role of photochemical reactions in the biogeochemical cycling of
TRANSCRIPT

Role of photochemical reactionsin the biogeochemical cycling
of detrital carbon in aquatic environments
Anssi Vähätalo
Department of Applied Chemistry and MicrobiologyDivision of Microbiology
University of HelsinkiFinland
Academic Dissertation in Microbiology
To be presented, with the permission of the Faculty of Agriculture and Forestryof the University of Helsinki, for public criticism in the Auditorium 2 at the
Viikki Infocentre, Viikinkaari 11 on 10 March, 2000, at 12 o'clock noon
Cover figure: Atomic force microscopic image of <0.1 µm filtered water of Lake Valkea-Kotinen sampled on 6 August 1996 displaying detrital and/or viral particles. Fifty µl of thefiltered water was allowed to evaporate to dryness on a crystal of mica (diameter of 1 cm).The image was taken with a multi mode atomic force microscope by the courtesy of dr. S. J.Everstein (Digital Instruments, Nano Scope IIIa, Scanning Probe Microscope Controller,imaging mode - tapping mode, with a silicon tip from Nanosensor).

2
Supervisors:Prof. Kalevi Salonen
Department of Biological and Environmental ScienceSection of Hydrobiology and Limnology
University of Jyväskylä
Prof. Mirja Salkinoja-SalonenDepartment of Applied Chemistry and Microbiology
Division of MicrobiologyUniversity of Helsinki
Reviewers:Prof. Jussi Kukkonen
Department of BiologyUniversity of Joensuu
Dr. Alasdair H. NeilsonSwedish Environmental Research Institute
Opponent:Prof. Dr. Hauke Harms
Department of Rural EngineeringSwiss Federal Institute of Technology
ISBN 951-45-9141-0 (PDF version)Helsingin yliopiston verkkojulkaisut, Helsinki 2000
This work was supported by grants from the Academy of Finland, Maj and Tor NesslingFoundation, University of Helsinki and Commission of European Communities (STEP-CT90-0112); by the Finnish Graduate Schools for Environmental Ecology, Ecotoxicology andEcotechnology and that for Environmental Science and Technology; and by CIMO.

3
To my teachers

4
CONTENTS
1. List of original papers and the author's contribution 12. Abbreviations and definitions 23. Abstract 34. Introduction 5
4.1. Detritus 54.2. The role of detritus in food webs 54.3. Biodegradation of detritus 64.4. Abiotic decomposition of detritus 84.5. Physical and chemical factors affecting the decomposition of detritus 94.6. Solar radiation and radiative transfer in water 104.7. Absorption of radiation 114.8. Photochemical reactions 124.9. Predicting the rates of photochemical reactions 134.10. Photochemical reactions related to the processing of detritus 144.11. Effects of solar radiation on detrivores 14
5. Aims of the study 186. Material & Methods 197. Results 20
7.1. Mineralization of DOC in humic lakes 207.2. Quantum yield for photochemical mineralization 217.3. Photochemical modification of DOC and it's effects on bacteria 227.4. Photochemical modification of detrital eelgrass 23
8. Discussion 268.1. The relative role of bacterioplankton and photochemical mineralization 268.2. Photomineralization and primary production 268.3. Photodegradation of vascular plant detritus 268.4. Photodegradation of DOM and bioavailability of photoproducts 278.5. Photochemical release of nitrogen and phosphorus 288.6. Photodegradation of structural lignin: implications for bacteria 288.7. Net effect of solar radiation on the decomposition of detritus 29
9. Conclusions 3010. Acknowledgements 3211. References 3412. Original papers 45

List of original papers
1
1. List of original papers
I. Salonen, K. and A. Vähätalo. 1994. Photochemical mineralisation of dissolved organicmatter in lake Skervatjern. Environment International 20: 307-312.
II. Vähätalo, A. V., M. Salkinoja-Salonen, P. Taalas, and K. Salonen. 2000. Spectrum of thequantum yield for photochemical mineralization of dissolved organic carbon in a humic lake.Limnology and Oceanography (in the press).
III. Vähätalo, A. V., K. Salonen, M. Salkinoja-Salonen and A. Hatakka. 2000. Photochemicalmineralization of synthetic lignin in lake water indicates rapid turnover of aromatic organicmatter under solar radiation. Biodegradation (in the press).
IV. Vähätalo, A. V., K. Salonen. 1996. Enhanced bacterial metabolism after exposure ofhumic lake water to solar radiation. Nordic Humus Newsletter 3: 36-43.
V. Vähätalo, A., M. Søndergaard, L. Schlüter, and S. Markager. 1998. The impact of solarradiation on the decomposition of detrital leaves of eelgrass (Zostera marina). MarineEcology Progress Series 170: 107-117.
The author's contribution
I. Anssi Vähätalo planned the experiments, conducted the measurements, analysed andinterpreted the results under supervision of Kalevi Salonen.
II. Anssi Vähätalo wrote the paper, developed the theoretical models, planned theexperiments, conducted all measurements, analysed and interpreted the results. Kalevi Salonenand Mirja Salkinoja-Salonen supervised the work. Petteri Taalas provided the ozone data.
III. Anssi Vähätalo wrote the paper, planned the experiments, conducted all measurements,analysed and interpreted the results. Kalevi Salonen and Mirja Salkinoja-Salonen supervisedthe work. Annele Hatakka provided synthetic
14C-lignin and contributed to the supervision ofthe use of 14C-labelled lignin.
IV. Anssi Vähätalo wrote the paper, planned the experiments, conducted all measurements,analysed and interpreted the results. Kalevi Salonen supervised the work.
V. Anssi Vähätalo wrote the paper, planned the experiments, conducted all measurementsexcluding pigment analyses, analysed and interpreted the results. Morten Søndergaardsupervised the work. Louise Schlüter analysed the pigments and Stiig Markager providedexpertise in the use of LI-1800 spectroradiometer.

Abbreviations
2
2. Abbreviations and definitions
CDOM Dissolved organic matter absorbing solar radiation at 290-700 nm.
Biodegradation Degradation mediated by organisms.
Degradation Reduction in the complexity of detritus.
Detritus Dead organic carbon, distinguishable from living organic and frominorganic carbon, lost by non-predatory means from any trophic level(egestion, excretion, secretion, etc.) or from sources external to theecosystem that enter and cycle in the system (allochthonous organiccarbon; Wetzel 1984).
Detrivore An organism utilizing detritus.
DIC Dissolved inorganic carbon (CO2, HCO3-, CO3
2-).
DOC Dissolved organic carbon.
DOM Dissolved organic matter.
Extracellular Enzymes located outside the cell membrane.enzyme
High molecular Detritus that must be cleaved outside the cell wall to low molecularweight detritus weight compounds prior penetration through the cell wall and
membrane. For polar compounds the high molecular weight means amolecular mass of ca. >500 g mol-1.
Humic substances A general category of naturally occurring, biogenous organicsubstances that can generally be characterized as being yellow to blackin colour, of high molecular weight, and refractory (Aiken et al. 1985).
Kd Vertical attenuation coefficient of the downward irradiance.
Low molecular Molar mass <500 g mol-1.weight detritus
Mineralization Degradation leading to inorganic constituents of detritus.
Transformation Modification of detritus (includes both increase and decrease incomplexity).
UV-B 280-315 nm.
UV-A 315-400 nm.
Visible radiation 400-700 nm.

Abstract
3
3. Abstract
In the biosphere, detritus is the largest poolof organic matter (2.5 × 1018 g C).Decomposition of detrital carbon covers 50to 100% of the carbon flux in aquatic foodwebs. In aquatic ecosystems mainlyphotochemical and biological reactionsdecompose detritus. The decomposition ofdetritus has two major implications incarbon cycling. (1) It supports the detritalfood web and (2) mineralizes organiccarbon.
In this study I quantified photochemicaldegradation of detritus in aquaticecosystems and evaluated the interactionsbetween photochemical and microbialprocesses enhancing the decomposition ofdetrital carbon. The decomposition ofdissolved organic carbon (DOC) wasstudied in two humic lakes: Lake Valkea-Kotinen in Finland and Lake Skervatjern inNorway. Decomposition of detrital leavesof eelgrass (Zostera marina, L) wasstudied with material collected fromRoskilde Fjord, Denmark. Decompositionof detritus was measured after exposingnatural or synthetic detritus to solarradiation and/or to the indigenousmicroorganisms.
I found that in Lake Valkea-Kotinen,planktonic microorganisms were the mostimportant mineralizers of DOC. In June toAugust, microbes mineralized 45 mg C m-3
d-1 and covered 88% of the total
mineralization of DOC in the 2-m deepepilimnion of Lake Valkea-Kotinen. At thedepth of 1 cm, solar radiation mineralizedDOC with a rate of 230 mg C m-3
d-1,which was four times higher than that ofmicrobial mineralization of DOC.Photochemical mineralization attenuatedsteeply with an increasing depth of thewater column in Lake Valkea-Kotinen aswell as in Lake Skervatjern. In LakeValkea-Kotinen, the vertical attenuation ofphotochemical mineralization was similar
to that of UV-A radiation (315-400 nm).Ninety % of the total photochemicalmineralization took place in the top 10 cmof the water column. The sensitivity ofDOC to photochemical mineralization washigher in water flowing to LakeSkervatjern than in epilimnetic water.
I used 14C-[ring]-labelled synthetic lignin
(14C-DHP) as a model compound forbiologically recalcitrant DOC. The
14C-DHP was blended in the Lake Valkea-Kotinen water, submerged to the depth of1 cm and exposed to solar radiation for 7d. As the result, solar radiation mineralized19% of 14C-labelled aromatic rings toinorganic carbon and increased the watersolubility of remaining part of the
14C-label.In darkness the mineralization of 14C-DHPby indigenous microbes was negligible.
The quantum yield for a photochemicalmineralization of DOC describes theproportion of absorbed photons resulting inmineralization of DOC. Quantum yield is akey parameter for predicting photo-chemical rates. I developed a novel methodfor determining the quantum yieldspectrum for the photochemical minerali-
zation of DOC (φλ) in Lake Valkea-
Kotinen in situ. The method describes φλas c × 10-dλ, where c (dimensionless) and d(nm-1) are positive constants. The
determination gave a value of φλ of 7.52 ×10-0.0122λ based on (1) the absorptionspectrum of chromophoric dissolvedorganic matter, (2) the photochemicalmineralization measured in situ at depth zand (3) the solar spectrum of photon fluxdensity at the same depth. The in situdetermined quantum yields agreed withthose determined in the laboratory at single
wavelengths. Applying a φλ of 7.52 × 10-
0.0122λ in the model resulted in asatisfactory prediction of photochemicalmineralization rates at the depths of 1-10cm as well as in the whole water column ofLake Valkea-Kotinen. The modelling

Abstract
4
revealed that the UV-A component ofsolar radiation was responsible for most ofthe mineralization of DOC in Lake Valkea-Kotinen.
The interactions between photochemicaland microbial reactions in the decom-position of DOC were studied with thewater of Lake Valkea-Kotinen. First the<0.2 µm filtered water was exposed tosolar radiation for 1 d or kept in darkness.Then the exposed and non-exposed waterwere spiked with indigenous bacteria andwere cultured for 4 d. The microbialmineralization of DOC was 2 to 5 foldhigher in the water exposed to solarradiation than in the non-exposed water.More dissolved phosphorus and nitrogenwere converted into particulate form in thewater exposed to solar radiation than in thenon-exposed water. The nutrient dynamicsindicated an increased production ofmicrobial biomass in the solar radiationexposed water. This result was supportedby microscopic data. The biovolume ofbacteria grown in the solar radiationexposed water was 1.7 times higher than ofthose grown in the non-exposed water keptin darkness.
To study photochemical decomposition ofdetrital eelgrass, I heat sterilized detritalleaves and exposed those to solar radiation.The leaves lost pigments and chromophoricmatter during 30-d exposure to solarradiation. Solar radiation enhanced the lossof organic matter from the leaves andinduced their fragmentation. After anexposure of the detrital leaves to solarradiation, the incorporation of 14C-leucineby a microbial biofilm introduced on theleaves was 2-4 times higher than thatwithout solar radiation.
This study showed that solar radiationdecomposed DOC and detrital leaves ofeelgrass both directly by photochemicalmineralization and indirectly by converting
recalcitrant organic matter more availableto microorganisms. The results suggest thatsolar radiation should be taken into accountin the decomposition of detritus.

Introduction
5
4. Introduction
4.1. Detritus
The biogeochemistry of carbon can beconceptually simplified as the cycling ofcarbon between inorganic and organicpools. The largest pool of organic matter inthe biosphere is detritus (2.5 × 1018 g C,Hedges 1992). A typical definition ofdetritus includes all the particulate organicmatter which once belonged to livingorganisms (Odum 1971). This matterincludes visually macroscopic objects likefallen tree trunks, detached leaves oranimal corpses as well as microscopicdetrital particles. In this work detritusincludes also dissolved organic matter inthe sense of Wetzel (1984): Detritus is alldead organic carbon, distinguishable fromliving organic and from inorganic carbon,lost by non-predatory means from anytrophic level (egestion, excretion,secretion, etc.) or from sources external tothe ecosystem that enter and cycle in thesystem (allochthonous organic carbon).
All biomolecules are potential sources ofdetritus. The most abundant biomoleculesin living organisms are cellulose and lignin(Hedges 1992) found in large amounts alsoamong particulate detritus in littoralecosystems. In the water column most ofthe detrital carbon is in the dissolved form.Dissolved to particulate organic carbonratio is typically 10:1 in lakes and oceans(Wetzel 1984). Only part (generally <50%)of dissolved detritus consists of clearlydefined biomolecules found in livingorganisms (carbohydrates, peptides, lipids,nucleic acids, lignin, and their monomers;Münster 1993). Typically, half of lakewater DOC is composed of humicsubstances, mainly the soluble fraction i.e.,fulvic acids (McKnight & Aiken 1998,Peuravuori & Pihlaja 1999). Humicsubstances are defined as a generalcategory of naturally occurring, biogenousorganic substances that can generally be
characterized as being yellow to black incolour, of high molecular weight, andrefractory (Aiken et al. 1985). Thedescription does not define the chemicalstructure of humic substances, except thathumic substances contain chromophoricmoieties. Most chemical knowledge ofhumic substances has been derived frommaterial isolated with non-ionicmacroporous resins at low pH. Theseresins bind large (>500 g mol-1) moleculeswith acidic functional groups. Thecompounds isolated in this way from freshwaters have molecular weights from 500 to1200 g mol-1, contain acidic functionalgroups (mainly carboxylic acids), consistmainly of carbon (54%), oxygen (40%) andhydrogen (4%), and are heterogeneous(Saski et al. 1996, McKnight & Aiken1998, Peuravuori & Pihlaja 1999). Thehumic substances may be formed viadegradative pathways from the remainingorganic material not readily biodegraded bydetrivores (Peuravuori & Pihlaja 1999). Inaddition, condensation of simplemonomeric compounds may producehumic substances (Larson & Hufnal 1980,Backlund 1992, Kieber et al. 1997).
4.2. The role of detritus in food webs
Food webs can be conceptually dividedinto grazer and detritus food webs on thebasis of the energy and carbon source(Wetzel et al. 1972). Living primaryproducers form the basis of a grazer foodweb. Organisms using detritus-based foodbelong to the detritus food web.Heterotrophic micro-organisms, bacteriaand fungi, are the most important detritusconsuming organisms detrivores (Peduzzi& Herndel 1991, Newell 1996). Theconceptual division of detritus in the foodwebs is useful in evaluating the role ofdetritus in carbon cycling. Due to omnivoryof organisms, the division of grazer anddetritus food webs and the determination ofthe trophic levels of organisms is often

Introduction
6
difficult (Polis & Strong 1996, Hessen1998).
Detrital carbon initially produced within theecosystem is autochthonous and includesthe dead components of food web at a siteand leakages from internal carbon transferprocesses (e.g., exudation, Chen &Wangersky 1996; egestion, Viitasalo et al.1999; sloppy feeding, Strom et al. 1997;viral lysis, Middelboe et al. 1996). Thedetritus food web in the water column isoften called the microbial loop when it isbased on autochthonous detritus (Mann1988). Detritus produced in one ecosystemand imported to another one isallochthonous. When detrivores consumeallochthonous material they form a linkbetween an aquatic environment and anallochthonous source. To emphasize thisand the important role of microbes in thisprocess a definition of the microbial linkwas described by Salonen et al. (1992).
The surface waters of oceans are the mostautochthonous aquatic environments. Evenin those environments >50% of carbonfixed by primary producers may flux viadetritus food web (a microbial loop; Mann1988, Fasham et al. 1999). The proportionof allochthonous and also the overallimportance of detritus increases along thegradient, open ocean - coastal waters -lakes - rivers. Due to allochthonousdetritus most aquatic systems areheterotrophic (Salonen et al. 1983, 1992,Del Giorgio et al. 1997).
4.3. Biodegradation of detritus
Detrital carbon covers 50 to 100% of thecarbon flux in aquatic food webs (Mann1988). The logistics of the production anddecomposition of detritus is important forunderstanding the functioning of aquaticecosystems. The decomposition of detritushas two major implications in carboncycling. (1) It supports the detrital food
web and (2) mineralizes organic carbon toavailable form for primary producers.
Detrivores are important degraders andmineralizers of detritus. Detrivores use apart of the detrital carbon for biosynthesisand utilize the chemical energy in thedetritus for their energetic needs.Detrivores utilize the energy released fromcleavage of covalent bonds only inside thecells. Thus detrital molecules mustpenetrate the cell membrane actively (polarmainly small molecules, 300-700 g mol-1;Page et al. 1989, Liu et al. 1993) orpassively (hydrophobic molecules, Neilson1994).
Biological mineralization of detritusrequires that following conditions are met(Schwarzenbach et al. 1993, Alexander1999):A) Environmental conditions must besuitable to support organisms.B) The detritus must be bioavailable.C) Large (>500 g mol-1) polar moleculesmust be cleaved prior uptake. Thecompounds or their cleavage productsmust be successfully transported throughcell membrane.D) The enzymes inside the cell must beable to process the detrital molecules.
Detrivores grow and survive almost in allconditions found in aquatic environments.Micro-organisms can live at temperaturesfrom -30°C to 113°C, at pH between 2 and11, and in saturated solution of salt inwater (Brock et al. 1994). Thus, theenvironmental conditions are seldom harshenough to block the activity of detrivoresin natural waters.
Detritus is bioavailable when physicalcontact between the detritus processingenzymes and detritus is possible (Alexander1999). For example, although fossil oil iseasily biodegradable by aerobic microbes, itis often not bioavailable, because the majoroil deposits are located outside the

