role of cd4+ foxp3+ regulatory t cells in protection ... · oxoplasmosis is caused by a protozoan...
TRANSCRIPT

Role of CD4� Foxp3� Regulatory T Cells in Protection Induced by aLive Attenuated, Replicating Type I Vaccine Strain of Toxoplasmagondii
Haroon Akbar,* Isabelle Dimier-Poisson, Nathalie Moiré
Université de Tours, UMR1282 Infectiologie et Santé Publique, Tours, France, and INRA, UMR1282 Infectiologie et Santé Publique, Nouzilly, France
Vaccination with the live attenuated Toxoplasma gondii Mic1.3KO strain induced long-lasting immunity against challenge withToxoplasma gondii type I and type II strains. The involvement of regulatory T cells (Tregs) in the protection mechanism wasinvestigated. Intraperitoneal injection of Mic1.3KO induced a weak and transient influx of CD4� Foxp3� T regulatory cells fol-lowed by recruitment/expansion of CD4� Foxp3� CD25� effector cells and control of the parasite at the site of infection. Thelocal and systemic cytokine responses associated with this recruitment of Tregs were of the TH1/Treg-like type. In contrast, in-jection of RH, the wild-type strain from which the vaccinal strain is derived, induced a low CD4� Foxp3� cell influx and uncon-trolled multiplication of the parasites at this local site, followed by death of the mice. The associated local and systemic cytokineresponses were of the TH1/TH17-like type. In addition, in vivo Treg induction in RH-infected mice with interleukin-2 (IL-2)/anti-IL-2 complexes induced control of the parasite and a TH1/Treg cytokine response similar to the response after Mic1.3KOvaccination. These results suggest that Tregs may contribute to the protective response after vaccination with Mic1.3KO.
Toxoplasmosis is caused by a protozoan parasite that infectshumans and other warm-blooded animals. Infections in hu-
mans are generally asymptomatic, although immunosuppressedpatients may exhibit severe symptoms. Similarly, primary infec-tion during pregnancy can lead to miscarriage and neonatal mal-formations. Toxoplasmosis can be transmitted to humans via in-gestion of oocysts or via the consumption of meat productscontaminated with tissue cysts (1). Effective vaccination of do-mestic livestock can therefore prevent human infection with Tox-oplasma gondii. It is possible to induce strong protection by im-munization with a live attenuated strain (2). Live attenuatedvaccine strain models are also useful for advancing the under-standing of the protective host immune response (3–8). However,the information available about the mechanism of protection in-volved after vaccination with a type I attenuated strain was ob-tained with nonreplicating, nonpersistent strains, such as strainscps-1 (3–7) and ts4 (8). A strain known as Mic1.3KO was obtainedin our laboratory by deleting the MIC1 and MIC3 genes (9). Thisstrain was derived from the highly virulent type I RH strain, and itsreduced invasion capacity in vitro was correlated with decreasedvirulence when injected into outbred Swiss OF1 mice (9). Type Istrains are characterized by the rapid dissemination of the parasiteand by a high parasite burden that results in death soon afterinfection by a single viable parasite in mice (10). High levels ofgamma interferon (IFN-�) were produced following infectionwith a type I parasite, and mice succumbed to uncontrolled para-site growth and associated inflammation (11, 12). The Mic1.3KOstrain with the MIC1 and MIC3 gene deletions showed reducedvirulence, lower levels of dissemination throughout tissues, andlower levels of IFN-� production than the parental RH strain afterinjection into mice (13).
In a model of lethal toxoplasmosis induced after oral adminis-tration of infection with a type II strain causing infection inC57BL/6 mice, overproduction of IFN-� was also responsible formortality and was correlated with a sharp decline in the percent-age of regulatory T cells (Tregs) just before death, supporting the
hypothesis of defective immunoregulation (14). Tregs are a sub-population of CD4� T cells, and their main function is to main-tain immune homeostasis and tolerance (15). They constitutivelyexpress the interleukin-2 (IL-2) receptor alpha chain (IL2R�), asurface receptor also known as CD25, and the intracellular forkhead box-p3 transcription factor (Foxp-3) marker (16). The roleof Tregs after infection with type II strains has been fully described(14, 17–21), and Tregs have been clearly implicated in the mortal-ity of C57BL/6 mice after oral infection in the lethal ileitis model(14). The collapse of Tregs is correlated with pathogenicity andoccurs only under highly pathogenic conditions since oral infec-tion of BALB/c mice with a type II strain did not induce a reduc-tion in the levels of Tregs (14). However, depletion of Tregs inthese mice resulted in morbidity associated with a high parasiteburden and increased ileal pathology compared to that in controlBALB/c mice (19), suggesting a role of Tregs in protection duringacute infection.
In the present study, we compared the involvement of Tregsafter infection with the vaccinal Mic1.3KO strain with that after
Received 19 February 2015 Returned for modification 25 March 2015Accepted 24 June 2015
Accepted manuscript posted online 29 June 2015
Citation Akbar H, Dimier-Poisson I, Moiré N. 2015. Role of CD4� Foxp3�
regulatory T cells in protection induced by a live attenuated, replicating type Ivaccine strain of Toxoplasma gondii. Infect Immun 83:3601–3611.doi:10.1128/IAI.00217-15.
Editor: J. H. Adams
Address correspondence to Nathalie Moiré, [email protected].
* Present address: Haroon Akbar, Department of Parasitology, University ofVeterinary and Animal Sciences, Lahore, Pakistan.
Supplemental material for this article may be found at http://dx.doi.org/10.1128/IAI.00217-15.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
doi:10.1128/IAI.00217-15
September 2015 Volume 83 Number 9 iai.asm.org 3601Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

infection with the parental lethal RH strain in an attempt to iden-tify their involvement in the protection induced by vaccination.We showed a small increase in the absolute CD4� Foxp3� Tregcount at the site of infection with Mic1.3KO followed by an in-crease in CD4� CD25� Foxp3� effector cells and control of theparasite. In contrast, the increase in the Treg count in RH-infectedmice was lower and the parasites were not controlled locally. Ourstudies show that CD4� Foxp3� Tregs are involved in protection,since specific expansion of these cells using IL-2/anti-IL-2 com-plexes in mice infected with RH induced a reduction in the para-site burden and a decrease in proinflammatory cytokine levels.These features were similar to those of mice infected withMic1.3KO, supporting the role of CD4� Foxp3� Tregs in the pro-tection induced by vaccination.
MATERIALS AND METHODSAnimals and parasites. Eight-week-old female Swiss OF1 and C57BL/6mice were obtained from Janvier (France). All experiments using animalswere approved by the local ethics committee (CEEA VdL) and registeredunder reference number 2011-06-6.
Two weeks before injection, RH tachyzoites harvested from the peri-toneal cavity of the mice were cultured on human foreskin fibroblast(HFF) monolayers (ATCC CRL-1634; American Type Culture Collec-tion), as previously described (13). Mic1.3KO parasites were obtained bytargeted disruption of the MIC1 and MIC3 genes in the �HX RH strain ofT. gondii, as previously described (9). Mic1.3KO tachyzoites were alsopropagated by serial passage on HFF monolayers. Tachyzoites freshlyharvested from a cell culture were counted using a Malassez countingchamber.
Mice were infected by intraperitoneal (i.p.) inoculation of 100 freshlyharvested tachyzoites diluted in 0.2 ml Dulbecco modified Eagle saline.
