review article computational insights into...
TRANSCRIPT

Review ArticleComputational Insights into Substrate and Site SpecificitiesCatalytic Mechanism and Protonation States of the CatalyticAsp Dyad of 120573-Secretase
Arghya Barman and Rajeev Prabhakar
Department of Chemistry University of Miami 1301 Memorial Drive Coral Gables FL 33146 USA
Correspondence should be addressed to Rajeev Prabhakar rprmiamiedu
Received 5 March 2014 Accepted 21 June 2014 Published 18 September 2014
Academic Editor Christoph Gabler
Copyright copy 2014 A Barman and R PrabhakarThis is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in anymedium provided the originalwork is properly cited
In this review information regarding substrate and site specificities catalytic mechanism and protonation states of the catalyticAsp dyad of 120573-secretase (BACE1) derived from computational studies has been discussed BACE1 catalyzes the rate-limiting stepin the generation of Alzheimer amyloid beta peptide through the proteolytic cleavage of the amyloid precursor protein Due to itsbiological functioning this enzyme has been considered as one of the most important targets for finding the cure for Alzheimerrsquosdisease Molecular dynamics (MD) simulations suggested that structural differences in the key regions (inserts A D and F andthe 10s loop) of the enzyme are responsible for the observed difference in its activities towards the WT- and SW-substrates Themodifications in the flap third strand and insert F regions were found to be involved in the alteration in the site specificity ofthe glycosylphosphatidylinositol bound form of BACE1 Our QM and QMMM calculations suggested that BACE1 hydrolyzed theSW-substrate more efficiently than the WT-substrate and that cleavage of the peptide bond occurred in the rate-determining stepThe results from molecular docking studies showed that the information concerning a single protonation state of the Asp dyad isnot enough to run an in silico screening campaign
1 Introduction
Alzheimerrsquos disease (AD) is a progressive neurological disor-der and common cause of dementia which affects 4ndash8 ofthe elderly population worldwide Scientific efforts to slowthe progression of AD could save society $ 12 trillion to$ 397 trillion a year by 2050 in cost The formation ofextracellular amyloid (or senile) plaques and intracellularneurofibrillary tangles at the hippocampus and cortical greyregions of the brain are considered to be the pathologicalhallmark for this life threatening disease It has also beenpostulated that plaques are not neurotoxic and more solublepartly aggregated complexes are the main cause of thisdisease These amyloid plaques are developed due to theaggregation and deposition of 1ndash4042 amino acid containingsmall peptides known as amyloid beta (A120573) peptide(s) [1 2]Inside the brain A120573peptide(s) are produced by the sequential
cleavage of the membrane bound amyloid precursor protein(APP) by two extremely critical enzymes called 120573-secretase(BACE1) and 120574-secretase
BACE1 belongs to a ubiquitous family of aspartyl pro-teases [3ndash6] which catalyzes the hydrolytic cleavage of a largenumber (sim80) of proteins that play critical roles in severalcellular and subcellular pathways [7 8] BACE1 cleaves theMet671-Asp672 amide bond of amyloid precursor protein(APP) at the extracellular space and initiates the formationof A120573 peptide(s) [9ndash11] In addition BACE1 can cleave adouble mutant (Lys670 rarr Asn and Met671 rarr Leu) of APPknown as the Swedish mutant (SW) with sixty times higherefficiency than the WT-substrate [12] Since BACE1 plays acritical role in the rate-limiting steps of A120573 production theinhibition of this enzyme became widely acknowledged toregulate the production of A120573 peptide(s) [13] Based on awealth of experimental data including genetics biochemical
Hindawi Publishing CorporationScientificaVolume 2014 Article ID 598728 11 pageshttpdxdoiorg1011552014598728
2 Scientifica
studies and animalmodeling this enzyme has been proposedto be a very promising target for the treatment of AD[9ndash14]
Structurally the ectodomain of BACE1 is composed ofseveral subregions that control the entry and orientation ofthe substrate at the active site The active site of BACE1contains two conserved Asp residues [15] that forms thecatalytic dyad This dyad has been implicated in the catalyticfunctioning of the entire family of aspartyl proteases includ-ing pepsin renin cathepsin D and HIV protease [16ndash23]The catalytic Asp dyad at the active site is covered over byan antiparallel hairpin-loop known as flap The mechanismsof flap closing and catalysis are of great significance due totheir involvement in human diseases such as AD During thecatalytic cycle the flap must open to allow the entrance ofthe substrate into the active site cleft and steer it towards thecatalytic Asp dyad to attain a reactive conformation In thisconformation the specific peptide bond(s) of the substrateis hydrolytically cleaved This type of gating mechanism hasbeen reported to be utilized by most of the members of theaspartyl protease family The dyad utilizes a general acid-base mechanism for the catalysis of peptide hydrolysis [17]Although theoretical X-ray and neutron diffraction datashow that the Asp residues in the catalytic dyad can switchprotonation states during the catalytic turnover [24ndash26] theeffect of the protonation states of these residues still remainsan intriguing issue in the development of successful drugdesigning strategies
Although experimental techniques can provide a greatdeal of information about the catalytic mechanism andprotonation states of the ionizable residues the atomic leveldescription of these complex chemical transformations is stillbeyond their reach These limitations can be overcome bymodern computational chemistry methods that can describethese complex processes at the atomistic level Computationalchemistry is a well-established field which has become anindispensable tool to study complex chemical and biochem-ical systems In the last decade applications of the densityfunctional theory (DFT) to study the chemical reactionsusing small systems have dominated the field Howeverthe main caveat of using DFT is the restricted number ofatoms (sim200) Therefore to study the larger system DFT(QM) has been coupled to the molecular mechanics (MM)potentials and implemented in hybrid QMMM(ONIOM)[27ndash33] method to study the catalytic mechanisms of theenzymes Applications of QMMM in biological systemshave shown tremendous success [34] On the other handmolecular dynamics (MD) simulations have become anessential part of current research to study the phase spacebehavior conformational changes of molecules and to cal-culate thermodynamic properties of systems [35 36] Alongwith MD MM based scoring functions have also beenincorporated into the docking engines andhave gained exten-sive use in modern computer aided drug design protocol[37]
In this review the current knowledge of structural andfunctional aspects of BACE1 in atomistic detail using multi-dimensional computational methods has been discussed
2 Structural Characteristics of BACE1
To date more than 290 crystal structures of BACE1 (Apoform and cocrystal with inhibitors) have been deposited intothe protein data bank (PDB) However the first cocrystalstructure of BACE1 with a hydroxyethylene (HE) basedtransition state isostere (OM99-2 and OM00-3) revealed thefirst evidence of BACE1 active site that contained the catalyticdyad (Asp32 and Asp228) at the center of the active site[15 38]The globular nature of BACE1 can be divided into N-andC-terminal domainsTheflap of this enzyme is composedof eleven residues (Val67-Glu77) and is positioned at theN-terminal domain A conserved Tyr71 residue is locatedat the flap which is found to adopt different conformationsin the presence and absence of inhibitors (Figure 1) Thereare several key functional regions namely 10s loop (Lys9-Tyr14) third strand (Lys107-Gly117) and insert A (Gly158-Leu167) that are present at the N-terminus Whereas insert B(Lys218-Asn221) insert C (Ala251-Pro258) insert D (Trp270-Thr274) insert E (Glu290-Ser295) and insert F (Asp311-Asp317) regions are located at the C-terminus these regionsfacilitate the entry and binding of different substrates atthe active site through their movements [15 39] At theactive site BACE1 contains two conserved water molecules(WAT1 and WAT2) After carefully analyzing 82 cocrystalstructures of aspartyl proteases the specific role of thesetwo water molecules was suggested by Andreeva and Rumsh[40] The WAT1 water located near the catalytic Asp dyadwas assigned to be most important as it is utilized in thehydrolytic cleavage of the peptide bond The second watermolecule (WAT2) participates in a continuous H-bondingnetwork and stabilizes the flap in the closed conformationthrough structural organization
3 Differential Substrate Specificity of BACE1
The induced fit model for enzyme-substrate interactions[41] suggests that the interaction of the substrate with anenzyme modulates the spatial orientation of the enzymersquosactive site residues into an optimal and precise conformationto accommodate the substrate Conformational preference ofthe enzyme-substrate complex is driven by the dynamics ofboth the enzyme and substrate It is not possible to resolve X-ray structure of a wild type- (WT-) enzyme in active formbound to substrate In the absence of this structure theatomistic level understanding of enzyme-substrate interac-tions remains elusive We have studied the role of proteindynamics in the formation of enzyme-substrate complex andsubstrate specificity of BACE1 involved in the generation ofA120573-peptides [23]
BACE1 is known to cleave the SW-substrate (Glu-Val-Asn-Leu-Asp-Ala-Glu-Phe) sixty times more efficiently thanthe WT-substrate (Glu-Val-Lys-Met-Asp-Ala-Glu-Phe) Weexplored the interaction profiles of BACE1 with the WT- andSW-substrates through 20 ns all-atom MD simulations in aTIP4P explicit water system using the OPLS-AA [42] forcefield Four different structures (WT-BACE1 SW-BACE1 anda 4-phenoxypyrrolidine-based BACE-1 inhibitor (compound11 PDB ID 2qmg [43]) bound BACE1) [44] along with the
Scientifica 3
Open flap
Closed flap
Tyr71
WAT2WAT1
Asp228 Asp32
C-terminal domain
Insert C
Insert E
Insert BInsert D
Insert F
Insert A
10s loop
Flap
N-terminal domain
Figure 1 Critical regions of BACE1 that participate in the substrate recognition catalytic Asp dyad conserved waters (WAT1 and WAT2)and orientation of Tyr71 in open and closed conformation of the flap
apo form of BACE1 were studied In order to capture thedynamic behavior of substrate recognition by the enzyme thestarting models were generated by placing the substrates andthe inhibitor inside the apo form of BACE1 (PDB ID1w50)[15 45] The representative structure derived from the com-pound 11-BACE1 simulations shows an accurate reproductionof the crystallographic orientation of the flap the 10s loopand BACE1- inhibitor interactions The root-mean-square(rms) deviation between the simulated and X-ray structureswas only 13 AThe simulation therefore served as a validationof the MD method and the parameters that were used forthe substrate bound BACE1 In the presence of the inhibitorand the substrate the flap closing event (Figure 2) occurredaround 3 ns and two water molecules facilitated the processAccording to a site-directed mutagenesis study electrostaticinteractions between the Glu residue of the substrate andArg307 residue of BACE1 play important roles in enhancingthe catalytic efficiency of the enzyme [46] In the BACE1-WTsimulation this interaction was lost due to the formation ofintramolecular salt-bridge between the Glu residue and theLys residue of the substrate (Figure 2) Therefore the lossof the Glu-Arg307 interaction was considered as one of thecauses for the diminished activity of the enzyme with theWT-substrate A comparison between three critical topologi-cal distances (C120572(Thr72)-C120573(Asp32) C120572(Thr72)-C120572(Thr329)and OG1(Thr72)-NH1(Arg235)) and the volume of the active
