liet al., 1998). First-pass metabolism of alcohol by gastricalcohol dehydrogenase accounts for the absorption of aconsiderable amount of alcohol in the stomach. As a resultof higher first-pass metabolism, BACs are lower in menthan in women younger than 50 years old (Frezza et al.,1990). It is unlikely that first-pass metabolism was a factorin our study, because alcohol was given by ip injection.
Although most sex comparisons in rodent models haveshown that females eliminate ethanol more quickly than domales (Eriksson, 1973; Wilson et al., 1984), this was not thecase in our study. In some studies that showed sex differ-
ences in ethanol elimination, female rats were shown tohave significantly different ethanol elimination rates frommales in five of eight inbred strains and in the N/NIH line(Erickson, 1984). Although we report no sex differences inethanol elimination rate in Long-Evans rats, we do showthat ethanol elimination is more variable among the fe-males than among the males. The most likely explanationfor this is sex-specific genetic differences in ethanol absorp-tion, distribution, or both among the females. Using labo-ratory animals, Fernandez et al. (1997) attempted to iden-tify sex-specific genetic factors whose interactions mightcontribute to a differential sensitivity to alcohol’s effects.
In our study, male Long-Evans rats demonstrated agreater initial sensitivity to ethanol than the females. Theduration of LORR was also longer in the males than in thefemales, again suggesting that males have an increasedsensitivity to ethanol. The duration of LORR is influencedby both the sensitivity of the brain to ethanol and by therate of metabolism. The outbred Long-Evans rats in ourstudy had a mean sleep time similar to that observed ininbred rat strains, but they showed slightly more variability.The mean sleep times in inbred rat strains (N/NIH) were100 � 13.8 min in males and 75.5 � 11 min in females(Deitrich and Baker, 1984). The difference in the variabilityin sleep time between the two studies may be explained bythe higher ethanol dose used in Deitrich and Baker’sstudy—3.5 g/kg—compared with the 3.0 g/kg ethanol doseused in our study.
The female and male rats in our study were young rats,almost 2 months old at the time of testing. We know thatboth age and sex affect the sensitivity of the brain toalcohol. Both older rats (Ott et al., 1985; York and Chan,1993) and humans (Dufour et al., 1992) are more impairedat similar blood concentrations than are younger rats andhumans. Therefore, we must be cautious when makingcomparisons between our results and those from previousstudies that used older rats.
Correlative analysis of ethanol-related traits in rat inbredstrains showed that there was an association among devel-opment of tolerance, initial sensitivity, and several physio-logical measures (Spuhler and Deitrich, 1984). Theyshowed that there was a significant negative correlationbetween sleep time and ethanol intake, i.e., the shortersleeping strains had a higher ethanol intake. Ethanol me-tabolism in the brain may also be affected by sex. Zimatkinet al. (2001b) have recently demonstrated that there was apositive correlation between ethanol-derived acetaldehydeaccumulation in vitro in the brain and the duration ofethanol-induced hypnosis in outbred rats. They also dem-onstrated that acetaldehyde accumulation from ethanolwas significantly greater in the brain homogenates from thebrains of animals with a high sensitivity to the hypnoticeffect of ethanol compared with animals genetically se-lected for low sensitivity (Zimatkin et al., 2001a). There-fore, the longer sleep time we observed in our male com-
Fig. 4. Combined BAC curve for all male Long-Evans rats that showed signsof LORR by 30 min (n � 29). Males were fasted for 12 hr and then given an ipinjection of 3.0 g/kg of ethanol. BAC (mg/dl) was measured as in Fig. 3. Time ismeasured in minutes after injection. BAC values were plotted against the times forRORR and 1 to 5 hr after RORR by regression. The combined BAC curvedemonstrates that there was little variability in ethanol elimination among themales (r2 � 0.881).
SEX DIFFERENCES IN ETHANOL-INDUCED HYPNOSIS AND HYPOTHERMIA 701

pared with our female rats in this study may be a result ofsex differences in ethanol metabolism.