Cell culture conditions and cytokine quantification. For cytokinedetection, splenocytes were recovered and purified as described previ-ously (13, 21) and stimulated for 72 h with 10 �g/ml Toxoplasma extract(TE) or with purified anti-CD3 (clone 145-2C11) (eBioscience) at 1 �g/ml. The cells (5 � 105) were seeded into 24-well plates in 1 ml RPMI 1640containing 5% fetal calf serum (FCS), and supernatants were collected at24, 48, and 72 h after activation.
Peritoneal exudate cells (PECs) were obtained from uninfected mice(day 0 controls) and from infected mice at 4, 7, and 11 days postinfection(dpi) by peritoneal lavage with 5 ml of ice-cold phosphate-buffered saline(PBS), as already described (13), and centrifuged to collect the superna-tants. The supernatants were kept frozen at �20°C until assayed for cyto-kines. The cytokines (IFN-�, IL-2, IL-10, IL12p40, IL12p70, and IL-23)and chemokines (CCL2, CCL3, and CCL20) in the serum, cell culturesupernatants, and peritoneal washes were quantified by enzyme-linkedimmunosorbent assay (ELISA) using a Ready-Set-Go kit (eBioscience)and R/D Duoset kits.
Cell surface staining and intracellular staining. PECs and spleno-cytes were obtained from mice on day 0 (noninfected controls) and on 4,7, and 11 dpi. Cells were washed once in PBS and counted to determinetotal viable cell numbers by trypan blue exclusion. Standard procedureswere used to stain 2 � 105 to 5 � 105 PECs and 106 splenocytes in 5% FCSin PBS and mouse Fc block (BD Biosciences) as previously described (21).Antibodies for the detection of CD4 (GK1.5), CD8 (eBioH35-17.2), CD25(PC61), CD69 (H1.2F3), Ki-67 (SolA15), and Foxp3 (FJK-16S) were pur-chased from eBioscience.
For IL-10 detection, splenocytes were seeded into 6-well plates at106/ml in a final volume of 5 ml RPMI 1640 containing 5% FCS andstimulated for 18 h with 10 �g/ml TE. The culture medium was thenremoved and replaced by fresh culture medium containing phorbol my-ristate acetate (50 ng/ml), ionomycin (1 �g/ml), and brefeldin A (5 �g/ml) for a further 4-h period. Antibodies for the detection of IL-10 werepurchased from BD Biosciences.
Cell acquisition was undertaken with a BD FACSCalibur cytometer,and cells were analyzed using CellQuest software (BD Bioscience).
In vivo Treg expansion. IL-2/anti-IL-2 complexes were prepared asdescribed in the literature (14, 20) with minor modifications. Recombi-nant IL-2 and anti-IL-2 monoclonal antibody (MAb; clone JES6-1A12)were obtained from eBioscience. IL-2 (1 �g) was mixed with the anti-IL-2MAb (10 �g), and the mixture was incubated for 15 min at room temper-ature prior to i.p. injection at 0, 3, and 5 dpi.
Statistical analyses. Differences between the groups were comparedusing the Mann-Whitney or Kruskal-Wallis nonparametric test followedby Dunn’s posttest using GraphPad Prism software. All statistical testswere two-sided, and a P value of 0.05 was considered statistically signif-icant in all tests. Some analyses were performed with StatXact software(Cytel Studio) using nonparametric exact tests with strata to take intoaccount variability between repetitions of the same experiment.
All data in the graphs are expressed as the median plus the range(unless otherwise specified).
RESULTSLack of control of the parasite was not associated with decreasedIFN-� production in mice infected with RH. The parasite burdenin the peritoneum was monitored by direct counting on days 4, 7,and 11 after inoculation of 100 tachyzoites of both strains (Fig.1A). On day 4, few parasites (�0.02 � 106) were found in eithergroup, and the difference between the two groups was not signif-icant. However, on day 7 parasite counts were significantly in-creased in the group infected with strain RH (referred to here asthe RH group; 16 � 106) but not in the group infected with strainMic1.3KO (referred to here as the Mic1.3KO group; 0.002 � 106).In the Mic1.3KO group, the parasite counts remained very loweven at 11 dpi, and the number of parasites decreased betweendays 4 and 11. Taken together, these results suggest that after day4 the parasite was being controlled at the local level in theMic1.3KO-infected mice but not in the RH-infected mice, wherethe parasite count increased exponentially until the animals died.This high parasite count at 7 dpi was correlated with a high cellmortality rate (between 25 and 90%, as determined by trypan blueexclusion) for PECs recovered from the RH mice. Similar resultswere obtained when using inbred C57BL/6 mice, in which lowerparasite counts were found in the Mic1.3KO group than in the RHgroup (see Table S1 in the supplemental material), suggesting thatthese mice were as resistant to the infection with the vaccinal strainas outbred mice.
We also measured the serum levels of systemic IFN-� at 4, 7,and 11 dpi (Fig. 1B). In control naive mice, the level of IFN-� wasbelow the detection threshold (data not shown). The serum levelsof IFN-� rose significantly following infection. In the RH group,the level of IFN-� increased significantly between days 4 and 7 andparalleled the level of parasite multiplication. In the Mic1.3KOgroup, levels also increased between days 4 and 7 and subsequentlyremained at similar levels until day 11. Taken together, these re-sults suggest that mortality in the RH group may not have origi-nated from a lack of IFN-� production but, as for type II-infectedC57BL/6 mice, may have originated from uncontrolled IFN-�production (14) or a lack of parasite control, or both.
The total numbers of PECs and splenocytes were checked at 4,7, and 11 dpi and compared to those of naive mice in response toMic1.3KO and to RH (data not shown). Both infected groupsdisplayed a slightly higher total PEC count after infection than thecontrol naive group. This difference was significant only at day 11for the Mic1.3KO-injected group. At 4 dpi, the number of spleen
Akbar et al.
3602 iai.asm.org September 2015 Volume 83 Number 9Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

cells increased in both infected groups compared to the number inthe naive group, although the difference was significant only forthe RH group. In the Mic1.3KO group, this increase was signifi-cant at 7 and 11 dpi. At 7 dpi, there were significantly higher spleencell counts in the Mic1.3KO group than in the control and RHgroups. In the RH group, the cell counts were also significantlylower than those at 4 dpi. This suggests that a significant immuneresponse was induced on day 4 after infection in both infectedgroups.
Kinetics of CD4� Foxp3� Tregs after infection. After infec-tion with a lethal dose of a type II strain of T. gondii, the numbersand frequencies of CD4� Foxp3� Tregs have been reported to bereduced at the site of infection and systemically in C57BL/6 mice,and this decrease is correlated with high levels of IFN-� produc-tion (14). We therefore followed the CD4� Foxp3� Tregs afterinfection at local (peritoneum) and systemic (spleen) sites. Threepopulations (CD25� Foxp3�, CD25� Foxp3�, and CD25�
Foxp3�) were observed within the CD4� gated population inboth groups. The percentage of total Foxp3� cells in the perito-neum at 4 dpi was slightly but nonsignificantly higher in the RHgroup than in the naive mice (Fig. 2A). Moreover, there was nosignificant difference in the numbers (data not shown) or percent-
ages of Foxp3� cells (Fig. 2A) between the two infected groups atthis time. The total percentage of Foxp3� cells at 4 dpi was signif-icantly greater in the Mic1.3KO group than in the naive group(Fig. 2A). The kinetics of both effector (CD4� Foxp3� CD25�)and regulator (CD4� Foxp3�) cells were followed until 11 dpi inthe Mic1.3KO group (Fig. 2B and C). This analysis could not beperformed for the RH group because there were high levels of cellmortality at 7 dpi, and the permeabilization step further increasedcell mortality to 100% (data not shown). The relative number ofregulatory Foxp3� cells at day 4 was significantly higher in theMic1.3KO group than in the naive group (Fig. 2B) (25.9 � 104
versus 1.8 � 104 for the naive group). This increase was transient,since there was no significant difference between the groups at 7and 11 dpi.