Introduction
7
biosphere. Even if detritus exists in thesame environment together with microbes,such as sediments, the bioavailability ofdetritus may be limited. For example, if(hydrophobic) detrital molecules are tightlysorbed onto (in)organic matrix, their(solubility and) bioavailability is low, andhigh threshold concentrations (104
g C m-3)of detrital molecules are needed to supportbacterial growth (Bosma et al. 1997).Structural barriers within particulatedetritus, including thick epidermis, cuticule,bark, cell walls, woody structures, mayprotect the labile parts of detritus and thuslimit their bioavailability (Enríques et al.1993). For example, detrital leaves ofeelgrass over 1 year old still containedintact chloroplasts, possibly because theywere sheltered by the structuralcomponents of leaves (Pellikaan 1982).Some microbes can overcome thestructural barriers of particulate detritus bythemselves (e.g., Porter et al. 1989,Barnabas 1992). Typically, macroscopicdetrivores (shredders, grinders) play a keyrole in breaking the structural barriers(Fenchel 1970, Robertson & Mann 1980,Harrison 1982).
Detrivores must cleave high molecularweight detritus with extracellular enzymes.Because detritus is often of high molecularweight (>500 g mol-1), the cleavage of highmolecular weight detritus is one of themost important factors controlling the useof detritus. Hydrolytic extracellularenzymes cleave predictable biopolymerslike proteins, nucleic acids, poly-saccharides and lipids (Münster & De Haan1998). A precise contact between thesubstrate and the catalytic site of anenzyme promotes thermodynamicallyfavorable hydrolysis. Hydrolytic enzymesdo not need external energy for thecatalysis and can be active independent oftheir host cells (Münster & De Haan 1998).Some of non-specific oxidizingextracellular enzymes can cleaveheterogeneous polymers like lignin and
humic substances. These enzymes activateoxygen or manganese(II) cation, whichattacks detritus molecules with doublebonds and aromatic structures (i.e.,electron rich regions). These enzymes needreducing cosubstrates (e.g., reducednicotinamide adenine dinucleotide) and areactive only in the vicinity of their host cells.Ligninolytic fungi and some bacteriaproduce non-specific oxidizing enzymes(Kirk & Farrell 1987). These enzymes areimportant in the decomposition ofparticulate detritus in the littoral(Raghukumar et al. 1999), but their role inthe water column is low (Münster et al.1998). Manganese(II) oxidizing bacteriamay utilize manganese oxides for theoxidation of humic substances (Sunda &Kieber 1994). These bacteria oxidizeMn(II) to Mn(IV). The reaction of humicsubstances with Mn(IV) may yield Mn(II)and oxidized low molecular weightproducts such as pyruvate, acetone,acetaldehyde and formaldehyde.
If a detritus molecule succeeds in bypassingthe cell wall (and outer membrane), it mustpenetrate the cell membrane to becomemetabolized. Specific enzymes that carryout cell metabolism process detritusmolecules inside the cell. It is possible thatan organism lacks catabolic enzymes fordetritus processing and that structurallycomplex detrital molecules may thereforeresist the activity of enzymes (Neilson1994). For example, biodegradability ofpolycyclic aromatic hydrocarbons bycatabolic enzymes decreases withincreasing number of fused aromatic rings.Similarly increased number of chlorinesubstituents typically decreases bio-degradability. The unfavourable structureof detritus may, thus, retard the activity ofthe degradative enzymes.
The rate of biodegradation is regulated byvarious biotic and abiotic factors. Forexample, low temperatures may decreasethe rate of biodegradation (Wiebe et al.

Introduction
8
1992, Tulonen 1993). Microbes maysuffer from lack of energy and nutrients inaquatic environments that are oftenconsidered oligotrophic (Morita 1997).Energy gain from the hydrolysis ofpolymers and subsequent uptake of thecleavage products should supply energy forthe biosynthesis of extracellular enzymes(Vetter et al. 1998). If the concentration ofdetritus is too low to support thebiosynthesis of extracellular enzymes, thebiodegradation of high molecular weightdetritus may cease. The nutrientstoichiometry is typically different in thedetritus from that in the organisms (Hessenet al. 1994). The lack of nitrogen and/orphosphorus often limits biodegradation(Enríques et al. 1993, Elser et al. 1995,Jansson et al. 1996, Skoog et al. 1999,Vrede et al. 1999). Toxic compounds, suchas phenolic acids, in detritus may furtherhamper biodegradation (Harrison 1982,Murray & Hodson 1986). Lack of essential(co)substrates may limit biodegradationeven in presence of abundant nutrient andenergy sources (as in the sediment of aneutrophic lake). For example, lack ofoxygen or other electron acceptors in theabsence of oxygen (like nitrate, nitrite,sulphate, oxidized iron(III) ormanganese(IV)) needed for respiratorymetabolisms typically reduce the rate ofbiodegradation.
Grazing, viral attack, competition and thebehaviour of organisms regulate the rate ofbiodegradation. Grazers and viruses caneffectively control the biomass ofdetrivores (Middelboe et al. 1996). Thepresence of grazers or viruses typicallystimulates, rather than retards, bio-degradation (Middelboe et al. 1996,Zweifel et al. 1996, Strom et al. 1997,Vrede et al. 1999). Intra- and interspecificcompetition between detrivores may limitbiodegradation. In natural environments thepresence of predator, the heterogeneity
Figure 1. Dissociation energy for somebonds (Schwarzenbach et al. 1993) andenergy of photons at 300-700 nm.
of environment and environmentaldisturbances reduce potential competition(Leibold 1996, Flöder & Sommer 1999).The behaviour of detrivores, such as thesearching and trapping of detritus increasesbiodegradation. For example, chemotaxisof bacterioplankton may increase theturnover of detritus even in an apparentlyhomogenous water column (Blackburn etal. 1997).
4.4. Abiotic decomposition of detritus
Abiotic chemical reactions may transformand mineralize detritus. Decompositionreactions are by definition typicallythermodynamically favourable (change inGibbs free energy is negative). Reactants,however, need an activation energy of 50-150 kJ mol-1 before an energeticallyfavourable chemical reaction takes place(Mill 1980, Schwarzenbach et al. 1993). Inthe biosphere temperatures and pressuresare often too low to activate thermalchemical reactions and therefore detritus isstable or degrades only slowly.
Absorption of a photon excites a detritalmolecule and may lead to photochemicaldecomposition. Absorption of photons atthe UV- and the visible range of spectrum
300 400 500 600 700
150
200
250
300
350
400
450
Wavelength (nm)
Ene
rgy
of p
hoto
ns (
kJ m
ol-1
)
C-Cl
C-C
C-O
N-H

Introduction
9
increases the energy content of a moleculeby 150-400 kJ mol-1 (Fig. 1). Excitedmolecules need only an activation energy of10 to 30 kJ mol-1 for chemical reactions,and therefore photochemical degradationof detritus is possible (Cooper et al. 1989,Schwarzenbach et al. 1993). The energy ofphotons in the UV- and visible range ofspectrum is high enough to break covalentbonds (Fig. 1 and 2).
Figure 2. Dissociation energy of carbon-carbon bonds in selected compounds.
Dissociation energies of carbon-carbonbonds range from 65 kJ mol-1 of tert-butyl-triphenylmethane to 962 kJ mol-1 of ethyne(Fig. 2, Dean 1992). A typical mean isaround 360 kJ mol-1 as is found for a singleC-C bond of alkene (Fig. 1, Dean 1992).Scission of carboxyl group(decarboxylation) leads directly tomineralization. Carboxyl group of phenylacetic acid decarboxylates with an energyof 285 kJ mol-1 and that of dibenzyl aceticacid with an energy of 249 kJ mol-1 (Dean1992). Thus, absorption of photons below
480 nm by detritus may lead tomineralization (Fig. 1).
4.5. Physical and chemical factorsaffecting the decomposition of detritus
The rate of detritus degradation dependson the environmental conditions. Forexample, alkaline hydrolysis of a detritusmolecule may proceed at pH 13, but isnegligible at pH 7. Biological reactions maybe fast at pH 7, but do not proceed at pH13 because organisms cannot stand thehigh pH. Photochemical reactions maydecompose detritus during the day whenthe sun is shining but not in darknessduring the night.
Physical factors may transform particulatedetritus without breaking covalent bonds.Wind, water movements, freezing anddrought may fragment particulate detritus(Robertson & Mann 1980, Peduzzi andHerndl 1991, Mateo and Romero 1996).Physical fragmentation assists biologicaldegradation of detritus by promoting closecontact between degradative enzymes andthe detritus.
Physical and chemical processes modifyand transport detritus. Detritus may exist inthe solid (particulate), the dissolved or thegaseous phase. Physico-chemical factorsregulate the transport of detritus betweenthese phases (sorption - desorption,dissolution - volatilization). Movements ofwater and air are important vehicles inredistributing detritus. For example,dissolved detritus may flocculate andsediment in the presence of air bubbles,high ionic strength, low pH or CaCO3
(Otsuki & Wetzel 1972, Johnson & Cooke1980, Petersen 1986, Yan et al. 1996,Schindler & Curtis. 1997, Donahue et al.1998). Sedimentation transports detritusfrom the aerobic illuminated surface watersto dark, cold and possibly anaerobicenvironments. The rates and mechanisms ofdecomposition in anaerobic sediments are
tert-butyltri-phenylmethane
O
OH
phenylaceticacid
O
OH
285 kJ/mol
65 kJ/mol
dibenzylaceticacid
249 kJ/mol
ethyne
962 kJ/mol
CHHC

Introduction
10
different from those in aerobic surfacewaters.
4.6. Solar radiation and radiative transferin water
Solar radiation powers photochemicalreactions in surface waters. The sun emits arather constant flux of radiation (1450 Wm-2) to the outer edge of the atmosphere ofearth. On earth surface solar radiationvaries a lot in response to latitude, month,local time, ozone content, cloud coverage,the precense of other atmospheric aerosols,water vapour, tropospheric pollutants andaltitude (Frederick and Lubin 1988, Zeppand Cline 1977, Blumthaler et al. 1992,Frederick et al. 1989, Madronich et al.1995). In most atmospheric situations theform of solar radiation spectrum is ratherconstant. The visible range of the spectrumcovers 50%, UV-A (315-400 nm) 6% andUV-B (280-315 nm) 0.1% of globalradiation (i.e., in the wavelength range of290 to 3000 nm). The proportion of UV-Bradiation is the most variable component ofspectrum and controlled to a large extentby the content of atmospheric ozone(Molina & Molina 1986).
The absorption of a single photon is theprimary event in photochemistry, andtherefore the solar radiation is oftenpresented as the photon flux density. Thenumber of photons reaching the surface ofEarth is high. For example, a square meterof Earth's surface receives 1024
photonsbelow 500 nm during a typical summernoon hour with a global radiation of 600 Wm-2. In contrast, during the night thephoton flux density is near zero. Thus, thetemporal photon flux density may vary1024-fold.
A full understanding of photochemicalreactions needs an accurate description ofthe spectral distribution, the intensity, andthe three dimensional distribution of theincident photon flux. When radiation enters
an aquatic ecosystem, it is partly reflectedby the surface of water and its angulardistribution changes. The inherent opticalproperties of the medium dictate theradiative transfer of radiation in the watercolumn. A beam attenuation coefficient (c,m-1) describes exhaustively the inherentoptical properties and is the sum of boththe absorbing (a, m-1) and scattering (b, m-
1) components (Gordon 1989):c(z) = a(z) + b(z) (1).For a complete description of scattering itis necessary to describe the volumescattering function (Gordon 1989):
b z z( ) ( ; ) sin= �2
0
π βπ
Θ Θ Θ d (2)
where β is the volume scattering function(m-1
sr-1) and Θ is the angle betweendirections specified by (θ', ϕ ') and (θ, ϕ).The θ and ϕ are the zenith and azimuthangles in a spherical coordinate system inwhich the the z axis is directed to the nadir,and the x- and y-axes are along the surfaceof the water body. Because photons fromall directions cause photochemicalreactions, the estimation of photochemicalreaction rates requires a description of thescalar photon flux density that is thephoton flux arriving at a given point fromall directions of the sphere.
Typically neither the inherent opticalproperties nor the three dimensionaldistribution function of the incidentradiation are known in detail. Often theoptical properties of a medium aredescribed by the vertical attenuationcoefficient of downward irradiance (Kd). Kd
is calculated from the measured irradiance(I) in situ at least at two different depths (zand z+i):Kd = ln (Iz/Iz+i) (3).Kd describes the attenuation of radiation ina medium, and takes into accountabsorption and scattering. Kd dependspartly on the distribution of light field at

Introduction
11
the moment of measurement and istherefore an apparent optical property.
Kd is a useful measure for the attenuationof solar radiation in different aquaticenvironments. For example, Kd at UV-Brange of the spectrum is for oceanic waters0.12-0.15 m-1, for coastal waters 0.27-0.86m-1
and for Baltic Sea Proper 3.0-3.5 m-1
(Kirk 1994). The Kd,UV-B of most fresh-water lakes and rivers is higher than that ofthe Baltic Sea, up to extremes of 466 m-1
(Kirk 1994, Granéli et al. 1998). In otherwords, for example, 10% of the surfaceirradiation at UV-B penetrates to a depthof 20 m in Sargasso Sea (Kd,310 = 0.116 m-
1), and to a depth of 0.0049 m in adystrophic lake (Kd,UV-B = 466 m-1). Theattenuation of solar radiation in particulatedetritus is very steep, and most (99.9%) ofthe solar radiation may attenuate within amillimeter or less (i.e., with Kd > 6900 m-
1). For example, Kd may range from 2300to 27 000 m-1
in microbial mats andsediments (Kühl et al. 1997). Thus, indifferent aquatic environments theattenuation of solar radiation may varyover five orders of magnitude.
4.7. Absorption of radiation
The absorption of photon is prerequisitefor photochemical reaction and may lead toelectronic excitation of a detritus molecule(Stumm 1992). Visible light and UV-radiation are sufficiently energetic to causeelectronic excitation (Bloom & Leenheer1989). At sub-molecular levelchromophores are responsible for theabsorption of UV- and of visible radiation(Bloom & Leenheer 1989). A chromo-phore is a covalently unsaturated groupfundamentally responsible for electronicabsorption (Miller 1953). Whereas a singlechromophore such as a simple C=C doublebond has an absorption at 185 nm, this isincreased to 217 nm in 1,3-butadiene, andto 273 nm in trans-cinnamic acid (Miller1953). The presence of groups such as OH,
NH2, or S with lone pairs of electronsgenerally increases both the wavelengthand the intensity of the absorption (Miller1953). Thus, a conjugation of choromo-phores makes an absorption of solarradiation (>290 nm) possible.
Chromophoric dissolved organic matter(CDOM) absorbs large part of thephotochemically active radiation in freshand coastal waters (300-500 nm, Cooper etal. 1989, Davies-Colley and Vant 1987).CDOM has a characteristic featurelesselectronic spectrum decreasing exponen-tially towards longer wavelength with littlesignal above 500 nm. The absorptionspectrum of humic substances arepractically identical to that of CDOM.Because humic substances are the mostimportant group of detritus and arechromophoric by definition, they contributemost to absorption. In natural waters minorabsorbers are e.g., the water itself (Arrigo1994, Smith & Baker 1978), nitrate &nitrite (Zuo & Deng 1998) and degradationproducts of pigments. Suspendedparticulate matter may containchromophoric substances such aschlorophylls, carotenoids and poly-unsaturated fatty acids (Zafiriou et al.1984). Absorption of radiation by particlesmay exceed that by dissolved matter inoceanic and very clear fresh waters, ineutrophic waters, as well in the areas wherewater movements keep large amount ofparticles in suspension (rivers, turbidlittoral). However, in most fresh andcoastal waters the radiation below 500 nmis absorbed by CDOM rather than byparticles.
The rates of excitation of dissolved detritalmolecules by photons can be estimatedwhen the solar photon flux density and theabsorption characteristic of water are bothtaken into account. For example, humiclakes typically contain 1 mol DOC-C m-3
and Kd below 500 nm is greater than 5 m-1
(Hoigné 1990). Thus, CDOM in the top

Introduction
12
meter of lake water absorbs most (>99%)of photons at wavelengths <500 nm. Atnoon, typically 1024
photons m-2 below 500
nm reaches the surface during one hour. Ifthe dissolved detrital molecules absorbingsolar radiation contain 10 carbon atoms,the number of chromophoric molecules is 6× 1022
m-3. In this case solar radiationexcites each chromophore 17 times duringa noon hour if the top meter is well mixed.Thus, solar radiation has a high potential tomodify CDOM in surface waters.
4.8. Photochemical reactions
Excited molecules lose their excitationenergy via physical or chemical quenching(Mialocq 1997). In natural waters most(>90%) of the excitation energy is lost asheat (Haag & Mill 1990). Energy may belost also through emitting a photon(fluorescence or phosphorescence) orthrough transfer of energy to anotherchemical species (sensitization; Mialocq1997). In the wavelength range of 290-700nm, a small fraction (< few percent) of theexcited molecules undergoes a chemicalreaction (Fig. 3). The proportion ofabsorbed photons finally leading to achemical reaction is given as the quantumyield (φλ):
φλ = mol of product
mol of absorbed photons(4).
For example, it can be calculated from theφ290 of 0.008 for the photoproduction ofDIC (Fig. 3, Gao & Zepp 1998) that one ofthe 125 absorbed photons at 290 nm willconvert DOC to DIC. The quantum yieldsdecrease exponentially with increasingwavelength (Fig. 3). In different waters, thevariation in quantum yield is within anorder of magnitude for a single type ofreaction at a single wavelength (Fig. 3).For example, the quantum yields for thephotoproduction of hydrogen peroxide
Figure 3. Apparent quantum yields forsome photochemical reaction products ofdissolved detritus at 250-550 nm.Photoproduction of singlet oxygen (1O2)from Lake Baldegg (Haag et al. 1984).Photoproduction of DIC from the SatillaRiver (Gao & Zepp 1998). Photo-production of carbon monoxide (CO) fromfive different freshwaters (Valentine &Zepp 1993, Gao & Zepp 1998). Photo-production of hydrogen peroxide (H2O2)from the Eastern Carribean (Moore et al.1993). Photoproduction of ethene fromseawater (Ratte et al. 1998). Photo-production of carbonyl sulfide (COS) fromthe North Sea, the Gulf of Mexico and thePacific Ocean (Zepp & Andreae 1994,Weiss et al. 1995). Photoproduction ofhydroxyl radicals (�OH) from theDelaware and the Cheaspeake Bays(Vaughan & Blough 1998).
ranged from 4 × 10-5 to 23 × 10-5
in 17freshwaters (Scully et al. 1996).
Photochemical reactions can occur insolution (homogenous) or on surfaces(heterogeneous) and can be classified intoindirect and direct reactions (Hoigné 1990,Schwarzenbach et al. 1993). Directreaction occurs when the initially excitedmolecule undergoes a chemical reaction,such as fragmentation, intramolecularrearrangement, isomerization, abstractionof a hydrogen atom or dimerization(Schwarzenbach et al. 1993). For example,direct photolysis decarboxylates diaryl-
250 300 350 400 450 500 5501E-8
1E-7
1E-6
1E-5
1E-4
1E-3
0.01
0.1
Wavelength (nm)
Qua
ntum
yie
ld
DIC CO H
2O
2
ethene COS
1O2
HO

