role of a sodium-dependent symporter homologue in …aac.asm.org/content/52/2/505.full.pdf ·...
TRANSCRIPT

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Feb. 2008, p. 505–512 Vol. 52, No. 20066-4804/08/$08.00�0 doi:10.1128/AAC.00504-07Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Role of a Sodium-Dependent Symporter Homologue in theThermosensitivity of �-Lactam Antibiotic Resistance and
Cell Wall Composition in Staphylococcus aureus�
Krzysztof Sieradzki, Marilyn Chung, and Alexander Tomasz*The Rockefeller University, 1230 York Avenue, New York, New York 10021
Received 13 April 2007/Returned for modification 2 July 2007/Accepted 15 November 2007
Expression of high-level �-lactam resistance is known to be thermosensitive in many methicillin-resistantStaphylococcus aureus (MRSA) strains, including strain COL, in which the high methicillin MIC for cultures grownat 37°C (800 �g/ml) was reduced to 12 �g/ml at 42°C. COL grew faster at 42°C than at 37°C and at the highertemperature produced cell walls of abnormal composition: there was an over-representation of the monomericmuropeptide without the oligoglycine chain and an increase in the representation of multimers that contained thiswall component as the donor molecule. Screening of a Tn551 insertional library for mutants, in which the high andhomogenous �-lactam antibiotic resistance of strain COL is retained at 42°C, identified mutant C245, whichexpressed high-level methicillin resistance and produced a cell wall of normal composition independent of thetemperature. The Tn551 inactivated gene was found, by homology search, to encode for a sodium-dependentsymporter, homologues of which are ubiquitous in both prokaryotic and eukaryotic genomes. Inactivation of thisputative symporter in several heteroresistant clinical MRSA isolates caused striking increases in the level of their�-lactam resistance.
The �-lactam-resistant phenotype of methicillin-resistantStaphylococcus aureus (MRSA) strains is known to dependsensitively not only on genetic determinants of resistance (i.e.,expression of the resistance gene mecA and the functionality ofthe so-called auxiliary genes) (10, 12, 17, 19) but also on theconditions of growth, such as the temperature, pH, and saltconcentration during challenge with the antibiotics (3, 5, 24,25). Reduced temperature of incubation is used routinely inmost diagnostic microbiological laboratories in order to opti-mize detection of MRSA in clinical specimens (6). How theseenvironmental factors affect the activity and/or expression ofthe genetic elements associated with �-lactam resistance is notknown, although in the case of higher temperatures, it hasbeen suggested that the observed reduced resistance levelsmay be related to decreased amounts of PBP2A in the cells(16, 22, 27).
Strain COL is one of the MRSA strains in which the level of�-lactam resistance is extremely temperature dependent: pre-liminary tests showed that growth of COL at 42°C, whichcaused a nearly 100-fold decrease in the methicillin MIC,caused only a minor reduction in the transcription of mecA. Inthe studies described here, we used strain COL as a model tobetter understand the mechanism of the temperature sensitiv-ity of antibiotic resistance.
MATERIALS AND METHODS
Bacterial strains and growth conditions. The laboratory S. aureus constructsand plasmids used in the present study are listed in Table 1. All S. aureus strains
were grown in tryptic soy broth (TSB; Difco, Detroit, MI) at 37 and 42°C withaeration. E. coli transformants were grown in Luria-Bertani medium (Difco) at37°C supplemented with 100 �g of ampicillin/ml. The growth medium of thetransposon mutant C245 and its backcrosses and complementation mutants wassupplemented with erythromycin (10 �g/ml) and chloramphenicol (25 �g/ml),respectively. Growth was followed by monitoring the absorbance (A620) using anLKB spectrophotometer (Pharmacia LKB Biotechnology, Inc., Sweden).
Determination of antibiotic susceptibility. Determination of antibiotic suscep-tibility was done by plating diluted overnight cultures on petri dishes containingtryptic soy agar (TSA; Difco) and serial (twofold) dilutions of the appropriateantibiotic according to a population analysis method described previously (26).Antibiotic MICs for the majority of cells were calculated as the lowest concen-tration of the antibiotic causing a 99.9% loss of the inoculum.
Isolation of total RNA and Northern blot hybridization. Overnight cultureswere inoculated into fresh TSB where they were grown to mid-log phase (A620 �0.6). Before being harvested, bacterial cells were stabilized for 10 min withRNAprotect bacteria reagent (Qiagen Gmbh, Hilden, Germany), and RNA wasextracted by using a FastRNA Blue isolation kit (Bio 101, Vista, CA) accordingto the manufacturer’s recommendations. After the concentration was adjustedwith a GeneQuant spectrophotometer (Pharmacia), RNA samples (5 �g)were resolved by electrophoresis on 1.2% agarose–0.66 M formaldehyde gelsin morpholinepropanesulfonic acid running buffer. Blotting of RNA onto aHybond N� membrane (Amersham, Arlington Heights, IL) was performedwith the Turbo Blotter neutral transfer system (Schleicher & Schuell, Keene,NH). For detection of transcripts, DNA probes corresponding to internalfragments of particular gene, amplified by using the GeneAmp PCR reagentkit with AmpliTaq DNA polymerase (Perkin-Elmer) and purified by using theQIAquick PCR purification kit (Qiagen), were labeled with [�-32P]dCTP bythe random prime method using a Ready-to-Go labeling kit (Amersham) andhybridized under high-stringency conditions. The blots were subsequentlywashed and autoradiographed.
