seasonal variation in age structure and spatial distribution of a savanna larval anuran assemblage...
TRANSCRIPT

Society for the Study of Amphibians and Reptiles
Seasonal Variation in Age Structure and Spatial Distribution of a Savanna Larval AnuranAssemblage in Central BrazilAuthor(s): Larissa Barreto and Glória MoreiraReviewed work(s):Source: Journal of Herpetology, Vol. 30, No. 1 (Mar., 1996), pp. 87-92Published by: Society for the Study of Amphibians and ReptilesStable URL: http://www.jstor.org/stable/1564716 .Accessed: 18/10/2012 17:10
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Society for the Study of Amphibians and Reptiles is collaborating with JSTOR to digitize, preserve and extendaccess to Journal of Herpetology.
http://www.jstor.org

SHORTER COMMUNICATIONS SHORTER COMMUNICATIONS
FoREsTER, R. P. 1942. The nature of the glucose reab- sorption process in the frog renal tubule. Evidence for intermittency of glomerular function in the intact animal. J. Cell. Comp. Physiol. 20:55-69.
LAYNE, J. R., JR. 1995. Seasonal variation in the cry- obiology of Rana sylvatica from Pennsylvania. J. Therm. Biol. (in press).
, AND M. L. FIRST. 1991. Resumption of phys- iological functions in the wood frog (Rana sylva- tica) following freezing. Amer. J. Physiol. 261:R134- R137.
, AND R. E. LEE, JR. 1995. Adaptations of frogs to survive freezing. Climate Research 5:53-59.
LEE, R. E., JR., AND J. P. COSTANZO. 1993. Integrated physiological responses promoting anuran freeze tolerance. In C. Carey, G. L. Florant, B. A. Wunder, and B. Horwitz (eds.), Life in the Cold III: Eco- logical, Physiological, and Molecular Mecha- nisms, pp. 501-510. Westview Press, San Francis- co.
,AND D. L. DENLINGER (eds.). 1991. Insects at Low Temperature. Chapman and Hall, New York.
MICELLI, S., C. MASSAGLI, AND E. GALLUCCI. 1983. Se- rosal and mucosal facilita the transport of urea in urinary bladder of Bufo bufo: evidence for an al- leged water uptake. Comp. Biochem. Physiol. 75A: 157-162.
SHOEMAKER, V. H., AND K. A. NAGY. 1977. Osmo- regulation in amphibians and reptiles. Ann. Rev. Physiol. 39:449-471.
STOREY, J. M., AND K. B. STOREY. 1985. Triggering of cryoprotectant synthesis by initiation of ice nu- cleation in the freeze tolerant frog, Rana sylvatica. J. Comp. Physiol. 156:191-195.
VANATTA, J. C., AND L. W. FRAZIER. 1982. The uptake of glucose by the skin of Rana pipiens. Comp. Bio- chem. Physiol. 72A:603-606.
Accepted: 18 October 1995.
Journal of Herpetology, Vol. 30, No. 1, pp. 87-92, 1996 Copyright 1996 Society for the Study of Amphibians and Reptiles
Seasonal Variation in Age Structure and Spatial Distribution of a
Savanna Larval Anuran Assemblage in Central Brazil
LARISSA BARRETO,1' AND GL6RIA MOREmA,2 IUniversidade Federal do Maranhdo, Departamento de Biologia, Largo dos Amores, 21, 65020-240, Sao Luis, MA, Brazil and 2INPA, Departamento de Ecologia, C. P. 478, 69083-000, Manaus, AM, Brazil.
Studies of tropical larval anuran communities are important in understanding several ecological aspects of the adult phase such as demography, reproductive strategies, and habitat selection (Heyer, 1973; Crump,
FoREsTER, R. P. 1942. The nature of the glucose reab- sorption process in the frog renal tubule. Evidence for intermittency of glomerular function in the intact animal. J. Cell. Comp. Physiol. 20:55-69.
LAYNE, J. R., JR. 1995. Seasonal variation in the cry- obiology of Rana sylvatica from Pennsylvania. J. Therm. Biol. (in press).
, AND M. L. FIRST. 1991. Resumption of phys- iological functions in the wood frog (Rana sylva- tica) following freezing. Amer. J. Physiol. 261:R134- R137.
, AND R. E. LEE, JR. 1995. Adaptations of frogs to survive freezing. Climate Research 5:53-59.
LEE, R. E., JR., AND J. P. COSTANZO. 1993. Integrated physiological responses promoting anuran freeze tolerance. In C. Carey, G. L. Florant, B. A. Wunder, and B. Horwitz (eds.), Life in the Cold III: Eco- logical, Physiological, and Molecular Mecha- nisms, pp. 501-510. Westview Press, San Francis- co.
,AND D. L. DENLINGER (eds.). 1991. Insects at Low Temperature. Chapman and Hall, New York.
MICELLI, S., C. MASSAGLI, AND E. GALLUCCI. 1983. Se- rosal and mucosal facilita the transport of urea in urinary bladder of Bufo bufo: evidence for an al- leged water uptake. Comp. Biochem. Physiol. 75A: 157-162.
SHOEMAKER, V. H., AND K. A. NAGY. 1977. Osmo- regulation in amphibians and reptiles. Ann. Rev. Physiol. 39:449-471.
STOREY, J. M., AND K. B. STOREY. 1985. Triggering of cryoprotectant synthesis by initiation of ice nu- cleation in the freeze tolerant frog, Rana sylvatica. J. Comp. Physiol. 156:191-195.
