rhizobia indigenous to the okavango region in sub …aem.asm.org/content/80/23/7244.full.pdf ·...
TRANSCRIPT

Rhizobia Indigenous to the Okavango Region in Sub-Saharan Africa:Diversity, Adaptations, and Host Specificity
Jann L. Grönemeyer, Ajinkya Kulkarni, Dirk Berkelmann, Thomas Hurek, Barbara Reinhold-Hurek
Department of Microbe-Plant Interactions, Faculty of Biology and Chemistry, Center for Biomolecular Interactions Bremen (CBIB), University of Bremen, Bremen, Germany
The rhizobial community indigenous to the Okavango region has not yet been characterized. The isolation of indigenous rhizo-bia can provide a basis for the formulation of a rhizobial inoculant. Moreover, their identification and characterization contrib-ute to the general understanding of species distribution and ecology. Isolates were obtained from nodules of local varieties of thepulses cowpea, Bambara groundnut, peanut, hyacinth bean, and common bean. Ninety-one of them were identified by BOX re-petitive element PCR (BOX-PCR) and sequence analyses of the 16S-23S rRNA internally transcribed spacer (ITS) and the recA,glnII, rpoB, and nifH genes. A striking geographical distribution was observed. Bradyrhizobium pachyrhizi dominated at sam-pling sites in Angola which were characterized by acid soils and a semihumid climate. Isolates from the semiarid sampling sitesin Namibia were more diverse, with most of them being related to Bradyrhizobium yuanmingense and Bradyrhizobium daqin-gense. Host plant specificity was observed only for hyacinth bean, which was nodulated by rhizobia presumably representingyet-undescribed species. Furthermore, the isolates were characterized with respect to their adaptation to high temperatures,drought, and local host plants. The adaptation experiments revealed that the Namibian isolates shared an exceptionally hightemperature tolerance, but none of the isolates showed considerable adaptation to drought. Moreover, the isolates’ performanceon different local hosts showed variable results, with most Namibian isolates inducing better nodulation on peanut and hyacinthbean than the Angolan strains. The local predominance of distinct genotypes implies that indigenous strains may exhibit a betterperformance in inoculant formulations.
The catchment area of the Okavango River extends across partsof the three sub-Saharan African countries of Angola, Na-
mibia, and Botswana and is mainly characterized by smallholderand subsistence farming. Since cultivation techniques remain un-derdeveloped, e.g., in terms of irrigation and the application ofagrochemicals or rhizobial inoculants, local farmers are con-fronted with low yields and decreasing soil fertilities (1). Pulseslike cowpea (Vigna unguiculata), Bambara groundnut (Vigna sub-terranea), peanut (Arachis hypogaea), hyacinth bean (Lablab pur-pureus), and common bean (Phaseolus vulgaris) are cultivated andoccasionally intercropped, but natural nodulation rates are oftenlow (2), presumably due to a seasonal reduction of the rhizobialpopulation caused by drought and heat. Thus, nitrogen input viasymbiotic N2 fixation might be very limited.
Field-grown cowpea plants in Ghana have been reported to fixmore than 200 kg N/ha (89% of plant N) (3). In contrast, values ofonly 4 to 29 kg N-fixed/ha (15 to 56% of plant N) were observed insemiarid southwestern Zimbabwe (4). In such areas, the applica-tion of a rhizobial inoculant may thus hold the potential to in-crease plant nutrition and soil fertility. Many effective rhizobialstrains have been identified and are available. However, it has beenobserved that rhizobial strains often perform poorly under condi-tions dissimilar to their original habitat (5–7) and that their effec-tiveness depends on environmental factors such as soil tempera-ture (5, 6, 8), soil pH (9), soil texture (7), and host plant variety(10), an issue which might be especially relevant for the Okavangoregion with its harsh climate, heterogeneous soils, and local plantvarieties. In this regard, Law et al. (7) already reported that aprominent inoculant strain had no effect on cowpea and peanutgrown in Botswana. Furthermore, soybean inoculant applicationincreased seed yields at only one out of three locations in SouthAfrica (9). Hence, inoculant formulations consisting of effective
indigenous rhizobia being adapted to the local conditions mayexhibit a better performance.
Most grain legume species grown in the Okavango region areknown to be almost exclusively nodulated by a wide range of Bra-dyrhizobium spp. which were originally grouped as heterogeneous“cowpea miscellaneous rhizobia” according to, and solely basedon, their host range. A more precise classification to discrete spe-cies is hampered by the exceptional conservation of the 16S rRNAgene sequence, routinely used as a marker for species discrimina-tion, in the genus Bradyrhizobium (e.g., see references 11 and 12).Thus, for a long time the genus consisted of only three describedspecies: Bradyrhizobium japonicum (13), already described in1896 by Kirchner; Bradyrhizobium elkanii (14); and Bradyrhizo-bium liaoningense (15). Alternative markers such as the 16S-23SrRNA intergenic spacer (internally transcribed spacer [ITS]) (11,12, 16, 17) and the multilocus sequence analysis (MLSA) ap-proach to bradyrhizobia (e.g., see references 18 to 20) alloweduncovering of a high species diversity inside the genus Bradyrhi-zobium. It currently consists of 18 validly published species (LPSNdatabase [21]).
Studies on soybean-nodulating bradyrhizobia already revealeda clear species geographical distribution related to soil tempera-
Received 21 July 2014 Accepted 10 September 2014
Published ahead of print 19 September 2014
Editor: C. R. Lovell
Address correspondence to Barbara Reinhold-Hurek, [email protected].
Supplemental material for this article may be found at http://dx.doi.org/10.1128/AEM.02417-14.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AEM.02417-14
7244 aem.asm.org Applied and Environmental Microbiology p. 7244 –7257 December 2014 Volume 80 Number 23
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

ture (8), soil pH (22), and climate (22–24). Furthermore, eventhough it is well established that the host plant compatibility of arhizobial strain is generally dependent on the quality of its symlocus (19, 25, 26), chromosomal backgrounds can also have adecisive influence on the selection by the host plant (27–29). Thus,the assignment of a bradyrhizobial strain to a distinct species cangenerate information on adaptational properties, which in turnaccount for a strain’s suitability to serve as an inoculant at a par-ticular site.
Here, we report on the distribution and adaptation of rhizobiain the Okavango region and the identification of strains that holdthe potential to be used for the inoculation of locally grown pulses.
MATERIALS AND METHODSSampling and isolation of rhizobia from root nodules. Root noduleswere collected from the grain legumes cowpea (Vigna unguiculata), Bam-bara groundnut (V. subterranea), peanut (Arachis hypogaea), hyacinthbean (Lablab purpureus), and common bean (Phaseolus vulgaris). Theywere stored in 2-ml glass vials with silica gel desiccant at room tempera-ture during the field trip and at 4°C upon arrival in Bremen, Germany.The surveyed plants were growing in different soils, mainly on small-holder farms in the Namibian Mashare and the Angolan Cusseque areas(Fig. 1 and Table 1). Sampling dates and sites and sampling and isolationprocedures on modified arabinose gluconate (MAG) (30) medium aregiven in the work of Grönemeyer et al. (2). The isolates were designatedconsidering the individual host plant or soil sample (first number), therespective nodule (second number), and an isolate number (third num-ber). In cases where only two numbers are given, they correspond to theindividual host plant and the isolate number. For detailed information onsoil properties and climate, see the work of Gröngröft et al. and of Weber(31, 32). Soils were transported to Germany in plastic bags to preventdrying and under cooling (around 4°C). For trapping of rhizobia, a tea-spoon of the respective soil sample was used to inoculate cowpea andBambara groundnut seedlings which were grown aseptically in Magentajars containing vermiculite supplemented with half-concentrated N-freeJensen’s medium (33) as described for the cross-inoculation experiments.Root nodules were taken 3 weeks postinoculation.
Samples were collected under the research and collection permits1569/2011, 1635/2011, 1757/2012, and 1780/2013 and export permits83786, 90409, and 90990.
BOX repetitive element PCR (BOX-PCR) and amplification of the16S-23S rRNA ITS, nifH, and housekeeping genes. The templates for allPCR applications were crude extracts. A portion of a bacterial colony wassuspended in 10 �l Lyse and Go PCR reagent (Thermo Fisher ScientificInc., Rockford, IL, USA), which was then subjected to thermal cyclingaccording to the manufacturer’s instructions to release genomic DNA.
The suspension was briefly centrifuged, and 2 �l of the supernatant servedas a template in one PCR mixture.
PCR mixtures were composed of 2.5 U MolTaq DNA polymerase(Molzym, Bremen, Germany), 5 �l of the respective 10� PCR buffer, 50�M (each) deoxynucleoside triphosphate (dNTP), 0.5 �M (each) primer,and 2 �l template in a total volume of 50 �l. The 16S-23S rRNA gene ITSwas amplified using the primer pair FGPS1490/FGPS132= under the cy-cling conditions described by Laguerre et al. (34) with increased annealingtemperature at 58°C. Partial amplification of glnII, recA, and rpoB frag-ments was achieved using the primer pairs and cycling parameters re-ported previously (19, 23). Amplification of nifH fragments was con-ducted with primers FGPH19/PolR (35) under the cycling conditionsdescribed by Demba-Diallo et al. (36).
For BOX-PCR, the reaction mix of 50 �l consisted of 2.5 U DreamTaqDNA polymerase (Thermo Fisher Scientific), 5 �l of the respective 10�PCR buffer, 100 �M (each) dNTP, 2 �M primer BOXA1R (37), and 2 �lof template. DNA amplification was carried out in a Biometra TProfes-sional Thermocycler (Biometra, Göttingen, Germany) conducting 35 cy-cles under the cycling conditions described previously (37). PCR productswere separated by horizontal gel electrophoresis on 1.5% (wt/vol) agarosegels in Tris-acetate-EDTA (TAE) buffer for 1.5 h at 90 V. Ethidium bro-mide-treated gels were scanned with a Typhoon FLA 9500 scanner (GEHealthcare Life Sciences).
