semantic priming and retrieval from lexical memory: … · semantic priming and retrieval from...
TRANSCRIPT

Journal of Experimental Psychology: General1977, Vol. 106, No. 3, 226-254
Semantic Priming and Retrieval from Lexical Memory: Rolesof Inhibitionless Spreading Activation and
Limited-Capacity Attention
James H. NeelyYale University
SUMMARY
Prior to each visually presented target letter string in a speeded word-nonwordclassification task, either BIRD, BODY, BUILDING, or xxx appeared as a primingevent. When the target was a word, it was (a) a name of a type of bird on mostBiRD-prime trials; (b) a name of part of a building on most BODY-prime trials;(c) a name of a part of the body on most BUiLDiNG-prime trials; (d) a name of atype of bird, part of a building, or part of the body equally often on xxx-primetrials. Thus, on BiRD-prime trials the subject expected the word target to be chosenfrom the same category as the category represented by the word prime itself (Non-shift), whereas on BODY-prime and BuiLDiNG-prime trials the subject's attentionwas to be shifted because he or she expected the word target to be chosen from acategory other than the category represented by the word prime itself (Shift).The word target was an exemplar of either the category the subject expected (Ex-pected) or a category the subject did not expect (Unexpected) and was eithersemantically related (Related) or semantically unrelated (Unrelated) to the wordprime. Thus, there were five different types of word-prime-word-target trials: (a)BiRD-robin (Condition Nonshift-Expected-Related) ; (b) BiRD-arm (ConditionNonshift-Unexpected-Unrelated) ; (c) BODY-door (Condition Shift-Expected-Un-related) ; (d) BODY-sparrow (Condition Shift-Unexpected-Unrelated); (e) BODY-heart (Condition Shift-Unexpected-Related). The stimulus onset asynchrony(SOA) between the prime and the target letter string varied between 250 and 2,000msec. At the 2,000-msec SOA, reaction times (RTs) on BiRD-robin type trials werefaster than RTs on xxx-prime trials (a facilitation effect), whereas RTs on BIRD-arm type trials were slower than RTs on xxx-prime trials (an inhibition effect).As SOA decreased, the facilitation effect on BiRD-robin trials remained constant,but the inhibition effect on BiRu-arm trials decreased until, at the 250-msec SOA,there was no inhibition. For the Shift conditions at the 2,000-msec SOA, facilitationwas obtained on BODY-door type trials and inhibition was obtained on BODY-sparrowtype trials. These two effects decreased in magnitude as the SOA decreased until,at the 250-msec SOA, there was no facilitation or inhibition. On BODY-heart typetrials, there was an inhibition effect at the 2,000 msec SOA, which decreased asthe SOA decreased until, at the 250-msec SOA, it became a facilitation effect. Forthe nonword targets, the facilitatory effects of the word primes decreased as SOAdecreased. These results were regarded as supporting the theory of Posner andSnyder that postulates two distinct components of attention: a fast automaticinhibitionless spreading-activation process and a slow limited-capacity conscious-attention mechanism.
226

SEMANTIC PRIMING OF LEXICAL MEMORY 227
Several recent theories of informationprocessing share the common assumptionthat retrieval from long-term memory isgoverned by the operation of two distinctprocesses. One of these processes operatesautomatically, is strategy free, and occurswithout depleting the resources of a limited-capacity central processor. The second ofthese processes is an intentional, strategy-dependent mechanism that taps the limitedresources of the central processor. Althoughthe relative degree of emphasis given to eachof these two processes varies among the dif-ferent theories, this generic two-process con-ceptualization of retrieval from long-termmemory is instantiated in the recent writingsof Anderson and Bower (1973) ; Collinsand Loftus (1975) ; Keele (1973) ; LaBergeand his associates (LaBerge, 1973a, 1973b,1975; LaBerge & Samuels, 1974) ; Posnerand Snyder (1975a, 1975b); Shiffrin andhis associates (Schneider & Shiffrin, 1977;Shiffrin, 1975; Shiffrin & Geisler, 1973;Shiffrin & Schneider, 1977) ; and Turvey(1974).
A further assumption of most of thesetheories is that the automatic process playsa dominant role in the retrieval of over-learned associations and the retrieval of in-formation from well-established memory
This article is based on a dissertation submittedto Yale University in partial fulfillment of therequirements for the PhD degree. I would like toacknowledge the guidance of the members of myadvisory committee—Robert G. Crowder (Chair-man), Alice F. Healy, and Robert A. Rescorla—and readers' committee—Wendell R. Garner(Chairman) and Michael Kubovy. Special thanksare due Wendell R. Garner for providing mewith the equipment necessary for conducting theresearch. I am grateful to James C. Bartlett andDonna Sutliff for several broad-ranging discus-sions of the theoretical issues raised by this re-search and to Carolyn Starr, Virginia Locke, JimBorzym, Alan Stahura, Nora Keane, GeorgeNobel, and Gail Grant, who assisted in the colla-tion of the data. I also wish to thank two anony-mous reviewers and Randall Engle for severalhelpful suggestions that led to a clearer expositionof the concepts and results presented in the paper.
Requests for reprints should be sent to JamesH. Neely, who is now at the Department of Psy-chology, University of South Carolina, Columbia,South Carolina 29208.
structures, whereas the limited-capacity cen-tral processor plays a dominant role in theretrieval of less well learned information.(Such an assumption has also been centralto earlier theories of skill acquisition, e.g.,Fitts, 1964.) Evidence for this assumptionhas been obtained in experiments reportedby LaBerge (1973a, 1975), Schneider &Shiffrin (1977), and Shiffrin & Schneider(1977). These experiments have shown thata mental operation that initially demandsattention becomes automated with extendedtraining.
Of those two-process theories mentionedabove that are particularly relevant to thepresent experiments, Posner and Snyder's(1975a) theory of attention is one of themore explicit ones in terms of specifying theproperties of each of the two processes andhow they operate in isolation and in con-junction with each other. In their theory, anautomatic spreading-activation process and alimited-capacity attentional mechanism areinvolved in the retrieval of information fromlong-term memory. Posner and Snyder sub-scribe to Morton's (1970) view that verballong-term memory contains logogens—mem-ory structures containing information storedabout events with which a person has hadconsiderable experience. These logogens arefed by banks of visual and auditory featuredetectors, and they are activated wheneverthe number of features feeding into themexceeds some threshold value. In terms ofa spatial metaphor, the logogens for seman-tically related words are assumed to belocated nearer one another in long-termmemory than are the logogens for semanti-cally unrelated words.
According to Posner and Snyder's(1975a) theoretical treatment of the spread-ing-activation process, a stimulus automati-cally activates its logogen, and this activa-tion automatically spreads to adjacent,semantically related logogens but not to re-mote, semantically unrelated logogens. Thisautomatic spreading-activation process hasthree major properties: (a) It is fast acting;(b) it can occur without intention or con-scious awareness; and (c) it does not affectthe retrieval of information stored in seman-

228 JAMES H. NEELY
tically unrelated logogens to which it has notspread. Thus, if a word that is to be pro-cessed is presented before there has been acomplete decay of the logogen activationproduced by a previously presented seman-tically related word, the activation level inthe logogen of the to-be-processed word willinitially be higher (due to the activation thathas spread to it) than it would have beenhad it been preceded by a semantically un-related word, and the result will be a facili-tation in its processing.
The limited-capacity conscious-attentionmechanism of the Posner-Snyder (1975a)theory can also produce a facilitation inprocessing, with the facilitation occurringfor those stimuli that activate logogens uponwhich attention is being focused. However,the three major properties of the limited-capacity attentional mechanism are oppositeto the three major properties of the auto-matic spreading-activation process. That is,the limited-capacity attentional mechanism(a) is slow acting; (b) cannot operate with-out intention and conscious awareness, and(c) inhibits the retrieval of informationstored in semantically unrelated logogensupon which it is not focused. Indeed, oneof the most crucial components of the Pos-ner-Snyder (197Sa) theory is the mannerin which the conscious-attention mechanismproduces inhibition in the retrieval of in-formation from unattended logogens. Ac-cording to their theory, the conscious-atten-tion mechanism does not inhibit the buildupof activation in unattended logogens butrather inhibits the readout of informationfrom unattended logogens. That is, if a to-be-processed stimulus activates a logogenthat is not contained in the subset of logo-gens upon which attention is focused, thebuildup of activation in this unattended logo-gen proceeds in the same manner it wouldhave if attention had not been misdirectedto other logogens; however, before the in-formation stored at the unattended logogencan be analyzed in preparation for responseinitiation, the conscious-attention readoutmechanism must be "shifted" to that logo-gen. Thus, in terms of the operation of theconscious-attention mechanism, a to-be-
processed word will be processed fasterwhen it follows a semantically related wordthan when it follows a semantically unre-lated word because the "distance" that at-tention must be shifted is shorter when at-tention is being shifted between semanticallyrelated logogens than between semanticallyunrelated logogens.
To evaluate their two-factor theory ofattention, Posner and Snyder (197Sb) con-ducted a letter-matching experiment and ananimal-name classification experiment inwhich the target array about which the sub-ject was to decide was preceded by a prim-ing event to which the subject made noovert response. The subject's conscious at-tention was controlled by variations in theprobability that the priming cue itself wouldappear as an element in the target array, andthe prior automatic activation of the targetelement was controlled by the presence orabsence of that particular target element asa priming cue. A baseline for assessing theconscious-attention and automatic-activationeffects produced by the priming cue was pro-vided by a neutral warning-signal primethat did not direct the conscious-attentionreadout mechanism to a specific set of logo-gens nor automatically activate any of thelogogens activated by the stimulus elementsof the target array. If, in comparison to thereaction times (RTs) to respond to thetarget array when it followed this neutralwarning-signal prime, the RTs were shorterfollowing a priming cue, the priming cuewas said to have facilitated the processing ofthe target array; if RTs were longer fol-lowing the priming cue, the priming cue wassaid to have inhibited the processing of thetarget array. The stimulus onset asynchrony(SOA) between the priming event and thetarget array was varied between 10 and 500msec to test if the facilitation produced bythe fast automatic-activation process wouldoccur at shorter SOAs than would the in-hibition produced by the more deliberatecommitment of conscious attention.
The data in the Posner and Snyder(1975b) experiments to be considered herefirst are for trials in which the subject wasto respond "yes"—that is, for trials in which

SEMANTIC PRIMING OF LEXICAL MEMORY 229
the letters in the target array matched inthe letter-matching task and for trials inwhich an animal name appeared in the targetarray in the animal-name classification task.For the 500-msec SOA, when the primingcue appeared in the target array, the pro-cessing of the target array was facilitatedregardless of whether the probability thatthe priming cue would appear in the targetarray was high or low; but when the prim-ing cue did not appear in the target array,the processing of the target array was in-hibited only when the probability that thepriming cue would appear in the target ar-ray was high enough that the subject's at-tention would have been misdirected to thepriming cue's logogen when the target arrayappeared. Thus, as predicted, facilitation oc-curred automatically, regardless of whetherthe subject was consciously using the prim-ing cue to direct his or her attention to thelogogen for a target element, whereas in-hibition occurred only when the subject wasmisdirecting attention in the high-probabilitycondition. Also, as predicted, the facilitationoccurred sooner following the presentationof the priming cue than did the inhibition.
Unfortunately, the interpretation of thedata from the yes trials was complicated bythe data for no trials—that is, for trials inwhich the letters in the target array did notmatch in the letter-matching task and fortrials in which an animal name did notappear in the target array in the animal-name classification task. When the primedid not appear in the target array for theno trials in the high-probability condition,the prime did not inhibit the processing ofthe target array, as would be predicted byan unembellished application of the Posner-Snyder (1975a) theory, but rather facili-tated the processing of the target array. Pos-ner and Snyder (1975b) suggested that thisfacilitation occurred because of the subject'stendency to respond "no" when the primewas not physically identical to any of theelements in the target array. Therefore, itwas unclear whether the inhibition effect forthe yes trials was being produced by a gen-eral inhibition associated with misdirected
attention or by a specific physical-matchstrategy. Posner and Snyder (1975a, 1975b)therefore acknowledged that their theorywould be more strongly supported if an in-hibition effect were obtained in an experi-ment in which the priming cue and the tar-get elements were not physically identical.
Of particular relevance to the present re-search is the manner in which the Posner-Snyder (1975a) theory accounts for a find-ing commonly obtained in experiments in-volving a lexical decision task, a task inwhich the subject must decide as quickly aspossible whether a visually presented letterstring is a common English word or a non-word. The commonly obtained finding insuch experiments (e.g., Meyer & Schvane-veldt, 1971; Meyer, Schvaneveldt, & Ruddy,Note 1) is that subjects are quicker to re-spond that a target letter string (e.g.,NURSE) is a word when the immediatelyprior target was a semantically related word(i.e., DOCTOR) than when it was a semanti-cally unrelated word (e.g., BREAD). Asnoted earlier, each of the two mechanisms ofthe Posner-Snyder (1975a) theory can ac-count for such a semantic-facilitation effect.Responses to NURSE would be faster follow-ing DOCTOR than following BREAD because ac-tivation would have automatically spreadfrom the logogen for DOCTOR to the logogenfor NURSE and because the distance thatattention must traverse in shifting from thelogogen for DOCTOR to the logogen for NURSEis less than the distance that it must tra-verse in shifting from the logogen for BREADto the logogen for NURSE.
As Posner and Snyder (1975a) havepointed out, the automatic spreading-activa-tion and limited-capacity attentional mecha-nisms of their theory correspond ratherclosely to the spreading-excitation and lo-cation-shifting models proposed by Schvane-veldt and Meyer (1973) to account for thesemantic-facilitation effect in the lexical de-cision task. On the basis of data obtainedin a series of experiments designed to dis-tinguish between these two models (Schva-neveldt & Meyer, 1973; Meyer et al., Note1), Schvaneveldt and Meyer (1973) con-cluded that the semantic-facilitation effect in