VANATTA, J. C., AND L. W. FRAZIER. 1982. The uptake of glucose by the skin of Rana pipiens. Comp. Bio- chem. Physiol. 72A:603-606.
Accepted: 18 October 1995.
Journal of Herpetology, Vol. 30, No. 1, pp. 87-92, 1996 Copyright 1996 Society for the Study of Amphibians and Reptiles
Seasonal Variation in Age Structure and Spatial Distribution of a
Savanna Larval Anuran Assemblage in Central Brazil
LARISSA BARRETO,1' AND GL6RIA MOREmA,2 IUniversidade Federal do Maranhdo, Departamento de Biologia, Largo dos Amores, 21, 65020-240, Sao Luis, MA, Brazil and 2INPA, Departamento de Ecologia, C. P. 478, 69083-000, Manaus, AM, Brazil.
Studies of tropical larval anuran communities are important in understanding several ecological aspects of the adult phase such as demography, reproductive strategies, and habitat selection (Heyer, 1973; Crump,
3 Present Address: INPA, Departamento de Ecolo- gia, C. P. 478, 69083-000, Manaus, AM, Brazil.
3 Present Address: INPA, Departamento de Ecolo- gia, C. P. 478, 69083-000, Manaus, AM, Brazil.
1974; Gascon, 1991). The temporal distribution pat- terns of larval anurans have shown that species ex- hibit seasonal reproductive cycles strongly correlated to rainfall both in tropical (Crump, 1971; Aichinger, 1987; Gascon, 1991; Donnelly and Guyer, 1994), and temperate habitats (Turnipseed and Altig, 1975; Cald- well, 1987). Consequently, recruitment periods for different species will be dependent on individual re- productive strategies and variation in yearly rainfall (Gascon, 1991; Pechmann et al., 1991). Moreira and Lima (1991) observed variation on juvenile recruit- ment periods of four litter anuran species related to their reproductive modes. Caldwell (1987) observed increased recruitment in years with high rainfall for two species of chorus frogs in South Carolina. Don- nelly and Guyer (1994) noted that most recruitment of juveniles occurred during the dry periods of the year in Costa Rica. One can suppose that overall ad- aptations to seasonality in amphibian reproduction will result in a series of behavioral, physiological, and morphological characters aimed at maximizing re- cruitment and juvenile survival. One adaptation to strongly seasonal environments is rapid larval de- velopment, thereby increasing the probability of avoiding unfavorable conditions either in the larval stage or in the early juvenile stage (Herreid and Kin- ney, 1967; Koskela 1973; Keen et al., 1984). Many fac- tors will influence larval development rate (Wilbur, 1977; Travis, 1981, 1984; Semlitsch and Caldwell, 1982; Keen et al., 1984), and temperature is one of the main determinants (Herreid and Kinney, 1967; Keen et al., 1984).
In this study, we surveyed a permanent pond in the highly seasonal tropical savanna region of Central Brazil to determine age structure, recruitment period (larval and juvenile), and spatial distribution of the larval anuran community. The species studied were the hylids Scinax sp. (rubra group), Hyla albopunctata and Hyla rubicundula, the microhylid Elachistocleis bi- color, the leptodactylid Physalaemus cuvieri, and the bufonid Bufo paracnemis. The Scinax species is an un- described species of the rubra group and will be re- ferred to as Scinax sp.
This study was conducted in the Reserva Biol6gica do Jardim Botanico (RBJB), located 35 km southeast of Brasilia (15?54'S; 47050'W), Distrito Federal, Brazil. The rainy season in this region occurs from October to March, and the dry season from April to September (Camargo, 1971, see Fig. 1). The dry season is pro- nounced; humidity can fall to 20% during the day, and air temperature can drop to 8 C during the night (pers. obs.). Data were collected in a permanent grass- land pond of approximately 400 m2, near a gallery forest. This pond includes two different microhabi- tats. The shallow environment is composed of the margin and shallow areas of the pond (maximum depth in rainy season = 32 cm), with a predominance of grasses. The deep environment is represented by the open water area itself; maximum depth during the rainy season is 61cm and the area is covered by aquatic macrophytes.
Tadpoles were collected twice a month, from Jan- uary 1992 to March 1993, at 22 collecting stations spread over shallow and deep environments of the pond. Eleven stations were distributed along the mar- gins of the pond (shallow areas), and 11 stations were distributed in deep areas. Tadpoles were collected using aquatic sweep nets (0.4 m diameter opening).
1974; Gascon, 1991). The temporal distribution pat- terns of larval anurans have shown that species ex- hibit seasonal reproductive cycles strongly correlated to rainfall both in tropical (Crump, 1971; Aichinger, 1987; Gascon, 1991; Donnelly and Guyer, 1994), and temperate habitats (Turnipseed and Altig, 1975; Cald- well, 1987). Consequently, recruitment periods for different species will be dependent on individual re- productive strategies and variation in yearly rainfall (Gascon, 1991; Pechmann et al., 1991). Moreira and Lima (1991) observed variation on juvenile recruit- ment periods of four litter anuran species related to their reproductive modes. Caldwell (1987) observed increased recruitment in years with high rainfall for two species of chorus frogs in South Carolina. Don- nelly and Guyer (1994) noted that most recruitment of juveniles occurred during the dry periods of the year in Costa Rica. One can suppose that overall ad- aptations to seasonality in amphibian reproduction will result in a series of behavioral, physiological, and morphological characters aimed at maximizing re- cruitment and juvenile survival. One adaptation to strongly seasonal environments is rapid larval de- velopment, thereby increasing the probability of avoiding unfavorable conditions either in the larval stage or in the early juvenile stage (Herreid and Kin- ney, 1967; Koskela 1973; Keen et al., 1984). Many fac- tors will influence larval development rate (Wilbur, 1977; Travis, 1981, 1984; Semlitsch and Caldwell, 1982; Keen et al., 1984), and temperature is one of the main determinants (Herreid and Kinney, 1967; Keen et al., 1984).
