seed fibre colour in gossypium and its possible significance in the evolution of domesticated...
TRANSCRIPT

Journal of Genetics Votume 63, Number 2 Dec. 19'77
SEED FIBRE COLOUR IN GOSSYPIUM AND ITS POSSIBLE SIGNIFICANCE IN THE EVOLUTION OF DOMESTICATED COTTONS
S. G. Stephens
Department of Genetics, North Carolina State University Raleigh, North Carolina 27607
A central element in the study of crop plant evolution is a comparison of the
cultivar with its nearest wild relatives. This not only iDvolves comparative
morphology, cytogenetics, taxonomy and other disciplines, but also consideration
of the geographicalranges of the wild and cultivated forms. I f the wild form
has all the characteristics of a "relic", i.e. existence in very small popula-
tions, occupying widely disjunct areas, then its present geographical range may
bear l i t t l e relation to the area occupied in the past. When wild forms and
cultivars belong to the same species, the evolutionary problem becomes magnified.
I t is fair ly easy to develop morphological and physiological criteria that
distinguish the wild form from the cultivar, but the same criteria are usually
not useful to distinguish authentic wild from feral types. In order to survive,
both wild and feral types must possess "wild" characteristics. These usually
involve efficient means of seed dispersal, capacity to produce large numbers of
small seeds rather than fewer numbers of large seeds, methods of delaying germi-
nation (hard seedcoatsor physiological dormancy), and perhaps also mechanisms
for regulating the flowering cycle in accordance with local habitat requirements
(photoperiodism, thermoperiodicity). Most of these characteristics contrast
sharply with those in a cultivar, grown directly or indirectly for seed production.
The nature of the problem is well illustrated by the two New World cultivated
species of cotton, G. barbadense L. and G. hirsutum L. All the primitive forms
of these species are perennial shrubs or trees, not tolerant of low temperatures
and therefore confined to tropical and sub-tropical regions.. G. barbadense
63

64 S e e d f i b r e c o I o u r in C o t t o n
occurs as a primitive cultivar in northern South America, extending through the
Antilles to Central America. G. hirsutum is found as a primitive cultivar in
Mexico, Central America, the Antilles, Venezuela and Colombia, with an outlying
pocket in northeast Brazil. "Wild" forms of both species are known: these
always occur in small localized populations in areas that seem to be well
removed from habitats associated with present human activities. They have a
curious geographical distribution. "Wild" forms of G. barbadense have been
found only in the Guayas region of northern Peru and Ecuador, and on the Galapagos
Islands. "Wild" forms of G. hirsutum, in contrast, occupy an extremely wide
geographical range. They occur in small localized populations from Venezuela
through several Caribbean islands to the Yucatan Peninsula, the Gulf Coast of
Mexico and the Florida Keys. They are also widely scattered on Pacific islands,
from the Revillagigedos in the east, Wake Island in the north, to Polynesia in
the south..Are these truly wild or feral? The question is important, because
i f one supposes them to be feral, i t is l ikely that their present distribution
has been influenced to a large extent by the migrations of human cultures. I f
truly wild, there is no necessary connection with human activities. Although
these alternatives have been studied at length (Hutchinson, 1951; Stephens, 1958,
1965, 1971; Fryxell, 1965) bearing in mind the possible relevance of the diverse
information provided by ocean currents, trade routes, prehistory and early
historical documents, there seemed to be no satisfactory way of deciding between
them. In these earlier discussions, the possible significance of seed fibre
colour in wild and cultivated cottons had been overlooked. I t is the purpose
of this paper to supply the deficiency.

