s. - jbc.org · mammalian tumor cells as well as to human and rat pneu- mocytes (15, 25). the ......
TRANSCRIPT

THE JOURNAL OF BIOLOGICAL CHEMISTRY 0 1989 by The American Society for Biochemistry and Molecular Biology, Inc.
Vol. 264, No. 10, Issue of April 5, pp. 5660-5668, 1989 Printed in U. S. A.
(Received for publication, August 10, 1988)
Arthur N. MayenoS, A. Jane CurranS, Robert L. Roberts$, and Christopher S. FooteSV From the Departments of $Chemistry and Biochemistry and §Pediatrics, University of Californiu, Los Angeles, California 90024
Human eosinophils preferentially utilize bromide to generate a brominating agent, even at physiological halide concentrations, where chloride (140 mM) is over 1000-fold greater than bromide (20-100 PM). Under the same conditions, neutrophils use chloride to gen- erate a chlorinating agent. The total amount of active halogen trapped by 1,3,5-trimethoxybenzene from eo- sinophils increases by over 2-fold as the added bromide concentration increases from 0 to 100 PM, with ap- proximately 40 nmol of halogen trapped per million cells at the highest bromide level. At least 25-35% of the oxygen consumed by stimulated eosinophils is di- rected toward the generation of halogenating species. Since the relative halogenating behavior of eosinophil peroxidase and neutrophil myeloperoxidase in this bromide range is essentially identical to that of the cells, the specificity of eosinophils toward bromide is intrinsic to eosinophil peroxidase and not to any special cellular properties. These results suggest that human eosinophils use bromide in vivo and that a deficiency of bromide may influence their ability to produce hal- ogenating agents.
In contrast to the neutrophil, whose role in preventing bacterial infections has been established, the importance of the eosinophil in human health and disease is not well under- stood. An increase in the number of eosinophils in the periph- eral blood may occur with parasitic infestations, allergic re- actions, or various malignancies. Eosinophils have a limited ability to kill invasive parasites, which is enhanced by com- plement and specific antibody (1-3). In diseases such as interstitial pulmonary fibrosis, large numbers of eosinophils accumulate in the lung and damage pulmonary tissues, caus- ing serious consequences to the host (4-10). Eosinophils are also responsible for the pathological changes that occur in hypereosinophilia syndrome and some allergic disorders (11- 13).
Eosinophils mediate damage to the parasite or the host by the release of granule constituents (which include major basic protein and eosinophil cationic protein) and by the generation of reactive oxygen intermediates, including superoxide anion and hydrogen peroxide (14, 15). The ability of both eosino- phils and neutrophils to make superoxide from oxygen may be dependent upon the same complex of oxidase proteins,
*This research was supported by National Science Foundation Grant CHE86-07464 and Cystic Fibrosis Foundation Grant G151 7- 02. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
11 To whom correspondence should be addressed Dept. of Chem- istry and Biochemistry, University of California, 405 Hilgard Ave., Los Angeles, CA 90024-1569.
since neither eosinophils nor neutrophils from chronic gran- ulomatous disease patients are able to produce superoxide (16,17). Normal eosinophils are able to produce two to three times as much superoxide as normal neutrophils; eosinophils from eosinophilic donors produce more superoxide than eo- sinophils from normals (18-21).
The superoxide generated in the cell dismutates to hydrogen peroxide which, in the presence of peroxidases, reacts with a halide to form a highly reactive halogen (X+, halogen in the formal +1 oxidation state), shown as hypochlorous acid in Scheme 1 (22). “Active” halogens occur in such species as hypohalous acids, N-haloamines, and diatomic halogens.
The peroxidation reaction is catalyzed by the enzymes eosinophil peroxidase (EPO) in eosinophils and myeloperox- idase (MPO) in neutrophils. Both peroxidases are packaged in lysosomes but are distinct gene products with different heme and protein moieties (23,24). Halogenating agents have been produced in cell-free systems by eosinophil peroxidase in the presence of halide and H202 and are very toxic to mammalian tumor cells as well as to human and rat pneu- mocytes (15, 25).
The chlorinating agents generated by neutrophils react with amines, such as ammonia and taurine, to give relatively stable N-chloramines (22). Since these chloramines retain the oxi- dizing equivalents (although with much lower reactivity), Thomas et al. (26-29) and Weiss et al. (30,31) have presented in uitro evidence which suggests that these long-lived oxidants may have significant biological consequences. Monochlora- mine is a potent microbicidal agent as is taurine chloramine, but to a lesser extent. The cytotoxic effects of hypochlorous acid, N-chloramines, and the myeloperoxidase/H202/halide system on Escherichia coli have been studied (32-34).
Although chloride and iodide both serve essential functions in animals, the biological and physiological role of bromide is unclear, even though bromide is found in all animal tissues at levels 50-100 times that of iodide (except in the thyroid, where this ratio is reversed) (35). Some marine organisms such as certain molluscs, corals, sponges, and algae utilize bromide in the biosynthesis of brominated natural products (36, 37).
Weiss et al. (38,39) reported that, in contrast to neutrophils, eosinophils preferentially use bromide instead of chloride to generate a halogenating oxidant despite a greater than 1000-
2 H* NADPH
2oz oxidase 20-z - +
Superoxide dismulase
EPOMPO 1 H * + C I ’
HOC1 + Hz0
SCHEME 1. Formation of hypochlorous acid from oxygen.
5660

Bromination by Eosinophils 5661
/Q + "X+" -*
CH30 OCH3 H* CH3O OCH,
TMB "active halogen" TMBX
SCHEME 2. Trimethoxybenzene (TMB) assay for halogenat- ing agents.
fold excess concentration of chloride, presenting the first experimental evidence of a role of bromide in mammalian systems. These findings were based on the ability of bromide to inhibit hypochlorous acid formation as shown by inhibition of taurine chloramine formation (40) and the incorporation of 82Br into exogenous proteins. Taurine bromamine has been found t o kill schistosomula of Schistosoma mansoni (41).
Radioactive 14C-labeled 1,3,5-trimethoxybenzene (TMB)' can be used as a trap for these agents (Scheme 2) (42). This assay is quantitative and can be used to determine the amount of active halogen produced from various sources as well as the rate of reaction with the trap. Micromolar amounts of halo- genation agents can be detected with this technique.
Using the TMB assay, we have investigated the generation of halogenating agents by human eosinophils and have con- firmed that eosinophils preferentially utilize bromide (even when the chloride/bromide ratio is greater than 20,000) and that the total active halogen production is significantly de- creased in the absence of bromide.