In this study, we surveyed a permanent pond in the highly seasonal tropical savanna region of Central Brazil to determine age structure, recruitment period (larval and juvenile), and spatial distribution of the larval anuran community. The species studied were the hylids Scinax sp. (rubra group), Hyla albopunctata and Hyla rubicundula, the microhylid Elachistocleis bi- color, the leptodactylid Physalaemus cuvieri, and the bufonid Bufo paracnemis. The Scinax species is an un- described species of the rubra group and will be re- ferred to as Scinax sp.
This study was conducted in the Reserva Biol6gica do Jardim Botanico (RBJB), located 35 km southeast of Brasilia (15?54'S; 47050'W), Distrito Federal, Brazil. The rainy season in this region occurs from October to March, and the dry season from April to September (Camargo, 1971, see Fig. 1). The dry season is pro- nounced; humidity can fall to 20% during the day, and air temperature can drop to 8 C during the night (pers. obs.). Data were collected in a permanent grass- land pond of approximately 400 m2, near a gallery forest. This pond includes two different microhabi- tats. The shallow environment is composed of the margin and shallow areas of the pond (maximum depth in rainy season = 32 cm), with a predominance of grasses. The deep environment is represented by the open water area itself; maximum depth during the rainy season is 61cm and the area is covered by aquatic macrophytes.
Tadpoles were collected twice a month, from Jan- uary 1992 to March 1993, at 22 collecting stations spread over shallow and deep environments of the pond. Eleven stations were distributed along the mar- gins of the pond (shallow areas), and 11 stations were distributed in deep areas. Tadpoles were collected using aquatic sweep nets (0.4 m diameter opening).
87 87

SHORTER COMMUNICATIONS
28
26 . ,
- Jp-*
.'
O
LU w
I~
bJ
24
22
20-
16
14-
12-
J F M A M J J A S O N D J F M A 1992 * 1993-
FIG. 1. Weekly rainfall and maximum and mini- mum mean air temperatures throughout the period of study.
In order to standardize sampling effort in each en- vironment, five sweeps were done at each sampling station. Tadpoles collected were fixed in 5% formalin. In the laboratory, the body length of the tadpoles was measured using vernier calipers. All tadpole species were staged according to Gosner (1960).
Sizes and stages of each larva captured were used to verify changes in age structure of the populations studied. These changes were observed by size classes and stage distribution of tadpoles captured through- out the year. In this study, the presence of tadpoles in early development stages (25 and 26 stages, Gosner, 1960), and small sizes (3 to 7 mm) was used to define the larval recruitment period. In the same manner, the presence of tadpoles in later development stages (from 40 to 45 stages, Gosner, 1960) was used to define the juvenile recruitment period. The distribution of size classes is presented graphically only for the two species that were present in the pond during the en- tire study, Scinax sp. and Hyla albopunctata.
23 - 23-
x
z z
-1
0
0 0
92 179 177 205 215 325 171
We analyzed the spatial distribution of tadpoles using the number of tadpoles of each species collected monthly in each of the two environments. A chi- square goodness of fit test was calculated to test for an association of tadpoles with the shallow or deep environment of the pond. Yates continuity correction factor was applied, as the degree of freedom was equal to one (Zar, 1984). For small samples (N < 25), we used the binomial test presented by Siegel (1975).
We recorded vocalizations of the species weekly during the high activity period, and twice a month during the low activity period, from January 1992 to March 1993.
Tadpoles were deposited in the collection of the Instituto Nacional de Pesquisas da Amaz6nia (INPA). Daily meteorological data such as precipitation, and maximum and minimum mean air temperatures, were obtained from January 1992 to March 1993 from the Meteorological Station at the Instituto Brasileiro de Geografia e Estatistica (IBGE), located ca. 5 km of the study area. A decline in rainfall and minimum and maximum air temperatures occurred especially from July to September (Fig. 1). Rainfall decreased in June, prior to the temperature decrease.
Scinax sp. and H. albopunctata showed seasonal vari- ation in size structure of their tadpole populations (Figs. 2 and 3). Small sizes of Scinax sp. and H. albo- punctata tadpoles were captured frequently during the rainy season in both years, (approximately from January 1992 to May 1992, and from November 1992 to March 1993). A decrease in small size frequencies for H. albopunctata was observed during the dry season (from June to October 1992, in Fig. 3); the same trend was observed for Scinax sp. Scinax sp. tadpoles of in- termediate size (from 7 to 15 mm) were captured fre- quently in the drier months from May to September 1992.
Later development stages of the six species studied were only encountered during the rainy season (Fig. 4), even though two of these species (Scinax sp. and H. albopunctata) reproduced and vocalized almost con- tinuously throughout the year (Fig. 5). For Scinax sp. and H. albopunctata, larval recruitment occurred every year, but juvenile recruitment occurred only during the rainy season (Fig. 4A, B). For the other four spe- cies, larval and juvenile recruitment only occurred during the rainy season.
