seasonal radiation interception, dry matter production and yield determination for a semi-leafless...
TRANSCRIPT

Ann. appl. Biol. (1987), 110, 413420 Printed in Great Brituin
413
Seasonal radiation interception, dry matter production and yield determination for a semi-leafless pea (Pisum safiuum) breeding selection under contrasting field conditions
BY M. C. HEATH* AND P. D. HEBBLETHWAITE Depurtment oj’ Agriculture and Horticulture, University of Nottingham, School of Agriculture,
Sutton Ronington, Loughborough, Leics. LEl2 SRD
(Accepted 4 September 1986)
SUMMARY
Radiation interception, dry matter accumulation, flower and pod production and yield were measured for a semi-leafless pea (Pisum satiuum) breeding selection (BS3) on three contrasting sites. Differences in soil moisture availability were largely responsible for a three-fold difference in yield between sites.
Radiation interception was related to dry matter production by calculating photosynthetic efficiencies. In the absence of lodging, crop canopies converted intercepted radiation into dry matter with constant efficiency ( E ) throughout the season; under conditions of moisture stress E was reduced. Serious lodging during the post-flowering period on one site resulted in a mean seasonal photosynthetic efficiency (E) 17% lower than E .
The ability of the pea crop canopy to intercept radiation was related also to yield components.
INTRODUCTION
A major agronomic problem associated with the combining (dried) pea (Pisum sutivurn L.) crop is inherent sensitivity to extremes of soil moisture contributing to yield variability (Heath & Hebblethwaite, 19851). The aim of this study was to measure growth, development and yield of combining peas at three contrasting sites differing in soil moisture availability and to relate these variables, where possible, to radiation interception.
One promising, semi-leafless breeding selection was chosen for study. Semi-leafless peas are currently of interest because of their reduced sensitivity to lodging (Heath & Hebblethwaite, 1984; 1985a).
MATERIALS A N D METHODS
One white, round-seeded, semi-leafless combining pea (Pisum sativum L.) breeding selection (BS3) developed by the John Innes Institute, Norwich was grown in c. 150 m2 plots on three sites of contrasting soil type at the University of Nottingham, School of Agriculture Farm. Plots were sown with a Nordsten combine drill at a target density of 100 plants m-2 on 1 April 1982 following spring cultivations.
The first site at Sutton Bonington (SB) consisted of a coarse, gravelly, sandy loam (Arrow series). The other two sites were located at Bunny Park Farm, approximately 11 km from Sutton Bonington. Bunny Park ~ Site A (BP-A) consisted of a shallow, surface-compacted, ‘droughty’ silty-clay (Worcester series, shallow phase) frequently subject to moisture stress
8TQ * Present address: MAFF/ADAS, Midlands and Western Regional Office, Woodthorne, Wolverhampton. WV6
6 1987 Association of Applied Biologists
414 M . C. H E A T H A N D P . D . HEBBLETHWAITE
throughout the season (Heath & Hebblethwaite, 19853). Bunny Park - Site B (BP-B) consisted of a deep, water-retentive clay loam (Worcester series, colluvial phase). The two sites at Bunny Park were located 0.1 km apart; BP-A was situated on an exposed knoll, BP-B at the foot of a gentle slope, close to a brook. Effective rooting depths measured using a modified'Wallingford'neutron probe (Bell, 1973; Long & French, 1967) were: SB, 30 cm; BP- A, 30cm and BP-B, > 5Ocm.
Intercepted radiation measurement and growth analysis methodology followed Heath & Hebblethwaite (19856). Intercepted radiation was measured until final growth analysis samples were harvested. Plot areas were combine harvested 6 - 7 days after final growth analysis. Flower and pod production were monitored on six labelled plants within each crop. The number of pods retained at plant maturity were assessed from three areas of 1.2 m2.