Sequencing and phylogenetic analysis. Amplification products wereeither purified using the QIAquick PCR purification kit (Qiagen, Hilden,Germany) or custom purified before Sanger sequencing (LGC Genomics,Berlin, Germany). Sequencing primers were the same forward primerspreviously used for PCR amplification, except for nifH (primer PolF)(35).
Multiple sequence alignments were generated using either ClustalWversion 5.1 (38) or MUSCLE (39) incorporated in the MEGA 5.2 software(40). For ITS sequences, ClustalW was used and the gap open/extensionpenalties were set to 20/6.66 for the pairwise and 50/6.66 for the multiplealignment, respectively. The alignment was visually inspected and manu-ally edited. Positions containing gaps were deleted in the phylogeneticanalysis, and distances were calculated from the number of differences assuggested by Willems et al. (12). For the analysis of protein-coding se-quences, the MUSCLE aligner was used under default settings. Thealigned housekeeping gene fragment sequences were manually trimmedto remove overhangs and analyzed individually as well as in a concate-nated fashion. The best-fit models of evolution were determined usingMODELTEST (41) integrated in MEGA5 software, and the substitutionmodel with the lowest Bayesian information criterion (BIC) (42) scorewas chosen for a maximum likelihood-based phylogenetic analysis. Thereliability of the tree topology was estimated by conducting a bootstraptest (43) with 500 pseudoreplicates.
Tests for temperature tolerance and osmotolerance of isolates. Theisolates’ maximum growth temperature was surveyed on MAG agar slants(at 28.5, 35.5, and 38.5°C � 0.5°C each) as well as in liquid cultures.Precultures grown at 28°C in MAG medium were adjusted to an opticaldensity at 600 nm (OD600) of 0.2, and 25 �l was used for inoculation of25-ml test cultures. Test cultures were grown on a rotary shaker (200 rpm)at 35.5°C (�0.5°C) or in an agitating water bath at higher temperatures(38, 40, and 42°C). Cultures were inspected for visible growth within 7days of incubation. To survey the isolates’ drought tolerance, MAG me-dium was supplemented with various amounts of polyethylene glycol(PEG) 6000 (Rotipuran; Carl Roth, Karlsruhe, Germany) (15, 20, 25, and30% [wt/vol]), and cultures were grown on a rotary shaker with 200 rpmat 28°C. Bacterial density was monitored at OD600 over a period of 3weeks.
Cross-inoculation experiments. The isolates were tested for theirability to nodulate cowpea variety Lutembwe as well as local varieties ofcowpea, Bambara groundnut, peanut, hyacinth bean, and common bean.Hand-sorted beans free of visible damage and of uniform size were surfacesterilized by rinsing them in 70% ethanol for 10 s followed by three wash-
FIG 1 Okavango region and core sampling sites.
Rhizobial Community Indigenous to the Okavango Region
December 2014 Volume 80 Number 23 aem.asm.org 7245
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

TA
BLE
1C
har
acte
rist
ics
ofis
olat
esan
dre
fere
nce
stra
ins
use
din
this
stu
dya
Isol
ate
orre
fere
nce
stra
in
No.
ofis
olat
esre
pres
ente
d
Ori
gin
Ph
ylog
eny
Max
imu
mte
mp
(°C
)c
Max
imu
mP
EG
(%)
Hos
tra
nge
d
Pla
nt
Cou
ntr
yLo
cati
onb
Soil
type
Lan
du
seC
ore
ITS
Solid
Liqu
idC
OW
BA
MH
YA
PE
AP
HA
Isol
ates
1-7
1P
EA
NA
MA
MA
DI
DA
II
3835
*20
AP
AB
NB
B3-
21
CO
WN
AM
CM
AD
IIA
II
3835
ND
CC
ND
ND
C4-
81
CO
WN
AM
CM
AD
IIA
II
3828
ND
AP
AN
DN
DB
6-8
1C
OW
NA
ME
MA
DI
DA
II
3835
20A
PA
BN
AP
ND
2-13
1P
EA
NA
MB
MA
DI
DA
III
I38
3520
AA
AA
PB
222-
11
CO
WN
AM
KK
SM
AD
ID
AII
III
I38
38N
DC
CC
CC
602-
11
BA
MN
AM
MO
FPD
AII
II38
38N
DA
AA
AB
582-
11
PE
AN
AM
MO
FPD
AII
II38
3820
AA
AP
AB
572-
11
PE
AN
AM
MO
FPD
AII
II38
38N
DN
DN
DN
DN
DN
D54
2-1
1B
AM
NA
MM
OFP
DA
IIII
3838
20A
AA
AB
541-
11
BA
MN
AM
MO
FPD
AII
IIN
DN
DN
DN
DN
DN
DN
DN
D55
1-1
1B
AM
NA
MM
OFP
DA
IIII
3835
20A
AB
AP
B26
1-1
2P
EA
NA
MK
KS
MA
DI
DA
VV
3835
20A
AN
DA
B21
1-1
1C
OW
NA
MK
KS
MA
DI
DA
IVIV
38N
D20
CC
CN
CC
B14
6-1
2B
AM
NA
MT
KS
BV
IVIV
ND
ND
ND
ND
ND
ND
ND
ND
282-
11
PE
AN
AM
KK
SM
AD
ID
AIV
IV38
40N
DA
AN
DN
DB
15C
4-1
2C
OW
NA
MU
KS
BV
IVIV
ND
ND
ND
ND
ND
ND
ND
ND
7-2
2C
OW
NA
MF
MA
DI
DA
IVIV
3840
ND
AP
AA
PA
PB
16-1
02
PE
AN
AM
JK
SD
AIV
IVN
DN
DN
DN
DA
ND
ND
B10
-21
CO
WN
AM
HK
SD
AIV
IV38
4020
AA
ND
ND
B1B
1-1
2B
AM
NA
MH
KS
DA
IVIV
ND
ND
ND
ND
ND
ND
ND
ND
363-
22
BA
MN
AM
HK
SD
AIV
IV38
40N
DA
AN
DN
DB
3B4-
11
BA
MN
AM
SO
FPF
IVIV
ND
ND
ND
ND
ND
ND
ND
ND
B14
4-2
1B
AM
NA
MT
KS
BV
IVIV
ND
ND
ND
ND
ND
ND
ND
ND
271-
12
PE
AN
AM
KK
SM
AD
ID
AIV
IV38
4020
AP
AP
AP
AP
B35
1-3
1C
OW
NA
MH
KS
DA
IVIV
3840
ND
ND
ND
ND
ND
ND
9-5
1B
AM
NA
MH
KS
DA
IVIV
3840
20A
AN
DN
DB
14-3
1C
OW
NA
MI
OFP
DA
VI
VI
3838
20A
PA
AA
PN
D34
1-1
2C
OW
NA
MH
KS
DA
VI
VI
3838
20A
AN
DA
B1B
T1-
11
BA
MN
AM
HK
SD
AV
IV
IN
DN
DN
DN
DN
DN
DN
DN
D35
3-3
2C
OW
NA
MH
KS
DA
VI
VI
3838
ND
AA
ND
ND
B37
1-1
2B
AM
NA
MH
KS
DA
VI
VI
3838
ND
ND
ND
ND
ND
ND
451-
42
CO
WA
NG
PD
AV
IIV
II38
380
AA
AP
BB
361-
11
BA
MN
AM
HK
SD
AX
IV(B
)N
D35
3520
AP
AA
AB
311-
11
HY
AN
AM
KK
SM
AD
ID
AV
III
VII
I35
2820
AP
AA
PA
PB
21B
2-1
1B
AM
NA
MW
OFP
IAV
III
VII
IN
DN
DN
DN
DN
DN
DN
DN
D32
2-1
2H
YA
NA
MK
KS
MA
DI
DA
ND
VII
I35
2820
AA
AP
AP
B30
2-1
2H
YA
NA
MK
KS
MA
DI
DA
VII
IV
III
3528
20N
DN
DN
DN
DN
D26
3-1
1P
EA
NA
MK
MA
DI
DA
IXIX
3835
20A
PA
AP
BN
B18
C2-
11
CO
WN
AM
VO
FPD
AIX
IX38
3820
AN
DA
PB
B18
C1-
11
CO
WN
AM
VO
FPD
AX
X35
3520
AA
PA
PA
B5-
102
HY
AN
AM
DM
AD
ID
AX
IX
I38
*35
0A
PA
AP
AP
B30
3-2
2H
YA
NA
MK
KS
DA
XI
XI
3535
0A
AA
PA
B
Grönemeyer et al.
7246 aem.asm.org Applied and Environmental Microbiology
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from
AD
1T2
3-1
1B
AM
AN
GY
DA
XII
XII
ND
ND
ND
ND
AB
BN
D40
2-2
1C
OW
AN
GN
DA
XII
IX
III
ND
28N
DN
DN
DN
DN
DN
D40
3-1
1C
OW
AN
GN
DA
XII
IX
III
3528
ND
AA
ND
ND
B44
1-1
1P
HA
AN
GP
DA
XII
IX
III
3528
ND
AA
BN
BN
B42
1-1
2P
HA
AN
GO
DA
XII
IX
III
3528
20A
PA
BN
BN
B41
1-2
2C
OW
AN
GN
DA
XII
IX
III
3528
20A
AN
DN
DB
492-
13
PH
AA
NG
QD
AX
III
XII
I35
2820
AA
ND
BB
F12-
11
BA
MA
NG
XF
XII
IX
III
ND
ND
ND
ND
ND
ND
ND
ND
511-
32
CO
WA
NG
QD
AX
III
XII
I35
*28
20A
AB
NB
NB
P14
1-1
1B
AM
AN
GZ
PX
III
XIV
35*
2820
AP
AP
BB
ND
471-
13
CO
WA
NG
PD
AX
III
XV
3528
ND
AA
ND
ND
BF1
1-1
1B
AM
AN
GX
FX
III
XV
ND
ND
ND
ND
ND
ND
ND
ND
461-
13
CO
WA
NG
PD
AX
III
XV
35*
2820
ND
AA
BN
B62
1-1
1C
OW
NA
MM
OFP
DA
Rhi
zobi
umsp
.R
hizo
bium
sp.