site showed that in comparison to theWT-substrate the flapwas more closed and the active site was more constrictedupon the binding of the SW-substrate In addition the num-ber of hydrogen bonds formed by the SW-substratewas foundto be two times (8ndash10) more than the bonds contributedby the WT-substrate The computed electrostatic bindingenergy of the SW-substrate was found to be 19 kcalmolmorefavorable than the WT-substrate In addition to substrate-enzyme interactions structures of inserts A D and F and the10s loop of the enzyme were also substantially different uponthe binding of these two substrates Overall these structuraldifferences between the WT- and SW-substrate explicitlyindicate that BACE1 demonstrated greater affinity for the SW-substrate and positioned it in a more bioactive conformationcompared to the WT-substrate
4 Differential Site Specificity of BACE1
In addition to the cleavage of Met-Asp amide bond of WT-APP at +1 site or 120573 site BACE1 can cleave the peptide bondbetween Tyr-Glu at +11 or 1205731015840 site and produce +11 A120573 speciesThis site specificity can be altered through themodification ofC-terminal domain of BACE1 Glycosylphosphatidylinositol(GPI) bound form of BACE1 (BACE1-GPI chimera) wasshown to have more 120573 site cleavage specificity compared to
4 Scientifica
2
18
16
14
12
1
0 5 10 15 20
Time (ns)
Dist
ance
(nm
)
(a)
0 5 10 15 20
Time (ns)
16
14
12
10
8
6
4
2
0
Num
ber o
f hyd
roge
n bo
nds
(b)
Lys321
Arg307
WT
Glu(P
Glu(P4)
4)
Asn233
Asn233Lys(P2)
Lys321
Val(P3)Val(P3)
SW
Arg307
Asn(P2)
(c)
Figure 2 (a) Time evolution of the topological distance between the C120572(Thr72) of flap tip and C120573(Asp32) of catalytic dyad (violet BACE1-compound 11 blue BACE1-SW substrate red BACE1-WT substrate and green Apo BACE1) (b) Number of hydrogen bonds formed betweenthe substrates and BACE1 (blue BACE1-SW substrate red BACE1-WT substrate) (c) Interaction of the substrate Glu with the Arg307 ofBACE1
the1205731015840 site OurMD simulation studies onBACE1 andBACE1-GPI showed that GPI could introduce significant changes inthe structure of BACE1The structural changes are associatedwith the flap third strand and insert F of the enzyme In thepresence of GPI the flapmoved to amore open conformationand strong hydrogen bond interactions between the thirdstrand and insert F brought them closer to each otherOn the other hand the flap remains in a relatively closedconformation and the two segments moved apart due to theloss of the hydrogen bond interactions in the WT-BACE1Overall these results suggest that GPI induces structuralmodifications thatmay regulate the site recognition of BACE1[47]
5 Catalytic Mechanism of BACE1
BACE1 cleaves specific peptide bonds of the substrates uti-lizing the catalytic Asp dyad This Asp dyad interacts withthe conserved water molecule (WAT1) present in the activesite and utilizes it for peptide hydrolysis Recent X-ray andneutron diffraction data and theoretical calculations on theaspartyl proteases show that during the catalytic cycle oneof the Asp residues is protonated and the second one isunprotonated [24ndash26 48] BACE1 catalyzes the hydrolytic
cleavage of the Met-Asp and Leu-Asp peptide bonds of theWT- and SW-substrate respectively
In the catalytic cycle BACE1 utilizes a two-step generalacidbasemechanism to cleave these peptide bonds (Figure 3)[17] In the first step from the reactant (I) the unprotonatedAsp228 functions as a base and abstracts a proton from thecatalytic water (WAT1) to generate a hydroxyl ion (OHminus)which subsequently makes a nucleophilic attack on thecarbonyl carbon of the peptide bond In this process Asp32acts as an acid and concomitantly donates its proton to thecarbonyl oxygen atom of the scissile peptide bond to generatethe gem-diol intermediate (II) In II two hydroxyl groups arecoordinated to the carbonyl carbon atomof the peptide bondIn the next step the two aspartate residues exchange theirfunctional roles Asp32 now functions as a base and abstractsa proton from the hydroxyl group (ndashOH) of II Here Asp228which initially acted as a base and became protonated in theprevious step plays the role of an acid by donating its protonto the amide nitrogen atom (ndashNH) of the scissile peptidebondThe cleavage process of the peptide bond occurs in thisprocess and generates separated amine (ndashNH
2) and carboxyl
(ndashCOOH) terminals (III)The hydrolysis of both WT- and SW-substrates was
investigated using two computational methods In the first
Scientifica 5
O
OH
H
O O
H
HN
O
HOOC HOOC
O
HOOC
O O
O H
H
O
H
HN
O
NH H
O H
O O
HO
I II III
Asp32 Asp32 Asp32
Asp228 Asp228 Asp228
NH2 NH2NH2
R1 R1
R2 R2R2
Ominus
Ominus
Ominus
C C C
R1
Figure 3 General acid-base mechanism utilized for peptide hydrolysis
ONIOMPruned model
196
219
00
50
100
150
200
250
00
Ener
gy (k
calm
ol)
Reaction coordinate
147
224
159171
minus32
minus57I TSI II TSII III
WT-substrate
(a)
ONIOMPruned model
161 172
00
50
100
150
200
250
00
Reaction coordinate
100
191
82
98
minus16
minus52I TSI II TSII III
SW-substrate
Ener
gy (k
calm
ol)
(b)
Figure 4 Computed barriers for the WT- and SW-substrate using pruned models (with the DFT method) and entire protein (with theONIOM (QMMM) method)
step of this study the most representative structures derivedfrom the BACE1-WT and BACE1-SW MD simulations wereutilized to develop pruned models of the enzyme-substratecomplexes In the second step these structures were used toinvestigate hydrolytic cleavage mechanisms through densityfunctional theory (DFT) [23] In the second study a moreaccurate hybrid QMMM (ONIOM) method was applied tostudy these mechanisms by including the whole protein inmodels [49]
According to the DFT study the formation of the gem-diol intermediate in the first step is the rate-determining step
of the entire mechanism for both substrates The reactionproceeds through a barrier of 224 kcalmol and 191 kcalmol(TSI) for the WT- and SW-substrate respectively Thegem-diol intermediate (II) for the SW-substrate was foundto be 80 kcalmol lower in energy than the one for theWT-substrate The stabilization of this intermediate is alsoreflected during the cleavage of the peptide bond throughtransition state TSII The cleavage of the peptide bond pro-ceeds through a barrier of 171 and 98 kcalmol for the WT-and SW-substrates respectivelyThe potential energy surface(PES) diagrams for these reactions are shown in Figure 4
6 Scientifica
Table 1 Ligands with chemical structure and PDB ID of cocrystal structure of BACE1
Ligands Chemical structures PDB ID
L1 (HE)
O
O
OS
O O
OOH
NH
NH H
NH
N
N2P4J
L2 (AE)O O O
F
NH2
N NH
NH
2FDP
L3 (TC)
O
OO
O
O
OH
N
NH
NH2
F
S
2IS0
L4 (CA)O
O
OHO
HN
HN
F
F
N
2QMF
L5 (RA)
O O O
O
O
NH
NN
H
HN
N
H
S
F
2QZL
L6 (AK)
O
O
OO
NNH HO
OH
S
N
NH 2WF4
Scientifica 7
Table 1 Continued
Ligands Chemical structures PDB ID
L7 (ABP)
O O
OO
Cl
OO
N
N
HS
NH2
2ZJM
L8 (AP)
O
N
HN
H2N
3HVG
The structural differences in the microenvironment of theactive site play a critical role in lowering the barrier for theSW-substrate Although the prunedmodels accurately repro-duced the experimentally observed barrier ca 180 kcalmolfor both substrates [50] contribution of the steric and elec-trostatic effects of the protein surrounding on the computedenergetics of the reaction was completely ignored Thereforeto incorporate the effect of the protein environment on theenergetics the hybrid QMMM (ONIOMB3LYPAmber)method was applied to investigate the hydrolytic cleavageof the WT- and SW-substrate The PES diagrams for thesereactions are shown in Figure 4 In this case the formationof the gem-diol intermediate occurs with barriers of 196 and161 kcalmol for the WT- and SW-substrates respectivelyThe inclusion of the electrostatic and steric effects of thesurrounding protein in the model reduced the barrier of thisstep by sim30 kcalmol for both substrates These reductionsare observed due to the alterations in the reaction coordinatesand microenvironment of the active site and are not due tothe long range structural reorganization in the enzyme Thecreation of the gem-diol intermediate in this step for the SW-substrate (100 kcalmol) is 47 kcalmol more favorable thanfor theWT-substrate (147 kcalmol)The cleavage of the pep-tide bond of the WT-substrate occurs through a calculatedbarrier of 219 kcalmol For the SW-substrate the barrier forthis process is reduced by 47 kcalmol that is 172 kcalmolfrom the reactant Results from theDFT andQMMMstudiesindicate that the formation of the gem-diol intermediateis the rate determining step of the entire mechanism forboth substrates The recent crystallographic [51] and Car-Parrinello MD simulations [20] studies on another memberof the aspartyl protease family HIV protease also suggestedthe cleavage of the peptide bond to be the rate-limiting stepIn our QMMM calculations the rate-limiting step for theSW-substrate is hard to distinguish due to a small difference(11 kcalmol) in barriers between the two steps The overall
reduction in the barrier for each step in the SW-substrate isalso in linewith the experimental observation [50] suggestingthat BACE1 can cleave the SW-substrate more efficiently thantheWT-substrateThese studies explicitly provided an atomiclevel understanding of the complex mechanism of peptidebond cleavage by BACE1
6 Effect of Protonation State of the CatalyticAsp Dyad in Ligand Design
The design of potent drug like molecules for the inhibitionof BACE1 is an intensive area of research However scien-tific efforts to develop BACE1 inhibitors with appropriatepharmacokinetic and pharmacodynamics properties havenot been successful [52 53] In search of novel inhibitors anumber of peptidic and nonpeptidic small molecules werecocrystallized with the ectodomain of BACE1 [52] Importantinformation such as ligand-protein interaction network andconformation of the ligand at the active site can be obtainedfrom cocrystal structures But a deeper understanding ofmicroenvironments such as the protonation states of theAspsor the ionizable groups is still beyond the scope of X-raycrystallography This information is absolutely necessary forthe design of novel specific inhibitors that can cross the bloodbrain barrier (BBB) Therefore current BACE1 inhibitordesign efforts intensively focused on the determination ofthe protonation states of these two Asp residues (Asp32 andAsp228) in the presence of a specific functional inhibitorscaffold [26 48 54ndash57]
To address the protonation states of the Asp dyad inthe presence of different scaffolds of inhibitors a moleculardocking study was conducted including the critical Aspdyad (Asp32 and Asp228) of BACE1 using eight chemicallydiverse inhibitors [58] In total eight structurally differentscaffolds (HE AE TC CA RA AK ABP and AP (Table 1))
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 Scientifica
studies and animalmodeling this enzyme has been proposedto be a very promising target for the treatment of AD[9ndash14]