EXPERIMENTAL PROCEDURES
Materials-[14C]Methy1 iodide (250 pCi, 3.6 mg) was from Du Pont- New England Nuclear. HEPES, bovine serum albumin (BSA; No. A- 4378, crystallized and lyophilized), glutathione (reduced), Zymosan A, and catalase (bovine liver, C-40) were from Sigma; bromine was from Mallinckrodt. Buffer components were: HEPES-buffered saline, (HBS; 10 mM HEPES, 135 mM NaC1,l mM CaCL, and 1 mM MgC12, pH 7.4), or phosphate-buffered saline (PBS; 10 mM sodium phos- phate, 135 mM NaCl, 1 mM CaClz, and 1 mM MgC12, pH 7.4). Buffers did not contain BSA unless specified. Phorbol myristate acetate (PMA) was from Sigma and stored at -15 "C dissolved in dimethyl sulfoxide (10 ng/pl). Polystyrene beads (1.0 pm diameter, monodis- persed latex, 5.69 X 10lO/ml) were from Polysciences. Beads with adsorbed PMA (called PMA-beads) were prepared immediately before use by mixing 50 ng of PMA/1 X 10" beads, pelleting the beads by centrifugation, and replacing the supernatant with buffer. Zymosan was opsonized by incubation in human serum (15 mg/ml) for 30 min at 37 "C in a rocking bath. The zymosan was then washed and resuspended at the same concentration in HEPES buffer without calcium or magnesium. Sodium hypobromite (NaOBr) was prepared as described previously (43) by dissolving bromine in aqueous NaOH (pH 12), kept at 4 "C, away from light, and used within 24 h of preparation. Concentration of sodium hypohalite (pH 12) was deter- mined spectrophotometrically using e = 345 M" cm-' at 330 nm for NaOBr and e = 350 M" cm" at 292 nm for NaOCl (44); concentra- tions of both were within 5% of those determined by iodometry. Syntheses of [methyl-"CITMB and TMB halides are described in another paper? TMB concentration was determined using c = 540 M" cm" at 265 nm.
Cells and Enzymes-Neutrophils (45) and eosinophils (46) from the blood of normal volunteers and patients with eosinophilia were isolated on discontinuous Percoll gradients as described previously. Procedures for obtaining blood samples were approved by the UCLA Human Subjects Protection Committee.
Peritoneal eosinophils were isolated as follows: peritoneal fluid was
The abbreviations used are: TMB, 1,3,5-trimethoxybenzene; BSA, bovine serum albumin; TMBX,, the corresponding halogenated TMB derivatives; HBS, HEPES-buffered saline; HEPES, N-2-hy- droxyethylpiperazine-N'-2-ethanesulfonic acid; PBS, phosphate- buffered saline; PMA, phorbol myristate acetate.
T. E. Goyne, A. N. Mayeno, G. J. Stoddard, A. Y. Craig, C. S. Foote, and R. I. Lehrer, manuscript in preparation.
obtained from pediatric patients undergoing continuous peritoneal dialysis in the Division of Nephrology in the Department of Pediatrics at UCLA. All patients required dialysis due to renal failure and were under 5 years of age. The patients selected for the study had a marked increase in the number of eosinophils (over 80% of peritoneal cells) in their peritoneal fluid, affording up to lo9 eosinophils/liter of fluid. The cause of eosinophilia is not known but is not in response to parasitic infestation or identifiable allergens.
Due to the high percentage of eosinophils in the peritoneal fluid, purification of eosinophils from peritoneal fluid differs from that of blood in that treatment with N-formylmethionylleucylphenylalanine is not required. In order to obtain highly enriched eosinophil popu- lations, the peritoneal cells were concentrated by centrifugation, washed in PBS, and then layered over discontinuous Percoll gradients similar to those prepared for the separation of blood eosinophils. Three concentrations of Percoll (55, 65, and 75%) were layered into a 15-ml polystyrene tube to form a discontinuous gradient using 3 ml of each concentration of Percoll/layer. Peritoneal cells (2-4 X 10' in 3 ml of PBS) were layered over the gradient and then spun at 400 X g for 15 min at room temperature. Erythrocytes pelleted to the bottom of the tube; the peritoneal leukocytes formed bands at the interfaces of the Percoll layers.
The band of cells between the 75 and 65% Percoll layers was >95% eosinophils, and the band between 65 and 55 layers was 50-95% eosinophils (remainder neutrophils and some lymphocytes) depend- ing on the particular patient. The band above the 55% layer was a mixture of eosinophils, neutrophils, lymphocytes, and macrophages. The different bands were then collected separately by pipette and washed in PBS prior to use in the assays. Cell samples >95% eosinophils were used in the assays, except where noted.
leukocytes were kindly provided by Dr. Robert I. Lehrer (UCLA, Myeloperoxidase and eosinophil peroxidase purified from human
Department of Medicine) and Dr. Gerald J. Gleich (Mayo Clinic and Research Foundation). The purity of the enzymes was determined from the Reinheitzahl values: A430/Am = 0.83 for myeloperoxidase A416/Am = 0.9 for eosinophil peroxidase. Acid-urea-polyacrylamide gel electrophoresis (47) and sodium dodecyl sulfate-polyacrylamide gel electrophoresis (48) revealed no detectable cross-contamination of eosinophil peroxidase by myeloperoxidase when stained with Coo- massie Blue or silver (49).
Oxygen Consumption-Oxygen consumption was determined with a Yellow Springs Instruments (YSI) model 5300 oxygen monitor with a Instech model 600b oxygen chamber (5356 micro oxygen chamber). Eosinophils or neutrophils and TMB (1 X 10" M) were stirred in the chamber at 37 "C for approximately 10 min to determine base-line consumption of oxygen. PMA-beads (4 X 10'/ml) were added to stimulate the cells.
TMB Assay-Assays for halogenation by cells were carried out in the oxygen electrode chamber or in microcentrifuge tubes. Cells (2 X 106/ml), TMB (1 X lo" M, "C = 0.5 nCi/ml), and any other compo- nents in HBS were allowed to pre-incubate at 37 "C for approximately 10 min and then stimulated with PMA-beads (4 X 1O8/rnl) or opson- ized zymosan (0.8 mg/ml). The microcentrifuge tubes were inverted every 5 min to prevent clumping of cells. After a fixed time, an aliquot (100 pl) of the reaction mixture was quenched by addition to sodium thiosulfate (2.4 pmol) and sodium azide (1 pmol). The quenched cell samples were centrifuged for 5 min on a microcentrifuge, and then the supernatant was analyzed for halogenation (see below). The enzymatic reactions were performed by equilibrating TMB (1 X 10" M), myeloperoxidase or eosinophil peroxidase (5 pg/ml), PMA-beads (1 X 109/ml), and bromide (various concentrations) a t 37 "C in glass test tubes for 5 min and then adding hydrogen peroxide (5 X M) to initiate the reaction. After 30 min, the reactions were quenched by addition to sodium thiosulfate and sodium azide. The beads were pelleted by centrifugation, and the supernatant analyzed for halogen- ation after filtration through Acro LC3S filters (Gelman). Halogen- ation by hypohalite was carried out in a similar manner: TMB and any other components were allowed to equilibrate at 37 "C for 10 min, and then either NaOCl or NaOBr was added. For slow addition conditions, the hypohalite was slowly added using a syringe pump through a finely pulled capillary tube over a period of 5 min to a well- stirred sample of TMB.