Introduction
13
acetic acids and N-[p-chloro-benzoyl]-5-methoxy-2-methylindole-3-acetic acid(Krogh & Wan 1992, Dabestani et al.1993).
Indirect photochemical reaction orphotosensitization occurs when energyfrom the initially excited molecule istransferred to another molecule, aquencher, which undergoes the chemicalreaction (Zepp et al. 1985, Hoigné 1990,Schwarzenbach et al. 1993, Mialocq 1997).In natural waters humic substances are thedominant absorbers and thus the mostimportant sensitizers for photo-chemicalreactions (Zepp et al. 1985). Energytransfer, for example, to oxygen may resultin transient reactive species like singletoxygen (1O2), superoxide anion (O2
-) andthe hydroxyl radical (HO·), or in the ratherstable hydrogen peroxide (H2O2; Hoigné1990, Scully et al. 1996). Afterphotochemical formation, the reactivespecies may undergo secondary thermalreactions (Cooper et al. 1989). It isimportant to notice that indirectphotochemical reactions may concern alsonon-chromophoric species. For example,indirect photochemical reaction candecarboxylate sulphur-containing aminoacids and indole-3-acetic acid (Bobrowskiet al. 1992, Brennan 1996).
Heterogeneous photochemical reactionstake place on the surfaces of particlessuspended in the water column or on thesediments of the photic zone. For example,hematite (α-Fe2O3) catalyses photo-chemical oxidation of oxalate to CO2
(Siffert & Sulzberger 1991) andlepidocrocite (γ-FeOOH) photocatalyzesdecomposition of ethylenediaminetetraacetic acid (EDTA) and of fulvic acid(Karametaxas et al. 1995, Voelker et al.1997). The oxidation of organic compoudson the surface of particles may occur viasemiconductor mechanism. Absorption of aphoton by a semiconductor particletransports an electron from a valence band
to a conduction band leaving behind anelectron vacancy, a photohole. A photo-hole seeks desperately for an electron andmay oxidize species on surface of thesemiconductor. A photoelectron in theconduction band may reduce adsorbedspecies. In the complexes of metal andorganic compounds, the photochemicaldecomposition may proceed via ligand tometal charge transfer (Siffert & Sulzberger1991, Karametaxas et al. 1995). Forexample, the presence of dissolved ironenhances photochemical degradation offulvic acids, dicarboxylic acids and α-ketoacids (Faust & Zepp 1993, Voelker et al.1997, Gao & Zepp 1998).
4.9. Predicting the rates of photochemicalreactions
The rates of photochemical reactions canbe estimated from the product of threeparameters: (1) scalar photon flux density,(2) the absorption of photons by themedium and (3) the quantum yield of thereaction (Zepp & Cline 1977, Miller 1998).The product of (1) and (2) will give therate of photon absorption. The quantumyield (3) reveals how many absorbedphotons will result in a photochemicalreaction. A mathematical presentation ofthe reaction rate at the depth z is:
reaction ratez =
λ
λ
min
max
� φλ Qs,z,λ aλ dλ (5)
where φλ is the quantum yield of reactionat λ (dimensionless), Qs,z,λ is the scalarphoton flux density at the depth z and thewavelength λ (mol of photons m-2
s-1), aλ isthe absorption coefficient of the medium atthe wavelength λ (m-1), λmin and λmax arethe minimum and maximum wave-lengths(nm) contributing to the photo-chemicalreaction. Since the three parameters of Eq.5 depend on wavelength, the prediction ofrate requires an integration overwavelengths. A photo-chemical reaction,like the mineralization of organic carbon

Introduction
14
(see 4.8.), may proceed via severalreaction mechanisms and each reaction
mechanism has its own φλ. The sum of
individual φλ will give the total φλ for thereaction.
4.10. Photochemical reactions related tothe processing of detritus
Photochemical reactions decrease thearomaticity of the humic substances andincrease the number of aliphatic structures,carbonyl and carboxylic groups (Kulovaaraet al. 1996). Although photochemicalreactions may increase the molecularweight of dissolved detritus (Backlund1992, Kieber et al. 1997), most often themolecular weight of dissolved detritusdecreases (Strome and Miller 1978, Geller1985, De Haan 1993, Hongve 1994). Themost abundant photoproduct is CO2
(Gjessing & Gjerdahl 1970, Chen et al.1978, Miller & Zepp 1995, Granéli et al.1996, 1998, Gao & Zepp 1998, Miller &Moran 1997, Moran & Zepp 1997). Theother low molecular weight compoundsinclude those given in Fig. 4. Many organicphotoproducts are substrates for bacteriaand will be biodegraded.
Photochemical reactions may mobilizedetritus bound nutrients. Photochemicalreactions release inorganic phosphorus andnitrogen from organic or from complexedcompounds (e.g., humic substance-iron-phosphorus complexes; Manny et al. 1971,Cotner & Heath 1990, Bushaw et al.1996). Photochemically released nutrientsmay provide nutrients for detrivores andthus increase biodegradation (Bushaw et al.1996).
The paragraphs above describe thereactions of bulk dissolved detritus. Anearly review by Roof (1982) describes thephotochemical degradation of 100antropogenic chemicals, but few studiesconcern chemically well characterized
natural compounds. Photodegradation ofsenescent phytoplankton leads todegradation of chlorophylls, chlorophyllphytyl chains, carotenoids, sterols,unsaturated fatty acids, and cyanobacterialtoxins (Rotani 1998, Choswell et al. 1999).Lignin-related photochemical reactionsresult in cleavage of methoxy groups andformation of quinones, hydroxyl, carboxyland keto groups (Weir et al. 1994, 1996,Argyropoulos & Sun 1996). Photo-chemical reactions break bonds betweenpolymer bound phenoxy monomers andrelease low molecular weight compounds(Gold et al. 1983, Castellan et al. 1987,Argyropoulos & Sun 1996). For example,Sun et al. (1998) identified 26 aliphaticacids after photodegradation of lignin.Solar radiation decomposes lignin in thedetrital leaves of seagrass and modifieslignin residues also in the water column(Opshal & Benner 1993, 1998).
While photochemical production of lowmolecular weight organic substrates andbioavailable nutrients may increasebiodegradation, opposite effects have alsobeen reported. Solar radiation decreasesthe biodegradation of protein and otherlabile detritus (Naganuma et al. 1996,Benner & Biddanda 1998, Tranvik &Kokalj 1998, Anesio et al. 1999,Obernosterer et al. 1999). UV radiationmay generate long (days) lived toxiccompounds, which reduce the growth ofphytoplankton (Hessen & Van Donk 1994)and of bacteria (Lund & Hongve 1994,Anesio et al. 1999).
4.11. Effects of solar radiation ondetrivores
Inside a cell photochemical reactions arenoxious for the highly structured bio-chemical machinery taking care of thefunctions of life. Solar UV-B radiationcauses photodimerization of adjacent
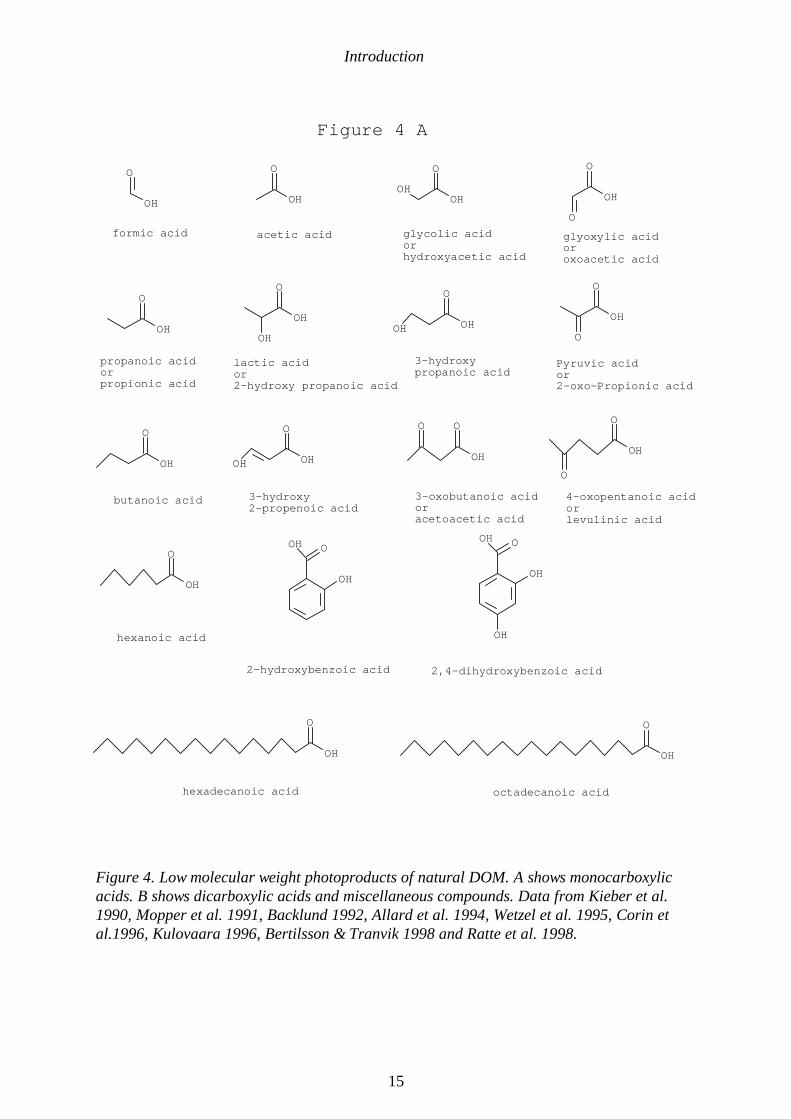
Introduction
15
Figure 4. Low molecular weight photoproducts of natural DOM. A shows monocarboxylicacids. B shows dicarboxylic acids and miscellaneous compounds. Data from Kieber et al.1990, Mopper et al. 1991, Backlund 1992, Allard et al. 1994, Wetzel et al. 1995, Corin etal.1996, Kulovaara 1996, Bertilsson & Tranvik 1998 and Ratte et al. 1998.
O
OH
OH
OH
O
formic acid
OH
O
acetic acid
OH
O
O
glyoxylic acidoroxoacetic acid
lactic acidor2-hydroxy propanoic acid
OH
O
propanoic acidorpropionic acid
OH
O
OH
glycolic acidorhydroxyacetic acid
OOH
OH
2-hydroxybenzoic acid
OOH
OH
OH
2,4-dihydroxybenzoic acid
hexanoic acid
hexadecanoic acid octadecanoic acid
O
OH
O
Pyruvic acidor2-oxo-Propionic acid
O
OHOH
3-hydroxypropanoic acid
O
OHOH
3-hydroxy2-propenoic acid
OH
O
O
4-oxopentanoic acidorlevulinic acid
OH
OO
3-oxobutanoic acidoracetoacetic acid
butanoic acid
OH
O
OH
O
OH
O
OH
O
Figure 4 A
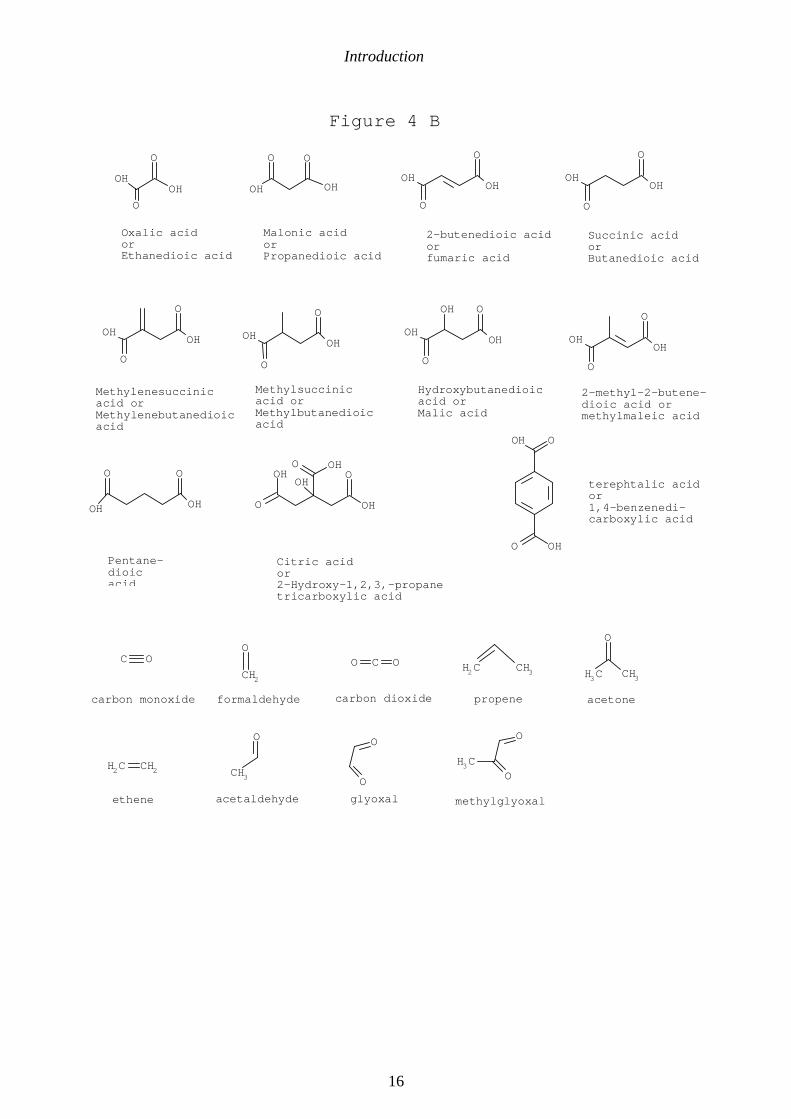
Introduction
16
OH
O
O
OH
Oxalic acidorEthanedioic acid
OH
O
OH
O
Malonic acidorPropanedioic acid
OH
O
OH
O
Succinic acidorButanedioic acid
OH
OOH
OHO
O
OH
Citric acidor2-Hydroxy-1,2,3,-propanetricarboxylic acid
OH
O
OH
O
Methylsuccinicacid orMethylbutanedioicacid
OH
O
OH
O
Methylenesuccinicacid orMethylenebutanedioicacid
OH
O
OH
O
2-butenedioic acidorfumaric acid
OH
OO
OH
Pentane-dioicacid
OH
O
OH
O
OH
Hydroxybutanedioicacid orMalic acid
OH
O
OH
O
2-methyl-2-butene-dioic acid ormethylmaleic acid
O OH
OOH
terephtalic acidor1,4-benzenedi-carboxylic acid
CH2
O
formaldehyde
acetaldehyde
O
O
O
CH3
carbon monoxide carbon dioxide acetone
H3C CH3
O
O
OH3C
methylglyoxal
H2C CH2
H2C CH3
ethene
propene
glyoxal
Figure 4 B
C O C OO