230 JAMES H. NEELY
the lexical decision task was to -be attributedto the spreading-activation process and wasnot to be attributed to the operation of thelimited-capacity attentional mechanism. Sincethe Posner-Snyder (1975a) theory assumesthat spreading-activation and limited-ca-pacity attention are complementary ratherthan mutually exclusive mechanisms, Posnerand Snyder (1975a) have argued that al-though the Meyer et al. and Schvaneveldtand Meyer data do indeed provide evidencefor the involvement of spreading activationin the semantic-facilitation effect, they donot rule out an involvement of the limited-capacity conscious-attention mechanism.
In an experiment designed to evaluatefurther the role of the limited-capacity atten-tional mechanism in the semantic-facilitationeffect, Neely (1976) adopted the method-ology of Posner and Snyder (1975b). Thetarget word to which the subject was tomake a lexical decision was preceded by aneutral warning signal on some trials and byeither a semantically related word prime ora semantically unrelated word prime onother trials. The RTs to word targets weremuch faster when the prime was a seman-tically related word than when the primewas a semantically unrelated word, thus rep-licating the semantic-facilitation effect. Moreimportantly, in comparison to when a wordtarget followed the neutral warning-signalprime, there was an inhibition in the proc-essing of a word target when it followed asemantically unrelated word prime and fa-cilitation in its processing when it followeda semantically related word prime. Withinthe framework of the Posner-Snyder(1975a) theory, the fact that a semanticallyunrelated word prime produced inhibitionsuggests that limited-capacity attention isinvolved in the semantic-facilitation effect.
Unfortunately, two other results of theNeely (1976) experiment raised interpre-tive problems. First of all, the magnitudeof the inhibitory effect of the semanticallyunrelated word prime remained constantover the three SO As (360, 600, and 2,000msec), whereas the facilitatory effect of thesemantically related word prime was greaterat the two longer SOAs than at the shortest
SOA. This result is problematic because,contrary to the Posner-Snyder (1975a)theory, the inhibition did not build up asthe SOA increased. The second result thatargued against limited-capacity attention asthe source of the inhibition produced by thesemantically unrelated word primes was thatthe processing of nonword targets was facili-tated by the word primes. If the primingword were indeed depleting some of the sub-ject's limited-capacity attention, there shouldhave been an inhibition effect for the non-word targets as well as for the semanticallyunrelated word targets. Neely (1976) arguedthat because of the structure of the experi-ment and the highly stereotyped primary-associative relationship between the primingwords and the semantically related targetwords (e.g., CAT-dog), subjects may haveadopted the strategy of generating a primaryassociate to the priming word (e.g., gener-ating, as the candidate for the target, "dog"to the priming word CAT), matching thisself-generated item with the target item andthen tending to respond "word" when thetwo items were the same and tending to re-spond "nonword" when the two items weredifferent. Such a strategy would have slowedRTs to semantically unrelated words andsped RTs to nonwords.
The purpose of the present research wasto provide a more rigorous test of the basicassumptions underlying the Posner-Snyder(197Sa) theory by elaborating on the Neely(1976) paradigm so as to permit a furtherclarification of the roles of limited-capacityattention and automatic spreading activationin the lexical decision task. There werethree important features of the present ex-periment's design. First, the presence orabsence of automatic spreading activationwas controlled by whether the prime andthe target were semantically related or se-mantically unrelated. Second, the presenceor absence of the operation of limited-ca-pacity attention was controlled by whetherthe prime and target were separated by along SOA, in which case the subject wouldbe given enough time to engage, focus, andcommit limited-capacity attention, or a shortSOA, in which case the subject would not

SEMANTIC PRIMING OF LEXICAL MEMORY 231
be given enough time to engage, focus, andcommit attention.
The third, and perhaps most important,feature of the present experiment is that itavoids an as yet unmentioned confoundingthat has occurred in previous semantic-priming experiments (e.g., Neely, 1976;Posner & Snyder, 1975b; Rosch, 1975).Since these previous experiments were de-signed so that the priming event that di-rected the subject's conscious attention tothe target's logogen was an event (i.e., thetarget item itself or a word semantically re-lated to the target word) that should havealso produced automatic activation in thetarget's logogen, these experiments haveconfounded the facilitatory effects of con-scious attention with the facilitatory effectsof automatic spreading activation. Thus, itis unclear how much of the facilitation ob-tained in these experiments was being pro-duced by the subject's having directed hisor her conscious attention to the target logo-gen and how much of the facilitation wasbeing produced by activation having auto-matically spread to the target logogen. Ofcourse, within the framework of the Posner-Snyder (197Sa) theory, a determination ofthe relative contribution of conscious at-tention to the facilitation effect can be madeby assessing the amount of inhibition ac-companying the facilitation effect. However,one need not resort to such an indirect mea-sure, since the two types of facilitation canbe unconfounded. The confounding isavoided in the present experiment 'by usinga priming word semantically unrelated tothe target word to direct the subject's at-tention to the area of lexical memory con-taining the logogen for the target word.As will become clear in the explication of thepresent experiment's complete design, somenovel observations of potential theoreticalimportance are occasioned when automatic-activation and conscious-attention facilitationeffects are unconfounded in this manner.
Design and Theoretical Predictions
Prior to each target letter string aboutwhich the subject was to make a lexical de-
cision, the subject saw as a priming eventxxx or the priming word BIRD, BODY, orBUILDING. The subjects were instructed asfollows:
When the priming word is BIRD, if the target thatfollows it is a word, it will be the name of atype of bird on most trials. However, when thepriming word is BODY, rather than expecting wordsthat are the names of parts of the human body,you should shift your attention to expect that ifthe target is a word it will be the name of some-thing to do with a building. In fact, when BODYis the priming event, if the target that follows itis a word, it will be the name of something to dowith a building on most trials. Likewise, when thepriming word is BUILDING, rather than expectingwords that are the names of parts of a building,you should shift your attention to expect that if thetarget is a word it will be the name of a part ofthe human body. In fact, when BUILDING is thepriming event, if the target that follows it is aword, it will be the name of a part of the humanbody on most trials. When xxx is the primingevent, if the target is a word it will equally oftenbe chosen from the three semantic categories bird,body, and building.
In accord with these instructions, whena word target followed the BIRD prime it wasa name of a type of bird on two thirds ofthe trials, a name of a part of the body onone sixth of the trials, and a name of a partof a building on one sixth of the trials. Whena word target followed the BODY prime itwas a name of a part of a building on twothirds of the trials, a name of a type of birdon one sixth of the trials, and a name of apart of the body on one sixth of the trials.When a word target followed the BUILDINGprime it was a name of a part of the body ontwo thirds of the trials, a name of a type ofbird on one sixth of the trials, and a name ofa part of a building on one sixth of the trials.
Three main variables are being manipu-lated by these instructions and these word-target trial types. The first variable iswhether the word prime is cuing the subjectto focus attention on exemplars of the cate-gory represented by the word prime itself,as was the case for the BIRD prime (Non-shift), or cuing the subject to shift and focusattention on exemplars of a category otherthan the category represented by theword prime itself, as was the case for theBODY and BUILDING primes (Shift). (Since,
232 JAMES H. NEELY
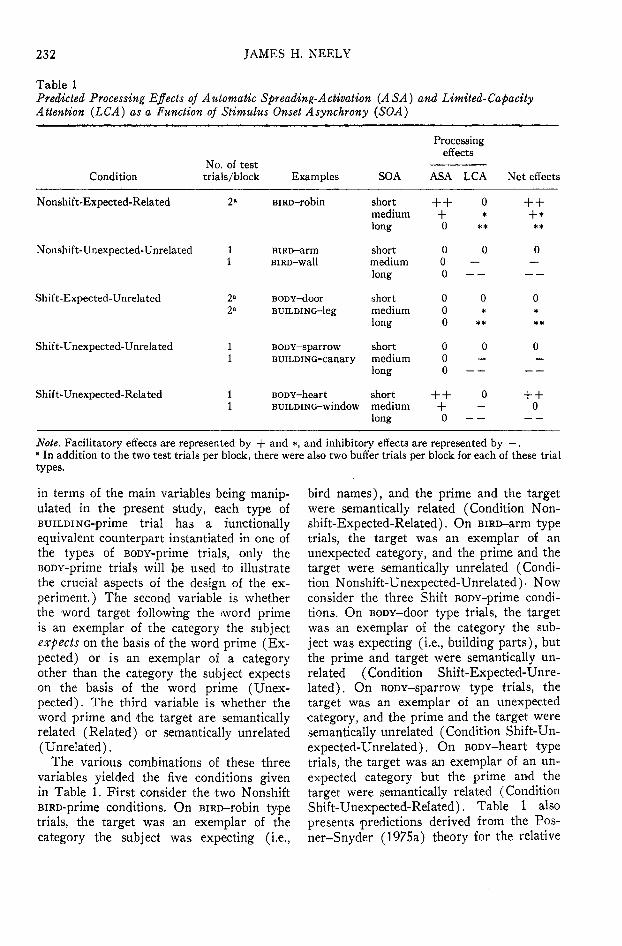
Table 1Predicted Processing Effects of Automatic Spreading-Activation (ASA) and Limited-CapacityAttention (LCA) as a Function of Stimulus Onset A synchrony (SOA)
Condition
Nonshift-Expected- Related
Nonshift-Unexpected-Unrelated
Shift-Expected-Unrelated
Shift-Unexpected-Unrelated
Shift-Unexpected-Related
No. of testtrials/block
2"
11
2"2"
11
11
Processingeffects
Examples
BiRD-robin
BiRD-armBIRD-Wall
BODY-doorBUILDING-leg
BODY-sparrowBUiLDiNG-canary
BODY-heartBUiLDiNG-window
SOA
shortmediumlong
shortmediumlong
shortmediumlong
shortmediumlong
shortmediumlong
ASA
+ ++0
000
000
000
+++0
LCA
0*
**
0—
0*
**
0—
0—
Net effects
+ ++ ***
0—
0***
0—
++0
Note. Facilitatory effects are represented by + and *, and inhibitory effects are represented by — ." In addition to the two test trials per block, there were also two buffer trials per block for each of these trialtypes.
in terms of the main variables being manip-ulated in the present study, each type ofBuiLDiNG-prime trial has a functionallyequivalent counterpart instantiated in one ofthe types of BODY-prime trials, only theBODY-prime trials will be used to illustratethe crucial aspects of the design of the ex-periment.) The second variable is whetherthe word target following the word primeis an exemplar of the category the subjectexpects on the basis of the word prime (Ex-pected) or is an exemplar of a categoryother than the category the subject expectson the basis of the word prime (Unex-pected). The third variable is whether theword prime and the target are semanticallyrelated (Related) or semantically unrelated(Unrelated).
The various combinations of these threevariables yielded the five conditions givenin Table 1. First consider the two NonshiftBiRD-prime conditions. On BiRD-robin typetrials, the target was an exemplar of thecategory the subject was expecting (i.e.,
bird names), and the prime and the targetwere semantically related (Condition Non-shift- Expected-Related). On BiRD-arm typetrials, the target was an exemplar of anunexpected category, and the prime and thetarget were semantically unrelated (Condi-tion Nonshift-Unexpected-Unrelated). Nowconsider the three Shift BODY-prime condi-tions. On BODY-door type trials, the targetwas an exemplar of the category the sub-ject was expecting (i.e., building parts), butthe prime and target were semantically un-related (Condition Shift-Expected-Unre-lated). On BODY-sparrow type trials, thetarget was an exemplar of an unexpectedcategory, and the prime and the target weresemantically unrelated (Condition Shift-Un-expected-Unrelated). On BODY-heart typetrials, the target was an exemplar of an un-expected category but the prime and thetarget were semantically related (ConditionShift-Unexpected-Related). Table 1 alsopresents predictions derived from the Pos-ner-Snyder (1975a) theory for the relative