High Performance Liquid Chromatography Conditions and Deter- mination of Halogenation-High performance liquid chromatography was carried out with a C-18 reversed-phase column (4.6 X 250 mm, Econosphere C-18, Alltech) with the following conditions: Mobile phase, acetonitrile/water (43:57); flow rate, 2.0 ml/min; wavelength monitored, 216 nm. 80 pl of the quenched reaction mixture was

5662 Bromination by Eosinophils combined with 20 pl of TMB standards, which contained TMB, TMBCl, TMBBr, TMBI (monoiodinated TMB), TMBBrCl (brom- chlorinated TMB), TMBClZ (dichlorinated TMB), and the TMBBr2 (dibrominated TMB), each at 2 X lo-' M. 80 pl of this mixture was injected into the sample loop. A guard column (Upchurch Uptight precolumn) packed with Perisorb RP-18 was used. Peaks and non- peak regions were separately collected into scintillation vials contain- ing 9 ml of ScintiVerse LC liquid scintillation mixture (Fisher). The samples were counted on a Beckman LS 8100 liquid scintillation counter with the activity given in counts/min. Halogenation was calculated as follows. After subtracting background activity for each fraction, the counts/min of all the fractions were totalled. The active bromine concentration was found by dividing the sum of the activities in the TMBBr and TMBBrCl fractions and twice the TMBBrz fraction by the total corrected activity, and multiplying this value by the initial TMB concentration, as shown below. Active chlorine was determined similarly.
activity (TMBBr + TMBBrCl + 2 TMBBrz) x [TMBl [Br+] =
Neutron Activation Analysis-Neutron activation analysis of HBS for bromine was performed by Drs. John T. Wasson and Gregory W. Kallemeyn (UCLA, Department of Earth and Space Science). HBS (5 ml) was evaporated under an IR lamp, then placed in a desiccator with Pz06 for 24 h. Analysis of the remaining solid revealed a bromine content of 46 k 3 pg/g solid, corresponding to a bromide concentration of 6.2 k 0.3 p~ in the buffer. In tables and figures, the values shown for bromide concentration refer to added bromide and do not include this "background" concentration of bromide.
total activity
RESULTS
Bromination by Purified Eosinophils or Neutrophils at Var- ious Bromide Concentrations-Since the concentration of bro- mide in human fluids ranges from about 20 to 100 pM (50), halogenation by these granulocytes was studied in this range. A typical series of runs is summarized in Table I. Eosinophils or neutrophils (2 X 106/ml) were pre-incubated at 37 "C with TMB and bromide (added at the various concentrations) in
TABLE I Halogenation by eosinophils and neutrophils at various bromide
concentrations Eosinophils or neutrophils (2 X 106/ml), TMB (1 X M), and
added bromide (at the given concentrations) in HBS were preincu- bated at 37 "C in the oxygen electrode chamber or microcentrifuge tubes for 10 min, and then stimulated with PMA-beads (4 X lO*/ml). Each reaction was allowed to proceed for the given time, then quenched by addition to sodium thiosulfate and sodium azide. The quenched cell samples were centrifuged for 5 min, and the supernatant analyzed. The results are typical of the three kinds of cells studied. NA, not applicable.
twe" time Reaction [Br-] CI' Br' Total X' CI+/Br+ X+/02 Cl-/Br-
nmol X+ trapped/l@ cells min PM
A 36 0 10.5 0.3 10.8 70 5 10.6 0.4 11.0 36 20 10.9 0.5 11.4 45 50 10.7 1.5 12.2 43 100 14.9 2.6 17.5
B 48 0 7.0 2.1 9.1 90 5 6.0 4.0 10.0 50 20 6.3 6.7 13.0 54 50 4.3 16.6 20.9 51 100 4.7 33.8 38.5
C 60 0 10.2 2.9 13.1 60 5 9.4 4.9 14.3 60 20 7.3 11.4 18.7 60 50 7.3 25.5 32.8 60 100 2.8 37.5 40.3
35.0 28.8 22.7 7.1 5.8 3.4 1.5 0.9 0.3 0.1 3.5 1.9 0.6 0.3 0.1
0.28 0.29 0.33 0.32 0.37 0.20 0.15 0.23 0.30
NDb 0.57
ND ND ND ND
NA 27,000 6,800 2,700 1,400 NA
27,000 6,800 2,700 1,400 NA
27,000 6,800 2,700 1,400
A, neutrophils, >95%, blood; B, eosinophils, >95%, peritoneal; C,
Oxygen uptake not determined. eosinophils, >97%, blood.
TABLE I1 Halogenation by eosinophils stimulated with opsonized zymosan The reaction conditions are the same as in Fig. 26 except that
various bromide concentrations were used. Reaction was quenched at 60 min. Values given are typical.
Bromide" C1+ Br' Total X+ CI'/Br+
P M nmol X' trapped/l@ cells 0 12.3 2.8 15.0 4.4 5 11.8
20 4.3 16.1 2.7
9.0 10.0 19.0 50
0.9 5.2 17.9 23.1 0.3
100 2.8 21.7 24.5 0.1 Added bromide; background level excluded (see "Experimental
Procedures").
HBS for about 5 min and then stimulated with PMA-beads (see "Experimental Procedures"). Eosinophils obtained from blood gave similar results to those from the peritoneal cavity. As the concentration of added bromide was increased from 0 to 100 pM, eosinophils showed a substantial increase in the absolute amount of bromination of TMB and a decrease in chlorination. The ratio of bromination to chlorination in- creased from less than 0.3 to greater than 7. At 100 p~ bromide, eosinophils produced up to approximately 40 nmol of hypohalous acid/million cells/h, but less than half that amount was formed when no added bromide was present. The difference arises primarily from increased bromination at the high bromide level, Eosinophils stimulated with opsonized zymosan gave essentially the same bromination/chlorination ratio in this bromide range (Table 11), although the total amount of halogenating agent trapped was significantly lower (-25 nmol/million cells). Neutrophils, on the other hand, showed only a slight increase in the amount of bromination over this range of bromide concentrations, while chlorination and the total amount of halogenation remained relatively constant. At bromide concentrations greater than or equal to 1 mM, neutrophils also showed significant bromination (Table 111).
Interestingly, some bromination is detected even with no added bromide. To ascertain whether or not HBS was the source of bromide impurities, the buffer was analyzed for bromine by neutron activation. The experiment revealed that the buffer contained 6.2 f 0.3 p~ bromine, presumably as b r ~ m i d e . ~ With a cell concentration of 2 X 106/ml, the maxi- mal amount of active bromine that can be formed from this amount of bromide is 3.1 nmol/106 cells, a value which is in good agreement with those observed. Therefore, bromide con- centrations as low as 6 p~ are utilized by eosinophils.
Experiments using mixtures of neutrophils and eosinophils gave the expected intermediate results (Table IV). At a given bromide concentration, as the eosinophil/neutrophil ratio in- creases, the trapped active bromine/active chlorine ratio also increases. These results also indicate that the presence of neutrophils does not interfere with the ability to measure active bromine.
Effect of Peroxidase Inhibitors on Halogenation by Eosim- phik+"he pivotal role of eosinophil peroxidase in the gen- eration of these halogenating agents by eosinophils is shown by the effect of peroxidase inhibitors on the trapping of active halogen (Fig. 1). The eosinophil peroxidase inhibitor azide (1 mM) completely abolished halogenation, while cyanide (1 mM) gave only partial inhibition, in accordance with the finding that azide is much better inhibitor of eosinophil peroxidase than cyanide (51). Other studies have shown that cyanide
Analysis by neutron activation can determine the amount of bromine but not its oxidation state.

Bromination b3 TABLE I11
Effect of high bromide concentration on neutrophil halogenation The reaction conditions are identical to those in Table I, except
for the following: Neutrophil concentration was 2.5 X 106/ml, and the stimulus used was PMA (100 ng/ml). Reaction time was 30 min. Values are the mean k range of duplicate experiments. NA, not applicable.
Bromide" C1+ Br+ Total X' Cl+/Br+ Cl-/Br-
F M nmol X+ trappedl lp cells 0 14.1 c 0.1 0.3 f 0.1 14.4 f 0.2 56.4 NA
100 12.8 & 0.4 1.6 f 0.2 14.4 f 0.6 8.0 1,400 1,000 9.0 k 0.1 7.0 f 0.6 16.0f 0.5 1.3 140
10,000 1.4 * 0.1 19.6 f 1.2 21.0 f 1.4 0.1 14
Added bromide; background level excluded (see "Experimental Procedures").