S.G. Stephens 65
SEED-FIBRE COLOUR IN CULTIVATED FORMS
In cul t ivars of G. hirsutum and G. barbadense, the seeds bear two kinds of f i b re :
the long l i n t f ibres that can be removed from the seeds and spun, and an undercoat
of short "fuzz" f ibres, f i rmly adherent to the seedcoat. The fuzz may cover the
seeds ent i re ly , as in many forms of G. hirsutum, or be reduced to a small t u f t
at the micropylar end of the seed, as in most cul t ivated forms of G. barbadense.
In both species, mutants occur in which the fuzzy undercoat is absent (naked
seed types).
In both species, var iet ies in modern cu l t iva t ion have white, less frequent ly,
cream coloured l i n t . Under low magnification (circa I0 X) and with ref lected
l igh t , the individual f ibres have a character ist ic s i l ve ry ref lect ion. On the
contrary, the fuzz in such white l in ted 'forms may be drab or bright green in
colour. Green l i n t is known only as a rare mutant in Upland cottons (annual forms
of G. hirsutum). The bright green colour in the newly opened boll is retained i f
the l i n t is harvested immediately and placed in cold storage. Otherwise the
colour changes rather quickly to a dull greyish brown, par t i cu la r ly under f i e l d
exposure. Green l i n t is always associated with green fuzz, though as noted
ea r l i e r , many white l inted forms have green fuzz. In a l l pr imi t ive cu l t ivars
the green colour is confined to the fuzz, except in certain strains of G. barbadense
in which the green colour extends a short distance into the bases of the otherwise
white l i n t f ibres. These are known as "green halo" types.
Primit ive cul t ivars of both species often have brown l i n t , which is always
associated with brown fuzz. The brown colour may vary from a deep chocolate or
mahogany, through reddish brown to a l i gh t tan. Under low magnification, individual
brown l i n t f ibres show considerable variat ion both in density and d is t r ibu t ion of
the pigment. Chocolate coloured f ibres tend to be most intensely and uniformly
pigmented. Lighter coloured f ibres may show only patches of pigment that have a

66 S e e d f i b r e coLour in C o t t o n
F i g u r e 1. T h e l o c a t i o n s of b r o w n and g r e e n p i g m e n t s in the l i n t f i b r e s of two c u l t t v a r s .
(l) G. barbadense: Piura "chocolate" fibre mounted in ZO percent sodium hydroxide.
(z)
(3)
T h e s a m e , wi th f i b r e w a i l p a r t l y d i g e s t e d in s u l p h u r [ c a c i d . No te the s o l i d c o r e of p i g m e n t .
G. hirsutum: Upland "Green Lint" fibre. Transverse section mounted in ZO percent sodium hydroxide. In the photograph the green pigment appears as a broad grey zone in the wall. The lumen (black in the photograph) is devoid of pigment.