In a previous study, we found a similar correlation be-tween shorter sleep time and ethanol intake in Long-Evansrats. In that study, females and males were given an adlibitum liquid chocolate diet containing 36% of the caloriesfrom 95% ethanol (Walker et al., 1980) for 10 weeks,followed by 39% of the calories from ethanol for an addi-tional 10 weeks (total of 20 weeks). The females hadshorter sleep times but consumed more ethanol per kilo-gram of body weight per day than did the males (Wenn etal., 2000, unpublished data).
In this study, the females and males had similar BACs atRORR and had the same metabolic and clearance rates.These results are consistent with those of Spuhler andDeitrich (1984), who reported that blood ethanol at RORRcorrelated significantly with preference and ethanol con-sumption and with metabolism and clearance; a higher rateof metabolism was correlated with a higher blood alcohollevel at RORR.
The degree of initial sensitivity to ethanol has beenproposed to constitute a factor capable of predicting sub-sequent levels of ethanol intake. Initial low sensitivity ofresponse to alcohol was associated with a high risk of futurealcoholism in men (Schuckit, 1994; Schuckit and Smith,1996) and in women (Schuckit et al., 2000). Results consis-tent with these reported in humans have also been reportedin selectively bred alcohol-preferring and -nonpreferringrats, such as the P (alcohol-preferring) and NP (alcohol-nonpreferring) (Kurtz et al., 1996) and sP (Sardinian alcohol-preferring) and sNP (Sardinian alcohol-nonpreferring) rats(Colombo et al., 2000), suggesting that the initial low sensi-tivity to alcohol also contributed to the development of highalcohol consumption in these rat lines. However, in otherselectively bred rat lines, the alcohol-preferring AA andalcohol-avoiding ANA rats, initial sensitivity to alcohol wasnot a predictor of alcohol intake (Lê and Kiianamaa, 1988).Differences in how well initial sensitivity to alcohol predictsalcohol preference in rat lines probably occurs because theserat lines have different genotypes involved in the developmentof alcohol preference.
On the basis of our results, we predicted that the females,who had a lower sensitivity to ethanol than the males,would consume more alcohol each day than the males. Aspart of an ongoing study, these same females and maleswere placed on an ethanol-containing liquid diet (the sameas previously mentioned). After 20 weeks on this diet, thefemales consumed consistently more alcohol per kilogrambody weight per day than did the males (Webb et al., 2001,unpublished data). It has been reported that alcohol intakeis correlated with the rate of alcohol elimination, althoughit seems that alcohol elimination by itself does not havedirect control over drinking. The rates of alcohol elimina-tion and voluntary alcohol consumption are strongly influ-enced by genetic factors (Kopun and Propping, 1977). Thegenetic factors involved in alcohol consumption seem to be
sex specific in Long-Evans rats. We know that geneticfactors and sex play an important role in the metabolismand behavioral actions of alcohol. Both genetic differencesand sex can influence whether or not certain levels ofalcohol produce pleasurable feelings or activation and re-duction of anxiety in some humans or animals, whereas inothers, the same levels of alcohol can have aversive effects,sedative effects, or no effect. In this study, the same level ofethanol produced greater sedative effects in the male Long-Evans rats than it did in the females. In future studies it willbe important to identify the genes responsible for the in-creased sedative affect that ethanol had in our males.
Tolerance to ethanol is a complex phenomenon, appear-ing in chronic, rapid, and acute forms that are largelydependent on the amount and schedule of ethanol expo-sure and the behavioral paradigm used to measure toler-ance (Khanna et al., 1993; Ruiz and Littleton, 1980). In thisstudy we did not directly measure BAC in the brain. Weused the duration of LORR and BACs in the tail at LORRand RORR to determine whether acute ethanol toleranceto the hypnotic effect of ethanol had occurred. We foundthat acute ethanol tolerance developed slower and in moreof the males than in the females. Although BAC was thesame in both females and males at RORR, the females hadhigher BACs at LORR. In many females, the BAC atRORR was lower than the BAC at LORR. Our datasupport the conclusion that there are sex differences in thedevelopment of acute ethanol tolerance. However, becauseLevitt et al. (1994) demonstrated that BAC measured fromthe tail vein lags behind BAC measured in arterial blood orin the jugular vein blood, we cannot be sure that our resultswould be the same if we had measured BAC in the braininstead of BAC in the tail vein at LORR and RORR.
