role of ccaat/enhancer-binding protein site in transcription of
TRANSCRIPT

of January 29, 2018.This information is current as
Peptide-1 and -3 Defensin GenesSite in Transcription of Human Neutrophil Role of CCAAT/Enhancer-Binding Protein
Yuko Tsutsumi-Ishii, Takeshi Hasebe and Isao Nagaoka
http://www.jimmunol.org/content/164/6/3264doi: 10.4049/jimmunol.164.6.3264
2000; 164:3264-3273; ;J Immunol
average*
4 weeks from acceptance to publicationSpeedy Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
?The JIWhy
Referenceshttp://www.jimmunol.org/content/164/6/3264.full#ref-list-1
, 23 of which you can access for free at: cites 52 articlesThis article
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2000 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on January 29, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Role of CCAAT/Enhancer-Binding Protein Site inTranscription of Human Neutrophil Peptide-1 and -3 DefensinGenes1,2
Yuko Tsutsumi-Ishii, Takeshi Hasebe, and Isao Nagaoka3
The human neutrophil defensins (human neutrophil peptides (HNPs)), major components of azurophilic granules, contribute toinnate and acquired host immunities through their potent antimicrobial activities and ability to activate T cells. Despite beingencoded by nearly identical genes, HNP-1 is more abundant in the granules than HNP-3. We investigated the regulation of HNP-1and HNP-3 expression at the transcriptional level using a promyelocytic HL-60 cell line. Luciferase analysis showed that tran-scriptional levels ofHNP-1 and HNP-3 promoters were equivalent and that an;200-bp region identical between promoters wassufficient for transcriptional activity. Furthermore, overlapping CCAAT/enhancer-binding protein (C/EBP) and c-Myb sites in theregion were found to be required for efficient transcription. Gel mobility shift assay demonstrated that C/EBPa predominantlybound to the C/EBP/c-Myb sites using HL-60 nuclear extracts. No specific binding to C/EBP/c-Myb sites was observed in nuclearextracts from mature neutrophils, which expressed neither C/EBPa protein nor HNP mRNAs. Taken together, these findingssuggest that the difference in the amounts of HNP-1 and HNP-3 peptides in neutrophils is caused by posttranscriptional regulationand that C/EBPa plays an important role in the transcription of HNP genes in immature myeloid cells. The Journal of Immu-nology,2000, 164: 3264–3273.
N eutrophils act as key effectors in host defenses againstmicrobial infection. They ingest and destroy invadingmicrobes through both oxidative and nonoxidative
mechanisms (1). The former mechanism depends on reactive ox-ygen intermediates produced by activated neutrophils (2, 3),whereas the latter is controlled by antimicrobial proteins, peptides,or both in cytoplasmic granules independent of reactive oxygenmetabolites (4). A number of microbicidal polypeptides have beenisolated from neutrophils of humans and various animal species(5–7). The most abundant are low-molecular-weight cationic pep-tides called “defensins,” which exhibit a broad spectrum of micro-bicidal activities against Gram-positive and Gram-negative bacte-ria, fungi, parasites, and viruses (8–10). The defensin family ofantimicrobial peptides are widely distributed among many mam-mals, insects, and plants (9–13), and have three or four intramo-lecular disulfide bonds which differ in the placement and connec-tivity of their conserved cysteine residues (9–13). The vertebratedefensins consist of two subfamilies, designeda andb defensins(10, 11).
In humans,a defensins (human neutrophil peptides (HNPs))4
are major components of the primary (azurophil) granules of neu-
trophils and comprise 30–50% of azurophil granule protein (8, 14,15). Among HNPs, HNP-1, HNP-2, and HNP-3 have almost iden-tical amino acid sequences (9). Both HNP-1 and HNP-3 are com-posed of 30 amino acid residues, with the first amino acid inHNP-1, alanine, substituted by aspartate in HNP-3. HNP-2, 29amino acids in length due to lack of the first amino acid, is as-sumed to be a proteolytic derivative of HNP-1, HNP-3, or both (9).HNP-1 and HNP-3 are encoded by nearly identical genes on chro-mosome 8; notwithstanding this similarity, HNP-1 is more abun-dant than HNP-3 in neutrophil granules (16–18). The fourth de-fensin, HNP-4, is the least abundant and except for conservedcysteine residues shows low homology with other HNPs (19–21).Despite their nearly identical amino acid sequences, HNPs exhibitdifferent biological properties in vitro. For example, althoughHNP-1, HNP-2, and HNP-4 can killCandida albicans, HNP-3cannot (20, 22). Interestingly, HNP-1 and HNP-2, but not HNP-3,have chemotactic activity for murine and human T cells and mono-cytes (16, 23). Moreover, HNP-1, HNP-2, and HNP-3 have beenshown to enhance T cell-dependent immune responses in vivo(24). Such diverse functions of HNPs are likely to contribute toboth innate and acquired host defenses.
In contrast to the abundance of information on the function ofthese peptides, their gene regulation is little known. HNP mRNAsare only expressed in immature bone marrow cells and HL-60human promyelocytic leukemia cells (25–28). A more recent re-port has indicated that PU.1 and an unknown Ets-like factor (IRD)are involved in the basal transcription ofHNP-1 gene (29). How-ever, given that several transcription factors such as CCAAT/en-hancer-binding proteins (C/EBPs), c-Myb, and AML1 (PEBP2/CBF) cooperatively regulate the efficient transcription of manymyeloid-specific genes (30–32), factors other than PU.1 and IRDmay also play a role in the transcriptional regulation of HNPs.Furthermore, it is unclear whether the difference in HNP-1 andHNP-3 contents in neutrophil granules is caused by transcriptionalregulation, posttranscriptional regulation, or both. Detailed analy-sis of HNP-1 andHNP-3 genes should therefore provide clues tothe regulation of HNP expression.
Department of Biochemistry, Juntendo University, School of Medicine, Tokyo, Japan
Received for publication July 26, 1999. Accepted for publication January 5, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by grants from Takeda Science Foundation and theAtopy (Allergy) Research Center, Juntendo University.2 The sequences of the HNP-1 and HNP-3 promoters have been deposited in theGenBank database under accession numbers AB025231 and AB025232, respectively.3 Address correspondence and reprint requests to Dr. Isao Nagaoka, Department ofBiochemistry, Juntendo University, School of Medicine, Hongo 2-1-1, Bunkyo-ku,Tokyo, 113-8421 Japan. E-mail address: [email protected] Abbreviations used in this paper: HNP, human neutrophil peptide; C/EBP, CCAAT/enhancer-binding protein; IRD, increased regulatory element of defensin expressionbinding during differentiation; EMSA, electrophoretic mobility shift assay; CDP,CCAAT displacement protein.
Copyright © 2000 by The American Association of Immunologists 0022-1767/00/$02.00
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

In the present study, we show thatHNP-1 and HNP-3 geneshave equivalent levels of promoter activity in HL-60 cells and that;200 bp of 59flanking sequence identical betweenHNP-1 andHNP-3 is sufficient for transcription activity. We further demon-strate that the binding of C/EBPa but not c-Myb protein to theoverlapping C/EBP and c-Myb sites is important for efficient tran-scription ofHNP promoters in HL-60 cells. Our findings suggestthat C/EBPa acts as a potent positive regulator of theHNP-1andHNP-3 genes.
Materials and MethodsCell culture
Human promyelocytic leukemia HL-60 cells (JCRB0085; Japanese Col-lection of Research Bioresource, Tokyo, Japan) and T lymphoblastic leu-kemia Jurkat cells (TIB-152; American Type Culture Collection, Manas-sas, VA) were maintained in RPMI 1640 medium (Nissui Pharmaceutical,Tokyo, Japan) supplemented with 10% FBS (Sanko, Tokyo, Japan), 100U/ml penicillin, and 0.1 mg/ml streptomycin (Sigma, St. Louis, MO) at37°C in 5% CO2. Cervical carcinoma HeLa cells (JCRB9004) were cul-tured in DMEM (Nissui Pharmaceutical) with 10% FBS, 100 U/ml peni-cillin, and 0.1 mg/ml streptomycin at 37°C.