Mutagenesis and selection of Tn551 mutants. The transposition experi-ment and selection of mutants were performed as described before (9). Theparental strain COL harboring the thermosensitive plasmid pRN3208 (car-rying Tn551 with an erythromycin resistance determinant) was grown over-night at 30°C and then diluted and plated at different concentrations on TSAcontaining erythromycin (100 �g/ml). The plates were incubated at 42°C for76 h. The approximate frequency of transposon insertion was 3 � 10�5. Inorder to cure cells from residual plasmid, bacteria were cultured for 48 h at42°C. Only erythromycin-resistant and cadmium-susceptible colonies wereused for further study.
* Corresponding author. Mailing address: The Rockefeller Univer-sity, Laboratory of Microbiology, 1230 York Ave., New York, NY10021. Phone: (212) 327-8277. Fax: (212) 327-8688. E-mail: [email protected].
� Published ahead of print on 3 December 2007.
505
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

Screening for mutants with altered methicillin resistance. The erythromycin-resistant, cadmium-sensitive colonies were tested sequentially in three stages. Inthe first screen, all colonies were streaked onto TSA plates containing erythro-mycin (10 �g/ml) and different concentrations of methicillin (0, 25, 50 and 100�g/ml). Strain COLpRN3208 was used as control. In the second screen, coloniesthat seemed to have altered methicillin resistance were grown overnight in TSBwith erythromycin (10 �g/ml), diluted (10�2), and tested on TSA plates contain-ing a 1.0-mg methicillin disk. After 24 h of incubation at 42°C the halos ofinhibition were measured. After this preliminary testing for altered resistance, athird screen was used to more precisely determine the methicillin resistancephenotype of mutants by using population analysis (26).
Transductional crosses and analysis of transductants. Transductionalcrosses were performed with phage 80� as described previously (20) using asa recipient the parental strain COL (cured from the plasmid pRN3208) andas a donor the newly isolated transposon mutant COL245. Transductantswere selected on plates that contained erythromycin at a final concentrationof 10 �g/ml. From the cross, 25 transductants were tested for increased levelsof methicillin resistance by the 1.0-mg methicillin disk method. Eventually,two transductants were selected for testing by PAP analysis for their antibi-otic resistance phenotypes.
Peptidoglycan preparation and analysis. Cell wall peptidoglycan was pre-pared, and muropeptide composition of peptidoglycan was analyzed by reversed-phase high-performance liquid chromatography (HPLC) as described previously(7), except that the alkaline phosphatase step was omitted.
Autolysis assay. Triton X-100-stimulated autolysis in glycine buffer (pH 8.0)was measured as previously described (8). Cells were grown exponentially to anabsorbance at 620 nm (A620) of about 0.3. The cultures were then rapidly chilled,and the cells were washed once with ice-cold distilled water and suspended to anoptical density at 620 nm of 1.0 in 50 mM glycine buffer supplemented with0.01% Triton X-100. Autolysis was measured during incubation at 37°C as thedecrease in A620 by using a model 340 spectrophotometer (Sequoia-TurnerCorp., Mountain View, CA).
DNA methods. Chromosomal DNA preparation and manipulations were per-formed by standard methods (4). Restriction enzymes were used as recom-mended by the manufacturer (New England Biolabs, Beverly, MA). DNA se-quencing was done at the Rockefeller University Protein/DNA TechnologyCenter by the BigDye terminator cycle sequencing method with either a 3700DNA analyzer for capillary electrophoresis or ABI Prism 377 DNA sequencersfor slab gel electrophoresis.
Sequencing and identification of Tn551 inactivated region. Inverse PCR wasperformed to isolate DNA region flanking the Tn551 insertion site, as previouslydescribed (28).
The chromosomal DNA preparations were digested with HindIII to comple-tion. The self-ligation of the HindIII digests was performed at a DNA concen-tration of 2 �g/ml overnight at 4°C, and the ligation mixture was then used as atemplate DNA for PCR amplification. The primer pairs were the same asdescribed before (11). DNA sequencing was performed at the Rockefeller Uni-versity Protein/DNA Technology Center with the Taq fluorescent dye terminatorsequencing method using a Perkin-Elmer/Applied Biosystems model 377 auto-mated sequencer. The obtained open reading frames and deduced amino acidsequences were analyzed with DNASTAR software and compared to the knownpeptides from the TIGR (The Institute for Genomic Research) and GenBankdatabases by using the BLAST algorithm.