173 96 104 60 41 103 106 144
19
I 5-
I I
7-
3- I
I ,I I I I 1 I I !I I
JAN FEB MAR APR MAY JUN JUL AUG SEP OCT NOV DEC JAN FEB MAR
1992 1993
FIG. 2. Size distribution of Scinax sp. tadpoles captured monthly from January 1992 to March 1993. The total number of individuals collected is indicated at the top of the figure. Bar width is proportional to the number of individuals collected.
Ils U n IC . 0~0
Min
t f
88
cl w
(A
Qt
ILJ
un
WR
,A ̂ S
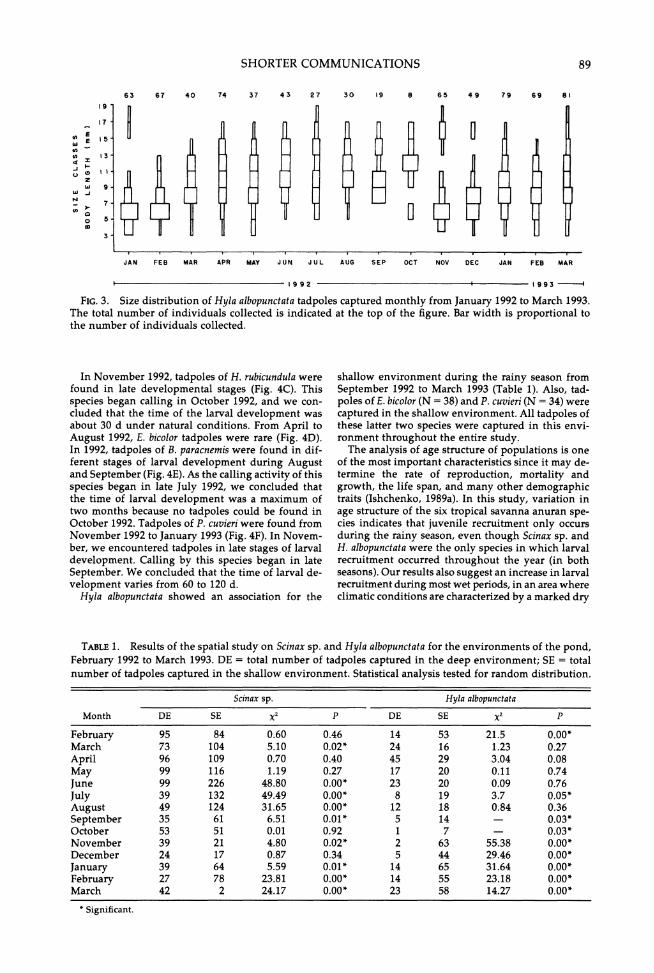
SHORTER COMMUNICATIONS
63 67 40 74 37 43 27 30 19 8 65 49 79 69 81
- 3 i
^"Cr^^ n4 JAN FE MAR PR MAY JUN JULSEP OCT NOV DEC AN FEB MAR JAN FEB MAR APR MAY JUN J U L AUG SEP OCT NOV DEC JAN FEB MAR
1992 I 993 -
FIG. 3. Size distribution of Hyla albopunctata tadpoles captured monthly from January 1992 to March 1993. The total number of individuals collected is indicated at the top of the figure. Bar width is proportional to the number of individuals collected.
In November 1992, tadpoles of H. rubicundula were found in late developmental stages (Fig. 4C). This species began calling in October 1992, and we con- cluded that the time of the larval development was about 30 d under natural conditions. From April to August 1992, E. bicolor tadpoles were rare (Fig. 4D). In 1992, tadpoles of B. paracnemis were found in dif- ferent stages of larval development during August and September (Fig. 4E). As the calling activity of this species began in late July 1992, we concluded that the time of larval development was a maximum of two months because no tadpoles could be found in October 1992. Tadpoles of P. cuvieri were found from November 1992 to January 1993 (Fig. 4F). In Novem- ber, we encountered tadpoles in late stages of larval development. Calling by this species began in late September. We concluded that the time of larval de- velopment varies from 60 to 120 d.
Hyla albopunctata showed an association for the
shallow environment during the rainy season from September 1992 to March 1993 (Table 1). Also, tad- poles of E. bicolor (N = 38) and P. cuvieri (N = 34) were captured in the shallow environment. All tadpoles of these latter two species were captured in this envi- ronment throughout the entire study.
The analysis of age structure of populations is one of the most important characteristics since it may de- termine the rate of reproduction, mortality and growth, the life span, and many other demographic traits (Ishchenko, 1989a). In this study, variation in age structure of the six tropical savanna anuran spe- cies indicates that juvenile recruitment only occurs during the rainy season, even though Scinax sp. and H. albopunctata were the only species in which larval recruitment occurred throughout the year (in both seasons). Our results also suggest an increase in larval recruitment during most wet periods, in an area where climatic conditions are characterized by a marked dry
TABLE 1. Results of the spatial study on Scinax sp. and Hyla albopunctata for the environments of the pond, February 1992 to March 1993. DE = total number of tadpoles captured in the deep environment; SE = total number of tadpoles captured in the shallow environment. Statistical analysis tested for random distribution.