Preparation of peripheral blood neutrophils
Peripheral blood obtained from healthy volunteers was heparinized, andneutrophils (purity. 95%) were isolated by Ficoll-Conray centrifugationafter dextran sedimentation of erythrocytes (28). In some experiments, neu-trophils were resuspended in RPMI 1640 at a final concentration of 53 106
cells/ml and stimulated with 1mg/ml LPS (fromEscherichia coliO127:B8;Difco, Detroit, MI) or 100 U/ml recombinant human TNF-a (Genzyme,Boston, MA) at 37°C for 24 h.
Isolation of HNP-1 and HNP-3 promoters
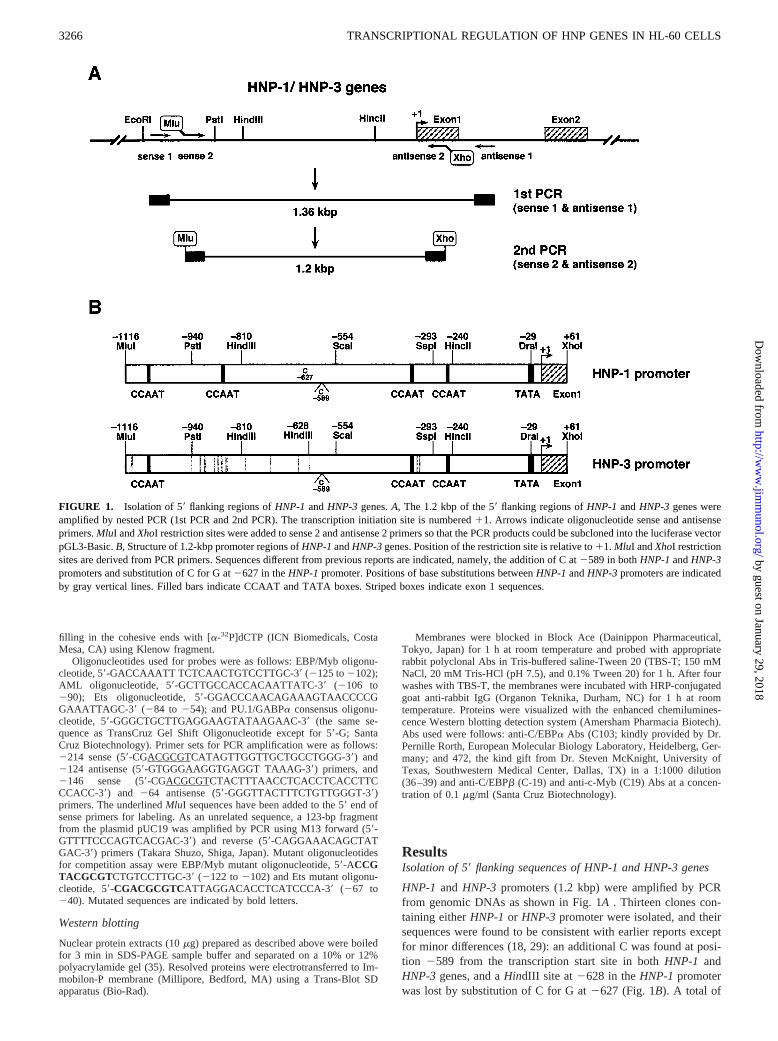
The sequence of the 59flanking region of exon 1 of humanHNP-1 andHNP-3genes was amplified from human genomic DNA by PCR based onpublished sequences as illustrated in Fig. 1A(18). The first PCR reactionwas performed with a primer set of sense 1 and antisense 1 on a thermalcycler model 480 (Perkin-Elmer, Norwalk, CT) after incubation at 94°C for2 min using 30 cycles of 30 s at 94°C, 30 s at 54°C, and 1 min at 72°C. Thefinal polymerization step was extended by an additional 5 min at 72°C. Thesecond PCR was conducted using the first reaction as a template with aprimer set of sense 2 and antisense 2. The synthesized 1.2-kbp fragmentsconsisted of the sequences from21116 to161 of HNP genes, and werecloned to a TA-cloning vector pT7Blue(R) (Novagen, Madison, WI). Plas-mid inserts were confirmed by sequencing using a Dye Terminator CycleSequencing kit FS and a model 373A DNA autosequencer (PE AppliedBiosystems Division, Foster City, CA). Base substitutions betweenHNP-1andHNP-3 promoters are shown in Fig. 1B.
Oligonucleotides used were sense 1 primer (59-AGCCCTGTTACAGGGGCTGC-39; located at21159 from the transcription start site),sense 2 primer containing aMluI site (59-CCGACGCGTCGTCCAAGGCAGGCAACTCAACCC-39; at 21116), antisense 1 primer (59-ACGTTCCCTAGCAGGGATTCACCCGC-39; at 1199), and antisense 2 primercontaining aXhoI site (59-CCGCTCGAGCGCAGGGTGACCAGAGAGGGCAGA-39; at 161). Underlining in the sequences indicate additionalrestriction sites.
Plasmid construction
The 1.2-kbpMluI-XhoI fragments ofHNP-1 or HNP-3 promoter frompT7Blue(R) clones were subcloned into the promoterless luciferase vectorpGL3-Basic (Promega, Madison, WI) and named21116/HNP1-Luc and21116/HNP3-Luc, respectively. A series of 59deletion plasmids shown inFig. 2Awas constructed as follows. To remove 59upstream sequences ofHNP promoters,21116/HNP1-Luc or21116/HNP3-Luc plasmids weredigested withSmaI (located in the polylinker site of pGL3-Basic) andSspI(position 2293 on theHNP promoters),HincII (2240), or DraI (229).Each plasmid was recircularized by ligation to generate2293/HNP-Luc,2240/HNP-Luc, and229/HNP-Luc, respectively. The2240/HNP-Lucplasmid was utilized as a template for PCR with appropriate sense primerswith MluI sequence at the 59end and antisense 2 primer to construct furtherdeletion derivatives,2133/HNP-Luc, 2111/HNP-Luc, 286/HNP-Luc,and258/HNP-Luc (Fig. 4A). Sequences of sense primers were as follows:2133 sense, 59-CTCGTACGCGTCCTTCCCAC-39(2133 to 2124);2111 sense, 59-ACCGTACGCGTCTGTCCTTGC-39(2111 to 2102);286 sense, 59-ACCGTACGCGTATGGACCCA-39(286 to 277); and
258 sense, 59-CGACGCGTCATTAGGACACCTCATCCCA-39(258 to240). Underlining in the sequences indicate theMluI site.
A 2133DAML/HNP-Luc plasmid containing a mutant AML1 site from2100 to295 was created by PCR using mutant AML1 primers, where anAML1 element (59-ACCACA-39) was replaced by anAgeI restriction site(59-ACCGGT-39) (Fig. 4A). PCR reactions were conducted with2133sense and mutant AML1 antisense (283 to2103; 59-CCATTAAATAATACCGGTGGC-39), and mutant AML1 sense (2103 to283; 59-GCCACCGGTATTATTTAATGG-39) and antisense 2 primers. Bold letters indi-cate theAgeI site. Amplified PCR products were digested withAgeI andligated to generate2133DAML/HNP-Luc. All deletion constructs wereconfirmed by sequencing.
Transfection and luciferase assay
HL-60 (1 3 107) cells were transfected with 20mg of luciferase reporterconstructs and 10mg of b-galactosidase expression vector pSV-b-galac-tosidase (Promega) in 500ml of serum-free RPMI 1640 medium by elec-troporation at 960mF and 280 V using a Gene Pulser apparatus (Bio-Rad,Hercules, CA) as described previously (33). Cells were incubated in 10 mlRPMI 1640 supplemented with 10% FBS for 6 h posttransfection.
HeLa cells were plated at 43 105 cells/60-mm dish. On the followingday, the cells were cotransfected with 8mg reporter plasmids and 2mgpSV-b-galactosidase with a calcium phosphate transfection system (LifeTechnologies, Rockville, MD) according to the manufacturer’s protocol.After 24 h, DNA precipitates were removed by washing, and the cells wereincubated for another 24 h at 37°C.