TABLE IV Halogenation by granulocyte mixtures
The reaction conditions are similar to those in Table I. Added bromide concentration = 100 phi. A, [PMN] = 2 X 106/ml; reaction time, 43 min. B, [PMN] = 8.8 X 106/ml; reaction time, 32 min. C, [PMN] = 9.6 X 10S/ml; reaction time, 32 min. D, [PMN] = 2 X lo6/ ml: reaction time. 1 h.
Composition
Eosinophils Neutrophils Cl+ Br' 'g,!' Cl+/Br'
. "+ t rappedl lp OI nmol X 70 cells
A <5 >95 14.9 2.6 17.5 5.73 B 41 52 13.3 14.3 27.6 0.93 C 75 25 5.5 30.5 36.0 0.18 D >98 <2 4.5 30.8 35.3 0.14
Inhibitor
FIG. 1. Effect of inhibitors on the halogenation by eosino- phils. Peritoneal or blood eosinophils (2 X 106/ml), TMB (1 X M), an inhibitor, and bromide (100 p~ added) in HBS were pre- incubated at 37 "C for 10 min, then stimulated with PMA-beads (4 X 108/ml). The reaction was allowed to proceed for 1 h and then quenched with sodium thiosulfate and sodium azide. The quenched cell samples were centrifuged for 5 min, and then the supernatant was analyzed. The values are the mean t S.D. of two or more experiments.
decreases (without completely inhibiting) the cytotoxic effects of eosinophils on LSTRA cells, a line of mouse ascites lym- phoma cells (25), and is the least effective of the heme-protein inhibitors in blocking the bactericidal and cytotoxic effects of the eosinophil peroxidase/HzOz/halide system (52). Catalase (25 pg/ml), which catalyzes the breakdown of hydrogen per- oxide to oxygen and water, decreased halogenation, whereas heat-inactivated catalase (25 pg/ml) only marginally affected trapping. Iodide at 5 p~ markedly reduced TMB halide for- mation but had negligible effect at 1 p ~ . Iodide itself may serve as a substrate for eosinophil peroxidase (53) or may be
I Eosinophils 5663
40i (a) PMA-beads - 1
I / 0 1 2 3 4 5 1
40 1 (b) Opsonized Zymosan 30
Cl'trapped -t Br+trapped - Total
- J - - 0 1 0 20 30 4 0 50 60
Time (min)
FIG. 2. Time course of halogenation by eosinophils stimu- lated with PMA-beads (a) or opsonized zymosan (b) . Peritoneal eosinophils (2 X 106/ml), TMB (1 X lo-' M), and bromide (100 pM added) in HBS were pre-incubated at 37 "C for 10 min and then stimulated with PMA-beads (4 X 108/ml) or opsonized zymosan (0.8 mg/ml). Aliquots were removed at the indicated times and quenched with sodium thiosulfate and sodium azide. The quenched cell samples were centrifuged for 5 min, and then the supernatant was analyzed. Inset, expanded view of first 5 min.
oxidized by hypohalous acid to i ~ d i n e . ~ At 5 mM, reduced glutathione, which is readily oxidized by hypohalous acid and hydrogen peroxide, completely inhibited halogenation, as did human serum (48% of the reaction volume). In the absence of a stimulus, little halogenation was detected.
Time Course of Halogenation of TMB by Eosinophils-Fig. 2, a and b show the time course of halogenation of TMB by eosinophils (in 100 p~ bromide/HBS) stimulated with PMA- beads and opsonized zymosan, respectively. With PMA-beads, bromination is relatively rapid, with about half the brominat- ing potential trapped in 10 min; the generation of halogenat- ing agents is not immediate but occurs after a lag period of at least 2.5 min (Fig. 2a, inset), consistent with the observed lag in oxygen uptake. Opsonized zymosan produced much slower bromination and a diminution of active halogen trapped. A small amount of slower chlorination continued up to about 30 min with both stimuli.
Addition of Bromide after Stimulation of Eosinophils with PMA-beads-The predilection of eosinophils for bromide is clearly illustrated when bromide is added 30 min after the cells have been stimulated by PMA-beads, as revealed in Fig. 3. When bromide (100 PM) was added 30 min after stimulation to cells initially in HBS containing 0 or 5 p~ added bromide, a marked increase in bromination occurred with nearly com- plete inhibition of further chlorination. Without added bro- mide, a small amount of bromination was found (from bro- mide impurities) at 30 min which remained unchanged there- after; chlorination continued for 60 min. All halogenation activity ceased by 60 min after stimulation. These results suggest that both NADPH oxidase and eosinophil peroxidase activity were still present even after 30 min.
Halogenation Related to Oxygen Uptake by Eosinophils and
Hypohalous acid reacts rapidly with iodide to give iodine; this reaction is the basis for iodometric studies. Iodine does not react with TMB.
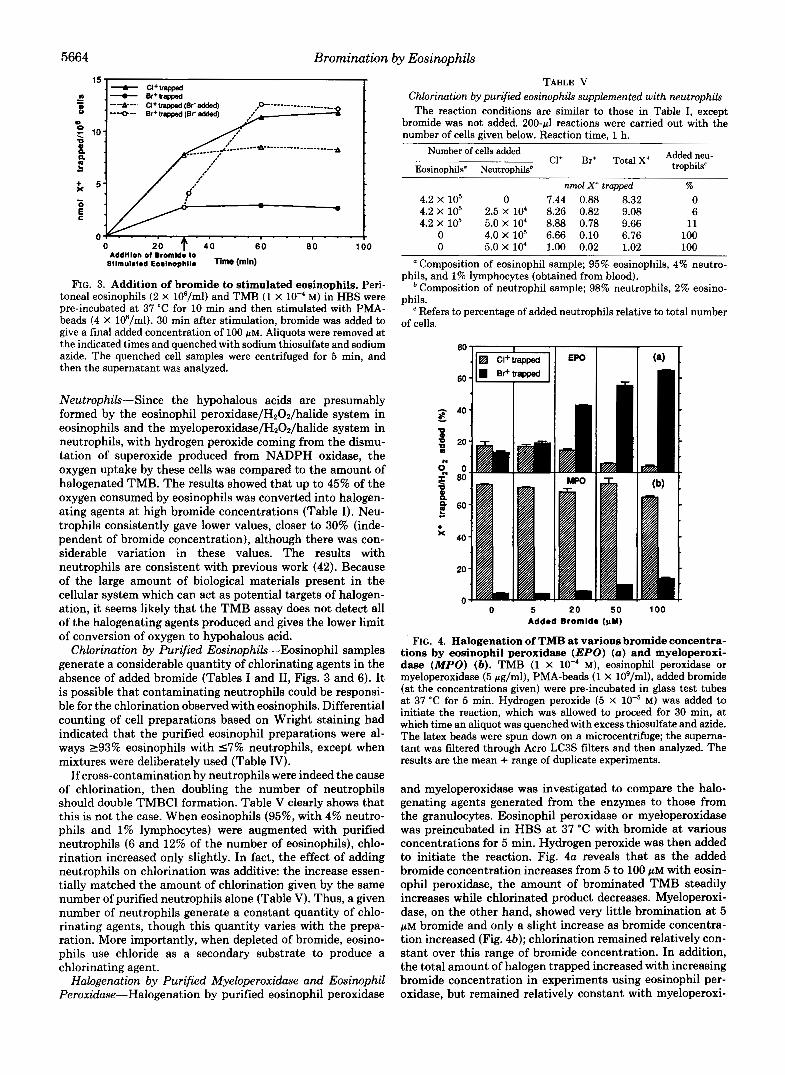
5664 Bromination by Eosinophils
0 20
Sllmulalad Eoalnophlla Time W n ) Addlllon 01 Bromld; lo
FIG. 3. Addition of bromide to stimulated eosinophils. Peri- toneal eosinophils (2 X 106/ml) and TMB (1 X M) in HBS were pre-incubated at 37 "C for 10 min and then stimulated with PMA- beads (4 X 108/ml). 30 min after stimulation, bromide was added to give a final added concentration of 100 PM. Aliquots were removed at the indicated times and quenched with sodium thiosulfate and sodium azide. The quenched cell samples were centrifuged for 5 min, and then the supernatant was analyzed.