Structurally the ectodomain of BACE1 is composed ofseveral subregions that control the entry and orientation ofthe substrate at the active site The active site of BACE1contains two conserved Asp residues [15] that forms thecatalytic dyad This dyad has been implicated in the catalyticfunctioning of the entire family of aspartyl proteases includ-ing pepsin renin cathepsin D and HIV protease [16ndash23]The catalytic Asp dyad at the active site is covered over byan antiparallel hairpin-loop known as flap The mechanismsof flap closing and catalysis are of great significance due totheir involvement in human diseases such as AD During thecatalytic cycle the flap must open to allow the entrance ofthe substrate into the active site cleft and steer it towards thecatalytic Asp dyad to attain a reactive conformation In thisconformation the specific peptide bond(s) of the substrateis hydrolytically cleaved This type of gating mechanism hasbeen reported to be utilized by most of the members of theaspartyl protease family The dyad utilizes a general acid-base mechanism for the catalysis of peptide hydrolysis [17]Although theoretical X-ray and neutron diffraction datashow that the Asp residues in the catalytic dyad can switchprotonation states during the catalytic turnover [24ndash26] theeffect of the protonation states of these residues still remainsan intriguing issue in the development of successful drugdesigning strategies
Although experimental techniques can provide a greatdeal of information about the catalytic mechanism andprotonation states of the ionizable residues the atomic leveldescription of these complex chemical transformations is stillbeyond their reach These limitations can be overcome bymodern computational chemistry methods that can describethese complex processes at the atomistic level Computationalchemistry is a well-established field which has become anindispensable tool to study complex chemical and biochem-ical systems In the last decade applications of the densityfunctional theory (DFT) to study the chemical reactionsusing small systems have dominated the field Howeverthe main caveat of using DFT is the restricted number ofatoms (sim200) Therefore to study the larger system DFT(QM) has been coupled to the molecular mechanics (MM)potentials and implemented in hybrid QMMM(ONIOM)[27ndash33] method to study the catalytic mechanisms of theenzymes Applications of QMMM in biological systemshave shown tremendous success [34] On the other handmolecular dynamics (MD) simulations have become anessential part of current research to study the phase spacebehavior conformational changes of molecules and to cal-culate thermodynamic properties of systems [35 36] Alongwith MD MM based scoring functions have also beenincorporated into the docking engines andhave gained exten-sive use in modern computer aided drug design protocol[37]
In this review the current knowledge of structural andfunctional aspects of BACE1 in atomistic detail using multi-dimensional computational methods has been discussed
2 Structural Characteristics of BACE1
To date more than 290 crystal structures of BACE1 (Apoform and cocrystal with inhibitors) have been deposited intothe protein data bank (PDB) However the first cocrystalstructure of BACE1 with a hydroxyethylene (HE) basedtransition state isostere (OM99-2 and OM00-3) revealed thefirst evidence of BACE1 active site that contained the catalyticdyad (Asp32 and Asp228) at the center of the active site[15 38]The globular nature of BACE1 can be divided into N-andC-terminal domainsTheflap of this enzyme is composedof eleven residues (Val67-Glu77) and is positioned at theN-terminal domain A conserved Tyr71 residue is locatedat the flap which is found to adopt different conformationsin the presence and absence of inhibitors (Figure 1) Thereare several key functional regions namely 10s loop (Lys9-Tyr14) third strand (Lys107-Gly117) and insert A (Gly158-Leu167) that are present at the N-terminus Whereas insert B(Lys218-Asn221) insert C (Ala251-Pro258) insert D (Trp270-Thr274) insert E (Glu290-Ser295) and insert F (Asp311-Asp317) regions are located at the C-terminus these regionsfacilitate the entry and binding of different substrates atthe active site through their movements [15 39] At theactive site BACE1 contains two conserved water molecules(WAT1 and WAT2) After carefully analyzing 82 cocrystalstructures of aspartyl proteases the specific role of thesetwo water molecules was suggested by Andreeva and Rumsh[40] The WAT1 water located near the catalytic Asp dyadwas assigned to be most important as it is utilized in thehydrolytic cleavage of the peptide bond The second watermolecule (WAT2) participates in a continuous H-bondingnetwork and stabilizes the flap in the closed conformationthrough structural organization
3 Differential Substrate Specificity of BACE1
The induced fit model for enzyme-substrate interactions[41] suggests that the interaction of the substrate with anenzyme modulates the spatial orientation of the enzymersquosactive site residues into an optimal and precise conformationto accommodate the substrate Conformational preference ofthe enzyme-substrate complex is driven by the dynamics ofboth the enzyme and substrate It is not possible to resolve X-ray structure of a wild type- (WT-) enzyme in active formbound to substrate In the absence of this structure theatomistic level understanding of enzyme-substrate interac-tions remains elusive We have studied the role of proteindynamics in the formation of enzyme-substrate complex andsubstrate specificity of BACE1 involved in the generation ofA120573-peptides [23]
BACE1 is known to cleave the SW-substrate (Glu-Val-Asn-Leu-Asp-Ala-Glu-Phe) sixty times more efficiently thanthe WT-substrate (Glu-Val-Lys-Met-Asp-Ala-Glu-Phe) Weexplored the interaction profiles of BACE1 with the WT- andSW-substrates through 20 ns all-atom MD simulations in aTIP4P explicit water system using the OPLS-AA [42] forcefield Four different structures (WT-BACE1 SW-BACE1 anda 4-phenoxypyrrolidine-based BACE-1 inhibitor (compound11 PDB ID 2qmg [43]) bound BACE1) [44] along with the
Scientifica 3
Open flap
Closed flap
Tyr71
WAT2WAT1
Asp228 Asp32
C-terminal domain
Insert C
Insert E
Insert BInsert D
Insert F
Insert A
10s loop
Flap
N-terminal domain
Figure 1 Critical regions of BACE1 that participate in the substrate recognition catalytic Asp dyad conserved waters (WAT1 and WAT2)and orientation of Tyr71 in open and closed conformation of the flap
apo form of BACE1 were studied In order to capture thedynamic behavior of substrate recognition by the enzyme thestarting models were generated by placing the substrates andthe inhibitor inside the apo form of BACE1 (PDB ID1w50)[15 45] The representative structure derived from the com-pound 11-BACE1 simulations shows an accurate reproductionof the crystallographic orientation of the flap the 10s loopand BACE1- inhibitor interactions The root-mean-square(rms) deviation between the simulated and X-ray structureswas only 13 AThe simulation therefore served as a validationof the MD method and the parameters that were used forthe substrate bound BACE1 In the presence of the inhibitorand the substrate the flap closing event (Figure 2) occurredaround 3 ns and two water molecules facilitated the processAccording to a site-directed mutagenesis study electrostaticinteractions between the Glu residue of the substrate andArg307 residue of BACE1 play important roles in enhancingthe catalytic efficiency of the enzyme [46] In the BACE1-WTsimulation this interaction was lost due to the formation ofintramolecular salt-bridge between the Glu residue and theLys residue of the substrate (Figure 2) Therefore the lossof the Glu-Arg307 interaction was considered as one of thecauses for the diminished activity of the enzyme with theWT-substrate A comparison between three critical topologi-cal distances (C120572(Thr72)-C120573(Asp32) C120572(Thr72)-C120572(Thr329)and OG1(Thr72)-NH1(Arg235)) and the volume of the active
site showed that in comparison to theWT-substrate the flapwas more closed and the active site was more constrictedupon the binding of the SW-substrate In addition the num-ber of hydrogen bonds formed by the SW-substratewas foundto be two times (8ndash10) more than the bonds contributedby the WT-substrate The computed electrostatic bindingenergy of the SW-substrate was found to be 19 kcalmolmorefavorable than the WT-substrate In addition to substrate-enzyme interactions structures of inserts A D and F and the10s loop of the enzyme were also substantially different uponthe binding of these two substrates Overall these structuraldifferences between the WT- and SW-substrate explicitlyindicate that BACE1 demonstrated greater affinity for the SW-substrate and positioned it in a more bioactive conformationcompared to the WT-substrate
4 Differential Site Specificity of BACE1
In addition to the cleavage of Met-Asp amide bond of WT-APP at +1 site or 120573 site BACE1 can cleave the peptide bondbetween Tyr-Glu at +11 or 1205731015840 site and produce +11 A120573 speciesThis site specificity can be altered through themodification ofC-terminal domain of BACE1 Glycosylphosphatidylinositol(GPI) bound form of BACE1 (BACE1-GPI chimera) wasshown to have more 120573 site cleavage specificity compared to
4 Scientifica
2
18
16
14
12
1
0 5 10 15 20
Time (ns)
Dist
ance
(nm
)
(a)
0 5 10 15 20
Time (ns)
16
14
12
10
8
6
4
2
0
Num
ber o
f hyd
roge
n bo
nds
(b)
Lys321
Arg307
WT
Glu(P
Glu(P4)
4)
Asn233
Asn233Lys(P2)
Lys321
Val(P3)Val(P3)
SW
Arg307
Asn(P2)
(c)
Figure 2 (a) Time evolution of the topological distance between the C120572(Thr72) of flap tip and C120573(Asp32) of catalytic dyad (violet BACE1-compound 11 blue BACE1-SW substrate red BACE1-WT substrate and green Apo BACE1) (b) Number of hydrogen bonds formed betweenthe substrates and BACE1 (blue BACE1-SW substrate red BACE1-WT substrate) (c) Interaction of the substrate Glu with the Arg307 ofBACE1
the1205731015840 site OurMD simulation studies onBACE1 andBACE1-GPI showed that GPI could introduce significant changes inthe structure of BACE1The structural changes are associatedwith the flap third strand and insert F of the enzyme In thepresence of GPI the flapmoved to amore open conformationand strong hydrogen bond interactions between the thirdstrand and insert F brought them closer to each otherOn the other hand the flap remains in a relatively closedconformation and the two segments moved apart due to theloss of the hydrogen bond interactions in the WT-BACE1Overall these results suggest that GPI induces structuralmodifications thatmay regulate the site recognition of BACE1[47]
5 Catalytic Mechanism of BACE1
BACE1 cleaves specific peptide bonds of the substrates uti-lizing the catalytic Asp dyad This Asp dyad interacts withthe conserved water molecule (WAT1) present in the activesite and utilizes it for peptide hydrolysis Recent X-ray andneutron diffraction data and theoretical calculations on theaspartyl proteases show that during the catalytic cycle oneof the Asp residues is protonated and the second one isunprotonated [24ndash26 48] BACE1 catalyzes the hydrolytic
cleavage of the Met-Asp and Leu-Asp peptide bonds of theWT- and SW-substrate respectively
In the catalytic cycle BACE1 utilizes a two-step generalacidbasemechanism to cleave these peptide bonds (Figure 3)[17] In the first step from the reactant (I) the unprotonatedAsp228 functions as a base and abstracts a proton from thecatalytic water (WAT1) to generate a hydroxyl ion (OHminus)which subsequently makes a nucleophilic attack on thecarbonyl carbon of the peptide bond In this process Asp32acts as an acid and concomitantly donates its proton to thecarbonyl oxygen atom of the scissile peptide bond to generatethe gem-diol intermediate (II) In II two hydroxyl groups arecoordinated to the carbonyl carbon atomof the peptide bondIn the next step the two aspartate residues exchange theirfunctional roles Asp32 now functions as a base and abstractsa proton from the hydroxyl group (ndashOH) of II Here Asp228which initially acted as a base and became protonated in theprevious step plays the role of an acid by donating its protonto the amide nitrogen atom (ndashNH) of the scissile peptidebondThe cleavage process of the peptide bond occurs in thisprocess and generates separated amine (ndashNH