3840
30C
NN
DC
NC
NA
P
543-
12
BA
MN
AM
MO
FPD
AR
hizo
bium
sp.
Rhi
zobi
umsp
.38
4030
CN
ND
CN
CA
P
432-
12
PH
AA
NG
PD
AR
hizo
bium
sp.
Rhi
zobi
umsp
.38
38*
25C
CN
CN
CA
P
433-
11
PH
AA
NG
PD
AR
hizo
bium
sp.
Rhi
zobi
umsp
.N
DN
DN
DN
DN
DN
DN
DN
D52
3-1
2P
HA
AN
GR
Rhi
zobi
umsp
.R
hizo
bium
sp.
ND
ND
ND
ND
ND
ND
ND
ND
531-
12
PH
AA
NG
RR
hizo
bium
sp.
Rhi
zobi
umsp
.38
38*
30C
CN
CC
NA
P
Ref
eren
cest
rain
sB
.hua
nghu
aiha
iens
eLM
G26
136
38*
28N
DA
AP
BB
NB
B.i
riom
oten
seLM
G24
129
35*
28N
DA
AP
ND
AP
BB
.jic
amae
LMG
2455
628
2820
AA
PN
DN
DB
B.l
abla
biL
MG
2557
235
*28
20A
AP
AA
BB
.lia
onin
gens
eL
MG
1823
038
*28
20C
CN
ND
ND
CB
.pac
hyrh
iziL
MG
2424
635
28N
DA
AP
BN
BN
BN
B.e
lkan
iiLM
G61
3435
2820
AA
PN
DN
DB
N
B.y
uanm
inge
nse
LMG
2182
738
3820
AA
PN
DB
BN
B.d
aqin
gens
eLM
G26
137
3835
20A
AN
DB
NB
Bra
dyrh
izob
ium
sp.s
trai
nU
SDA
3384
3535
*20
AP
AA
AB
Bra
dyrh
izob
ium
sp.s
trai
nU
SDA
3451
3528
ND
AA
AA
BB
rady
rhiz
obiu
msp
.str
ain
USD
A38
3438
*35
ND
AA
ND
ND
Ba
Abb
revi
atio
ns:
ND
,not
dete
rmin
ed;C
OW
,cow
pea;
BA
M,B
amba
ragr
oun
dnu
t;H
YA
,hya
cin
thbe
an;P
EA
,pea
nu
t;P
HA
,com
mon
bean
;AN
G,A
ngo
la;N
AM
,Nam
ibia
;MA
DI,
Mas
har
eA
gric
ult
ura
lDev
elop
men
tIn
stit
ute
;KS,
Kal
ahar
isan
ds;O
FP,o
ldfl
oodp
lain
soil;
DA
,dry
lan
dag
ricu
ltu
re;I
A,i
rrig
atio
nag
ricu
ltu
re;B
V,B
ush
veld
;F,f
allo
waf
ter
subs
iste
nce
farm
ing.
bSa
mpl
ing
loca
tion
sas
desc
ribe
din
the
wor
kof
Grö
nem
eyer
etal
.(2)
.c
Ast
eris
ksin
dica
tew
eak
grow
th.
dA
,eff
ecti
ven
odu
lati
on;B
,in
effe
ctiv
en
odu
lati
on(s
eeth
ete
xt);
C,n
on
odu
lati
on;s
upe
rscr
ipt
P,A
RA
posi
tive
;su
pers
crip
tN
,AR
An
egat
ive.
Rhizobial Community Indigenous to the Okavango Region
December 2014 Volume 80 Number 23 aem.asm.org 7247
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

ings in sterile distilled water and immersion in 2.5% sodium hypochloritefor 6 min. Seeds were again washed five times in sterile distilled water,followed by soaking in water for at least 2 h. To ensure that the seeds werethoroughly surface sterilized, randomly chosen beans were placed onMAG agar plates and inspected for microbial growth within 3 days. Sur-face-disinfected seeds were allowed to pregerminate on water agar (1.5%[wt/vol]) in the dark at 28°C for up to 4 days (depending on bean), plantedinto Magenta jars (Magenta Corp., Chicago, IL, USA) containing sterilewashed vermiculite supplemented with 100 ml of 0.5� Jensen’s medium(33), and inoculated with 109 cells of the respective isolate in 1 ml of a 1%(wt/vol) sucrose solution. Each isolate was assayed at least in duplicate,with uninoculated control plants. The plants were maintained under con-trolled conditions with day/night (11.5-h/12.5-h) temperatures of 28°C/25°C and lighting at 16.4 klx. The humidity was set to 60%, and plantswere watered once a week with sterile distilled water. After 3 (cowpea,Bambara groundnut, and hyacinth bean) to 4 (peanut and common bean)weeks of growth, the presence and appearance of root nodules were con-trolled. To test for nitrogenase activity, an acetylene reduction assay(ARA) on whole roots was carried out in sealed 80-ml glass tubes with10% (vol/vol) acetylene in the headspace. After 4 h of incubation in thedark at 28°C, a 1-ml gas sample was transferred to 3-ml Exetainers (LabcoLtd., Lampeter, Wales, United Kingdom) and stored until gas chromato-graphic quantification of ethylene.
Nucleotide sequence accession numbers. Sequences were submittedto GenBank (accession no. KM378245 to KM378555).
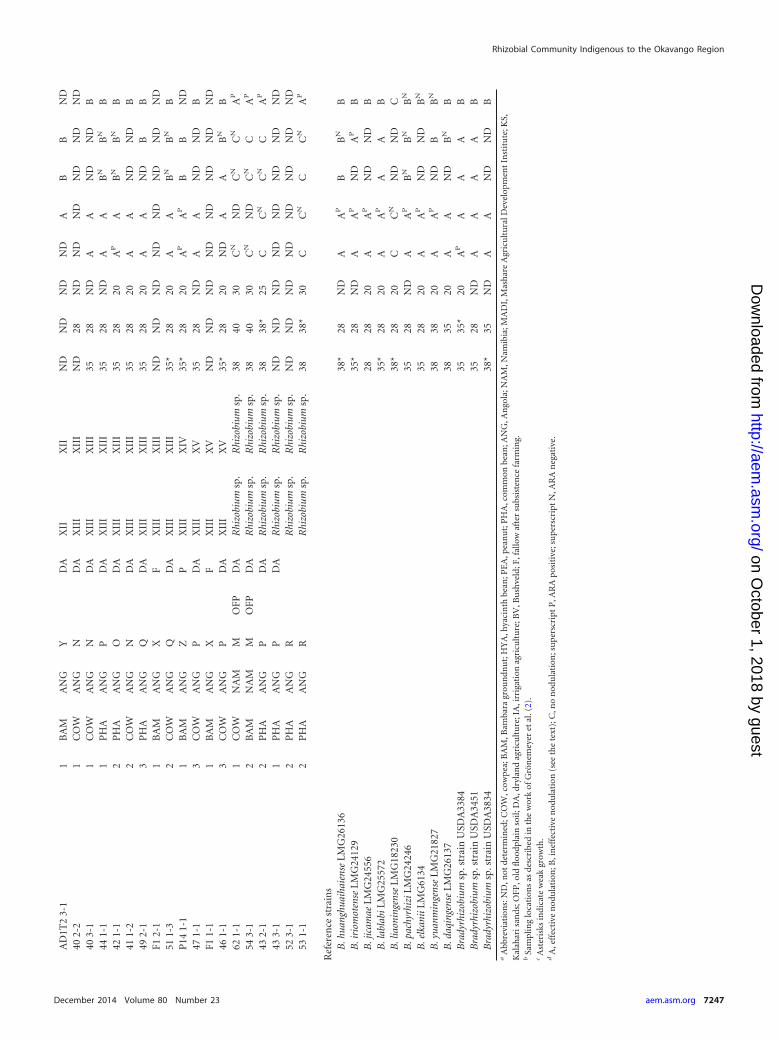
RESULTSIsolation of root nodule rhizobia from the Okavango region. Atotal of 91 putative rhizobia (60 from Namibia, 31 from Angola)were isolated from 86 individual root nodules of various pulses(Table 1). The 75 isolates from nodules collected in the fields werecomplemented by 16 isolates obtained from trapping from soilsamples. Among others, the trapping experiment added some An-golan rhizobia trapped by Bambara groundnut, which was notgrown on fields sampled in Angola.
As expected, the majority of the isolates (75/78) obtained fromcowpea, Bambara groundnut, peanut, and hyacinth bean wereslow growing, with single colonies becoming visible after 5 to 7days of incubation on MAG agar plates, indicating that they arebradyrhizobia. However, from two nodules (sampled from plants54 and 62), only fast-growing isolates with colony morphologytypical for Rhizobium sp. were obtained (isolates 54 3-1 and 621-1), while for another nodule from plant 54, some fast-growingcolonies (which turned out to be genotypically identical with 543-1) were observed on plates dominantly populated by slow-growing isolate 54 1-1. From common bean, most strains (7/13)were fast growing, with single colonies appearing after 1 to 2 daysof growth as expected, but slow growers were isolated also.
All isolates were initially screened for the presence of the nifHgene, and a PCR amplification product with the expected size ofabout 450 bp was observed for all of the 91 isolates except forisolate 22 2-1. As colony morphology was similar to that of bra-dyrhizobia, this strain was included in the subsequent analysis.
Grouping of isolates by BOX-PCR fingerprinting. In order toidentify similar isolates and to select representatives for furtherstudies, the BOX-PCR fingerprinting technique was applied. Theresulting patterns were highly reproducible and were rather influ-enced by PCR parameters (e.g., the thermal cycler used) than bythe PCR template (data not shown). The patterns of slow-growingisolates originating from the same individual nodule were alwaysidentical (Fig. 2), thus indicating their clonality. Most of the othervery similar patterns belonged to isolates from the same individual
plant. However, in some rare cases identical to very similar pat-terns were observed for isolates obtained from different individualplants (34 1-1 and 35 3-3; 30 2-1 and 31 1-1) as well as fromdifferent plant species (54 2-1 and 58 2-1) at different samplingsites (10-2 and 28 2-1). These results indicated that some rhizobialgenotypes are competitive in nodulation of several host plants atvarious locations. In contrast, the isolation of different genotypesfrom nodules of the same plant was rare, indicating that the indi-vidual plants were preferentially nodulated by a group of veryclosely related rhizobia at a distinct location.