RESULTS
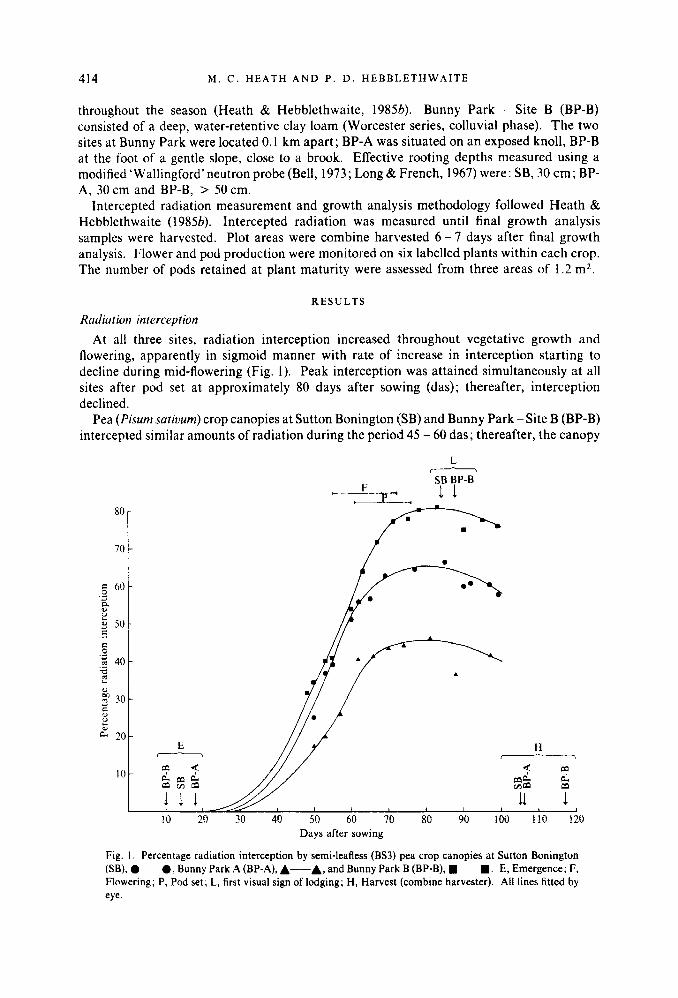
Radiution interception At all three sites, radiation interception increased throughout vegetative growth and
flowering, apparently in sigmoid manner with rate of increase in interception starting to decline during mid-flowering (Fig. I). Peak interception was attained simultaneously at all sites after pod set at approximately 80 days after sowing (das); thereafter, interception declined.
Pea (Pisurn satiourn) crop canopies at Sutton Bonington (SB) and Bunny Park - Site B (BP-B) intercepted similar amounts of radiation during the period 45 - 60 das; thereafter, the canopy
80 -
70 -
g 6 0 - .- a n U ; 5 0 - .- C .- .- 2 4 0 -
2
a 30- E 2
U
0 00
I
k 20-
10 -
/i A- /r' / A A
E H
d F -
2 gLa& E% m ! l l 11 1
I 10 20 30 40 50 60 70 80 90 100 110 120
///
Days after sowing
Fig. I . Percentage radiation interception by semi-leafless (BS3) pea crop canopies at Sutton Bonington (SB), 0-0, Bunny Park A (BP-A), A-A, and Bunny Park B (BP-B), .-. . E, Emergence; F, Flowering; P, Pod set; L, first visual sign of lodging; H, Harvest (combine harvester). All lines fitted by eye.
Radiation interception and crop growth in pea 415
700
600
500 - E on 400
2
$ 300
v
M .-
6 200
100
at BP-B intercepted considerably more radiation than the canopy at SB (Fig. 1). The crop canopy at Bunny Park - Site A (BP-A) intercepted considerably less radiation than the other two sites throughout the season.