Scinax sp. Hyla albopunctata Month DE SE x2 P DE SE x2 P
February 95 84 0.60 0.46 14 53 21.5 0.00* March 73 104 5.10 0.02* 24 16 1.23 0.27 April 96 109 0.70 0.40 45 29 3.04 0.08 May 99 116 1.19 0.27 17 20 0.11 0.74 June 99 226 48.80 0.00* 23 20 0.09 0.76 July 39 132 49.49 0.00* 8 19 3.7 0.05* August 49 124 31.65 0.00* 12 18 0.84 0.36 September 35 61 6.51 0.01* 5 14 - 0.03* October 53 51 0.01 0.92 1 7 - 0.03* November 39 21 4.80 0.02* 2 63 55.38 0.00* December 24 17 0.87 0.34 5 44 29.46 0.00* January 39 64 5.59 0.01* 14 65 31.64 0.00* February 27 78 23.81 0.00* 14 55 23.18 0.00* March 42 2 24.17 0.00* 23 58 14.27 0.00*
* Significant.
w <n
UJ
N
-1
U)
19
17 -
5-
13
9-
7-
5-
3-
E E
I- (9 z
0 O g
89
I I

SHORTER COMMUNICATIONS
45-
40-
35-
30-
25-
Scinox sp.
I
() w 3-
I I
40-
35-
30-
25- . I
V t I II I I I I I I II I -
I J FM A MJ J A S 0 N D J F M
1 1992 I 1993-I
47-
45-
43- 41-
39-
37-
35- 33-
31-
29- 27-
25-
? H. rubiaundula
I 47- @ 45-
() w
9 C/)
43- 41-
39- 37-
35-
33-
31-
29-
27-
25-
H. albopuncta
I I I I I I J F M A M J J A S 0 N D J F M
1 1992 I 1993-I
E. bicolor
I I. 1
F N1D J F - 1992 1 1993-1
B. parcnemis
I I V) w
C,,
M A M J A S O N D F
i 1992 1 1993
41-
39-
37-
35-
33-
31-
29-
27-
25-
I I A I I ID A M J J A S 0 N D
? P. cuvieri
J F MAMJ J AS I
O I I
i IM J F M A M J J A S 0 N D J F M
t 1992 t 1992 1993-
FIG. 4. Variation of larval development stages encountered from January 1992 to March 1993. A. Scinax
sp., B. Hyla albopunctata, C. Hyla rubicundula, D. Elachistocleis bicolor, E. Bufo paracnemis, F. Physalaemus cuvieri. The dots represent the presence of tadpoles in only one development stage.
season. In fluctuating environments, larval growth and development strategies are directly linked with future reproductive success of the individual and, consequently, with the stability of the population (Ishchenko, 1989b). Rainfall distribution may also in- fluence the recruitment period in some amphibians with aquatic tadpoles (Semlitsch and Wilbur, 1988). Berven (1990) observed an increase in juvenile sur- vival of wood frogs (Rana sylvatica) in wet periods. Based on this information, we project that larval and
juvenile recruitment in anurans can be related to many other factors such as regional climatic conditions (e.g., rainfall and/or temperature), pond hydroperiod, etc. These hypotheses were not tested.
It was not possible to determine the length of larval development under natural conditions for Scinax sp. and H. albopunctata because tadpoles in early devel- opment stages were normally encountered year-round. Nevertheless, tadpoles of H. albopunctata were only encountered in stages 25 and 26 during approxi-
U) uJ 4
Un
cn Ii
40-
30-
20-
UJ 4 1- (0
10-
2u I . . . . . . . . . . . . .
I , I I II I I I I
90
I

SHORTER COMMUNICATIONS
EB-
PC-
0 w 0 a
a BP-
HA-
SR-
HR-
I I I I I I I I t I I I I I I
FMAM J JASO NDJ FMA 1992 1993
FIG. 5. Vocalization activity period of anurans studied from January 1992 to March 1993. HR = Hyla rubicundula, SR = Scinax sp., HA = Hyla albopunctata, BP = Bufo paracnemis, PC = Physalaemus cuvieri, EB = Elachistocleis bicolor.
mately 6 months (from February to July 1992). This suggests that the larval development period of this species is prolonged.
Differences in morphological, physiological, and behavioral characteristics of species are among the major factors that influence the distribution of the tadpoles in ponds. In our study, Scinax sp. and H. albopunctata tadpoles showed seasonal variation in spatial distribution. There was a clear association of H. albopunctata with the shallow environment during the rainy season, whereas E. bicolor and P. cuvieri tad- poles were found in this environment during the entire study. The association of H. albopunctata, E. bi- color, and P. cuvieri tadpoles with the shallow envi- ronment might be related to the selection of micro- habitats with higher temperatures. Andrade (1987) observed the preference of H. albopunctata tadpoles for shallow and warm environments. She suggested that the selection of these microhabitats might favor higher body metabolism and consequently decrease the time to metamorphosis.
Some studies suggest that anuran tadpoles seem to select different temperatures among microhabitats (e.g., Lucas and Reynolds, 1967; Woolmuth and Crawshaw, 1988). However, other factors of natural habitats such as sunlight, predators, parasites, food, water depth, and oxygen tension can modify habitat use of larval amphibians (Lucas and Reynolds, 1967; Resetarits and Wilbur, 1991). Different habitat pref- erences could also arise due to differing physiological requirements among species or developmental stages of tadpoles. It could also be the result of differences in predation pressure among microhabitats (reviewed in Alford, 1986), as well as competitive pressures, since microhabitat selection may decrease competi- tion (Schoener, 1974).
Acknowledgments.-This paper represents part of a master's thesis by Larissa Barreto presented to the program of Ecology (University of Brasilia). We are grateful to A. Sebben, C. A. Schwartz, E. N. Ferroni, W. Oliveira, W. C. Silveira and D. Curado for labo- ratory assistance; and G. V. Andrade for helpful com- ments on the manuscript. C. Gascon provided valu- able suggestions in this study. We also thank T. Oliv- eira for revisions in English. Evaldo Azuelos and Glaucio da Silva (INPA) drew the figures. This re- search was supported in part by funds from WWF
(Brasilia-BR/grant number SR-024). Additional sup- port was provided by a governmental foundation of the Maranhao State (FundaCao de Amparo a Pesquisa do Maranhao).