To analyze luciferase andb-galactosidase activities, cells were lysed in200ml PicaGene Reporter Lysis Buffer LUC (Toyo Ink, Tokyo, Japan) andsonicated on ice for 10 s (ultrasonic disrupter; Tomy Seiko, Tokyo, Japan).Protein concentration of cell extracts was determined with a bicinchoninicacid protein assay kit (Pierce, Rockford, IL). Luciferase activity was mea-sured as relative light units using a PicaGene luciferase assay kit (ToyoInk) and a Lumat LB9501 luminometer (Berthold, Wildbad, Germany).b-Galactosidase activity was measured with a Galacto-Light kit (Tropix,Bedford, MA), and transfection efficiency was normalized to the level ofb-galactosidase activity.
Preparation of nuclear extracts
Nuclear extracts were prepared as described by Dignam et al. (34), withminor modifications. Briefly, 13 108 cells were washed twice with PBSand lysed in lysis buffer (10 mM HEPES (pH 7.9), 10 mM KCl, 1 mMEDTA, 1.5 mM MgCl2, 0.5% Nonidet P-40, 1 mM DTT, 1 mM PMSF, 5mg/ml leupeptin, and 5mg/ml pepstatin) on ice for 10 min. Nuclei pelletswere washed once with the same buffer except Nonidet P-40. After incu-bation with extraction buffer (10 mM HEPES (pH 7.9), 420 mM NaCl, 1mM EDTA, 1.5 mM MgCl2, 20% glycerol, 1 mM DTT, 1 mM PMSF, 5mg/ml leupeptin, and 5mg/ml pepstatin) at 4°C for 20 min, nuclei werecentrifuged at 12,0003 g for 20 min at 4°C. The resultant nuclear extractswere immediately subjected to SDS-PAGE/Western blotting or stored at280°C for gel retardation assay. Protein content in the nuclear extracts wasmeasured with a bicinchoninic acid protein assay kit (Pierce).
Electrophoretic mobility shift assay (EMSA)
Nuclear extracts (10mg) were mixed with a32P-labeled probe (describedin the next section; 53 104 cpm, 10–20 fmol) in 15ml of a binding buffercontaining 10 mM HEPES (pH 7.9), 50 mM KCl, 5 mM MgCl2, 1 mMEDTA, 5% glycerol, 1 mM DTT, 1 mM PMSF, 1mg/ml leupeptin, 1mg/mlpepstatin, and 2mg poly(dI-dC)zpoly(dI-dC) (Amersham Pharmacia Bio-tech AB, Uppsala, Sweden) for 20 min on ice. The reaction mixtures wereapplied to a native 5% polyacrylamide gel in 0.53TBE (44.5 mM Tris,44.5 mM boric acid, and 1 mM EDTA (pH 8.3)) at 180 V for 90 min at4°C. The gels were dried and exposed to Fuji RX-U x-ray film (Fuji PhotoFilm, Tokyo, Japan) at280°C. For competition assay, a 20–50-fold molarexcess of unlabeled oligonucleotides and PCR products were preincubatedin reaction mixture for 15 min on ice. For Ab supershift experiments, 1mlof rabbit polyclonal Abs to C/EBPa [14-AA], C/EBPb [C-19], C/EBPb[D198], c-Myb [C-19], and PU.1 [T-21] or normal rabbit IgG was added tothe reaction mixture 20 min before probe addition. All specific Abs (Tras-Cruz Gel Supershift reagents, 1 mg/ml) were obtained from Santa CruzBiotechnology (Santa Cruz, CA).
Oligonucleotides for EMSA
Synthetic oligonucleotides or PCR products were used as probes forEMSA. The oligonucleotides were designed to generate a single 59-G over-hang to each end after annealing with their compliments. PCR productswere digested withMluI to generate a 59-CGCG overhang. Double-stranded oligonucleotides and digested PCR fragments were labeled by
3265The Journal of Immunology
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
filling in the cohesive ends with [a-32P]dCTP (ICN Biomedicals, CostaMesa, CA) using Klenow fragment.
Oligonucleotides used for probes were as follows: EBP/Myb oligonu-cleotide, 59-GACCAAATT TCTCAACTGTCCTTGC-39(2125 to2102);AML oligonucleotide, 59-GCTTGCCACCACAATTATC-39(2106 to290); Ets oligonucleotide, 59-GGACCCAACAGAAAGTAACCCCGGAAATTAGC-39 (284 to 254); and PU.1/GABPa consensus oligonu-cleotide, 59-GGGCTGCTTGAGGAAGTATAAGAAC-39(the same se-quence as TransCruz Gel Shift Oligonucleotide except for 59-G; SantaCruz Biotechnology). Primer sets for PCR amplification were as follows:2214 sense (59-CGACGCGTCATAGTTGGTTGCTGCCTGGG-39) and2124 antisense (59-GTGGGAAGGTGAGGT TAAAG-39) primers, and2146 sense (59-CGACGCGTCTACTTTAACCTCACCTCACCTTCCCACC-39) and 264 antisense (59-GGGTTACTTTCTGTTGGGT-39)primers. The underlinedMluI sequences have been added to the 59 end ofsense primers for labeling. As an unrelated sequence, a 123-bp fragmentfrom the plasmid pUC19 was amplified by PCR using M13 forward (59-GTTTTCCCAGTCACGAC-39) and reverse (59-CAGGAAACAGCTATGAC-39) primers (Takara Shuzo, Shiga, Japan). Mutant oligonucleotidesfor competition assay were EBP/Myb mutant oligonucleotide, 59-ACCGTACGCGTCTGTCCTTGC-39(2122 to2102) and Ets mutant oligonu-cleotide, 59-CGACGCGTCATTAGGACACCTCATCCCA-39 (267 to240). Mutated sequences are indicated by bold letters.
Western blotting
Nuclear protein extracts (10mg) prepared as described above were boiledfor 3 min in SDS-PAGE sample buffer and separated on a 10% or 12%polyacrylamide gel (35). Resolved proteins were electrotransferred to Im-mobilon-P membrane (Millipore, Bedford, MA) using a Trans-Blot SDapparatus (Bio-Rad).
Membranes were blocked in Block Ace (Dainippon Pharmaceutical,Tokyo, Japan) for 1 h at room temperature and probed with appropriaterabbit polyclonal Abs in Tris-buffered saline-Tween 20 (TBS-T; 150 mMNaCl, 20 mM Tris-HCl (pH 7.5), and 0.1% Tween 20) for 1 h. After fourwashes with TBS-T, the membranes were incubated with HRP-conjugatedgoat anti-rabbit IgG (Organon Teknika, Durham, NC) for 1 h at roomtemperature. Proteins were visualized with the enhanced chemilumines-cence Western blotting detection system (Amersham Pharmacia Biotech).Abs used were follows: anti-C/EBPa Abs (C103; kindly provided by Dr.Pernille Rorth, European Molecular Biology Laboratory, Heidelberg, Ger-many; and 472, the kind gift from Dr. Steven McKnight, University ofTexas, Southwestern Medical Center, Dallas, TX) in a 1:1000 dilution(36–39) and anti-C/EBPb (C-19) and anti-c-Myb (C19) Abs at a concen-tration of 0.1mg/ml (Santa Cruz Biotechnology).
ResultsIsolation of 59flanking sequences of HNP-1 and HNP-3 genes
HNP-1 and HNP-3 promoters (1.2 kbp) were amplified by PCRfrom genomic DNAs as shown in Fig. 1A . Thirteen clones con-taining eitherHNP-1 or HNP-3 promoter were isolated, and theirsequences were found to be consistent with earlier reports exceptfor minor differences (18, 29): an additional C was found at posi-tion 2589 from the transcription start site in bothHNP-1 andHNP-3 genes, and aHindIII site at 2628 in theHNP-1 promoterwas lost by substitution of C for G at2627 (Fig. 1B). A total of
FIGURE 1. Isolation of 59flanking regions ofHNP-1 andHNP-3 genes.A, The 1.2 kbp of the 59flanking regions ofHNP-1 andHNP-3 genes wereamplified by nested PCR (1st PCR and 2nd PCR). The transcription initiation site is numbered11. Arrows indicate oligonucleotide sense and antisenseprimers.MluI andXhoI restriction sites were added to sense 2 and antisense 2 primers so that the PCR products could be subcloned into the luciferase vectorpGL3-Basic.B, Structure of 1.2-kbp promoter regions ofHNP-1andHNP-3genes. Position of the restriction site is relative to11. MluI andXhoI restrictionsites are derived from PCR primers. Sequences different from previous reports are indicated, namely, the addition of C at2589 in bothHNP-1andHNP-3promoters and substitution of C for G at2627 in theHNP-1promoter. Positions of base substitutions betweenHNP-1andHNP-3promoters are indicatedby gray vertical lines. Filled bars indicate CCAAT and TATA boxes. Striped boxes indicate exon 1 sequences.