- = 1 1
-
-
-
-
-
-
- 10 20 30 40 SO 60 70 80 90 100 110
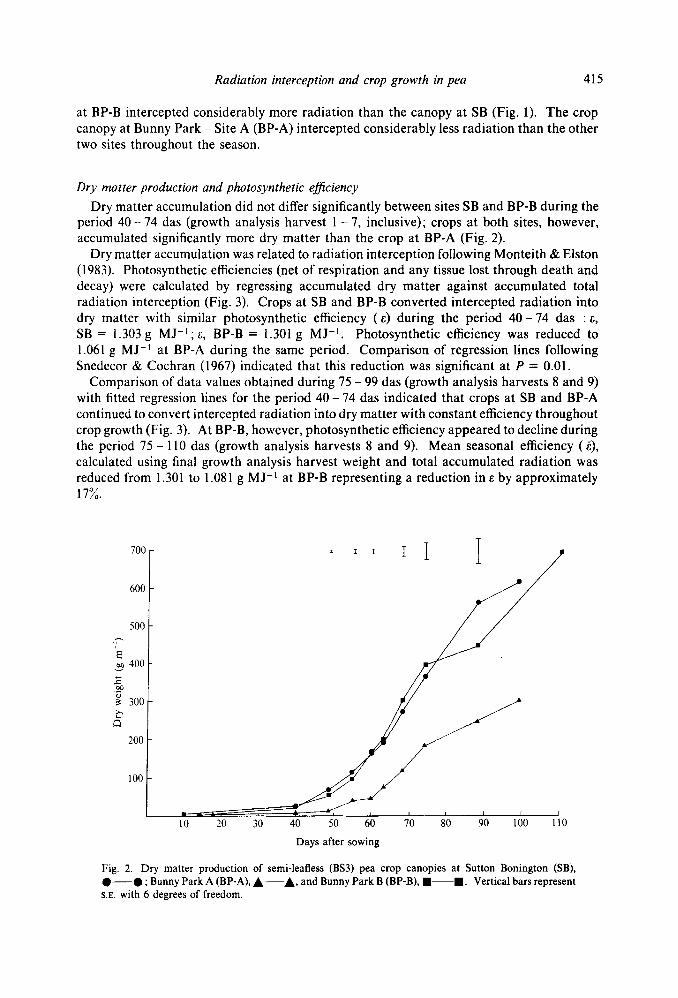
Dry matter production and photosynthetic eficiency Dry matter accumulation did not differ significantly between sites SB and BP-B during the
period 40 - 74 das (growth analysis harvest 1 - 7, inclusive); crops at both sites, however, accumulated significantly more dry matter than the crop at BP-A (Fig. 2).
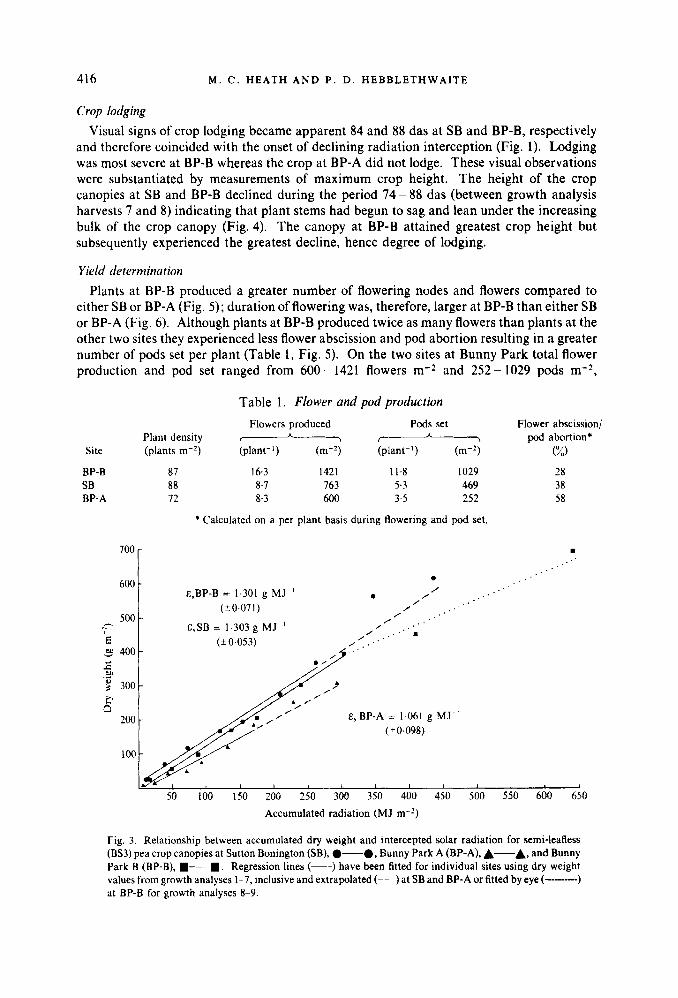
Dry matter accumulation was related to radiation interception following Monteith & Elston (1983). Photosynthetic efficiencies (net of respiration and any tissue lost through death and decay) were calculated by regressing accumulated dry matter against accumulated total radiation interception (Fig. 3). Crops at SB and BP-B converted intercepted radiation into dry matter with similar photosynthetic efficiency ( E ) during the period 40 - 74 das : E ,
SB = 1.303 g MJ-I ; E, BP-B = 1.301 g MJ-I. Photosynthetic efficiency was reduced to 1.061 g MJ-I at BP-A during the same period. Comparison of regression lines following Snedecor & Cochran (1967) indicated that this reduction was significant at P = 0.01.
Comparison of data values obtained during 75 - 99 das (growth analysis harvests 8 and 9) with fitted regression lines for the period 40 - 74 das indicated that crops at SB and BP-A continued to convert intercepted radiation into dry matter with constant efficiency throughout crop growth (Fig. 3). At BP-B, however, photosynthetic efficiency appeared to decline during the period 75 - 110 das (growth analysis harvests 8 and 9). Mean seasonal efficiency (a), calculated using final growth analysis harvest weight and total accumulated radiation was reduced from 1.301 to 1.08 1 g MJ-' at BP-B representing a reduction in E by approximately 17%.