Neutrophik-Since the hypohalous acids are presumably formed by the eosinophil peroxidase/H202/halide system in eosinophils and the myeloperoxidase/H202/halide system in neutrophils, with hydrogen peroxide coming from the dismu- tation of superoxide produced from NADPH oxidase, the oxygen uptake by these cells was compared to the amount of halogenated TMB. The results showed that up to 45% of the oxygen consumed by eosinophils was converted into halogen- ating agents at high bromide concentrations (Table I). Neu- trophils consistently gave lower values, closer to 30% (inde- pendent of bromide concentration), although there was con- siderable variation in these values. The results with neutrophils are consistent with previous work (42). Because of the large amount of biological materials present in the cellular system which can act as potential targets of halogen- ation, it seems likely that the TMB assay does not detect all of the halogenating agents produced and gives the lower limit of conversion of oxygen to hypohalous acid.
Chlorination by Purified Eosinophils-Eosinophil samples generate a considerable quantity of chlorinating agents in the absence of added bromide (Tables I and 11, Figs. 3 and 6). It is possible that contaminating neutrophils could be responsi- ble for the chlorination observed with eosinophils. Differential counting of cell preparations based on Wright staining had indicated that the purified eosinophil preparations were al- ways 293% eosinophils with 5 7 % neutrophils, except when mixtures were deliberately used (Table IV).
If cross-contamination by neutrophils were indeed the cause of chlorination, then doubling the number of neutrophils should double TMBCl formation. Table V clearly shows that this is not the case. When eosinophils (95%, with 4% neutro- phils and 1% lymphocytes) were augmented with purified neutrophils (6 and 12% of the number of eosinophils), chlo- rination increased only slightly. In fact, the effect of adding neutrophils on chlorination was additive: the increase essen- tially matched the amount of chlorination given by the same number of purified neutrophils alone (Table V). Thus, a given number of neutrophils generate a constant quantity of chlo- rinating agents, though this quantity varies with the prepa- ration. More importantly, when depleted of bromide, eosino- phils use chloride as a secondary substrate to produce a chlorinating agent.
Halogenation by Purified Myeloperoxidase and Eosinophil Peroxidase-Halogenation by purified eosinophil peroxidase
TABLE V Chlorination by purified eosinophils supplemented with neutrophils The reaction conditions are similar to those in Table I, except
bromide was not added. 200-pl reactions were carried out with the number of cells given below. Reaction time, 1 h.
Number of cells added cl+ Br+ Total x+ Addedneu- Eosinouhils" NeutroDhilsb trophils'
~ nmol X+ trapped % 4.2 X lo5 0 7.44 0.88 8.32 0 4.2 X lo5 2.5 X lo' 8.26 0.82 9.08 6 4.2 X lo5 5.0 X lo4 8.88 0.78 9.66 11
0 4.0 X lo5 6.66 0.10 6.76 100 0 5.0 X lo4 1.00 0.02 1.02 100
Composition of eosinophil sample; 95% eosinophils, 4% neutro-
Composition of neutrophil sample; 98% neutrophils, 2% eosino-
Refers to percentage of added neutrophils relative to total number
phils, and 1% lymphocytes (obtained from blood).
phils.
of cells.
0 5 2 0 5 0 100 Added Bromide (pY)
FIG. 4. Halogenation of TMB at various bromide concentra- tions by eosinophil peroxidase (EPO) (a) and myeloperoxi- dase (MPO) (6). TMB (1 X 10" M), eosinophil peroxidase or myeloperoxidase (5 pg/ml), PMA-beads (1 X 109/ml), added bromide
at 37 "C for 5 min. Hydrogen peroxide (5 X M) was added to (at the concentrations given) were pre-incubated in glass test tubes
initiate the reaction, which was allowed to proceed for 30 min, at which time an aliquot was quenched with excess thiosulfate and azide. The latex beads were spun down on a microcentrifuge; the superna- tant was filtered through Acro LCBS filters and then analyzed. The results are the mean + range of duplicate experiments.
and myeloperoxidase was investigated to compare the halo- genating agents generated from the enzymes to those from the granulocytes. Eosinophil peroxidase or myeloperoxidase was preincubated in HBS at 37 "C with bromide at various concentrations for 5 min. Hydrogen peroxide was then added to initiate the reaction. Fig. 4a reveals that as the added bromide concentration increases from 5 to 100 PM with eosin- ophil peroxidase, the amount of brominated TMB steadily increases while chlorinated product decreases. Myeloperoxi- dase, on the other hand, showed very little bromination at 5 PM bromide and only a slight increase as bromide concentra- tion increased (Fig. 46); chlorination remained relatively con- stant over this range of bromide concentration. In addition, the total amount of halogen trapped increased with increasing bromide concentration in experiments using eosinophil per- oxidase, but remained relatively constant with myeloperoxi-

Bromination by Eosinophils
TABLE VI Halogenation of TMB by NaOCl in the presence of bromide
TMB (1 X M) and bromide were allowed to equilibrate in HBS at 37 "C for 5 min. Then, NaOCl (2 X M, final concentration) was added slowly (see "Experimental Procedures"). The reaction was allowed to proceed for 15 min after initial addition and then was quenched with sodium thiosulfate and analyzed. The results are the mean + range of duplicate experiments.
Bromide" Cl' Br+ Total Cl'/Br'
P M % X' trappedlNaOC1 added 0 83.7 f 0.2 1.4 k 0.1 85.1 f 0.3 60 5 86.2 +. 0.7 0.4 f 0.1 86.6 f 0.8 215
20 85.1 +. 0.6 1.3 f 0.6 86.4 f 1.1 65 50 82.3 2.4 1.7 +. 0.4 84.0 f 2.8 48
100 81.4 f 2.0 4.7 f 0.6 86.1 f 2.6 17
Added bromide; background level excluded (see "Experimental Procedures").
TABLE VI1 Effect of buffer on halogenation by NaOBr
TMB (1 X 10" M) in one of the buffers was allowed to equilibrate at 37 "C for 5 min. Then, NaOBr (2 X M, final concentration) was added slowly, under reaction conditions identical to Table V. Results are the mean + range of duplicate experiments.
Buffer c1+ Br+ Total Cl+/Br' % X + trapped/NaOBr added
HBS 0.8 f 0.2 98.0 f 2.3 96.8 f 2.5 0.01 PBS 0.6 t 0.3 85.1 f 4.6 85.7 f 4.9 0.01 HEPES only 1.6 t 0.2 91.6 f 1.6 93.2 f 1.8 0.02 Phosphate buffer 0.8 f 0.5 97.6 f 0.9 98.4 f 1.4 0.01
dase. Calculations showed that eosinophil peroxidase had utilized all the available bromide at 5 PM, as had eosinophils.