3266 TRANSCRIPTIONAL REGULATION OF HNP GENES IN HL-60 CELLS
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
15 base differences were located upstream of aSspI site at2293betweenHNP-1andHNP-3promoters, and;300 bp of the prox-imal region was identical between the two promoters. As shown inFig. 1B, there were three (HNP-1) or two (HNP-3) CCAAT boxeswithin the distal region (21116 to2293), whereas one CCAAT boxand a TATA box were found within the identical region (18, 29).
Transcription potentials of HNP-1 and HNP-3 promoters in HL-60 cells
To examine the promoter activity ofHNP-1 and HNP-3, the 59flanking region from21116 to161 of both genes was introduced
into the luciferase reporter vector to form21116/HNP1-Luc and21116/HNP3-Luc, respectively (Fig. 2A).
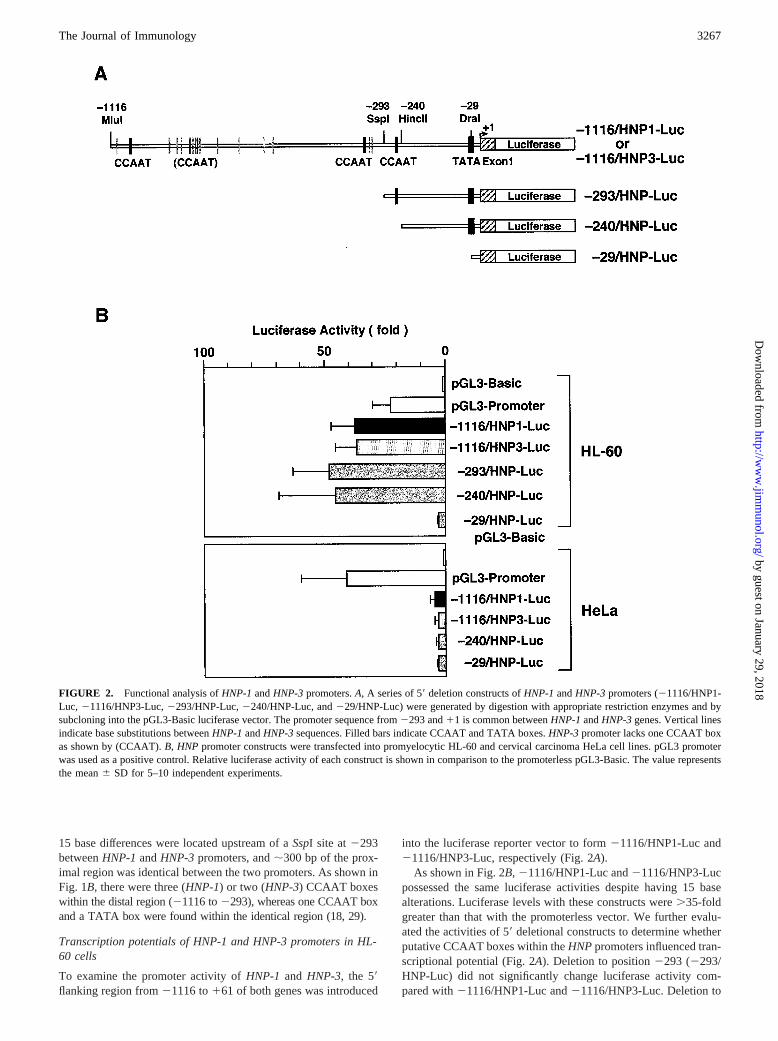
As shown in Fig. 2B,21116/HNP1-Luc and21116/HNP3-Lucpossessed the same luciferase activities despite having 15 basealterations. Luciferase levels with these constructs were.35-foldgreater than that with the promoterless vector. We further evalu-ated the activities of 59deletional constructs to determine whetherputative CCAAT boxes within theHNPpromoters influenced tran-scriptional potential (Fig. 2A). Deletion to position2293 (2293/HNP-Luc) did not significantly change luciferase activity com-pared with21116/HNP1-Luc and21116/HNP3-Luc. Deletion to
FIGURE 2. Functional analysis ofHNP-1andHNP-3promoters.A, A series of 59deletion constructs ofHNP-1andHNP-3promoters (21116/HNP1-Luc, 21116/HNP3-Luc,2293/HNP-Luc,2240/HNP-Luc, and229/HNP-Luc) were generated by digestion with appropriate restriction enzymes and bysubcloning into the pGL3-Basic luciferase vector. The promoter sequence from2293 and11 is common betweenHNP-1andHNP-3genes. Vertical linesindicate base substitutions betweenHNP-1andHNP-3sequences. Filled bars indicate CCAAT and TATA boxes.HNP-3promoter lacks one CCAAT boxas shown by (CCAAT).B, HNPpromoter constructs were transfected into promyelocytic HL-60 and cervical carcinoma HeLa cell lines. pGL3 promoterwas used as a positive control. Relative luciferase activity of each construct is shown in comparison to the promoterless pGL3-Basic. The value representsthe mean6 SD for 5–10 independent experiments.
3267The Journal of Immunology
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
position 2240 (2240/HNP-Luc) showed retained activity,whereas additional deletion to position229, which removed apotential TATA box, decreased promoter activity by.90%. Theseresults indicate that base substitutions and CCAAT boxes were oflittle consequence in the promoter activity ofHNP-1 andHNP-3genes, and that the proximal promoter region between2240 and229 contains positivecis-acting element(s) interacting with tran-scription factors in HL-60 cells.
In contrast, in the cervical carcinoma HeLa cell line, which doesnot express HNP mRNAs, full-lengthHNP-1or HNP-3promotersdisplayed only 3-fold stimulation over the promoterless vector(Fig. 2B). Moreover, these promoter activities were not altered bythe deletion to position229.
Taken together, these findings suggest thatHNP-1 andHNP-3genes are equally transcribed, and that both HNP genes are tran-scriptionally regulated in a myeloid-specific manner.
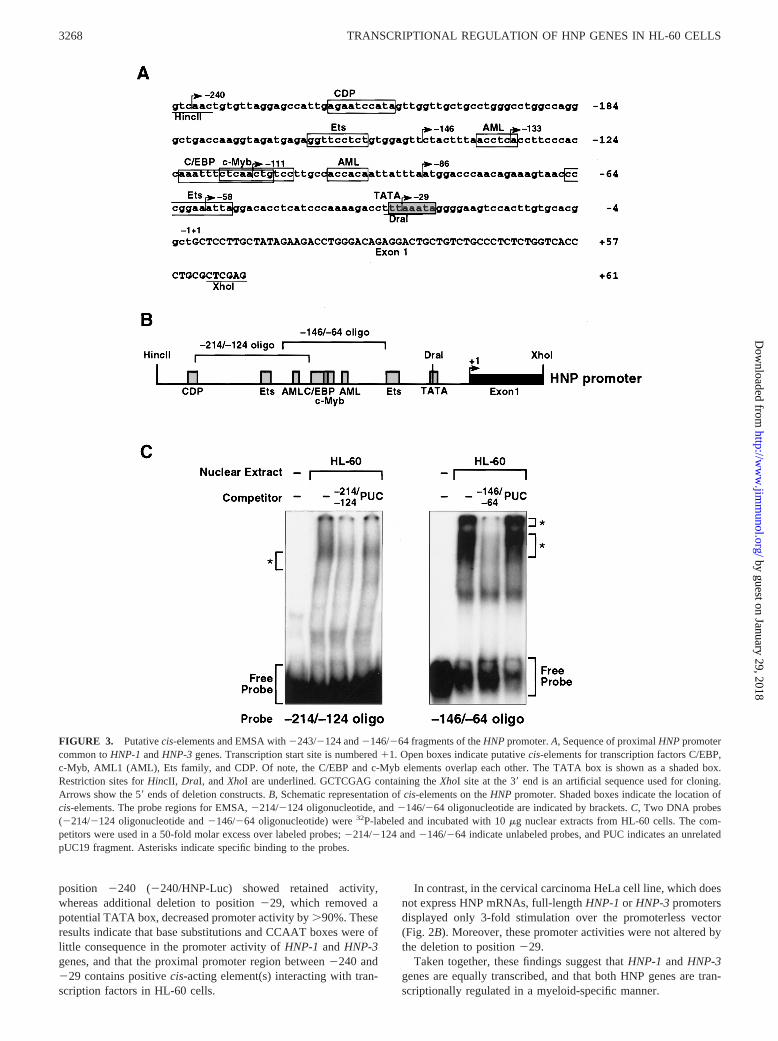
FIGURE 3. Putativecis-elements and EMSA with2243/2124 and2146/264 fragments of theHNPpromoter.A, Sequence of proximalHNPpromotercommon toHNP-1andHNP-3genes. Transcription start site is numbered11. Open boxes indicate putativecis-elements for transcription factors C/EBP,c-Myb, AML1 (AML), Ets family, and CDP. Of note, the C/EBP and c-Myb elements overlap each other. The TATA box is shown as a shaded box.Restriction sites forHincII, DraI, andXhoI are underlined. GCTCGAG containing theXhoI site at the 39end is an artificial sequence used for cloning.Arrows show the 59ends of deletion constructs.B, Schematic representation ofcis-elements on theHNP promoter. Shaded boxes indicate the location ofcis-elements. The probe regions for EMSA,2214/2124 oligonucleotide, and2146/264 oligonucleotide are indicated by brackets.C, Two DNA probes(2214/2124 oligonucleotide and2146/264 oligonucleotide) were32P-labeled and incubated with 10mg nuclear extracts from HL-60 cells. The com-petitors were used in a 50-fold molar excess over labeled probes;2214/2124 and2146/264 indicate unlabeled probes, and PUC indicates an unrelatedpUC19 fragment. Asterisks indicate specific binding to the probes.