Days after sowing
Fig. 2. Dry matter production of semi-leafless (BS3) pea crop canopies at Sutton Bonington (SB), 0-0 ; Bunny Park A (BP-A), A-A, and Bunny Park B (BP-B), m-m. Vertical bars represent S.E. with 6 degrees of freedom.
416 M . C . HEATH A N D P . D . HEBBLETHWAITE
600
500 h
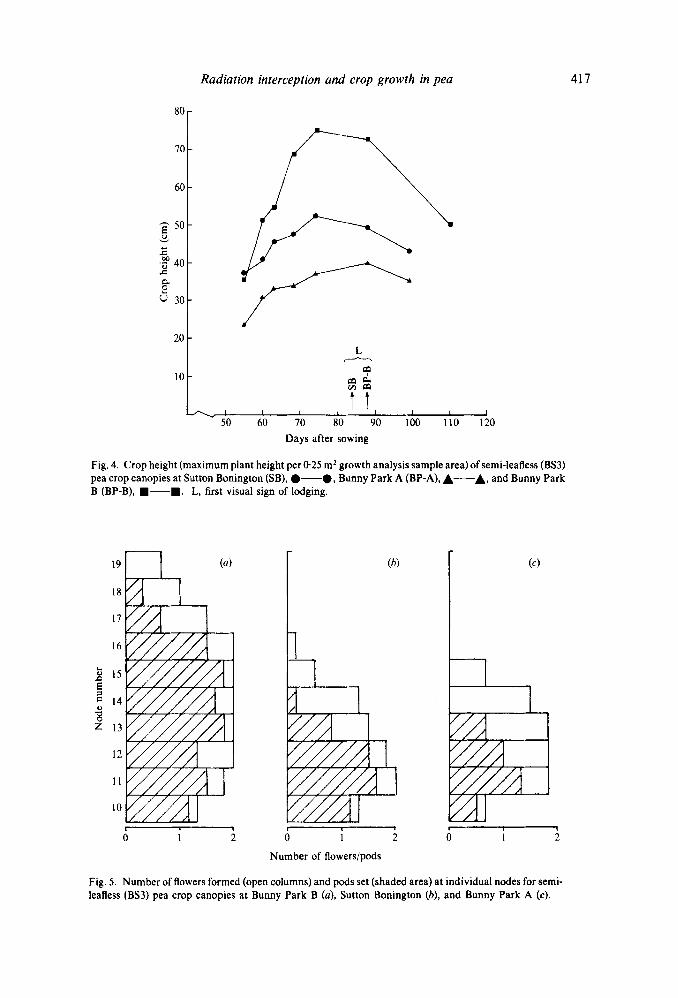
Crop lodging Visual signs of crop lodging became apparent 84 and 88 das at SB and BP-B, respectively
and therefore coincided with the onset of declining radiation interception (Fig. I ) . Lodging was most severe at BP-B whereas the crop at BP-A did not lodge. These visual observations were substantiated by measurements of maximum crop height. The height of the crop canopies at SB and BP-B declined during the period 74 - 88 das (between growth analysis harvests 7 and 8) indicating that plant stems had begun to sag and lean under the increasing bulk of the crop canopy (Fig. 4). The canopy at BP-B attained greatest crop height but subsequently experienced the greatest decline, hence degree of lodging.
Yield determination Plants at BP-B produced a greater number of flowering nodes and flowers compared to
either SB or BP-A (Fig. 5); duration of flowering was, therefore, larger at BP-B than either SB or BP-A (Fig. 6). Although plants at BP-B produced twice as many flowers than plants at the other two sites they experienced less flower abscission and pod abortion resulting in a greater number of pods set per plant (Table I , Fig. 5). On the two sites at Bunny Park total flower production and pod set ranged from 600- 1421 flowers m-2 and 252- 1029 pods m-*,