Introduction
17
pyrimidine bases and photohydration ofcytosine (Jeffrey et al. 1996, Weinbauer etal. 1999). Other, less frequent DNA-damage includes intra- and inter-strandcross links, single-strand breaks, protein-DNA cross links and base adducts (Audic& Giacomoni 1993, Miller et al. 1999).Many bacteria (25 to 65% of marineisolates) carry lysogenic prophages (Jiang& Paul 1998). After exposure to UV-radiation, prophages may initiate a lyticcycle and destroy the bacterial host (Jiang& Paul 1998). UV-radiation is known todamage enzymes and decrease their activity(Müller-Niklas et al. 1995, Zigman et al.1996). On the other hand, UV-radiationmay increase enzyme activity in humicwaters, possibly by breaking the complexesbetween humic substances and enzymes(Wetzel et al. 1995). The negative effectsof UV-radiation may decrease the bacterialproduction in surface waters (Herndl et al.1993, Müller-Niklas et al. 1995, Pakulskiet al. 1998). Solar radiation causesselective losses of UV-sensitivepopulations (Bothwell et al. 1994). TheUV-resistant populations survive and mayincrease in density. Thus solar radiation isan important factor determining thestructure of a biological community(Bothwell et al. 1994).
Organisms can protect themselves andrecover from the negative effects of solarradiation (Herndl et al. 1993, Müller-Niklaset al. 1995, Pakulski et al. 1998). Catalase,superoxide dismutase, glutathionereductase and glutathione peroxidaseprotect cells against reactive oxygenspecies. Antioxidants (like carotenoids andflavonoids) serve the same function(Krasnovsky 1994). Protective pigments(carotenoids, mycosporin, melanin) absorbharmful UV-radiation and can act asinternal sun screens (Zellmer 1995).However, at the level of a single bacterialcell (radius <1µm), the optical depth ofprotective pigments may be too narrow tobe effective (Garcia-Pichel 1994). In the
water column CDOM absorbs effectivelysolar UV and forms an effective externalsun screen. Some organisms can escapeharmful solar radiation (Williamson 1995).For example, vertically migratingzooplankton enter the surface waters atnight and spend the daytime in deep water.After photodamage, cells may recover e.g.,by the action of DNA repair enzymes(Herndl et al. 1993, Müller-Niklas et al.1995, Pakulski et al. 1998, Miller et al.1999).
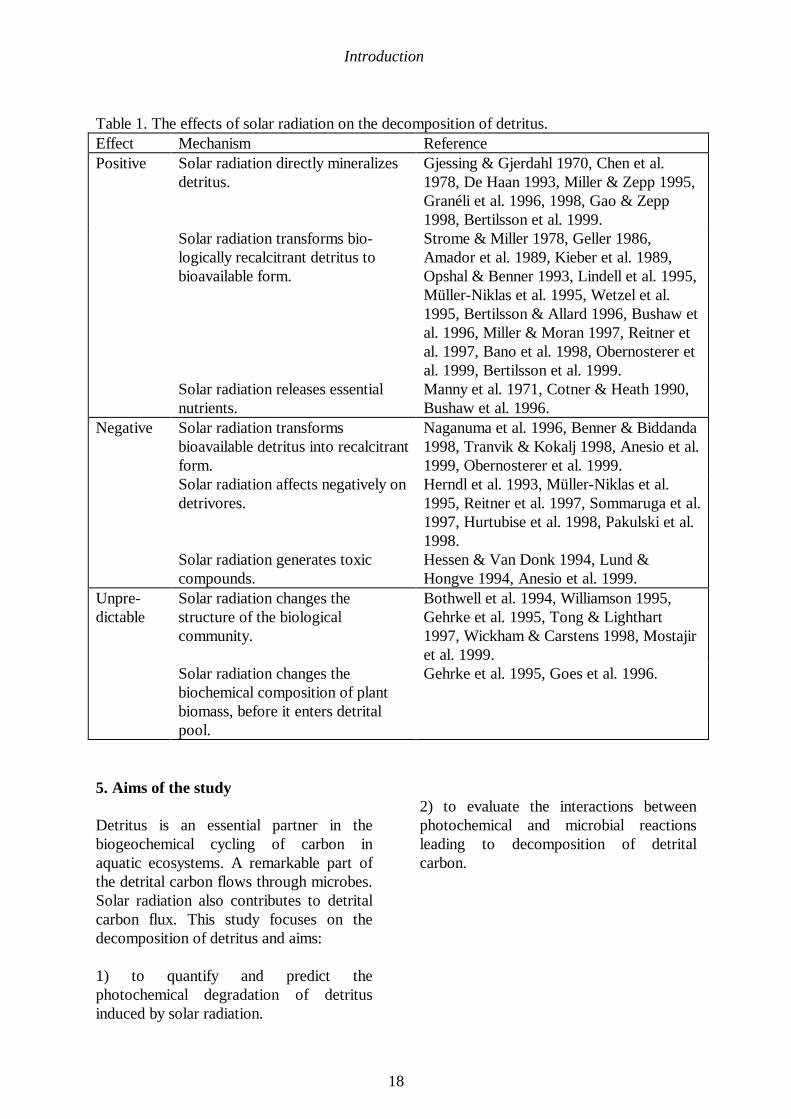
The chapters 4.10. and 4.11. indicate thatsolar radiation affects the rate ofdecomposition in positive, or negative orhitherto unpredictable ways. These effects(summarized in Table 1) act simulta-neously in natural environments. Theimportance of each mechanism and theoverall effect of solar radiation depends onthe ecological conditions in eachecosystem.
Introduction
18
Table 1. The effects of solar radiation on the decomposition of detritus.Effect Mechanism ReferencePositive Solar radiation directly mineralizes
detritus.Gjessing & Gjerdahl 1970, Chen et al.1978, De Haan 1993, Miller & Zepp 1995,Granéli et al. 1996, 1998, Gao & Zepp1998, Bertilsson et al. 1999.
Solar radiation transforms bio-logically recalcitrant detritus tobioavailable form.
Strome & Miller 1978, Geller 1986,Amador et al. 1989, Kieber et al. 1989,Opshal & Benner 1993, Lindell et al. 1995,Müller-Niklas et al. 1995, Wetzel et al.1995, Bertilsson & Allard 1996, Bushaw etal. 1996, Miller & Moran 1997, Reitner etal. 1997, Bano et al. 1998, Obernosterer etal. 1999, Bertilsson et al. 1999.
Solar radiation releases essentialnutrients.
Manny et al. 1971, Cotner & Heath 1990,Bushaw et al. 1996.
Negative Solar radiation transformsbioavailable detritus into recalcitrantform.
Naganuma et al. 1996, Benner & Biddanda1998, Tranvik & Kokalj 1998, Anesio et al.1999, Obernosterer et al. 1999.
Solar radiation affects negatively ondetrivores.
Herndl et al. 1993, Müller-Niklas et al.1995, Reitner et al. 1997, Sommaruga et al.1997, Hurtubise et al. 1998, Pakulski et al.1998.
Solar radiation generates toxiccompounds.
Hessen & Van Donk 1994, Lund &Hongve 1994, Anesio et al. 1999.
Unpre-dictable
Solar radiation changes thestructure of the biologicalcommunity.
Bothwell et al. 1994, Williamson 1995,Gehrke et al. 1995, Tong & Lighthart1997, Wickham & Carstens 1998, Mostajiret al. 1999.
Solar radiation changes thebiochemical composition of plantbiomass, before it enters detritalpool.
Gehrke et al. 1995, Goes et al. 1996.
5. Aims of the study
Detritus is an essential partner in thebiogeochemical cycling of carbon inaquatic ecosystems. A remarkable part ofthe detrital carbon flows through microbes.Solar radiation also contributes to detritalcarbon flux. This study focuses on thedecomposition of detritus and aims:
1) to quantify and predict thephotochemical degradation of detritusinduced by solar radiation.
2) to evaluate the interactions betweenphotochemical and microbial reactionsleading to decomposition of detritalcarbon.

19
Table 2. The study sites, the detritus and a source for further information.Site Detritus Further informationA humic Lake Skervatjern, Norway DOC of lake water Gjessing 1992A humic Lake Valkea-Kotinen, Finland DOC of lake water Rask et al. 1998, Keskitalo et al. 1998
14C-[ring]-labelled lignin Faix et al. 1985, Eriksson et al. 1990Marine littoral in Roskilde Fjord, Denmark Eelgrass (Zostera marina, L.) Kamp-Nielsen 1992, Pellikaan 1984
Table 3. The summary of the methods.Target Method Further informationDissolved inorganic carbon Acidify-purge-infrared method, Universal carbon analyser Salonen 1981Organic carbon Combustion infrared method, Universal carbon analyser and
Shimadzu TOC-5000Salonen 1979,Søndergaard & Middelboe 1993
Phosphate Ascorbic acid method, AKEA-autoanalyser, APHA 1998Nitrate + nitrite Cadmium reduction method, AKEA-autoanalyser, APHA 1998Ammonium Phenate method, Colorimetry, APHA 1998Total nitrogen and phosphorus Persulfate digestion, AKEA-autoanalyser APHA 1998Ash free dry weight Ignition for 2 h at 550°C, an electrobalancePigments Shimadzu LC-10A HPLC Jeffrey 1997, Wright et al. 1991Absorption of CDOM Shimadzu UV-2100 and UV-2501PC spectrophotometers Bricaud et al. 1981Optics of leaves LI-1800 spectroradiometer with an intergrating sphere,
Shimadzu UV-160A spectrophotometerLi-Cor 1982,Vähätalo et al. 1998
Colour of water Absorption at 420 nm, Cobolt-plantinum standards APHA 1998Spectroradiometry Macam SR991 spectroradiometer Http://www.macam.comPhoton flux density (400-700 nm) Li-Cor, LI-190Sa quantum sensor Li-Cor 1982Global radiation BPW 20 photodiode, Hortimic Oy Strangeways 1996Ozone M-124 ozonometer, Brewer spectrophotometerQuantum yield in laboratory Increase in DIC, Chemical actinometry Becker et al. 1976Quantum yield in situ Increase in DIC, Quasi-measured spectra Vähätalo et al. 2000Respiration Dark bottle method, Increase in DIC APHA 1998Biovolume of bacteria Epifluorescence microscopy Bergström et al. 1986Bacterial activity Incorporation of 14C-leucine Hollibaugh & Wong 199214C-activity Wallac 1400 liquid scintillation counter

Results
20
6. Material & Methods
The sites and the detritus used in this studyare shown in Table 2. Decomposition ofdetritus was studied by exposing thedetritus to solar radiation or to indigenousmicro-organisms of the study sites. Themethods used in the decomposition studiesare summarized in Table 3.
7. Results
7.1. Mineralization of DOC in humic lakes
To quantify the biological mineralization ofDOC, I measured microbial respiration in<2 µm filtered water (12 g of DOC-C m-3)of the humic Lake Valkea-Kotinen,Finland. Planktonic microbes mineralized45 mg of DOC m-3
d-1 (Fig. 3 in III) and
470 mg of DOC m-3 in 7 d during the
summer (Table 2 in III). Microorganisms,thus, mineralized <4% of the DOC in aweek (Fig. 2 in III) indicating biologicalrecalcitrance of the bulk of DOC in LakeValkea-Kotinen.
To compare with the microbialmineralization, I also determined the solarradiation induced photochemical minera-lization in Lake Valkea-Kotinen. Solarradiation mineralized 230 mg of DOC m-3
d-1 (Fig. 3 in II, Fig. 3 in III) and 2200 mg
DOC m-3 in 7 d (Table 2 in III) at the depth
of 1 cm. Thus, the photochemical minera-lization exceeded the microbial minera-lization of DOC four fold at the depth of 1cm (Fig. 3 in II, Fig. 3 and Table 2 in III).The magnitude of photomineralization wassimilar in another humic Lake Skervatjern,Norway (Figs. 1 and 4 in I). The DOC inthe water flowing into Lake Skervatjernwas the most sensitive for photochemicalmineralization followed by the DOC of thehypolimnetic and of the epilimnetic waterof the lake (Figs. 3 and 5 in I). Theseresults suggest that the rate ofphotochemical minera-lization of DOC canbe high near the surface of humic lakes and
that the sensitivity of DOC forphotochemical mineralization may vary indifferent parts of the watershed (Figs. 3and 5 in I).
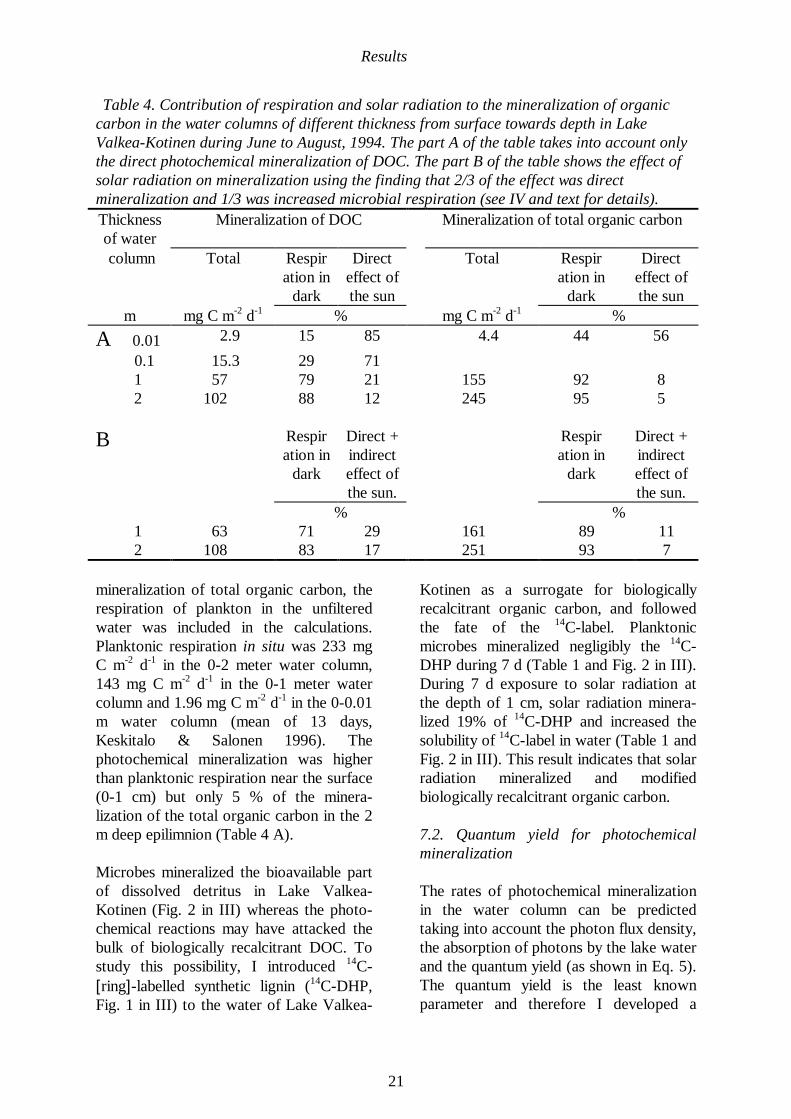
To compare the contributions of photo-chemical mineralization of DOC andplanktonic respiration to the mineralizationof organic carbon in the water column ofLake Valkea-Kotinen, the depth integratedmineralization yields were calculated forthe summer 1994 (Table 4 A). Themineralization of DOC was considered toconsist of two components, the photo-chemical mineralization of DOC and themicrobial respiration in <1 µm filteredwater. The dark microbial respiration was4.5 mg C m-2
d-1, when integrated over thetop 10 cm (mean of 10 days, Fig. 3 in III).This rate of microbial respiration was usedalso to calculate the dark microbialrespiration in Lake Valkea-Kotinen overother depths to 2 m (Table 4 A). Therespiration was measured in darkness fromthe water collected in early morningsrepresenting the minimal effect of solarradiation. The depth-integrated photo-chemical mineralization of DOC wascalculated from pmz = 270 mg C m-3
d-1 e-
23 z, where the 270 mg C m-3 d-1
is the rateof mineralization extrapolated to z = 0 m,pmz is the mineralization rate at the depth zand 23 m-1
is the vertical attenuationcoefficient for the photo-chemicalmineralization (mean of 11 days, Fig. 3 inII). The contribution of photochemicalminera-lization to the mineralization ofDOC was high near (0-10 cm) the lakesurface and attenuated steeply with anincreasing depth (Table 4 A, Fig. 3 in II,see also Figs. 1 and 4 in I for LakeSkervatjern). In the 2 m deep epilimnion ofLake Valkea-Kotinen photochemicalmineralization of DOC was 12 mg of C m-2
d-1 (Fig. 4 in II) meaning that
bacterioplankton was the dominant catalystof DOC mineralization in this layer (Table4 A). To further quantify the role ofphotochemical reactions in the
Results
21
Table 4. Contribution of respiration and solar radiation to the mineralization of organiccarbon in the water columns of different thickness from surface towards depth in LakeValkea-Kotinen during June to August, 1994. The part A of the table takes into account onlythe direct photochemical mineralization of DOC. The part B of the table shows the effect ofsolar radiation on mineralization using the finding that 2/3 of the effect was directmineralization and 1/3 was increased microbial respiration (see IV and text for details).Thicknessof water
Mineralization of DOC Mineralization of total organic carbon
column Total Respiration in
dark
Directeffect ofthe sun
Total Respiration in
dark
Directeffect ofthe sun
m mg C m-2 d-1 % mg C m-2
d-1 %
A 0.01 2.9 15 85 4.4 44 56
0.1 15.3 29 71 1 57 79 21 155 92 8 2 102 88 12 245 95 5
B Respiration in
dark
Direct +indirecteffect ofthe sun.
Respiration in
dark
Direct +indirecteffect ofthe sun.
% % 1 63 71 29 161 89 11 2 108 83 17 251 93 7
mineralization of total organic carbon, therespiration of plankton in the unfilteredwater was included in the calculations.Planktonic respiration in situ was 233 mgC m-2
d-1 in the 0-2 meter water column,
143 mg C m-2 d-1
in the 0-1 meter watercolumn and 1.96 mg C m-2
d-1 in the 0-0.01
m water column (mean of 13 days,Keskitalo & Salonen 1996). Thephotochemical mineralization was higherthan planktonic respiration near the surface(0-1 cm) but only 5 % of the minera-lization of the total organic carbon in the 2m deep epilimnion (Table 4 A).
Microbes mineralized the bioavailable partof dissolved detritus in Lake Valkea-Kotinen (Fig. 2 in III) whereas the photo-chemical reactions may have attacked thebulk of biologically recalcitrant DOC. Tostudy this possibility, I introduced
14C-[ring]-labelled synthetic lignin (14C-DHP,Fig. 1 in III) to the water of Lake Valkea-
Kotinen as a surrogate for biologicallyrecalcitrant organic carbon, and followedthe fate of the
14C-label. Planktonicmicrobes mineralized negligibly the
14C-DHP during 7 d (Table 1 and Fig. 2 in III).During 7 d exposure to solar radiation atthe depth of 1 cm, solar radiation minera-lized 19% of 14C-DHP and increased thesolubility of 14C-label in water (Table 1 andFig. 2 in III). This result indicates that solarradiation mineralized and modifiedbiologically recalcitrant organic carbon.
7.2. Quantum yield for photochemicalmineralization
The rates of photochemical mineralizationin the water column can be predictedtaking into account the photon flux density,the absorption of photons by the lake waterand the quantum yield (as shown in Eq. 5).The quantum yield is the least knownparameter and therefore I developed a

Results
22
method for determining the spectrum ofquantum yield for the photo-chemical
mineralization of DOC (φλ, II). The keyissue in the method was the description ofφλ as c × 10-dλ, where c (dimensionless)and d (nm-1) are positive constants. It was
possible to determine φλ based on (1) theabsorption spectrum of CDOM (Fig. 7 A inII), (2) the in situ measured photochemicalmineralization at the depth z (Fig. 3 in II),and (3) the solar spectrum of photon fluxdensity at the same depth (Fig. 7 C in II).
In Lake Valkea-Kotinen φλ was 7.52 × 10-
0.0122λ (Fig. 7 B in II) describing thequantum yield spectrum for thephotochemical mineralization of DOCbetween 300 nm and 700 nm. The quantum
yields calculated from the φλ of 7.52 × 10-
0.0122λ agreed with the quantum yieldsdetermined in a laboratory at the singlewavelengths of 320, 355 and 390 nm(Table 2 in II). This agreement suggeststhat the developed method resulted in a
reliable determination of φλ in situ.
I tested whether Eq. 5 with the observed φλcorrectly predicted the rates of photo-chemical mineralization of DOC in LakeValkea-Kotinen. The predicted rates ofphotochemical mineralization agreed withthe measured ones at the depths of 1-10 cmand in the whole water column (Figs. 8-9 inII). The modelling with Eq. 5 revealed thatthe UV-A part of solar radiationmineralized DOC most (Fig. 7 E and Table3 in II). The attenuation of UV-A in thewater column described roughly theattenuation of photochemical minera-lization (compare Figs. 2, 3 and 7 in II).
Thus, the φλ applied in Eq. 5 allowed areliable prediction of the rates of photo-chemical mineralization and gave spectraldetails of photochemical mineralization ofDOC.
7.3. Photochemical modification of DOCand it's effects on bacteria
I used a microbial assay for quantifying thechanges in the bioavailability of DOCduring exposure to solar radiation. Theexposure of <0.2 µm filtered water fromLake Valkea-Kotinen to solar radiation fora day resulted in the mineralization of 250-800 mg of DOC m-3
d-1 (Fig. 1 in IV).
Mineralization was negligible in the non-exposed water kept in dark (Fig. 1 in IV).The light exposed and the non-exposedwaters were spiked with indigenousmicrobes from Lake Valkea-Kotinen (<1µm filtered water) and cultured for 4 d indarkness. During culture, the bacteriamineralized 2 to 5 fold more of DOC in thesolar radiation exposed water than in thenon-exposed water (Figs. 5-6 in IV). Thus,solar radiation mineralized DOC indirectlyby enhancing microbial minera-lization ofDOC.
The effect of solar radiation on themineralization of DOC, thus consisted oftwo components, (1) the direct photo-chemical mineralization of DOC and (2) theindirect microbial mineralization of DOC,which increased after the solar radiationexposure of DOM. To quantify the relativemagnitude of the direct and the indirectcomponents, I compared the magnitude ofphotochemical minera-lization of DOC(Fig. 1 in IV) to that of the solar radiationstimulated increase in the microbialrespiration during the 4 d long culturing(Fig. 6 in IV). The respiration was higherby 159-189 mg C m-3
in the solar radiationexposed water than in the non-exposedwater collected from 0-1 m depth in Juneto August (Fig. 6 in IV). This increasedrespiration was 46-51% (mean 49%, n = 3)of the photo-chemical mineralization ofDOC during the initial irradiation (319-372mg C m-3; see Figs. 1 and 6 in IV). Thisresult indicates that the totalphotochemically induced mineralization ofDOC consisted of 2/3 from the directphotochemical mineralization and of 1/3from the enhanced microbial respiration.