LITERATURE CITED
AICHINGER, M. 1987. Annual activity patterns of anu- rans in a seasonal Neotropical environment. Oec- ologia (Berlin) 71:583-592.
ALFORD, R. A. 1986. Habitat use and positional be- havior of anuran larvae in a Northern Florida tem- porary pond. Copeia 1986:408-423.
ANDRADE, G. V. 1987. Reproducao e vida larvaria de anuros (Amphibia) em poSa de area aberta na Serra do Japi, Estado de Sao Paulo. Unpubl. M.S. Thesis, Campinas State Univ., Sao Paulo.
BERVEN, K. A. 1990. Factors affecting population fluc- tuations in larval and adult stages of the wood frog Rana sylvatica Ecology 7:1599-1608.
CALDWELL, J. P. 1987. Demography and life history of two species of chorus frogs (Anura: Hylidae) in South Carolina. Copeia 1987:114-127.
CAMARGO, A.G. 1971. Clima do Cerrado. In Simp6sio Sobre o Cerrado, Edgard Blucher publ., Sao Paulo.
CRUMP, M. L. 1971. Quantitative analysis of the eco- logical distribution of a tropical herpetofauna. Misc. Publ. Univ. Kansas Mus. Nat. Hist. 3:1-62.
. 1974. Reproductive strategies in a tropical anuran community. Misc. Publ. Univ. Kansas Mus. Nat. Hist. 61:1-68.
DONNELLY, M. A., AND C. GUYER. 1994. Patterns of reproduction and habitat use in an assemblage of Neotropical hylid frogs. Oecologia (Berlin) 98:291- 302.
DUELLMAN, W. E., AND L. TRUEB. 1986. Biology of Amphibians. McGraw-Hill, New York.
GASCON, C. 1991. Population- and community-level analyses of species ocurrences of Central Ama- zonian rainforest tadpoles. Ecology 72:1731-1746.
GOSNER, K. L. 1960. A simplified table for staging anuran embryos and larvae with notes on iden- tification. Herpetologica 16:183-190.
HEYER, W. R. 1973. Ecological interactions of frog larvae at a seasonal tropical location in Thailand. J. Herpetol. 7:337-361.
HERREID, C. F. II, AND S. KINNEY. 1967. Survival of Alaskan woodfrog Rana sylvatica larvae. Ecology 47:1039-1041.
ISHCHENKO, V. G. 1989a. Population biology of am- phibians. Sov. Sci. Rev. F. Phisiol. Gen. Biol. 1989: 119-155.
. 1989b. Ecological mechanisms for ensuring stability in a population of amphibians. Soviet Ecol. 20:69-75.
KEEN, W. H., J. TRAVIS, AND J. JUILIANNA. 1984. Larval growth in three sympatric Ambystoma salamander species: species differences and the effects of tem- perature. Can. J. Zool. 62:1043-1047.
KOSKELA, P. 1973. Duration of the larval stage, growth and migration in Rana temporaria in two ponds in Northern Firland in relation to environmental fac- tors. Ann. Zool. Fennici 10:414-418.
LUCAS, E. A., AND W. A. REYNOLDS. 1967. Temper- ature selection by amphibian larvae. Physiol. Zool. 40:159-171.
MOREIRA, G., AND A. LIMA. 1991. Seasonal patterns of juvenile recruitment and reproduction in four
91

SHORTER COMMUNICATIONS SHORTER COMMUNICATIONS
species of leaf litter frogs in central Amaz6nia. Herpetologica 47:295-300.
PECHMANN, J. H. K., D. E. SCOTT, R. D. SEMLITSCH, J. P. CALDWELL, L. J. VIrr, AND J. W. GIBBONS. 1991. Declining amphibian populations: the problem of separating humans impacts from natural fluctua- tions. Science 253:892-895.
RESETARITS, W. J., JR., AND H. M. WILBUR. 1991. Call- ing site choice by Hyla chrysoscelis: effect of pred- ators, competitors, and oviposition sites. Ecology 72:778-786.
SCHOENER, T. W. 1974. Resource partitioning in eco- logical communities. Science 185:27-39.
SEMLITSCH, R. D., AND J. P. CALDWELL. 1982. Effects of density on growth, metamorphosis, and sur- vivorship in tadpoles of Scaphiopus holbrooki. Ecol- ogy 63:905-911.
, AND H. M. WILBUR. 1988. Effects of pond drying time on the metamorphosis and survival in the salamander Ambystoma talpoideum. Copeia 1988:978-983.
SIEGEL, S. 1975. Estatistica Nao-Param6trica para as Ciencias do Comportamento. McGraw-Hill do Bra- sil, Sao Paulo.
TRAVIS, J. 1981. Control of larval growth variation in a population of Pseudacris triseriata (Anura: Hy- lidae). Evolution 35:423-432.
. 1984. Anuran size at metamorphosis: ex- perimental test of a model based on intraspecific competition. Ecology 65:1155-1160.
TURNIPSEED, G., AND R. ALTIG. 1975. Population den- sity and age structure of three species of hylid tadpoles. J. Herpetol. 9:287-291.