-
-
Table 1. Flower and pod production Flowers produced Pods set Flower abscission/
Plant density & & pod abortion, Site (plants m-*) (plant-') (mm2) (plant-') (m-3 (%)
87 16.3 1421 11.8 1029 28 88 8.7 763 5.3 469 38 72 8.3 600 3.5 252 58
* Calculated on a per plant basis during flowering and pod set.
b
/ &,BP-B = 1.301 g MJ-l b / /
( 5 0,071) /
E,SB = 1.303 g MJ ' ' : (* 0.053)
/ , :.' ' . '
E , BP-A = 1,061 g MJ-' (+0.098)
50 100 150 200 250 300 350 400 450 500 550 600 650 Accumulated radiation (MJ m-2)
Fig. 3. Relationship between accumulated dry weight and intercepted solar radiation for semi-leafless (BS3) pea crop canopies at Sutton Bonington (SB), .--a, Bunny Park A (BP-A), A-& and Bunny Park B (BP-B), W-W. Regression lines (-) have been fitted for individual sites using dry weight values from growth analyses 1-7, inclusive and extrapolated (---)at SB and BP-A or fitted by eye (---------) at BP-B for growth analyses 8-9.
Radiation interception and crop growth in pea
10
417
L r---.
- 2 %
I I I I 1 I
Days after sowing
Fig. 4. Crop height (maximum plant height per 0.25 m2 growth analysis sample area) of semi-leafless (BS3) pea crop canopies at Sutton Bonington (SB), 0-0, Bunny Park A (BP-A), A-A, and Bunny Park B (BP-B), W-.. L, first visual sign of lodging.
R
f, 1 i 6 1 i Number of flowers/pods
I 8 i 0 1 2
Fig. 5 . Number of flowers formed (open columns) and pods set (shaded area) at individual nodes for semi- leafless (BS3) pea crop canopies at Bunny Park B (a), Sutton Bonington (b), and Bunny Park A (c).