The fast-growing isolates and the isolates obtained from thetrapping experiment were analyzed separately (data not shown).Fifty-six slow-growing and six fast-growing isolates, representingeither a distinct BOX-PCR pattern or an individual plant, wereselected for the subsequent analyses.
Phylogenetic classification of rhizobia. A first classificationwas based on sequence comparisons of the 16S-23S rRNA geneITS region. As expected, the ITS regions varied in length (approx-imately 800 to 1,000 bp), resulting in sections with large gaps inthe alignment. Phylogenetic trees computed with and without in-clusion of gaps led to similar but slightly different topologies (datanot shown). As Willems et al. (12) observed a better agreementwith their results from DNA-DNA hybridizations when gaps inthe ITS alignment were not penalized, we adopted this approach.GenBank accession numbers of reference sequences are listed inthe supplemental material (see Table S1 in the supplemental ma-terial). The resulting phylogenetic tree was based on 634 positionsand separated (100% and 98% bootstrap support) the B. japoni-cum well from the B. elkanii lineage (20, 23), the latter also show-ing good bootstrap support of the subclades (Fig. 3). The isolateswere highly diverse, forming several distinct clusters distributedover both major clades. As most of these clusters were composedof isolates obtained from different plant species and often evenshared identical ITS sequences, the plant origin had only a minorinfluence on the relatedness of the isolates. Exceptions were therhizobia obtained from hyacinth bean.
Interestingly, a geographic distribution was observed, with allAngolan rhizobia apart from isolate 45 1-4 clustering in proximityto Bradyrhizobium pachyrhiziT on the B. elkanii branch. For mostNamibian isolates, the ITS sequence had highest similarity to theones of Bradyrhizobium daqingenseT, Bradyrhizobium yuanmin-genseT, or Bradyrhizobium iriomotenseT located on the B. japoni-cum branch. The remaining Namibian isolates showed remark-ably low relatedness to any bradyrhizobial species type strain,suggesting that they represent novel species. This was especiallyevident for the isolates 26 3-1 and 18C 2-1 early branching fromthe B. elkanii lineage as well as for isolates 5-10 and 30 3-2.
The isolates were grouped by their sequence identity to facilitatecomparisons with the phylogeny of concatenated housekeepinggenes and to select representatives for the adaptation experiments.The genospecies threshold of 95.5% ITS sequence identity proposedpreviously (44) was not chosen because higher similarities amongdifferent species were observed in our sequence comparisons: Brady-rhizobium betaeT shared 96.7% and 96.0% of its ungapped ITSsequence with Bradyrhizobium canarienseT and BradyrhizobiumdiazoefficiensT, respectively. Furthermore, some other pairs of typestrains showed similarities slightly above 95.5%, which were B.canarienseT and Bradyrhizobium cytisiT (95.9%), B. canarienseT
and B. diazoefficiensT (95.9%), B. betaeT and B. liaoningenseT
(95.6%), B. cytisiT and B. japonicumT (95.9%), Bradyrhizobium
Grönemeyer et al.
7248 aem.asm.org Applied and Environmental Microbiology
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

lablabiT and Bradyrhizobium jicamaeT (95.7%), and B. yuanmin-genseT and Bradyrhizobium huanghuaihaienseT (95.6%), respec-tively. Therefore, the threshold was set to 97%, resulting in 15distinct clusters with �98% sequence identity among the isolateswithin one cluster. Isolates from clusters IV and XIII showed highsequence similarities to B. yuanmingenseT (96.8 to 97.1%) and B.pachyrhiziT (98.4 to 98.7%), respectively, strongly supporting theidea that they are members of the corresponding species. In con-trast, the sequence identity to the closest related species type strainwas rather low for clusters IX (88.7 to 88.9%) and XI (91.7%).
To better support this classification, the evolutionary history ofpartial sequences of the glnII, recA, and rpoB genes was analyzedindividually and in a concatenated fashion. The proportion of theindividual genes in the 1,625-bp-long concatenated sequences was505 bp for glnII, 381 bp for recA, and 739 bp for rpoB. The best-fitmodel of evolution evaluated by MODELTEST was the generaltime reversible model with a gamma-distributed rate heterogene-ity and invariant sites (GTR �G �I). The calculated phylogenetictree corroborated geographic distribution of the isolates, as theclustering appeared to be very similar to the one obtained fromITS sequence analysis (Fig. 4). Some inconsistencies could befound in early branching inside the B. japonicum lineage, mostprobably deriving from the fact that the rapidly evolving ITS lacks
resolution at higher taxonomic ranks. One conspicuous differencewas the separation of isolates 2-13 and 22 2-1 from previous clus-ter III, with B. liaoningenseT showing a close relationship to isolate22 2-1. While the divergence of 2-13 and 22 2-1 was observed foreach individual housekeeping gene (see Fig. S1 to S3 in the sup-plemental material), the shift of B. liaoningenseT was mainly basedon rpoB phylogeny (Fig. S3).
The range of sequence variation of concatenated sequences wassimilar to the one observed for the ITS sequences, but in several cases,the resolution was slightly increased, discriminating isolates withidentical ITS sequences. The sequence identity between species typestrains was up to 96.3% (B. lablabiT and B. jicamaeT); thus, theisolates were grouped by 97% sequence identity. The resulting 14groups were almost identical to the ITS-based ones, giving goodsupport in the overall classification of the isolates. Exceptions werethe change of isolate 2-13 to cluster I and merging of ITS clustersXIII to XV. One additional cluster resulted from isolate 36 1-1 notoccurring in ITS phylogeny. For 36 1-1, the ITS PCR yielded twoproducts slightly different in size which were thus not assigned inFig. 3, indicating that this isolate is one of the rare Bradyrhizobiumspecies which exhibit more than one rRNA operon (44). Thus,assignment based on housekeeping genes may be more reliable.
Additionally, the relatedness of the isolates’ symbiotic islands
FIG 2 BOX-PCR fingerprints of isolates from nodules. Solid and dashed lines indicate identical or very similar genotypes, respectively.
Rhizobial Community Indigenous to the Okavango Region
December 2014 Volume 80 Number 23 aem.asm.org 7249
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

FIG 3 Neighbor-joining phylogram inferred from ITS sequences of bradyrhizobial isolates and species type strains. Rhodopseudomonas palustrisT was includedas an outgroup strain. A bootstrap value is indicated when a given node appeared in �50% of 500 pseudoreplicates. The following symbols indicate the plantorigin of the isolates: �, Namibian cowpea; Œ, Namibian Bambara groundnut; �, Namibian peanut; }, Namibian hyacinth bean; p, Angolan cowpea; o,Angolan Bambara groundnut; �, Angolan common bean. Isolates labeled with “(soil)” were trapped from soil samples. Roman numerals cluster isolates with�97% sequence identity. The scale bar indicates the number of base differences per sequence.
7250 aem.asm.org Applied and Environmental Microbiology
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

FIG 4 Maximum likelihood phylogram inferred from glnII-recA-rpoB sequence concatemers based on the GTR �G �I model. Rhodopseudomonas palustrisBisB5 was included as an outgroup strain. A bootstrap value is indicated when a given node appeared in �50% of 500 pseudoreplicates. The following symbolsindicate the plant origin of the isolates: �, Namibian cowpea; Œ, Namibian Bambara groundnut; �, Namibian peanut; }, Namibian hyacinth bean;p, Angolancowpea;o, Angolan Bambara groundnut; �, Angolan common bean. Isolates labeled with “(soil)” were trapped from soil samples. Roman numerals clusterisolates with �97% sequence identity. The scale bar indicates the number of substitutions per site.
Rhizobial Community Indigenous to the Okavango Region
December 2014 Volume 80 Number 23 aem.asm.org 7251
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

was investigated. As several studies reported a congruent phylog-eny among nodulation genes and nifH (45–47), the latter wasregarded to be representative. The final alignment consisted of 286nucleotides, and the Tamura 3-parameter model with gamma-distributed evolutionary rates among sites (T92 �G) was the best-fit model according to MODELTEST analysis. The resulting phy-logenetic tree showed a slightly differing topology from theprevious ones (Fig. 5), which might be due to some strain se-quences not included in both alignments or to lateral gene transferof nifH. Three main clusters were observed, the first one (A) in-cluding most Namibian isolates, the second one (B) exclusivelyharboring Angolan isolates, and a third one (C) containing theNamibian isolates from the previous B. elkanii branch which weremainly obtained from the newly introduced hyacinth bean.
Adaptation to temperature and water stress. The rhizobiawere subjected to high growth temperatures and water stress toevaluate their adaptation to the harsh Okavango climatic condi-tions. Nine bradyrhizobial species type strains, which showed thehighest degree of relatedness to the isolates in the phylogeneticanalyses, were included as references. Furthermore, stress toler-ance of the indigenous rhizobia was compared to that of threeUSDA strains (U.S. Department of Agriculture, Beltsville, MD,USA), which are recommended for the inoculation of legumes insub-Saharan Africa (Table 1).