S.G. Stephens 67
brassy reflection, as contrasted with the silvery reflection of a white fibre.
Development and location of the pigments. Brown pigment is deposited in the
lumen of the fibre, while the green pigment seems to be interfaced between the
cellulose layers of the fibre wall (Kerr, 1936). The location of the brown pigment
can be seen most readily in whole mounts of chocolate coloured fibres (Figure l - l ) .
I t is preferable to make cross-sections of the fibres to locate the green pigment
(Figure I-3).
The chemical nature of the pigments is presently unknown. They only become
clearly visible in the fibres a few days before the boll opens. Younger fibres,
irrespective of their colour at maturity, are colourless'and contain large
quantities of leucoanthocyanins. Levings (unpublished) was able to identify two
of these as leucocyanidin and leucodelphinidin, by acid hydrolysis and chromato-
graphic matching with authentic samples of cyanidin and delphinidin,, respectively.
At maturity, the leucoanthocyanins disappear from green and white fibres but are
retained, at least in part, in brown fibres. I f the leucoanthocyanins are
precursors of one or both pigments, i t is possible that the lat ter may be polymerized
tannin-like substances. Further analysis is hampered by their insolubi l i ty and
d i f f icu l ty of extracting them from the fibre.
Genetic basis for seed-fibre colour. Dominant genes at a minimum of three loci
are individually capable of producing the brown pigment. Details have been
published elsewhere (Hutchinson, 1946a; Stephens, 1955; Rhyne, 1957; Endrizzi
and Kohel, 1966) and wi l l not be reviewed here. I t is worth noting, however, that
although multiple loci are involved, the usual situation is for any particular
brown linted strain to carry only one of the alternative genes that have been
identified. Thus, in crossing white and brown linted strains, as they occur in
nature, monofactorial inheritance is the rule.
In contrast to the multiple loci involved with brown pigment, only one is
known to control green colour. The L_9_gene, that produces green pigment in both
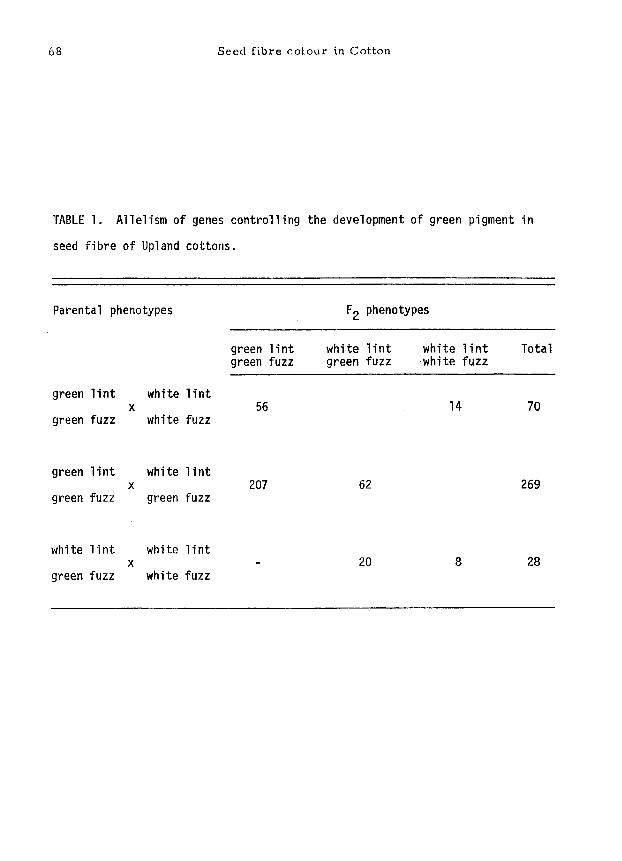
68 Seed fibre co[our in Cotton
TABLE I. Allelism of genes controlling the development of green pigment in
seed fibre of Upland cottons.
Parental phenotypes
green l in t white l in t x
green fuzz white fuzz
F 2 phenotypes
green l in t white l in t white l i n t Total green fuzz green fuzz -white fuzz
56 14 70
green l in t white l in t x
green fuzz green fuzz 207 62 269
white l in t white l in t x
green fuzz white fuzz 20 8 28