Reaction of NaOCl with TMB in HEPES in the Presence of Bromide-A possible explanation for the formation of hypo- bromous acid is that bromide is oxidized by hypochlorous acid. This reaction has been studied in detail (54-56). To determine to what extent this reaction could be taking place in our system, NaOCl was added to TMB in HEPES-buffered saline containing various amounts of bromide. Table VI shows that even at 100 PM bromide, only a small percentage of TMB bromides are formed, suggesting that this reaction is not significant under our assay conditions. The trapping of NaOCl by TMB appears to be faster than reaction with bromide to produce hypobromous acid. With HEPES-buffered saline, TMB was able to trap about 87% of the NaOCl added. NaOCl is known to react with HEPES'; this side reaction is probably the reason that NaOCl is not trapped quantitatively, as it is in PBS.
Reaction of NaOBr with TMB in HEPES and Phosphate Buffers in the Presence and Absence of Chloride-Hypobro- mous acid is known to decompose rather rapidly (57), but hypobromite is fairly stable at pH 12 (58) when kept refrig- erated and in the dark. Decomposition is catalyzed by sunlight (59). The effectiveness of TMB as a trap for NaOBr was studied in various buffers (Table VII). In HBS and phosphate- only buffer, NaOBr was trapped essentially quantitatively, although in HBS, a significant fraction of dibrominated TMB was found. 10 mM HEPES buffer gave similar results though some oxidizing equivalents were lost. PBS gave some dibrom- inated product and showed some loss of hypobromite. These results show that TMB is an effective trap for hypobromite even in HEPES buffer.
Inhibition of Halogenation of TMB by Bovine Serum Albu- min; Effect of HEPES-Since TMB must trap halogenating
A. N. Mayeno, A. J. Curran, R. L. Roberts, and C. S. Foote, unpublished results.
5665
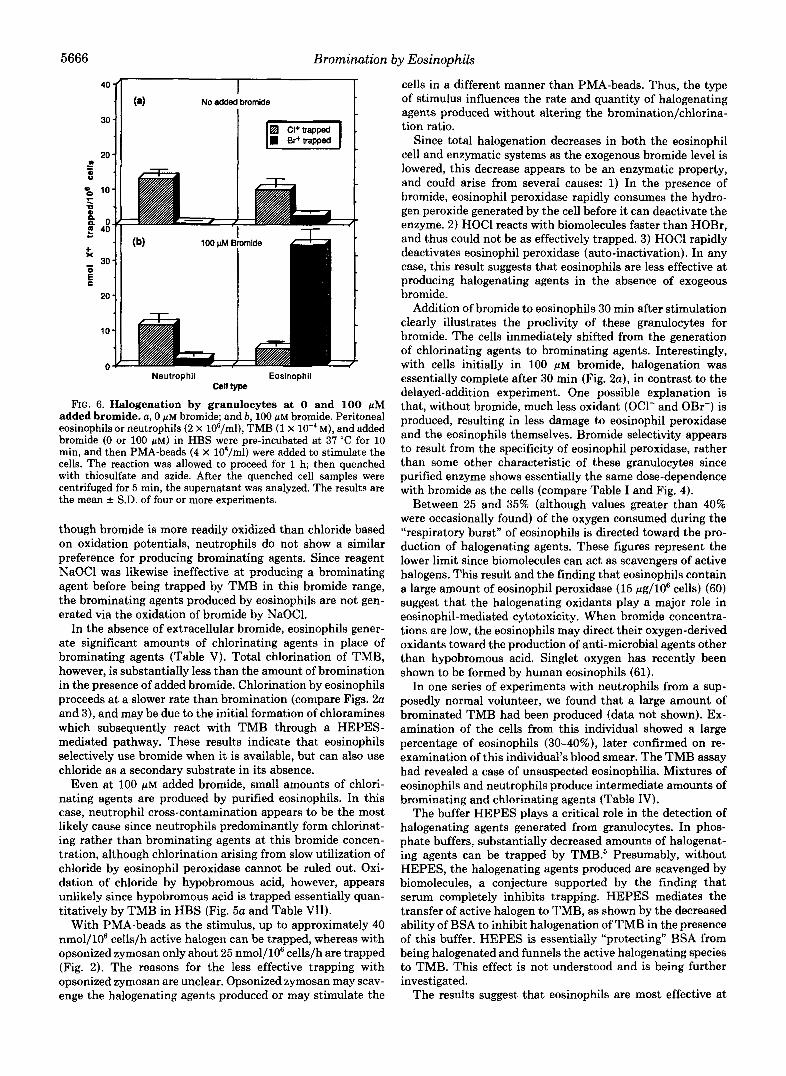
5666 Bromination by Eosinophils
40 I (a) No added bromide I
20 301 - + X
0 - 30 E
20
10
n Neutrophil Eosinophil
Cell type
FIG. 6. Halogenation by granulocytes at 0 and 100 pM added bromide. a, 0 p~ bromide; and b, 100 pM bromide. Peritoneal eosinophils or neutrophils (2 X 106/ml), TMB (1 X M), and added bromide (0 or 100 p ~ ) in HBS were pre-incubated at 37 “C for 10 min, and then PMA-beads (4 X 108/ml) were added to stimulate the cells. The reaction was allowed to proceed for 1 h; then quenched with thiosulfate and azide. After the quenched cell samples were centrifuged for 5 min, the supernatant was analyzed. The results are the mean f S.D. of four or more experiments.
though bromide is more readily oxidized than chloride based on oxidation potentials, neutrophils do not show a similar preference for producing brominating agents. Since reagent NaOCl was likewise ineffective at producing a brominating agent before being trapped by TMB in this bromide range, the brominating agents produced by eosinophils are not gen- erated via the oxidation of bromide by NaOC1.
In the absence of extracellular bromide, eosinophils gener- ate significant amounts of chlorinating agents in place of brominating agents (Table V). Total chlorination of TMB, however, is substantially less than the amount of bromination in the presence of added bromide. Chlorination by eosinophils proceeds at a slower rate than bromination (compare Figs. 2a and 3) , and may be due to the initial formation of chloramines which subsequently react with TMB through a HEPES- mediated pathway. These results indicate that eosinophils selectively use bromide when it is available, but can also use chloride as a secondary substrate in its absence.
Even at 100 p~ added bromide, small amounts of chlori- nating agents are produced by purified eosinophils. In this case, neutrophil cross-contamination appears to be the most likely cause since neutrophils predominantly form chlorinat- ing rather than brominating agents at this bromide concen- tration, although chlorination arising from slow utilization of chloride by eosinophil peroxidase cannot be ruled out. Oxi- dation of chloride by hypobromous acid, however, appears unlikely since hypobromous acid is trapped essentially quan- titatively by TMB in HBS (Fig. 5a and Table VII).
With PMA-beads as the stimulus, up to approximately 40 nmol/106 cells/h active halogen can be trapped, whereas with opsonized zymosan only about 25 nmol/106 cells/h are trapped (Fig. 2). The reasons for the less effective trapping with opsonized zymosan are unclear. Opsonized zymosan may scav- enge the halogenating agents produced or may stimulate the
cells in a different manner than PMA-beads. Thus, the type of stimulus influences the rate and quantity of halogenating agents produced without altering the bromination/chlorina- tion ratio.