SEMANTIC PRIMING OF LEXICAL MEMORY 233
amounts of facilitation and inhibition pro-duced by the automatic spreading-activationprocess and the limited-capacity attentionalmechanism in each of these five word-targetconditions as a function of SO A. To clarifythe predictions shown in Table 1 it will benecessary to enumerate the assumptions un-derlying them.
The assumptions underlying the predic-tions for the automatic spreading-activationeffects were as follows:
1. Automatic spreading activation has aneffect only when the prime and the targetare semantically related. Thus, zeroes ap-pear for all SOAs for all of the Unrelatedconditions.
2. Automatic spreading activation pro-duces only facilitatory effects and no inhibi-tory effects. Thus, all nonzero effects are +•
3. Automatic spreading activation decaysunless the subject makes a conscious effortto maintain it via rehearsal. Thus, the facili-tatory effect of automatic spreading activa-tion decreases as SO A increases. For pres-ent purposes, it will be assumed that thelong SOA is sufficiently long that there willhave been a complete decay of automaticspreading activation. Evidence supportingthe assumption that automatic spreading ac-tivation decays has been obtained in ex-periments by Loftus (1973), Meyer et al.(Note 1), and Warren (1972).
The assumptions underlying the predic-tions for the effects of limited-capacity at-tention were as follows:
4. Limited-capacity attention produces afacilitatory effect (designated as * ratherthan + to distinguish it from the facilitatoryeffect produced by automatic spreading ac-tivation) when the target is an exemplarof the category the subject expects, and itproduces an inhibitory effect when the targetis an exemplar of an unexpected category.Thus, *s are present in the two Expectedconditions and — s are present in the threeUnexpected conditions.
5. The effects of limited-capacity atten-tion increase with increasing SOA. Forpresent purposes, it will be assumed thatthe short SOA is sufficiently short that lim-ited-capacity attention has not yet been com-
j1
.NS-Ex-RyiBIRD-robin)
\ S-Ex-U\ (BODY- door)
S-Ux-l(BODY-span
(BIRD-arm)
Short Medium
SOALong
Figure 1. Amount of facilitation (+) or inhibi-tion ( —) predicted for word targets in word-prime conditions as a function of stimulus onsetasynchrony (SOA). (NS-Ex-R= Nonshift-Ex-pected-Related; NS-Ux-U = Nonshift-Unexpected-Unrelated; S-Ex-U = Shift-Expected-Unrelated;S-Ux-R = Shift-Unexpected-Related; S-Ux-U =Shift-Unexpected-Unrelated.)
mitted. Thus, there is a zero entry in thelimited-capacity attention column for all con-ditions at the short SOA.
The following assumptions were made tocompute the net effects:
6. The facilitatory effects produced byautomatic spreading activation and limited-capacity attention are additive. To simplifymatters, it will be assumed that + = *.
7. The facilitatory effects of automaticspreading activation and the inhibitory ef-fects of limited-capacity attention are addi-tive. To further simplify matters, it will beassumed that + and — exactly cancel eachother out in Condition Shift-Unexpected-Related at the intermediate SOA such thatthe net effect is zero.
Figure 1 presents a graphic display ofthe net effects predicted by the Posner-Sny-der (1975a) theory in conjunction with thesimplifying assumptions mentioned above.To permit a direct comparison of these pre-dictions with the obtained results, the short,medium, and long SOAs are given thevalues used in the experiment.
Of course, to derive predictions for RTs,

234 JAMES H. NEELY
additional assumptions will need to be madeconcerning the mapping of various magni-tudes of the hidden underlying facilitatoryand inhibitory processes into the RTs theexperimenter observes. Two further assump-tions were made in this regard:
8. The mapping of underlying processinto RTs is some unspecified monotonicfunction.
9. The scales underlying the mapping offacilitation into RTs and of inhibition intoRTs are not necessarily comparable. Thatis, when a given prime (e.g., BODY) pro-duces facilitation only through the operationof limited-capacity attention (as in Condi-tion Shift-Expected-Unrelated), the amountof observed facilitation measured in milli-seconds for that prime will not necessarilybe equal to the amount of observed limited-capacity attentional inhibition measured inmilliseconds for that prime (i.e., in Condi-tion Shift-Unexpected-Unrelated). How-ever, under such circumstances, if the purelimited-capacity attentional facilitatory ef-fect for a given prime is statistically sig-nificant or nonsignificant, the inhibitory ef-fect for that prime should likewise be statis-tically significant or nonsignificant, and viceversa.
Given these two assumptions, the mostimportant aspect of Figure 1 is the patternof the facilitation and inhibition effects pre-dicted for the various conditions and nottheir absolute magnitudes.
The five most important aspects of thepredicted pattern of results shown in Figure1 are the following:
1. The facilitation effect in ConditionNonshift-Expected-Related (BiRD-robin) isflat as a function of SOA, whereas the fa-cilitation effect in Condition Shift-Expected-Unrelated (soDY-door) starts at zero andincreases with increasing SOA until it con-verges with the facilitation effect obtainedfor the BiRD-robin trials. Of course, the factthat the facilitation effect for BODY-doortrials follows a different time course thanthe facilitation effect for BiRD-robin trialsdoes not necessarily mean that spreadingactivation is not involved in the facilitationeffect obtained for BODY-door trials. When
BODY is the prime, the subject most prob-ably uses the attentional mechanism to re-trieve the BODY-"building" association, thusactivating the "building" logogen. This ac-tivation would begin to spread to logogenssemantically related to "building", such as"door." Once the "building" logogen hasbeen activated via retrieval of the unauto-mated BODY-"building" association, the sub-ject probably uses the attentional mechanismto maintain activation in the "building" lo-gogen (and in the "bird" logogen followingthe BIRD prime) so that it does not decayduring the 2,000-msec SOA. However, theimportant point is that the activation in the"building" logogen would not have auto-matically occurred when BODY was presentedbecause the BODY-"building" association isnot a well-learned, automated associationlike the BiRD-"bird" association.
2. The inhibition effects in ConditionsNonshift-Unexpected-Unrelated (BiRD-arm)and Shift-Unexpected-Unrelated (BODY-sparrow) are equivalent, starting at zero atthe short SOA and growing in magnitudewith increasing SOA.
3. The BIRD prime produces a facilitationeffect for BiRD-robin trials at the short SOA,but this facilitation is not accompanied byan inhibition effect for BiRD-arm trials at theshort SOA. On the other hand, the BODYprime produces a facilitation effect for BODY-door trials only if at the same time it is pro-ducing an inhibition effect for BODY-sparrowtrials. Facilitation in BiRD-robin trials is notnecessarily accompanied by inhibition inBiRD-arm trials because the BiRD-robin fa-cilitation is based on an overlearned, auto-mated association. However, facilitation inBODY-door trials is always accompanied byinhibition in BODY-sparrow trials becausethe BODY-door facilitation is not based on anoverlearned, automated association andtherefore must involve the commitment oflimited-capacity attention.
4. Perhaps the most interesting predictionmade by the Posner-Snyder (1975a) theoryis that in Condition Shift-Unexpected-Re-lated (soDY-heart), there should be a netfacilitation effect at the short SOA becauseof activation automatically spreading from

SEMANTIC PRIMING OF LEXICAL MEMORY 235
the "body" logogen to the semantically re-lated "heart" logogen and because there hasnot been enough time allowed by the shortSOA for limited-capacity attention to beginexerting its counteracting inhibitory influ-ence. However, as the SOA increases, theautomatic spreading activation decays, andthe limited-capacity attention mechanism be-gins to exert its inhibitory influence, so thatthe net effect is inhibitory at the long SOA.
5. The final aspect to note about the pre-dictions shown in Figure 1 is that the curvefor the BODY-heart trials is far above thecurve for the BODY-sparrow trials at theshort SOA because of the facilitation effectproduced by automatic spreading activationdue to the close semantic relationship be-tween BODY and heart. However, as SOA in-creases and automatic spreading activationdecays because attention is shifted to the"building" logogen, the curve for BODY-hearttrials converges with the curve for BODY-sparrow trials.
To the degree that the data obtained forthe word-prime-word-target conditions cor-respond to the five major predictions of thePosner-Snyder (1975a) theory displayed inFigure 1 and outlined above, the basic as-sumptions of the Po-sner-Snyder theory willbe supported. Any discrepancies between thepredicted results shown in Figure 1 and theobtained results will call into question thebasic assumptions of the Posner-Snydertheory and will demand that these assump-tions be abandoned or modified to accountfor such discrepancies.
Method
Stimulus Materials
So that enough observations per subject percondition could be obtained without requiring thata word target be repeated in the experiment, wordtargets were chosen from the exemplars of threelarge semantic categories given in the Battig andMontague (1969) norms—birds, parts of the body,and parts of a building. The pronounceable non-word targets were matched with the word targetsby changing one letter in a word matched to eachof the word targets on the basis of frequency ofoccurrence in the language, number of letters, andnumber of syllables. Nonword targets were formedin this manner because James (1975) has arguedthat subjects must access the meaning of a word
target in order to make a lexical decision when-ever they cannot classify words and nonwords onthe basis of structural differences. The set of wordtargets from each semantic category was split intotwo sets of equal size according to category domi-nance in the Battig and Montague norms: a high-category-dominance (HCD) set and a low-cate-gory-dominance (LCD) set. Since the nonwordtargets were matched with the word targets, theytoo were distinguished by their category member-ship and category dominance, but for the nonwordtargets these are pseudodistinctions unless theyare confounded with the structural characteristicsthat formed the basis for the matching of theword and nonword targets (cf. Landauer &Streeter, 1973).
Test-List Construction
To insure that the results could not be attributedto differences in the categories used in a givencondition, a base list was constructed for each ofthree different category-assignment (CA) sets.Across the three CA sets, each of the primingwords occurred in the nonshift condition, and eachpriming word cued each of the two remainingsemantic categories when it occurred in a Shiftcondition. A base list consisted of six blocks of60 trials: 30 word-target trials (10 from eachcategory) and their 30 matched nonword-targettrials. There were 36 word-prime trials per block,with each of the three word primes appearing in12 trials, and 24 xxx-prime trials. Twelve of the36 word-prime trials were buffer trials: 6 word-target trials and their matching set of 6 nonword-target trials. As noted in Table 1, the 6 word-prime-word-target buffer trials represented in-stances of the two different Expected conditions.A block always began with 2 buffer trials, withthe remaining 10 buffer trials being randomlyscattered throughout the rest of the trials in thatblock.
In addition to the 12 word-prime buffer trials,each block of 60 trials contained 48 test trialsfrom which the data to be reported here weretaken. Twenty-four of these trials were word-prime trials (12 word-target trials and their 12matched nonword-target trials) and 24 were xxx-prime trials (12 word-target trials and their 12matched nonword-target trials). The constitutionof the 12 word-prime-word-target test trials foreach block is represented in Table 1. It can beseen that in each block Condition Shift-Expected-Unrelated was represented in 4 trials, whereasthe remaining four conditions were represented in2 trials each. As a consequence of the nonword-target trials being matched with the word-targettrials, there were 8 Shift trials and 4 Nonshifttrials for the 12 word-prime-nonword-target testtrials. For the 12 xxx-prime-word-target testtrials, 4 word targets were chosen from each of thethree semantic categories, and their matching non-word targets were used for the 12 xxx-prime-

236 JAMES H. NEELY
nonword-target test trials. Across blocks, wordtargets and nonword targets from each of thethree categories were equally divided betweenHCD and LCD items within each condition.Within and across blocks, the presentation orderof the target items was a pseudorandom sequencewith the first-order sequential probabilities for con-dition, target lexicality, and category membershipbeing equated as nearly as possible.
Seven other lists were derived from each ofthe base lists for each of the three different CAsets by reordering the priming events for the testtrials so that the order of presentation of thetarget events was identical for all lists. Thus,altogether there were 24 different lists, that is, 8lists X 3 CA sets. Across these 24 lists, the neteffect for the test trials was that each wordtarget and each nonword .target appeared equallyoften following a given word prime and the xxxprime, and each word-prime condition involved thethree different word primes BIRD, BODY, and BUILD-ING. Thus, none of the theoretically crucial dif-ferences between the various priming conditionscan be attributed to item effects.
Practice-List Construction
The targets in both of the two practice listswere words on half of the trials and nonwords onthe other half of the trials. For the 4(Ktrial firstpractice list, there were two different primingevents, ANIMAL and xxx, and the target wordswere selected from the Battig and Montague(1969) categories animals and metals. The ANIMALand xxx primes were both equally often followedby word targets and nonword targets, but the wordtarget following the ANIMAL prime was always ananimal name, whereas the word target followingthe xxx prime was an animal name on half ofthe trials and a metal name on the other half ofthe trials. The first practice list was constructedin this manner so that the subject would learnthat the word prime provides information aboutthe semantic nature of the word target, whereasthe xxx prime does not provide such information,and that neither the word prime nor the xxxprime provides information as to the lexicality ofthe target that follows it.
The stimulus materials for the 36-trial secondpractice list were comparable to those of the testlist. However, only word primes appeared, witheach of the three word primes being presentedequally often and always being followed by aword target chosen from the expected category onword-target trials.
Subjects
One hundred twenty-two native English speakersin the Yale community were individually testedand received either experimental credits for anintroductory psychology course or $4 each fortheir participation in the 14-hr, session. The data
from two subjects were discarded because one ofthem failed to respond on several trials and theother could not describe midway through the ses-sion the attention-shifting strategy she was to beadopting.
Procedure
General instructions were read to all subjectsprior to the first practice list. Each trial beganwith the subject pressing down with his dominanthand a home-base key between the two responsekeys. The response key on the subject's dominantside and nondominant side was always assigned tothe "word" and "nonword" response, respectively.Each trial consisted of two slides, the primingslide and the target slide, successively rear pro-jected on translucent Mylar near the center of a12 X 16 cm aperture. The uppercase priming eventand the lowercase target event appeared in identi-cal positions in the aperture. The priming slidewas exposed for ISO msec, and the SOA wasvaried according to the duration of the dark blankinterval between the offset of the priming slideand the onset of the target slide, which was dis-played for 1,000 msec for all trials. During the4-sec inter.trial interval, the experimenter informedthe subject if an error had been made on theprevious trial. The exposure durations of the primeand the target slides were controlled by Lafayetteshutters and BRS/LVE Digi-Bit solid-statetimers. The RTs were measured to the nearestmillisecond.
Before the second practice list, the three differ-ent instructional sets appropriate to the three CAsets were introduced. In addition to informing thesubject explicitly about the attention-shifting strat-egies that should be adopted with each prime, theseinstructions emphasized how important it was forthe subject to attempt to adopt these strategies onevery trial. The SOA was always 2,000 msec forthe first two practice lists.
Following the second practice list, the SOAvaried between 250 and 2,000 msec both betweenand within subjects. In the 2,000-alone group, theSOA was held constant at 2,000 msec for alltrials. Each of the 24 subjects in the 2,000-alonegroup received a different one of the 24 lists—8lists X 3 CA sets—described above. For the three2,000-mixed groups, the 2,000-msec SOA was ran-domly intermixed with a shorter SOA. Theshorter SOAs were 250, 400, and 700 msec forGroups 250-2,000, 400-2,000, and 700-2,000, respec-tively. For the three 2,000-mixed groups, the shortand long SOAs were superimposed on the stimulussequence with the constraint that within each blockeach condition would occur with a short and longSOA and that across blocks both HCD and LCDitems would equally often be represented withineach condition at the short and long SOAs. Aresult of these constraints was that a short or longSOA followed itself more often (around 55% ofthe time) than it followed the opposite SOA. Since