2) and carboxyl
(ndashCOOH) terminals (III)The hydrolysis of both WT- and SW-substrates was
investigated using two computational methods In the first
Scientifica 5
O
OH
H
O O
H
HN
O
HOOC HOOC
O
HOOC
O O
O H
H
O
H
HN
O
NH H
O H
O O
HO
I II III
Asp32 Asp32 Asp32
Asp228 Asp228 Asp228
NH2 NH2NH2
R1 R1
R2 R2R2
Ominus
Ominus
Ominus
C C C
R1
Figure 3 General acid-base mechanism utilized for peptide hydrolysis
ONIOMPruned model
196
219
00
50
100
150
200
250
00
Ener
gy (k
calm
ol)
Reaction coordinate
147
224
159171
minus32
minus57I TSI II TSII III
WT-substrate
(a)
ONIOMPruned model
161 172
00
50
100
150
200
250
00
Reaction coordinate
100
191
82
98
minus16
minus52I TSI II TSII III
SW-substrate
Ener
gy (k
calm
ol)
(b)
Figure 4 Computed barriers for the WT- and SW-substrate using pruned models (with the DFT method) and entire protein (with theONIOM (QMMM) method)
step of this study the most representative structures derivedfrom the BACE1-WT and BACE1-SW MD simulations wereutilized to develop pruned models of the enzyme-substratecomplexes In the second step these structures were used toinvestigate hydrolytic cleavage mechanisms through densityfunctional theory (DFT) [23] In the second study a moreaccurate hybrid QMMM (ONIOM) method was applied tostudy these mechanisms by including the whole protein inmodels [49]
According to the DFT study the formation of the gem-diol intermediate in the first step is the rate-determining step
of the entire mechanism for both substrates The reactionproceeds through a barrier of 224 kcalmol and 191 kcalmol(TSI) for the WT- and SW-substrate respectively Thegem-diol intermediate (II) for the SW-substrate was foundto be 80 kcalmol lower in energy than the one for theWT-substrate The stabilization of this intermediate is alsoreflected during the cleavage of the peptide bond throughtransition state TSII The cleavage of the peptide bond pro-ceeds through a barrier of 171 and 98 kcalmol for the WT-and SW-substrates respectivelyThe potential energy surface(PES) diagrams for these reactions are shown in Figure 4
6 Scientifica
Table 1 Ligands with chemical structure and PDB ID of cocrystal structure of BACE1
Ligands Chemical structures PDB ID
L1 (HE)
O
O
OS
O O
OOH
NH
NH H
NH
N
N2P4J
L2 (AE)O O O
F
NH2
N NH
NH
2FDP
L3 (TC)
O
OO
O
O
OH
N
NH
NH2
F
S
2IS0
L4 (CA)O
O
OHO
HN
HN
F
F
N
2QMF
L5 (RA)
O O O
O
O
NH
NN
H
HN
N
H
S
F
2QZL
L6 (AK)
O
O
OO
NNH HO
OH
S
N
NH 2WF4
Scientifica 7
Table 1 Continued
Ligands Chemical structures PDB ID
L7 (ABP)
O O
OO
Cl
OO
N
N
HS
NH2
2ZJM
L8 (AP)
O
N
HN
H2N
3HVG
The structural differences in the microenvironment of theactive site play a critical role in lowering the barrier for theSW-substrate Although the prunedmodels accurately repro-duced the experimentally observed barrier ca 180 kcalmolfor both substrates [50] contribution of the steric and elec-trostatic effects of the protein surrounding on the computedenergetics of the reaction was completely ignored Thereforeto incorporate the effect of the protein environment on theenergetics the hybrid QMMM (ONIOMB3LYPAmber)method was applied to investigate the hydrolytic cleavageof the WT- and SW-substrate The PES diagrams for thesereactions are shown in Figure 4 In this case the formationof the gem-diol intermediate occurs with barriers of 196 and161 kcalmol for the WT- and SW-substrates respectivelyThe inclusion of the electrostatic and steric effects of thesurrounding protein in the model reduced the barrier of thisstep by sim30 kcalmol for both substrates These reductionsare observed due to the alterations in the reaction coordinatesand microenvironment of the active site and are not due tothe long range structural reorganization in the enzyme Thecreation of the gem-diol intermediate in this step for the SW-substrate (100 kcalmol) is 47 kcalmol more favorable thanfor theWT-substrate (147 kcalmol)The cleavage of the pep-tide bond of the WT-substrate occurs through a calculatedbarrier of 219 kcalmol For the SW-substrate the barrier forthis process is reduced by 47 kcalmol that is 172 kcalmolfrom the reactant Results from theDFT andQMMMstudiesindicate that the formation of the gem-diol intermediateis the rate determining step of the entire mechanism forboth substrates The recent crystallographic [51] and Car-Parrinello MD simulations [20] studies on another memberof the aspartyl protease family HIV protease also suggestedthe cleavage of the peptide bond to be the rate-limiting stepIn our QMMM calculations the rate-limiting step for theSW-substrate is hard to distinguish due to a small difference(11 kcalmol) in barriers between the two steps The overall
reduction in the barrier for each step in the SW-substrate isalso in linewith the experimental observation [50] suggestingthat BACE1 can cleave the SW-substrate more efficiently thantheWT-substrateThese studies explicitly provided an atomiclevel understanding of the complex mechanism of peptidebond cleavage by BACE1
6 Effect of Protonation State of the CatalyticAsp Dyad in Ligand Design
The design of potent drug like molecules for the inhibitionof BACE1 is an intensive area of research However scien-tific efforts to develop BACE1 inhibitors with appropriatepharmacokinetic and pharmacodynamics properties havenot been successful [52 53] In search of novel inhibitors anumber of peptidic and nonpeptidic small molecules werecocrystallized with the ectodomain of BACE1 [52] Importantinformation such as ligand-protein interaction network andconformation of the ligand at the active site can be obtainedfrom cocrystal structures But a deeper understanding ofmicroenvironments such as the protonation states of theAspsor the ionizable groups is still beyond the scope of X-raycrystallography This information is absolutely necessary forthe design of novel specific inhibitors that can cross the bloodbrain barrier (BBB) Therefore current BACE1 inhibitordesign efforts intensively focused on the determination ofthe protonation states of these two Asp residues (Asp32 andAsp228) in the presence of a specific functional inhibitorscaffold [26 48 54ndash57]
To address the protonation states of the Asp dyad inthe presence of different scaffolds of inhibitors a moleculardocking study was conducted including the critical Aspdyad (Asp32 and Asp228) of BACE1 using eight chemicallydiverse inhibitors [58] In total eight structurally differentscaffolds (HE AE TC CA RA AK ABP and AP (Table 1))
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Scientifica 3
Open flap
Closed flap
Tyr71
WAT2WAT1
Asp228 Asp32
C-terminal domain
Insert C
Insert E
Insert BInsert D
Insert F
Insert A
10s loop
Flap
N-terminal domain
Figure 1 Critical regions of BACE1 that participate in the substrate recognition catalytic Asp dyad conserved waters (WAT1 and WAT2)and orientation of Tyr71 in open and closed conformation of the flap
apo form of BACE1 were studied In order to capture thedynamic behavior of substrate recognition by the enzyme thestarting models were generated by placing the substrates andthe inhibitor inside the apo form of BACE1 (PDB ID1w50)[15 45] The representative structure derived from the com-pound 11-BACE1 simulations shows an accurate reproductionof the crystallographic orientation of the flap the 10s loopand BACE1- inhibitor interactions The root-mean-square(rms) deviation between the simulated and X-ray structureswas only 13 AThe simulation therefore served as a validationof the MD method and the parameters that were used forthe substrate bound BACE1 In the presence of the inhibitorand the substrate the flap closing event (Figure 2) occurredaround 3 ns and two water molecules facilitated the processAccording to a site-directed mutagenesis study electrostaticinteractions between the Glu residue of the substrate andArg307 residue of BACE1 play important roles in enhancingthe catalytic efficiency of the enzyme [46] In the BACE1-WTsimulation this interaction was lost due to the formation ofintramolecular salt-bridge between the Glu residue and theLys residue of the substrate (Figure 2) Therefore the lossof the Glu-Arg307 interaction was considered as one of thecauses for the diminished activity of the enzyme with theWT-substrate A comparison between three critical topologi-cal distances (C120572(Thr72)-C120573(Asp32) C120572(Thr72)-C120572(Thr329)and OG1(Thr72)-NH1(Arg235)) and the volume of the active
site showed that in comparison to theWT-substrate the flapwas more closed and the active site was more constrictedupon the binding of the SW-substrate In addition the num-ber of hydrogen bonds formed by the SW-substratewas foundto be two times (8ndash10) more than the bonds contributedby the WT-substrate The computed electrostatic bindingenergy of the SW-substrate was found to be 19 kcalmolmorefavorable than the WT-substrate In addition to substrate-enzyme interactions structures of inserts A D and F and the10s loop of the enzyme were also substantially different uponthe binding of these two substrates Overall these structuraldifferences between the WT- and SW-substrate explicitlyindicate that BACE1 demonstrated greater affinity for the SW-substrate and positioned it in a more bioactive conformationcompared to the WT-substrate
4 Differential Site Specificity of BACE1
In addition to the cleavage of Met-Asp amide bond of WT-APP at +1 site or 120573 site BACE1 can cleave the peptide bondbetween Tyr-Glu at +11 or 1205731015840 site and produce +11 A120573 speciesThis site specificity can be altered through themodification ofC-terminal domain of BACE1 Glycosylphosphatidylinositol(GPI) bound form of BACE1 (BACE1-GPI chimera) wasshown to have more 120573 site cleavage specificity compared to
4 Scientifica
2
18
16
14
12
1
0 5 10 15 20
Time (ns)
Dist
ance
(nm
)
(a)
0 5 10 15 20
Time (ns)
16
14
12
10
8
6
4
2
0
Num
ber o
f hyd
roge
n bo
nds
(b)
Lys321
Arg307
WT
Glu(P
Glu(P4)
4)
Asn233
Asn233Lys(P2)
Lys321
Val(P3)Val(P3)
SW
Arg307
Asn(P2)
(c)
Figure 2 (a) Time evolution of the topological distance between the C120572(Thr72) of flap tip and C120573(Asp32) of catalytic dyad (violet BACE1-compound 11 blue BACE1-SW substrate red BACE1-WT substrate and green Apo BACE1) (b) Number of hydrogen bonds formed betweenthe substrates and BACE1 (blue BACE1-SW substrate red BACE1-WT substrate) (c) Interaction of the substrate Glu with the Arg307 ofBACE1
the1205731015840 site OurMD simulation studies onBACE1 andBACE1-GPI showed that GPI could introduce significant changes inthe structure of BACE1The structural changes are associatedwith the flap third strand and insert F of the enzyme In thepresence of GPI the flapmoved to amore open conformationand strong hydrogen bond interactions between the thirdstrand and insert F brought them closer to each otherOn the other hand the flap remains in a relatively closedconformation and the two segments moved apart due to theloss of the hydrogen bond interactions in the WT-BACE1Overall these results suggest that GPI induces structuralmodifications thatmay regulate the site recognition of BACE1[47]
5 Catalytic Mechanism of BACE1
BACE1 cleaves specific peptide bonds of the substrates uti-lizing the catalytic Asp dyad This Asp dyad interacts withthe conserved water molecule (WAT1) present in the activesite and utilizes it for peptide hydrolysis Recent X-ray andneutron diffraction data and theoretical calculations on theaspartyl proteases show that during the catalytic cycle oneof the Asp residues is protonated and the second one isunprotonated [24ndash26 48] BACE1 catalyzes the hydrolytic
cleavage of the Met-Asp and Leu-Asp peptide bonds of theWT- and SW-substrate respectively