S.G. Stephens 69
l int and fuzz fibres, has only been found inUpland cottons. Earlier work on
the inheritance of green fuzz is conflicting (Harland, 1939) and the relation
of green fuzz to green l int does not seem to have been reported previously. The
data given in Table l show that in Upland cottons green l int and green fuzz are
allel ic: Lg and Ig F, respectively.
As far as I know, combinations of L~with any of the brown l in t genes are
not found in nature. In Upland cottons the experimental combination of green
and brown l int genes produces a brown phenotype which becomes a dull, dingy brown
in storage. The dingy appearance is due, presumably, to the fading and colour
change of the green pigment. In the absence of the green pigment the brown
colour remains clear and relatively bright.
SEED FIBRE COLOUR IN WILD FORMS
The seeds of wild forms of G. barbadense and G. hirsutum, like their cultivated
relatives, bear both l int and fuzz fibres, though differentiation into adherent
fuzz and non-adherent l int is not so clear-cut. Mutants which reduce or remove
the fuzz are very rare in wild forms. A type lacking both fuzz and l in t is found
in some forms of the wild Galapagos cotton, G. barbadense vat. darwinii. Hutchinson,
and in the Polynesian wild form of G. hirsutum the fuzz is very sparse or absent.
In the Asiatic cottons, G. arboreum L. and G. herbaceum L., only the latter species
is known in a wild form.
No other species of Gossypium has cultivated forms. With one exception, the
seed fibres are undifferentiated and more or less strongly adherent to the seedcoat.
The exception is the Brazilian species, G. mustelinum Watt (= G. caicoense Aranha
et al . ) , recently described by Pickersgill et al. (1975). In this species
differentiation of the seed fibres resembles that found in wild forms of
G. barbadense and G. hirsutum.
A casual inspection of the seed fibres of all wild forms of Gossypium gives
the impression that they vary from dull brown to a greyish white in colour,
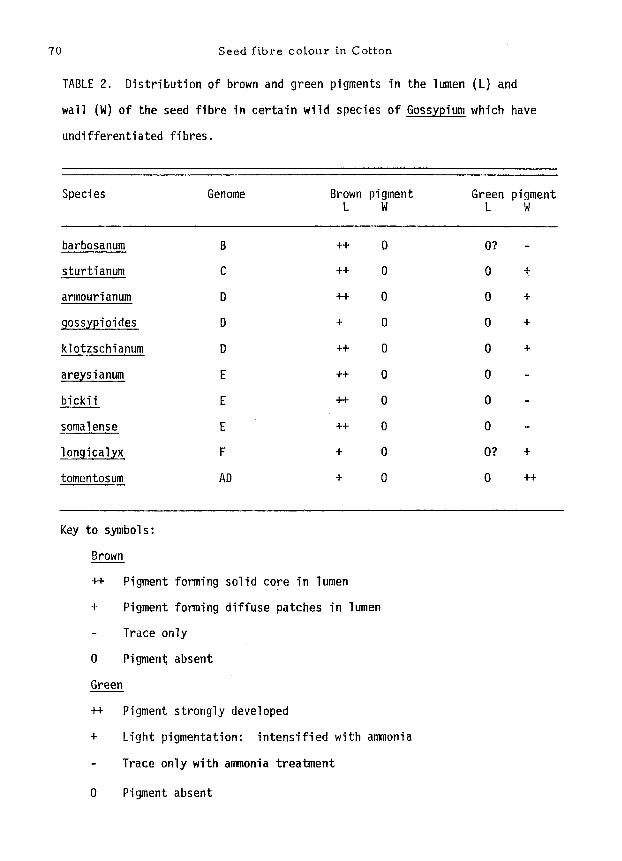
70 Seed fibre cotour in Cotton
TABLE 2. Distribution of brown and green pigments in the lumen (L) and
wall (W) of the seed fibre in certain wild species of Gossypium which have
undifferentiated fibres.
Species Genome Brown pigment Green pigment L W L W
barbosanum B ++ 0 O? -
sturtianum C ++ 0 0 +
armourianum D ++ 0 0 +
9ossypioides D + 0 0 +
klotzschianum D ++ 0 0 +
areysianum E ++ 0 0 -
bicki i E ++ 0 0 -
somalense E ++ 0 0 -
longicalyx F + 0 O? +
tomentosum AD + 0 0 ++
Key to symbols:
Brown
++ Pigment forming solid core in lumen
+ Pigment forming diffuse patches in lumen
Trace only
0 Pigment absent
Green
++ Pigment strongly developed
+ Light pigmentation: intensif ied with ammonia
- Trace only with ammonia treatment
0 Pigment absent