SEMANTIC PRIMING OF LEXICAL MEMORY 237
preliminary analyses of the data from the 2,000-alone group indicated that the amount of facilita-tion or inhibition seen in a particular conditionwas not substantially affected by CA set, subjectsin the 2,000-mixed groups were exposed to listsfrom only two of the three CA sets, and a givenlist within a CA set was used twice, so that aparticular target appeared in a given condition atboth SOAs. Thus, each of the 32 subjects in eachof the 2,000-mixed groups received a different oneof 32 lists: 8 lists X 2 CA sets X 2 SOA orders.
A third practice list, a repetition of the second,was given to the three 2,000-mixed groups but notto the 2,000-alone group, in order to introduce thesubjects to the two SOAs they would be receivingin the test series. The long SOA was given onhalf of the trials and the short SOA was givenon the remaining half of the trials, with eachpriming event and the word and nonword targetsoccurring equally often at the long and shortSOAs. The subjects were told that even thoughit might be difficult to shift their attention duringthe short interval, it was extremely important thatthey make every effort to do so and that, sincethe order of presentation of the long and shortintervals would be randomly determined, theyshould not try to anticipate what the interval fora given trial would be, and they should try tostart shifting their attention as soon as the prim-ing event was shown.
The test series followed either the third practicelist for the 2,000 mixed groups or the second prac-tice list for the 2,000-alone group. The subjectswere told that on a very few trials they would re-ceive a word target chosen from a category otherthan the one from which they expected the wordtargets to be chosen, given the particular primingevent, and that when this happened the word targetwould equally often come from the two differentunexpected categories. It was strongly emphasizedthat they were to try to avoid anticipating whethera given trial would contain a word target chosenfrom an expected or an unexpected category andthat they should fervently try to adopt the ap-propriate attention-shifting strategy on every trial.The subjects were also told to avoid the strategyof responding "nonword" if the target letter stringdid not belong to the expected category because ifthey adopted such a strategy they would maketoo many errors on trials in which the word targetwas chosen from an unexpected category. A S-min.rest period occurred between the third and fourthblocks of the test series. Following this break, thesubject was asked to review the attentional strategythat was to be adopted for each prime.
Results
Facilitation and Inhibition for Word-Prime-Word-Target Trials
The data for correct responses to theword targets for the various conditions at
the 2,000-msec SOA for the 2000-alonegroup and the three 2,000-mixed groups aredisplayed in Table 2. Since the xxx-primetrials used to compute a facilitation or in-hibition score for a given word-prime con-dition involved exemplars of only those cate-gories represented by the word targets inthat particular word-prime condition, twodifferent xxx-prime control conditions wereused to compute the facilitation or inhibitionscores for the word-prime conditions.
All reported t values were computed usingthe appropriate error term from an overallanalysis of variance as the error estimate.Unless otherwise specified, all differencesreferred to as statistically significant wereassociated with two-tailed p values less than.01. (These two general rules apply to thenonword-target data as well.)
Group 2,000-alone. An inspection ofTable 2 reveals that for the 2,000-aloneSOA a facilitation effect was obtained ineach of the two Expected conditions, aninhibition effect was obtained in each of thethree Unexpected conditions, and these fa-cilitation and inhibition effects were sub-stantial and statistically different from zero.It can also be seen in Table 2 that theerror rates were very low for the 2,000-aloneSOA, that is, only 3.8% averaged across allconditions. Since most errors were madeto LCD word targets (6.7%) and virtuallyno errors were made to HCD word targets(.9%), and since subjects often commentedthat they made errors to some of the LCDwords (e.g., joists, egret, and osprey) be-cause these words were not in their vocabu-lary, the error data are not particularly in-formative measures of processing difficultyand will not be considered further.
An analysis of variance with one between-subjects factor, CA, and two within-subjectsfactors, category dominance and condition,was conducted on the facilitation and inhibi-tion scores for the word-prime conditions.The effect of condition was highly signifi-cant, F(4, 84) = 46.85, MSe = 4414. How-ever, subsequent t tests revealed that therewas no difference in the facilitation scoresfor the two Expected conditions nor werethere differences among the inhibition scores

238 JAMES H. NEELY
for the three Unexpected conditions (all ts< 1.1). Although the CA X Condition in-teraction was marginally significant, F(8,84) = 2.18, MSe = 4414, p = .05, when thedata were broken down according to CAsubgroups, the smallest facilitation effect for
any Expected condition in any CA sub-group was +33 msec, and the smallest in-hibition effect for any Unexpected conditionin any CA subgroup was —34 msec. Theamount of facilitation or inhibition did notdepend on category dominance as is evi-
Table 2Reaction Times (RTs) (in msec) and Error Percentages to Word Targets in Terms of MeanFacilitation (+) or Inhibition (—) Scores for Word-Prime Conditions and of Mean RTs andError Percentages for XXX-Prime Conditions for the 2,000-msec Stimulus Onset Asynchrony
Group
Condition
Word prime
NS-Ex-R NS-Ux-U S-Ex-U S-Ux-U S-Ux-R
XXX prime
x-a x-b
2,000-aloneRTSEMn
% errorsN
250-2,000RTSEMn&
% errorsN
400-2,000RTSEn»a
% errorsN
700-2,000RTSEitn*
% errorsN
+47***6.9
23***-.2
275
+ 1411.1
22*+.2
188
+33*13.022*
+3.2186
+26*13.722*
+ 1.6186
_72***8.4
23***-4.6266
_70***13.526**
-9.5165
-56***10.326**
-5.2171
-57***12.024**
-6.5171
+56***7.4
24***+.1
553
+30***8.4
24**-1.6369
+49***8.7
26**-2.0365
+48***9.9
25**+.6
374
-72***13.623***
-4.6268
-77***12.528***
-1.5177
-68***14.027***
-4.2175
-100***13.429***
-2.6180
_59***14.219*
-2.5271
-63***17.125**
-8.8169
-43***14.426**
-6.6174
-53***11.224**
-5.1176
716
——2.4
1,098
765— .—
4.5 ,720
749
——4.2719
760
——2.7731
718
——2.0
550
759——1.3
372
736——1.4
372
739—
—2.7
362
Note. Facilitation and inhibition scores were derived by subtracting word-prime RTs (error percentages)from xxx-prime RTs (error percentages). Facilitation or inhibition scores for Conditions NS-Ex-R andS-Ux-U are based on Condition x-a, and facilitation or inhibition scores for Conditions NS-Ux-U,S-Ex-U and S-Ux-R are based on Condition x-b. Word-prime-word-target conditions = Nonshift-Expected-Related (NS-Ex-R), Nonshift-Unexpected-Unrelated (NS-Ux-U), Shift-Expected-Unrelated (S-Ex-U), Shift-Unexpected-Unrelated (S-Ux-U), and Shift-Unexpected-Related (S-Ux-R). n = number of subjects, out of24, showing the facilitation or inhibition effect. N = total number of correct-response observations remain-ing after exclusion of errors and shutter-failure trials. Variations in N greater than 30 are due to differences inmaximum number of observations possible for different conditions.
* Number of subjects, out of 32, showing the facilitation or inhibition effect.* p < .05, one-tailed.* p < .05, two-tailed.
**p < .01, two-tailed.***p < .001, two-tailed.

SEMANTIC PRIMING OF LEXICAL MEMORY 239
denced by the nonsignificant Condition XCategory Dominance interaction, F(4, 84)= 1.36.
The word-target data for the 2,000-aloneSOA provide some useful general informa-tion about the present paradigm. First ofall, the fact that facilitation was obtainedin Condition Nonshift-Expected-Related(siRD-robin) indicates that in the lexical de-cision task a semantic-facilitation effect canbe produced by a category-exemplar rela-tionship between the priming word and thetarget word as well as by the previouslyemployed primary-associative relationshipbetween the priming word and the targetword (e.g., CAT-dog and DocroR-nurse).Furthermore, within the framework of thePosner-Snyder (1975a) theory, the inhibi-tion obtained in Condition Nonshift-Unex-pected-Unrelated (BiRD-arm) serves as asource of evidence that conscious attentionwas partially involved in the facilitation ef-fect obtained for BiRD-robin trials. The fa-cilitation effect obtained in Condition Shift-Expected-Unrelated (BODY-door) and theinhibition effects obtained in the two Shift-Unexpected conditions (Boov-sparrow andBODY-heart) are methodologically importantbecause they verify that the subjects didindeed shift their attention during the 2,000-msec SOA interval. Also, the fact that themagnitude of the inhibition obtained forBODY-heart trials was not significantlysmaller than the magnitude of the inhibitionobtained for BODY-sparrow trials indicatesthat the automatic activation spreading fromthe "body" logogen to the "heart" logogenhad decayed during the 2,000-msec SOA.
Groups 2,000-mixed. As can be seen inTable 2, the word-target data for the 2,000-msec SOA showed the same pattern for allthree 2,000-mixed groups, that is, facilita-tion in the two Expected conditions andinhibition in the three Unexpected condi-tions, and this pattern was very similar tothe pattern that emerged in the 2,000-alonedata, but with the magnitude of the facilita-tion and inhibition effects somewhat reducedfrom that in the 2,000-alone data. This simi-larity in the pattern of results for the 2,000-msec SOA for all groups is important be-
cause it indicates that subjects were adopt-ing similar strategies at the 2,000-msec SOAindependently of the presence and durationof the shorter intermixed SOAs. Thus, itwould seem to be more parsimonious to at-tribute any differences in the pattern of re-sults at the shorter SOAs to changes in theoperation of attentional mechanisms withtime rather than to the subjects' adoptingcompletely different strategies for the dif-ferent SOAs in the different groups.
An analysis of variance with two between-subjects factors, CA and group, and twowithin-subjects factors, category dominanceand condition, was conducted on the facili-tation and inhibition scores for the 2,000-msec SOAs from the three 2,000-mixedgroups. The contention that the same pat-tern of data held true for all three 2,000-mixed groups was supported by the nonsig-nificant Group X Condition interaction (F< 1). The only effect to approach statisticalsignificance was the highly significant maineffect of condition, F(4, 360) = 66.99, MS*= 8750. Unlike the 2,000-alone data, partof this effect was attributable to there beinga larger inhibition effect in Condition Shift-Unexpected-Unrelated (BODY-sparrow) thanin Condition Shift-Unexpected-Related(BODY-heart) |>(360) = 3.01; sign test, 58+ and 38 -; p < .05, one-tailed] or inCondition Nonshift-Unexpected-Unrelated(BiRD-arm) [*(360) =2.15, p < .05; signtest, 59 + and 37 -, p < .05]. The factthat the inhibition effect was smaller forBODY-heart trials than for BODY-sparrowtrials indicates that the automatic activationspreading from the "body" logogen to the"heart" logogen had not completely decayedduring the 2,000-msec SOA for the 2,000-mixed groups. The fact that the inhibitioneffect was smaller for BiRD-arm trials thanfor BODY-sparrow trials may be attributableto the subjects' paying more attention to theShift prime than to the Nonshift prime be-cause of the emphasis given to the shiftingstrategy by the instructions. Further sup-port for the contention that subjects paidmore attention to the Shift primes than tothe Nonshift prime is provided by the factthat the facilitation effect in Condition Shift-