Acquisition of acute ethanol tolerance to the intoxicatingeffects of ethanol, in humans as well as in animals, isattributed to a rapid adaptation of neuronal processes thatcauses a decreased response to the effects of ethanol and isinfluenced by both genetic and environmental factors (Er-win and Deitrich, 1996). Most of the females in this studydid not demonstrate a decreased response to ethanol, sug-gesting that neuronal adaptation had not occurred. It iswidely recognized that men and women may differ in themanner and rate of development of alcoholism and thatthese differences may have a physiological basis (Mello,1980). Perhaps failure to demonstrate neuronal adaptationto the effects of alcohol causes women to have reducedbrain size (Hommer et al., 1996), or perhaps the reducedbrain size (increased brain atrophy) influences neuronaladaptive abilities and causes women to experience greatermemory deficits over a shorter period of time and withlower alcohol consumption than men.
In addition to the BAC at LORR and the duration ofLORR, the hypothermic response to ethanol can be used asa measure of ethanol sensitivity. The initial sensitivity ofrats to hypothermic ethanol varies according to rat strainand between females and males of a particular strain. Fe-
702 WEBB ET AL.

male and male Long-Evans rats showed a similar sensitivityto the hypothermic response to ethanol, but the recoveryfrom hypothermia was slower in the females. Alcohol actscentrally, as does any other anesthetic, to abolish all ther-moregulatory control functions (Myers, 1981) and seems toexert a direct central effect on nerve cells comprising thethermoregulatory mechanism located within the anteriorhypothalamic, preoptic area (Huttunen and Myers, 1985).Therefore, it is possible that females recover more slowlyfrom ethanol-induced hypothermia because ethanol has agreater or longer-lasting effect on central thermoregulatorymechanisms.
In summary, female and male Long-Evans rats demon-strated individual and sex differences in initial responses toethanol that are similar to those reported in other ratstrains and in humans. Therefore, Long-Evans rats are anexcellent animal model for studying sex differences in ini-tial ethanol sensitivity, the hypnotic response of ethanol,and ethanol-induced hypothermia.
REFERENCES
Collins AC, Yeager TN, Lebsack ME, Panter SS (1975) Variations inalcohol metabolism: influence of sex and age. Pharmacol BiochemBehav 3:973–978.
Colombo C, Agabio R, Carai MAM, Lobina C, Pani M, Reali R, Vacca G,Gessa GL (2000) Different sensitivity to ethanol in alcohol-preferringsP and -nonpreferring sNP rats. Alcohol Clin Exp Res 24:1603–1608.
Crippens D, White ML, George MA, Jaworski JN, Brunner LJ, LancasterFE, Gonzales RA (1999) Gender differences in blood levels, but notbrain levels, of ethanol in rats. Alcohol Clin Exp Res 23:414–420.
Deitrich RA, Baker RC (1984) Initial sensitivity of rat inbred strains toacute alcohol. Alcohol Clin Exp Res 8:487–490.
Dufour M, Archer L, Gordis E (1992) Alcohol and the elderly. ClinGeriatr Med 8:127–141.
Erickson CK (1984) Ethanol clearance in nine inbred rat strains. AlcoholClin Exp Res 8:491–494.
Eriksson CJP (1973) Ethanol and acetaldehyde metabolism in rat strainsgenetically selected for their ethanol preference. Biochem Pharmacol22:2283–2292.
Erwin VG, Deitrich RA (1996) Genetic selection and characterization ofmouse lines for acute functional tolerance to ethanol. J Pharmacol ExpTher 279:1310–1317.
Fernandez JR, Vogler G, Tarantino LM (1997) Sex-specific QTL influ-ences in alcohol-related phenotypes: analysis of an F2 population. Be-hav Genet 27:589.
Frezza M, Di Padova C, Pozzato G, Terpin M, Baraona E, Lieber CS(1990) High blood alcohol levels in women: the role of decreased gastricalcohol dehydrogenase activity and first-pass metabolism. N Engl J Med322:95–99.
Hommer D, Momenan R, Rawlings R, Ragan P, Williams W, Rio D,Eckardt M (1996) Decreased corpus callosum size among alcoholicwomen. Arch Neurol 53:359–363.
Hutchins JB, Allen DL, Cole-Harding LS, Wilson JR (1981) Behavioraland physiological measures for studying ethanol dependence in mice.Pharmacol Biochem Behav 15:55–59.