Similar results were obtained using C57BL/6 mice (see Fig. S1in the supplemental material). C57BL/6 mice infected with theRH strain showed reduced percentages of Foxp3� cells at the locallevel compared to those in the mice infected with strainMic1.3KO. The percentages of Foxp3� cells in the spleens of thetwo groups were similar.
The numbers and percentages of effector cells at day 4 wereslightly and nonsignificantly higher in the Mic1.3KO group thanin the naive mice (Fig. 2C). However, the numbers of CD4�
Foxp3� CD25� cells increased exponentially between days 0 and11 (Fig. 2C) and at 7 and 11 dpi were significantly different fromthose at day 0. This high CD4� Foxp3� CD25� cell count at thelocal level was correlated with low parasite counts in Mic1.3KO-infected mice.
The total percentage (Fig. 2D) and absolute number of theFoxp3� population in the spleen (data not shown) did not differsignificantly between the two groups at 7 dpi. However, signifi-cantly higher percentages of CD25� Foxp3� cells were found inthe RH group. The ratio of the percentage of Foxp3�-expressingCD25 cells to the total percentage of Foxp3� cells was similarbetween the naive and the Mic1.3KO groups (0.61 and 0.60, re-spectively) but was significantly higher for the RH group (about0.71) than for both the naive and the Mic1.3KO groups (Fig. 2E).
We investigated the expression of phenotypic markers associ-ated with activation (CD69) and proliferation (Ki-67). Cells wereanalyzed directly ex vivo from the peritoneal cavity and spleen byflow cytometry on day 7 (Tables 1 and 2).
There was no significant difference in the percentages of CD4�
and CD8� cells in PECs between the control and the infectedgroups (Table 1). The percentage of CD69� cells in PECs wassignificantly higher in the RH group than in the naive group (19.9versus 5.8%) at 7 dpi (Table 1). The percentage of CD69� cells insplenocytes was also significantly higher in the RH group (11.8%)than in both the naive (3.7%) and the Mic1.3KO (1.9%) groups.The kinetics of the expression of CD69 on splenocytes were mon-itored in both groups up to 11 dpi (for the Mic1.3KO groupalone), and this higher level was only seen at 7 dpi in the RH group(data not shown). The level had not increased at 11 dpi in theMic1.3KO group, suggesting that CD69 expression may be corre-lated with the mortality observed in the RH group.
Ki-67 expression was used to follow the proliferation of PECs andsplenocytes after infection with both strains. Ki-67 expression wassignificantly higher in the PECs of both infected groups than in thoseof the naive control group (16.4% in the control group versus 62.8and 52.7% in the RH and Mic1.3KO groups, respectively). The dif-ference between the two infected groups was not significant. Similar
FIG 1 A lack of parasite control in the RH group is not correlated with lowlevels of IFN-� secretion. (A) Parasites were counted directly after peritoneallavage at the indicated times postinfection with 100 RH or Mic1.3KOtachyzoites. Results are expressed as the median plus range. Cumulative datafrom four different experiments are shown (4 to 6 mice per group per timepoint per experiment). (B) Sera were recovered at the indicated times postin-fection, and the IFN-� levels were quantified. Results are representative ofthose from three independent experiments with 5 or 6 mice per group per timepoint in which mice were injected with 100 tachyzoites of RH or Mic1.3KO. **,P 0.01 using the Kruskal-Wallis test followed by Dunn’s posttest; ***, P 0.001 using the Kruskal-Wallis test followed by Dunn’s posttest.
CD4� Foxp3� Cells and Vaccination against Toxoplasmosis
September 2015 Volume 83 Number 9 iai.asm.org 3603Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

results were obtained for splenocytes, with higher percentages of Ki-67-expressing (Ki-67�) cells being detected in both infected groups(14.0% in the control group versus 24.3 and 37.6% in the RH andMic1.3KO groups, respectively), but the difference between the con-trol and RH groups was not significant.
The expression of CD25 was monitored in the CD4� andCD8� splenocytes and compared to that in the naive group (Table 2).
Both the CD4� and CD8� populations expressed CD25 at 7 dpi.Moreover, the percentage of CD25� cells was not significantlydifferent between the naive and infected groups. However, thedifference between the two infected groups was significant for theCD4� population and was correlated with the higher level of ex-pression of CD25 on Foxp3� cells observed in the RH group.
Although the proliferation rates were similar in both groups,the RH group showed features of uncontrolled inflammation,with expression of high levels of CD69 molecules.
Induction of proinflammatory chemokines and cytokinesafter infection. To determine whether protection may be due toequilibrium between effector and regulator cells, chemokines andcytokines associated with TH1/Tregs were quantified at the locallevel (peritoneum) and in the sera of both infected groups at 7 dpi.
Chemokine levels in the peritoneal washes and in the sera werequantified at 7 dpi. In the sera, only CCL2 was above the detectionthreshold and the level was higher in the RH group (Fig. 3A). Thelevels of CCL2, CCL3, and CCL20 in the peritoneal washes weresignificantly higher in the RH group than in the Mic1.3KO group(Fig. 3B to D).
Systemic (sera) and local (peritoneum) IL-12 and IL-23 levels
FIG 2 CD4� Foxp3� Tregs were recruited/expanded before CD4� Foxp3� CD25� T effector cells at the local site after infection with Mic1.3KO but not at thesystemic level. PECs recovered at the indicated times postinfection and splenocytes recovered at 7 dpi were stained with anti-CD4 anti-CD25 and anti-Foxp3MAbs and analyzed by flow cytometry. (A) Percentage of the total Foxp3� population among CD4� cells recovered at 4 dpi. *, P 0.05 using the Kruskal-Wallistest. (B and C) For calculation of the count, the absolute number of lymphocytes was evaluated by multiplying the total PEC count by the percentage of thelymphocyte gate compared to the whole population. Tregs were defined as Foxp3� CD25� and Fopx3� CD25� cells within the CD4� population (B), and Teffector cells (T eff) were defined as Foxp3� CD25� cells within the CD4� population (C). The data presented are representative of those from two independentexperiments with 5 mice per group per injection time point. *, P 0.05 using the Kruskal-Wallis test followed by Dunn’s posttest; **, P 0.01 using theKruskal-Wallis test followed by Dunn’s posttest. (D) Percentage of the total Foxp3� and CD25� Foxp3�population within CD4� splenocytes at 7 dpi with RHor Mic1.3KO tachyzoites. Results are representative of those from four independent experiments with 4 to 6 mice per group. *, P 0.05 using the Kruskall-Wallistest followed by Dunn’s posttest. (E) Ratio of the percentage of Foxp3� cells expressing CD25 compared to the total percentage of Foxp3� cells. Results arecumulative data from 4 different experiments. *, P 0.05 using the Kruskall-Wallis test followed by Dunn’s posttest.