240 JAMES H. NEELY
+60 n
-B+40'
E
T+20-
|
I
1
-20-
^-40H
1-60-
-80-
,NS-Ex-R/(BIRD-robin)
S-Ux-UIBODY-sparrow)
250 350 450 550SOA (in msec)
650 750
Figure 2. Amount of facilitation (+) or inhibi-tion (—) (in msec) for word targets in word-prime conditions as a function of stimulus onsetasynchrony (SOA) (in msec). (NS-Ex-R = Non-shift-Expected-Related; NS-Ux-U = Nonshift-Un-expected-Unrelated; S-Ex-U = Shift-Expected-Unrelated; S-Ux-R = Shift-Unexpected-Related;S-Ux-U = Shift-Unexpected-Unrelated.)
Expected-Unrelated (BODY-door) was mar-ginally larger than the facilitation effect inCondition Nonshift-Expected-Related (BIRD-robin), *(360) = 1.87, p < .05, one-tailed;sign test, 57 + and 39 -, p < .05, one-tailed.
Of course, the most important aspect ofthe data from the 2,000-mixed groups isthe time course of the development of thefacilitation and inhibition effects seen foreach of the conditions at • the 2,000-msecSOA as the SOA was increased from 250msec to 2,000 msec. Given the design of thepresent experiment, there are two ways toevaluate changes in facilitation and inhibi-tion with changing SOA. First, in each of thethree 2,000-mixed groups, the magnitude ofthe facilitation or inhibition effect for a givencondition at the 2,000-msec SOA can becompared with the magnitude of the facilita-tion or inhibition effect in the same conditionat the short SOA, with SOA as a within-subjects factor. The second method ofevaluating the changes in facilitation and
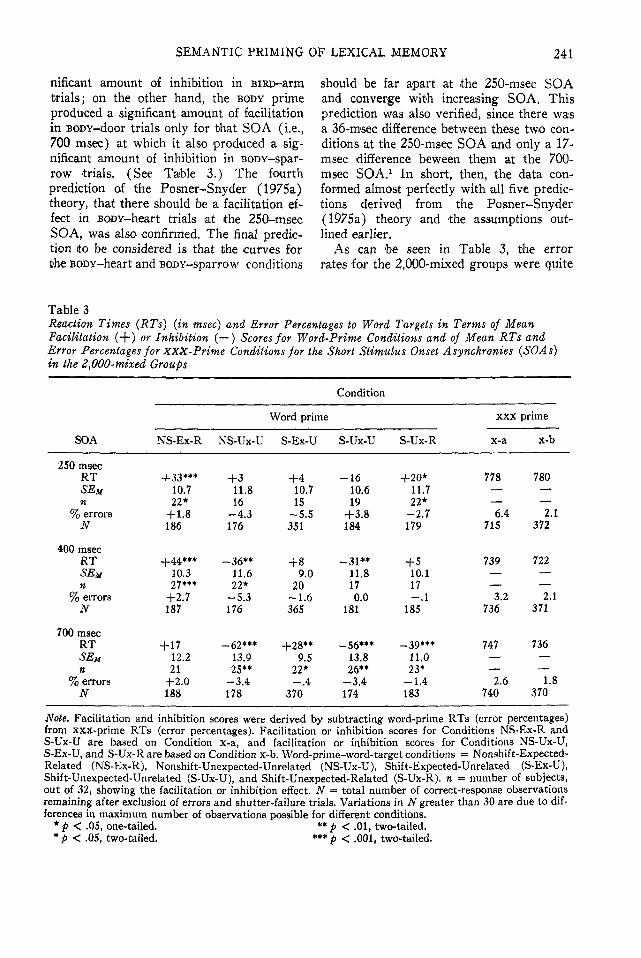
inhibition with changing SOA is to comparethe magnitude of the facilitation or inhibitioneffect for a given condition at the 250-,400-, and 700-rosec SOAs, with SOA as abetween-subjects factor. Since an analysis ofvariance conducted on the data from Group700-2,000 indicated that the amount of fa-cilitation or inhibition in a given conditionwas the same for the 700- and 2,000-msecSOAs, an adequate picture of changes infacilitation or inhibition as the SOA in-creases from 250 msec to 2,000 msec is pro-vided by comparisons of the magnitudes ofthe facilitation or inhibition effects obtainedfor the various conditions at the 250-, 400-,and 700-msec SOAs. The data for correctresponses to the word targets for the variousconditions at the 250-, 400-, and 700-msecSOAs are shown in Table 3 in the sameformat as the data presented in Table 2.
The facilitation and inhibition effects forthe RTs are displayed in Figure 2 so thatthe pattern of the obtained results can bedirectly compared with the pattern of pre-dicted results displayed in Figure 1. Recallthat there were five important aspects ofthe predictions derived from the Posner-Snyder (1975a) theory. The first predictionwas that the facilitation effect for BiRD-robintrials would be flat as a function of SOAand that the magnitude of the facilitationeffect for BODY-door trials would start atzero at the 250-msec SOA and increasewith increasing SOA until it converged withthe facilitation effect for the BiRD-robintrials. As can be seen in Figure 2, this pre-diction was confirmed by the data. The sec-ond prediction was that the magnitude ofthe inhibition effects for the BODY-sparrowand BiRD-arm trials would be equivalent,with both originating at zero at the 250-msecSOA and increasing with increasing SOA.This prediction was borne out by the data,since the 16-msec inhibition effect obtainedfor the BODY-sparrow trials at the 250-msecSOA was not statistically significant. (SeeTable 3.) The third prediction confirmedby the data was that the BIRD prime pro-duced a significant amount of facilitation inBiRD-robin trials at the 250-msec SOA, butat that same SOA it did not produce a sig-
SEMANTIC PRIMING OF LEXICAL MEMORY 241
nifkant amount of inhibition in BiRD-armtrials; on the other hand, the BODY primeproduced a significant amount of facilitationin BODY-door trials only for that SOA (i.e.,700 msec) at which it also produced a sig-nificant amount of inhibition in BODY-spar-row trials. (See Table 3.) The fourthprediction of the Posner-Snyder (197Sa)theory, that there should be a facilitation ef-fect in BODY-heart trials at the 250-msecSOA, was also confirmed. The final predic-tion to be considered is that the curves forthe BODY-heart and BODY-sparrow conditions
should be far apart at the 250-msec SOAand converge with increasing SOA, Thisprediction was also verified, since there wasa 36-msec difference between these two con-ditions at the 250-msec SOA and only a 17-msec difference beween them at the 700-msec SOA.1 In short, then, the data con-formed almost perfectly with all five predic-tions derived from the Posner-Snyder(I975a) theory and the assumptions out-lined earlier.
As can be seen in Table 3, the errorrates for the 2,000-<mixed groups were quite
Table 3Reaction Times (RTs) (in msec) and Error Percentages to Word Targets in Terms of MeanFacilitation (+) or Inhibition (—) Scores for Word-Prime Conditions and of Mean RTs andError Percentages for XXX-Prime Conditions for the Short Stimulus Onset A synchronies (SO As)in the 2,000-mixed Groups
Condition
Word prime xxx prime
SOA NS-Ex-R NS-Ux-U S-Ex-U S-Ux-U S-Ux-R x-a x-b
250 msecRTSEMn
% errorsN
400 msecRTiSfijtfn
% errorsN
700 msecRTSEMn
% errorsN
+33***10.722*
+1.8186
+44***10.327***
+2.7187
+ 1712.221
+2.0188
+311.816
-4.3176
-36**11.622*
-5.3176
-62***13.925**
-3.4178
+410.715
-5.5351
+89.0
20-1.6365
+28**9.5
22*-.4
370
-1610.619
+3.8184
-31**11.8170.0
181
-56***13.826**
-3.4174
+20*11.722*
-2.7179
+510.117
1
185
-39***11.023*
-1.4183
778—
—6.4
715
739
——3.2
736
747
——2.6
740
780
——2.1
372
722
——2.1
371
736—
—1.8370
Note, Facilitation and inhibition scores were derived by subtracting word-prime RTs (error percentages)from xxx-prime RTs (error percentages). Facilitation or inhibition scores for Conditions NS-Ex-R andS-Ux-U are based on Condition X-a, and facilitation or inhibition scores for Conditions NS-Ux-U,S-Ex-U, and S-Ux-R are based on Condition x-b. Word-prime-word-target conditions = Nonshift-Expected-Related (NS-Ex-R), Nonshift-Unexpected-Unrelated (NS-Ux-U), Shift-Expected-Unrelated (S-Ex-U),Shift-Unexpected-Unrelated (S-Ux-U), and Shift-Unexpected-Related (S-Ux-R). n = number of subjects,out of 32, showing the facilitation or inhibition effect. N — total number of correct-response observationsremaining after exclusion of errors and shutter-failure trials. Variations in N greater than 30 are due to dif-ferences in maximum number of observations possible for different conditions.
*p < .05, one-tailed. ** p < .01, two-tailed.* p < .05, two-tailed. *** p < .001, two-tailed.

242 JAMES H. NEELY
low, 4.4% averaged across all conditionsand all SOAs. Also, as in the 2,000-alonegroup, there were many more errors madeto LCD words (7.4%) than to HCD words(1.2%). Thus, the error data will not begiven further consideration.
A single analysis of variance was per-formed on only the data for the 250-, 400-,and 700-msec SOAs with SOA a between-subjects factor, and separate analyses ofvariance were conducted for the three dif-ferent 2,000-mixed groups. In all analyses,CA was a between-subjects variable andcategory dominance and condition werewithin-subjects variables. When the data forthe 250-, 400-, and 700-msec SOAs wereanalyzed, the SOA X Condition interactionwas significant, F(8, 360) = 3.41, MSe -7125. When the data were analyzed sepa-rately for the three different 2,000-mixedgroups, the SOA X Condition interactionwas significant for Groups 250-2,000 and400-2,000, F(4, 120) = 15.63, MS, =7885, and F(4, 120) = 5.60, MSe = 7208,respectively, but, as noted earlier, it wasnot significant for Group 700-2,000, F(4,120) = 2.23. In all analyses, the main effectsof category dominance and CA were neversignificant nor did these two factors par-ticipate in any statistically significant in-teractions.2
Table 4 presents for each of the fiveconditions the t values for comparisons ofthe magnitudes of facilitation or inhibitionobtained at the two long SOAs with themagnitudes of facilitation or inhibition ob-tained at each of the two short SOAs. Table4 also presents, for each of the five condi-tions for the within-subjects comparisonsinvolving the 2,000-msec SOA, the numberof -subjects who showed a facilitation or in-hibition effect at the short SOA that wassmaller than the facilitation or inhibitioneffect obtained at the 2,000-msec SOA. Ofparticular relevance are the differences be-tween the shortest (250-msec) SOA andthe two long SOAs. The main conclusionto be drawn from Table 4 is that, with theexception of Condition Nonshift-Expected-Related (siRD-robin), at least one of thecomparisons between the 250-msec SOA
and the two longer SOAs was highly signifi-cant or approached statistical significance.Thus, these statistical analyses support thestatements made above about the changesin the amount of facilitation or inhibitionobserved in a given condition as the SOAdecreased from 700 or 2,000 msec to 250msec. These analyses also support the con-
1 It should be noted that the fact that there wasstatistically less (i.e., 29 msec less) inhibition inCondition Shift-Unexpected-Related than in Con-dition Shift-Unexpected-Unrelated for the 2,000-msec SOA in the 2,000-mixed groups is prob-lematic for the interpretation that automaticspreading activation has decayed within 700 msecin the 2,000-mixed groups. However, the decayof spreading-activation interpretation is supportedby the fact that there was no statistical differencein the amounts of inhibition obtained for ConditionsShift-Unexpected-Related and Shift-Unexpected-Unrelated for the 2,000-msec SOA in the 2,000-alone group. Given the previously obtained evi-dence for a decay of spreading activation (Loftus,1973; Warren, 1972; Meyer, Schvaneveldt, &Ruddy, Note 1) and the present study's supportiveevidence in the form of statistically equivalent in-hibition effects in Conditions Shift-Unexpected-Related and Shift-Unexpected-Unrelated at the700-msec and 2,000-alone SOAs and in the formof a flat facilitation effect as a function of SOA inCondition Nonshift-Expected-Related, it wouldseem that the problematic finding of statisticallyless inhibition in Condition Shift-Unexpected-Re-lated than in Condition Shift-Unexpected-Unre-lated for the 2,000-msec SOA in the 2,000-mixedgroups can be largely discounted, at least forpresent purposes. Nevertheless, future researchshould be conducted to help clarify this one minordeviation from the predicted pattern of results.
2 There was one exception to this general con-clusion. In Group 400-2,000, the Category Domi-nance X Condition interaction just barely met con-ventional standards for statistical significance, F(4, 120) =2.47, # = .05. However, this interactionis of little interest because all of the means forthe Expected conditions were facilitation effectsand all of the means for the Unexpected conditionswere inhibition effects for both HCD and LCDitems. Furthermore, this interaction most certainlywould not have been statistically significant had amore conservative analysis of variance been con-ducted with categories and items treated as ran-dom factors (cf. Clark, 1973). Categories anditems were not treated as random factors in thepresent analyses because Clark's arguments forsuch a treatment are less compelling when eachcategory and each item occurs in each condition oftheoretical interest, as was the case in the presentexperiment.

SEMANTIC PRIMING OF LEXICAL MEMORY 243
Table 4Results of t Tests Comparing Facilitation orInhibition Scores for EachWord-Prime-Word-Target Condition at the250-msec and 400-msec Stimulus OnsetA synchronies (SOAs) with Facilitation orInhibition Scores at the 700-msec and2,000-msec SO As
Condition
NS-Ex-R
NS-Ux-U
S-Ex-U
S-Ux-U
S-Ux-R
Short SOA
250/n
400tn
250tn
400tn
250tn
400*n
250/n
400tn
250/n
400tn
Long
700"
-1.11— .
-1.84*—
4.36***
—
1.72*
—
1.61**
—
1.38— .
2.63**—
1.98*—
3.94***
—
2.92**—
SOA
2,000"
-1.1314
<115
4.32***24**
1.2721
1.5022*
2.78**24**
3.58**26**
2.46**21
4.90***25**
3.19**23*
Note. Word-prime-word-target conditions = Non-shift-Expected-Related (NS-Ex-R), Nonshift-Un-expected-Unrelated (NS-Ux-U), Shift-Expected-Unrelated (S-Ex-U), Shift-Unexpected-Unrelated(S-Ux-U), and Shift-Unexpected-Related (S-Ux-R).n — number of subjects, out of 32, showing the effect.The t values are negative if the facilitation or inhibi-tion effect obtained at the longer SOA was smallerthan the facilitation or inhibition effect obtained atthe shorter SOA.
B df = 360, between-subjects comparison.b df = 120, within-subjects comparison.* p < .05, one-tailed.
** p < .06, one-tailed.* p < .05, two-tailed.
** p < .01, two-tailed.*** p < .001, two-tailed.
elusion that the magnitude of the facilitationeffect for the BiRO-robin trials did not varywith changes in SOA.8
In addition to the effects of SOA uponfacilitation or inhibition within a condition,certain between-conditions comparisons atthe various SOAs are of theoretical impor-tance. The t values associated with thesecomparisons are presented in Table 5 alongwith the number of subjects, out of 32,showing the observed effect. As can be seenin Table 5, the t tests and the sign testssupport the conclusions made above that(a) there was a difference in the magnitudesof the facilitation effects in Conditions Shift-Expected-Unrelated (BODY-door) and Non-shift-Expected-Related (siED-robin) at thetwo short SOAs but not at the 700-msecSOA; (b) the inhibition effects in Condi-tions Shift-Unexpected-Unrelated (BODY-sparrow) and Nonshift-Unexpected-Unre-lated (BiRD-arm) were equivalent at allthree SOAs; and (c) there was less in-hibition in Condition Shift-Unexpected-Re-lated (BODY-heart) than in Condition Shift-Unexpected-Unrelated (soDY-sparrow) atthe 250- and 400-msec SOAs but not at the700-msec SOA. In short, the statistical anal-yses support all five of the theoretically im-portant conclusions drawn above.
RTs for xxx-Prime-Word-Target Trials
Although the major theoretical thrust ofthe data is provided by the facilitation andinhibition scores, a few passing commentsneed to be made about the RTs to the wordtargets when they followed the xxx primes.For all four groups, the 2,000-alone groupand the three 2,000-mixed groups, the CA XCondition (x-a vs. x-b) interaction wassignificant: for the 2,000-alone group, F(2,21) = 5.27, p < .025, and for the 2,000-
a Actually as can be seen in Table 4, there wassome evidence that the facilitation effect in Con-dition Nonshift-Expected-Related was actuallygreater at the 400-msec SOA than at the 700-msecSOA (p<.OS, one-tailed). Such an effect can beaccounted for by assuming that the facilitationeffect produced by automatic spreading activationis slightly greater than the facilitation effect pro-duced by limited-capacity attention (viz., by as-suming that + > * in Table 1).