Since total halogenation decreases in both the eosinophil cell and enzymatic systems as the exogenous bromide level is lowered, this decrease appears to be an enzymatic property, and could arise from several causes: 1) In the presence of bromide, eosinophil peroxidase rapidly consumes the hydro- gen peroxide generated by the cell before it can deactivate the enzyme. 2) HOCl reacts with biomolecules faster than HOBr, and thus could not be as effectively trapped. 3) HOCl rapidly deactivates eosinophil peroxidase (auto-inactivation). In any case, this result suggests that eosinophils are less effective at producing halogenating agents in the absence of exogeous bromide.
Addition of bromide to eosinophils 30 min after stimulation clearly illustrates the proclivity of these granulocytes for bromide. The cells immediately shifted from the generation of chlorinating agents to brominating agents. Interestingly, with cells initially in 100 p~ bromide, halogenation was essentially complete after 30 min (Fig. 2a), in contrast to the delayed-addition experiment. One possible explanation is that, without bromide, much less oxidant (OC1- and OBr-) is produced, resulting in less damage to eosinophil peroxidase and the eosinophils themselves. Bromide selectivity appears to result from the specificity of eosinophil peroxidase, rather than some other characteristic of these granulocytes since purified enzyme shows essentially the same dose-dependence with bromide as the cells (compare Table I and Fig. 4).
Between 25 and 35% (although values greater than 40% were occasionally found) of the oxygen consumed during the “respiratory burst” of eosinophils is directed toward the pro- duction of halogenating agents. These figures represent the lower limit since biomolecules can act as scavengers of active halogens. This result and the finding that eosinophils contain a large amount of eosinophil peroxidase (15 pg/106 cells) (60) suggest that the halogenating oxidants play a major role in eosinophil-mediated cytotoxicity. When bromide concentra- tions are low, the eosinophils may direct their oxygen-derived oxidants toward the production of anti-microbial agents other than hypobromous acid. Singlet oxygen has recently been shown to be formed by human eosinophils (61).
In one series of experiments with neutrophils from a sup- posedly normal volunteer, we found that a large amount of brominated TMB had been produced (data not shown). Ex- amination of the cells from this individual showed a large percentage of eosinophils (30-40%), later confirmed on re- examination of this individual’s blood smear. The TMB assay had revealed a case of unsuspected eosinophilia. Mixtures of eosinophils and neutrophils produce intermediate amounts of brominating and chlorinating agents (Table IV).
The buffer HEPES plays a critical role in the detection of halogenating agents generated from granulocytes. In phos- phate buffers, substantially decreased amounts of halogenat- ing agents can be trapped by TMB? Presumably, without HEPES, the halogenating agents produced are scavenged by biomolecules, a conjecture supported by the finding that serum completely inhibits trapping. HEPES mediates the transfer of active halogen to TMB, as shown by the decreased ability of BSA to inhibit halogenation of TMB in the presence of this buffer. HEPES is essentially “protecting” BSA from being halogenated and funnels the active halogenating species to TMB. This effect is not understood and is being further investigated.
The results suggest that eosinophils are most effective at

Bromination by Eosinophils 5667
producing a halogenating agent in bromide-rich environ- ments, and that their halogenating ability is hampered in bromide-poor surroundings. Under normal circumstances, this may have little effect since bromide is usually found extracellularly (62,63), normally at 20-100 pM (50). However, patients receiving long term total parenteral nutrition showed a substantial decrease in the amount of bromide in their serum (as low as 8% of levels in control serum) because of the low levels of bromide in the fluids they were receiving (64). Conversely, bromide levels in all tissues and fluids can be substantially raised by increased dietary bromine intakes (65). As eosinophils are involved in combating metazoan parasites, further investigation comparing the effectiveness of hypo- bromous acid and hypochlorous acid in killing helminths and other targets of eosinophils, as well as a study of their relative cytotoxic effects, may be worthwhile. The effects of dietary bromide administration in selected parasitic disorders may also warrant investigation.
Acknowledgments-We thank Drs. Robert I. Lehrer and Gerald J. Gleich for providing neutrophil and eosinophil peroxidases, respec- tively, Drs. John T. Wasson and Gregory W. Kallemeyn for the neutron activation studies, Drs. E. Richard Stiehm and Isidro Salusky for allowing their patients to participate in the study, Sylvia S. L. Harwig for performing the electrophoresis procedures, Drs. Robert I. Lehrer and Tammy E. Kilgore for their helpful suggestions and criticisms in preparing this manuscript, and Lisa Chen for her tech- nical assistance.
REFERENCES
1. Capron, M., Nogueira-Queiroz, J. A., Papin, J. P., and Capron, A. (1984) Cell Zmmunol. 83,60-72
2, Capron, M., Spiegelberg, H. L., Prin, L., Bennich, H., Butter- worth, A. E., Pierce, R. J., Ali Ouaissi, M., and Capron, A. (1984) J. Zmmunol. 132 , 462-468
3. Hamada, A,, and Greene, B. M. (1987) J. Zmmunol. 138, 1240- 1245
4. Davis, W. B., Fells, G. A., Sun, X.-H., Gadek, J. E., Venet, A., and Crystal, R. G. (1984) J. Clin. Invest. 74 , 269-278
5. Dor, P. J., Ackerman, S. J., and Gleich, G. J. (1984) Am. Rev. Respir. Dis. 130,1072-1077
6. De Monchy, J. G. R., Kauffman, H. F., Venge, P., Koeter, G. H., Jansen, H. M., Sluiter, H. J., and De Vries, K. (1985) Am. Reu. Respir. Dis. 131,373-376
7. Gonzalez, E. B., Swedo, J. L., Rajaraman, S., Daniels, J. C., and Grant, J. A. (1987) J. Allergy Clin. Zmmunol. 79, 755-762
8. Cluzel, M., Damon, M., Chanez, P., Bousquet, J., de Paulet, A. C., Michel, F. B., and Godard, Ph. (1987) J. Allergy Clin. Zmmunol. 80,195-201
9. Pinkston, P., Vijayan, V. K., Nutman, T. B., Rom, W. N., O’Donnell, K. M., Cornelius, M. J., Kumaraswami, V., Ferrans, V. J., Takemura, T., Yenokida, G., Thiruvengadam, K. V., Tripathy, S. P., Ottesen, E. A,, and Crystal, R. G. (1987) J. Clin. Invest. 80, 216-225
10. Hallgren, R., Samuelsson, T., Venge, P., and Modig, J. (1987) Am. Rev. Respir. Dis. 136,639-642
11. Fauci, A. S., Harley, J. B., Roberts, W. C., Ferrans, V. J., Gralnick, H. R., and Bjornson, B. H. (1982) Ann. Intern. Med. 97, 7% 92
12. Parrillo, J. E., Borer, J. S., Henry, W. L., Wolff, S. M., and Fauci, A. S. (1979) Am. J. Med. 67,572-582
13. Shult, P. A., Graziano, F. M., and Busse, W. W. (1986) J. Allergy Clin. Zmmunol. 77, 702-708
14. Ackerman, S. J., Loegering, D. A., Venge, P., Olsson, I., Harley, J. B., Fauci, A. S., and Gleich, G. J. (1983) J. Zmmunol. 131 ,
15. Agosti, J. M., Altman, L. C., Ayars, G. H., Loegering, D. A., Gleich, G. J., and Klebanoff, S. J. (1987) J. Allergy Clin. Zmmunol. 79,496-504
16. Segal, A. W., Garcia, R., Goldstone, A. H., Cross, A. R., and Jones, 0. T. G. (1981) Biochem. J. 196,363-367
17. Roberts, R. L., Ohno, Y., and Gallin, J . I. (1986) Pediatr. Res.
18. Shult, P. A., Graziano, F. M., Wallow, I. H., and Busse, W. W.
2977-2982
20,378-380
(1985) J. Lab. Clin. Med. 106,638-645 19. Learn, D. B., and Brestel, E. P. (1982) Agents Actions 12 , 485-
488 20. Pretreccia, D. C., Nauseef, W. M., and Clark, R. A. (1987) J.