TABLE 1 PECs from the RH-infected group of mice presenting anactivated phenotype after infectionb
Group
% of cells that were:
CD4� CD8� CD69� Ki-67�
Naive 8.1 4.1 0.82 0.74 5.8 4.2 16.4 3.4RH 15.4 2.4 2.8 1.1 19.9 5.1a 62.8 3.5a
Mic1.3KO 16.1 3.1 3.3 2.5 11.0 3.3 52.7 7a
a The difference between the infected group and the control group using the Kruskal-Wallis test followed by Dunn’s posttest was significant (P 0.05).b PECs obtained at 7 dpi were stained with different MAbs, as indicated, and analyzedby cytometry. Results are presented as means standard deviations and arerepresentative of those from two independent experiments with 4 to 6 mice per group.
Akbar et al.
3604 iai.asm.org September 2015 Volume 83 Number 9Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

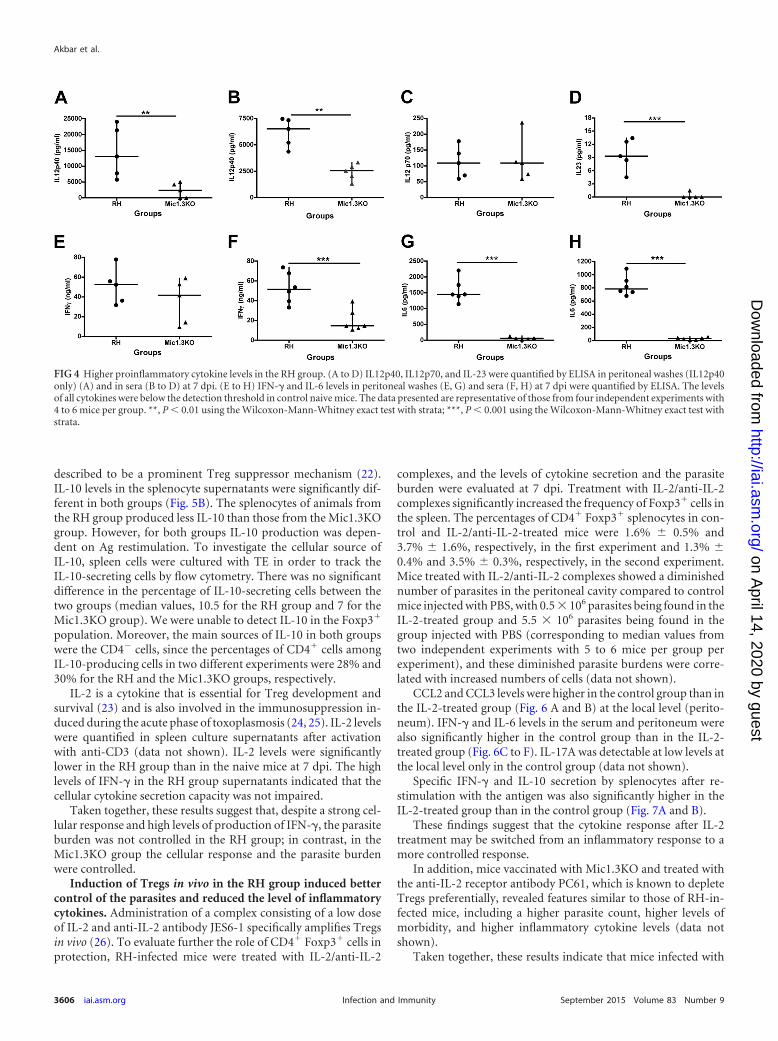
were also quantified at 7 dpi (Fig. 4). The levels of the IL12p40subunit in the peritoneal washes (Fig. 4A) and in the serum (Fig.4B) were significantly higher for the RH group. IL12p70 was un-detectable in the peritoneal washes, and the levels of the biologicalform of this cytokine were not significantly different between thetwo groups (Fig. 4C). IL-23 was detected at low levels in the sera ofRH-infected mice but not in the sera of Mic1.3KO-infected mice(Fig. 4D). The IL-23 level in the peritoneal washes was below thethreshold of detection.
As IL12p70 directs the differentiation of naive T cells in TH1cells and IL-23 is linked to TH17 differentiation and pathogeny,cytokines related to TH1 and TH17 cells were then quantified.
Serum IFN-� levels were significantly higher in the RH groupthan in the Mic1.3KO group (Fig. 4E). Slightly higher levels ofIFN-� in the peritoneal washes were observed for the RH group,but the difference from the Mic1.3KO group was not statisticallysignificant (Fig. 4F). IL-6 levels were also significantly higher inthe sera and peritoneal washes of the RH group than in those of the
Mic1.3KO group (Fig. 4G and H). IL-17A levels were at the detec-tion limit for the RH group and below the detection limit for theMic1.3KO group (data not shown).
Taken together, these results suggest that the responses of theRH group were more inflammatory, and the RH group revealed aTH1/TH17-like response, whereas the Mic1.3KO group showed amore regulated TH1/Treg-like response.
To evaluate the TH1/Treg response further, antigen (Ag)-spe-cific IFN-� production in spleen cell supernatant was measuredfollowing 48 h of restimulation with soluble Toxoplasma antigens(TE) (Fig. 5A). The IFN-� levels were below the detection thresh-old on day 4 in both groups (data not shown). At day 7, the levelswere very high in the RH group and significantly different betweenthe two groups. Cells from the RH group produced higher levels ofIFN-� than those from the Mic1.3KO group with or without TEstimulation.
IL-10 plays an important role in the suppressive function ofantigen-specific Tregs, and IL-10 production by Tregs has been
TABLE 2 Splenocytes of the RH-infected group of mice presenting an activated phenotype after infectionc
Group
% of cells that were:
CD69� CD25� CD4� CD25� CD8� CD25� Ki-67�
Naive 3.7 2.7 3.8 0.6 2.8 0.6 0.9 0.6 14.0 5.5RH 11.8 5a,b 6.5 0.8b 3.4 1.1b 1.5 0.6 24.3 3.4Mic1.3KO 1.9 1.2 2.0 0.7 1.2 0.7 0.8 0.6 37.6 4.9a
a The difference between the infected groups and the control group using the Kruskal-Wallis test was significant (P 0.05).b The difference between the two infected groups using the Kruskal-Wallis test followed by Dunn’s posttest was significant (P 0.05).c Splenocytes obtained at 7 dpi were stained with different MAbs, as indicated, and analyzed by cytometry. Results are presented as means standard deviations and arerepresentative of those from three independent experiments with 4 to 6 mice per group.
FIG 3 Higher chemokine levels in the RH group. Chemokines (CCL2, CCL3, CCL20) in sera (A) and peritoneal washes (B to D) were quantified at 7 dpi withRH or Mic1.3KO tachyzoites. The levels of all chemokines were below the detection threshold in control naive mice. The data presented are representative ofthose from five independent experiments with 5 to 6 mice per group. **, P 0.01 using the Wilcoxon-Mann-Whitney exact test with strata; ***, P 0.001 usingthe Wilcoxon-Mann-Whitney exact test with strata.