In the catalytic cycle BACE1 utilizes a two-step generalacidbasemechanism to cleave these peptide bonds (Figure 3)[17] In the first step from the reactant (I) the unprotonatedAsp228 functions as a base and abstracts a proton from thecatalytic water (WAT1) to generate a hydroxyl ion (OHminus)which subsequently makes a nucleophilic attack on thecarbonyl carbon of the peptide bond In this process Asp32acts as an acid and concomitantly donates its proton to thecarbonyl oxygen atom of the scissile peptide bond to generatethe gem-diol intermediate (II) In II two hydroxyl groups arecoordinated to the carbonyl carbon atomof the peptide bondIn the next step the two aspartate residues exchange theirfunctional roles Asp32 now functions as a base and abstractsa proton from the hydroxyl group (ndashOH) of II Here Asp228which initially acted as a base and became protonated in theprevious step plays the role of an acid by donating its protonto the amide nitrogen atom (ndashNH) of the scissile peptidebondThe cleavage process of the peptide bond occurs in thisprocess and generates separated amine (ndashNH
2) and carboxyl
(ndashCOOH) terminals (III)The hydrolysis of both WT- and SW-substrates was
investigated using two computational methods In the first
Scientifica 5
O
OH
H
O O
H
HN
O
HOOC HOOC
O
HOOC
O O
O H
H
O
H
HN
O
NH H
O H
O O
HO
I II III
Asp32 Asp32 Asp32
Asp228 Asp228 Asp228
NH2 NH2NH2
R1 R1
R2 R2R2
Ominus
Ominus
Ominus
C C C
R1
Figure 3 General acid-base mechanism utilized for peptide hydrolysis
ONIOMPruned model
196
219
00
50
100
150
200
250
00
Ener
gy (k
calm
ol)
Reaction coordinate
147
224
159171
minus32
minus57I TSI II TSII III
WT-substrate
(a)
ONIOMPruned model
161 172
00
50
100
150
200
250
00
Reaction coordinate
100
191
82
98
minus16
minus52I TSI II TSII III
SW-substrate
Ener
gy (k
calm
ol)
(b)
Figure 4 Computed barriers for the WT- and SW-substrate using pruned models (with the DFT method) and entire protein (with theONIOM (QMMM) method)
step of this study the most representative structures derivedfrom the BACE1-WT and BACE1-SW MD simulations wereutilized to develop pruned models of the enzyme-substratecomplexes In the second step these structures were used toinvestigate hydrolytic cleavage mechanisms through densityfunctional theory (DFT) [23] In the second study a moreaccurate hybrid QMMM (ONIOM) method was applied tostudy these mechanisms by including the whole protein inmodels [49]
According to the DFT study the formation of the gem-diol intermediate in the first step is the rate-determining step
of the entire mechanism for both substrates The reactionproceeds through a barrier of 224 kcalmol and 191 kcalmol(TSI) for the WT- and SW-substrate respectively Thegem-diol intermediate (II) for the SW-substrate was foundto be 80 kcalmol lower in energy than the one for theWT-substrate The stabilization of this intermediate is alsoreflected during the cleavage of the peptide bond throughtransition state TSII The cleavage of the peptide bond pro-ceeds through a barrier of 171 and 98 kcalmol for the WT-and SW-substrates respectivelyThe potential energy surface(PES) diagrams for these reactions are shown in Figure 4
6 Scientifica
Table 1 Ligands with chemical structure and PDB ID of cocrystal structure of BACE1
Ligands Chemical structures PDB ID
L1 (HE)
O
O
OS
O O
OOH
NH
NH H
NH
N
N2P4J
L2 (AE)O O O
F
NH2
N NH
NH
2FDP
L3 (TC)
O
OO
O
O
OH
N
NH
NH2
F
S
2IS0
L4 (CA)O
O
OHO
HN
HN
F
F
N
2QMF
L5 (RA)
O O O
O
O
NH
NN
H
HN
N
H
S
F
2QZL
L6 (AK)
O
O
OO
NNH HO
OH
S
N
NH 2WF4
Scientifica 7
Table 1 Continued
Ligands Chemical structures PDB ID
L7 (ABP)
O O
OO
Cl
OO
N
N
HS
NH2
2ZJM
L8 (AP)
O
N
HN
H2N
3HVG
The structural differences in the microenvironment of theactive site play a critical role in lowering the barrier for theSW-substrate Although the prunedmodels accurately repro-duced the experimentally observed barrier ca 180 kcalmolfor both substrates [50] contribution of the steric and elec-trostatic effects of the protein surrounding on the computedenergetics of the reaction was completely ignored Thereforeto incorporate the effect of the protein environment on theenergetics the hybrid QMMM (ONIOMB3LYPAmber)method was applied to investigate the hydrolytic cleavageof the WT- and SW-substrate The PES diagrams for thesereactions are shown in Figure 4 In this case the formationof the gem-diol intermediate occurs with barriers of 196 and161 kcalmol for the WT- and SW-substrates respectivelyThe inclusion of the electrostatic and steric effects of thesurrounding protein in the model reduced the barrier of thisstep by sim30 kcalmol for both substrates These reductionsare observed due to the alterations in the reaction coordinatesand microenvironment of the active site and are not due tothe long range structural reorganization in the enzyme Thecreation of the gem-diol intermediate in this step for the SW-substrate (100 kcalmol) is 47 kcalmol more favorable thanfor theWT-substrate (147 kcalmol)The cleavage of the pep-tide bond of the WT-substrate occurs through a calculatedbarrier of 219 kcalmol For the SW-substrate the barrier forthis process is reduced by 47 kcalmol that is 172 kcalmolfrom the reactant Results from theDFT andQMMMstudiesindicate that the formation of the gem-diol intermediateis the rate determining step of the entire mechanism forboth substrates The recent crystallographic [51] and Car-Parrinello MD simulations [20] studies on another memberof the aspartyl protease family HIV protease also suggestedthe cleavage of the peptide bond to be the rate-limiting stepIn our QMMM calculations the rate-limiting step for theSW-substrate is hard to distinguish due to a small difference(11 kcalmol) in barriers between the two steps The overall
reduction in the barrier for each step in the SW-substrate isalso in linewith the experimental observation [50] suggestingthat BACE1 can cleave the SW-substrate more efficiently thantheWT-substrateThese studies explicitly provided an atomiclevel understanding of the complex mechanism of peptidebond cleavage by BACE1
6 Effect of Protonation State of the CatalyticAsp Dyad in Ligand Design
The design of potent drug like molecules for the inhibitionof BACE1 is an intensive area of research However scien-tific efforts to develop BACE1 inhibitors with appropriatepharmacokinetic and pharmacodynamics properties havenot been successful [52 53] In search of novel inhibitors anumber of peptidic and nonpeptidic small molecules werecocrystallized with the ectodomain of BACE1 [52] Importantinformation such as ligand-protein interaction network andconformation of the ligand at the active site can be obtainedfrom cocrystal structures But a deeper understanding ofmicroenvironments such as the protonation states of theAspsor the ionizable groups is still beyond the scope of X-raycrystallography This information is absolutely necessary forthe design of novel specific inhibitors that can cross the bloodbrain barrier (BBB) Therefore current BACE1 inhibitordesign efforts intensively focused on the determination ofthe protonation states of these two Asp residues (Asp32 andAsp228) in the presence of a specific functional inhibitorscaffold [26 48 54ndash57]
To address the protonation states of the Asp dyad inthe presence of different scaffolds of inhibitors a moleculardocking study was conducted including the critical Aspdyad (Asp32 and Asp228) of BACE1 using eight chemicallydiverse inhibitors [58] In total eight structurally differentscaffolds (HE AE TC CA RA AK ABP and AP (Table 1))
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 Scientifica
2
18
16
14
12
1
0 5 10 15 20
Time (ns)
Dist
ance
(nm
)
(a)
0 5 10 15 20
Time (ns)
16
14
12
10
8
6
4
2
0
Num
ber o
f hyd
roge
n bo
nds
(b)
Lys321
Arg307
WT
Glu(P
Glu(P4)
4)
Asn233
Asn233Lys(P2)
Lys321
Val(P3)Val(P3)
SW
Arg307
Asn(P2)
(c)
Figure 2 (a) Time evolution of the topological distance between the C120572(Thr72) of flap tip and C120573(Asp32) of catalytic dyad (violet BACE1-compound 11 blue BACE1-SW substrate red BACE1-WT substrate and green Apo BACE1) (b) Number of hydrogen bonds formed betweenthe substrates and BACE1 (blue BACE1-SW substrate red BACE1-WT substrate) (c) Interaction of the substrate Glu with the Arg307 ofBACE1
the1205731015840 site OurMD simulation studies onBACE1 andBACE1-GPI showed that GPI could introduce significant changes inthe structure of BACE1The structural changes are associatedwith the flap third strand and insert F of the enzyme In thepresence of GPI the flapmoved to amore open conformationand strong hydrogen bond interactions between the thirdstrand and insert F brought them closer to each otherOn the other hand the flap remains in a relatively closedconformation and the two segments moved apart due to theloss of the hydrogen bond interactions in the WT-BACE1Overall these results suggest that GPI induces structuralmodifications thatmay regulate the site recognition of BACE1[47]
5 Catalytic Mechanism of BACE1
BACE1 cleaves specific peptide bonds of the substrates uti-lizing the catalytic Asp dyad This Asp dyad interacts withthe conserved water molecule (WAT1) present in the activesite and utilizes it for peptide hydrolysis Recent X-ray andneutron diffraction data and theoretical calculations on theaspartyl proteases show that during the catalytic cycle oneof the Asp residues is protonated and the second one isunprotonated [24ndash26 48] BACE1 catalyzes the hydrolytic
cleavage of the Met-Asp and Leu-Asp peptide bonds of theWT- and SW-substrate respectively
In the catalytic cycle BACE1 utilizes a two-step generalacidbasemechanism to cleave these peptide bonds (Figure 3)[17] In the first step from the reactant (I) the unprotonatedAsp228 functions as a base and abstracts a proton from thecatalytic water (WAT1) to generate a hydroxyl ion (OHminus)which subsequently makes a nucleophilic attack on thecarbonyl carbon of the peptide bond In this process Asp32acts as an acid and concomitantly donates its proton to thecarbonyl oxygen atom of the scissile peptide bond to generatethe gem-diol intermediate (II) In II two hydroxyl groups arecoordinated to the carbonyl carbon atomof the peptide bondIn the next step the two aspartate residues exchange theirfunctional roles Asp32 now functions as a base and abstractsa proton from the hydroxyl group (ndashOH) of II Here Asp228which initially acted as a base and became protonated in theprevious step plays the role of an acid by donating its protonto the amide nitrogen atom (ndashNH) of the scissile peptidebondThe cleavage process of the peptide bond occurs in thisprocess and generates separated amine (ndashNH
2) and carboxyl
(ndashCOOH) terminals (III)The hydrolysis of both WT- and SW-substrates was
investigated using two computational methods In the first
Scientifica 5
O
OH
H
O O
H
HN
O
HOOC HOOC
O
HOOC
O O
O H
H
O
H
HN
O
NH H
O H
O O
HO
I II III
Asp32 Asp32 Asp32
Asp228 Asp228 Asp228
NH2 NH2NH2
R1 R1
R2 R2R2
Ominus
Ominus
Ominus
C C C
R1
Figure 3 General acid-base mechanism utilized for peptide hydrolysis
ONIOMPruned model
196
219
00
50
100
150
200
250
00
Ener
gy (k
calm
ol)
Reaction coordinate
147
224
159171
minus32
minus57I TSI II TSII III
WT-substrate
(a)
ONIOMPruned model
161 172
00
50
100
150
200
250
00
Reaction coordinate
100
191
82
98
minus16
minus52I TSI II TSII III
SW-substrate
Ener
gy (k
calm
ol)
(b)
Figure 4 Computed barriers for the WT- and SW-substrate using pruned models (with the DFT method) and entire protein (with theONIOM (QMMM) method)
step of this study the most representative structures derivedfrom the BACE1-WT and BACE1-SW MD simulations wereutilized to develop pruned models of the enzyme-substratecomplexes In the second step these structures were used toinvestigate hydrolytic cleavage mechanisms through densityfunctional theory (DFT) [23] In the second study a moreaccurate hybrid QMMM (ONIOM) method was applied tostudy these mechanisms by including the whole protein inmodels [49]