Results
23
I used the above estimate to recalculate themineralization by solar radiation in the 1and 2 m thick water columns of LakeValkea-Kotinen (Table 4 B). Since thedirect photochemical mineralization ofDOC was 12 mg C m-2
d-1 in the water
column of Lake Valkea-Kotinen (Fig. 4 inII), the indirect component was calculatedto be half of that, 6 mg C m-2
d-1 (Table 4
B). When this is applied into thecalculations, the sum of direct and indirecteffects of solar radiation on themineralization of DOC was 17 % in the 2m thick water column of Lake Valkea-Kotinen (Table 4 B).
During the culturing of the microbes fromLake Valkea-Kotinen in the exposed andnon-exposed waters, the concentration ofdissolved (permeate of GF/F-filter)phosphorus and nitrogen decreased, andthe amount of particulate (retained byGF/F-filter) nitrogen and phosphorusincreased (Figs. 2-3 in IV). These changesin the nutrient pools indicated theconsumption of dissolved nutrients bybacteria and the simultaneous generation ofparticulate nutrients bound to bacterialbiomass. The concentrations of particulatenitrogen and phosphorus were typically 1.5times higher in the solar radiation exposedlake water than in the non-exposed waterkept in darkness (Figs. 2-4 in IV). Thisindicates increased production of bacterialbiomass in the solar radiation exposedwater. Microscopic counting of bacterialbiomass gave similar result. After the 2-4 dof culturing, the biovolume of the bacterialcommunity in the water exposed to solarradiation was 1.7 fold higher than in thenon-exposed water kept in dark (Fig. 5).Thus, the exposure of water from LakeValkea-Kotinen to solar radiation increasedthe production of bacteria.
During the culturing of the microbes fromLake Valkea-Kotinen, the concentration ofphosphate decreased below the detection
limit of the method (<2 mg PO4-P m-3, Fig.2 of IV) indicating that bacteria scavengedinorganic phosphorus into the biomass. Atthe beginning of the experiments, theC:N:P mass ratio in the DOM was2500:75:1 (n = 12) and far greater than the100:20:1 found on the GF/F filters (n = 12)representing bacterial biomass. Since theconcentration of phosphorus was low bothin DOM and in inorganic form (Fig. 2 inIV), it was plausible that the growth ofbacteria had been limited by phosphorus.
7.4. Photochemical modification ofdetrital eelgrass
I expected that solar radiation may affect,not only the dissolved detritus in lakewater, but also the particulate detritus inwater. To investigate this, I exposed darkbrown heat sterilized detrital leaves ofeelgrass (Zostera marina) to solar radiation(Fig. 6; Figs. 2 and 3 in V). During a 30 dexposure to solar radiation the detritalleaves bleached, but no bleaching wasobserved in the non-exposed leaves (Fig. 6;Figs. 4-5 in V). The leaves lost theirchlorophyll a already after an exposure of<5 days to solar radiation (Fig. 6 in V) andphotobleached leaves were devoid ofpigments (Fig. 3 and Table 1 in V). Solarradiation enhanced loss of organic matterfrom the leaves (Fig. 7 and Table 2 in V).The loss was caused mainly due to leachingof dissolved organic compounds (Table 2and Fig. 8 C in V) and to a small extentdue to fragmentation (Fig. 9 in V). Thus,solar radiation alone decomposed detritalleaves of eelgrass.
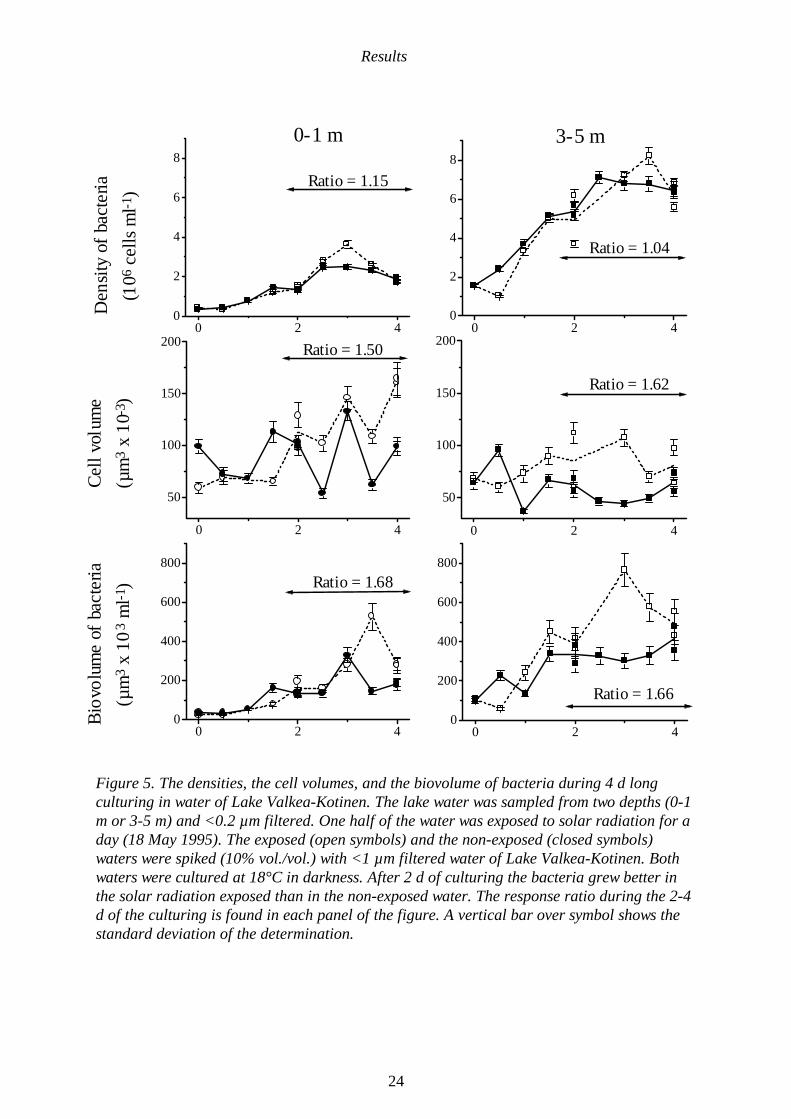
Results
24
Figure 5. The densities, the cell volumes, and the biovolume of bacteria during 4 d longculturing in water of Lake Valkea-Kotinen. The lake water was sampled from two depths (0-1m or 3-5 m) and <0.2 µm filtered. One half of the water was exposed to solar radiation for aday (18 May 1995). The exposed (open symbols) and the non-exposed (closed symbols)waters were spiked (10% vol./vol.) with <1 µm filtered water of Lake Valkea-Kotinen. Bothwaters were cultured at 18°C in darkness. After 2 d of culturing the bacteria grew better inthe solar radiation exposed than in the non-exposed water. The response ratio during the 2-4d of the culturing is found in each panel of the figure. A vertical bar over symbol shows thestandard deviation of the determination.
0 2 40
2
4
6
8
0 2 4
50
100
150
200
Ratio = 1.62
0 2 40
200
400
600
800
Ratio = 1.66
Ratio = 1.68
Ratio = 1.04
Ratio = 1.15
3-5 m0-1 m
Bio
volu
me
of b
acte
ria
(µm
3 x
10 3
ml-1
)
Cel
l vol
ume
(µm
3 x
10-3
)D
ensi
ty o
f ba
cter
ia
(106
cel
ls m
l-1)
0 2 40
200
400
600
800
0 2 40
2
4
6
8
0 2 4
50
100
150
200Ratio = 1.50

Results
25
Figure 6. Pieces of detrital eelgrass initially (0 d), after 30 d incubation in darkness andafter 30 d exposure to solar radiation.
To evaluate the impact of solar radiationon a microbial biofilm, the sterile leavesafter solar radiation exposure were spikedwith indigenous microbes from thesampling site (<GF/F filtered seawaterfrom Roskilde Fjord, Denmark). Microbialactivity was measured as the incorporationof 14C-leucine into microbial macro-molecules after 6 d of culturing in the dark.
Incorporation of 14C-leucine by themicrobial biofilm on leaf surfaces was 2-3fold higher on the solar radiation exposedleaves than on the leaves kept in the dark(Fig. 10 in V). The result indicates thatsolar radiation converted the detrital leavesinto a more suitable habitat fordevelopment of a microbial biofilm.
Initial 0 d
30 d exposureto solar radiation
30 d incubationin darkness

Discussion
26
8. Discussion
8.1. The relative role of bacterioplanktonand photochemical mineralization
The microbial mineralization of organiccarbon in the epilimnion of a lake is thesum of photochemical mineralization andthe respiration of plankton. Based onsimultaneous in situ measurements of theDIC production by microbes, by theplankton and by photomineralization overthree summer months, the contribution ofplankton to the mineralization of organiccarbon was >88% in the 2 m thickepilimnion of the humic Lake Valkea-Kotinen (61°N, Table 4 A). Granéli et al.(1996) used similar methods during a singlesummer day and showed that planktoncontributed 90 to 93% to themineralization of the total organic carbonin the epilimnions of four Swedish lakes(57°N). These lakes ranged from a clear (0g Pt m-3) to humic water lakes (140 g Ptm-3), and had epilimnetic depths from 1.5to 7.5 m (Granéli et al. 1996). Reitner et al.(1997) calculated from the rates of oxygenconsumption that 90% of the minera-lization of DOC was due to bacterio-plankton in the 1 meter deep brownwatered Lake Neusiedl (Austria, 47°N)dominated by Phragmites australis stands.The conclusion from this study, and thoseof Granéli et al. (1996) and Reitner et al.(1997) is that bacterioplankton, rather thanphotochemical mineralization, dominatedthe total mineralization of DOC in theepilimnion of the boreal and temperatelakes that have been studied.
8.2. Photomineralization and primaryproduction
The concentration of DIC was 0.2-0.3 g Cm-3
in the surface of the lakes Valkea-Kotinen and Lake Skervatjern during thesummer (I, II, Keskitalo et al. 1998). Thedaily photochemical mineralization of DOC
doubled the concentration of DIC (Fig. 3 inII, Figs. 1, 3-4 in I). The concentration ofDIC is a parameter in the determination ofprimary production with the
14CO2 method(APHA 1998). If the doubling of DIC byphotochemical mineralization is ignoredduring the determination of primaryproduction, the
14CO2 method willunderestimate the primary production by afactor of 0.5. The error is largest in thesurface of acidic humic lakes, where theconcentration of DIC is low and the rate ofphotochemical mineralization high (I, II).
8.3. Photodegradation of vascular plantdetritus
The sterile detrital leaves of eelgrass lost30% of their organic matter in 30 d after aglobal radiation load of 870 MJ m-2, whilethe loss in darkness was 10% during thesame period (Fig. 7 in V). Detrital leaves ofPhragmites australis lost 35% of dryweight in non-sterile microcosms after theglobal radiation load of ca. 900 MJ m-2
(Denward & Tranvik 1998). The loss indark microcosms was 25%, which is lessthan in the microcosms exposed to fullsolar radiation (Denward & Tranvik 1998).The detrital leaves of Typha latifolia,exposed to global radiation of ca. 470 MJm-2
at the depth of 20-25 cm in a Talldegawetland, lost 15 % of dry weight, and only10% in the shade (Neely & Wetzel 1997).Carbon budgets made for the decom-position of eelgrass and of Phragmitessuggest that leaching of DOC accountedfor most (>85%) of the solar radiation-induced mass loss from the leaves (Table 2in V; Denward & Tranvik 1998).
Lignin is ubiquitous in vascular plants,where it accounts for the structuralintegrity of the plant (Eriksson et al. 1990).Lignin is susceptible to photochemicaldecomposition (Eriksson et al. 1990,Opsahl & Benner 1993, III). Photo-chemical decomposition of lignin possibly

Discussion
27
weakened the structural integrity of theeelgrass and the Phragmites leaves, andaccounted for the solar radiation-inducedleaching of leaves (V; Denward & Tranvik1998). The solar radiation induced leachingtransforms vascular plant detritus into adissolved form and makes it available forplanktonic bacteria (V; Denward &Tranvik 1998). Our data and those ofNeely & Wetzel (1997) and Denward &Tranvik (1998) suggest that photochemicaldecomposition of lignin may mediate thesolar radiation enhanced mass loss indetrital vascular plants.
8.4. Photodegradation of DOM andbioavailability of photoproducts
The results of this study show that solarradiation transformed the DOM of lakewater into biologically labile form andincreased thereby the microbialmineralization of the DOC and bacterialgrowth (Figs. 5-6 in IV). The methodsemployed in this study (IV) includedirradiation of <0.2 µm filtered water,subsequent spiking of the filtered waterwith the lake water bacterial communityand the determination of bacterial growthas biovolume and cell numbers during theculturing. These methods have beenapplied in other studies where the bacterialgrowth was measured also as theincorporation of radiolabelled thymidine orleucine. Irradiation of DOM was found toaffect bacterial growth minimally (Reche etal. 1998, Obernosterer et al. 1999),positively (Strome & Miller 1978, Geller1986, Lindell et al. 1995, Bushaw et al.1996, Miller & Moran 1997, Bano et al.1998, Obernosterer et al. 1999) ornegatively (Naganuma et al. 1996, Benner& Biddanda 1998, Tranvik & Kokalj 1998,Obernosteres et al. 1999). All thesecontrasting aspects of solar radiation onbacterial growth were observed in the workof Obernosterer et al. 1999. Solar radiationdid not affect the incorporation of [3H]-leucine in Caribbean water (Ober-nosterer
et al. 1999) whereas in the Straits ofGibraltar and in the Aegean Sea solarradiation increased the incorporation of[3H]-thymidine in the mesopelagic water(the depth of 200-350 m, possiblydominated by recalcitrant DOM). Theopposite effect was observed when thewater was amended with a labile substrate(protein). At the same geographic areas,solar radiation was found to decrease theincorporation of [3H]-thymidine in surfacewater during the chlorophyll maximumwhen labile algae-derived DOM possiblydominated the bioavailable substrates forbacteria (Obernosterer et al. 1999). Otherstudies have found out that solar radiationtransformed DOM into recalcitrant formwhen biologically labile substrates werepresent during irradiation (protein,Naganuma et al. 1996; or algae-derivedDOM, Benner & Biddanda 1998, Tranvik& Kokalj 1998). Irradiation produced labilesubstrates from DOM dominated by humicsubstances (IV, Strome & Miller 1978,Geller 1986, Lindell et al. 1995, Bushaw etal. 1996, Miller & Moran 1997, Bano et al.1998). Thus, the impact of solar radiationon the bioavailability of DOM may dependon the biological lability of irradiatedDOM. The results obtained from theFinnish lake and some, but not all othergeographic areas suggest that in humicwaters solar radiation mainly transformsDOM into a more bioavailable form (IV,Strome & Miller 1978, Geller 1986, Lindellet al. 1995, Bushaw et al. 1996, Miller &Moran 1997, Bano et al. 1998).
Our results showed that in the surface (0-1m) water of Lake Valkea-Kotinen, thetransformation of DOM by solar radiationresulted in a two-fold increase inrespiration compared to the biomassproduction, while no such effect wasobserved in the non-exposed DOM (seeFigs. 4 and 6 in IV). These results suggestthat the growth efficiency of bacteriagrown in photochemically transformedhumic DOM was 50% lower than of those

Discussion
28
grown in the unmodified DOM. Miller &Moran (1997) and Bano et al. (1998)assumed that the bacterial growthefficiency for the irradiated and the non-irradiated waters in a salt marsh and inOkefenokee Swamp, Georgia USA werethe same when they calculated microbialmineralization of DOC by bacterialsecondary production. These calculationsmay, however, have been biased due topossible differences in the bacterial growthefficiencies in irradiated and non-irradiatedwaters.
8.5. Photochemical release of nitrogen andphosphorus
Although photochemical reactions canliberate nutrients (nitrogen and phos-phorus) from organic compounds and fromcolloids (Cotner & Heath 1990, Bushaw etal. 1996), this has not been observed in allstudies (Fig. 3 in IV, Bertilsson et al.1999), or at least the amount of phos-phorus released was low (Fig. 2 in IV). Theso far identified organic photo-products aredevoid of nitrogen and phosphorus (Fig.4). The C:N:P ratio found in the DOM ofLake Valkea-Kotinen was far from thatfound in the microbial biomass. It can bededuced that if the photochemicallytransformed DOM has the same C:N:Pratio as the initial DOM, it can serve mainlyas a source for carbon, though not fornitrogen or phosphorus, for the bacteria. Inagreement with this interpretation, thephotochemical produc-tion of organiccompounds devoid of nitrogen andphosphorus, sometimes exceeded 10-foldthe secondary production of bacteria in thesurface water of lakes Fiolen andSkärhultsjön (Bertilsson & Tranvik 1998).Without nutrient amendment, the bacterialproduction in the <GF/F-filtered water ofthe humic Old Man McMullen Pond wassimilar in the non-exposed water to that insolar radiation exposed water for 32 to 196h (or 27 to 279 mol of photosyntheticallyactive radiation, Reche et al. 1998). In the
presence of inorganic phosphorus andnitrogen, bacterial production wasconsistently higher in irradiated than innon-exposed water (Reche et al. 1998).Our observation that the bacterialproduction did not always increase in theirradiated water of Lake Valkea-Kotinen(Fig. 4 in IV) is in agreement with theresults of Reche et al. (1998). However thebacterial respiration was consistently higherin the irradiated than in the non-irradiatedwater (Fig. 5-6 in IV). The results from theFinnish lake and those of obtainedelsewhere suggest that solar radiation leadsto a differential increase in respirationcompared to that of biomass production inhumic waters with low concentration of Nand P (IV; Reche 1998, Bertilsson &Tranvik 1999 and Bertilsson et al. 1999).
8.6. Photodegradation of structural lignin:implications for bacteria
In the water of Lake Valkea-Kotinen, solarradiation increased microbial activity (Figs.2-6 in IV). This was possibly caused by theliberated nutrients and labile substrates formicrobes (Cotner & Heath 1990, Bushawet al. 1996, see Fig. 4 and references therefor labile substrates). The nutrient effectscan not explain the increased microbialactivity on leaves of the eelgrass, becausethe leaves were transferred in the sameautoclaved <0.2 µm filtered water ofRoskilde Fjord prior to the measurement ofmicrobial activity. Photochemicaldegradation of lignin may, however, haveenhanced the activity of microbes on theleaves (Opshal & Benner 1993). Thestructural integrity of the leavespresumably collapsed upon loss of ligninfacilitating the fragmentation of the leaves(Fig. 9 in V). These processes may havepromoted colonization of the leaves bybacteria and intensified the contact betweenbacterial enzymes and detritalmacromolecules.