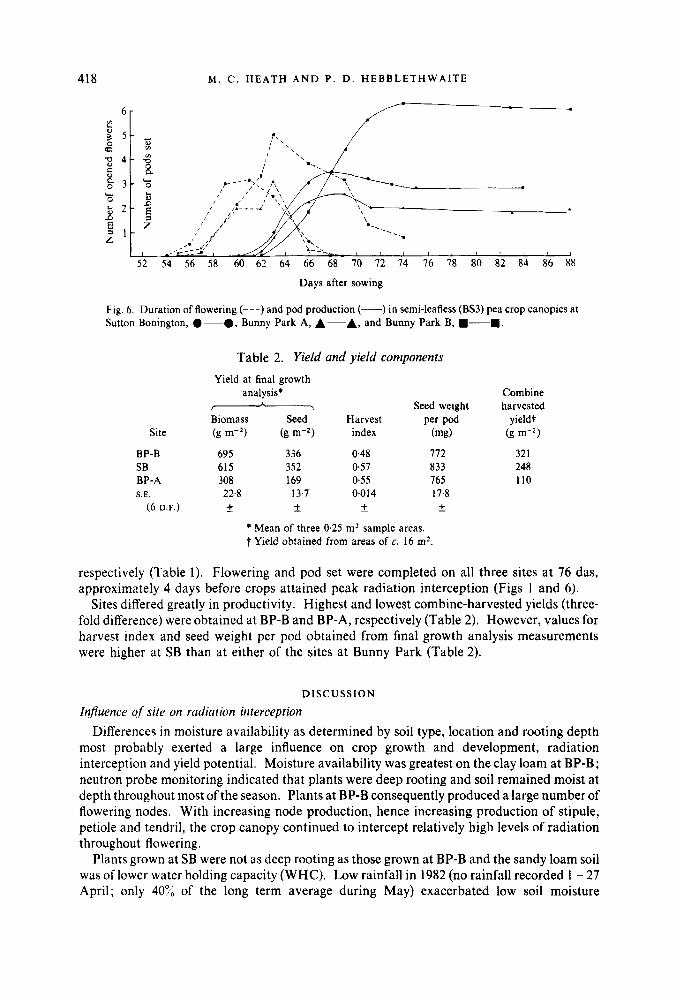
41 8 M . C . H E A T H A N D P . D . H E B B L E T H W A I T E
6 - -
.
1 1 I 1 I I f
52 54 56 58 60 62 64 66 68 70 72 74 76 78 80 82 84 86 88
Days after sowing
Fig. 6. Duration of flowering (---) and pod production (-) in semi-leafless (BS3) pea crop canopies at Sutton Bonington, 0-0, Bunny Park A, A-A, and Bunny Park B, .-..
Table 2. Yield and yield components Yield at final growth
analysis* I 1
Biomass Seed Harvest Site (g m-*) (g m-2) index
BP-B 695 336 0.48 SB 615 352 0.57 BP-A 308 169 0.55 S.E. 224 13-7 0.014
(6 D.F.) * L *
Combine Seed weight harvested
Per Pod yield? (mg) (g m-’)
772 321 833 248 765 110 17.8 ?
* Mean of three 0.25 m2 sample areas. t Yield obtained from areas of c. 16 m2.
respectively (Table 1). Flowering and pod set were completed on all three sites at 76 das, approximately 4 days before crops attained peak radiation interception (Figs 1 and 6).
Sites differed greatly in productivity. Highest and lowest combine-harvested yields (three- fold difference) were obtained at BP-B and BP-A, respectively (Table 2). However, values for harvest index and seed weight per pod obtained from final growth analysis measurements were higher at SB than at either of the sites at Bunny Park (Table 2).