CD4� Foxp3� Cells and Vaccination against Toxoplasmosis
September 2015 Volume 83 Number 9 iai.asm.org 3605Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from
described to be a prominent Treg suppressor mechanism (22).IL-10 levels in the splenocyte supernatants were significantly dif-ferent in both groups (Fig. 5B). The splenocytes of animals fromthe RH group produced less IL-10 than those from the Mic1.3KOgroup. However, for both groups IL-10 production was depen-dent on Ag restimulation. To investigate the cellular source ofIL-10, spleen cells were cultured with TE in order to track theIL-10-secreting cells by flow cytometry. There was no significantdifference in the percentage of IL-10-secreting cells between thetwo groups (median values, 10.5 for the RH group and 7 for theMic1.3KO group). We were unable to detect IL-10 in the Foxp3�
population. Moreover, the main sources of IL-10 in both groupswere the CD4� cells, since the percentages of CD4� cells amongIL-10-producing cells in two different experiments were 28% and30% for the RH and the Mic1.3KO groups, respectively.
IL-2 is a cytokine that is essential for Treg development andsurvival (23) and is also involved in the immunosuppression in-duced during the acute phase of toxoplasmosis (24, 25). IL-2 levelswere quantified in spleen culture supernatants after activationwith anti-CD3 (data not shown). IL-2 levels were significantlylower in the RH group than in the naive mice at 7 dpi. The highlevels of IFN-� in the RH group supernatants indicated that thecellular cytokine secretion capacity was not impaired.
Taken together, these results suggest that, despite a strong cel-lular response and high levels of production of IFN-�, the parasiteburden was not controlled in the RH group; in contrast, in theMic1.3KO group the cellular response and the parasite burdenwere controlled.
Induction of Tregs in vivo in the RH group induced bettercontrol of the parasites and reduced the level of inflammatorycytokines. Administration of a complex consisting of a low doseof IL-2 and anti-IL-2 antibody JES6-1 specifically amplifies Tregsin vivo (26). To evaluate further the role of CD4� Foxp3� cells inprotection, RH-infected mice were treated with IL-2/anti-IL-2
complexes, and the levels of cytokine secretion and the parasiteburden were evaluated at 7 dpi. Treatment with IL-2/anti-IL-2complexes significantly increased the frequency of Foxp3� cells inthe spleen. The percentages of CD4� Foxp3� splenocytes in con-trol and IL-2/anti-IL-2-treated mice were 1.6% 0.5% and3.7% 1.6%, respectively, in the first experiment and 1.3% 0.4% and 3.5% 0.3%, respectively, in the second experiment.Mice treated with IL-2/anti-IL-2 complexes showed a diminishednumber of parasites in the peritoneal cavity compared to controlmice injected with PBS, with 0.5 � 106 parasites being found in theIL-2-treated group and 5.5 � 106 parasites being found in thegroup injected with PBS (corresponding to median values fromtwo independent experiments with 5 to 6 mice per group perexperiment), and these diminished parasite burdens were corre-lated with increased numbers of cells (data not shown).
CCL2 and CCL3 levels were higher in the control group than inthe IL-2-treated group (Fig. 6 A and B) at the local level (perito-neum). IFN-� and IL-6 levels in the serum and peritoneum werealso significantly higher in the control group than in the IL-2-treated group (Fig. 6C to F). IL-17A was detectable at low levels atthe local level only in the control group (data not shown).
Specific IFN-� and IL-10 secretion by splenocytes after re-stimulation with the antigen was also significantly higher in theIL-2-treated group than in the control group (Fig. 7A and B).
These findings suggest that the cytokine response after IL-2treatment may be switched from an inflammatory response to amore controlled response.
In addition, mice vaccinated with Mic1.3KO and treated withthe anti-IL-2 receptor antibody PC61, which is known to depleteTregs preferentially, revealed features similar to those of RH-in-fected mice, including a higher parasite count, higher levels ofmorbidity, and higher inflammatory cytokine levels (data notshown).
Taken together, these results indicate that mice infected with
FIG 4 Higher proinflammatory cytokine levels in the RH group. (A to D) IL12p40, IL12p70, and IL-23 were quantified by ELISA in peritoneal washes (IL12p40only) (A) and in sera (B to D) at 7 dpi. (E to H) IFN-� and IL-6 levels in peritoneal washes (E, G) and sera (F, H) at 7 dpi were quantified by ELISA. The levelsof all cytokines were below the detection threshold in control naive mice. The data presented are representative of those from four independent experiments with4 to 6 mice per group. **, P 0.01 using the Wilcoxon-Mann-Whitney exact test with strata; ***, P 0.001 using the Wilcoxon-Mann-Whitney exact test withstrata.
Akbar et al.
3606 iai.asm.org September 2015 Volume 83 Number 9Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

RH and treated with IL-2/anti-IL-2 complexes showed featuressimilar to those of mice vaccinated with Mic1.3KO (a reducedparasite count, a more regulated TH1 response), suggesting a rolefor Tregs in the protection developed with this vaccine strain.
DISCUSSION
The aim of this study was to investigate the role of CD4� Foxp3�
T regulatory cells (Tregs) in the protection induced by an attenu-ated replicating type I Mic1.3KO strain. Type I strains are uni-formly lethal except after attenuation, which leads to lower levelsof replication and dissemination in the host and the developmentof long-term immunity, as for type II strains. Several studies havebeen performed to examine the role of Tregs using a susceptibleC57BL/6 mouse strain (14, 17, 18, 20) and a type II Toxoplasmastrain. Decreased Foxp3� Treg counts at local and systemic siteswere associated with such mortality in C57BL/6-susceptible mice
after lethal oral infection with the type II strain (14). This decreaseoccurred very shortly before the animals died and was correlatedwith virulence. When the dose and the route of infection werechanged and the mice recovered (14, 18), the decrease was onlytransient. In the present study, we investigated the Foxp3� Tregsafter infection with the attenuated Mic1.3KO parasite and com-pared them with the Tregs from the parental RH type I strain,from which Mic1.3KO is derived. In contrast to the results ob-tained after infection with the type II strain (14), the mortalityinduced by infection with the parental RH strain was not associ-ated with a decrease in CD4� Foxp3� Treg numbers or frequen-cies. Neither the number nor the frequency differed from that inthe naive control group. A lack of Treg reduction seemed to con-tradict the uncontrolled immune response observed in the RHgroup and previous studies (14, 18), showing that a decrease inTregs was associated with mortality. However, we cannot excludethe possibility that this decrease appeared just before death or thatit was significant only at the local site. Oldenhove et al. (14)showed a highly significant reduction in Foxp3� Treg frequencyin both the intestine (local) and the spleen at 10 dpi. However, thereduction in Treg splenocytes was significant only at 12 dpi, whichwas the day that the mice died.
In our experiment, a weak increase in Tregs was observed atthe local site after infection with the vaccinal strain. This in-crease was small and transient and was followed by increases inCD4� CD25� T effector cells, which were correlated with par-asite control. This increase in Treg number was limited to localsites, since both the numbers and percentages of CD4� Foxp3�
T cells remained unchanged in spleens until 7 dpi in both in-fected groups compared to those in naive control mice. How-ever, the percentage of CD4� Foxp3� splenocytes expressingCD25 was slightly higher in the RH group than in theMic1.3KO group on day 7. Tenorio et al. (24) reported that,after infection with a type II strain of T. gondii, Tregs wereactivated and showed increased expression of CD25; in addition,there were increased percentages of Tregs expressing CD25. Tregsexpanded in vivo following IL-2 injection also expressed higherlevels of CD25 (27).