Discussion
29
8.7. Net effect of solar radiation on thedecomposition of detritus
In this study, detritus was exposed to solarradiation in the absence of detrivores. Innatural environments solar radiation willaffect also detrivores, either negatively orin an unpredictable way (Table 1). Bacte-rioplankton of Lake Valkea-Kotinen will beexposed to solar UV-radiation only in thetop layer of 10 cm due to the highabsorptivity of the lake water (Figs. 2 and 7in II). Consequently the negative effect ofsolar radiation on bacterioplankton musthave been small in Lake Valkea-Kotinen. Inthe marine littoral 10% of the surface UV-B radiation may penetrate to the depth of1.5 m and detrivores thus may be exposedto high UV-radiation (Kuhn et al. 1999).Maybe the detrivores in such marine littoralare adapted to their environment and sufferless than expected from the adverse effectsof solar UV-radiation (Rawlings 1996, Hillet al. 1997). If this were the case, solarradiation could enhance the decompositionof DOC in a humic lake and aged vascularplant detritus in the littoral zone.

Conclusions
30
9. Conclusions
1. Planktonic micro-organisms werequantitatively the most important minera-lizers of DOC in a humic Lake Valkea-Kotinen, Finland. During the summer,microbes mineralized 45 mg C m-3
d-1 and
covered 88% of the total mineralization ofDOC in the 2 m deep epilimnion of LakeValkea-Kotinen.
2. During summer in Lake Valkea-Kotinen,the rate of solar radiation inducedphotochemical mineralization of DOC was230 mg C m-3
d-1 at the depth of 1 cm. At
that depth, the rate of photochemicalmineralization of DOC was four timeshigher than that of microbial minera-lization of DOC.
3. In two humic lakes (Valkea-Kotinen,Finland and Skervatjern, Norway) thephotochemical mineralization attenuatedsteeply with increasing depth of the watercolumn. In Valkea-Kotinen, the verticalattenuation of photochemical minera-lization was similar to the verticalattenuation of UV-A radiation, and most(90%) of the photochemical mineralizationoccurred in the top 10 cm of the watercolumn. Due to the steep attenuation, thesummer time photochemical minera-lizationcovered only 12% of the totalmineralization of DOC in the epilimnion ofLake Valkea-Kotinen.
4. The sensitivity of DOC to photo-chemical mineralization varied in differentparts of the watershed of Lake Skervatjern.The DOC flowing into the lake from thecatchment area was the most sensitive forphotochemical mineralization.
5. Solar radiation mineralized 19% of 14C-labelled aromatic rings of 14C-[ring]-labelled synthetic lignin (14C-DHP) toDI14C and increased the water solubility of14C-label. This occurred when the
14C-DHPwas mixed to the water of Lake Valkea-
Kotinen and exposed to solar radiation for7 d at the depth of 1 cm.
6. A novel method was developed fordetermining the quantum yield spectrumfor the photo-chemical mineralization of
DOC (φλ) in Lake Valkea-Kotinen in situ.
The method describes φλ as c × 10-dλ,where c (dimensionless) and d (nm-1) arepositive constants. The determination gave
a value of φλ of 7.52 × 10-0.0122λ based on(1) the absorption spectrum of CDOM, (2)the in situ measured photochemicalmineralization at the depth z and (3) thesolar spectrum of photon flux density at thesame depth. The quantum yieldsdetermined in situ agreed with thosedetermined in the laboratory at singlewavelengths.
7. Applying a φλ of 7.52 × 10-0.0122λ in themodel resulted in a satisfactory predictionof the photochemical mineralization rates atthe depths of 1-10 cm and in the wholewater column of Lake Valkea-Kotinen. Themodelling revealed that the UV-A part ofsolar radiation mineralized DOC of LakeValkea-Kotinen most.
8. The microbial mineralization of DOCwas 2 to 5 fold higher in the water of LakeValkea-Kotinen exposed to solar radiationfor a day than in the non-exposed waterkept in darkness.
9. More dissolved phosphorus and nitrogenwere converted to particulate form in thewater of Valkea-Kotinen exposed to solarradiation for a day than in the non-exposedwater kept in the dark during microbialculturing. The nutrient dynamics indicatedan increased production of microbialbiomass in the solar radiation exposedwater. This result was supported bymicroscopic data. The biovolume ofbacteria grown in the solar radiationexposed water was 1.7 times higher than of

Conclusions
31
those grown in the non-exposed waterkept in darkness.
10. Solar radiation alone decomposeddetrital leaves of eelgrass. The leaves lostpigments and chromophoric matter during30 d exposure to solar radiation. Solarradiation enhanced the loss of organicmatter from the leaves and induced theirfragmentation.
11. The incorporation of 14C-leucine by amicrobial biofilm introduced on the detritalleaves was 2-4 times higher following anexposure to solar radiation compared to abiofilm on leaves not-exposed to solarradiation. Thus, the exposure of detritalleaves to solar radiation increased theactivity of microbial biofilm on the leaves.

Acknowledgements
32
10. Acknowledgements
I dedicate this thesis to my teachers. With ateacher I do not only mean the teachers Ihave met during the lengthy period I havespent in schools and university, but also mycolleagues, friends, relatives and parents.All of you have taught me skills requiredfor preparing this thesis.
Kalevi Salonen introduced me the topic ofthe thesis. He taught me the principles oflimnology and guided me to the challengingarea of ecosystem studies. Kale’s ideas andexperimental set-ups were clever and novel.If the results of an experiment turned out tobe a disaster and I lost my faith, theoptimism of Kale never failed. If I havesucceeded to catch a bit of the optimismand of Kale’s innovative ideas, I havelearned a lot.
I have had a privilege to been supervisedalso by Mirja Salkinoja-Salonen. She gavefull support in my work and taught to methe principles of scientific work. Herpersonal investments on my scientificeducation have been great. On the basis ofthese investments I have a solid ground tocontinue my work.
Jussi Kukkonen and Alasdair H. Neilsonreviewed my thesis and gave valuablecomments, which improved the quality ofthe thesis considerably.
I enjoyed my work at the Lammi BiologicalStation. The open atmosphere amongpeople at Lammi ensured good scientificdiscussion and friendly relationships. Theheads of the station, Jaakko Syrjämäki andLauri Arvola, were always kind andunderstanding related to the progress of thiswork. I owe my warmest gratitude to themembers of the project, nutrient cycling inhumic lakes, Marko Järvinen, UweMünster, Taavi Aalto, Eeva Huitu andMarkit Likolammi. I am grateful to JormaKeskitalo for assisting in the fieldwork and
patiently delivering data related to LakeValkea-Kotinen. Jaakko Vainionpää andRiitta Ilola helped me hundreds of times inthe lab. I thank Anne Ojala for globalradiation measurements. Ilpo Hakala helpedme to use computers effectively. LeilaTuominen cheered me up during every visitat Lammi and took reliably care fromvarious practical things. The work atLammi was pleasant, because food wasdelicious, rooms were clean, technicalproblems were always solved by “the boys”,and the sauna at the shore of Lake Pääjärviwas wonderful.
My relatives at Lammi, Jaakko (†) & AnneliVähätalo, Keijo & Maija-Liisa Vähätalo,and Ulla & Antti Kartano provided meseveral opportunities for relaxation andpleasant non-scientific discussion.
I wrote my thesis in the MSS (MirjaSalkinoja-Salonen) research group. I wantto express my gratitude to the present andformer group members: Eija Saski, Kimmo& Irmgard Suominen, Rainer & JoannaPeltola, Raimo Mikkola, Ira Tsitko, TuijaPirttijärvi, Jouni Jokela, Riitta Boeck,Marko Kolari, Maria Andersson, KatriMattila, Minna Laine, Liisa Nohynek,Eszter Kostyál, Jaro Nuutinen, ChristophWittmann, Timo Nieminen, Jukka Kurola,etc. You shared with me parties as well aseveryday work including the negotiations atthe coffee and lunch table.
I was lucky to study decomposition ofeelgrass in the Freshwater BiologicalStation at Hillerød, Denmark undersupervison of Morten Søndergaard. I thankMorten for fluent and fruitful co-operation.I am grateful to Louise Schlüter and StiigMarkager for pigment and optical analysis.
I am grateful for Helge Lemmetyinen forsharing his laboratory and knowledge withme at the Technical University of Tampere.Elina Vuorimaa offered her help in thepractical work at Tampere. I thank Tomi

Acknowledgements
33
Vähätalo, Päivi Valta, Keijo & Maija-LiisaVähätalo for housing at Tampere.
I once had a dream. It was high molecularweight labelled model compoundrepresenting biologically recalcitrant humicmatter. Annele Hatakka made my dreamcome true in the form of 14C-DHP. I thankAnnele and her group members PekkaOivanen, Kari Steffen and Marja Tuomelafor 14C-DHP and valuable practicalinstructions in the use of it.
Petteri Taalas and Martti Heikinheimohelped me to understand radiative transferin atmosphere. Antti Herlevi gave valuableinformation of underwater optics. HeikkiHaario introduced me the Matlab software.Tom Frisk gave me first insight tomodelling. Vappu Pennanen explained methe contribution of iron to the colour oflake water. Sirpa Kleemola gave iron datafrom Lake Valkea-Kotinen. Timo Nieminenand Jukka Järvinen delivered outflow dataof Valkea-Kotinen. Erja Huovila explainedme the principles of Washi-making. Jari Yli-Kauhaluoma assisted me in the search ofdissociation energies. Donald Smartchecked a language of early drafts of some
manuscripts. Pirjo Tuomi almost prepared aNature-paper with me. Anna-Mari Venteläshared her expertise in image analysis.
I am grateful to Jürg Hoigné, SilvioCanonica and Barbara Sulzberger for awarm reception during my short butimportant visit in EAWAG, Switzerland. Iam grateful to Swedish colleagues, WilhelmGranéli, Måns Lindell, Måns Denward andStefan Bertilsson, for stimulating discussionand help in the preparation of papers. Ithank Maaret Kulovaara and Nina Corin forsharing their expertise in chemistry with me.
My student mates, especially those from thebiology course of 1984, were pleasantcompanions during studies as well as freetime activities. I am grateful to my friendsin the field of ornithology. Your companionprepared me to scientific criticisms andprovided unforgettable moments with birds.
I thank my parents Anne & Aatos Vähätalofor advice concerning general survival andfull support during my studies. I thank myfiancée Merja Taiminen for her love,patience and understanding during ourrelationship lasting over this study.

References
34
11. References
Aiken, G. R., D. M. McKnight, R. L. Wershaw,and P. MacCarthy (eds.) 1985. Humic substancesin soil, sediment, and water - geochemistry,isolation, and characterization. John Wiley &Sons. 692 pp.
Alexander, M. 1985. Biodegradation of organicchemicals. Environ. Sci. Technol. 18: 106-111.
Alexander, M. 1999. Biodegradation andbioremediation. Second edition. Academic Press.San Diego.
Allard, B., H. Borén, C. Pettersson, and G. Zhang.1994. Degradation of humic substances by UVirradiation. Environ. Internat. 20: 97-101.
Amador, J. A., M. Alexander, and R. Zika. 1989.Sequential photochemical and microbialdegradation of organic molecules bound to humicacid. Appl. Environ. Microbiol. 55: 2843-2849.
Anesio, A. M., C. M. Denward, L. J. Tranvik, andW. Granéli. 1999. Decreased bacterial growth onvascular plant detritus due to photochemicalmodification. Aquat. Microb. Ecol. 17: 159-165.
APHA. 1998. Standard methods for theexamination of water and wastewater. 20th edition.American Public Health Association. WashingtonDC.
Argyropoulos D. S., and Y. Sun. 1996.Photochemically induced solid-state degradation,condensation, and rearrangement reactions inlignin model compounds and milled wood lignin.Photochem. Photobiol. 64: 510-517.
Arrigo, K. R. 1994. The impact of ozone depletionon phytoplankton growth in the Southern Ocean:large-scale spatial and temporal variability. Mar.Ecol. Prog. Ser. 114: 1-12.
Audic, A., and P. U. Giacomoni. 1993. DNAnicking by ultraviolet radiation is enhanced in thepresence of iron and of oxygen. Photochem.Photobiol. 57: 508-512.
Backlund, P. 1992. Degradation of aquatic humicmaterial by ultraviolet light. Chemosphere 25:1869-1878.
Bano, N., M. A. Moran, and R. E. Hodson. 1998.Photochemical formation of labile organic matter
from two components of dissolved organic carbonin a freshwater. Aquat. Microb. Ecol. 16: 95-102.
Barnabas, A. D. 1992. Bacteria on and within leafblade epidermal cells of the seagrassThalassodendron ciliatum (Forssk.) Den Hartog.Aquat. Bot. 43: 257-266.
Becker, H. G. O., H. Böttcher, F. Dietz, A. V.El`cov, V. M. Grebenkina, A. Hantcshmann, H.Haufe, G. Roewer, K. Schiller, O. P. Studzinskij,P. Thomas, and H.-J. Timpe. 1976. Einführung indie Photochemie. Deutscher Verlag derWissenschaften, Berlin. p. 182-195.
Benner, R., and B. Biddanda. 1998. Photochemicaltransformation of surface and deep marinedissolved organic matter: Effects on bacterialgrowth. Limnol. Oceanogr. 43: 1373-1378.
Bergström, I., A. Heinänen, and K. Salonen. 1986.Comparison of acridine orange, acriflavine andbisbenzimide stains for enumeration of bacteria inclear and humic waters. Appl. Environ. Microbiol.51: 664-447.
Bertilsson, S., and Allard, B. 1996. Sequentialphotochemical and microbial degradation ofrefractory dissolved organic matter in a humicfreshwater system. Arch. Hydrobiol./Advanc.Limnol. 48: 133-141.
Bertilsson, S., and L. Tranvik. 1998.Photochemically produced carboxylic acids assubstrates for freshwater bacterioplankton. Limnol.Oceanogr. 43: 885-895.
Bertilsson, S., R. Stepanauskas, R. Cuadros-Hansson, W. Granéli, J. Wikner, and L. Tranvik.1999. Photochemically induced changes inbioavailable carbon and nitrogen pools in a borealwatershed. Aquat. Microb. Ecol. 19: 47-56.
Blackburn, N., F. Azam, and Å. Hagström. 1997.Spatially explicit simulations of a microbial foodweb. Limnol. Oceanogr. 42: 613-622.
Bloom, P. R., and J. A. Leenheer. 1989.Vibrational, electronic, and high-energyspectroscopic methods for characterizing humicsubstances. In: M. H. B. Hayes, P. MacCarthy, R.L. Malcolm, and R. S. Swift (eds.) John Wiley &Sons Ltd, New York, p. 411-446.
Blumthaler, M., W. Ambach and W. Rehwald.1992. Solar UV-A and UV-B radiation fluxes attwo Alpine stations at different altitudes. Theor.Appl. Climatol. 46: 39-44.