3268 TRANSCRIPTIONAL REGULATION OF HNP GENES IN HL-60 CELLS
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Requirement for overlapping C/EBP and c-Myb sites and Ets-like element for activation of the HNP promoter
To localizecis-acting elements within theHNP promoter, the se-quence from2240 and229 was scanned with the MatInspectorversion 2.2 computer program (40, 41). Putative binding sites forC/EBP, c-Myb, AML1, Ets, and CCAAT displacement protein(CDP), which are related to myeloid-specific gene expression,were contained within the region;200 bp upstream of the TATAbox (Fig. 3A) (30–32, 42).
We analyzed whether the nuclear factors bound to the predictedsites ofHNP promoter. EMSA was performed with aa-32P-la-beled DNA fragment containing the HNP promoter (Fig. 3B) andnuclear extracts prepared from HL-60 cells. As represented in Fig.3C, nuclear protein(s) in HL-60 cells strikingly bound to the2146/264 oligonucleotide containing putative C/EBP, c-Myb, andAML1 sites. Formation of the two major complexes was specifi-cally inhibited by the addition of unlabeled probe, but not by thatof unrelated pUC19 fragment (Fig. 3C,right panel). Weak bindingto the fragment from2214 to 2124 containing distal Ets and
AML1 sites was developed, which competed with the excess un-labeled2214/2124 fragment but not with the unrelated pUC19fragment (Fig. 3C,left panel). These findings indicate that an;200-bp region of HNP promoter contains multiple binding sitesfor transcription factors.
Next, to elucidate which element(s) were functionally important toHNP promoter activity, successive 59truncates and a point mutantwere created from2240/HNP-Luc using PCR (Fig. 4A) and assessedin HL-60 cells (Fig. 4B). Deletion to position2133 showed a ten-dency to increased promoter activity compared with2240/HNP-Luc,suggesting the loss of negative regulatory elements. Deletion to po-sition 2111, which removed overlapping C/EBP and c-Myb sites,resulted in a remarkable decrease in promoter activity. Interestingly,mutation or deletion of the proximal AML1 site at2100(2133DAML and 286/HNP-Luc) had little effect on promoter ac-tivity compared with those of2133 and2111/HNP-Luc, respec-tively. Additional deletion to position258, which removed the bind-ing site for Ets family proteins, resulted in a further decrease inactivity. Promoter activity was virtually abolished by deletion of a
FIGURE 4. Luciferase activity of 59deletion and point mutation constructs ofHNP promoter in HL-60 cells.A, Successive deletion constructs ofputativecis-elements and a mutant of the AML1 site were generated by PCR and by subcloning to the pGL3-Basic vector. Shaded boxes indicate thelocation of putativecis-elements on theHNP promoter. The crossed box of2133DAML/HNP-Luc shows a mutated AML1 element.B, Deletion and pointmutation constructs were transiently transfected into HL-60 cells. Average luciferase activities were generated from 5 to 10 separate experiments. SD ofthe mean is indicated by error bars. Data are presented as the percentage of activity ofHNP promoter constructs vs that of2240/HNP-Luc (100%).
3269The Journal of Immunology
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
putative TATA box (at229). These results indicate that in addition tothe TATA box, the overlapping C/EBP and c-Myb sites and the Etssite are required for transcriptional activation of theHNP promoter.Moreover, they suggest that the overlapping C/EBP and c-Myb sitesseem to be more important than the Ets site in positive regulation ofHNP promoter.
Binding of C/EBPa to the C/EBP site on the HNP promoterwith HL-60 nuclear extracts
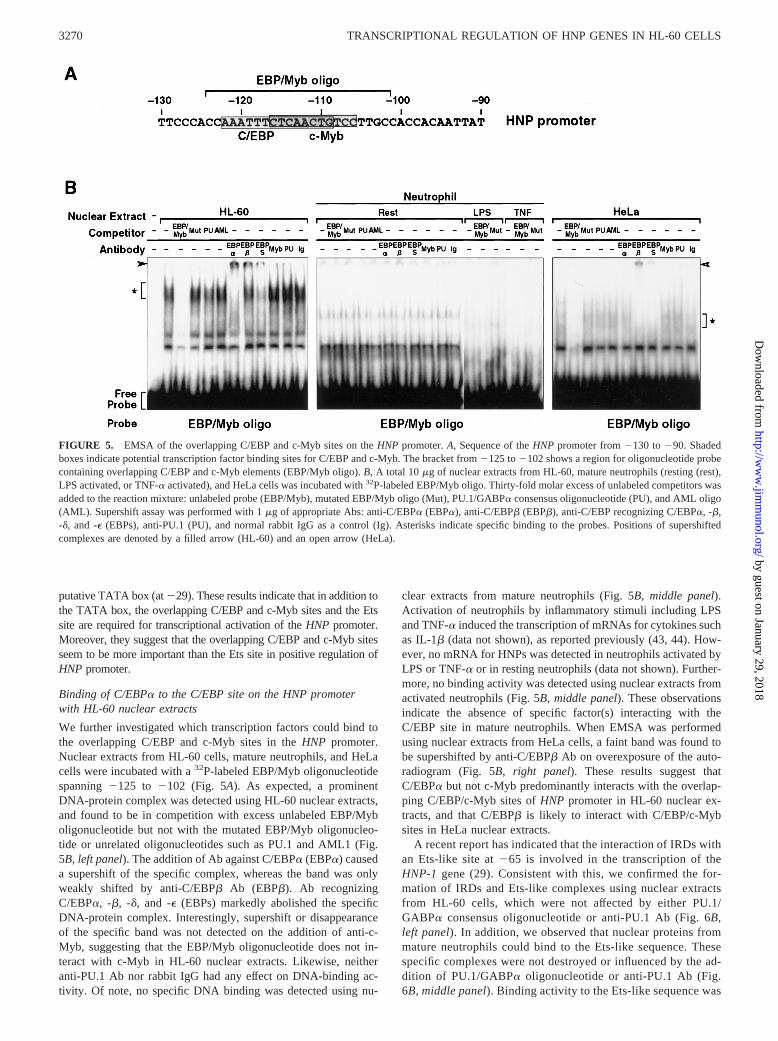
We further investigated which transcription factors could bind tothe overlapping C/EBP and c-Myb sites in theHNP promoter.Nuclear extracts from HL-60 cells, mature neutrophils, and HeLacells were incubated with a32P-labeled EBP/Myb oligonucleotidespanning2125 to 2102 (Fig. 5A). As expected, a prominentDNA-protein complex was detected using HL-60 nuclear extracts,and found to be in competition with excess unlabeled EBP/Myboligonucleotide but not with the mutated EBP/Myb oligonucleo-tide or unrelated oligonucleotides such as PU.1 and AML1 (Fig.5B, left panel). The addition of Ab against C/EBPa (EBPa) causeda supershift of the specific complex, whereas the band was onlyweakly shifted by anti-C/EBPb Ab (EBPb). Ab recognizingC/EBPa, -b, -d, and -e (EBPs) markedly abolished the specificDNA-protein complex. Interestingly, supershift or disappearanceof the specific band was not detected on the addition of anti-c-Myb, suggesting that the EBP/Myb oligonucleotide does not in-teract with c-Myb in HL-60 nuclear extracts. Likewise, neitheranti-PU.1 Ab nor rabbit IgG had any effect on DNA-binding ac-tivity. Of note, no specific DNA binding was detected using nu-
clear extracts from mature neutrophils (Fig. 5B, middle panel).Activation of neutrophils by inflammatory stimuli including LPSand TNF-a induced the transcription of mRNAs for cytokines suchas IL-1b (data not shown), as reported previously (43, 44). How-ever, no mRNA for HNPs was detected in neutrophils activated byLPS or TNF-a or in resting neutrophils (data not shown). Further-more, no binding activity was detected using nuclear extracts fromactivated neutrophils (Fig. 5B,middle panel). These observationsindicate the absence of specific factor(s) interacting with theC/EBP site in mature neutrophils. When EMSA was performedusing nuclear extracts from HeLa cells, a faint band was found tobe supershifted by anti-C/EBPb Ab on overexposure of the auto-radiogram (Fig. 5B,right panel). These results suggest thatC/EBPa but not c-Myb predominantly interacts with the overlap-ping C/EBP/c-Myb sites ofHNP promoter in HL-60 nuclear ex-tracts, and that C/EBPb is likely to interact with C/EBP/c-Mybsites in HeLa nuclear extracts.