Huttunen P, Myers RD (1985) Release of norepinephrine from the rat’shypothalamus perfused with alcohol: relation to body temperature.Alcohol 2:683–691.
Kalant H (1971) Absorption, diffusion, distribution, and elimination ofethanol: effects on biological membranes, in The Biology of Alcoholism(Vol 1: Biochemistry) (Kissin B, Begleiter H eds), pp 21–46. PlenumPress, New York.
Khanna JM, Shah G, Weiner J, Wu PH, Kalant H (1993) Effect of NMDAreceptor antagonists on rapid tolerance to ethanol. Eur J Pharmacol230:23–31.
Kopun M, Propping P (1977) The kinetics of ethanol absorption andelimination in twins and supplementary repetitive experiments in sin-gleton subjects. Eur J Clin Pharmacol 11:337–344.
Kurtz DL, Stewart RB, Zweifer M, Li T-K, Froehlich JC (1996) Geneticdifferences in tolerance and sensitization to the sedative/hypnotic ef-fects of alcohol. Pharmacol Biochem Behav 53:585–591.
Kwo PY, Ramachandani VA, O’Connor S, Amann D, Carr LG, Sandra-segaran K, Kopecky KK, Li T-K (1998) Gender differences in alcoholmetabolism: relationship to liver volume and effect of adjusting for bodymass. Gastroenterology 115:1552–1557.
Lancaster FE (1994) Gender differences in the brain: implications for thestudy of human alcoholism. Alcohol Clin Exp Res 18:740–746.
Lancaster FE (1995) Gender differences in animal studies: implicationsfor the study of human alcoholism, in Recent Developments in Alcohol-ism (Vol 12: Alcoholism and Women) (Galanter M ed), pp 209–215.Plenum Press, New York.
Lê AD, Kiianamaa K (1988) Characteristics of ethanol tolerance in alco-hol drinking (AA) and alcohol avoiding (ANA) rats. Psychopharmacol-ogy 94:479–483.
Levitt MD, Furne J, DeMaster EG (1994) Magnitude, origin, and impli-cations of the discrepancy between blood ethanol concentrations of tailvein and arterial blood of the rat. Alcohol Clin Exp Res 18:1237–1241.
Levitt MD, Li R, DeMaster EG (1997) Use of measurements of ethanolabsorption from stomach and intestine to assess human ethanol metab-olism. Am J Physiol 36:G951–G957.
Li T-K, Beard JD, Orr WE, Kwo PY, Ramchandani VA (1998) Genderand ethnic differences in alcohol metabolism (abstract). Alcohol ClinExp Res 22:771–772.
Lieber CS (2000) Ethnic and gender differences in ethanol metabolism.Alcohol Clin Exp Res 24:417–418.
Marshall A, Kingstone D, Boss M, Morgan M (1983) Ethanol eliminationin males and females: relationship to menstrual cycle and body compo-sition. Hepatology 3:701–706.
Mello NK (1980) Some behavioral and biological aspects of alcoholproblems in women, in Alcohol and Drug Problems in Women (Vol 5:Research Advances in Alcoholism and Drug Problems) (Kalant OJ ed), pp263–298. Plenum Press, New York.
Middaugh LD, Frackelton WF, Boggan WO, Onofrio A, Shepherd CL(1992) Gender differences in the effects of ethanol on C57BL/6 mice.Alcohol 9:257–260.
Myers AD (1981) Alcohol’s effect on body temperature: hypothermia orpoikilothermia? Brain Res Bull 7:209–220.
Ott JF, Hunter BE, Walker DW (1985) The effect of age on ethanolmetabolism and on the hypothermic and hypnotic responses to ethanolin the Fischer 344 rat. Alcohol Clin Exp Res 9:59–65.
Rivier C (1993) Female rats release more corticosterone than males inresponse to alcohol: influence of circulating sex steroids and possibleconsequences for blood alcohol levels. Alcohol Clin Exp Res 17:854–859.
Rivier C, Rivest S, Vale W (1992) Alcohol-induced inhibition of LHsecretion in intact and gonadectomized male and female rats: possiblemechanisms. Alcohol Clin Exp Res 16:935–941.