References
35
Boavida, M. J., and R. G. Wetzel. 1998.Inhibition of phosphatase activity by dissolvedhumic substances and hydrolytic reactivation bynatural ultraviolet light. Freshw. Biol. 40: 285-293.
Bobrowski, K., B. Marciniak, and G. L. Hug.1992. 4-carboxybenzophenone-sensitizedphotooxidation of sulfur-containing amino acids.Nanosecond laser flash photolysis and pulseradiolysis studies. J. Am. Chem. Soc. 114: 10279-10288.
Bosma, T. N. P., P. J. M. Middeldorp, G. Schraa,and A. J. B. Zehnder. 1997. Mass transferlimitation of biotransformation: Quantifyingbioavailability. Environ. Sci. Techol. 31: 248-252.
Bothwell, M. L., D. M. J. Sherbot, and C. M.Pollock. 1994. Exosystem response to solarultraviolet-B radiation: Influence of trophic-levelinteractions. Science 265: 97-100.
Brennan, T. M. 1996. Decarboxylation of indole-3-acetic acid and inhibition of growth in Avenastativa seedlings by plant-derived photosensitizers.Photochem. Photobiol. 64: 1001-1006.
Bricaud, A., A. Morel, and L. Prieur. 1981.Absorption by dissolved organic matter of the sea(yellow substance) in the UV and visible domains.Limnol. Oceanogr. 26: 43-53.
Brock, T. D., M. T. Madigan, J. M. Martinko, andJ. Parker. 1994. Biology of microorganisms.Seventh edition. Prentice-Hall InternationalEditions. London.
Bushaw, K. L., R. G. Zepp, M. A. Tarr, D.Schultz-Jander, R. A. Bourbonniere, R. E. Hodson,W. L. Miller, D. A. Bronk and M. A. Moran.1996. Photochemical release of biologicallyavailable nitrogen from aquatic dissolved organicmatter. Nature 381: 404-407.
Castellan A., C. Vanucci, and H. Bouas-Laurent.1987. Photochemical degradation of lignin throughβC-O bond cleavage of non phenolic benzyl arylether units. A study of the photochemistry ofβ(2',4',6'-trimethyl-phenoxy)-3,4 dimethoxytoluene. Holzforschung 41: 231-238.
Chen, Y., S. U. Khan, and M. Schnitzer. 1978.Ultraviolet irradiation of dilute fulvic acidsolutions. Soil Sci. Soc. Am. J. 42: 292-296.
Chen, W., and P. J. Wangersky. 1996. Productionof dissolved organic carbon in phytoplanktoncultures as measured by high-temperature catalyticoxidation and ultraviolet photo-oxidation methods.J. Plankton Res. 18: 1201-1211.
Choswell, R. K., G. R. Shawn, G. Eaglesham, M.J. Smith, R. L. Norris, A. A. Seawright, and M. R.Moore. 1999. Stability of cylindrospermopsin, thetoxin from the cyanobacterium, Cylindrosperm-opsis raciborskii: Effect of pH, temperature, andsunlight on decomposition. Environ. Toxicol. 14:155-161.
Cooper, W. J., R. G. Zika, R. G. Petasne and A. M.Fischer. 1989. Sunlight-induced photo-chemistryof humic substances in natural waters: majorreactive species. In: I. H. Suffet and P. MacCarthy(eds.). Influence of aquatic humic substances onfate and treatment of pollutants. AmericanChemical Society Symposium Series 219.Washington, D.C. p. 333-362.
Corin, N., P. Backlund, and M. Kulovaara. 1996.Degradation products formed during UV-irradiation of humic waters. Chemosphere 33: 245-255.
Cotner, J. B. Jr., and R. T. Heath. 1990. Iron redoxeffects on photosensitive phosphorus release fromdissolved humic materials. Limnol. Oceanogr. 35:1175-1181.
Dabestani, R., R. H. Sik, D. G. Davis, G. Dubay,and C. F. Chignell. 1993. Spectroscopic studies ofcutaneous photosensitizing agents. XVIII.Indometacin. Photobiol. Photochem. 58: 367-373.
Davies-Colley, R. J. and W. N. Vant. 1987.Absorption of light by yellow substance infreshwater lakes. Limnol. Oceanogr. 32: 416-425.
Dean, J. A. 1992. Lange's Handbook of Chemistry(14th edition). McGraw Hill, New York. p. 4.25.
De Haan, H. 1993. Solar UV-light penetration andphotodegradation of humic substances in peatylake water. Limnol. Oceanogr. 38: 1072-1076.
Del Giorgio, P. A., J. J. Cole, and A. Cimbleris.1997. Respiration rates in bacteria exceedphytoplankton production in unproductive aquaticsystems. Nature 385: 148-151.
Denwar, C. M. T., and L. J. Tranvik. 1998. Effectsof solar radiation on aquatic macrophyte litterdecomposition. Oikos 82: 51-58.

References
36
Donahue, W. F., D. W. Schindler, S. J. Page, andM. P. Stainton. 1998. Acid-induced changes in
DOC quality in an experimental whole-lakemanipulation. Environ. Sci. Technol. 32: 2954-2960.
Elser, J. J., L. B. Stabler, and R. P. Hassett. 1995.Nutrient limitation of bacterial growth and rates ofbacteriovory in lakes and oceans: a comparativestudy. Aquat. Microb. Ecol. 9: 105-110.
Enríques, S., C. M. Duarte, and K. Sand-Jensen.1993. Patterns in decomposition rates amongphytosynthetic organisms: the importance ofdetritus C:N:P content. Oecologia 94: 457-471.
Eriksson K.-E. L., R. A. Blanchette, and P. Ander.1990. Microbial and enzymatic degradation ofwood and wood components. Springer Verlag,Berlin, Heidelberg. p. 232.
Faix O., M. D. Mozuch, and T. K. Kirk. 1985.Degradation of gymnosperm (guaiacyl) vs.angiosperm (syringyl/guaiacyl) lignins byPhanerochaete chrysosporium. Holzforschung 39:203-208.
Fasham, M. J. R., P. W. Boyd, and G. Savidge.1999. Modeling the relative contributions ofautotrophs and heterotrophs to carbon flow at aLagrangian JGOFS station in the NortheastAtlantic: The importance of DOC. Limnol.Oceanogr. 44: 80-94.
Faust, B. C., and R. G. Zepp. 1993.Photochemistry of aqueous iron(III)-polycarboxylate complexes: Roles in the chemistryof atmosphere and surface waters. Environ. Sci.Technol. 27: 2517-2522.
Fenchel, T. 1970. Studies on the decomposition oforganic detritus from the turtle grass Thalassiatestudinum. Limnol. Oceanogr. 15: 14-20.
Flöder, S., and U. Sommer. 1999. Diversity inplankton communities: An experimental test of theintermediate disturbance hypothesis. Limnol.Oceanogr. 44: 1114-1119.
Frederick, J. E. and D. Lubin. 1988. The budget ofbiologically active ultraviolet radiation in the earthatmosphere system. J. Geophys. Res. 93: 3825-3832.
Frederick, J. E., H. E. Snell and E. K. Haywood.1989. Solar ultraviolet radiation at the earth'ssurface. Photochem. Photobiol. 50: 443-450.
Gao, H., and R. G. Zepp. 1998. Factors influencingphotoreactions of dissolved organic matter in acoastal river of the Southeastern United States.Environ. Sci. Technol. 32: 2940-2946.
Garcia-Pichel, F. 1994. A model for internal self-shading in planktonic organisms and itsimplications for the usefulness of ultravioletsunscreens. Limnol. Oceanogr. 39: 1704-1717.
Gehrke, C., U. Johanson, T. V. Callaghan, D.Chadwick, and C. H. Robinson. The impact ofenhanced ultraviolet-B radiation on litter qualityand decomposition processes in Vaccinium leavesfrom the subarctic. Oikos. 72: 312-222.
Geller, A. 1985. Light-induced conversion ofrefractory, high molecular weight lake waterconstituents. Schweiz. Z. Hydrol. 47: 21-26.
Geller, A. 1986. Comparison of mechanismsenhancing biodegradability of refractory lake waterconstituents. Limnol. Oceanogr. 31: 755-764.
Gjessing, E. T. 1992. The HUMEX project:experimental acidification of a catchment and itshumic lake. Environ. Int. 18: 535-543.
Gjessing, E. T., and T. Gjerdahl. 1970. Influenceof ultra-violet radiation on aquatic humus. Vatten2: 144-145.
Gold M. H., H. Kutsuki, and M. A. Morgan. 1983.Oxidative degradation of lignin by photochemicaland chemical radical generating systems.Photochem. Photobiol. 38: 647-651.
Goes, J. I., N. Handa, S. Taguchi, T. Hama, and H.Saito. 1996. Metabolism of neutralmonosaccharide constituents of storage andstructural carbohydrates in natural assemblages ofmarine phytoplankton exposed to ultravioletradiation. Limnol. Oceanogr. 41: 1478-1489.
Gordon, H. R. 1989. Can the Lambert-Beer law beapplied to the diffuse attenuation coefficient ofocean water? Limnol. Oceanogr. 34: 1389-1409.
Granéli, W., M. Lindell, and L. Tranvik. 1996.Photo-oxidative production of dissolved inorganiccarbon in lakes of different humic content. Limnol.Oceanogr. 41: 698-706.
Granéli, W., M. Lindell, B. Marcal de Faria, andF. de Assis Esteves. 1998. Photoproduction ofdissolved inorganic carbon in temperate andtropical lakes - dependence on wavelength band

References
37
and dissolved organic carbon concentration.Biogeochemistry 43: 175-195.
Haag, W. R., J. Hoigné, E. Gassman, and A. M.Braun. 1984. Singlet oxygen in surface waters -part II: Quantum yields of its production by somenatural humic materials as a function ofwavelength. Chemosphere 13: 641-650.
Haag, W. R., and T. Mill. 1990. Survey ofsunlight-produced transient reactants in surfacewaters. In: N. V. Blough, R. G. Zepp (eds.).Effects of solar ultraviolet radiation onbiogeochemical dynamics in aquatic environments.Technical Report. Woods Hole: Woods Hole MAOceanographic Institution. p. 82-88.
Harrison, P.G. 1982. Control of microbial growthand amphipod grazing by water-solublecompounds from leaves of Zostera marina. Mar.Biol. 67: 225-230.
Harrison, P.G., and K. H. Mann. 1975. Detritusformation from eelgrass (Zostera marina L.): Therelative effects of fragmentation, leaching anddecay. Limnol. Oceanogr. 20: 924-934.
Hedges, J. I. 1992. Global biogeochemical cycles:progress and problems. Mar. Chem. 29: 67-93.
Herndl, G. J., G. Müller-Niklas and J. Frick. 1993.Major role of ultraviolet-B in controllingbacterioplankton growth in the surface layer of theocean. Nature 361: 717-719.
Hessen, D. O. 1998. Food webs and carbon cyclingin humic lakes. In: D. O. Hessen, and L. J. Tranvik(eds.). Aquatic humic substances. Springer-Verlag,Berlin Heidelberg. p. 287-315.
Hessen, D. O., and E. Van Donk. 1994. Effects ofUV-radiation of humic water on primary andsecondary production. Water Air Soil Poll. 75:325-338.
Hessen, D. O., K. Nygaard, K. Salonen, and A.Vähätalo. 1994. The effects of substratestoichiometry on microbial activity and carbondegradation in humic lakes. Environ. Internat. 20:67-76.
Hill, W. R., S. M. Dimick, A. E. McNamara, andC. A. Branson. 1997. No effects of ambient UVradiation detected in periphyton and grazers.Limnol. Oceanogr. 42: 769-774.
Hoigné, J. 1990. Formulation and calibration ofenvironmental reaction kinetics; oxidations by
aqueous photooxidants as an example. In: W.Stumm (ed.). Aquatic chemical kinetics. JohnWiley & Sons, Inc. p. 43-70.
Hollibaugh, J.T., and P. S. Wong. 1992. Ethanol-extractable substrate pools and the incorporation ofthymidine, L-leucine, and other substrates bybacterioplankton. Can. J. Microbiol. 38: 605-613.
Hongve, D. 1994. Sunlight degradation of aquatichumic substances. Acta Hydrochim. Hydrobiol. 22:117-120.
Hurtubise, R. D., J. E. Havel, and E. E. Little.1998. The effects of ultraviolet-B radiation onfreshwater invertebrates: Experiments with a solarsimulator. Limnol. Oceanogr. 43: 1082-1088.
Jansson, M., P. Blomquist, A. Jonsson, and A.-K.Bergström. 1996. Nutrient limitation ofbacterioplankton, autotrophic and mixotrophicphytoplankton, and heterotrophic nanoflagellatesin Lake Örträsket. Limnol. Oceanogr. 41: 1552-1559.
Jeffrey, S. W. 1997. Preparation of chlorophyllstandards. In: Jeffrey, S. W., R. F. C. Mantoura,and S. W. Wright (eds.) Phytoplankton pigmentsin oceanography: guidelines to modern methods.Unesco Paris, p. 207-238.
Jeffrey, W. H., R. J. Plefger, P. Aas, S. Hager, R.B. Coffin, R. Von Haven, and D. L. Mitchell.1996. Diel and depth profiles of DNAphotodamage in bacterioplankton exposed toambient solar ultraviolet radiation. Mar. Ecol.Prog. Ser. 137: 283-291.
Jiang, S. C., and J. H. Paul. 1998. Significance oflysogeny in the marine environment: Studies withisolates and a model of lysogenic phageproduction. Microb. Ecol. 35: 235-243.
Johnson, B. D., and R. C. Cooke. 1980. Organicparticle and aggregate formation resulting from thedissolution of bubbles in seawater. Limnol.Oceanogr. 25: 653-661.
Kamp-Nielsen, L. 1992. Benthic-pelagic couplingof nutrient metabolism along an estuarineeutrophication gradient. Hydrobiologia 235/236:457-470.
Karametaxas, G., S. J. Hug, and B. Sulzberger.1995. Photodegradation of EDTA in the precenceof lepidocrocite. Environ. Sci. Technol. 29: 2992-3000.

References
38
Keskitalo, J., and K. Salonen. 1996. Integratedmonitoring studies of plankton in the Lake
Valkea-Kotinen in 1994 (In Finnish), Report 16.Finnish Environment Agency. Helsinki.
Keskitalo J., K. Salonen, and A.-L. Holopainen.1998. Long-term fluctuations in environmentalconditions, plankton and macrophytes in a humiclake, Valkea-Kotinen. Boreal Environ. Res. 3:251-262.
Kieber, D. J., J. McDaniel, and K. Mopper. 1989.Photochemical source of biological substrates insea water: implications for carbon cycling. Nature341: 637-639.
Kieber, R. J., X. Zhou, and K. Mopper. 1990.Formation of carbonyl compounds from UV-induced photodegradation of humic substances innatural waters: Fate of riverine carbon in the sea.Limnol. Oceanogr. 35: 1503-1515.
Kieber, R. J., L. H. Hydro, and P. J. Seaton. 1997.Photooxidation of triglycerides and fatty acids inseawater: Implication toward the formation ofmarine humic substances. Limnol. Oceanogr. 42:1454-1462.
Kirk, J. T. O. 1994. Optics of UV-B radiation innatural waters. Arch. Hydrobiol. Beih. 43: 1-16.
Kirk, T. K., and R. L. Farrell. 1987. Enzymatic"combustion": the microbial degradation of lignin.Ann. Rev. Microbiol. 41: 465-505.
Krasnovsky, A. A. Jr. 1994. Singlet molecularoxygen and primary mechanisms of photo-oxidative damage of chloroplasts. Studies based ondetection of oxygen and pigment phosphorescence.P. Roy. Soc. Edinb. 102B: 219-235.
Krogh, E. and P. Wan. 1992.Photodecarboxylation of diarylacetic acids inaqueous solution: enhanced photogeneration ofcyclically conjugated eight π electron carbanions.J. Am. Chem. Soc. 144: 705-712.
Kuhn, P., H. Brownman, B. McArthur, and J.-F.St-Pierre. 1999. Penetration of ultraviolet radiationin the waters of the estuary and Gulf of St.Lawrence. Limnol. Oceanogr. 44: 710-716.
Kulovaara, M. 1996. Light-induced degradation ofaquatic humic substances by simulated sunlight.Intern. J. Environ. Anal. Chem. 62: 85-95.
Kulovaara, M., N. Corin, P. Backlund, and J.Tervo. 1996. Impact of UV254-radiation on aquatichumic substances. Chemosphere. 33: 783-790.
Kühl, M., C. Lassen, and N. P. Revsbech. 1997. Asimple light meter measurements of PAR (400 to700 nm) with fiber-optic microprobes: applicationfor P vs E0(PAR) measurements in a microbialmat. Aquat. Microb. Ecol. 13: 197-207.
Langford, C. H., and J. H. Carey. 1987.Photocatalysis by inorganic components of naturalwater systems. In: R.G. Zika, and W. J. Cooper(eds.) Photochemistry of environmental aquaticsystems. American Chemical Society, WashingtonDC, p. 225-239.
Larson, R. A., and J. H. Hufnal. 1980. Oxidativepolymerization of dissolved phenols by soluble andinsoluble inorganic species. Limnol. Oceanogr. 25:505-512.
Leibold, M. A. 1996. A graphical model ofkeystone predators in food webs: Trophicregulation of abundance, incidence, and diversitypatterns in communities. Am. Nat. 147: 784-812.
Li-Cor. 1982. Radiation measurements andinstrumentation. Lic-Cor. Inc. Lincoln, Nebraska,USA.
Lindell, M. J., W. Granéli, and L. J. Tranvik.1995. Enhanced bacterial growth in response tophotochemical transformation of dissolved organicmatter. Limnol. Oceanogr. 40: 195-199.
Liu, J., J. M. Rutz, J. B. Felix, and P. E. Klebba.1993. Permeability properties of a large gatedchannel within the ferric enterobactin receptor,FepA. P. Natl. Acad. Sci. U.S.A. 90: 10653-10657.
Lund, V., and D. Hongve. 1994. Ultravioletirradiated water containing humic substancesinhibits bacterial metabolism. Wat. Res. 28: 1111-1116.
Madronich, S., R. L. McKenzie, M. M. Caldwelland L. O. Björn. 1995. Changes in ultravioletradiation reaching the earth's surface. Ambio 24:143-152.
Mann, K. H. 1988. Production and use of detritusin various freshwater, estuarine, and coastalmarine ecosystems. Limnol. Oceanogr. 33: 910-930.
Manny, B. A., M. C. Miller, and R. G. Wetzel.1971. Ultraviolet combustion of dissolved organic