A recent report has indicated that the interaction of IRDs withan Ets-like site at265 is involved in the transcription of theHNP-1 gene (29). Consistent with this, we confirmed the for-mation of IRDs and Ets-like complexes using nuclear extractsfrom HL-60 cells, which were not affected by either PU.1/GABPa consensus oligonucleotide or anti-PU.1 Ab (Fig. 6B,left panel). In addition, we observed that nuclear proteins frommature neutrophils could bind to the Ets-like sequence. Thesespecific complexes were not destroyed or influenced by the ad-dition of PU.1/GABPa oligonucleotide or anti-PU.1 Ab (Fig.6B, middle panel). Binding activity to the Ets-like sequence was
FIGURE 5. EMSA of the overlapping C/EBP and c-Myb sites on theHNP promoter.A, Sequence of theHNP promoter from2130 to290. Shadedboxes indicate potential transcription factor binding sites for C/EBP and c-Myb. The bracket from2125 to2102 shows a region for oligonucleotide probecontaining overlapping C/EBP and c-Myb elements (EBP/Myb oligo).B, A total 10mg of nuclear extracts from HL-60, mature neutrophils (resting (rest),LPS activated, or TNF-a activated), and HeLa cells was incubated with32P-labeled EBP/Myb oligo. Thirty-fold molar excess of unlabeled competitors wasadded to the reaction mixture: unlabeled probe (EBP/Myb), mutated EBP/Myb oligo (Mut), PU.1/GABPa consensus oligonucleotide (PU), and AML oligo(AML). Supershift assay was performed with 1mg of appropriate Abs: anti-C/EBPa (EBPa), anti-C/EBPb (EBPb), anti-C/EBP recognizing C/EBPa, -b,-d, and -e (EBPs), anti-PU.1 (PU), and normal rabbit IgG as a control (Ig). Asterisks indicate specific binding to the probes. Positions of supershiftedcomplexes are denoted by a filled arrow (HL-60) and an open arrow (HeLa).
3270 TRANSCRIPTIONAL REGULATION OF HNP GENES IN HL-60 CELLS
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

decreased in nuclear extracts from neutrophils stimulated withLPS or TNF-a (Fig. 6B, middle panel). Differences in EMSA-binding patterns between HL-60 cells and neutrophils may in-dicate that IRDs include several Ets family proteins. No suchspecific complex with the Ets-like sequence was detected innuclear extracts from HeLa cells (Fig. 6B, right panel).
Expression pattern of transcription factors in HL-60 cells andneutrophils
We analyzed levels of transcription factors likely to be involved inHNP expression by Western blot analysis (Fig. 7). Both C/EBPaand C/EBPb proteins were abundantly expressed in HL-60 nuclei(Fig. 7A). In contrast, neither C/EBPa nor C/EBPb were detectedin nuclear extracts from resting or activated neutrophils (Fig. 7A).c-Myb protein could not be detected in HL-60 and neutrophil nu-clear extracts, although c-Myb was apparent in Jurkat nuclear ex-tracts used as a positive control (Fig. 7B).
DiscussionFour different HNPs are exclusively expressed in neutrophils andcontribute to both innate and acquired immune systems (9, 24).Even though HNP-1 and HNP-3 peptides are encoded by nearlyidentical genes, their activities and contents are considerably dif-ferent (8, 16, 18). In the present study, we isolatedHNP-1 and
HNP-3promoters and investigated how each gene is transcription-ally controlled.
Our data clearly showed thatHNP-1andHNP-3promoters (1.2kbp) had the same luciferase activities in HL-60 cells despite hav-ing 15 nucleotide differences (Fig. 2B), suggesting that transcrip-tion of HNP-1 andHNP-3 genes is equal. Thus, the difference inpeptide contents between HNP-1 and HNP-3 appears to be due toposttranscriptional and/or posttranslational modifications. Onepossible explanation is that HNP-2 may be selectively produced byproteolysis of HNP-3 but not HNP-1, on the basis of earlier reportsindicating that HNP-1 content (50%) in the granules is equal to thesum of those of HNP-2 (30%) and HNP-3 (20%) (16, 24).
Interestingly, promoter activities ofHNP-1 and HNP-3 weremarkedly low in nonhematopoietic HeLa cells compared withHL-60 cells (Fig. 2B). This result suggests that transcription ofboth HNP genes is regulated in a myeloid-specific manner. Noneof the putative CCAAT boxes were required forHNP promoteractivity, and the 200-bp promoter sequence identical betweenHNP-1andHNP-3, immediately upstream region from the TATAbox, was sufficient for the promoter activity in HL-60 cells. Com-puter analysis of the proximalHNP promoter indicated the puta-tive cis-elements for C/EBP family, c-Myb, AML1, Ets family,and CDP (40, 41). Consistent with the computer prediction, EMSArevealed that HL-60 nuclear extracts contained multiple nuclear
FIGURE 6. EMSA of the Ets-like element on theHNP promoter.A, Sequences of theHNP promoter from280 to 220. Shaded boxes indicate theputative Ets-like element and TATA box. A region of oligonucleotide probe (Ets oligo) spanning284 to254 is shown by a bracket.B, A total 10mg ofnuclear extracts from HL-60, mature neutrophils (resting (rest), LPS activated, or TNF-a activated), and HeLa cells was incubated with32P-labeled Ets oligoprobe. Unlabeled oligonucleotides were added in a 25-fold molar excess as a competitor: Ets oligo (Ets), mutated Ets oligo (Mut), and PU.1/GABPa oligo(PU1). In supershift experiments, 1mg of anti-PU.1 Ab (PU) or normal rabbit IgG was added to the reaction mixture. Asterisks indicate specific bindingto the probes.
3271The Journal of Immunology
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

factors interacting with the sequences of the proximalHNP pro-moter (2214 to2124 and2146 to264) (Fig. 3C).
Sequential deletion from2240 to2133 tended to enhanceHNPpromoter activity compared with that of2240/HNP-Luc (Fig. 4).This suggests the presence of negative regulatory element(s)within the region. One potent candidate seems to be the CDP-binding sequence at2220. CDP is implicated in the transcriptionalrepression of myeloid-specific genes such as gp91phox and lacto-ferrin (45–47). It is also possible that other negative element(s)function within the region, because weak but specific binding wasobserved using the fragment (2214 to2124) lacking the CDP-binding sequence (Fig. 3C).