WILBUR, H. M. 1977. Density-dependent aspects of growth and metamorphosis in Bufo americanus. Ecology 58:196-200.
WOLLMUTH, L. P., AND L. I. CRAWSHAW. 1988. The effect of development and season on temperature selection in bullfrog tadpoles. Physiol. Zool. 61: 461-469.
ZAR, J. H. 1984. Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, New Jersey.
Accepted: 19 October 1995.
Journal of Herpetology, Vol. 30, No. 1, pp. 92-94, 1996 Copyright 1996 Society for the Study of Amphibians and Reptiles
Food Habits of an Alpine Population of the Sand Viper (Vipera ammodytes)
LUCA LUISELLI, Department of Animal and Human Biol- ogy, University of Rome "La Sapienza," via A. Borelli 50, 1-00161 Rome, Italy.
The diet of the European vipers (genus Vipera), ex- cept for the primarily insectivorous Vipera ursinii (Agrimi and Luiselli, 1992; Baron, 1992), is character- ized by a remarkable ontogenetic change: juveniles feed mainly on ectothermic prey (mainly lizards), whereas adults feed on homeothermic prey (mainly small mammals) (Saint Girons, 1979, 1980, 1983; Bea et al., 1992). The most intensively studied species (Vi- pera berus and Vipera aspis) did not show any remark-
species of leaf litter frogs in central Amaz6nia. Herpetologica 47:295-300.
PECHMANN, J. H. K., D. E. SCOTT, R. D. SEMLITSCH, J. P. CALDWELL, L. J. VIrr, AND J. W. GIBBONS. 1991. Declining amphibian populations: the problem of separating humans impacts from natural fluctua- tions. Science 253:892-895.
RESETARITS, W. J., JR., AND H. M. WILBUR. 1991. Call- ing site choice by Hyla chrysoscelis: effect of pred- ators, competitors, and oviposition sites. Ecology 72:778-786.
SCHOENER, T. W. 1974. Resource partitioning in eco- logical communities. Science 185:27-39.
SEMLITSCH, R. D., AND J. P. CALDWELL. 1982. Effects of density on growth, metamorphosis, and sur- vivorship in tadpoles of Scaphiopus holbrooki. Ecol- ogy 63:905-911.
, AND H. M. WILBUR. 1988. Effects of pond drying time on the metamorphosis and survival in the salamander Ambystoma talpoideum. Copeia 1988:978-983.
SIEGEL, S. 1975. Estatistica Nao-Param6trica para as Ciencias do Comportamento. McGraw-Hill do Bra- sil, Sao Paulo.
TRAVIS, J. 1981. Control of larval growth variation in a population of Pseudacris triseriata (Anura: Hy- lidae). Evolution 35:423-432.
. 1984. Anuran size at metamorphosis: ex- perimental test of a model based on intraspecific competition. Ecology 65:1155-1160.
TURNIPSEED, G., AND R. ALTIG. 1975. Population den- sity and age structure of three species of hylid tadpoles. J. Herpetol. 9:287-291.
WILBUR, H. M. 1977. Density-dependent aspects of growth and metamorphosis in Bufo americanus. Ecology 58:196-200.
WOLLMUTH, L. P., AND L. I. CRAWSHAW. 1988. The effect of development and season on temperature selection in bullfrog tadpoles. Physiol. Zool. 61: 461-469.
ZAR, J. H. 1984. Biostatistical Analysis. Prentice-Hall, Englewood Cliffs, New Jersey.
Accepted: 19 October 1995.
Journal of Herpetology, Vol. 30, No. 1, pp. 92-94, 1996 Copyright 1996 Society for the Study of Amphibians and Reptiles
Food Habits of an Alpine Population of the Sand Viper (Vipera ammodytes)
LUCA LUISELLI, Department of Animal and Human Biol- ogy, University of Rome "La Sapienza," via A. Borelli 50, 1-00161 Rome, Italy.
The diet of the European vipers (genus Vipera), ex- cept for the primarily insectivorous Vipera ursinii (Agrimi and Luiselli, 1992; Baron, 1992), is character- ized by a remarkable ontogenetic change: juveniles feed mainly on ectothermic prey (mainly lizards), whereas adults feed on homeothermic prey (mainly small mammals) (Saint Girons, 1979, 1980, 1983; Bea et al., 1992). The most intensively studied species (Vi- pera berus and Vipera aspis) did not show any remark-
able geographic variation in this general feeding pat- tern (Luiselli and Agrimi, 1991; Luiselli and Anibaldi, 1991; Monney, 1990, 1993; Luiselli et al., 1995). Data on the sand viper (Vipera ammodytes), one of the larg- est and highly venomous European vipers, are very scarce and mostly anecdotal (see Bruno, 1983), despite its broad geographic distribution (Arnold and Burton, 1978). In this paper I give detailed information on the feeding habits of a population of sand vipers of a mountainous locality in northeastern Italy.
All data were collected between 1987 and 1994 si- multaneously to other research on alpine snakes car- ried out in the Carnic Alps (Capula and Luiselli, 1994). The study area (Pontebba, about 750 m a.s.l., Carnic Alps, Province of Udine) is characterized by detrital- alluvial cones, with abundant stones and rocky spots, at the limits of Abieti-Fagetum woods, where sand vi- pers are locally quite common (average density = 15- 22 adults per ha). Vipers were captured by hand, sexed by analyzing external tail morphology and coloration features, palpated to obtain stomach or fecal contents, measured for length (total length, to the nearest 0.1 cm) and body mass (to the nearest 0.1 g), permanently marked by scale-clipping, and then released. When the food item was in very good condition, its biomass (fresh weight) was measured. For testing whether di- etary ontogenetic change occurs in this taxon, I di- vided the captured vipers into three size categories: (1) specimens of 18-30 cm TL (0-2 yrs of age), (2) specimens of 30-50 cm TL (2-4 yr of age), (3) speci- mens of more than 50 cm TL (>4 yr of age). The respective niche breadths were measured using Lev- ins (1968) index (parameter B,), and the niche overlap between two categories using Pianka's (1973) sym- metric equation (parameter Ojk). Age determination was based on mark-recapture records.