To test their ability to grow at high temperatures, rhizobia weregrown on solid (28, 35, and 38°C) as well as in liquid medium at28, 35, 38, 40, and 42°C. On solid media, higher maximal growthtemperatures were reached (Table 1), which was already observedelsewhere (48). Most type strains and all Angolan bradyrhizobiaexcept for 45 1-4 did not grow in liquid medium at 35°C, whichwas also the case for two of the USDA strains. However, for severalspecies type strains the observed temperature tolerances were in-consistent with their original description, which might be a matterof different growth media. In contrast, most Namibian isolatesproliferated at 38°C, with some even tolerating growth tempera-tures as high as 40°C. The Namibian isolates most sensitive to hightemperatures were the ones obtained from hyacinth bean belong-ing to cluster VIII and one isolate from ITS cluster I. The pheno-types were for most strains in agreement with the genotypicgrouping. To investigate the isolates’ drought tolerance, waterstress was induced by the addition of PEG 6000 to the liquidgrowth medium at final concentrations of 15, 20, 25, and 30%(wt/vol). In a pretest, the influence of PEG on growth rates withinthe first 3 days postinoculation was monitored for a selection ofisolates. In the presence of lower PEG concentrations (15%),growth kinetics were characterized by longer lag phases (or celldeath due to osmotic shock), after which doubling times ap-proached the ones observed without PEG (not shown), Thus,these conditions were not selected for differentiation. At PEG con-centrations of 20%, longer lag phases were detected and doublingtimes in the exponential phase were strongly increased. In order totest the isolates’ general ability to grow under different concentra-tions of PEG, we thus finally used extended incubation times.Rhizobium bacteria were grown for 1 week, and slow-growing Bra-dyrhizobium bacteria were surveyed over a period of 3 weeks. Al-most all bradyrhizobial cultures became stationary within 1 weekof growth at 15% PEG and 2 weeks of growth at 20% PEG, andnone of the bradyrhizobial isolates showed detectable growth athigher PEG concentrations (Table 1). As similar results were ob-tained for the species type strains, no adaptation exclusive to in-
digenous rhizobia was detected. In contrast, three isolates (bothhyacinth bean isolates forming cluster XI and isolate 45 1-4) werenot even growing at 15% PEG and therefore were highly suscep-tible to water stress. All isolates identified as Rhizobium sp. toler-ated higher PEG concentrations of up to 30%.
Cross-inoculation of groups of isolates. The isolates weretested for their ability to nodulate a range of locally grownlegumes, mostly belonging to the “cowpea miscellany” cross-in-oculation group (cowpea, Bambara groundnut, hyacinth bean,and peanut). The only exception was the common bean, belong-ing to the bean cross-inoculation group. In agreement with thecross-inoculation group concept, isolates which were identified asRhizobium sp. formed nitrogen-fixing nodules on common beanbut failed to nodulate all other plants (Table 1).
Among the bradyrhizobial isolates, three (3-2, 21 1-1, and 22 2-1)did not induce root nodules on any of the legumes tested, indicatingthat they are not symbiotic. This was especially surprising for isolates3-2 and 21 1-1, which possessed the nifH gene and were highly relatedto nodulating isolates. Furthermore, B. liaoningenseT failed to nod-ulate any legume under the selected conditions. All other brady-rhizobia induced root nodules on both Vigna species (cowpea andBambara groundnut) and were actively fixing nitrogen as deter-mined by acetylene reduction assays of nodulated roots.
Variable results were observed for hyacinth bean and peanut.Even though almost all isolates were able to nodulate hyacinthbean, remarkable variation in nodule size was observed, correlat-ing with previously defined groups. All isolates located on branchC in nifH phylogeny (groups VIII to XI) induced 10 to 30 bignodules of up to 5 mm in diameter, while for most isolates fromnifH branch A (groups II to VII), nodules appeared to be smaller(up to 3 mm in diameter). Nevertheless, the nitrogen fixationactivities determined by the ARA were similar (data not shown).In contrast, when Angolan isolates from nifH branch B (groupsXIII to XV except for isolate 46 1-1) were used for inoculation,root nodules remained tiny (mostly about 1 mm in diameter), andno nitrogen fixation could be detected with a detection limit cor-responding to 50 ppm ethylene in 80 ml of headspace. This resultwas corroborated by the observation that the plants were appar-ently suffering nitrogen shortage, exhibiting light green leaves andceasing to grow upon prolonged time (data not shown).
For peanut, the results were similar to the ones observed forthe hyacinth bean: while the bulk of isolates induced effectivenodulation, nodules did not fully develop and no nitrogen fix-ation activity was measured when peanuts were inoculatedwith Angolan isolates (see Fig. S4 in the supplemental mate-rial). Moreover, isolates from two more groups, which wereITS groups I and IX located on nifH branches A and C, respec-tively, failed to nodulate. However, moderate nodulation andsome nitrogenase activity could be detected for isolate 6-8 fromgroup I. Again, the different degrees of effectiveness could beclearly seen from various green color intensities of the leavesafter 4 weeks of growth (not shown).
Apart from the four strains mentioned above, which did notnodulate any plant, all bradyrhizobia induced whitish and irregu-larly shaped nodule-like structures on common bean, as previ-ously observed (49). The plants showed clear symptoms of Nshortage, and no N2 fixation could be detected by ARA (see Fig. S4in the supplemental material).
Grönemeyer et al.
7252 aem.asm.org Applied and Environmental Microbiology
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

FIG 5 Maximum likelihood phylogram inferred from nifH sequences based on the T92 �G model. Rhodopseudomonas palustrisT was included as an outgroupstrain. A bootstrap value is indicated when a given node appeared in �50% of 500 pseudoreplicates. The following symbols indicate the plant origin of theisolates: �, Namibian cowpea; Œ, Namibian Bambara groundnut; �, Namibian peanut; }, Namibian hyacinth bean;p, Angolan cowpea;o, Angolan Bambaragroundnut; �, Angolan common bean. Isolates labeled with “(soil)” were trapped from soil samples. The scale bar indicates the number of substitutions per site.Grouping is not based on a sequence identity threshold.
December 2014 Volume 80 Number 23 aem.asm.org 7253
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

DISCUSSION
Our study reports on the rhizobial population in the OkavangoRiver region, which is characterized by a harsh climate, heteroge-neous soils, and local pulse varieties. Since no inoculation historyis known at the sampled sites, the isolates are expected to representthe indigenous rhizobial population that is adapted to the localenvironmental conditions. Thus, our results contribute to thegeneral understanding of regional species abundance. Moreover,they provide a basis for the formulation of a rhizobial inoculantmatching the local settings at the best.
As could be expected from the widely recognized compatibilityof the sampled pulses, the spectrum of isolates comprised bothBradyrhizobium sp. and Rhizobium sp., which were mainly ob-tained from “cowpea group” pulses and common bean, respec-tively. However, occasionally Rhizobium bacteria were isolatedfrom cowpea as well as Bradyrhizobium species from commonbean. Even though other studies reported having isolated fast-growing rhizobia nodulating cowpea (50–52), all our Rhizobiumisolates failed to nodulate any plant except for the common beanand did thus not represent the actual symbionts. This might alsobe the case for Bradyrhizobium spp. obtained from common bean.However, the cross-inoculation experiments revealed that all bra-dyrhizobial isolates induced ineffective nodules on the commonbean. Even though it was not tested whether these structures werepseudonodules or contained rhizobia, the rather unusual isolationof bradyrhizobia from common bean could also be explained bythe sampling of some ineffective nodules. In rare cases, the rhizo-bia did not effectively nodulate their original host plant under theselected conditions. For some peanut isolates, it was confirmedthat they were symbionts as they nodulated cowpea and Bambaragroundnut, which might be affected by the experimental growthconditions.
The bradyrhizobial population in the Okavango region ap-peared to be diverse, being divided into 15 groups based on un-gapped ITS sequence comparisons. While it is best to considermore than one gene for taxonomic inferences (53) with some-times-limited threshold suitability (54), bradyrhizobial strainswith more than 95.5% ungapped ITS sequence similarity usuallybelong to the same genospecies (44). However, a higher thresholdof 97% was chosen here, since the common genospecies cutoffvalue would have merged several species type strains in our se-quence comparisons. Although our analyses of physiologicalproperties were limited to maximum growth temperature, PEGtolerance, and host range, members of the same group mostlybehaved uniformly and could be clearly discriminated fromclosely related groups of isolates; whether they indeed representdifferent species needs to be elucidated in future. For example,isolates from cluster IV showed a maximal growth temperature of40°C, which was 35°C for cluster V and 38°C for cluster VI andVII, the latter two being differentiable by their susceptibility toPEG. ITS clusters XIII to XV were phenotypically similar as well,supported by being grouped together in core gene analysis. Dis-similar maximum growth temperatures of isolates 2-13 and 22 2-1supported their separation as indicated by phylogeny of house-keeping genes.
A clear, striking geographic distribution of bradyrhizobial spe-cies was uncovered. Except for one genotype (45 1-4), all Angolanbradyrhizobia were located on the B. elkanii major clade in thephylogenetic analyses, most of them being closely related to B.
pachyrhiziT. These isolates clustered together in all phylogeneticanalyses and behaved uniformly and identically to B. pachyrhiziT,indicating that they belong to the same species. Albeit promiscu-ous cowpea was grown and a high pedodiversity was observed(55), B. pachyrhizi appears to be a predominant species at thesampled sites of the Cusseque area in Angola. Reports on B. pachy-rhizi are rare, and not much is yet known about its distributionand ecology. A high proportion of B. pachyrhizi bacteria was de-tected under different land use systems in Mexican tropics usingcowpea and siratro (Macroptilium atropurpureum) as trap plants(56). The Mexican sampling sites were characterized by a warm,humid climate and acid soils (pH 4 to 6), strikingly matching thewarm, semihumid climate with an annual mean rainfall of 987mm and an annual mean temperature of 20.4°C (57) in theCusseque area. Almost all soils were also extremely acidic (pH 3.5to 4.4). Except for peaty wetland soils, they are characterized bysmall amounts of nitrogen and carbon (55). Even though the orig-inal host plant Pachyrhizus erosus (54) was usually present at thesampling sites in Mexico, these correlations indicate that B. pachy-rhizi is most competitive and can be preferentially found undersuch conditions, which is in line with its capability to grow at lowpH (pH 4.5) (54). In another report, Bradyrhizobium strainSEMIA 6099, a commercial inoculant strain in Brazil, was identi-fied as B. pachyrhizi (20). Possessing a core gene equipment wellmatching the environmental conditions, such effective B. pachy-rhizi strains might be the most promising candidates for the for-mulation of an inoculant for the Angolan Cusseque area. Interest-ingly, the B. pachyrizi isolates showed only low effectiveness in thecross-inoculation experiments on peanut, which was not culti-vated on the sampled fields.