According to the DFT study the formation of the gem-diol intermediate in the first step is the rate-determining step
of the entire mechanism for both substrates The reactionproceeds through a barrier of 224 kcalmol and 191 kcalmol(TSI) for the WT- and SW-substrate respectively Thegem-diol intermediate (II) for the SW-substrate was foundto be 80 kcalmol lower in energy than the one for theWT-substrate The stabilization of this intermediate is alsoreflected during the cleavage of the peptide bond throughtransition state TSII The cleavage of the peptide bond pro-ceeds through a barrier of 171 and 98 kcalmol for the WT-and SW-substrates respectivelyThe potential energy surface(PES) diagrams for these reactions are shown in Figure 4
6 Scientifica
Table 1 Ligands with chemical structure and PDB ID of cocrystal structure of BACE1
Ligands Chemical structures PDB ID
L1 (HE)
O
O
OS
O O
OOH
NH
NH H
NH
N
N2P4J
L2 (AE)O O O
F
NH2
N NH
NH
2FDP
L3 (TC)
O
OO
O
O
OH
N
NH
NH2
F
S
2IS0
L4 (CA)O
O
OHO
HN
HN
F
F
N
2QMF
L5 (RA)
O O O
O
O
NH
NN
H
HN
N
H
S
F
2QZL
L6 (AK)
O
O
OO
NNH HO
OH
S
N
NH 2WF4
Scientifica 7
Table 1 Continued
Ligands Chemical structures PDB ID
L7 (ABP)
O O
OO
Cl
OO
N
N
HS
NH2
2ZJM
L8 (AP)
O
N
HN
H2N
3HVG
The structural differences in the microenvironment of theactive site play a critical role in lowering the barrier for theSW-substrate Although the prunedmodels accurately repro-duced the experimentally observed barrier ca 180 kcalmolfor both substrates [50] contribution of the steric and elec-trostatic effects of the protein surrounding on the computedenergetics of the reaction was completely ignored Thereforeto incorporate the effect of the protein environment on theenergetics the hybrid QMMM (ONIOMB3LYPAmber)method was applied to investigate the hydrolytic cleavageof the WT- and SW-substrate The PES diagrams for thesereactions are shown in Figure 4 In this case the formationof the gem-diol intermediate occurs with barriers of 196 and161 kcalmol for the WT- and SW-substrates respectivelyThe inclusion of the electrostatic and steric effects of thesurrounding protein in the model reduced the barrier of thisstep by sim30 kcalmol for both substrates These reductionsare observed due to the alterations in the reaction coordinatesand microenvironment of the active site and are not due tothe long range structural reorganization in the enzyme Thecreation of the gem-diol intermediate in this step for the SW-substrate (100 kcalmol) is 47 kcalmol more favorable thanfor theWT-substrate (147 kcalmol)The cleavage of the pep-tide bond of the WT-substrate occurs through a calculatedbarrier of 219 kcalmol For the SW-substrate the barrier forthis process is reduced by 47 kcalmol that is 172 kcalmolfrom the reactant Results from theDFT andQMMMstudiesindicate that the formation of the gem-diol intermediateis the rate determining step of the entire mechanism forboth substrates The recent crystallographic [51] and Car-Parrinello MD simulations [20] studies on another memberof the aspartyl protease family HIV protease also suggestedthe cleavage of the peptide bond to be the rate-limiting stepIn our QMMM calculations the rate-limiting step for theSW-substrate is hard to distinguish due to a small difference(11 kcalmol) in barriers between the two steps The overall
reduction in the barrier for each step in the SW-substrate isalso in linewith the experimental observation [50] suggestingthat BACE1 can cleave the SW-substrate more efficiently thantheWT-substrateThese studies explicitly provided an atomiclevel understanding of the complex mechanism of peptidebond cleavage by BACE1
6 Effect of Protonation State of the CatalyticAsp Dyad in Ligand Design
The design of potent drug like molecules for the inhibitionof BACE1 is an intensive area of research However scien-tific efforts to develop BACE1 inhibitors with appropriatepharmacokinetic and pharmacodynamics properties havenot been successful [52 53] In search of novel inhibitors anumber of peptidic and nonpeptidic small molecules werecocrystallized with the ectodomain of BACE1 [52] Importantinformation such as ligand-protein interaction network andconformation of the ligand at the active site can be obtainedfrom cocrystal structures But a deeper understanding ofmicroenvironments such as the protonation states of theAspsor the ionizable groups is still beyond the scope of X-raycrystallography This information is absolutely necessary forthe design of novel specific inhibitors that can cross the bloodbrain barrier (BBB) Therefore current BACE1 inhibitordesign efforts intensively focused on the determination ofthe protonation states of these two Asp residues (Asp32 andAsp228) in the presence of a specific functional inhibitorscaffold [26 48 54ndash57]
To address the protonation states of the Asp dyad inthe presence of different scaffolds of inhibitors a moleculardocking study was conducted including the critical Aspdyad (Asp32 and Asp228) of BACE1 using eight chemicallydiverse inhibitors [58] In total eight structurally differentscaffolds (HE AE TC CA RA AK ABP and AP (Table 1))
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Scientifica 5
O
OH
H
O O
H
HN
O
HOOC HOOC
O
HOOC
O O
O H
H
O
H
HN
O
NH H
O H
O O
HO
I II III
Asp32 Asp32 Asp32
Asp228 Asp228 Asp228
NH2 NH2NH2
R1 R1
R2 R2R2
Ominus
Ominus
Ominus
C C C
R1
Figure 3 General acid-base mechanism utilized for peptide hydrolysis
ONIOMPruned model
196
219
00
50
100
150
200
250
00
Ener
gy (k
calm
ol)
Reaction coordinate
147
224
159171
minus32
minus57I TSI II TSII III
WT-substrate
(a)
ONIOMPruned model
161 172
00
50
100
150
200
250
00
Reaction coordinate
100
191
82
98
minus16
minus52I TSI II TSII III
SW-substrate
Ener
gy (k
calm
ol)
(b)
Figure 4 Computed barriers for the WT- and SW-substrate using pruned models (with the DFT method) and entire protein (with theONIOM (QMMM) method)
step of this study the most representative structures derivedfrom the BACE1-WT and BACE1-SW MD simulations wereutilized to develop pruned models of the enzyme-substratecomplexes In the second step these structures were used toinvestigate hydrolytic cleavage mechanisms through densityfunctional theory (DFT) [23] In the second study a moreaccurate hybrid QMMM (ONIOM) method was applied tostudy these mechanisms by including the whole protein inmodels [49]
According to the DFT study the formation of the gem-diol intermediate in the first step is the rate-determining step
of the entire mechanism for both substrates The reactionproceeds through a barrier of 224 kcalmol and 191 kcalmol(TSI) for the WT- and SW-substrate respectively Thegem-diol intermediate (II) for the SW-substrate was foundto be 80 kcalmol lower in energy than the one for theWT-substrate The stabilization of this intermediate is alsoreflected during the cleavage of the peptide bond throughtransition state TSII The cleavage of the peptide bond pro-ceeds through a barrier of 171 and 98 kcalmol for the WT-and SW-substrates respectivelyThe potential energy surface(PES) diagrams for these reactions are shown in Figure 4
6 Scientifica
Table 1 Ligands with chemical structure and PDB ID of cocrystal structure of BACE1
Ligands Chemical structures PDB ID
L1 (HE)
O
O
OS
O O
OOH
NH
NH H
NH
N
N2P4J
L2 (AE)O O O
F
NH2
N NH
NH
2FDP
L3 (TC)
O
OO
O
O
OH
N
NH
NH2
F
S
2IS0
L4 (CA)O
O
OHO
HN
HN
F
F
N
2QMF
L5 (RA)
O O O
O
O
NH
NN
H
HN
N
H
S
F
2QZL
L6 (AK)
O
O
OO
NNH HO
OH
S
N
NH 2WF4
Scientifica 7
Table 1 Continued
Ligands Chemical structures PDB ID
L7 (ABP)
O O
OO
Cl
OO
N
N
HS
NH2
2ZJM
L8 (AP)
O
N
HN
H2N
3HVG
The structural differences in the microenvironment of theactive site play a critical role in lowering the barrier for theSW-substrate Although the prunedmodels accurately repro-duced the experimentally observed barrier ca 180 kcalmolfor both substrates [50] contribution of the steric and elec-trostatic effects of the protein surrounding on the computedenergetics of the reaction was completely ignored Thereforeto incorporate the effect of the protein environment on theenergetics the hybrid QMMM (ONIOMB3LYPAmber)method was applied to investigate the hydrolytic cleavageof the WT- and SW-substrate The PES diagrams for thesereactions are shown in Figure 4 In this case the formationof the gem-diol intermediate occurs with barriers of 196 and161 kcalmol for the WT- and SW-substrates respectivelyThe inclusion of the electrostatic and steric effects of thesurrounding protein in the model reduced the barrier of thisstep by sim30 kcalmol for both substrates These reductionsare observed due to the alterations in the reaction coordinatesand microenvironment of the active site and are not due tothe long range structural reorganization in the enzyme Thecreation of the gem-diol intermediate in this step for the SW-substrate (100 kcalmol) is 47 kcalmol more favorable thanfor theWT-substrate (147 kcalmol)The cleavage of the pep-tide bond of the WT-substrate occurs through a calculatedbarrier of 219 kcalmol For the SW-substrate the barrier forthis process is reduced by 47 kcalmol that is 172 kcalmolfrom the reactant Results from theDFT andQMMMstudiesindicate that the formation of the gem-diol intermediateis the rate determining step of the entire mechanism forboth substrates The recent crystallographic [51] and Car-Parrinello MD simulations [20] studies on another memberof the aspartyl protease family HIV protease also suggestedthe cleavage of the peptide bond to be the rate-limiting stepIn our QMMM calculations the rate-limiting step for theSW-substrate is hard to distinguish due to a small difference(11 kcalmol) in barriers between the two steps The overall
reduction in the barrier for each step in the SW-substrate isalso in linewith the experimental observation [50] suggestingthat BACE1 can cleave the SW-substrate more efficiently thantheWT-substrateThese studies explicitly provided an atomiclevel understanding of the complex mechanism of peptidebond cleavage by BACE1
6 Effect of Protonation State of the CatalyticAsp Dyad in Ligand Design
The design of potent drug like molecules for the inhibitionof BACE1 is an intensive area of research However scien-tific efforts to develop BACE1 inhibitors with appropriatepharmacokinetic and pharmacodynamics properties havenot been successful [52 53] In search of novel inhibitors anumber of peptidic and nonpeptidic small molecules werecocrystallized with the ectodomain of BACE1 [52] Importantinformation such as ligand-protein interaction network andconformation of the ligand at the active site can be obtainedfrom cocrystal structures But a deeper understanding ofmicroenvironments such as the protonation states of theAspsor the ionizable groups is still beyond the scope of X-raycrystallography This information is absolutely necessary forthe design of novel specific inhibitors that can cross the bloodbrain barrier (BBB) Therefore current BACE1 inhibitordesign efforts intensively focused on the determination ofthe protonation states of these two Asp residues (Asp32 andAsp228) in the presence of a specific functional inhibitorscaffold [26 48 54ndash57]
To address the protonation states of the Asp dyad inthe presence of different scaffolds of inhibitors a moleculardocking study was conducted including the critical Aspdyad (Asp32 and Asp228) of BACE1 using eight chemicallydiverse inhibitors [58] In total eight structurally differentscaffolds (HE AE TC CA RA AK ABP and AP (Table 1))
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 Scientifica
Table 1 Ligands with chemical structure and PDB ID of cocrystal structure of BACE1
Ligands Chemical structures PDB ID
L1 (HE)
O
O
OS
O O
OOH
NH
NH H
NH
N
N2P4J
L2 (AE)O O O
F
NH2
N NH
NH
2FDP
L3 (TC)
O
OO
O
O
OH
N
NH
NH2
F
S
2IS0
L4 (CA)O
O
OHO
HN
HN