S . G . S t e p h e n s 71
depending on the species. For crit ical examination i t is necessary to examine
microscopically the fibres from newly-opened bolls. Samples obtained From
original collections in nature are frequently faded and discoloured through
isolation and weathering. Further loss of the original colour occurs during
storage. In some wild species, e.~. G. tomentosum Nutt. (= G. sandvicense Parl.),
G. mustelinum, G. raimon~ii Ulb., G. aridum Rose & Standley, G. armourianum Kearney,
and G. harknessii Brandegee, both green and brown colours are clearly visible
without microscopic examination. Hutchinson (1946b) extracted types with a very
dilute green l int and bright green fuzz by backcrossing G. armourianum to white
linted G. barbadense. The transfe~ed gene was shown to be linked with "crinkle"
(cr) known to be linked with Lg C When G. armourianum, G. mustelinum, G. tomentosum
and G. barbadense yar. darwinii are backcrossed to white linted G. hirsutum, green
fuzz (not. green lint~ is transferred as a simple dominant gene (Stephens,
unpublished). Thus i t is l ikely, though unproven, that the green Colour of
several wild forms is controlled by an allele at the L~locus.
In some wild forms the presence of one or both pigments is d i f f icul t to
verify without microscopic examination. The data to be presented were obtained
by mounting fibres from newly-opened bolls in 20 percent sodium hydroxide, and
examining them at c. 650 X. Sodium hydroxide expands the fibre walls and
intensifies the green pigment.
The distribution of pigments in the undifferentiated fibres of ten wild
species is summarized in Table 2, and in Table 3 similar data obtained from
eight wild forms, with fibres differentiated into fuzz and l in t , are presented.
In a rather crude fashion an attempt was made to record not only the location,
but also the intensity of the pigments, according to the key below Table 2.
Inspection of these tables leads to the following conclusions:-

7Z Seed fibre co[our in CoLton
(a) In a l l the forms examined, the f ibres contained both brown and green
pigments; whether the f ibres were undi f ferent iatea (Table 2) or d i f fe ren t ia ted
into fuzz and l i n t (Table 3).
(b) In most cases i t could be determined with confidence that brown pigment
was present in the lumen, and green pigment confined to the wal l .
(c) Both pigments tend to be more strongly developed in fuzz f ib res than
in l i n t f ibres (Table 3).
The variations in in tensi ty of brown pigmentation in a range of f ib re types are
i l lus t ra ted in Figure 2. The microphotographs were made with a yellow f i l t e r ,
and the dark areas indicate brown pigment.
TABLE 3. Distr ibut ion of brown and green pigments in the lumen (L) and
wall (W) of l i n t and fuzz f ibres in certain wi ld forms of Gossypium.
Al l forms l is ted belong to theAD genome group.
Species and type Brown pigments in Green pigments in l i n t fuzz l i n t fuzz L W L W L W L W
mustelinum - o + o o - o ++
barbadense
Guayas wild - o ++ o o + o ++
Tumbes wild - o + o o - o
vat. darwinii - o ++ o o - o ++
hirsutum
Venezuela wi ld + o + o o - o +
Puerto Rico wi ld - o + o o - o +
Yucatan wild - o + o o + o ++
Marquesas wi ld + o + o o - o +

S . G . S t e p h e n s 73
F i g u r e 2. B r o w n
(i) G.
(z) G.
{3) G.
(4) G.
{5} G.
{6) G.
(7) G.
(8) G.
{9} G.
{I0) G.
~-I l)
{iz)
E a c h division
p i g m e n t s in a r a n g e of G o s s y p i u m s e e d - f i b r e s .
a r m o u r i a n u m K e a r n e y
k l o t z s c h i a n u m A n d e r s o n
g o s s y p i o i d e s (Ulb.) S t a n d i e y
s t u r t i a n u m W i l l i s
s o m a l e n s e (Gurke ) H u t c h i n s o n
longicalyx Hutchinson & Lee
tomentos tlm Nutt
mustelinum Watt -- fuzz fibre
hirsutum race morrilli Hutchinson -- fuzz fibre
hirsutum race latifolium Hutchinson -- lint fibre
G. muste[inum Watt ~ tint fibre
G. hirsutum race morrilli Hutchinson -- lint fibre
on the scale equals 10 microns.