Sequence analysis of smr1 (TIGR COL locus SA0501). Based on the prelim-inary sequence obtained, DNA fragments containing smr1 (for suppressor ofmethicillin resistance) were amplified from the chromosomal DNA of the pa-rental strain COL by using the primers KS-smr-F0 (5�-TGGCGC AAC ACTATC CTT-3�) and KS-smr-R0 (5�-TTC GGG TCC CAA TGT ATG A-3�) under
the following conditions: 94°C for 5 min, followed by 30 cycles of 94°C for 30 s,52°C for 30 s, and 72°C for 4 min, with a final extension step of 72°C for 6 min.
Complementation of mutant C245. A 2.6-kb fragment containing smr1 wasamplified from the COL chromosome with the primers KS-smr-XbaI-PF
FIG. 1. Effect of growth temperature on the phenotypic expressionof methicillin resistance (A) and growth rate (B) in strain COL. Forthe determination of methicillin resistance, cultures grown overnight inTSB were plated at different cell concentrations on agar plates con-taining twofold concentrations of methicillin. Colonies were countedafter 48 h of incubation at 37 and 42°C. Growth rates were measuredin TSB cultures, which were monitored by determining the A620.
TABLE 1. Laboratory derivatives of strain COL and plasmids
Strain Relevant propertiesa Source or reference
COL Homogeneous Mcr at 37°C, heterogeneous Mcr at 42°C, Ems mecA RU collectionCOLmec- COL with removed SCCmec cassette 21COLmec-245 COLmec- (mutagenized smr1::Tn551) Mcs Emr This studyCOLpRN3208 COL with pRN3208 (Tn551) Rep(Ts); Mcr Emr Cd r 15C245 COL (mutagenized smr1::Tn551), homogeneous Mcr at 42°C, Emr This studyC245Td COL (smr1::Tn551, backcross from C245), homogeneous Mcr at 42°C, Emr This studyC245KS20 C245Td with pGCKS20; heterogeneous Mcr at 42°C, Emr Cmr This studypGC2 E. coli-S. aureus shuttle vector, Apr Cmr Constructed by P. MatthewspGCKS20 pGC2 carrying XbaI-AvaI-restricted 2.5-kb smr1 insert This study
a Abbreviations: Mc, methicillin; Emr, erythromycin; Cd, cadmium; Cm, chloramphenicol; Ap, ampicillin. Superscripts: r, resistant; s, sensitive.
506 SIERADZKI ET AL. ANTIMICROB. AGENTS CHEMOTHER.
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

(5�-GTA TCA CTG TCT CTA GAG CAC CAA AGG AAA AG-3�; under-lined letters indicate the introduced restriction site) and KS-smr-PR (5�-CTGACG CGG ACC CTA TGC CTG TAT TA-3�) using a High-Fidelity PCRSystem (Roche Diagnostics, Indianapolis, IN). The amplified smr1 fragmentand shuttle vector pGC2 were digested with AvaI and XbaI and ligated withT4 DNA ligase (Roche Diagnostics), generating pGCKS20. Cloning wasperformed in Escherichia coli XL1-Blue (Stratagene, La Jolla, CA), and the
suitable plasmid was selected from among the ampicillin-resistant transfor-mants by its molecular size verification and PCR analysis; one transformantwas picked, and the amplified replicative plasmid pGCKS20 was then intro-duced into RN4220 by electroporation (29) and subsequently transferred toC245 by transduction, generating C245KS20. For selection, ampicillin (100�g/ml) was used in E. coli and chloramphenicol (25 �g/ml) was used in S.aureus.
FIG. 2. HPLC elution muropeptide profiles of strain COL grown at 42°C (A) and 37°C (B). Separation of S. aureus muropeptides was carriedout by reversed-phase HPLC. Muropeptides were prepared and analyzed after reduction, as described previously (7). The chemical structuresassigned to the muropeptides have been published elsewhere (7).
FIG. 3. Schematic representation of the Tn551 insertion site in the disrupted open reading frame in strain C245. Genomic DNA fragments,containing the investigated gene, after self-ligation of the HindIII digests were amplified by the PCR and then sequenced. The lower part of thefigure shows the amino acid sequence deduced from the nucleotide sequence. The orientation of the open reading frame is indicated by thearrowhead.