Leukocyte Biol. 41 , 283-288 21. Bass, D. A., Grover, W. H., Lewis, J. C., Szejda, P., DeChatelet,
L. R., and McCall, C. E. (1980) J. Clin. Znuest. 6 6 , 1265-1273 22. Test, S. T., and Weiss, S. J. (1986) Adu. Free Radical Bwl. Med.
23. Olsson, I., Persson, A. M., Stromberg, K., Winqvist, I., Tai, P. C., and Spry, C. J. F. (1985) Blood 66,1143-1148
24. Lepelley, P., Zandecki, M., Paquet, S., Lerche, B., Estienne, M. H., Fenaux, P., Torpier, G., and Cosson, A. (1987) Eur. J.
2, 91-116
HAmatol. 39,77-81- 25. Jong, E. C., and Klebanoff, S. J. (1980) J. Zmmunol. 124,1949-
1953
Clin. Invest. 72,441-454
Bwl. Chem. 269,6757-6765
E. L. (1984) J. Biol. Chem. 269,10404-10413
M. (1985) J. Biol. Chem. 260,3321-3329
Weiss, S. J. (1984) J. Clin. Znuest. 74,1341-1349
26. Thomas, E. L., Grisham, M. B., and Jefferson, M. M. (1983) J.
27. Grisham, M. B., Jefferson, M. M., and Thomas, E. L. (1984) J.
28. Grisham, M. B., Jefferson, M. M., Melton, D. F., and Thomas,
29. Thomas, E. L., Grisham, M. B., Melton, D. F., and Jefferson, M.
30. Test, S. T., Lampert, M. B., Ossanna, P. J., Thoene, J. G., and
31. Weiss, S. J., Lampert, M. B., and Test, S. T. (1983) Science 2 2 2 ,
32. Albrich, J. M., Gilbaugh, J. H., 111, Callahan, K. B., and Hurst,
33. Thomas, E. L. (1979) Infect. Zmmunol. 23 , 522-531 34. Thomas, E. L., and Aune, T. M. (1978) Antimicrob. Agents
Chemother. 13,1006-1010 35. Nielsen, F. H. (1986) in Trace Elements in Human and Animal
Nutrition (Mertz, W., ed) Vol. 2,5th Ed., pp. 426-430, Academic Press, New York
36. Downs, A. J., and Adams, C. J. (1973) in Comprehensive Inorganic Chemistry (Bailar, J. C., Emeleus, H. J., Nyholm, R., and Trotman-Dickenson, A. F., eds) pp. 1107-1594, Pergamon Press, Oxford
37. Farkas-Himsley, H., and Jolles, G. H. (1966) in Bromine and Its Compounds (Jolles, Z. E., ed) pp. 487-555, Academic Press, London
38. Roos. D., Eckmann, C. M., and Weiss, S. J. (1985) Eur. J. Clin.
625-628
J. K. (1986) J. Clin. Znuest. 78,177-184
Znuest.. 15, A51
S. (1986) Science 234, 200-203 39. Weiss, S. J., Test, S. T., Eckmann, C. M., Roos, D., and Regiani,
40. Weiss, S. J., Klein, R., Slivaka, A., and Wei, M. (1982) J. Clin.
41. Yazdanbakhsh, M., Eckmann, C. M., and Roos, D. (1987) Am. J.
42. Foote, C. S., Goyne, T. E., and Lehrer, R. I. (1983) Nature 301,
43. Engel, P., Oplatka, A,, and Perlmutter-Hayman, B. (1954) J. Am. Chem. SOC. 76,2010-2015
44. Soulard, M., Bloc, F., and Hatterer, A. (1981) J. C. S. Dalton Trans. 12,2300-2310
45. Harbeck, R. J., Hoffman, A. A., Redecker, S., Biundo, T., and Kurnick, J . (1982) Clin. Zmmunol. Zmmunopathol. 23,682-690
46. Roberts, R. L., and Gallin, J. I. (1985) Blood 66,433-440 47. Selsted, M. E., Szklarek, D., and Lehrer, R. I. (1984) Infect.
48. Fling, S. P., and Gregerson, D. S. (1986) Anal. Biochem. 1 6 6 ,
49. Wray, W., Boulikas, T., Wray, V. P., and Hancock, R. (1981) Anal. Biochem. 118 , 197-203
50. Holzbecher, J., and Ryan, D. E. (1980) Clin. Biochem. 13, 277- 278
51. Migler, R., and DeChatelet, L. R. (1978) Biochem. Med. 19 , 16- 26
52. Klebanoff, S. J., Jong, E. C., and Henderson, W. R., Jr. (1980) in The Eosinophil in Health and Disease (Mahmoud, A. A. F., Austen, K. F., and Simon, A. S., eds) pp. 99-111, Grune & Stratton, New York
53. Milger, R., DeChatelet, L. R., and Bass, D. A. (1978) Blood 6 1 ,
54. Jolley, R. L., and Carpenter, J. H. (1983) in Water Chlorination: Environmental Impact and Health Effects (Jolley, R. L., Brungs,
Invest. 70,598-607
Trop. Med. Hyg. 3 7 , 106-110
715-716
Zmmunol. 46,150-154
83-88
445-456

5668 Bromination by Eosinophils
W. A., Cotruvo, J. A., Cumming, R. B., Mattice, J. S., and 60. Carlson, M. G. Ch., Peterson, C. G. B., and Venge, P. (1985) J. Jacobs, V. A., eds) Vol. 4, Book One, pp. 3-47, Ann Arbor Immuml. 134, 1875-1879 Sciences, Michigan 61. Kanofsky, J. R., Hoogland, H., Wever, R., and Weiss, S. J. (1988)
55. Farkas, L., Lewin, M., and Bloch, R. (1949) J. Am. Chem. SOC. J. Biol. Chem. 263,9692-9696 62. Cole, B. T., and Patrick, H. (1958) Arch. Biochem. Biophys. 7 4 ,
56. Kumar, K., and Margerum, D. W. (1987) Inorg. Chem. 26,2706- 357-361 2711 63. Hellerstein, S., Kaiser, C., Darrow, D. D., and Darrow, D. C.
57. Lewin, M., and Avrahimi, M. (1955) J. Am. Chern. SOC. 77,4491- 4498
(1960) J. Clin. Inuest. 39,282-287 64. Dahlstrom, K. M., Ament, M. E., Medhin, M. G., and Meurling,
58. Chapin, R. M. (1934) J. Am. Chem. SOC. 56,2211-2215 S. (1986) J. Pediatr. 109,625-630 59. Macalady, D. L., Carpenter, J. H., andMoore, C. A. (1977) Science 65. Winnek, P. S., and Smith, A. H. (1937) J. Biol. Chem. 121,345-
71,1988-1991
195,1335-1337 352