74 Seed fibre co[our in Cotton
DISCUSSION
The presence of both green and brown pigments in the undifferentiated fibres of
all the wild species examined (Table 2), and in the fuzz and l in t fibres of all
wild T~rms of the cultivated species (Table 3), shows that coloured fibre is an
ancestral primitive t rai t . With the outstanding exception of the green l in t
mutant (Lg) in Upland cottons, green pigment seems to have been reduced or
eliminated completely from the l in t fibres of cultivated forms. Green fuzz,
often reduced to a small tuft at the micropylar end of the seed, has been retained
in modern varieties of G. barbadense, but is found only in primitive cultivars
of G. hirsutum. When green pigment is present in both fuzz and l in t , i t is much
more strongly developed in the former. One could interpret the loss of green
pigment in cultivation as a progressive sequence: f i rs t , the loss of the pigment
from the l int fibre (as in G. barbadense) and later, the loss of the pigment from
both fuzz and l in t (as in modern varieties of G. hirsutum~.
The limited information presently available suggests that the green colour
of f~bres in several wild forms is determined by a single dominant gene. When
the gene is transferred to cultivated forms of G. barbadense, the colour is
expressed very weakly in the l in t and intensely in the fuzz fibres. Transference
of the gene to cultivated forms of G. hirsutum produces coloured fuzz only. The
gene responsible may be identical with l~ F of Upland cottons or an allele of the
same series. The reduced expression of the gene when transferred to cultivated
forms shows that the genetic milieu in which i t operates is different in wild
and cultivated forms, even when these belong to the same species. Is the
difference in genetic milieu a by-product of human selection? From the human
point of view, a green fibre is deleterious for two reasons -- (a) i t fades to
an unattractive grey colour (b) its presence in the fibre wall impairs the tensile
properties of the fibre (Neely, 1943; Richmond, 1943). Thus under human selection
there would be a cumulative tendency to select types with whiter and stronger l i n t

S.G. Stephens 75
fibres; types in which the green lint gene had a low expressiVity, or alternatively,
types in which it had mutated to white (]g). On this interpretation one would
regardS(only known as a unique mutant in Upland cottons) as a "super-green"
allele, capable of overriding the genetic milieu, h~seems to have no counterpart
in wild forms and may be a relatively recent mutant in cultivation.
The genetic basis for the development of brown pigment is more complex, in
the sense that several alternative loci are involved. Nevertheless, there is no
firm evidence thatmore than one locus is operating in any naturally occurring
type that has been investigated. In crossing brown with white linted types,
monofactorial inheritance is the rule, though the locus involved may differ from
race to race and species to species. From a modern point of view, brown l int is
as deleterious as green l int , since both have inferior fdbre strength. But
archaeological evidence (Stephens and Moseley, 1974; S~ephens, 1975) suggests that
white linted forms may not have been available during the earliest stages of
domestication. In several archaeological sites in coastal Peru, all cotton fibres
recovered in the form of raw cotton wads, seed cotton in mummy wrappings, or
textile scraps, varied in colour from light tan to dark brown. Today, dooryard
cottons with brown l int are common in Peru and Ecuador, and in living memory
naturally brown fibres have been used in texti le patterns. In contrast to the
fibres of wild forms, the colours of dooryard cottons occur in clear shades of
brown and reddish brown. The dingy brown colour of the wild fibre is missing --
presumably due to the loss of green pigment associated with wild type fibres.
I f the interpretation outlined above is substantially correct, one would
expect the fibres of authentic wild forms to contain both pigments. On the
other hand, there would seem to be no valid reason for both pigments to be
retained in feral forms. I f the green pigment was lost during the early stages
of domestication, all subsequent escapes from cultivation should be either
brown or white linted forms. All the wild forms of G. hirsutum and G. barbadense