DISCUSSION
Influence of site on rudiation interception Differences in moisture availability as determined by soil type, location and rooting depth
most probably exerted a large influence on crop growth and development, radiation interception and yield potential. Moisture availability was greatest on the clay loam at BP-B; neutron probe monitoring indicated that plants were deep rooting and soil remained moist at depth throughout most of the season. Plants at BP-B consequently produced a large number of flowering nodes. With increasing node production, hence increasing production of stipule, petiole and tendril, the crop canopy continued to intercept relatively high levels of radiation throughout flowering.
Plants grown at SB were not as deep rooting as those grown at BP-B and the sandy loam soil was of lower water holding capacity (WHC). Low rainfall in 1982 (no rainfall recorded 1 - 27 April; only 40% of the long term average during May) exacerbated low soil moisture

Radiation interception and crop growth in pea 419
availability. Consequently, the crop at SB would have experienced increasing moisture stress during flowering, the stage when peas are sensitive to moisture stress (Salter, 1963; Salter & Williams, 1967). Fewer flowering nodes were produced at SB compared to BP-B; conse- quently, interception during flowering did not increase as rapidly at SB compared to BP-B.
At BP-A plants were similarly shallow rooting and the soil type was typically of low WHC (Thomasson & Robson, 1967). Thus, the crop at BP-A, similar to SB, would have experienced increasing moisture stress. However, additional stress factors were experienced at BP-A. Peas were drilled into an open, cloddy, surface-compacted seedbed (typical of shallow soils developed on Keuper Marl) in an exposed situation: crop establishment and growth were impaired from the onset and interception remained at a relatively low level throughout the season.
Seasonal radiation interception, dry matter production and photosynthetic eficiency Pea (Pisum satiuum) is an indeterminate species and on two of the three sites (SB, BP-A)
where crop lodging was not a serious problem, semi-leafless pea crops converted intercepted radiation into dry matter with constant efficiency ( E ) throughout the growing season (vegetative, flowering and pod fill). In contrast, the photosynthetic efficiency of determinate species, such as cereals, declines towards the end of the growing season, due to the cessation of leaf production at flowering (Monteith & Elston, 1983). The reduction in t a t BP-A (a stressed site) in comparison to SB (a site less subject to stress) is in agreement with our previous findings (Heath & Hebblethwaite, 19856).
At the third site (BP-B) photosynthetic efficiency appeared to decline during the post- flowering period, resulting in a mean seasonal efficiency ( E ) which was only 83% of the efficiency ( E ) achieved previously. Severe lodging which occurred at BP-B most likely accounted for much of the decline in efficiency. Lodged pea plants lying on top of one another are likely to be less effective at utilising intercepted radiation for dry matter production. Furthermore, losses in dry matter from rotting during the post-flowering period are likely to be greater within a lodged compared to a standing crop. The decline in photosynthetic efficiency at BP-B may have been larger than actually measured; collapse of plants across a solarimeter located at the base of a lodged canopy will tend to overestimate crop canopy interception.
Seasonal radiation interception and yield determination Yield potential in peas is represented by the number of pods set prior to pod fill.
Environmental stress (soil moisture stress, poor soil physical conditions) appeared to influence the amount of flower abscission and pod abortion prior to pod fill. Flower and pod losses were highest on the most stressed site (BP-A) and lowest on the least stressed site (BP-B). The extent to which yield potential is realised will depend on the ability of the crop canopy to retain pods during pod fill and supply them with photosynthate to swell developing ovules (Meadley & Milbourn, 1970; 1971). In this study the yield potential of crops grown on all three sites was determined just prior to the onset of declining radiation interception. The occurrence of further pod loss during pod fill at all three sites would most likely result from declining radiation interception hence photosynthate availability.
The ability of the crop canopy to utilise intercepted radiation appeared to influence the supply of photosynthate to pods. Seed weight per pod and harvest index were highest at the site (SB) with the greatest photosynthetic efficiency during pod fill; reduced values for both components at the stressed (BP-A) site indicated a reduction in photosynthetic potential. The apparent decline in photosynthetic efficiency that occurred during pod fill at BP-B, presumably due to crop lodging, most probably influenced values of seed weight per pod and harvest index which were lower than at SB. Thus, in the absence of lodging ex-combine seed yield at BP-B may have been even higher than measured.