In contrast, B. pachyrhizi was not found at the Namibian sam-pling sites in the Mashare area, which were characterized by asemiarid climate with a lower annual mean rainfall of 571 mm andhigher annual mean temperature of 22.3°C (32). Soils in theMashare area are mostly characterized by low nutrient content aswell, but higher pH variability, most being only slightly acidic (pHof approximately 4 to 6) (31). Commonly grown pulses are morediverse, with mostly cowpeas but also Bambara groundnuts andpeanuts. Concordantly, the Namibian bradyrhizobia were phylo-genetically and physiologically more diverse, with the bulk beingrelated to B. yuanmingense and B. daqingense. While B. daqingensewas described just recently (58) and information is limited, B.yuanmingense seems to be widespread in the subtropics and trop-ics and was found, e.g., in China (59), India and Mexico (23), Peru(60), Botswana (61), and South Africa (10). Moreover, the Na-mibian isolates were very diverse even within the groups, as couldbe seen in the BOX-PCR fingerprinting. The overall higher diver-sity among Namibian isolates might to some extent be explainedby a higher diversity of legume plants; however, most indigenousNamibian pulses obviously shared the same bradyrhizobial spe-cies (Table 1 or phylogenetic trees). Some influence on the nodu-lating bradyrhizobial population could be found in soil heteroge-neity and land use, but often members of one group originatedfrom different locations under different agricultural practices(Table 1). Semiarid conditions might be another relevant factoraccounting for the higher diversity. The diversity of bradyrhizo-bial nodulating populations was found to increase under water-limited conditions (62). Even though more diverse, almost all Na-mibian isolates shared the adaptational trait of being able to growat much higher temperatures than Angolan isolates and most spe-
Grönemeyer et al.
7254 aem.asm.org Applied and Environmental Microbiology
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

cies type strains. Remarkably, the only species type strains show-ing similar temperature adaptation in our tests, which were some-times inconsistent with the literature, were B. yuanmingenseT andB. daqingenseT, which might explain the frequent detection of B.yuanmingense in warm regions. The high temperature tolerance ofNamibian isolates of up to 40°C makes it a prerequisite for prom-ising inoculant strains in this region. In this regard, only inoculantstrain USDA3834, originally isolated in Nigeria, showed moderateadaptation to high temperatures and grew up to 35°C.
Despite the relatively high diversity in the overall spectrum ofisolates, none of them fell into the cluster of B. japonicum, B.diazoefficiens, B. cytisi, B. canariense, and B. liaoningense. Otherstudies already indicated that these species might be rare in sub-Saharan Africa, since Steenkamp et al. (61) detected neither B.japonicum, B. canariense, nor B. liaoningense in their survey oncowpea- and peanut-nodulating strains in Botswana and SouthAfrica. Furthermore, in a comprehensive study on rhizobia nod-ulating cowpea with nine genotypes conducted in Ghana, Bo-tswana, and South Africa, B. liaoningense was not detected and B.japonicum was found only on one variety grown in South Africa(10). In the former study, it was speculated that the absence of B.canariense and B. liaoningense in the isolate spectrum might be dueto their inability to nodulate cowpea and peanut. B. liaoningenseT
indeed failed to nodulate any of the local plant varieties tested inour cross-inoculation experiment. Also, the rather pH-neutralsoils which were sampled might be relevant for lack of B. liaonin-gense (61), since it is assumed that B. canariense and B. liaoningensetolerate acidic and prefer alkaline soils, respectively (22, 63). Thebroad range of soil pHs at our sampling sites indicates that thismight not be the only crucial factor. Besides the geographical im-pact on species abundance, an influence of host plants was ob-served only for hyacinth bean, which was, under natural condi-tions, exclusively nodulated by two bradyrhizobial species; eventhough the cross-inoculations revealed that the hyacinth bean canbe nodulated by a wide range of indigenous bradyrhizobia. Fur-thermore, these strains yielded the best nodulation rates on hya-cinth bean under laboratory conditions (data not shown) andwere only distantly related to all other bradyrhizobia. Hence, theymost probably represent yet-undescribed species being welladapted to hyacinth beans. Since the hyacinth bean was newlyintroduced to the Okavango region, one can assume that theseBradyrhizobium spp. came along with the seeds, which is in agree-ment with the high susceptibility of one of the groups (VIII) tohigh growth temperatures, while the other one (XI) showed mod-erate temperature tolerance. Moreover, their nifH sequences dis-tinguished them from those of almost all other Namibian isolates,depicting a different geographical origin of their sym locus. Espe-cially in view of the fact that most studied strains from sub-Saha-ran Africa shared relatively similar nodA sequences which are lo-cated on one clade in nodA phylogeny (61), this result isremarkable. However, uncommon nifH sequences were also ob-served for a third group of strains (IX), and the hyacinth beanisolates provoked good nodulation on all indigenous Okavangopulses, which was not the case for many other strains, especiallywith respect to peanut. As one of the two groups (VIII) could alsobe trapped from a nearby maize field soil, it may spread and/orpersist in Namibian soils for at least a short period of time. Thebetter competitiveness of these strains on hyacinth bean illustratesthe benefit of being well adapted to a respective local host plant,
which has to be taken into account when introducing exogenousstrains as inoculants.
Several of our groups behaved uniquely and separated wellfrom other species in the phylogeny, indicating that they representnovel species. Among these groups, especially the taxonomic po-sition of group IX needs more clarification. ITS sequence identityin GenBank (July 2014) was found to be maximal at 86%, and thephylogeny reconstruction resulted in an early and deep branchingfrom the B. elkanii lineage for both the ITS and the housekeepinggenes. The respective ITS sequences obviously interfered with areliable phylogeny reconstruction since removing them from theanalysis yielded a much better concerned bootstrap support (in-creased from 57 to 98). Thus, from the genotype it was not evencertain if these isolates are bradyrhizobia. However, partial 16SrRNA gene sequence identity to bradyrhizobial sequences inGenBank was 99.8%, and nifH phylogeny supports a recent lateraltransfer from bradyrhizobia. Since their physiology is typical ofthe genus Bradyrhizobium, they may represent a novel bradyrhi-zobial lineage apart from the established B. japonicum and B.elkanii lineages. The existence of more such lineages was also in-dicated in a recent study (64).
Most well-studied bradyrhizobial species which have beenknown for quite some time could be isolated from different con-tinents and climatic zones and are considered to be cosmopolitan(23). Since the geographical distribution of bradyrhizobia is facil-itated by anthropogenic long-distance dispersal via seeds and in-oculants, the statement “everything is everywhere, but, the envi-ronment selects” might be especially true for rhizobia. This wasalso observed in our study owing to the isolation of B. pachyrhizi-related strains from an African environment resembling its origi-nal isolation source in Mexico. Furthermore, the presumable in-vasion of a nonindigenous species via the introduction of hyacinthbean illustrates that altering the environmental conditions maylead to a different selection by the environment.
Many prominent species, however, were not detected in our andin other surveys on sub-Sahara African bradyrhizobia, implying thatthey are less competitive under the specific environmental condi-tions. In contrast, phylogenetic analyses and comparisons of our ITSsequences with the GenBank database indicate the presence of en-demic bradyrhizobial species: except for the isolates from groups VIIIand X and the Angolan groups clustering in proximity to B. elkaniiT
and B. pachyrhiziT, all our ITS sequences either showed low iden-tity to sequences in GenBank with 86 to 94% identity at the max-imum (VI, 93%; VII, �93%; IX, 86%; XI, �94%), or theymatched sequences obtained from other sub-Saharan countries atthe best (I, 99% Senegal; II, 99% Botswana; III, 100% Senegal; IV,100% Senegal; V, 98% Senegal). In the latter cases, the maximalidentity to non-sub-Saharan sequences was often rather low (I,94% India; II, 95% China; V, �90% India). Thus, either many ofour Namibian isolates may represent local genotypes or they be-long to genotypes widespread in, and potentially restricted to,sub-Saharan countries. Similar results were observed in a recentsurvey on Australian bradyrhizobia (64), where it was found thatmost isolates were distinct from non-Australian bradyrhizobia,presumably due to geographic isolation and plant endemicity.
The geographic isolation by the Sahara Desert may have provideda basis for allopatric speciation. Heterogeneous microclimates andedaphic factors may have resulted in an unknown, highly specializedendemic species diversity in sub-Saharan Africa even at a smallerscale. This situation is supported by the fact that almost all our Na-
Rhizobial Community Indigenous to the Okavango Region
December 2014 Volume 80 Number 23 aem.asm.org 7255
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

mibian strains share an exceptional high-temperature tolerance eventhough they can be differentiated genotypically and phenotypicallyamong themselves and other bradyrhizobial species. Recently, it wasfound that two cowpea rhizobial genotypes dominated in northernand southern Senegal, respectively, and it was hypothesized that bothrepresent novel species (62).
In conclusion, we observed a predominance of distinct geno-types which were to date solely found in sub-Saharan Africawhereat in some cases the geographic distribution may be evenmore local. In view of the assumption that “the environment se-lects,” these results imply that the indigenous bradyrhizobia arehighly adapted to exclusive environmental conditions and shouldthus be preferred for the formulation of an inoculant.
ACKNOWLEDGMENTS
This work was supported by grants by the BMBF (Federal Ministry ofEducation and Research) in the framework of the project “TFO: The fu-ture Okavango” (01LL0912G) and “SASSCAL: Southern African ScienceService Centre for Climate Change and Adaptive Land Management” (01LG 1201D) to B.R.-H. and T.H.
We thank Bruce Kasoana and Roberth Mukuya for help with collec-tion of pulses.
REFERENCES1. Pröpper M, Gröngröft A, Falk T, Eschenbach A, Fox T, Gessner U,
Hecht J, Hinz MO, Hoettich C, Hurek T, Kangombe FN, Keil M, KirkM, Clever M, Mills A, Mukuya R, Namwoonde NE, Overmann J,Petersen A, Reinhold-Hurek B, Schneiderat U, Strohbach BJ, Lück-Vogel M, Wisch U. 2010. Causes and perspectives of land-cover changethrough expanding cultivation in Kavango, p 2–31. In Jürgens N, Schmie-del U, Hoffman T (ed), Biodiversity in southern Africa, vol 3. Implicationsfor land use and management. Klaus Hess, Göttingen, Germany.