VOL. 52, 2008 SODIUM-DEPENDENT SYMPORTER HOMOLOGUE IN S. AUREUS 507
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
RESULTS AND DISCUSSION
Effect of temperature on �-lactam resistance, growth rate,and cell wall composition. Overnight cultures of strain COLwere plated for population analysis and incubated at either 37or 42°C to evaluate the effect of incubation temperature on theexpression of methicillin resistance. The increase in incubationtemperature caused major and multiple alterations in theproperties of strain COL. The high-level methicillin resistanceat 37°C (MIC � 800 �g/ml) was reduced to 12 �g/ml at 42°Cand was accompanied by a change from a homogeneous to aheterogeneous population structure (Fig. 1A). The growth rateof the culture increased (Fig. 1B), and there were major alter-ations in the muropeptide composition of the cell wall pepti-doglycan (Fig. 2).
Bacterial cells grown at 42°C overproduced muropeptide 1,a disaccharide pentapeptide monomer free of the pentaglycinebranch, which is normally present in very limited relativeamounts in most staphylococcal muropeptides. In addition,there was also an increase in the proportion of multimers,
which contained this glycine-free muropeptide as the originaldonor molecule (see peaks 9, 11, and 14 through 18 in Fig. 2A).
The loss of resistance appeared to be specific for �-lactaminhibitors: cultures grown at 37°C versus 42°C had the sameMICs of bacitracin (50 �g/ml), D-cycloserine (50 �g/ml), andtetracycline (100 �g/ml). There was slight increase in the van-comycin MIC (from 1.5 to 3.0 �g/ml) and decrease in thefosfomycin MIC (from 50 to 25 �g/ml) at the higher temper-ature.
These observations do not allow one to establish a causalrelationship among the multiple changes that accompany theshift in the incubation temperature from 37 to 42°C. However,the increased growth rate at 42°C in parallel with the decreasedlevel of resistance was reminiscent of the “fitness cost” (slowergrowth rate) described by several investigators in bacteria car-rying genetic determinants of antibiotic resistance (2). Thisprompted us to search for transposon mutants in which thetemperature dependence of methicillin resistance and growthrate was altered.
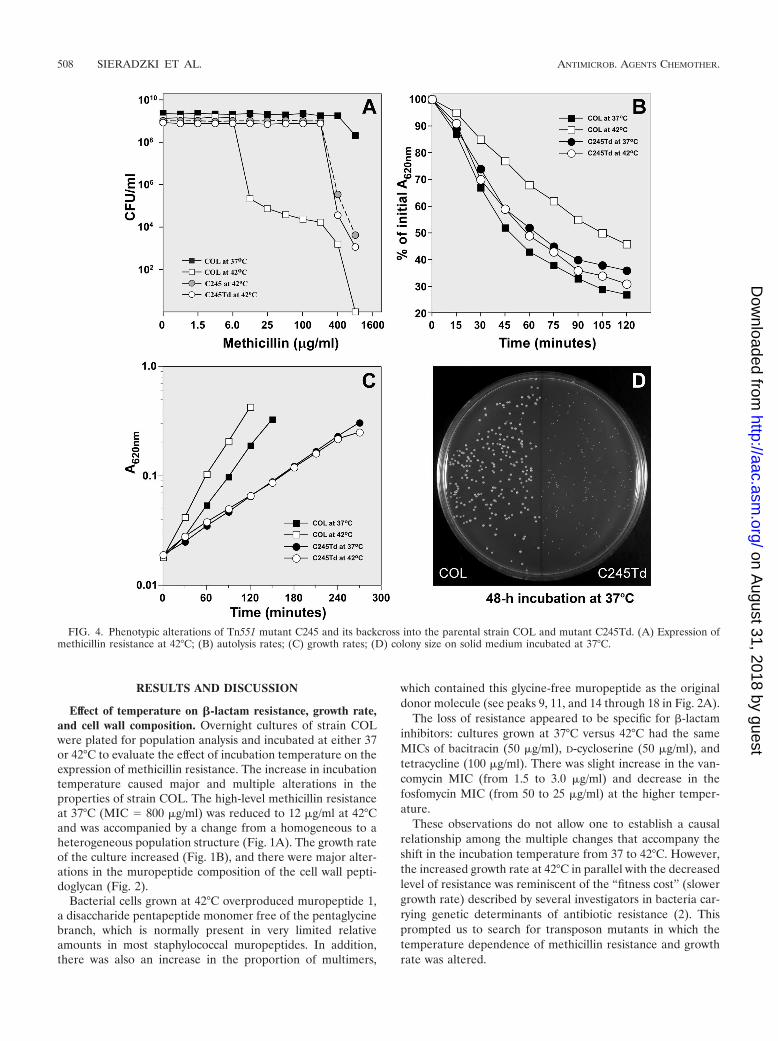
FIG. 4. Phenotypic alterations of Tn551 mutant C245 and its backcross into the parental strain COL and mutant C245Td. (A) Expression ofmethicillin resistance at 42°C; (B) autolysis rates; (C) growth rates; (D) colony size on solid medium incubated at 37°C.