It is notable that the overlapping C/EBP and c-Myb sites (2122to 2106) are required for both transcription and protein binding inHL-60 cells (Figs. 4Band 5B). Our data further revealed thatC/EBPa rather than C/EBPb interacted with the C/EBP site ofHNP promoter in HL-60 cells (Fig. 5B), despite the fact that bothC/EBPa and C/EBPb were detected in HL-60 nuclear extracts(Fig. 7A). In contrast, neither C/EBPa nor C/EBPb was containedin nuclear extracts from not only resting but also activated neu-trophils (Fig. 7A). Interestingly, a feeble complex of C/EBPb withthe C/EBP site was observed using nuclear extracts from HeLacells (Fig. 5B), although HeLa cells showed nearly negligible tran-scriptional activity of theHNP promoter (Fig. 2B). However,Western blot analysis revealed that HeLa cells expressed C/EBPbbut not C/EBPa (data not shown). Taken together, these findingsindicate that C/EBPa may play an important role in the transcrip-tion of HNP genes in HL-60 cells, whereas C/EBPb is unlikely toplay a role in either HL-60 or HeLa cells despite its expression.Furthermore, c-Myb does not seem to participate inHNPpromoteractivity in HL-60 cells, because no c-Myb was detected in nuclearextracts from HL-60 cells or neutrophils (Fig. 7B).
AML1 proteins facilitate the transcriptional activity of myeloidpromoters via interaction with other adjacent transcription factors(30, 48, 49). In the HNP promoter, the consensus AML1 sequence(at 2102) lies between the C/EBP and Ets-like sites. However,disruption of the AML1 site had little effect on the promoter ac-tivity of HNPgenes in HL-60 cells (Fig. 4B). Thus, the AML1 siteis unlikely to be involved in the regulation ofHNP geneexpression.
Ma et al. (29) have reported the importance of IRD binding tothe Ets-like site at265 on theHNP promoter . We observed thatdeletion of this site caused a further 30% decrease in promoter
activity (Fig. 4B), and that IRDs interacted with the Ets-like se-quence using nuclear extracts from HL-60 cells and mature neu-trophils (Fig. 6). Furthermore, we confirmed the presence of PU.1,which is implicated in the basal transcription ofHNP genes, innuclear extracts from both HL-60 cells and neutrophils (data notshown and Ref. 25). The finding that mature neutrophils expressEts family factors including IRDs and PU.1 but not HNP mRNAsfurther support the important role of C/EBPa in the transcriptionof HNP genes in immature myeloid cells.
Human b defensins, hBD-1 and hBD-2, have been recentlyidentified from plasma and various epithelial tissues (11, 50–52).Although a defensins (HNPs) andb defensins are encoded bydifferent genes, these genes share a common evolutionary origin(53, 54). Of note, hBD-2 expression is induced by stimuli such asbacteria and proinflammatory cytokine TNF-a, thereby evokingthe important functions of hBD-2 in acute inflammation (51, 53).Future investigation of transcriptional regulation of not onlyHNPbut alsob defensingenes will afford valuable information aboutthe role of the defensin family in the innate and acquired immunesystems.
AcknowledgmentsWe thank Dr. Pernille Rorth (European Molecular Biology Laboratory) forproviding C103 anti-C/EBPa Ab and Dr. Steven L. McKnight (Universityof Texas, Southwestern Medical Center) for 472 C/EBPa antiserum.
References1. Lehrer, R. I., T. Ganz, M. E. Selsted, B. M. Babior, and J. T. Curnutte. 1988.
Neutrophils and host defense.Ann. Intern. Med. 109:127.2. Klebanoff, S. 1992. Oxygen metabolites from phagocytes. InInflammation: Basic
Principles and Clinical Correlates, 2nd Ed. J. I. Gallin, I. M. Goldstein, andR. Snyderman, eds. Raven Press, New York, p. 541.
3. Hampton, M. B., A. J. Kettle, and C. C. Winterbourn. 1998. Inside the neutrophilphagosome: oxidants, myeloperoxidase, and bacterial killing.Blood 92:3007.
4. Elsbach, P., and J. Weiss. 1992. Oxygen-independent antimicrobial systems ofphagocytes. In Inflammation: Basic Principles and Correlates, 2nd Ed.J. I. Gallin, I. M. Goldstein, and R. Snyderman, eds. Raven Press, New York,p. 603.
5. Ganz, T., and R. I. Lehrer. 1997. Antimicrobial peptides of leukocytes.Curr.Opin. Hematol. 4:53.
6. Levy, O. 1996. Antibiotic proteins of polymorphonuclear leukocytes.Eur.J. Haematol. 56:263.
7. Gabay, J. E., and R. P. Almeida. 1993. Antibiotic peptides and serine proteasehomologs in human polymorphonuclear leukocytes: defensins and azurocidin.Curr. Opin. Immunol. 5:97.
8. Ganz, T., M. E. Selsted, D. Szklarek, S. S. Harwig, K. Daher, D. F. Bainton, andR. I. Lehrer. 1985. Defensins: natural peptide antibiotics of human neutrophils.J. Clin. Invest. 76:1427.
FIGURE 7. Western blot of nuclear extracts from HL-60 cells and mature neutrophils. Nuclear extracts (10mg) from HL-60 cells and neutrophils(resting (rest), and LPS activated, or TNF-a activated) were resolved by 12% (A) or 10% (B) SDS-PAGE. Western blotting was performed usinganti-C/EBPa and anti-C/EBPb Abs (A) or anti-c-Myb Ab (B). Nuclear extracts from Jurkat cells were used as positive control inB. Positions of molecularsize markers are shown in kDa on the right. Specific bands are indicated by arrows with molecular mass (kDa).
3272 TRANSCRIPTIONAL REGULATION OF HNP GENES IN HL-60 CELLS
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

9. Lehrer, R. I., A. K. Lichtenstein, and T. Ganz. 1993. Defensins: antimicrobial andcytotoxic peptides of mammalian cells.Annu. Rev. Immunol. 11:105.
10. Lehrer, R. I., and T. Ganz. 1999. Antimicrobial peptides in mammalian and insecthost defense.Curr. Opin. Immunol. 11:23.
11. Diamond, G., and C. L. Bevins. 1998.b-Defensins: endogenous antibiotics of theinnate host defense response.Clin. Immunol. Immunopathol. 88:221.
12. Garcıa-Olmedo, F., A. Molina, J. M. Alamillo, and P. la Rodorıguez-Palenzue´.1999. Plant defense peptides.Biopolymers 47:479.
13. Hoffman, J. A., and C. Hetru. 1992. Insect defensins: inducible antibacterial pep-tides.Immunol. Today 13:411.
14. Ganz, T. 1987. Extracellular release of antimicrobial defensins by human poly-morphonuclear leukocytes.Infect. Immun. 55:568.
15. Rice, W. G., T. Ganz, J. J. Kinkade, M. E. Selsted, R. I. Lehrer, andR. T. Parmley. 1987. Defensin-rich dense granules of human neutrophils.Blood70:757.
16. Chertov, O., D. F. Michiel, L. Xu, J. M. Wang, K. Tani, W. J. Murphy,D. L. Longo, D. D. Taub, and J. J. Oppenheim. 1996. Identification of defensin-1,defensin-2, and CAP37/azurocidin as T-cell chemoattractant proteins releasedfrom interleukin-8-stimulated neutrophils.J. Biol. Chem. 271:2935.
17. Sparkes, R. S., M. Kronenberg, C. Heinzmann, K. A. Daher, I. Klisak, T. Ganz,and T. Mohandas.1989. Assignment of defensin gene(s) to human chromosome8p23.Genomics 5:240.
18. Linzmeier, R., D. Michaelson, L. Liu, and T. Ganz. 1993. The structure of neu-trophil defensin genes.FEBS Lett. 321:267.
19. Palfree, R. G., L. C. Sadro, and S. Solomon. 1993. The gene encoding the humancorticostatin HP-4 precursor contains a recent 86-base duplication and is locatedon chromosome 8.Mol. Endocrinol. 7:199.
20. Wilde, C. G., J. E. Griffith, M. N. Marra, J. L. Snable, and R. W. Scott. 1989.Purification and characterization of human neutrophil peptide 4, a novel memberof the defensin family.J. Biol. Chem. 264:11200.