The lower increase in the numbers of Tregs in the RH group atthe local site of infection may explain the difference between theMic1.3KO and RH groups. In resistant BALB/c mice, Tregs arethought to reduce immunopathology since depletion of these cellsinduces morbidity due to the increased production of proinflam-matory cytokines and higher parasite burden after oral infection(19). In this study, mice of both the resistant BALB/c and suscep-tible C57BL/6 strains had higher CD4� Foxp3� counts at the locallevel at 6 days postinfection than naive mice, and the absolutecount in the resistant strain was higher than that in the susceptiblestrain. This suggests that differences in Treg counts may accountfor the differences in mortality. This may also have been true inour experiment, since the Treg count was lower for the RH groupthan for the Mic1.3KO group.
Treg expansion/recruitment and accumulation at local sites ofinfection have been extensively described in various parasite in-fections, especially in chronic infections (28).
After infection, the microenvironment may also influence theability of Tregs to determine the outcome of an immune responsetoward tolerance or immunity. TH17 and Treg differentiation isclosely related, and in the presence of proinflammatory cytokines,
FIG 5 Strong systemic, nonspecific IFN-� production and low levels of IL-10production induced by infection with RH. Splenocytes were recovered at 7 dpiwith RH or Mic1.3KO tachyzoites and stimulated with medium or with para-site extract (TE). Culture supernatants were collected after a 48-h stimulationperiod. IFN-� (A) and IL-10 (B) levels were determined by ELISA. The datapresented are representative of those from four independent experiments with4 to 6 mice per group.*, P 0.05 using the Kruskall-Wallis test followed byDunn’s posttest; **, P 0.01 using the Kruskall-Wallis test followed by Dunn’sposttest.
CD4� Foxp3� Cells and Vaccination against Toxoplasmosis
September 2015 Volume 83 Number 9 iai.asm.org 3607Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

transforming growth factor �-induced Foxp3 expression is re-duced, favoring the differentiation of TH17 (29).
Although IL-17A was undetectable in our study, several cyto-kines or chemokines induced in TH17 differentiation or inducedby IL-17 were preferentially expressed in the RH group. CCL2,CCL3, and CCL20 levels were higher in RH-infected mice than inMic1.3KO-infected mice both at the local level and at the systemiclevel. CCL20 is produced by TH17 cells (30), and IL-17 stimulatesthe production of CCL2 and IL-6 (29). CCL2 is produced early
after infection and plays an essential role in resistance to acutetoxoplasmosis by recruitment of Gr1-positive monocytes to thesite of injection (31). Dal Secco et al. (32) also showed that Tregsinhibit CCL2 and CCL3 production by dendritic cells in vivo andlimit the recruitment of inflammatory cells.
Higher levels of IL-6 and IL-23 were found in the RH groupthan in the Mic1.3KO group. Both cytokines are involved in TH17differentiation. Moreover, TH17 cells that differentiate in thepresence of IL-23 are pathogenic (33). The pathological role of
FIG 6 Higher chemokine and proinflammatory cytokine levels in the RH-infected control group than the RH-infected and IL-2-treated group. (A and B) Thechemokine CCL2 (A) and CCL3 (B) levels in peritoneal washes were quantified at 7 dpi. (C to F) IFN-� (C, D) and IL-6 (E, F) levels in peritoneal washes (C, E)and sera (D, F) were quantified at 7 dpi by ELISA. The control and IL-2 groups were injected by the i.p. route with PBS and IL-2/anti-IL-2 antibody, respectively.Results are expressed as the median plus range. The data presented are representative of those from two independent experiments with 5 to 6 mice per group. *,P 0.05; **, P 0.01; ***, P 0.001 using Wilcoxon-Mann Whitney exact test with strata on the cumulative data of the two experiments.
Akbar et al.
3608 iai.asm.org September 2015 Volume 83 Number 9Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

IL-17 has been shown in ocular toxoplasmosis. Sauer et al. (34)demonstrated that the balance between TH17 and TH1 responseswas crucial for the outcome of infection. They also showed a del-eterious TH17 response at the local level after primary infectionwhich was reduced after reinfection. TH2/Treg responses wereenhanced under the reinfection condition (35).
Contrary to the TH1/TH17-type signature after RH infection,the cytokine signature after Mic1.3KO infection in our study wasof the TH1/Treg type.
We observed a higher level of IL-10 in the spleen cell superna-tant of the Mic1.3KO group than in that of the RH group. IL-10 isan immunosuppressive cytokine, and the common feature of alleffector Tregs is the expression of IL-10 (22). Most of the IL-10-secreting splenocytes of both infected groups (75%) were CD4�,and within the CD4 population, the secreting cells were Foxp3�.This result is consistent with previous results reported by Jankovicet al. (36), who showed that the main producer of IL-10 during T.gondii infection within a CD4 population was the Foxp3� IFN-�-positive cell population.
IL-2 is essential for Treg development and survival (23) and isalso known to inhibit TH17 and to favor Treg differentiation.After anti-CD3 activation in vitro, a decrease in the IL-2 level wasobserved in the RH group compared to that in the control naivegroup and the Mic1.3KO group. The low level of IL-2 in the RHgroup may have resulted from the greater consumption of IL-2rather than from lower levels of IL-2 release, since cells were notimpaired in IFN-� production. This low IL-2 level may thereforebe correlated with the higher level of expression of CD25 byFoxp3� cells. This reduced IL-2 level in cells from animals infectedwith T. gondii has been described by others (14, 19, 24). A reducednumber of Tregs during the acute phase of T. gondii infection is aconsequence of a reduced IL-2 availability (14, 19). IL-2 is alsoinvolved in the transient immunosuppression observed duringacute toxoplasmosis (24, 25).
IFN-� levels were higher in the serum, peritoneal cavity, andspleen cell supernatant of the RH group. However, the productionof IFN-� by splenocytes of the RH group was not antigen depen-dent and was not increased by antigen restimulation. These find-ings suggest that splenocytes from the RH group were in an acti-vated state, and the expression of CD69 by these cells further
supports this hypothesis. Expression was not observed until day 7,1 to 3 days before the mice died. CD69 expression was not ob-served in the Mic1.3KO group even at day 11. Expression ofCD69� T cells is known to be localized at sites of chronic inflam-mation and at sites of an active immune response in vivo (37). Therole of CD69 has not yet been elucidated, and previous findingshave suggested that CD69 is an activating molecule, although re-cent studies have also reported that it has regulatory functions.CD69 regulates immune and inflammatory responses by acting asa brake on the differentiation of TH17 effector cells (38).
In summary, the protective response after infection with theattenuated type I-derived Mic1.3KO strain may originate from aTH1 response regulated by an appropriate Treg response. Themortality induced by infection with the parental RH strain mayhave been due to a TH1/TH17-type response.