508 SIERADZKI ET AL. ANTIMICROB. AGENTS CHEMOTHER.
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

Selection for Tn551 mutants. Strain COL harboring thethermosensitive plasmid pRN3208 (carrying Tn551 with theerythromycin resistance determinant) was used to generate ashotgun library of Tn551 mutants as described in Materials andMethods. The library was then screened for mutants that wereable to express high-level and homogeneous methicillin resis-tance at 42°C. Transposon mutant C245, exhibiting this phe-notype, was identified among over a thousand initially screenedTn551 inserts. Mutant C245 grown at either 37 or 42°C pro-duced homogeneously resistant cultures with methicillin MICsno lower than 400 �g/ml. Backcrossing of the Tn551-inacti-vated gene into the parental strain COL produced transduc-tant C245Td, which expressed a resistance phenotype virtuallyidentical to that of the original transposon mutant.
Sequencing and identification of the suppressor of methi-cillin resistance (smr1). The genetic determinant in mutantC245 was amplified and sequenced, and the obtained openreading frame and the deducted amino acid sequence (Fig. 3)were compared to sequences of known polypeptides in theTIGR and GenBank databases by using the BLAST algorithm.The homology search in the TIGR database for the S. aureusstrain COL yielded (with 100% peptide similarity) identifica-tion of the locus SA0501. Further search in the GenBankpeptide databases, based on the statistical significance of se-quence similarity, showed that the protein in question showedsignificant homology with the large neurotransmitter sodiumsymporter family, homologues of which can be found amongboth eukaryotes and prokaryotes (13).
Since SA0501 seems to play a negative role in terms ofmethicillin resistance, we propose to name this previously un-characterized gene suppressor of methicillin resistance 1(smr1).
Phenotype of the C245 transposon mutant. The methicillinresistance profile of strain COL and its transductant derivativecarrying the C245 mutation was tested at 37 and 42°C in par-allel with the rates of autolysis and growth rates of thesestrains at the two temperatures. The methicillin resistanceprofiles of C245 and C245Td transductant showed virtually
identical high-level and homogeneous resistance at bothtemperatures (Fig. 4A).
Figure 4B shows that autolysis—a property closely associ-ated with the response of bacteria to �-lactam antibiotics—wasalso altered in the C245Td transductant. Cells of strain COLgrown at 42°C (and tested at 37°C) autolyzed slower than cellsgrown at 37°C. Autolysis rates were relatively fast and wereidentical for cultures of C245Td mutant grown at both tem-peratures.
The C245 transductant showed identical slow growth rates atboth temperatures (Fig. 4C). The slow growth of the transduc-tant C245Td was also evidenced by the small size of its colonieson solid medium (Fig. 4D).
In contrast to the parental strain COL, C245Td producedcell walls of identical (i.e., normal) muropeptide profiles atboth temperatures, and the representation of the glycine-freemuropeptide monomer and its oligomeric derivatives was re-duced to the levels seen in COL grown at 37°C (Fig. 5).
Thus, each of the phenotypes that were temperature depen-dent in the parental strain COL became temperature indepen-dent in the transposon mutant. Although these observations donot allow the sorting out of the pleiomorphic phenotypes intoa sequential order, the critical role of the smr1 gene is quiteevident.
Effects of temperature shift on transcription levels of mecA,pbpB, mtgA, and femX. Several previous studies suggested thatthe reduced levels of methicillin resistance in cultures grown atelevated temperatures might be related to the lowered produc-tion of PBP2A and/or PBP2 at higher temperatures (16, 22,27). We tested by Northern analysis the transcription of mecA,pbpB, mtgA, and femX—four genes that are involved with the�-lactam resistance phenotypes in S. aureus—in strain COLand its C245 transductant grown at either 37 or 42°C (Fig. 6).A modest reduction in the signal intensity for each gene wasdetectable in the COL samples grown at 42°C. However, thesame difference was also seen in mutant C245 grown at thesame temperatures, although the resistant phenotype in C245is no longer temperature dependent.
FIG. 5. (A) Normalized HPLC elution muropeptide profile of strain C245Td grown at 42°C. (B) As a control, peptidoglycan of the same straingrown at 37°C was also analyzed.
VOL. 52, 2008 SODIUM-DEPENDENT SYMPORTER HOMOLOGUE IN S. AUREUS 509
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

Temperature-dependent changes in the muropeptide com-position of an isogenic methicillin-susceptible derivative ofstrain COL. The SCCmec type I cassette was removed fromstrain COL by the method of Katayama et al. (14) to generateCOLmec- (21). Incubation of the strain at 37 and 42°C repro-duced the same effects on growth rate and cell wall composi-tion, as already documented for the methicillin-resistantparent strain COL. HPLC elution profiles of enzymatic pepti-doglycan digests from COLmec- showed temperature-depen-dent differences in muropeptide composition identical to thoseshown in Fig. 2 for strain COL. Also, similar to the what weobserved with strain COL, the C245 transductant of COLmec-
produced peptidoglycan of identical (normal) composition atboth temperatures (Fig. 7). These observations make it evenless likely that the temperature-dependent changes observed inresistance level involved changes in the transcription and/ortranslation of the resistance gene mecA.