21. Gabay, J. E., R. W. Scott, D. Campanelli, J. Griffith, C. Wilde, M. N. Marra,M. Seeger, and C. F. Nathan. 1989. Antibiotic proteins of human polymorpho-nuclear leukocytes.Proc. Natl. Acad. Sci. USA 86:5610.
22. Lehrer, R. I., T. Ganz, D. Szklarek, and M. E. Selsted. 1988. Modulation of thein vitro candidacidal activity of human neutrophil defensins by target cell me-tabolism and divalent cations.J. Clin. Invest. 81:1829.
23. Territo, M. C., T. Ganz, M. E. Selsted, and R. Lehrer. 1989. Monocyte-chemo-tactic activity of defensins from human neutrophils.J. Clin. Invest. 84:2017.
24. Lillard, J. W. J., P. N. Boyaka, O. Chertov, J. J. Oppenheim, and J. R. McGhee.1999. Mechanisms for induction of acquired host immunity by neutrophil peptidedefensins.Proc. Natl. Acad. Sci. USA 96:651.
25. Daher, K. A., R. I. Lehrer, T. Ganz, and M. Kronenberg. 1988. Isolation andcharacterization of human defensin cDNA clones.Proc. Natl. Acad. Sci. USA85:7327.
26. Nagaoka, I., S. Yomogida, S. Nakamura, A. Someya, K. Iwabuchi, andT. Yamashita. 1992. Evaluation of the expression of the cationic peptide gene invarious types of leukocytes.FEBS Lett. 302:279.
27. Nagaoka, I., Y. Tsutsumi-Ishii, S. Yomogida, and T. Yamashita. 1997. Isolationof cDNA encoding guinea pig neutrophil cationic antibacterial polypeptide of 11kDa (CAP11) and evaluation of CAP11 mRNA expression during neutrophilmaturation.J. Biol. Chem. 272:22742.
28. Nagaoka, I., M. Hirata, K. Sugimoto, Y. Tsutsumi-Ishii, A. Someya, K. Saionji,and J. Igari. 1998. Evaluation of the expression of human CAP18 gene duringneutrophil maturation in the bone marrow.J. Leukocyte Biol. 64:845.
29. Ma, Y., Q. Su, and P. Tempst. 1998. Differentiation-stimulated activity binds anETS-like, essential regulatory element in the human promyelocytic defensin-1promoter.J. Biol. Chem. 273:8727.
30. Friedman, A. D. 1996. Regulation of immature myeloid cell differentiation byPEBP2/CBF, Myb, C/EBP and Ets family members.Curr. Top. Microbiol. Im-munol. 211:149.
31. Clarke, S., and S. Gordon. 1998. Myeloid-specific gene expression.J. LeukocyteBiol. 63:153.
32. Tenen, D. G., R. Hromas, J. D. Licht, and D. E. Zhang. 1997. Transcriptionfactors, normal myeloid development, and leukemia.Blood 90:489.
33. Pahl, H. L., T. C. Burn, and D. G. Tenen. 1991. Optimization of transient trans-fection into human myeloid cell lines using a luciferase reporter gene.Exp. He-matol. 19:1038.
34. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983. Accurate transcriptioninitiation by RNA polymerase II in a soluble extract from isolated mammaliannuclei.Nucleic Acids Res. 11:1475.
35. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4.Nature 227:680.
36. Radomska, H. S., C. S. Huettner, P. Zhang, T. Cheng, D. T. Scadden, andD. G. Tenen. 1998. CCAAT/enhancer binding protein alpha is a regulatory switchsufficient for induction of granulocytic development from bipotential myeloidprogenitors.Mol. Cell. Biol. 18:4301.
37. Scott, L. M., C. I. Civin, P. Rorth, and A. D. Friedman. 1992. A novel temporalexpression pattern of three C/EBP family members in differentiating myelomono-cytic cells.Blood 80:1725.
38. Landschulz, W. H., P. F. Johnson, E. Y. Adashi, B. J. Graves, andS. L. McKnight. 1988. Isolation of a recombinant copy of the gene encodingC/EBP.Genes Dev. 2:786.
39. Birkenmeier, E., B. Gwynn, S. Howard, J. Jerry, J. Girdon, W. Landschulz, andS. L. McKnight. 1989. Tissue-specific expression, developmental regulation, andgenetic mapping of the gene encoding CCAAT/enhancer binding protein.GenesDev. 3:1146.
40. Quandt, K., K. Frech, H. Karas, E. Wingender, and T. Werner. 1995. MatInd andMatInspector: new fast and versatile tools for detection of consensus matches innucleotide sequence data.Nucleic Acids Res. 23:4878.
41. Heinemeyer, T., X. Chen, H. Karas, A. E. Kel, O. V. Kel, I. Liebich,T. Meinhardt, I. Reuter, F. Schacherer, and E. Wingender. 1999. Expanding theTRANSFAC database towards an expert system of regulatory molecular mech-anisms.Nucleic Acids Res. 27:318.
42. Lenny, N., J. J. Westendorf, and S. W. Hiebert. 1997. Transcriptional regulationduring myelopoiesis.Mol. Biol. Rep. 24:157.
43. Cassatella, M. A., S. Gasperini, F. Calzetti, P. P. McDonald, and G. Trinchieri.1995. Lipopolysaccharide-induced interleukin-8 gene expression in human gran-ulocytes: transcriptional inhibition by interferon-g. Biochem. J. 310:751.
44. Palma, C., A. Cassone, D. Serbousek, C. A. Pearson, and J. Y. Djeu. 1992.Lactoferrin release and interleukin-1, interleukin-6, and tumor necrosis factorproduction by human polymorphonuclear cells stimulated by various lipopoly-saccharides: relationship to growth inhibition ofCandida albicans. Infect. Im-mun. 60:4604.
45. Khanna, G. A., T. Zibello, S. Kolla, E. J. Neufeld, and N. Berliner. 1997. CCAATdisplacement protein (CDP/cut) recognizes a silencer element within the lacto-ferrin gene promoter.Blood 90:2784.
46. Luo, W., and D. G. Skalnik. 1996. CCAAT displacement protein competes withmultiple transcriptional activators for binding to four sites in the proximalgp91phox promoter.J. Biol. Chem. 271:18203.
47. Lievens, P. M., J. J. Donady, C. Tufarelli, and E. J. Neufeld. 1995. Repressoractivity of CCAAT displacement protein in HL-60 myeloid leukemia cells.J. Biol. Chem. 270:12745.
48. Britos-Bray, M., and A. D. Friedman. 1997. Core binding factor cannot syner-gistically activate the myeloperoxidase proximal enhancer in immature myeloidcells without c-Myb.Mol. Cell. Biol. 17:5127.
49. Westendorf, J. J., C. M. Yamamoto, N. Lenny, J. R. Downing, M. E. Selsted, andS. W. Hiebert. 1998. The t(8;21) fusion product, AML-1-ETO, associates withC/EBP-a, inhibits C/EBP-a-dependent transcription, and blocks granulocytic dif-ferentiation.Mol. Cell. Biol. 18:322.
50. Bensch, K. W., M. Raida, H. J. Magert, P. Schulz-Knappe, and W. G. Forssmann.1995. hBD-1: a novelb-defensin from human plasma.FEBS Lett. 368:331.
51. Harder, J., J. Barteks, E. Christophers, and J.-M. Schrodeer. 1997. A peptideantibiotic from human skin.Nature 387:861.
52. Zhao, C. Q., I. Wang, and R. I. Lehrer. 1996. Widespread expression ofb-de-fensin HBD-1 in human secretory glands and epithelial cells.FEBS Lett. 396:319.
53. Liu, L., L. Wang, H. P. Jia, C. Zhao, H. H. Q. Heng, B. C. Schutte,P. B. McCray Jr., and T. Ganz. 1998. Structure and mapping of the humanb-defensin HBD-2 gene and its expression at sites of inflammation.Gene 222:237.
54. Liu, L., C. Zhao, H. H. Heng, and T. Ganz. 1997. The humanb-defensin-1 anda-defensins are encoded by adjacent genes: two peptide families with differingdisulfide topology share a common ancestry.Genomics 43:316.
3273The Journal of Immunology
by guest on January 29, 2018http://w
ww
.jimm
unol.org/D
ownloaded from