When the balance was in favor of Tregs by using IL-2/anti-IL-2complexes, we demonstrated that treated mice infected with RHshowed features (a lower parasite burden and TH1/Treg cytokinesand chemokines) similar to those of mice vaccinated withMic1.3KO. These complexes work by inducing and expandingperipheral Tregs (27). Treatment of C57BL/6 mice with IL-2/anti-IL-2 during T. gondii infection with a lethal dose of type II cystswas shown to prevent the loss of Tregs and reduce the level ofmorbidity, but it induced a higher parasite burden in the brain(14). The lower parasite burden that we observed at the local site inthe group in which Tregs were induced is surprising and seemscontradictory to the earlier findings (14). However, a higher par-asite burden in the lamina propria tissue of Treg-depleted BALB/cmice after oral infection has also been reported (19). The authorsargued that this high parasite burden may be an indirect effect ofthe high levels of proinflammatory cytokines at the local site caus-ing tissue destruction. This may also have been true in our exper-iments, since RH infection induced high levels of IFN-� at theinfection site. Alternatively, very high levels of IFN-� and the con-stitutive production of IFN-� may lead to the exhaustion of theresponse as a result of negative feedback or by saturation of theIFN receptor, leading to impairment of the IFN-�-induced re-sponse. This could also be related to a failure to induce Ag-specificIFN-� production, which may be ineffective in parasite control.
In conclusion, this study provides evidence that Tregs may
FIG 7 Systemic, antigen-specific IFN-� and IL-10 production after treatment with IL-2/anti-IL-2 in RH-infected mice. Splenocytes were recovered at 7 dpi withRH tachyzoites and stimulated for 72 h with TE. Culture supernatants were collected after a 48-h stimulation period. The data presented are representative ofthose from two independent experiments with 5 to 6 mice per group. The control and IL-2 groups were injected by the i.p. route with PBS and IL-2/anti-IL-2antibody, respectively. Values were corrected by subtraction of the level in control wells without antigen. IFN-� (A) and IL-10 (B) levels were measured byELISA.*, P 0.05 using the Wilcoxon-Mann-Whitney exact test with strata on the cumulative data from the two experiments.
CD4� Foxp3� Cells and Vaccination against Toxoplasmosis
September 2015 Volume 83 Number 9 iai.asm.org 3609Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

have contributed to protecting mice after infection with an atten-uated, replicating type I strain of T. gondii. It is important to de-termine whether Tregs have a beneficial or detrimental role inprotection against toxoplasmosis in order to design an efficientvaccine. Although a complex mechanism may underlie the pro-tection induced by an attenuated type I strain, we showed that achange in the absolute number of Tregs may be crucial in deter-mining whether animals are protected or whether they will suc-cumb.
ACKNOWLEDGMENTS
Haroon Akbar is the recipient of funding from the Higher EducationCommission, Islamabad, Pakistan.
We thank S. Bigot and T. Papin for their technical assistance. We aregrateful to F. Debierre-Grockiego for critical reading of the manuscriptand providing useful comments. We especially thank Doreen Raine(translator) for revision of the manuscript.
REFERENCES1. Tenter AM, Heckeroth AR, Weiss LM. 2000. Toxoplasma gondii: from
animals to humans. Int J Parasitol 30:1217–1258. http://dx.doi.org/10.1016/S0020-7519(00)00124-7.
2. Jongert E, Roberts CW, Gargano N, Förster-Waldl E, Förster-Wald E,Petersen E. 2009. Vaccines against Toxoplasma gondii: challenges andopportunities. Mem Inst Oswaldo Cruz 104:252–266. http://dx.doi.org/10.1590/S0074-02762009000200019.
3. Fox BA, Bzik DJ. 2002. De novo pyrimidine biosynthesis is required forvirulence of Toxoplasma gondii. Nature 415:926 –929. http://dx.doi.org/10.1038/415926a.
4. Gigley JP, Fox BA, Bzik DJ. 2009. Cell-mediated immunity to Toxo-plasma gondii develops primarily by local Th1 host immune responses inthe absence of parasite replication. J Immunol 182:1069 –1078. http://dx.doi.org/10.4049/jimmunol.182.2.1069.
5. Gigley JP, Fox BA, Bzik DJ. 2009. Long-term immunity to lethal acute orchronic type II Toxoplasma gondii infection is effectively induced in ge-netically susceptible C57BL/6 mice by immunization with an attenuatedtype I vaccine strain. Infect Immun 77:5380 –5388. http://dx.doi.org/10.1128/IAI.00649-09.
6. Jordan KA, Wilson EH, Tait ED, Fox BA, Roos DS, Bzik DJ, Dziersz-inski F, Hunter CA. 2009. Kinetics and phenotype of vaccine-inducedCD8� T-cell responses to Toxoplasma gondii. Infect Immun 77:3894 –3901. http://dx.doi.org/10.1128/IAI.00024-09.
7. Tait ED, Kimberly AJ, Dupont CD, Harris TH, Gregg B, Wilson EH,Pepper M, Dzierszinski F, Roos DS, Hunter CA. 2010. Virulence ofToxoplasma gondii is associated with distinct dendritic cell responses andreduced numbers of activated CD8� T cells. J Immunol 185:1502–1512.http://dx.doi.org/10.4049/jimmunol.0903450.
8. Gazzinelli RT, Hakim FT, Hieny S, Shearer GM, Sher A. 1991. Syner-gistic role of CD4� and CD8� T lymphocytes in IFN-gamma productionand protective immunity induced by an attenuated Toxoplasma gondiivaccine. J Immunol 146:286 –292.
9. Cérède O, Dubremetz JF, Soête M, Deslée D, Vial H, Bout D, Lebrun M.2005. Synergistic role of micronemal proteins in Toxoplasma gondii viru-lence. J Exp Med 201:453–463. http://dx.doi.org/10.1084/jem.20041672.
10. Saej JP, Boyle JP, Boothroyd JC. 2005. Differences among the threemajor strains of Toxoplasma gondii and their specific interactions with theinfected host. Trends Parasitol 21:476 – 481. http://dx.doi.org/10.1016/j.pt.2005.08.001.
11. Gavrilescu LC, Denkers EY. 2001. IFN� overproduction and high levelapoptosis are associated with high but not low virulence Toxoplasma gondiiinfection. J Immunol 167:902–909. http://dx.doi.org/10.4049/jimmunol.167.2.902.
12. Mordue DG, Monroy F, La Regina M, Dinarello CA, Sibley LD. 2001.Acute toxoplasmosis leads to lethal overproduction of Th1 cytokines. J Im-munol 167:4574–4584. http://dx.doi.org/10.4049/jimmunol.167.8.4574.
13. Moiré N, Dion S, Lebrun M, Dubremetz JF, Dimier-Poisson I. 2009.Mic1.3KO tachyzoite a live attenuated vaccine candidate against toxoplas-mosis derived from a type I strain shows features of type II strain. ExpParasitol 123:111–117. http://dx.doi.org/10.1016/j.exppara.2009.06.003.
14. Oldenhove G, Bouladoux N, Wohlfert EA, Hall JA, Chou D, Santos LD,O’Brien S, Blank R, Lamb E, Natarajan S, Kastenmayer R, Hunter C,Grigg ME, Belkaid Y. 2009. Decrease of Foxp3(�) Treg cell number andacquisition of effector cell phenotype during lethal infection. Immunity31:772–786. http://dx.doi.org/10.1016/j.immuni.2009.10.001.
15. Sakaguchi S, Yamaguchi T, Nomura T, Ono M. 2008. Regulatory T cellsand immune tolerance. Cell 133:775–787. http://dx.doi.org/10.1016/j.cell.2008.05.009.