F
F
N
2QMF
L5 (RA)
O O O
O
O
NH
NN
H
HN
N
H
S
F
2QZL
L6 (AK)
O
O
OO
NNH HO
OH
S
N
NH 2WF4
Scientifica 7
Table 1 Continued
Ligands Chemical structures PDB ID
L7 (ABP)
O O
OO
Cl
OO
N
N
HS
NH2
2ZJM
L8 (AP)
O
N
HN
H2N
3HVG
The structural differences in the microenvironment of theactive site play a critical role in lowering the barrier for theSW-substrate Although the prunedmodels accurately repro-duced the experimentally observed barrier ca 180 kcalmolfor both substrates [50] contribution of the steric and elec-trostatic effects of the protein surrounding on the computedenergetics of the reaction was completely ignored Thereforeto incorporate the effect of the protein environment on theenergetics the hybrid QMMM (ONIOMB3LYPAmber)method was applied to investigate the hydrolytic cleavageof the WT- and SW-substrate The PES diagrams for thesereactions are shown in Figure 4 In this case the formationof the gem-diol intermediate occurs with barriers of 196 and161 kcalmol for the WT- and SW-substrates respectivelyThe inclusion of the electrostatic and steric effects of thesurrounding protein in the model reduced the barrier of thisstep by sim30 kcalmol for both substrates These reductionsare observed due to the alterations in the reaction coordinatesand microenvironment of the active site and are not due tothe long range structural reorganization in the enzyme Thecreation of the gem-diol intermediate in this step for the SW-substrate (100 kcalmol) is 47 kcalmol more favorable thanfor theWT-substrate (147 kcalmol)The cleavage of the pep-tide bond of the WT-substrate occurs through a calculatedbarrier of 219 kcalmol For the SW-substrate the barrier forthis process is reduced by 47 kcalmol that is 172 kcalmolfrom the reactant Results from theDFT andQMMMstudiesindicate that the formation of the gem-diol intermediateis the rate determining step of the entire mechanism forboth substrates The recent crystallographic [51] and Car-Parrinello MD simulations [20] studies on another memberof the aspartyl protease family HIV protease also suggestedthe cleavage of the peptide bond to be the rate-limiting stepIn our QMMM calculations the rate-limiting step for theSW-substrate is hard to distinguish due to a small difference(11 kcalmol) in barriers between the two steps The overall
reduction in the barrier for each step in the SW-substrate isalso in linewith the experimental observation [50] suggestingthat BACE1 can cleave the SW-substrate more efficiently thantheWT-substrateThese studies explicitly provided an atomiclevel understanding of the complex mechanism of peptidebond cleavage by BACE1
6 Effect of Protonation State of the CatalyticAsp Dyad in Ligand Design
The design of potent drug like molecules for the inhibitionof BACE1 is an intensive area of research However scien-tific efforts to develop BACE1 inhibitors with appropriatepharmacokinetic and pharmacodynamics properties havenot been successful [52 53] In search of novel inhibitors anumber of peptidic and nonpeptidic small molecules werecocrystallized with the ectodomain of BACE1 [52] Importantinformation such as ligand-protein interaction network andconformation of the ligand at the active site can be obtainedfrom cocrystal structures But a deeper understanding ofmicroenvironments such as the protonation states of theAspsor the ionizable groups is still beyond the scope of X-raycrystallography This information is absolutely necessary forthe design of novel specific inhibitors that can cross the bloodbrain barrier (BBB) Therefore current BACE1 inhibitordesign efforts intensively focused on the determination ofthe protonation states of these two Asp residues (Asp32 andAsp228) in the presence of a specific functional inhibitorscaffold [26 48 54ndash57]
To address the protonation states of the Asp dyad inthe presence of different scaffolds of inhibitors a moleculardocking study was conducted including the critical Aspdyad (Asp32 and Asp228) of BACE1 using eight chemicallydiverse inhibitors [58] In total eight structurally differentscaffolds (HE AE TC CA RA AK ABP and AP (Table 1))
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Scientifica 7
Table 1 Continued
Ligands Chemical structures PDB ID
L7 (ABP)
O O
OO
Cl
OO
N
N
HS
NH2
2ZJM
L8 (AP)
O
N
HN
H2N
3HVG
The structural differences in the microenvironment of theactive site play a critical role in lowering the barrier for theSW-substrate Although the prunedmodels accurately repro-duced the experimentally observed barrier ca 180 kcalmolfor both substrates [50] contribution of the steric and elec-trostatic effects of the protein surrounding on the computedenergetics of the reaction was completely ignored Thereforeto incorporate the effect of the protein environment on theenergetics the hybrid QMMM (ONIOMB3LYPAmber)method was applied to investigate the hydrolytic cleavageof the WT- and SW-substrate The PES diagrams for thesereactions are shown in Figure 4 In this case the formationof the gem-diol intermediate occurs with barriers of 196 and161 kcalmol for the WT- and SW-substrates respectivelyThe inclusion of the electrostatic and steric effects of thesurrounding protein in the model reduced the barrier of thisstep by sim30 kcalmol for both substrates These reductionsare observed due to the alterations in the reaction coordinatesand microenvironment of the active site and are not due tothe long range structural reorganization in the enzyme Thecreation of the gem-diol intermediate in this step for the SW-substrate (100 kcalmol) is 47 kcalmol more favorable thanfor theWT-substrate (147 kcalmol)The cleavage of the pep-tide bond of the WT-substrate occurs through a calculatedbarrier of 219 kcalmol For the SW-substrate the barrier forthis process is reduced by 47 kcalmol that is 172 kcalmolfrom the reactant Results from theDFT andQMMMstudiesindicate that the formation of the gem-diol intermediateis the rate determining step of the entire mechanism forboth substrates The recent crystallographic [51] and Car-Parrinello MD simulations [20] studies on another memberof the aspartyl protease family HIV protease also suggestedthe cleavage of the peptide bond to be the rate-limiting stepIn our QMMM calculations the rate-limiting step for theSW-substrate is hard to distinguish due to a small difference(11 kcalmol) in barriers between the two steps The overall
reduction in the barrier for each step in the SW-substrate isalso in linewith the experimental observation [50] suggestingthat BACE1 can cleave the SW-substrate more efficiently thantheWT-substrateThese studies explicitly provided an atomiclevel understanding of the complex mechanism of peptidebond cleavage by BACE1
6 Effect of Protonation State of the CatalyticAsp Dyad in Ligand Design
The design of potent drug like molecules for the inhibitionof BACE1 is an intensive area of research However scien-tific efforts to develop BACE1 inhibitors with appropriatepharmacokinetic and pharmacodynamics properties havenot been successful [52 53] In search of novel inhibitors anumber of peptidic and nonpeptidic small molecules werecocrystallized with the ectodomain of BACE1 [52] Importantinformation such as ligand-protein interaction network andconformation of the ligand at the active site can be obtainedfrom cocrystal structures But a deeper understanding ofmicroenvironments such as the protonation states of theAspsor the ionizable groups is still beyond the scope of X-raycrystallography This information is absolutely necessary forthe design of novel specific inhibitors that can cross the bloodbrain barrier (BBB) Therefore current BACE1 inhibitordesign efforts intensively focused on the determination ofthe protonation states of these two Asp residues (Asp32 andAsp228) in the presence of a specific functional inhibitorscaffold [26 48 54ndash57]
To address the protonation states of the Asp dyad inthe presence of different scaffolds of inhibitors a moleculardocking study was conducted including the critical Aspdyad (Asp32 and Asp228) of BACE1 using eight chemicallydiverse inhibitors [58] In total eight structurally differentscaffolds (HE AE TC CA RA AK ABP and AP (Table 1))
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
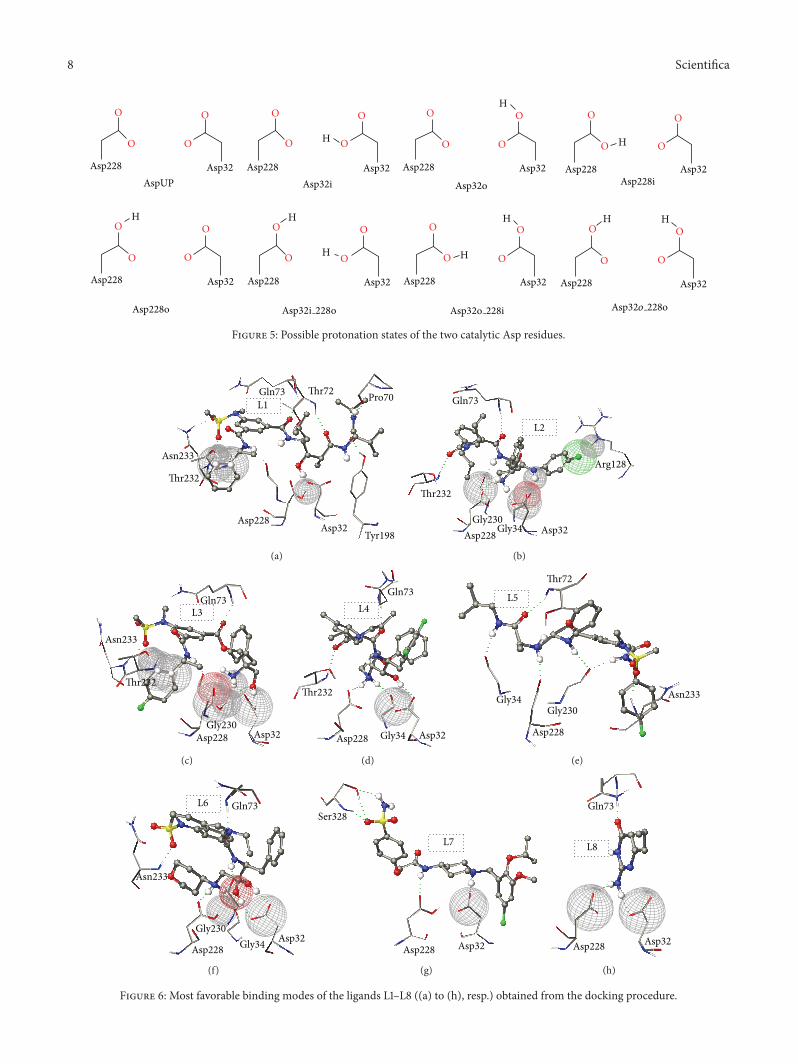
8 Scientifica
O
O O
O O
O O
O
H
O
O O
OH
O
O H O
O
O
O
H
O
O O
O
H
O
O
H
O
O H O
OH
O
O
H
O
OH
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
Asp228 Asp32 Asp228 Asp32 Asp228 Asp32 Asp228 Asp32
AspUP Asp32i Asp32o Asp228i
Asp228o Asp32i 228o Asp32o 228i Asp32o 228o
Figure 5 Possible protonation states of the two catalytic Asp residues
L1Gln73 Thr72 Pro70
Asn233
Thr232
Asp228Asp32
Tyr198
(a)
Asp228 Asp32
Arg128
Gln73
L2
Thr232
Gly34Gly230
(b)
L3Gln73
Asn233
Thr232
Gly230Asp228 Asp32
(c)
Gln73
Thr232
Asp228 Asp32
L4
Gly34
(d)
Thr72
Gly34Gly230
Asn233
Asp228
L5
(e)
Asn233
Asp228 Gly34 Asp32
L6 Gln73
Gly230
(f)
Asp32Asp228
L7
Ser328
(g)
Gln73
L8
Asp228 Asp32
(h)
Figure 6 Most favorable binding modes of the ligands L1ndashL8 ((a) to (h) resp) obtained from the docking procedure
Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Scientifica 9
[43 56 59ndash64] containing inhibitors were self-docked intothe corresponding X-ray structures using the two differentdocking engines AutoDock 40 [65 66] and Glide XP [6768] In these procedures eight possible protonation statesof the catalytic Asps (AspUP Asp32i Asp32o Asp228iAsp228o Asp32i 228o Asp32o 228i and Asp32o 228o) wereconsidered (Figure 5) The results obtained from these dock-ing simulations were analyzed using the following threeparameters (i) rms deviation of the docked ligand pose fromthe crystallographic orientation of the ligand that shouldbe below 2 A (ii) ligand docking score the more negativethe score is the more favorable the interaction with BACE1is and (iii) minimum deviation of the critical atoms thatinteract with the Asp dyad of the docked pose from the X-ray structure
The possible protonation states were considered only ifthese three criterions were satisfied The self-docking studyclearly showed that the binding mode of these inhibitorsdepends on the protonation states of the two Asp residuesThemonoprotonated Asp228i state was found to be preferredby HE (L1) TC (L3) AK (L6) and ABP (L7) based inhibitorswhereas RA (L5) prefers the monoprotonated Asp32o stateThe AE (L2) CA (L4) and AP (L8) inhibitors favored thedideprotonated AspUP state The most favorable dockingposes are shown in Figure 6 Although some discrepancieswere observed in predicting the protonation state of theL3 ligand the predicted docked scores showed a goodcorrelation (R2 value of gt 075) between the calculated bind-ing affinities and experimental data Moreover the resultsobtained from this study also showed that besides favoringa single protonation state multiple protonation states couldalso be favorable for a single ligand It is interesting to notethat five out of eight ligands (L1 L2 L3 L4 and L5) showedhigh preference for a diprotonated state The possibilities ofmultiple protonation states also agree with a previous studythat suggested that the protonation states of the Asp dyadcan alter under different pH conditions in the presence ofthe diverse inhibitors [69] The results obtained from thisstudy clearly indicate that considering a single protonationstate of these two critical Asp residues is not sufficientand the most favored states for definite chemical scaffoldmust be determined prior to conducting a virtual screeningcampaign
7 Summary and Future Perspective
This review summarizes the application of computationalmethodologies to elucidate the dynamics substrate speci-ficity and catalytic mechanism of one of the most criticalenzymes (BACE1) involved in the pathogenesis of AD Inaddition the effects of protonation states of the catalytic Aspresidues have been discussed in developing novel inhibitorsagainst this enzyme Although the results obtained fromthese studies support experimental observations the accu-racy of the structural information concerning the enzyme-substrate interactions requires additional advancement In