Impact of smr1 inactivation on the expression of oxacillinresistance in clinical heteroresistant MRSA isolates with dif-ferent genetic backgrounds. Inactivation of smr1 in the back-ground of COL caused two prominent effects: (i) eliminationof the temperature dependence of growth rate and (ii) anincrease in the methicillin MIC. Table 2 summarizes the resultsof a screen in which several clinical MRSA strains with differ-
FIG. 6. Transcription levels of mecA, pbpB, femX, and mtgA in theparental strain COL and the Tn551 mutant C245 grown at 37 and42°C. mRNA was extracted from mid-log-phase cultures, resolved byelectrophoresis on agarose-formaldehyde gels and then located afterhybridization with 32P-labeled DNA probes as described in Materialsand Methods.
FIG. 7. HPLC elution profiles of muropeptides isolated from strains COLmec- and COLmec-245 grown at 42°C (A and C) and 37°C (B and D),respectively.
TABLE 2. Effect of inactivation of the symporter gene smr1 on theexpression of oxacillin resistance in MRSA strainsa
Strain MLST SCCmectype
Clonalcomplex
37°C 42°C
Growth MIC Growth MIC
COL 250 I 8 �� �256 ��� 6.0C245Td 250 I 8 � �256 � �256D19455 247 I 8 �� 1.0 ��� 0.25D19455-T2 247 I 8 � 64 � 0.25E2125 247 I 8 �� 1.0 ��� 0.25E2125-T2 247 I 8 � 128 � 0.50E3005 247 I 8 �� 1.5 ��� 0.50E3005-T4 247 I 8 � 12 � 0.38MW2 1 IV 1 �� 0.75 ��� 0.75MW2-T 1 IV 1 � 8.0 � 0.75301 239 III 8 �� �256 ��� 1.0301-T6 239 III 8 � �256 � 16319 247 I 8 �� �256 ��� 0.75319-T1 247 I 8 � �256 � �256404 247 I 8 �� �256 ��� 1.0404-T3 247 I 8 � �256 � �256RUSA11 8 I 8 �� 0.50 ��� 0.19RUSA11-T3 8 I 8 � 4.0 � 0.25
a MICs were determined from the E-test results after 24 h of incubation at theindicated temperatures.
510 SIERADZKI ET AL. ANTIMICROB. AGENTS CHEMOTHER.
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

ent geographic origins, isolation dates, and genetic back-grounds were compared for their growth rates and oxacillinMICs when grown at 37 or 42°C. All but one of the strainstested belonged to clonal complex 8. In parallel, we analyzedtransductants of the same strains, in which smr1 was inacti-vated. Although the degrees of increase in the oxacillin MICsof the transductants clearly varied from strain to strain, im-proved resistance and elimination of the temperature depen-dence of growth rates was evident in each transductant. TheHPLC analysis of strains 319 and 404 revealed that, as in COL,inactivation of smr1 also caused a normalization of the cell wallmuropeptide composition produced by these strains at 42°C(data not shown).
Our results show that the central genetic determinant ofmethicillin resistance—the mecA gene—is only modestlydownregulated when COL cultures are grown at 42°C. A sim-ilar drop in the mecA transcription level could also be observedin the resistant mutant C245. Thus, in this case, the slightlydecreased cellular amount of PBP2A is not likely to accountfor the drastic reduction of the methicillin resistance in strainCOL. The same is true for the other tested genes that poten-tially contribute to methicillin resistance (pbpB, femX, andmtgA): they remain unaltered, and their transcription levels arecomparable for both the parental strain COL and the resistantmutant C245.
When trying to answer the question of what is the mecha-nism of the reduction of methicillin resistance in Staphylococ-cus aureus at 42°C, one has to remember that this is a temper-ature that already triggers the heat shock response, inducingmultiple transcriptional changes, which may, in turn, causeextensive metabolic alterations in the bacterial cells. Andersonet al. (1) found that heat-shocked S. aureus cells (at 42°C)
overexpressed 98 genes at the expense of 42 other genes thatwere downregulated.
As for the role of the inactivated putative symporter inantibiotic resistance, one may consider two alternatives. (i)The putative symporter may catalyze the uptake of an impor-tant amino acid, or related nitrogenous compounds, utilized bythe bacterial cells. Inactivation of that gene, vital for the inter-mediary metabolism, would slow down bacterial growth, whichmay allow bacterial cells to take full advantage of their slow-paced resistance protein PBP2A. Otherwise, in fast-growingcells, PBP2A, as a low-affinity penicillin-binding protein/poorcatalyst for the transpeptidation reaction, may lag behind otherenzymes participating in cell wall biosynthesis. This possibilityis suggested by the fact that the substantially improved resis-tance levels of the symporter transductants (originating fromthe fast-growing and only modestly resistant clinical MRSAisolates) could be achieved even at low temperatures.