244 JAMES H. NEELY
Table 5Results of t Tests /or Theoretically CriticalComparisons BetweenWord-Prime-Word-Target Conditions as aFunction of Stimulus Onset Asynchrony (SOA)
SOA (in msec)
Comparison 250 400 700
S-Ex-U vs. NS-Ex-R
tn
S-Ux-U vs. NS-Ux-U
tn
S-Ux-R vs. S-Ux-U
1.95* 2.45* <123* 24** 15
1.29 <113 19
2.45* 2.41*24** 22*
17
1.1420
Note. Word-prime word-target conditions = Non-shift-Expected-Related (NS-Ex-R), Nonshift-Un-expected-Unrelated (NS-Ux-U), Shift-Expected-Unrelated (S-Ex-U), Shift-Unexpected-Unrelated(S-Ux-U), Shift-Unexpected-Related (S-Ux-R). »= number of subjects, out of 32, showing the effect.* p < .05, one-tailed.* p < .05, two-tailed.
** p < .01, two-tailed.
mixed groups, all three Fs(l, 30) > 26.0.These differences in the xxx-prime RTs asa function of CA group merely demonstratethat exemplars from certain 'semantic cate-gories were more difficult to classify aswords than were the exemplars from othersemantic categories. Since such differencesare necessarily taken into account in thecomputation of the facilitation and inhibitionscores for the word-prime conditions, theyare theoretically uninteresting. Categorydominance was the only other variable thathad a statistically significant effect on RTsto the word targets following the xxxprime: for the 2,000-alone group, F(l, 21)= 127.50, and for the three 2,000-mixedgroups, all three Fs(l, 30) > 130.0. Aver-aged across all groups the mean RT forHCD words was 68 msec faster than themean RT for LCD words. Since the effectof category dominance is most probably aword-frequency effect (e.g., Stanners, Jas-trzembski, & Westbrook, 1975), it is not
particularly relevant to the theoretical issuesunder consideration in this article.
Discussion of Word-Target Data
The word-target data confirmed all fiveof the major predictions derived from Pos-ner and Snyder's (1975a) two-factor theoryof attention and therefore offer striking sup-port for that theory. In addition, the word-target data have two other interesting im-plications.
First, the present data -help to clarifywhether activation of a prime's logogen au-tomatically spreads to semantically relatedlogogens even when the subject is usingthat prime to direct his attention to seman-tically unrelated logogens. This issue wasnot adequately addressed by Posner andSnyder's (1975b) experiments. That is, al-though there was evidence in their experi-ments for automatic activation when theprobability was low that a priming cuewould appear in the target array, subjectssaid that they did not use the priming eventto direct their attention to a logogen otherthan the one activated by the priming event.Rather, they tried to ignore the prime. Sincefacilitation was obtained in this condition,automatic activation apparently occurredwithout intention on the part of the sub-ject. However, since subjects did not con-sciously use the prime to direct their at-tention to a different logogen, there was noevidence that the automatic activation of alogogen did indeed proceed normally evenwhen the subject's attention was focused ona different logogen. In fact, in the animal-name classification experiment in which thesubject was focusing his attention on thelogogen for the priming word CAT when thetarget word dog was presented, there wasno facilitation of the processing of the targetword dog when it followed the semanticallyrelated priming word CAT. Instead, therewas an inhibition of the processing of dogunder such conditions. Since the SOA be-tween the priming event and the target ar-ray was long enough (i.e., 500 msec) toallow the subject to focus his attention onthe logogen for CAT before the target dog

SEMANTIC PRIMING OF LEXICAL MEMORY 245
appeared, the subject would have needed toshift his attention from the "cat" logogento the "dog" logogen before he could takeadvantage of the activation having spreadto the "dog" logogen from the "cat" logo-gen. This required shift in attention couldhave consumed enough time to override thefacilitatory effect of the automatic activa-tion spreading from the "cat" logogen to the"dog" logogen. Consequently, the obtainednet inhibition effect could have occurred de-spite the fact that automatic activation hadindeed spread from the "cat" logogen to the"dog" logogen as predicted by the Posner-Snyder (1975a) theory.
The data from Condition Shift-Unex-pected-Related (soDY-heart) in the presentexperiment supports this analysis of whyPosner and Snyder (197Sb) found an in-hibition effect when CAT was the prime fora target array containing dog and there wasa high probability that the priming worditself would appear in the target array. Sinceinhibition was obtained for sooy-heart trialsat the 700-msec SOA and facilitation wasobtained for BODY-heart trials at the 250-msec SOA, if Posner and Snyder (1975b)had used a shorter SOA, presumably theytoo would have obtained a facilitation effectrather than the inhibition effect they ob-tained with their 500-msec SOA.
A second implication of the present dataconcerns the rates at which facilitation andinhibition build up within the conscious-at-tention mechanism. Posner and Snyder(197Sa) and other theorists have not ad-dressed this issue explicitly, probably be-cause there are no relevant data, sinceprevious experiments have confounded auto-matic-activation and conscious-attention fa-cilitation effects. In the present experiment,these two effects were unconfounded, anddata from Conditions Shift-Unexpected-Un-related (BODY-sparrow) and Shift-Expected-Related (soDY-door) indicated that con-scious-attention inhibition built up fasterthan conscious-attention facilitation. Thisconclusion is based on the observation thatat the 400-msec SOA there was a substan-tial and statistically .significant 31-msec in-hibition effect for BODY-nsparrow trials but
only a small and nonsignificant 8-msec fa-cilitation effect for soDY-door trials. Ofcourse, these results may merely reflect afailure to detect a facilitation effect forBODY-door trials at the 400-nisec SOA, butif future research should further substantiatethis asymmetry in the buildup of conscious-attention facilitation and conscious-attentioninhibition, it would be theoretically quiteimportant.
One way in which this asymmetry is im-portant vis a vis the Posner-Snyder(197Sa) theory is that it demonstrates thatwhen a prime produces both inhibition andfacilitation effects, the inhibition effects donot necessarily build up more slowly thando the facilitation effects. One could there-fore conclude with a greater degree of cer-tainty that in the Posner and Snyder(197Sb) experiments, facilitation precededinhibition only because the facilitation wasbeing produced by the fast automatic-activa-tion process, whereas the inhibition wasbeing produced by the slower acting con-scious-attention mechanism, and not becausefacilitation effects in general always pre-cede inhibition effects.
Another theoretical implication of thefinding that conscious-attention facilitationbuilds up more slowly than conscious-atten-tion inhibition is that the conscious-attentionfacilitation is being produced by activationspreading from the logogen upon which at-tention is being focused—once attention hasbeen shifted to that logogen. That is, con-scious-attention inhibition is being producedduring the time that attention is beingshifted to a logogen that was not automati-cally activated by the word prime, whereasconscious-attention facilitation is being pro-duced by activation spreading from thatlogogen—but only after attention has beenfully shifted to that logogen.
Facilitation and Inhibition for Word-Prime-Nonward-Target Trials
The data for correct responses to thenonword targets for the various conditionsand SOAs are given in Tables 6 and 7 inthe same format as were the word-target
246 JAMES H. NEELY
Table 6Reaction Times (RTs) (in msec) and ErrorPercentages to Nonword Targets in Terms ofMean Facilitation (+) or Inhibition (—)Scores for Word-Prime Conditions and ofMean RTs and Error Percentages forXXX-Prime Conditions for the 2,000-msecStimulus Onset Asynchrony
Condition
Word prime xxx prime
Group
2,000-alone
RTSEnn
% errorsN
250-2,000
RTSEu»»
% errorsN
Shift
+19**6.0
19*-.3
1,107
+23**7.2
22*-.5
723
Non-shift
+21**6.6
167
557
+1811.217
-2.4358
Shift
852—
—2.01,102
904
——
2.3726
Non-shift
851—
—2.1
547
891—
—1.9
358
400-2,000
RTSEMre»
% errorsN
700-2,000
RTSEitn*
% errorsN
+21**6.1
22*-.6
722
+ 14*5.8
20+.4
739
+89.7
22*.0
366
+612.317
+1.6369
873
——
1.9730
870——
1.8738
868
——1.4
367
866. ——2.9
365
Note. Facilitation ( + ) and inhibition ( — ) scoreswere derived by subtracting word-prime RTs (errorpercentages) from corresponding xxx-prime RTs(error percentages), n = number of subjects, out of24, showing the effect. N = total number of cor-rect-response observations remaining after exclusionof errors and shutter-failure trials. Variations in Ngreater than 30 are due to differences in maximumnumber of observations possible for different condi-tions.• Number of subjects, out of 32, showing the facilita-tion or inhibition effect.* p < .05, one-tailed.* p < .05, two-tailed.
**p < .01, two-tailed.
data in Tables 2 and 3. A discussion of thetheoretical implications of the facilitation orinhibition scares for the word-prime-non-word-target conditions will be deferred untilafter these data have been described.
Group 2,000-alone. As can be seen inTable 6, a relatively large and statisticallysignificant facilitation effect of nearly 20msec was obtained at the 2,000-alone SOAwhen a nonword target followed a wordprime, with the magnitude of the facilitationeffects for the Shift and Nonshift conditionsdiffering 'by less than 3 msec. Since errorrates for the nonword targets were evenlower than they were for the word targets(i.e., only 2.3% averaged across all condi-tions, with a 1.9% error rate for HCD non-words and a 2.7% error rate for LCD non-words) , they are of little interest.
An analysis of variance, with CA as a be-tween-subjects factor and category domi-nance and condition as within-subjects fac-tors, was conducted on the 'facilitation scoresfor the word-prime conditions. The resultsof this analysis indicated that the size of thefacilitation effect was not affected by anyof these factors nor by their various com-binations.
Groups 2,000-mixed. As was the casefor the word-target data for the 2,000-mixedgroups, the effects .in the word-prime condi-tions at the 2,000-msec SOA were not dif-ferent for the nonword targets in Groups250-2,000, 400-2,000, and 700-2,000 (F <1). As with the 2,000-alone SOA, a facilita-tion effect was obtained for both word-primeconditions. However, this facilitation effectwas statistically significant only for the Shiftconditions (p < .05), with 64 of the 96subjects showing the effect.
As can be seen in Table 7, for the shorterSOAs the pattern of data was relativelyinconsistent, with both facilitation and in-hibition effects being obtained for each ofthe two word-prime conditions but at dif-ferent SOAs. Only one of these effects wasstatistically significant—the facilitation ef-fect in Condition Nonshift at the 400-msecSOA (p < .05)—but only 19 of the 32 sub-jects showed the effect. Error rates were toolow to be informative—2.6% overall, 1.9%

SEMANTIC PRIMING OF LEXICAL MEMORY 247
for HCD nonwords, and 3.2% for LCDnonwords.
For each of the three 2,000-mixed groups,a separate analysis of variance was performedfor the word-prime RT data with CA as abetween-subjects factor and category domi-nance, condition, and SOA as within-sub-jects factors. The only effect associated witha p value less than .05 was SOA in Group250-2,000, F(l, 30) = 5.54, MS» = 5742.Averaged across the Shift and Nonshiftconditions, there was a 2-msec inhibitioneffect at the 250-msec SOA and a 21-msecfacilitation effect at the 2,000-msec SOA.Tihe results of these analyses permit oneconclusion pertinent to the theoretical issuesunder consideration here: Although SOAhad no consistent effect on the facilitationeffects obtained in the word-prime-nonword-target conditions, these facilitation effectswere larger at the 2,000-msec SOA than atthe 250-msec SOA.
RTs for xxx-Prime-Nonword-TargetTrials
Although the data of primary concern forthe nonword targets are the facilitation andinhibition effects produced by the wordprimes, one comment needs to be made re-garding the RTs to the nonword targetsfollowing the xxx prime. For all fourgroups, the only effect to reach conventionallevels of statistical significance was the effectof category dominance: for the 2,000-alonegroup, F(l, 21) = 10.31, and for the three2,000-mixed groups, all three Fs(l, 30) >22.0. Since the category-dominance effectfor the nonword targets following the xxxprime replicates the category-dominance ef-fect for the word targets following the xxxprime, part of the category-dominance effectfor the word targets may have been pro-duced by structural differences that wereconfounded with category dominance. How-ever, since the category-dominance effectwas much larger for the word targets (68msec) than for the nonword targets (18msec), it may be that frequency per se wasoperating in conjunction with structuralproperties to produce the category-domi-nance effect for the word targets. It should
also be noted in this regard that Stannerset al. (1975) have obtained frequency ef-fects for words and nonwords similar to theones obtained here.
Discussion of Nonward-Target Data
Since the principal finding for the word-prime—nonword-target trials was that the
Table 7Reaction Times (RTs) (in msec) and ErrorPercentages to Nonword Targets in Terms ofMean Facilitation (+) or Inhibition (—)Scores for Word-Prime Conditions and ofMean RTs and Error Percentages forXXX-Prime Conditions for the Short StimulusOnset Asynchronies (SOAs) in the 2,000-mixedGroups
Condition
Word prime XXX prime
SOA
250 msec
RTSEMn
% errorsN
400 msec
RTSEMn
% errorsN
Shift
+ 15.7
20-1.2715
+127.8
19+.4
724
Non-shift
-410.616
-1.4351
+20*10.219
-1.3362
Shift
896— .—3.4
722
870
——3.8
722
Non-shift
891
——3.2
356
857
——2.4
362
700 msec
RTSEMn
% errorsN
-138.8
20+.5
725
+ 149.1
19.0
373
856 857
2.4 1.8735 367
Note. Facilitation ( + ) and inhibition ( — ) scoreswere derived by subtracting word-prime RTs (errorpercentages) from corresponding xxx-prime RTs(error percentages). n = number of subjects, out of32, showing the effect. N = total number of correct-response observations remaining after exclusion oferrors and shutter-failure trials. Variations in Ngreater than 30 are due to differences in maximumnumber of observations possible for different condi-tions.* p < .05, two-tailed.