Ruiz JA, Littleton JM (1980) Glucocorticoids play an important role inthe rapid development of functional tolerance to ethanol. Subst AlcoholActions Misuse 1:415–422.
Schmidt W, Popham RE (1980) Sex differences in mortality. A compari-son of male and female alcoholics, in Alcohol and Drug Problems inWomen (Vol 5: Research Advances in Alcohol and Drug Problems)(Kalant OJ ed), pp 365–384. Plenum Press, New York.
Schuckit MA (1994) Low level of response to alcohol as a predictor offuture alcoholism. Am J Psychiatry 151:184–189.
Schuckit MA, Smith TL (1996) An 8-year follow-up of 450 sons ofalcoholic and control subjects. Arch Gen Psychiatry 53:202–210.
SEX DIFFERENCES IN ETHANOL-INDUCED HYPNOSIS AND HYPOTHERMIA 703

Schuckit MA, Smith TL, Kalmijn J, Tsuang J, Hesselbrock V, Bucholz K(2000) Response to alcohol in daughters of alcoholics: a pilot study anda comparison with sons of alcoholics. Alcohol Alcohol 35:242–248.
Seitz HK, Xu Y, Simanowski UA, Osswald B (1992) Effect of age andgender on in vivo ethanol elimination, hepatic alcohol dehydrogenaseactivity, and NAD� availability in F344 rats. Res Exp Med 192:205–212.
Spuhler K, Deitrich RA (1984) Correlative analysis of ethanol-relatedphenotypes in rat inbred strains. Alcohol Clin Exp Res 8:480–484.
Sturtevant FM (1976) Chronopharmacokinetics of ethanol. I. Review ofthe literature and theoretical considerations. Chronobiologia 3:237–262.
Taylor JL, Dolhert N, Friedman L, Mumenthaler M, Yesavage JA (1996)Alcohol elimination and simulator performance of male and femaleaviators: a preliminary report. Aviat Space Environ Med 67:407–413.
Thomasson HR (1995) Gender differences in alcohol metabolism: physi-ological response to ethanol, in Recent Developments in Alcoholism (Vol12: Alcoholism and Women) (Galanter M ed), pp 163–179. PlenumPress, New York.
Walker DW, Barnes DE, Riley JN, Hunter BE, Zornetzer SF (1980) Theneurotoxicity of chronic alcohol consumption: an animal model, in
Psychopharmacology of Ethanol (Sandler M ed), pp 17–31. Raven Press,New York.
Widmark EMP (1933) Verteilung und Umwandlung des Athylakohols imOrganism des Hundes. Biochem Z 267:128–134.
Wilkinson P (1980) Sex differences in morbidity of alcoholics, in Alcoholand Drug Problems in Women (Vol 5: Research Advances in Alcohol andDrug Problems) (Kalant OJ ed), pp 331–364. Plenum Press, New York.
Wilson JR, Erwin VG, McClearn GE (1984) Effects of Ethanol. I. Acutemetabolic tolerance and ethnic differences. Alcohol Clin Exp Res8:226–232.
York JL, Chan AW (1993) Age-related differences in sensitivity to alcoholin the rat. Alcohol Clin Exp Res 17:864–869.
Zimatkin SM, Liopo AV, Satanovskaya VI, Bardina-And LR, DeitrichRA (2001a) Relationship of brain ethanol metabolism to the hypnoticeffect of ethanol. II: Studies in selectively bred rats and mice. AlcoholClin Exp Res 25:982–988.
Zimatkin SM, Liopo AV, Slychenkov VS, Deitrich RA (2001b) Relation-ship of brain ethanol metabolism to the hypnotic effect of ethanol. I:Studies in outbred rats. Alcohol Clin Exp Res 25:976–981.
Erratum
In the January 2000 issue of Alcoholisn: Clinical and Experimental Research, the article “Quantitive Trait Loci AffectingEthanol Sensitivity in BXD Recombinant Inbred Mice,” by Kaitlin E. Browman and John C. Crabbe (volume 24, pages17–23) contained an error.
The ethanol dose actually used for the screen test was 2.5 g/kg, not the 2.0 g/kg dose stated in the article. The dosedeclared in the article for the loss or righting reflex test was correct.
704 WEBB ET AL.