(ii) Alternatively, the putative symporter may be involved inthe sensing and global transcriptional response to the temper-ature shifts and, as such, when inactivated, it would not bepossible for bacterial cells to switch their metabolism in re-sponse to the changing temperature. In this case, the expres-sion of factors involved in cell wall biosynthesis (�-lactam re-sistance) would continue undisturbed. The “corrected”features of the symporter mutants, i.e., methicillin resistance,cell wall composition, and growth and autolysis rates, which arephenotypic manifestations of complex interactions betweennumerous genes involved in cell wall synthesis and metabolism,seem to confirm this hypothesis. This scenario seems to bequite possible since some sodium-dependent symporters areknown components of the regulatory feedback control systems.For instance, an Na�/glycine betaine symporter was found to
FIG. 8. Effect of smr1complementation on oxacillin resistance of MRSA strain C245Td. Bacterial cultures grown overnight in TSB (COL), TSBsupplemented with 10 �g of erythromycin/ml (C245Td), or 10 �g of chloramphenicol/ml (C245KS20) were diluted to approximately 5 � 107
CFU/ml, swabbed onto TSA plates, and allowed to dry. Oxacillin E-test strips were then applied to the agar surface. The inhibitory effects ofoxacillin on the growth of COL (A), C245Td (B), and C245KS20 (C) were determined after 18 h of incubation at 42°C.
VOL. 52, 2008 SODIUM-DEPENDENT SYMPORTER HOMOLOGUE IN S. AUREUS 511
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from

be involved with the protection of Listeria monocytogenesagainst osmotic and thermal shock (18). The accumulation ofglycine betaine may lead to a regulatory feedback: the Na�/glycine betaine uptake carrier BetP from Corynebacterium glu-tamicum has recently been shown to act both as an osmoreg-ulator and directly as an osmosensor (23).
Whatever the cellular function(s) of the sodium-dependentsymporter homologue may turn out to be in S. aureus, it isbeyond speculation that alterations in smr1 functionality are atleast in part responsible for the methicillin resistance pheno-type in MRSA strains. This conclusion is confirmed by theresults of the complementation experiment illustrated in Fig. 8.Complementation of the C245Td mutant with plasmid-en-coded smr1 literally abolished its high level of �-lactam resis-tance at 42°C and restored the parent-like (COL) phenotype.
Experiments are currently in progress to clarify the mecha-nism of action of the symporter in �-lactam resistance and cellwall synthesis.
ACKNOWLEDGMENT
This study was supported in part by a grant 2 RO1AI045738 fromthe U.S. Public Health Service.
REFERENCES
1. Anderson, K. L., C. Roberts, T. Disz, V. Vonstein, K. Hwang, R. Overbeek,P. D. Olson, S. J. Projan, and P. M. Dunman. 2006. Characterization of theStaphylococcus aureus heat shock, cold shock, stringent, and SOS responsesand their effects on log-phase mRNA turnover. J. Bacteriol. 188:6739–6756.
2. Andersson, D. I., and B. R. Levin. 1999. The biological cost of antibioticresistance. Curr. Opin. Microbiol. 2:489–493.
3. Annear, D. I. 1968. The effect of temperature on resistance of Staphylococcusaureus to methicillin and some other antibiotics. Med. J. Aust. 1:444–446.
4. Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, J. G. Seidman, J. A.Smith, and K. Struhl (ed.). 1995. Current protocols in molecular biology, p.241–242. John Wiley & Sons, Inc., New York, NY.
5. Barber, M. 1964. Naturally occurring methicillin-resistant staphylococci.J. Gen. Microbiol. 35:183–190.
6. CLSI. 2007. Performance standards for antimicrobial susceptibility testing.CLSI approved standard M100–S17. Clinical and Laboratory Standards In-stitute, Wayne, PA.
7. De Jonge, B. L. M., Y.-S. Chang, D. Gage, and A. Tomasz. 1992. Peptidogly-can composition of a highly methicillin-resistant Staphylococcus aureusstrain: the role of penicillin binding protein 2A. J. Biol. Chem. 267:11248–11254.
8. De Jonge, B. L. M., H. de Lencastre, and A. Tomasz. 1991. Suppression ofautolysis and cell wall turnover in heterogeneous Tn551 mutants of amethicillin-resistant Staphylococcus aureus strain. J. Bacteriol. 173:1105–1110.
9. De Lencastre, H., and A. Tomasz. 1994. Reassessment of the number ofauxiliary genes essential for expression of high-level methicillin resistance inStaphylococcus aureus. Antimicrob. Agents Chemother. 38:2590–2598.
10. De Lencastre, H., B. L. M. de Jonge, P. R. Matthews, and A. Tomasz. 1994.Molecular aspects of methicillin resistance in Staphylococcus aureus. J. An-timicrob. Chemother. 33:7–24.