References
39
nitrogen compounds in lake waters. Limnol.Oceanogr. 16: 71-85.
Mateo, M.A., and J. Romero. 1996. Evaluatingseagrass leaf litter decomposition: an experimentalcomparison between litter-bag and oxygen-uptakemethods. J. Exp. Mar. Biol. Ecol. 202: 97-106.
McKnight, D. M., and G. R. Aiken. 1998. Sourcesand age of aquatic humus. In: D. O. Hessen, and L.J. Tranvik (eds.): Aquatic Humic Substances,Springer-Verlag, Berlin Heidelberg. p. 9-39.
Mialocq, J. C. 1997. Fundamentals of interactionbetween light and matter. In: M. Chanon (ed.)Homogenous Photocatalysis. John Wiley & SonsLtd, p. 16-51.
Middelboe, M., N. O. G. Jørgensen, and N. Kroer.1996. Effect of viruses on nutrient turnover andgrowth efficiency of noninfected marinebacterioplankton. Appl. Environ. Microbiol. 62:1991-1997.
Mill, T. 1980. Chemical and photo oxidation. In:O. Hutzinger (ed.). The handbook ofenvironmental chemistry. Vol. 2, Part A, p. 77-104.
Miller, F. A. 1953. Applications of infrared andultraviolet spectra to organic chemistry. In: H.Gilman (ed.) Organic chemistry Vol III. JohnWiley Inc., New York.
Miller, W. L. 1998. Effects of UV radiation onaquatic humus: photochemical principles andexperimental considerations In: D. O. Hessen, andL. J. Tranvik. (eds.). Aquatic humic substances.Springer-Verlag. p. 125-143.
Miller, W. L., and R. G. Zepp. 1995.Photochemical production of dissolved inorganiccarbon from terrestrial organic matter:Significance to the oceanic organic carbon cycle.Geophys. Res. Lett. 22: 417-420.
Miller, W. L., and M. A. Moran. 1997. Interactionof photochemical and microbial processes in thedegradation of refractory dissolved organic matterfrom a coastal marine environment. Limnol.Oceanogr. 42: 1317-1324.
Miller, R. V., W. Jeffrey, D. Michell, and M.Elasri. 1999. Bacterial responses to ultravioletlight. ASM News 65: 535-541.
Molina, L. T., and M. J. Molina. 1986. Absoluteabsorption cross sections of ozone in the 185- to
350-nm wavelength range. J. Geophys. Res. 91:14501-14508.
Moore, C. A., C. T. Farmer, and R. G. Zika. 1993.Influence of the Orinoco River on hydrogenperoxide distribution and production in the easternCaribbean. J. Geophys. Res. 98: 2289-2298.
Mopper, K., X. Zhou, R. J. Kieber, D. J. Kieber, R.J. Sikorski, and R. D. Jones. 1991. Photochemicaldegradation of dissolved organic carbon and itsimpact on the oceanic carbon cycle. Nature 353:60-62.
Moran, M. A., and R. G. Zepp. 1997. Role ofphotoreactions in the formation of biologicallylabile compounds from dissolved organic matter.Limnol. Oceanogr. 42: 1307-1316.
Morita, R. Y. 1997. Bacteria in oligotrophicenvironments. Chapman & Hall. New York.
Mostajir, B., S. Demers, S. de Mora, C. Belzile, J.-P. Chanut, M. Gosselin, S. Roy, P. Z. Villegas, J.Fauchot, and J. Bouchard, D. Bird, P. Monfort,and M. Levasseur. 1999. Experimental test of theeffects of ultraviolet-B radiation in a planktoniccommunity. Limnol. Oceanogr. 44: 586-596.
Müller-Niklas, G., A. Heissenberger, S. Puškaric,and G. J. Herndl. 1995. Ultraviolet-B radiation andbacterial metabolism in coastal waters. Aquat.Microb. Ecol. 9: 111-116.
Münster, U. 1993. Concentrations and fluxes oforganic carbon substrates in the aquaticenvironment. Antonie Leeuwenhoek 63: 243-274.
Münster, U., E. Heikkinen, K. Salonen, and H. DeHaan. 1998. Tracing of peroxidase activity inhumic lake water. Acta Hydrochim. Hydrobiol. 26:158-166.
Münster, U., and H. De Haan. 1998. The role ofmicrobial extracellular enzymes in thetransformation of dissolved organic matter inhumic waters. In: D. O. Hessen, and L. J. Tranvik(eds.). Aquatic humic substances. Springer-Verlag,Berlin Heidelberg. p. 199-257.
Murray, R. E., and R. E. Hodson. 1986. Influenceof macrophyte decomposition on growth rate andcommunity structure of Okefenokee swampbacterioplankton. Appl. Environ. Microbiol. 51:293-301.
Naganuma, T., S. Konishi, T. Inoue, T. Nakane,and S. Sukizaki. 1996. Photodegradation or

References
40
photoalteration? Microbial assay of the effect ofUV-B on dissolved organic matter. Mar. Ecol.
Prog. Ser. 135: 309-310.
Neely, R. K., and R. G. Wetzel. 1997. Autumnalproduction by bacteria and autotrophs attached toTypha latifolia L. detritus. J. Freshwat. Ecol. 12:253-267.
Neilson, A. H. 1994. Organic chemicals in theaquatic environment. Distribution, persistence andtoxicity. Lewis Publishers, CRC Press, Boca Raton,U.S.A.
Newell, S. Y. 1996. Established and potentialimpacts of eukaryotic mycelial decomposers inmarine/terrestrial ecotones. J. Exp. Mar. Biol.Ecol. 200: 187-206.
Obernosterer, I., B. Reitner, and G. J. Herndl.1999. Contrasting effects of solar radiation ondissolved organic matter and its bioavailability tomarine bacterioplankton. Limnol. Oceanogr. 44:1645-1654.
Odum, E. P. 1971. Fundamentals of ecology, 3rdedition. W.B. Saunders Company, Philadelphia. p.10.
Opsahl, S., and R. Benner. 1993. Decompositionof senescent blades of the seagrass Halodulewrightii in a subtropical lagoon. Mar. Ecol. Prog.Ser. 94: 191-205.
Opsahl, S., and R. Benner. 1998. Photochemicalreactivity of dissolved lignin in river and oceanwaters. Limnol. Oceanogr. 43: 1297-1304.
Otsuki, A., and R. G. Wetzel. 1973. Interaction ofyellow organic acids with calcium carbonate infreshwater. Limnol. Oceanogr. 18: 490-493.
Page, W. J., G. Huyer, M. Muyer, and E. A.Worobec. 1989. Characterization of the porins ofCampylobacter jejuni and Campylobacter coli andimplications for antibiotic susceptibility.Antimicrobial Agents and Chemotherapy 33: 297-303.
Pakulski, J. D., P. Aas, W. Jeffrey, M. Lyons, L.Von Waasenberger, D. Mitchell, and R. Coffin.1998. Influence of light on bacterioplank-tonproduction and respiration in a subtropical coralreef. Aquat. Microb. Ecol. 14: 137-148.
Peduzzi, P. and G. J. Herndl. 1991. Decompositionand significance of seagrass leaf litter (Cymodoceanodosa) for the microbial food web in coastal
waters (Gulf of Trieste, Northern Adriatic Sea).Mar. Ecol. Prog. Ser. 71: 163-174.
Pellikaan, G.C. 1982. Decomposition processes ofeelgrass, Zostera marina L. Hydrobiol. Bull. 16:83-92.
Pellikaan, G. C. 1984. Laboratory experiments oneelgrass (Zostera marina L.) decomposition. Neth.J. Sea Res. 18: 360-383.
Petersen, R. C. Jr. 1986. In situ particle generationin a southern Swedish stream. Limnol. Oceanogr.31: 432-473.
Peuravuori, J., and K. Pihlaja. 1999.Characterization of aquatic humic substances. In:J. Keskitalo, and P. Eloranta (eds.): Limnology ofhumic waters. Backhuys Publishers, Leiden, TheNetherlands, p. 11-39.
Polis, G. A., and D. R. Strong. 1996. Food webcomplexicity and community dynamics. Am. Nat.147: 813-846.
Porter, D., S. Y. Newell, W. L. Lingle. 1989.Tunneling bacteria in decaying leaves of aseagrass. Aquat. Bot. 35: 395-401.
Raghukumar, C., T. M. D´souza, R. G. Thorn, andC. A. Reddy. 1999. Lignin-modifying enzymes ofFlavodon flavus, a basidiomycete isolated from acoastal marine environment. Appl. Environ.Microbiol. 65: 2103-2111.
Rask, M., A.-L. Holopainen, A. Karusalmi, R.Niinioja, J. Tammi, L. Arvola, J. Keskitalo, I.Blomqvist, S. Heinimaa, C. Karppinen, K. Salonenand J. Sarvala. 1998. An introduction to thelimnology of the Finnish integrated monitoringlakes. Boreal Environ. Res. 3: 263-274.
Ratte, M., O. Bujok, A. Spitzy, and J. Rudolph.1998. Photochemical alkene formation in seawaterfrom dissolved organic carbon: Results formlaboratory experiments. J. Geophys. Res. 103:5707-5717.
Rawlings, T. A. 1996. Shields against ultravioletradiation: an additional protective role for the eggcapsules of benthic marine gastropods. Mar. Ecol.Prog. Ser. 136: 81-95.
Reche, I., M. L. Pace, and J. J. Cole. 1998.Interactions of photobleaching and inorganicnutrients in determining bacterial growth oncolored dissolved organic carbon. Microb. Ecol.36: 270-280.

References
41
Reitner, B., G. J. Herndl, and A. Herzig. 1997.Role of ultraviolet-B radiation on photochemicaland microbial oxygen consumption in a humic-richshallow lake. Limnol. Oceangr. 42: 950-960.
Robertson, A. I., and K. H. Mann. 1980. The roleof isopods and amphipods in the initialfragmentation of eelgrass detritus in Nova Scotia,Canada. Mar. Biol. 59: 63-69.
Roof, A. A. M. 1982. Aquatic photochemistry. In:O. Hutzinger (ed.) The Handbook ofenvironmental chemistry. Vol. 2. Part B. Springer-Verlag. Berlin. p. 43-70.
Rotani, J.-F. 1998. Photodegradation ofunsaturated fatty acids in senescent cells ofphytoplankton: photoproduct structuralidentification and mechanistic aspects. J.Photochem, Photobiol. A: Chem. 114: 37-44.
Salonen. K. 1979. A versatile method for the rapidand accurate determination of carbon by hightemperature combustion. Limnol. Oceanogr. 24:177-183.
Salonen, K. 1981. Rapid and precise determinationof total inorganic carbon and some gases by hightemperature combustion. Wat. Res. 15: 403-406.
Salonen, K., K. Kononen, and L. Arvola. 1983.Respiration of plankton in two small, polyhumiclakes. Hydrobiologia 101: 65-70.
Salonen, K., and T. Hammar. 1986. On theimportance of dissolved organic matter in thenutrition of zooplankton in some lake waters.Oecologia (Berl) 8: 246-353.
Salonen, K., P. Kankaala, T. Tulonen, T. Hammar,M. James, T.-R. Metsälä, and L. Arvola. 1992.Plankton food chains of a highly humic lake II. Amesocosm experiment in summer duringdominance of heterotrophic processes.Hydrobiologia 229: 143-157.
Saski, E. K., A. Vähätalo, K. Salonen, and M. S.Salkinoja-Salonen. 1996. Mesocosm simulation onsediment formation induced by biologically treatedbleach kraft pulp mill wastewater in freshwaterrecipients. In: M. R. Servos, K. R. Munkittrick, J.H. Carey, and B. J. van der Kraak (eds).Environmental fate and effects of pulp and papermill effluents. p. 261-270.
Schindler, D. W., and P. J. Curtis. 1997. The roleof DOC in protecting freshwaters subjected to
climatic warming and acidification from UVexposure. Biogeochemistry 36: 1-8.
Schwarzenbach, R. P., P. M. Gschwend, and D. M.Imboden 1993. Environmental organic chemistry.John Wiley & Sons, Inc.
Scully, N. M., D. J. McQueen, D. R. S. Lean, andW. J. Cooper. 1996. Hydrogen peroxide formation:The interaction of ultraviolet radiation anddissolved organic carbon in lake waters along a 43-75°N gradient. Limnol. Oceanogr. 41: 540-548.
Siffert, Ch., and B. Sulzberger. 1991. Light-induced dissolution of hematite in the presence ofoxalate: A case study. Langmuir 7: 1627-1634.
Skoog, A., B. Biddanda, and R. Benner. 1999.Bacterial utilization of dissolved glucose in theupper water column of the Gulf of Mexico.Limnol. Oceanogr. 44: 1625-1633.
Smith, R. C., and K. S. Baker. 1978. Opticalclassification of natural waters. Limnol. Oceanogr.23: 260-267.
Sommaruga, R., I. Obernosterer, G. J. Herndl, andR. Psenner. 1997. Inhibitory effect of solarradiation on thymidine and leucine incorporationby freshwater and marine bacterioplankton. Appl.Environ. Microbiol. 63: 4178-4184.
Strangeways. I. C. 1996. Back to basics: The 'met.Enclosure': Part 3 - Radiation. Weather 52: 43-49.
Strom, S. L., R. Benner, S. Ziegler, and M. J.Dagg. 1997. Planktonic grazers are a potentiallyimportant source of marine dissolved organiccarbon. Limnol. Oceanogr. 42: 1364-1374.
Strome, D. J., and M. C. Miller. 1978. Photolyticchanges in dissolved humic substances. Verh.Internat. Verein. Limnol. 20: 1248-1254.
Stumm, W. 1992. Chemistry of the solid-waterinterface. Chapter 10 - Heterogeneousphotochemistry. John Wiley & Sons, Inc. NewYork. p. 337-367.
Sun Y.-P., K. L. Nguyen, and A. F. A. Wallis.1998. Ring-opened products from reaction oflignin model compounds with UV-assistedperoxide. Holzforschung 52: 61-66.
Sunda, W. G., and D. J. Kieber. 1994. Oxidationof humic substances by manganese oxides yieldslow-molecular-weight organic substrates. Nature367: 62-64.

References
42
Søndergaard, M., and M. Middelboe. 1993.Measurements of particulate organic carbon: a noteon the use of glass fiber (GF/F) and Anodisc®filters. Arch Hydrobiol 127: 73-85.
Tranvik, L., and S. Kokalj. 1998. Decreasedbiodegradability of algal DOC due to interactiveeffects of UV radiation and humic matter. Aquat.Microb. Ecol. 14: 301-307.
Tong, Y., and B. Lighthart. 1997. Solar radiationis shown to select for pigmented bacteria in theambient outdoor atmosphere. Photochem.Photobiol. 65: 103-106.
Tulonen, T. 1993. Bacterial production in amesohumic lake estimated from [14C]leucineincorporation rate. Microb. Ecol. 26: 201-217.
Valentine, R. L., and R. G. Zepp. 1993. Formationof carbon monoxide from the photodegradation ofterrestrial dissolved organic carbon in naturalwaters. Environ. Sci. Technol. 27: 409-412.
Vaughan, P. P., and N. V. Blough. 1998.Photochemical formation of hydroxyl radical byconstituents of natural waters. Environ. Sci.Technol. 32: 2947-2953.
Vetter, Y. A., J. W. Deming, P. A. Jumars, and B.B. Krieger-Brockett. 1998. A predictive model ofbacterial foraging by means of freely releasedextracellular enzymes. Microb. Ecol. 36: 75-92.
Voelker, B. M., F. M. M. Morel, and B.Sulzberger. 1997. Iron redox cycling in surfacewaters: Effect of humic substances and light.Environ. Sci. Technol. 31: 1004-1011.
Vrede, K., T. Vrede, A. Isaksson, and A. Karlsson.1999. Effects of nutrients (phosphorus, nitrogen,and carbon) and zooplankton on bacterioplanktonand phytoplankton - a seasonal study. Limnol.Oceanogr. 44: 1616-1624.
Vähätalo, A., M. Søndergaard, L. Schlüter, and S.Markager. 1998. The impact of solar radiation onthe decomposition of detrital leaves of eelgrass(Zostera marina). Mar. Ecol. Prog. Ser. 170: 107-117.
Vähätalo, A. V., M. Salkinoja-Salonen, P. Taalas,and K. Salonen. 2000. Spectrum of the quantumyield for photochemical mineralization ofdissolved organic carbon in a humic lake. Limnol.Oceanog. (in the press).
Weir, N. A., J. Arct, A. Ceccarelli, and A. Siren.1994. Some photo-reactions of substitutedpoly(acrylophenones). Eur. Polym. J. 30: 701-706.
Weir, N. A., J. Arct, A. Ceccarelli, and D.Puumala. 1996. Long-wave photochemistry of di-methoxylated compounds related to lignin. Polym.Degrad. Stab. 52: 119-129.
Weiss, P. S., S. S. Andrews, J. E. Johnson, and O.C. Zafiriou. 1995. Photoproduction of carbonylsulfide in south Pacific Ocean waters as a functionof irradiation wavelength. Geophys. Res. Lett. 22:215-218.
Wetzel, R. G., P. R. Rich, M. C. Miller, and H. L.Allen. 1972. Metabolism of dissolved andparticulate detrital carbon in a temperate hard-water lake. Mem. Ist. Ital. Idrobiol. 29: 185-243.
Wetzel, R. G. 1984. Detrital dissolved andparticulate organic carbon functions in aquaticecosystems. Bull. Mar. Sci. 35: 503-509.
Wetzel, R. G., P. G. Hatcher and T. S. Bianchi.1995. Natural photolysis by ultraviolet irradianceof recalcitrant dissolved organic matter to simplesubstrates for rapid bacterial metabolism. Limnol.Oceanogr. 40: 1369-1380.
Weinbauer, M. G., S. W. Wilhelm, C. A. Suttle, R.J. Pledger, and D. L. Mitchell. 1999. Sunlight-induced DNA damage and resistance in naturalviral communities. Aquat. Microb. Ecol. 17: 111-120.
Wickhamn S., and M. Carstens. 1998. Effects ofultraviolet-B radiation on two arctic microbial foodwebs. Aquat. Microb. Ecol. 16: 163-171.
Wiebe, W. J., W. M. Sheldon Jr., and L. R.Pomeroy. 1992. Bacterial growth in the cold:Evidence for an enhanced substrate requirement.Appl. Envrion. Mirobiol. 58: 359-364.
Viitasalo, M., M. Rosenberg, A.-S. Heiskanen, andM. Koski. 1999. Sedimentation of copepod fecalmaterial in the coastal northern Baltic Sea: Wheredid all the pellets go? Limnol. Oceanogr. 44: 1388-1399.
Williamson, G. E. 1995. What role does UV-Bradiation play in freshwater ecosystems? Limnol.Oceanogr. 40: 386-392.
Wright S. W., S. W. Jeffrey, R. F. C. Mantoura, C.A. Llewellyn, T. Bjørnland, D. Repeta, and N.Welschmeyer. 1991. Improved HPLC method for

References
43
the analysis of chlorophylls and carotenoids frommarine phytoplankton. Mar. Ecol. Prog. Ser. 77:
183-196.
Zafiriou, O. C., J. Joussot-Dubien, R. G. Zepp, andR. G. Zika. 1984. Photochemistry of naturalwaters. Environ. Sci. Technol. 18: 358-371.
Zellmer, I. D. 1995. UV-B-tolerance of alpine andarctic Daphnia. Hydrobiologia 307: 153-159.
Zepp, R. G. and D. M. Cline. 1977. Rates of directphotolysis in aquatic environment. Environ. Sci.Technol. 11: 359-366.
Zepp, R. G., P. F. Schlotzhauer, and R. M. Sink.1985. Photosensitized transformations involvingelectronic energy transfer in natural waters: Roleof humic substances. Environ. Sci. Technol. 19:74-81.
Zepp, R. G., and M. O. Andreae. 1994. Factorsaffecting the photochemical production of carbonyl
sulfide in seawater. Geophys. Res. Lett. 21: 2813-2816.
Zigman, S., J. Reddan, J. B. Schultz, and T.McDaniel. 1996. Structural and functional changesin catalase induced by near-UV radiation.Photochem. Photobiol. 63: 818-824.
Zuo, Y., and Y. Deng. 1998. The near-UVabsorption constants for nitrite ion in aqueoussolution. Chemosphere 36: 181-188.
Zweifel, U. L., N. Blackburn, and Å. Hagström.1996. Cycling of marine dissolved organic matter.I. An experimental system. Aquat. Microb. Ecol.11: 65-77.
Yan, N. D., W. Keller, N. M. Scully, D. R. S.Lean, and P. J. Dillon. 1996. Increased UV-Bpenetration in a lake owing to drought-inducedacidification. Nature 381: 141-143.