76 S e e d fLbre coLour Ln C o t t o n
so far examined resemble the other w~Jd species of Gossypium in having fibres
that contain both pigments.
SUMMARY
Two genetic systems control the pigmentation of seed fibres in Gossypium. In
the New World amphidiploid cottons, and probably too in many of the wild diploid
species, the green colour is controlled by members of a series that includes a
minimum of three alleles: Lg (green l i n t ) , Ig F (green fuzz), Ig (white). Lg
as a mutant in Upland cottons, w h i l e ~ (or a similar al lele) is only known
is probably common to wild and primitive cultivars of G. hirsutum and G. barbadens(
G. tomentosum, G. mustelinum, and several wild diploid species. The recessive,
Ig, has not been found in any wild form of Gossypi~n.
At least three independent loci control the development of brown pigment,
but their homologies have not been analyzed systematically. No wild form of
Gossypium has been found to lack brown pigment completely.
All wild forms so far examined have fibres that contain both brown and
green pigments. I t is suggested that the loss of the green pigment and the
retention of brown pigment in the l i n t fibres of primitive cult ivars may have
been influenced by human selection. This favours the view that wild forms of
cultivated species are original ly wild -- not feral -- types.

S . G . S t e p h e n s 77
ACKNOWLEDGMENTS
Paper number 4403 of the Journal Series of the North Carolina Agr icu l tura l
Experiment Station, Raleigh, North Carolina. Work supported by NSF GB7769.
Permission to c i te the unpublished f indings of Dr. C. S. Levings, Genetics
Department, is gra te fu l ly acknowledged.
REFERENCES
ENDRIZZI, J. E. AND KOHEL, R. J. (1966). Use of telosomes in mapping three
chromosomes in cotton. Genetics 54:535-550.
FRYXELL, P. A. (1965). Stages in the evolution of Gossypium L. Advancin 9
Frontiers of Plant Sciences 10:31-56.
HARLAND, S. C. (1939). The genetics of cotton. Jonathan Cape, London. 193 pp.
HUTCHINSON, J. B. (1946a). The inheritance of brown l in t in New World cottons.
Jour. Genet. 47:295-309.
. . . . . . . . . . . . . . . . (1946b). The crinkled dwarf allelomorph series in the New
World cottons. Jour. Genet. 47:178-207.
HUTCHINSON, J. B. (1951). Intra-specific differentiation in Gossypiumhirsutum.
Heredity 5:161-193.
KERR~ T. (1936). The structure of the growth r ings in the secondary wall of
the cotton hair . Protoplasma 2_]_7:229-241.
NEELY, J. W. (1943). Relation of green l i n t to l i n t index in Upland cotton.
Jour. Agric. Res. 66:293-306.
PICKERSGILL, B., BARRETT, S.C.H., AND DARDANO DE ANDRADE-LIMA. (1975). Wild
cotton in northeast Brazi l . Biotropica ~:42-54.
RHYNE, C. L. (1957). Duplicated linkage groups in cotton as indicated by
the Y9-! and Dw loc i . Jour. Hered. 48:59-62.
RICHMOND, T. R. (1943). Inheritance of green and brown l i n t in Upland cotton.
J. Amer. Soc. Agron. 35:967-975.

78 Seed fibre co[our in Cotton
STEPHENS, S. G. (1955). Linkage inUpland cotton. Genetics 40:903-917.
. . . . . . . . . . . . . . (1958). Factors affecting seed dispersal in Gossypium.
North Carolina Agric. Expt. Sta~ Tech. Bul. No. 131:I-32.
. . . . . . . . . . . . . . (1965). The effects of domestication on certain seed and
fiber properties of perennial forms of cotton, Go.ss%piu m hirsutum L.
Amer. Nat. 49:355-371.
. . . . . . . . . . . . . . (1971). Some problems of interpretin9 transoceanic dispersal
of the New World cotton in Man across the Sea. University of Texas Press,
Austin & London. Carroll L. Riley, ed. 552 pp.
. . . . . . . . . . . . . . . (1975). A re-examination of the cotton remains from Huaca
Prieta, North Coastal Peru. Amer. Antiquity 40:406-419.
STEPHENS, S. G. AND MOSELEY, M. E. (1974). Early domesticated cottons from
archaeological sites in central coastal Peru. Amer. Antiquity 39:109-122.