11. De Lencastre, H., S. W. Wu, M. G. Pinho, A. M. Ludovice, S. Filipe, S.Gardete, R. Sobral, S. Gill, M. Chung, and A. Tomasz. 1999. Antibioticresistance as a stress response: complete sequencing of a large number ofchromosomal loci in Staphylococcus aureus strain COL that impact on theexpression of resistance to methicillin. Microb. Drug Resist. 5:163–175.
12. Hartman, B. J., and A. Tomasz. 1984. Low-affinity penicillin binding proteinassociated with �-lactam resistance in Staphylococcus aureus. J. Bacteriol.158:513–516.
13. Jung, H. 2001. Towards the molecular mechanism of Na�/solute symport inprokaryotes. Biochim. Biophys. Acta 1505:131–143.
14. Katayama, Y., T. Ito, and K. Hiramatsu. 2000. A new class of geneticelement, staphylococcus cassette chromosome mec, encodes methicillin re-sistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 44:1549–1555.
15. Kornblum, J., B. J. Hartman, R. P. Novick, and A. Tomasz. 1986. Conversionof a homogeneously methicillin-resistant strain of Staphylococcus aureus toheterogeneous resistance by Tn551-mediated insertional inactivation. Eur.J. Clin. Microbiol. 5:714–718.
16. Madiraju, M. V., D. P. Brunner, and B. J. Wilkinson. 1987. Effects oftemperature, NaCl, and methicillin on penicillin-binding proteins, growth,peptidoglycan synthesis, and autolysis in methicillin-resistant Staphylococcusaureus. Antimicrob. Agents Chemother. 31:1727–1733.
17. Matthews, P., and A. Tomasz. 1990. Insertional inactivation of the mec genein a transposon mutant of a methicillin-resistant clinical isolate of Staphylo-coccus aureus. Antimicrob. Agents Chemother. 34:1777–1779.
18. Mendum, M. L., and L. T. Smith. 2002. Characterization of glycine betaineporter I from Listeria monocytogenes and its roles in salt and chill tolerance.Appl. Environ. Microbiol. 68:813–819.
19. Murakami, K., and A. Tomasz. 1989. Involvement of multiple genetic de-terminants in high-level methicillin-resistance in Staphylococcus aureus. J.Bacteriol. 171:874–879.
20. Oshida, T., and A. Tomasz. 1992. Isolation and characterization of a Tn551-autolysis mutant of Staphylococcus aureus. J. Bacteriol. 174:4952–4959.
21. Pereira, S. F., A. O. Henriques, M. G. Pinho, H. de Lencastre, and A.Tomasz. 2007. The role of PBP1 in the cell division of Staphylococcus aureus.J. Bacteriol. 189:3525–3531.
22. Reynolds, P. E., and D. F. Brown. 1985. Penicillin-binding proteins of beta-lactam-resistant strains of Staphylococcus aureus. Effect of growth conditions.FEBS Lett. 192:28–32.
23. Rubenhagen, R., S. Morbach, and R. Kramer. 2001. The osmoreactive be-taine carrier BetP from Corynebacterium glutamicum is a sensor for cyto-plasmic K�. EMBO J. 20:5412–5420.
24. Sabath, L. D., and S. J. Wallace. 1971. Factors influencing methicillin resis-tance in staphylococci. Ann. N. Y. Acad. Sci. 182:258–266.
25. Sabath, L. D. 1977. Chemical and physical factors influencing methicillinresistance of Staphylococcus aureus and Staphylococcus epidermidis. J. Anti-microb. Chemother. 3(Suppl. C):47–51.
26. Tomasz, A., S. Nachman, and H. Leaf. 1991. Stable classes of phenotypicexpression in methicillin-resistant clinical isolates of staphylococci. Antimi-crob. Agents Chemother. 35:124–129.
27. Ubukata, K., N. Yamashita, and M. Konno. 1985. Occurrence of a beta-lactam-inducible penicillin-binding protein in methicillin-resistant staphylo-cocci. Antimicrob. Agents Chemother. 27:851–857.
28. Wu, S., H. de Lencastre, and A. Tomasz. 1999. The Staphylococcus aureustransposon Tn551: complete nucleotide sequence and transcriptional anal-ysis of the expression of the erythromycin resistance gene. Microb. DrugResist. 5:1–7.
29. Wu, S., H. de Lencastre, A. Sali, and A. Tomasz. 1996. A phosphoglu-comutase-like gene essential for the optimal expression of methicillin resis-tance in Staphylococcus aureus: molecular cloning and DNA sequencing.Microb. Drug Resist. 2:277–286.
512 SIERADZKI ET AL. ANTIMICROB. AGENTS CHEMOTHER.
on August 31, 2018 by guest
http://aac.asm.org/
Dow
nloaded from