248 JAMES H. NEELY
word primes facilitated nonword decisionsat the 2,000-msec SOA but not at the 250-msec SOA, the crucial theoretical issue ishow to accommodate the word primes' fa-cilitatory effect on nonword decisions. In theabsence of compelling data forcing the con-clusion that the word primes affect theword and nonword RTs by totally inde-pendent mechanisms, it would seem more.parsimonious to assume that the wordprimes influence the word and nonword RTsvia the operation of similar mechanisms.Within the framework of the Posner-Sny-der (1975a) theory, the word prime shouldaffect nonword RTs through the operationof the limited-capacity attentional mecha-nism but not through the operation of theautomatic spreading-activation process. Sincethe word-target inhibition produced by theword prime is in some sense a "pure" mea-sure of the operation of the limited-capacityattentional mechanism (see Table 1), itwould seem that the best strategy for in-corporating the nonword facilitation effectinto the Posner-Snyder (1975a) theorywould be to consider some alternative ex-planations of how the limited-capacity at-tentional mechanism might produce inhibi-tion for the word-prime-word-target trials,and then see how these alternative explana-tions fare with the nonword-target data.When such a strategy is adopted, the non-word-target data become quite importanttheoretically in that they discriminate amongalternative explanations of how the limited-capacity attentional mechanism operates toproduce inhibition in the two Unexpected-Unrelated word-prime-word-target condi-tions BiRD-arm and BODY-sparrow.
As noted in the introduction of this ar-ticle, one interpretation of the word-prime-word-target inhibition effect is that the wordprime depletes the limited resources of at-tention that must be used to "read out",from the target word's logogen, the informa-tion necessary for making the lexical deci-sion. This limited-resources interpretation ofthe word-prime—word-target inhibition effectcorresponds rather closely to the generalviews of Posner and Snyder (1975a). In thepresent experiment, -when the word prime
was a Shift BODY prime, the limited re-sources of attention would have been de-pleted by the acts of retrieving .the unauto-mated BODY-"building" association and con-sciously maintaining the activation in the"building" logogen so that the activationspreading from .that logogen would not de-cay during the 2,000-msec SOA; when theword prime was the Nomshift BIRD prime,the limited resources of attention would havebeen depleted by the subject's consciouslymaintaining activation in the "bird" logogen.However, when xxx was the prime, thesubject presumably would not have initiatednor maintained activation at a particularlogogen, and therefore the xxx prime wouldnot have depleted the limited resources ofattention. Thus, according to this view, RTsto word targets were slower following theword primes than following the xxx primebecause following the word primes there wasless attentional capacity remaining to readout, from the word-target logogen, the in-formation necessary for making the lexicaldecision.
What does this limited-resources viewpredict for the nonword targets? It predictsthat the word primes should produce an in-hibition effect for RTs to nonword targets.If the nonword response is made after allword logogens have been searched withoutfinding a lexical entry corresponding to thetarget letter string (cf. Forster & Bednall,1976), the search should take longer tocomplete following the word primes thanfollowing the xxx prime because of theword primes' greater depletion of the limitedattentional resources needed to conduct thesearch. The limited-resources view also pre-dicts an inhibition effect for word-prime-nonword-target trials even if the nonwordresponse is based on some process otherthan an unsuccessful exhaustive search ofthe word logogens. For example, some re-searchers (James, 1975; Meyer & Schvane-veldt, 1971; Taft & Forster, 1975) haveargued that both words and pronounceablenonwords have logogens or "lexical entries"stored in long-term memory. Since pro-nounceable nonwords also have lexical en-tries, when pronounceable nonwords are

SEMANTIC PRIMING OF LEXICAL MEMORY 249
used as distractors, finding a lexical entryfor the target letter string does not providesufficient information for making a "word"response. Consequently James (1975) hasargued that under such circumstances thesubject must "look up the meaning" of theletter string in order to make the lexicaldecision. But even if this meaning-look-upview of the lexical decision process is cor-rect, RTs to nonword targets should stillbe inhibited by the word primes because theywould have depleted some of the limited re-sources necessary for a meaning look-up.Therefore, regardless of whether a "non-word" response is based on an unsuccessfulsearch or a failure to find a meaning in ameaning-look-up stage of processing, thelimited-resources interpretation of the word-prime-word-target inhibition effect predictsthat there should also be an inhibition effecton word-prime-nonword-target trials. Ob-viously, this prediction of the limited-re-sources view was contravened by the facilita-tion effect that actually obtained on word-prime-nonword-target trials.
It also should be noted that, according tothis limited-resources view, the limited-ca-pacity attentional mechanism produces fa-cilitation on word-target trials through theinitiation and/or maintenance of spreadingactivation (cf. Collins & Loftus, 1975). Thisview was discussed earlier in this article—in the introduction, and in the discussionof the different rates at which conscious-attention facilitation and conscious-attentioninhibition appeared to build up in the pres-ent experiment. The fact that the facilitationeffect for the nonword targets tends to ruleout this particular limited-resources inter-pretation of the inhibition effect for theUnexpected-Unrelated word-target condi-tions does not necessarily compel one toabandon the view that the conscious-atten-tion mechanism produces facilitation throughthe initiation and/or maintenance of spread-ing activation; one could argue that con-scious attention produces facilitation andinhibition through two different mechanisms.However, such an argument is unparsimoni-ous and should not be accepted in the ab-sence of compelling evidence for it. Although
the apparent difference in the .rates of thebuildup of the conscious-attention facilitationand conscious-attention inhibition obtainedin the present experiment is suggestive ofconscious-attention facilitation and conscious-attention inhibition being produced by dif-ferent mechanism.s, it is not compelling evi-dence for their being produced by differentmechanism's (see earlier discussion). Thus,at present, it is necessary to conclude, forthe sake of parsimony, that the conscious-attention 'facilitation obtained in the presentexperiment was not produced by the limited-capacity attentional mechanism's initiatingand/or maintaining spreading activation inthe logogen on which attention was beingfocused. However, future research should beconducted in order to decide this issue moreconclusively. In the same vein, an addi-tional point should be made before ex-amining two other explanations (i.e., theorder-of-search and semantic-matching ex-planations) of the inhibition effect in theUnexpected-Unrelated word-target condi-tions. The point is that both of these ex-planations can account for conscious-atten-tion facilitation and conscious-attention in-hibition by appealing to only a single under-lying mechanism. Thus, they both meet thecriterion of parsimony discussed in thisparagraph.
According to order-of-search interpreta-tion of the word-prime-word-target inhibi-tion effect, the subject uses the word primeand limited-capacity attention to determinethe order in which the three semantic cate-gories employed in the experiment will besearched. When the xxx prime is presentedthe subject focuses attention on a categorythat has been randomly selected from amongthe three categories, and by chance this ran-domly selected category will be the categorycontaining the word target on one third ofthe trials. However, when the word-primeis presented and the subject has been givenenough time to focus his attention on thecategory that is cued by that word prime,in the two Unexpected-Unrelated conditionshis or her attention will always be focused ona category that does not contain the wordtarget. Consequently, an inhibition effect will

250 JAMES H. NEELY
be obtained for the word targets in the twoUnexpected-Unrelated conditions becausethe subject will have always searchedthrough the wrong category before searchingthe category containing the target word, anda conscious-attention facilitation effect willbe obtained for the word targets in the twoExpected conditions because the subject willalways be searching the correct categoryfirst.
What does this order-of-search mecha-nism predict for the nonword targets? Ifone assumes that a nonword response isbased on an unsuccessful exhaustive searchof the word logogens, there should be nodifference in RT-s to nonword targets fol-lowing the word primes and the xxx primebecause following both, the subject will haveexhaustively searched all three categories,with the only difference between the word-prime and xxx-prime trials being the orderin which the categories were searched. Thus,the facilitation effect that obtained in thepresent experiment for the word-prime-non-word-target trials is problematic for the viewthat the word prime determined the order inwhich the categories were searched.
Since both the limited-resources and theorder-of-search accounts of the conscious-attention inhibition effect (and the con-scious-attention facilitation effect) failed toaccommodate -the word primes' facilitatoryeffect upon RTs to the nonword targets,what is required is an alternative conceptual-ization of the mechanism by which limited-capacity attention produces the word-primeword-target inhibition effect. A clue to whatthis mechanism might be is provided by theobservation that the present word-prime-nonword-target facilitation effect corre-sponds to the negative-response results ob-tained by Posner and Snyder (1975b) andNeely (1976). Recall that in these previousexperiments, negative-response RTs werefaster following the letter prime or the wordprime than following the neutral prime.Both Posner and Snyder (197Sb) and Neely(1976) attributed this result to the sub-jects' using limited-capacity attention tomatch the prime (or an item internallygenerated to the prime) with the target and
then tending to respond "yes" or "word"when there was a match and "no" or "non-word" when there was a mismatch. In anattempt to minimize the likelihood that thesubject would adopt such a matching strat-egy in the present experiment, the wordprimes were chosen so as to cue the subjectsto expect words chosen from very largecategories. Presumably, it would be veryunlikely that the subject could generate allof the exemplars of these large semanticcategories and then respond on the basis ofwhether the target matched or mismatchedwith these internally generated exemplars.
Nevertheless, the subjects could haveadopted some sort of semantic-matchingstrategy (cf. Collins & Loftus, 1975; Smith,Shoben, & Rips, 1974). According to thissemantic-matching hypothesis, the subjecthas a tendency to respond on the basis ofthe similarity between the semantic featuresof the expectancy generated, via consciousattention, to the word prime and the seman-tic features activated by the target letterstring. If the semantic features of the ex-pectancy generated to the word prime andthe semantic features activated by the targetletter string are quite similar, the subjectwould have a tendency to respond "word",thus facilitating, via conscious attention,"word" responses in the Expected condi-tion; if they are dissimilar, the subjectwould have a tendency to respond "non-word", thus facilitating "nonword" re-sponses to nonword targets and inhibiting"word" responses to word targets in theUnexpected conditions.
Although the subjects in the present ex-periment were explicitly instructed not toadopt such a semantic-matching strategy(see last paragraph of Procedure section),the structure of the experiment was suchthat it would have been beneficial for themto have violated this proscription. That is,since a word target was a member of an un-expected category on only one third of theword-prime-word-target trials and since aword target followed a word prime on onlyone half of the word-prime trials, the sub-jects could have responded on the basis ofwhether the semantic features of the expec-