Statistical analyses were done on the SYSTAT-PC package (Wilkinson, 1988), with a set at 5%. Means are followed by +one standard deviation.
I examined 141 vipers, 38 belonging to size category (1), 43 to size category (2) and 60 to size category (3). From these vipers, I obtained 24,28, and 52 food items respectively (pooling both fecal pellets and stomach contents). The relative number of snakes with and without prey differed significantly among the three size-classes of snakes (X2 = 9.03, df = 3, P = 0.011), with the largest snake category differing from both of the other two in pairwise comparisons.
The mean TL of 77 males was 45.8 + 17.27 cm, whereas the mean TL of 65 females was 48.1 + 17.87. These means did not differ significantly (t-test, df = 139, P > 0.4). The largest male was 78.1 cm long, and the largest female was 76.3 cm long.
Dietary data are summarized in Table 1. There were significant differences in the dietary composition of the three size categories (P < 0.001, X2 test). Lacertid lizards represented 100% of the diet of juvenile vipers (category 1), 82.2% of the diet of subadult vipers (cat- egory 2), and 9.5% of the diet of adult vipers (category 3). Conversely, small mammals were not preyed at all by vipers of size category (1), but represented 17.8% of the diet of vipers of category (2) and 75.1% of the diet of vipers of category (3). The food niche of adult vipers was less specialized than that of juveniles (B, = 0.297 versus 0.138) but more than that of subadults (B, = 0.600). Niche overlap was very high between juvenile and subadult vipers (Ojk = 0.966), but low between subadult and adult vipers (O,j = 0.297) and
able geographic variation in this general feeding pat- tern (Luiselli and Agrimi, 1991; Luiselli and Anibaldi, 1991; Monney, 1990, 1993; Luiselli et al., 1995). Data on the sand viper (Vipera ammodytes), one of the larg- est and highly venomous European vipers, are very scarce and mostly anecdotal (see Bruno, 1983), despite its broad geographic distribution (Arnold and Burton, 1978). In this paper I give detailed information on the feeding habits of a population of sand vipers of a mountainous locality in northeastern Italy.
All data were collected between 1987 and 1994 si- multaneously to other research on alpine snakes car- ried out in the Carnic Alps (Capula and Luiselli, 1994). The study area (Pontebba, about 750 m a.s.l., Carnic Alps, Province of Udine) is characterized by detrital- alluvial cones, with abundant stones and rocky spots, at the limits of Abieti-Fagetum woods, where sand vi- pers are locally quite common (average density = 15- 22 adults per ha). Vipers were captured by hand, sexed by analyzing external tail morphology and coloration features, palpated to obtain stomach or fecal contents, measured for length (total length, to the nearest 0.1 cm) and body mass (to the nearest 0.1 g), permanently marked by scale-clipping, and then released. When the food item was in very good condition, its biomass (fresh weight) was measured. For testing whether di- etary ontogenetic change occurs in this taxon, I di- vided the captured vipers into three size categories: (1) specimens of 18-30 cm TL (0-2 yrs of age), (2) specimens of 30-50 cm TL (2-4 yr of age), (3) speci- mens of more than 50 cm TL (>4 yr of age). The respective niche breadths were measured using Lev- ins (1968) index (parameter B,), and the niche overlap between two categories using Pianka's (1973) sym- metric equation (parameter Ojk). Age determination was based on mark-recapture records.
Statistical analyses were done on the SYSTAT-PC package (Wilkinson, 1988), with a set at 5%. Means are followed by +one standard deviation.
I examined 141 vipers, 38 belonging to size category (1), 43 to size category (2) and 60 to size category (3). From these vipers, I obtained 24,28, and 52 food items respectively (pooling both fecal pellets and stomach contents). The relative number of snakes with and without prey differed significantly among the three size-classes of snakes (X2 = 9.03, df = 3, P = 0.011), with the largest snake category differing from both of the other two in pairwise comparisons.
The mean TL of 77 males was 45.8 + 17.27 cm, whereas the mean TL of 65 females was 48.1 + 17.87. These means did not differ significantly (t-test, df = 139, P > 0.4). The largest male was 78.1 cm long, and the largest female was 76.3 cm long.
Dietary data are summarized in Table 1. There were significant differences in the dietary composition of the three size categories (P < 0.001, X2 test). Lacertid lizards represented 100% of the diet of juvenile vipers (category 1), 82.2% of the diet of subadult vipers (cat- egory 2), and 9.5% of the diet of adult vipers (category 3). Conversely, small mammals were not preyed at all by vipers of size category (1), but represented 17.8% of the diet of vipers of category (2) and 75.1% of the diet of vipers of category (3). The food niche of adult vipers was less specialized than that of juveniles (B, = 0.297 versus 0.138) but more than that of subadults (B, = 0.600). Niche overlap was very high between juvenile and subadult vipers (Ojk = 0.966), but low between subadult and adult vipers (O,j = 0.297) and
92 92