16. Sakaguchi S, Sakaguchi N, Asano M, Itoh M, Toda M. 1995. Immuno-logic self-tolerance maintained by activated T cells expressing IL-2 recep-tor alpha-chains (CD25). Breakdown of a single mechanism of self-tolerance causes various autoimmune diseases. J Immunol 155:1151–1164.
17. Couper KN, Lanthier PA, Perona-Wright G, Kummer LW, Chen W,Smiley ST, Mohrs M, Johnson LL. 2009. Anti-CD25 antibody-mediateddepletion of effector T cell populations enhances susceptibility of mice toacute but not chronic Toxoplasma gondii infection. J Immunol 182:3985–3994. http://dx.doi.org/10.4049/jimmunol.0803053.
18. Tenorio EP, Olguín JE, Fernández J, Vieyra P, Saavedra R. 2010.Reduction of Foxp3� cells by depletion with the PC61 mAb induces mor-tality in resistant BALB/c mice infected with Toxoplasma gondii. J BiomedBiotechnol 2010:786078. http://dx.doi.org/10.1155/2010/786078.
19. Morampudi V, De Craeye S, Le Moine A, Detienne S, Braun MY,D’Souza S. 2011. Partial depletion of CD4� CD25� Foxp3� T regulatorycells significantly increases morbidity during acute phase Toxoplasma gon-dii infection in resistant BALB/c mice. Microbes Infect 13:394 – 404. http://dx.doi.org/10.1016/j.micinf.2011.01.006.
20. Benson A, Murray S, Divakar P, Burnaevskiy N, Pifer R, Forman J,Yarovinsky F. 2012. Microbial infection-induced expansion of effector Tcells overcomes the suppressive effects of regulatory T cells via an IL-2deprivation mechanism. J Immunol 188:800 – 810. http://dx.doi.org/10.4049/jimmunol.1100769.
21. Akbar H, Germon S, Berthon P, Dimier-Poisson I, Moiré N. 2012.Depletion of CD25� cells during acute toxoplasmosis does not signifi-cantly increase mortality in Swiss OF1 mice. Mem Inst Oswaldo Cruz107:155–162. http://dx.doi.org/10.1590/S0074-02762012000200002.
22. Cretney E, Kallies A, Nutt SL. 2013. Differentiation and function ofFoxp3(�) effector regulatory T cells. Trends Immunol 34:74 – 80. http://dx.doi.org/10.1016/j.it.2012.11.002.
23. Malek TR, Bayer AL. 2004. Tolerance, not immunity, crucially dependson IL-2. Nat Rev Immunol 9:665– 674.
24. Tenorio ED, Fernández J, Castellanos C, Olguín JE, Saavedra R. 2011.CD4�Foxp3� regulatory T cells mediate Toxoplasma gondii induced T-cell suppression through an IL-2-related mechanism but independently ofIL-10. Eur J Immunol 41:1–13. http://dx.doi.org/10.1002/eji.201090072.
25. Salinas N, Olguín JE, Castellanos C, Saavedra R. 2014. T cell suppressionin vitro during Toxoplasma gondii infection is the result of IL-2 competi-tion between Tregs and T cells leading to death of proliferating T cells.Scand J Immunol 79:1–11. http://dx.doi.org/10.1111/sji.12120.
26. Webster KE, Walters S, Kohler RE, Mrkvan T, Boyman O, Surh CD,Grey ST, Sprent J. 2009. In vivo expansion of T reg cells with IL-2-mAbcomplexes: induction of resistance to EAE and long-term acceptance ofislet allografts without immunosuppression. J Exp Med 206:751–760.http://dx.doi.org/10.1084/jem.20082824.
27. Shevach EM. 2012. Application of IL-2 therapy to target T regulatorycell function. Trends Immunol 33:626 – 632. http://dx.doi.org/10.1016/j.it.2012.07.007.
28. Belkaid Y, Blank RB, Suffia I. 2006. Natural regulatory T cells andparasites: a common quest for host homeostasis. Immunol Rev 212:287–300. http://dx.doi.org/10.1111/j.0105-2896.2006.00409.x.
29. Onishi RM, Gaffen SL. 2010. Interleukin-17 and its target genes: mech-anisms of interleukin-17 function in disease. Immunology 129:311–321.http://dx.doi.org/10.1111/j.1365-2567.2009.03240.x.
30. Louten J, Boniface K, de Waal Malefyt R. 2009. Development andfunction of TH17 cells in health and disease. J Allergy Clin Immunol123:1004 –1011. http://dx.doi.org/10.1016/j.jaci.2009.04.003.
31. Robben PM, La Regina M, Kuziel WA, Sibley LD. 2005. Recruitment ofGr-1� monocytes is essential for control of acute toxoplasmosis. J ExpMed 201:1761–1769. http://dx.doi.org/10.1084/jem.20050054.
32. Dal Secco V, Soldani C, Debrat C, Asperti-Boursin F, Donnadieu E,Viola A, Sarukhan A. 2009. Tunable chemokine production by antigenpresenting dendritic cells in response to changes in regulatory T cell fre-
Akbar et al.
3610 iai.asm.org September 2015 Volume 83 Number 9Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from

quency in mouse reactive lymph nodes. PLoS One 4:e7696. http://dx.doi.org/10.1371/journal.pone.0007696.
33. Croxford AL, Mair F, Becher B. 2012. IL-23: one cytokine in control ofautoimmunity. Eur J Immunol 42:2263–2273. http://dx.doi.org/10.1002/eji.201242598.
34. Sauer A, Pfaff AW, Villard O, Creuzot-Garcher C, Dalle F, Chiquet C,Pelloux H, Speeg-Schatz C, Gaucher D, Prevost G, Bourcier T, CandolfiE. 2012. Interleukin 17A as an effective target for anti-inflammatory andantiparasitic treatment of toxoplasmic uveitis. J Infect Dis 206:1319 –1329.http://dx.doi.org/10.1093/infdis/jis486.
35. Sauer A, Rochet E, Lahmar I, Brunet J, Sabou M, Bourcier T, CandolfiE, Pfaff AW. 2013. The local immune response to intraocular Toxoplasmare-challenge: less pathology and better parasite control through Treg/Th1/
Th2 induction. Int J Parasitol 43:721–728. http://dx.doi.org/10.1016/j.ijpara.2013.04.004.
36. Jankovic D, Kullberg MC, Feng CG, Goldszmid RS, Collazo CM,Wilson M, Wynn TA, Kamanaka M, Flavell RA, Sher A. 2007. Conven-tional T-bet(�)Foxp3(�) Th1 cells are the major source of host-protective regulatory IL-10 during intracellular protozoan infection. J ExpMed 204:273–283. http://dx.doi.org/10.1084/jem.20062175.
37. Sancho D, Gomez M, Sanchez-Madrid F. 2004. CD69 is an immunoregu-latory molecule induced following activation. Trends Immunol 26:136–140.
38. Martín P, Gómez M, Lamana A, Cruz-Adalia A, Ramírez-Huesca M,Ursa MA, Yáñez-Mo M, Sánchez-Madrid F. 2010. CD69 association withJak3/Stat5 proteins regulates Th17 cell differentiation. Mol Cell Biol 30:4877– 4889. http://dx.doi.org/10.1128/MCB.00456-10.
CD4� Foxp3� Cells and Vaccination against Toxoplasmosis
September 2015 Volume 83 Number 9 iai.asm.org 3611Infection and Immunity
on April 14, 2020 by guest
http://iai.asm.org/
Dow
nloaded from