420 M . C . HEATH A N D P . D . HEBBLETHWAITE
CONCLUSIONS
This study illustrated the important influence of site and soil conditions (particularly soil moisture availability) on crop growth (dry matter production), development (node and flower production) and yield determination (pod production, retention and fill) in pea.
An attempt was made to relate radiation interception firstly with crop growth and development and secondly, to pod retention and pod fill.
In the absence of lodging intercepted radiation was converted into dry matter with constant photosynthetic efficiency throughout the entire growing season. This behaviour for pea (indeterminate growth habit) contrasted previously documented behaviour for cereals (determinate growth habit) where photosynthetic efficiency declined towards the end of the growing season.
Yield potential (pods set prior to pod fill) was determined just prior to the onset of declining radiation interception; pod abortion during pod fill coincided with and most likely resulted from declining photosynthate supply. Occurrence of crop lodging on one site appeared to reduce the ability of the crop canopy to supply photosynthate to pods.
ACKNOWLEDGEMENTS
Financial assistance for this work from the Agricultural and Food Research Council and co- operation from the John Innes Institute, Norwich is gratefully acknowledged. The authors also wish to thank Dr C. F. Green for valuable discussion during the course of this work; Mrs S. Manison, Mr J. Travers and Mr D. Hodson for technical assistance, and Mr J. Craigon for statistical advice received.
REFERENCES BELL, J . P. (1973). Neutron probe practice. Report 19, Institute of Hydrology, Wallingford, U.K. HEATH, M. c. & HEBBLETHWAITE, P. D. (1984). A basis for improving the dried pea crop. Outlook on
Agriculture 13, 195 - 202. HEATH, M. C. & HEBBLETHWAITE, P. D. (1985~). Agronomic problems associated with the pea crop. Ch. 2.
In The Pea Crop - a Basis for Improvement. Eds P. D. Hebblethwaite, M. C. Heath and T. C. K. Dawkins. University of Nottingham Faculty of Agricultural Science 40th Easter School in Agricultural Science. London : Butterworths, 508 pp.
HEATH, M. c. & HEBBLETHWAITE, P. D. (19856). Solar radiation interception by leafless, semi-leafless and leafed peas (Pisum sativum) under contrasting field conditions. Annals of Applied Biology 107,
LONG, I. F. & FRENCH, B. K. (1967). Measurements of soil moisture in the field by neutron moderation. Journal of’ Soil Science 18, 149 - 166.
MEADLEY, J . T. & MILBOURN, G. M. (1970). The growth of vining peas. 11. The effect of density of planting. Journal of Agricultural Science, Cambridge 74, 273 - 278.
MEADLEY, I. T. & MILBOURN, G. M. (1971). The growth of vining peas. 111. The effect of shading on abscission of flowers and pods. Journal of Agricultural Science, Cambridge 17, 103 - 108.
MONTEITH, I. L. & ELSTON, I. (1983). Performance and productivity of foliage in the field. In The Growth and Functioning of Leaoes, pp. 499 - 518. Eds J. E. Dale and F. L. Milthorpe. London: Cambridge University Press.
SALTER, P. I. (1963). The effect of wet or dry soil conditions at different growth stages on the components of yield of a pea crop. Journal of Horticultural Science 38, 321 - 334.
SALTER, P. J. & WILLIAMS, I. B. (1967). The effect of irrigation on pea crops grown at different plant densities. Journal of Horticultural Science 42, 59 - 66.
SNEDECOR, G. w. & COCHRAN, N. G. (1967). Statistical Methods, 6th Edition, pp. 432 - 436. Iowa: Iowa State University Press.
THOMASSON, A. I. & ROBSON, J . D. (1967). The moisture regimes of soils developed on Keuper Marl. Journal of Soil Science 18, 329 - 340.
309 - 318.
(Receiued 20 July 198-5)