2. Grönemeyer J, Berkelmann D, Mubyana-John T, Haiyambo D, Chim-wamurombe P, Kasaona B, Hurek T, Reinhold-Hurek B. 2013. A surveyfor plant-growth-promoting rhizobacteria and symbionts associated withcrop plants in the Okavango region of Southern Africa. Biodivers. Ecol.5:287–294. http://dx.doi.org/10.7809/b-e.00282.
3. Dakora FD, Aboyinga RA, Mahama Y, Apaseku J. 1987. Assessment ofN2 fixation in groundnut (Arachis hypogaea L.) and cowpea (Vigna un-guiculata L. Walp) and their relative N contribution to a succeeding maizecrop in northern Ghana. MIRCEN J. 3:389 –399.
4. Ncube B, Twomlow SJ, van Wijk MT, Dimes JP, Giller KE. 2007.Productivity and residual benefits of grain legumes to sorghum undersemi-arid conditions in southwestern Zimbabwe. Plant Soil 299:1–15.http://dx.doi.org/10.1007/s11104-007-9330-5.
5. Roughley RJ. 1970. The influence of root temperature, Rhizobium strainand host selection on the structure and nitrogen-fixing efficiency of theroot nodules of Trifolium subterraneum. Ann. Bot. (Lond.) 34:631– 646.
6. Zhang H, Prithiviraj B, Charles TC, Driscoll BT, Smith DL. 2003. Lowtemperature tolerant Bradyrhizobium japonicum strains allowing im-proved nodulation and nitrogen fixation of soybean in a short season(cool spring) area. Eur. J. Agron. 19:205–213. http://dx.doi.org/10.1016/S1161-0301(02)00038-2.
7. Law IJ, Botha WJ, Majaule UC, Phalane FL. 2007. Symbiotic andgenomic diversity of ‘cowpea’ bradyrhizobia from soils in Botswana andSouth Africa. Biol. Fertil. Soils 43:653– 663. http://dx.doi.org/10.1007/s00374-006-0145-y.
8. Suzuki Y, Adhikari D, Itoh K, Suyama K. 2014. Effects of temperature oncompetition and relative dominance of Bradyrhizobium japonicum andBradyrhizobium elkanii in the process of soybean nodulation. Plant Soil374:915–924. http://dx.doi.org/10.1007/s11104-013-1924-5.
9. Botha WJ, Jaftha JB, Bloem JF, Habig JH, Law IJ. 2004. Effect of soilbradyrhizobia on the success of soybean inoculant strain CB 1809. Micro-biol. Res. 159:219 –231. http://dx.doi.org/10.1016/j.micres.2004.04.004.
10. Pule-Meulenberg F, Belane AK, Krasova-Wade T, Dakora FD. 2010.Symbiotic functioning and bradyrhizobial biodiversity of cowpea (Vignaunguiculata L. Walp.) in Africa. BMC Microbiol. 10:89. http://dx.doi.org/10.1186/1471-2180-10-89.
11. van Berkum P, Fuhrmann JJ. 2000. Evolutionary relationships among thesoybean bradyrhizobia reconstructed from 16S rRNA gene and internallytranscribed spacer region sequence divergence. Int. J. Syst. Evol. Microbiol.50:2165–2172. http://dx.doi.org/10.1099/00207713-50-6-2165.
12. Willems A, Coopman R, Gillis M. 2001. Comparison of sequence anal-ysis of 16S-23S rDNA spacer regions, AFLP analysis and DNA-DNA hy-bridizations in Bradyrhizobium. Int. J. Syst. Evol. Microbiol. 51:623– 632.
13. Jordan DC. 1982. Transfer of Rhizobium japonicum Buchanan 1980 toBradyrhizobium gen. nov., a genus of slow-growing, root nodule bacteriafrom leguminous plants. Int. J. Syst. Bacteriol. 32:136 –139. http://dx.doi.org/10.1099/00207713-32-1-136.
14. Kuykendall LD, Saxena B, Devine TE, Udell SE. 1992. Genetic diversityin Bradyrhizobium japonicum Jordan 1982 and a proposal for Bradyrhizo-bium elkanii sp. nov. Can. J. Microbiol. 38:501–505. http://dx.doi.org/10.1139/m92-082.
15. Xu LM, Ge C, Cui Z, Li J, Fan H. 1995. Bradyrhizobium liaoningense sp.nov., isolated from the root nodules of soybeans. Int. J. Syst. Bacteriol.45:706 –711. http://dx.doi.org/10.1099/00207713-45-4-706.
16. Vinuesa P, Rademaker JLW, Debruijn FJ, Werner D. 1998. Genotypiccharacterization of Bradyrhizobium strains nodulating endemic woodylegumes of the Canary Islands by PCR-restriction fragment length poly-morphism analysis of genes encoding 16S rRNA (16S rDNA) and 16S-23SrDNA intergenic spacers, repetitive extragenic palindromic PCR genomicfingerprinting, and partial 16S rDNA sequencing. Appl. Environ. Micro-biol. 64:2096 –2104.
17. Doignon-Bourcier F, Willems A, Coopman R, Laguerre G, Gillis M, deLajudie P. 2000. Genotypic characterization of Bradyrhizobium strainsnodulating small Senegalese legumes by 16S-23S rRNA intergenic genespacers and amplified fragment length polymorphism fingerprint analy-ses. Appl. Environ. Microbiol. 66:3987-3997. http://dx.doi.org/10.1128/AEM.66.9.3987-3997.2000.
18. Stepkowski T, Moulin L, Krzyzanska A, McInnes A, Law IJ, HowiesonJ. 2005. European origin of Bradyrhizobium populations infecting lupinsand serradella in soils of Western Australia and South Africa. Appl. Envi-ron. Microbiol. 71:7041–7052. http://dx.doi.org/10.1128/AEM.71.11.7041-7052.2005.
19. Vinuesa P, Silva C, Werner D, Martínez-Romero E. 2005. Populationgenetics and phylogenetic inference in bacterial molecular systematics: theroles of migration and recombination in Bradyrhizobium species cohesionand delineation. Mol. Phylogenet. Evol. 34:29 –54. http://dx.doi.org/10.1016/j.ympev.2004.08.020.
20. Menna P, Barcellos FG, Hungria M. 2009. Phylogeny and taxonomy of adiverse collection of Bradyrhizobium strains based on multilocus sequenceanalysis of the 16S rRNA gene, ITS region and glnII, recA, atpD and dnaKgenes. Int. J. Syst. Evol. Microbiol. 59:2934 –2950. http://dx.doi.org/10.1099/ijs.0.009779-0.
21. Parté AC. 2014. LPSN—list of prokaryotic names with standing in no-menclature. Nucleic Acids Res. 42:D613–D616. http://dx.doi.org/10.1093/nar/gkt1111.
22. Adhikari D, Kaneto M, Itoh K, Suyama K, Pokharel BB, Gaihre YK.2012. Genetic diversity of soybean-nodulating rhizobia in Nepal in rela-tion to climate and soil properties. Plant Soil 357:131–145. http://dx.doi.org/10.1007/s11104-012-1134-6.
23. Vinuesa P, Rojas-Jiménez K, Contreras-Moreira B, Mahna SK, PrasadBN, Moe H, Selvaraju SB, Thierfelder H, Werner D. 2008. Multilocussequence analysis for assessment of the biogeography and evolutionarygenetics of four Bradyrhizobium species that nodulate soybeans on theAsiatic continent. Appl. Environ. Microbiol. 74:6987– 6996. http://dx.doi.org/10.1128/AEM.00875-08.
24. Risal CP, Yokoyama T, Ohkama-Ohtsu N, Djedidi S, Sekimoto H. 2010.Genetic diversity of native soybean bradyrhizobia from different topo-graphical regions along the southern slopes of the Himalayan Mountainsin Nepal. Syst. Appl. Microbiol. 33:416 – 425. http://dx.doi.org/10.1016/j.syapm.2010.06.008.
25. Sullivan JT, Ronson CW. 1998. Evolution of rhizobia by acquisition of a500-kb symbiosis island that integrates into a phe-tRNA gene. Proc. Natl.Acad. Sci. U. S. A. 95:5145–5149. http://dx.doi.org/10.1073/pnas.95.9.5145.
26. Perret X, Staehelin C, Broughton WJ. 2000. Molecular basis of symbioticpromiscuity. Microbiol. Mol. Biol. Rev. 64:180 –201. http://dx.doi.org/10.1128/MMBR.64.1.180-201.2000.
27. Yang S, Tang F, Gao M, Krishnan HB, Zhu H. 2010. R gene-controlled
Grönemeyer et al.
7256 aem.asm.org Applied and Environmental Microbiology
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from

host specificity in the legume-rhizobia symbiosis. Proc. Natl. Acad. Sci.U. S. A. 107:18735–18740. http://dx.doi.org/10.1073/pnas.1011957107.
28. Devine TE, Kuykendall LD. 1996. Host genetic control of symbiosis insoybean (Glycine max L.). Plant Soil 186:173–187. http://dx.doi.org/10.1007/BF00035072.
29. Parker MA. 1999. Mutualism in metapopulations of legumes and rhizo-bia. Am. Nat. 153:S48 –S60. http://dx.doi.org/10.1086/303211.
30. van Berkum P. 1990. Evidence for a third uptake hydrogenase phenotypeamong the soybean bradyrhizobia. Appl. Environ. Microbiol. 56:3835–3841.
31. Gröngröft A, Luther-Mosebach J, Landschreiber L, Eschenbach A. 2013.Mashare soils. Biodivers. Ecol. 5:105–108. http://dx.doi.org/10.7809/b-e.00259.
32. Weber T. 2013. Mashare climate. Biodivers. Ecol. 5:103–104. http://dx.doi.org/10.7809/b-e.00258.
33. Jensen HL. 1942. Nitrogen fixation in leguminous plants. I. General char-acters of non-nodule bacteria isolated from species of Medicago and Tri-folium in Australia. Proc. Linn. Soc. N. S. W. 67:98 –108.