SEMANTIC PRIMING OF LEXICAL MEMORY 251
tancy generated to the priming word weresimilar to the semantic features activated bythe target letter string; in doing so, theywould have been correct and .would havebenefited on five sixths of the word-primetrials. Thus, given the design of the presentexperiment, the semantic-match explanationof the inhibition effect for the word-prime-word-target trials in the Unexpected condi-tions and the facilitation effect for the word-prime-nonword-target trials seems to be areasonable one. Furthermore, the fact thatthe facilitation effect for the nonword targetswas greatest at the 2,000-msec SOA sug-gests that the facilitation effect for the non-word targets depended on the commitmentof slow-acting conscious attention.
Unfortunately, the issue of whether theinhibition effect obtained for the Unexpectedword-target conditions can be solely attrib-uted to the operation of a semantic-matchingstrategy remains open because a substantialinhibition effect of 56 msec was obtainedin Condition Shift-Unexpected-Unrelated atthe 700-msec SOA, at which there was noevidence that the subject was using a se-mantic-matching strategy to facilitate hisnonword responses in the Shift nonword-target condition. In fact, at the 700-msecSOA for the nonword responses in the Shiftnonword-target condition, there was a 13-msec inhibition effect—an effect that, thoughstatistically nonsignificant, certainly did notprovide evidence for the operation of a se-mantic-matching strategy. Thus, at the 700-msec SOA, an inhibition effect occurred inCondition Shift-Unexpected-Unrelated eventhough in the Shift nonword-target condi-tion there was no evidence for the concur-rent involvement of a semantic-matchingstrategy. Despite this counterevidence at the700-msec SOA, the overall pattern of dataseems to indicate that the facilitation effectfor the nonword targets and the inhibitioneffects for the word targets in the Unex-pected conditions can 'be attributed to a se-mantic-matching strategy. Of course, theremay be alternative explanations that canaccount for the word-prime-word-target in-hibition effect and the word-prime-nonword-target facilitation effect by appealing to the
operation of only a single mechanism or ofseparate mechanisms, but one advantage ofthe present appeal to the semantic-matchingstrategy is that this strategy has also beenshown to be useful in explaining a broadrange of data obtained in other experimentsinvolving speeded retrieval of informationfrom semantic 'memory (cf. Collins & Lof-tus, 1975; Smith et al., 1974).
General Discussion
A general, model-free implication of thepresent experiment is that a priming eventcan produce two distinctly different types offacilitation effects in the processing of sub-sequent words. One kind of facilitation ef-fect 'occurred rather quickly following theprime's presentation. It was not accom-panied by an inhibition in the processing ofa target word semantically unrelated to thetarget word whose processing was facili-tated, and it depended on the prime andtarget being semantically related. The sec-ond kind of facilitation effect occurred at alater time following the prime's presentation.It was always accompanied by an inhibitionin the processing of a target word semanti-cally unrelated to the target word whoseprocessing was facilitated, and it did notdepend on the semantic relationship betweenthe prime and the target but rather de-pended on the probability with which a par-ticular class of words followed a particularpriming event within the experimental situ-ation. Thus, these two distinct facilitationeffects can be taken as support for Tulving's(1972) distinction between semantic andepisodic processes and as support for thegeneral class of theories described in theintroduction of this article. However, giventhe LaBerge (1973a, 1975), Schneider &Shiffrin (1977), and Shiffrin & Schneider(1977) results that an attention-demandingoperation can become automated with ex-tended training, if much more training hadbeen given in the present paradigm, thefacilitation effect in Condition Shift-Ex-pected-Unrelated presumably would havebecome automated and would have occurredsooner after the presentation of the primeand without a concomitant inhibition effect

252 JAMES H. NEELY
in Condition Shift-Unexpected-Unrelated.Nevertheless, the important point is thatsome brand of two-factor theory is de-manded by the present data, which .were col-lected in the early stages of training.
A second general implication of the pres-ent research is that a priming stimulus canproduce a facilitation in the subsequentprocessing of more than just itself (e.g.,Posner & Snyder, 1975b) or a few highlyrelated primary associates (e.g., Meyer &Schvaneveldt, 1971; Neely, 1976). Althoughother previous research has demonstratedthat there is an overlap in the activationpatterns produced by a category name andits exemplars, most of this previous researcheither has used only a few category exem-plars, all having high category dominance(e.g., Warren, 1972), or has involved atask in which the subject was required todecide whether the target stimulus was amember of a particular semantic category(e.g., Rosch, 1975). The present experimentis different from this previous research be-cause it employed several exemplars of lowcategory dominance in a task in which sub-jects were to decide whether the target letterstring was a word or a (pronounceable)nonword, a task that should have requireda meaning look-up (cf. James, 1975) butthat should not have necessarily requiredthat the subject classify the stimulus on thebasis of its membership in a particular se-mantic category. Consequently, the presentexperiment demonstrates that a category-name prime will facilitate the subsequentprocessing of its good and poor exemplarsto the same degree, even when they are tar-gets in a task in which the subject is notlogically required to access information oncategory membership.
This finding extends the generality of aresult obtained by Rosch (1975). She toofound that a category-name prime facili-tated the processing of its good and poorexemplars to the same degree, but she useda task in which subjects were to decidewhether two nonidentical target words wereboth members of the same semantic cate-gory. However, she also found that whenthe two target words were identical, a cate-
gory-name prime facilitated the processingof its good exemplars and inhibited the proc-essing of its poor exemplars at SOAsgreater than 400 msec, with the inhibitioneffect occurring when the subjects weremaking a category-membership decision butnot when they were making a physical-iden-tity decision. A general conclusion that canbe drawn from Roach's results is that acategory-name prime will produce an equaldegree of facilitation in the subsequent proc-essing of its good and poor exemplars when-ever the subject is making some sort ofsemantic decision and cannot respond on thebasis of physical identity. The present resultsare in accord with that general conclusion,but 'Such a conclusion raises the followingquestion: In what sense is the automaticactivation spreading if both high- and low-category-dominance items are facilitated toan equal degree? Given the available data,it is difficult to answer this question ade-quately. However, one possibility is that ifmuch shorter SOAs had been used, onewould have found that high-category-domi-nance items are facilitated earlier followingthe presentation of the category-name primethan are the low-category-dominance items.
Another general implication of the presentresearch relates to how a consciously gener-ated expectancy can be used to modulateperformance in a binary classification task.On -the basis of the word primes' facilitatingnonword responses, it can be argued thatsubjects were adopting a semantic-matchingstrategy in which they matched the semanticfeatures included in the expectancies theygenerated to the word prime with the se-mantic features activated by the target, eventhough they were explicitly told to avoidthis strategy. The fact that subjects appar-ently utilized such a strategy in the presentlexical-decision paradigm indicates that sucha matching strategy is not restricted to situa-tions in which the prime and the target arephysically identical (e.g., Posner & Snyder,1975b) nor to tasks in which the subjectsmust retrieve relatively specific semantic in-formation about a word (e.g., Collins &Loftus, 1975; Smith et al., 1974). Thus, itseems that conscious expectancies may in-

SEMANTIC PRIMING OF LEXICAL MEMORY 253
fluence performance via .some sort of gen-eral matching process, with the features onwhich the match is based being determinedby the particular stimulus materials used.
Finally, the present data have implica-tions for methodologies that have been usedto study structure in semantic memory. Onesuch commonly used methodology is thefree-classification task, in which subjectssort words into piles according to similarityin their meanings. For example, Miller(1972) has used such a procedure to deter-mine the underlying semantic structure forverbs of motion. On the basis of these free-classification results, Miller concluded thatthese verbs are categorized according torelatively subtle (at least in this author'sview) semantic features such as velocity,direction, and medium of the motion. How-ever, given that the subjects were allowed avirtually unlimited amount of time and wereforced to use some sort of categorizationschema, it does not necessarily follow thatthe verbs that were placed in the same cate-gory in such an experiment would automati-cally facilitate one another in a short-SOApriming paradigm similar to the one used inthe present experiment. That is, althoughfree-classification and similarity-rating para-digms provide information about semanticcomponents that a subject can utilize whengiven certain task demands and enough timeto make conscious comparisons and infer-ences, they may not provide informationabout semantic structures connected by auto-mated associative links. Of course, the pres-ent paradigm can be used to provide infor-mation about such automated associativelinks. If two items mutually facilitate theprocessing of each other without at the sametime inhibiting the processing of other items,these two items can be considered to belinked by a well-established automated linkin semantic memory. If, on the other hand,two items mutually facilitate the processingof each other only if they inhibit the process-ing of other items, the facilitation may beconsidered to be the result of a consciousinference and not the result of those twoitems' being connected by an automated as-sociative link in semantic memory.
Conclusion
The purpose of the foregoing general dis-cussion has been to delineate the model-freeimplications of the present research. Themain import of the data reported here isthat they seem to demand a theory that•postulates at least two independent proc-esses. They also have implications for themanner in which conscious expectancies de-velop and -for how people use expectanciesto modulate their performance in a binaryclassification task. And finally, the presentresults have methodological implications forthe study of the organization and structureof semantic memory. But equally as impor-tant as these general implications is the factthat the present data provide strong sup-port for a specific theory of attention thathas been advanced by Posner and Snyder(197Sa). When one allows for a semantic-matching strategy, the data conform almostperfectly to their theory. Thus, for the timebeing, the concepts of automatic spreadingactivation and limited-capacity conscious at-tention seem firmly enough rooted in abroad enough data base that they shouldserve as a useful point of departure for fu-ture research and theory construction.
Reference Note
1. Meyer, D. E., Schvaneveldt, R. W., & Ruddy,M. G. Activation of lexical memory. Paper pre-sented at the meeting of the Psychonomic So-ciety, St. Louis, November 1972.
References
Anderson, J. R., & Bower, G. H. Human associa-tive memory. Washington, D.C.; Winston, 1973.
Battig, W. F., & Montague, W. E. Category normsfor verbal items in 56 categories: A replicationand extension of the Connecticut category norms.Jowrnal of Experimental Psychology Mono-graph, 1969, 80(3, Pt. 2).
Clark, H. H. The language-as-fixed-effect fallacy:A critique of language statistics in psychologicalresearch. Journal of Verbal Learning and Ver-bal Behavior, 1973, 12, 335-359.
Collins, A. M., & Loftus, E. F. A spreading-activation theory of semantic processing. Psy-chological Review, 1975, 82, 407-428.
Fitts P. M. Perceptual-motor skill learning. InA. W. Melton (Ed.), Categories of humanlearning. New York: Academic Press, 1964.

254 JAMES H. NEELY
Forster, K. I., & Bednall, E. S. Terminating andexhaustive search in lexical access. Memory &Cognition, 1976, 4, 53-61.
James, C. T. The role of semantic information inlexical decisions. Journal of Experimental Psy-chology: Hitman Perception and Performance,1975, 1, 130-136.
Keele, S. W. Attention and human performance.Pacific Palisades, Calif.: Goodyear, 1973.
LaBerge, D. Attention and the measurement ofperceptual learning. Memory & Cognition, 1973,1, 268-276. (a)
LaBerge, D. Identification of the time to switchattention: A test of a serial and parallel modelof attention. In S. Kornblum (Ed.), Attentionand performance IV. New York: AcademicPress, 1973. (b)
LaBerge, D. Acquisition of automatic processingin perceptual and associative learning. In P.M. A. Rabbitt & S. Domic (Eds.), Attentionand performance V. New York: Academic Press,1975.
LaBerge, D., & Samuels, S. J. Toward a theoryof automatic information processing in reading.Cognitive Psychology, 1974, 6, 293-323.
Landauer, T. K., & Streeter, L. A. Structural dif-ferences between common and rare words: Fail-ure of equivalence assumption for theories ofword recognition. Journal of Verbal Learningand Verbal Behavior, 1973, 12, 119-131.
Loftus, E. F. Activation of semantic memory.American Journal of Psychology, 1973, 86, 331-337.
Meyer, D. E, & Schvaneveldt, R. W. Facilitationin recognizing pairs of words: Evidence of adependence between retrieval operations. Jour-nal of Experimental Psychology. 1971, 90, 227-234.
Miller, G. A. English verbs of motion: A casestudy in semantics and lexical memory. In A.W. Melton & E. Martin (Eds.), Coding proc-esses in human memory. Washington, D.C.:V. H. Winston, 1972.
Morton, J. A functional model for memory. InD. A. Norman (Ed.), Models of human memory.New York: Academic Press, 1970.
Neely, J. H. Semantic priming and retrieval fromlexical memory: Evidence for facilitatory andinhibitory processes. Memory & Cognition, 1976,4, 648-654.
Posner, M. L, & Snyder, C. R. R. Attention andcognitive control. In R. L. Solso (Ed.), In-formation processing and cognition: The Loyolasymposium. Hillsdale, NJ.: Erlbaum, 1975. (a)
Posner, M. L, & Snyder, C. R. .R. Facilitationand inhibition in the processing of signals. In
P. M. A. Rabbitt & S. Domic (Eds.), Attentionand performance V. New York: AcademicPress, 1975. (b)
Rosch, E. Cognitive representations of semanticcategories. Journal of Experimental Psychology:General, 1975, 104, 192-233.
Schneider, W., & Shiffrin, R. M. Controlled andautomatic human information processing: I. De-tection, search, and attention. Psychological Re-view, 1977, 84, 1-66.
Schvaneveldt, R. W., & Meyer, D. E. Retrievaland comparison processes in semantic memory.In S. Kornblum (Ed.), Attention and perform-ance IV. New York: Academic Press, 1973.
Shiffrin, R. M. The locus and role of attentionin memory systems. In P. M. A. Rabbitt & S.Domic (Eds.), Attention and performance V.New York: Academic Press, 1975.
Shiffrin, R. M., & Geisler, W. S. Visual recog-nition in a theory of information processing.In R. L. Solso (Ed.), Contemporary issues incognitive psychology: The Loyola symposium.Washington, D.C.: V. H. Winston, 1973.
Shiffrin, R. M., & Schneider, W. Controlled andautomatic human information processing: II.Perceptual learning, automatic attending, and ageneral theory. Psychological Review, 1977, 84,127-190.
Smith, E. E., Shoben, E. J., & Rips, L. J. Struc-ture and process in semantic memory: A fea-ture model for semantic decisions. Psychologi-cal Review, 1974, SI, 214-241.
Stanners, R. F., Jastrzembski, J. E., & Westbrook,A. Frequency and visual quality in a word-nonword classification task. Journal of VerbalLearning and Verbal Behavior, 1975, 14, 259-264.
Taft, M., & Forster, K. I. Lexical storage andretrieval of prefixed words. Journal of VerbalLearning and Verbal Behavior, 1975, 14, 638-647.
Tulving, E. Episodic and semantic memory. InE. Tulving & W. Donaldson (Eds.), Organ-ization of memory. New York: Academic Press,1972.
Turvey, M. T. Constructive theory, perceptualsystems, and tacit knowledge. In W. B. Weimer& D. S. Palermo (Eds.), Cognition and thesymbolic processes. Hillsdale, NJ.: Erlbaum,1974.
Warren, R. E. Stimulus encoding and memory.Journal of Experimental Psychology, 1972, 94,90-100.
Received January 14, 1976 •