34. Laguerre G, Mavingui P, Allard MR, Charnay MP, Louvrier P, Mazu-rier SI, Rigottier-Gois L, Amarger N. 1996. Typing of rhizobia by PCRDNA fingerprinting and PCR-restriction fragment length polymorphismanalysis of chromosomal and symbiotic gene regions—application to Rhi-zobium leguminosarum and its different biovars. Appl. Environ. Micro-biol. 62:2029 –2036.
35. Poly F, Monrozier LJ, Bally R. 2001. Improvement in the RFLP proce-dure for studying the diversity of nifH genes in communities of nitrogenfixers in soil. Res. Microbiol. 152:95–103. http://dx.doi.org/10.1016/S0923-2508(00)01172-4.
36. Demba Diallo M, Willems A, Vloemans N, Cousin S, Vandekerckhove TT,de Lajudie P, Neyra M, Vyverman W, Gillis M, Van der Gucht K. 2004.Polymerase chain reaction denaturing gradient gel electrophoresis analysis ofthe N2-fixing bacterial diversity in soil under Acacia tortilis ssp. raddiana andBalanites aegyptiaca in the dryland part of Senegal. Environ. Microbiol.6:400 – 415. http://dx.doi.org/10.1111/j.1462-2920.2004.00577.x.
37. Versalovic J, Schneider M, de Bruijn FJ, Lupski JR. 1994. Genomicfingerprinting of bacteria using repetitive sequence-based polymerasechain reaction. Methods Mol. Cell. Biol. 5:25– 40.
38. Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA,McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, ThompsonJD, Gibson TJ, Higgins DG. 2007. Clustal W and Clustal X version 2.0.Bioinformatics 23:2947–2948. http://dx.doi.org/10.1093/bioinformatics/btm404.
39. Edgar RC. 2004. MUSCLE: multiple sequence alignment with high accu-racy and high throughput. Nucleic Acids Res. 32:1792–1797. http://dx.doi.org/10.1093/nar/gkh340.
40. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011.MEGA5: molecular evolutionary genetics analysis using maximum likeli-hood, evolutionary distance, and maximum parsimony methods. Mol.Biol. Evol. 28:2731–2739. http://dx.doi.org/10.1093/molbev/msr121.
41. Posada D, Crandall KA. 1998. MODELTEST: testing the model ofDNA substitution. Bioinformatics 14:817– 818. http://dx.doi.org/10.1093/bioinformatics/14.9.817.
42. Schwarz G. 1978. Estimating the dimension of a model. Ann. Stat. 6:461–464. http://dx.doi.org/10.1214/aos/1176344136.
43. Felsenstein J. 1985. Confidence limits on phylogenies: an approach usingthe bootstrap. Evolution 39:783–791. http://dx.doi.org/10.2307/2408678.
44. Willems A, Munive A, de Lajudie P, Gillis M. 2003. In most Bradyrhi-zobium groups sequence comparison of 16S-23S rDNA internal tran-scribed spacer regions corroborates DNA-DNA hybridizations. Syst.Appl. Microbiol. 26:203–210. http://dx.doi.org/10.1078/072320203322346056.
45. Aserse AA, Räsanen LA, Aseffa F, Hailemariam A, Lindström K. 2012.Phylogenetically diverse groups of Bradyrhizobium isolated from nodulesof Crotalaria spp., Indigofera spp., Erythrina brucei and Glycine max grow-ing in Ethiopia. Mol. Phylogenet. Evol. 65:595– 609. http://dx.doi.org/10.1016/j.ympev.2012.07.008.
46. Menna P, Hungria M. 2011. Phylogeny of nodulation and nitrogen-fixation genes in Bradyrhizobium: supporting evidence for the theory ofmonophyletic origin, and spread and maintenance by both horizontal andvertical transfer. Int. J. Syst. Evol. Microbiol. 61:3052–3067. http://dx.doi.org/10.1099/ijs.0.028803-0.
47. Zhang XX, Guo HJ, Wang R, Sui XH, Zhang YM, Wang ET, Tian CF,Chen WX. 2014. Genetic divergence of Bradyrhizobium strains nodulat-
ing soybeans as revealed by multilocus sequence analysis of genes insideand outside the symbiosis islands. Appl. Environ. Microbiol. 80:3181–3190. http://dx.doi.org/10.1128/AEM.00044-14.
48. Kishinevsky BD, Sen D, Weaver RW. 1992. Effect of high root temper-ature on Bradyrhizobium-peanut symbiosis. Plant Soil 143:275–282. http://dx.doi.org/10.1007/BF00007883.
49. Michiels J, Dombrecht B, Vermeiren N, Xi C, Luyten E, VenderleydenJ. 1998. Phaseolus vulgaris is a non-selective host for nodulation. FEMSMicrobiol. Ecol. 26:193–205. http://dx.doi.org/10.1111/j.1574-6941.1998.tb00505.x.
50. Mpepereki S, Wollum AG, Makonese F. 1996. Diversity in symbioticspecificity of cowpea rhizobia indigenous to Zimbabwean soils. Plant Soil186:167–171. http://dx.doi.org/10.1007/BF00035071.
51. Martins LMV, Neves MCP, Rumjanek NG. 1997. Growth characteristicsand symbiotic efficiency of rhizobia isolated from cowpea nodules of thenorth-east region of Brazil. Soil Biol. Biochem. 29:1005–1010. http://dx.doi.org/10.1016/S0038-0717(96)00215-5.
52. Zhang WT, Yang JK, Yuan TY, Zhou JC. 2007. Genetic diversity andphylogeny of indigenous rhizobia from cowpea [Vigna unguiculata (L.)Walp.]. Biol. Fertil. Soils 44:201–210. http://dx.doi.org/10.1007/s00374-007-0196-8.
53. Rivas R, Martens M, de Lajudie P, Willems A. 2009. Multilocus se-quence analysis of the genus Bradyrhizobium. Syst. Appl. Microbiol. 32:101–110. http://dx.doi.org/10.1016/j.syapm.2008.12.005.
54. Ramírez-Bahena MH, Peix A, Rivas R, Camacho M, Rodríguez-NavarroDN, Mateos PF, Martinez-Molina E, Willems A, Vélazquez E. 2009.Bradyrhizobium pachyrhizi sp. nov. and Bradyrhizobium jicamae sp. nov.,isolated from effective nodules of Pachyrhizus erosus. Int. J. Syst. Evol.Microbiol. 59:1929 –1934. http://dx.doi.org/10.1099/ijs.0.006320-0.
55. Gröngröft A, Luther-Mosebach J, Landschreiber L, Eschenbach A. 2013.Cusseque soils. Biodivers. Ecol. 5:51–54. http://dx.doi.org/10.7809/b-e.00245.
56. Ormeño-Orrillo E, Rogel-Hernández MA, Lloret L, López-López A,Martínez J, Barois I, Martínez-Romero E. 2012. Change in land usealters the diversity and composition of Bradyrhizobium communitiesand led to the introduction of Rhizobium etli into the tropical rain forest ofLos Tuxtlas (Mexico). Microb. Ecol. 63:822– 834. http://dx.doi.org/10.1007/s00248-011-9974-9.
57. Weber T. 2013. Cusseque climate. Biodivers. Ecol. 5:45– 46. http://dx.doi.org/10.7809/b-e.00243.
58. Wang JY, Wang R, Zhang YM, Liu HC, Chen WF, Wang ET, Sui XH,Chen WX. 2013. Bradyrhizobium daqingense sp. nov., isolated from soy-bean nodules. Int. J. Syst. Evol. Microbiol. 63:616 – 624. http://dx.doi.org/10.1099/ijs.0.034280-0.
59. Yao ZY, Kan FL, Wang ET, Wei GH, Chen WX. 2002. Characterizationof rhizobia that nodulate legume species of the genus Lespedeza and de-scription of Bradyrhizobium yuanmingense sp. nov. Int. J. Syst. Evol.Microbiol. 52:2219 –2230. http://dx.doi.org/10.1099/ijs.0.01408-0.
60. Ormeño-Orrillo E, Vinuesa P, Zúñiga-Davila D, Martínez-Romero E.2006. Molecular diversity of native bradyrhizobia isolated from lima bean(Phaseolus lunatus L.) in Peru. Syst. Appl. Microbiol. 29:253–262. http://dx.doi.org/10.1016/j.syapm.2005.09.002.
61. Steenkamp ET, Stepkowski T, Przymusiak A, Botha WJ, Law IJ. 2008.Cowpea and peanut in southern Africa are nodulated by diverse Bradyrhi-zobium strains harboring nodulation genes that belong to the large pan-tropical clade common in Africa. Mol. Phylogenet. Evol. 48:1131–1144.http://dx.doi.org/10.1016/j.ympev.2008.04.032.
62. Krasova-Wade T, Ndoye I, Braconnier S, Sarr B, de Lajudie P, Neyra M.2003. Diversity of indigenous bradyrhizobia associated with three cowpeacultivars (Vigna unguiculata (L.) Walp.) grown under limited and favour-able water conditions in Senegal (West Africa). Afr. J. Biotechnol. 2:13–22.
63. Vinuesa P, Léon-Barrios M, Silva C, Willems A, Jarabo-Lorenzo A, Pérez-Galdona R, Werner D, Martínez-Romero E. 2005. Bradyrhizobium canar-iense sp. nov., an acid-tolerant endosymbiont that nodulates endemicgenistoid legumes (Papilionoideae: Genisteae) from the Canary Islands,along with Bradyrhizobium japonicum bv. genistearum, Bradyrhizobiumgenospecies alpha and Bradyrhizobium genospecies beta. Int. J. Syst. Evol.Microbiol. 55:569 –575. http://dx.doi.org/10.1099/ijs.0.63292-0.
64. Stepkowski T, Watkin E, McInnes A, Gurda D, Gracz J, Steenkamp ET.2012. Distinct Bradyrhizobium communities nodulate legumes native totemperate and tropical monsoon Australia. Mol. Phylogenet. Evol. 63:265–277. http://dx.doi.org/10.1016/j.ympev.2011.12.020.
Rhizobial Community Indigenous to the Okavango Region
December 2014 Volume 80 Number 23 aem.asm.org 7257
on October 1, 2018 by guest
http://aem.asm
.org/D
ownloaded from