this aspect the application of more sophisticated computa-tional approaches such as meta- accelerated dissipative par-ticle and polarizable dynamics simulations will be very use-ful In addition to substrate-protein interaction the studies ofcatalytic mechanisms can be further improved by employingmore sampling methods and incorporating the hydrophobicinteractions Therefore coupling of QMMM method withMD simulations and incorporation of the dispersion termin the DFT methods will provide more accurate energeticsFinally for the prediction of the protonation state of thecatalytic Asp more accurate free energy based calculationssuch as application of the orthogonal space random walk(OSRW) method or constant pH simulation methods will bevery useful
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
Financial support from the James and Esther King Biomedi-cal Research Program of the Florida State HealthDepartment(DOH Grant no 08KN-11) to Rajeev Prabhakar is greatlyacknowledged Computational resources from the Center forComputational Science (CCS) at the University of Miami aregreatly appreciated The authors thank Thomas Paul and AliSawani for their assistance in the preparation of the paper
References
[1] Z Suo J Humphrey A Kundtz et al ldquoSoluble Alzheimers120573-amyloid constricts the cerebral vasculature in vivordquo Neuro-science Letters vol 257 no 2 pp 77ndash80 1998
[2] T Pillot B Drouet S Queille et al ldquoThe nonfibrillar amyloid120573-peptide induces apoptotic neuronal cell death involvementof its C-terminal fusogenic domainrdquo Journal of Neurochemistryvol 73 no 4 pp 1626ndash1634 1999
[3] D B Northrop ldquoFollow the protons a low-barrier hydro-gen bond unifies the mechanisms of the aspartic proteasesrdquoAccounts of Chemical Research vol 34 no 10 pp 790ndash797 2001
[4] D R Davies ldquoThe structure and function of the asparticproteinasesrdquo Annual Review of Biophysics and BiophysicalChemistry vol 19 pp 189ndash215 1990
[5] M Miller M Jaskolski J K Mohana Rao J Leis and AWlodawer ldquoCrystal structure of a retroviral protease provesrelationship to aspartic protease familyrdquo Nature vol 337 no6207 pp 576ndash579 1989
[6] M A Navia P M D Fitzgerald B M McKeever et alldquoThree-dimensional structure of aspartyl protease from humanimmunodeficiency virus HIV-1rdquo Nature vol 337 no 6208 pp615ndash620 1989
[7] F Gruninger-Leitch D Schlatter E Kung P Nelbock and HDobeli ldquoSubstrate and inhibitor profile of BACE (120573-secretase)and comparison with other mammalian aspartic proteasesrdquoJournal of Biological Chemistry vol 277 no 7 pp 4687ndash46932002
10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 Scientifica
[8] M L Hemming J E Elias S P Gygi and D J Selkoe ldquoIdenti-fication of beta-secretase (BACE1) substrates using quantitativeproteomicsrdquo PLoS ONE vol 4 no 12 p e8477 2009
[9] R Yan M J Blenkowski M E Shuck et al ldquoMembrane-anchored aspartyl proteasewithAlzheimerrsquos disease120573-secretaseactivityrdquo Nature vol 402 no 6761 pp 533ndash537 1999
[10] R Vassar B D Bennett S Babu-Khan et al ldquo120573-Secretasecleavage of Alzheimerrsquos amyloid precursor protein by thetransmembrane aspartic protease BACErdquo Science vol 286 no5440 pp 735ndash741 1999
[11] S Sinha J P Anderson R Barbour et al ldquoPurification andcloning of amyloid precursor protein 120573-secretase from humanbrainrdquo Nature vol 402 no 6761 pp 537ndash540 1999
[12] H Cai Y Wang D McCarthy et al ldquoBACE1 is the major 120573-secretase for generation of A120573 peptides by neuronsrdquo NatureNeuroscience vol 4 no 3 pp 233ndash234 2001
[13] Y Luo B Bolon S Kahn et al ldquoMice deficient in BACE1 theAlzheimerrsquos 120573-secretase have normal phenotype and abolished120573-amyloid generationrdquo Nature Neuroscience vol 4 no 3 pp231ndash232 2001
[14] J Hardy ldquoAmyloid the presenilins and Alzheimerrsquos diseaserdquoTrends in Neurosciences vol 20 no 4 pp 154ndash159 1997
[15] L Hong G Koelsch X Lin et al ldquoStructure of the proteasedomain ofmemapsin 2 (120573-secretase) complexedwith inhibitorrdquoScience vol 290 no 5489 pp 150ndash153 2000
[16] D C Chatfield K P Eurenius and B R Brooks ldquoHIV-1protease cleavage mechanism a theoretical investigation basedon classicalMD simulation and reaction path calculations usinga hybrid QMMM potentialrdquo Journal of Molecular StructureTHEOCHEM vol 423 no 1-2 pp 79ndash92 1998
[17] K Suguna E A Padlan C W Smith W D Carlson andD R Davies ldquoBinding of a reduced peptide inhibitor to theaspartic proteinase from Rhizopus chinensis implications fora mechanism of actionrdquo Proceedings of the National Academyof Sciences of the United States of America vol 84 no 20 pp7009ndash7013 1987
[18] M Cascella C Micheletti U Rothlisberger and P CarlonildquoEvolutionary conserved functional mechanics across pepsin-like and retroviral aspartic proteasesrdquo Journal of the AmericanChemical Society vol 127 no 11 pp 3734ndash3742 2005
[19] S Bjelic and J Aqvist ldquoCatalysis and linear free energy rela-tionships in aspartic proteasesrdquo Biochemistry vol 45 no 25 pp7709ndash7723 2006
[20] S Piana D Bucher P Carloni and U Rothlisberger ldquoReactionmechanism of HIV-1 protease by hybrid car-parrinelloclassicalMD simulationsrdquo Journal of Physical Chemistry B vol 108 no30 pp 11139ndash11149 2004
[21] J Trylska P Grochowski and J A McCammon ldquoThe role ofhydrogen bonding in the enzymatic reaction catalyzed by HIV-1 proteaserdquo Protein Science vol 13 no 2 pp 513ndash528 2004
[22] R Singh A Barman and R Prabhakar ldquoComputationalinsights into aspartyl protease activity of presenilin 1 (ps1)generating alzheimer amyloid 120573-peptides (a12057340 and a12057342)rdquoJournal of Physical Chemistry B vol 113 no 10 pp 2990ndash29992009
[23] A Barman S Schurer and R Prabhakar ldquoComputationalmodeling of substrate specificity and catalysis of the120573-secretase(BACE1) enzymerdquo Biochemistry vol 50 no 20 pp 4337ndash43492011
[24] L Coates P T Erskine S Mall et al ldquoX-ray neutron and NMRstudies of the catalytic mechanism of aspartic proteinasesrdquoEuropean Biophysics Journal vol 35 no 7 pp 559ndash566 2006
[25] L Coates H-F Tuan S Tomanicek et al ldquoThe catalyticmechanism of an aspartic proteinase exploredwith neutron andX-ray diffractionrdquo Journal of theAmericanChemical Society vol130 no 23 pp 7235ndash7237 2008
[26] R Rajamani and C H Reynolds ldquoModeling the protonationstates of the catalytic aspartates in 120573-secretaserdquo Journal ofMedicinal Chemistry vol 47 no 21 pp 5159ndash5166 2004
[27] T Matsubara S Sieber and K Morokuma ldquoA test of the newldquointegrated MO + MMrdquo (IMOMM) method for the conforma-tional energy of ethane and n-butanerdquo International Journal ofQuantum Chemistry vol 60 no 6 pp 1101ndash1109 1996
[28] F Maseras and K Morokuma ldquoIMOMM a new integrated abinitio + molecular mechanics geometry optimization schemeof equilibrium structures and transition statesrdquo Journal ofComputational Chemistry vol 16 no 9 pp 1170ndash1179 1995
[29] S Humbel S Sieber andKMorokuma ldquoThe IMOMOmethodintegration of different levels of molecular orbital approxima-tions for geometry optimization of large systems test for n-butane conformation and SN2 reaction RCl+Clminusrdquo Journal ofChemical Physics vol 105 no 5 pp 1959ndash1967 1996
[30] M Svensson S Humbel and K Morokuma ldquoEnergetics usingthe single point IMOMO (integrated molecular orbital+molec-ular orbital) calculations choices of computational levels andmodel systemrdquo Journal of Chemical Physics vol 105 no 9 pp3654ndash3661 1996
[31] M Svensson S Humbel R D J Froese T Matsubara SSieber and K Morokuma ldquoONIOM a multilayered integratedMO + MM method for geometry optimizations and singlepoint energy predictions A test for diels-alder reactions andPt(P(t-Bu)3)2+H2 oxidative additionrdquo The Journal of PhysicalChemistry vol 100 pp 19357ndash19363 1935
[32] T Vreven and K Morokuma ldquoOn the Application of theIMOMO (integrated molecular orbital + molecular orbital)Methodrdquo Journal of Computational Chemistry vol 21 no 16 pp1419ndash1432 2000
[33] S Dapprich I Komaromi K S Byun K Morokuma and M JFrisch ldquoA new ONIOM implementation in Gaussian98 Part IThe calculation of energies gradients vibrational frequenciesand electric field derivativesrdquo Journal of Molecular StructureTHEOCHEM vol 461-462 pp 1ndash21 1999
[34] H M Senn and WThiel ldquoQMMMmethods for biomolecularsystemsrdquoAngewandte Chemie vol 48 no 7 pp 1198ndash1229 2009
[35] J D Durrant and J A McCammon ldquoMolecular dynamicssimulations and drug discoveryrdquo BMC Biology vol 9 article 712011
[36] D W Borhani and D E Shaw ldquoThe future of moleculardynamics simulations in drug discoveryrdquo Journal of Computer-Aided Molecular Design vol 26 no 1 pp 15ndash26 2012
[37] H Alonso A A Bliznyuk and J E Gready ldquoCombiningdocking and molecular dynamic simulations in drug designrdquoMedicinal Research Reviews vol 26 no 5 pp 531ndash568 2006
[38] L Hong R T Turner III G Koelsch D Shin A K Ghoshand J Tang ldquoCrystal structure of memapsin 2 (120573-secretase) incomplex with an inhibitor OM00-3rdquo Biochemistry vol 41 no36 pp 10963ndash10967 2002
[39] B Xiong X-Q Huang L-L Shen et al ldquoConformationalflexibility of 120573-secretase molecular dynamics simulation andessential dynamics analysisrdquo Acta Pharmacologica Sinica vol25 no 6 pp 705ndash713 2004
[40] N S Andreeva and L D Rumsh ldquoAnalysis of crystal structuresof aspartic proteinases on the role of amino acid residues
Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Scientifica 11
adjacent to the catalytic site of pepsin-like enzymesrdquo ProteinScience vol 10 no 12 pp 2439ndash2450 2001
[41] D E Koshland ldquoApplication of a theory of enzyme specificityto protein synthesisrdquo Proceedings of the National Academy ofSciences vol 44 pp 98ndash104 1958
[42] W L Jorgensen and J Tirado-Rives ldquoThe OPLS [optimizedpotentials for liquid simulations] potential functions for pro-teins energy minimizations for crystals of cyclic peptides andcrambinrdquo Journal of the American Chemical Society vol 110 no6 pp 1657ndash1666 1988
[43] U Iserloh J Pan A W Stamford et al ldquoDiscovery of an orallyefficaceous 4-phenoxypyrrolidine-based BACE-1 inhibitorrdquoBioorganic and Medicinal Chemistry Letters vol 18 no 1 pp418ndash422 2008
[44] U Iserloh YWu J N Cumming et al ldquoPotent pyrrolidine- andpiperidine-based BACE-1 inhibitorsrdquo Bioorganic and MedicinalChemistry Letters vol 18 no 1 pp 414ndash417 2008
[45] S Patel L Vuillard A Cleasby C W Murray and J YonldquoApo and inhibitor complex structures of BACE (120573-secretase)rdquoJournal of Molecular Biology vol 343 no 2 pp 407ndash416 2004
[46] R T Turner III G Koelsch L Hong P Castenheira A Ghoshand J Tang ldquoSubsite specificity of memapsin 2 (120573-secretase)implications for inhibitor designrdquo Biochemistry vol 40 no 34pp 10001ndash10006 2001
[47] K S Vetrivel A Barman Y Chen et al ldquoLoss of cleavage at 1205731015840-site contributes to apparent increase in 120573-amyloid peptide (A120573)secretion by 120573-secretase (BACE1)-glycosylphosphatidylinositol(GPI) processing of amyloid precursor proteinrdquo The Journal ofBiological Chemistry vol 286 no 29 pp 26166ndash26177 2011
[48] N Yu S A Hayik B Wang N Liao C H Reynolds and K MMerz Jr ldquoAssigning the protonation states of the key aspartatesin 120573-secretase using QMMM X-ray structure refinementrdquoJournal of Chemical Theory and Computation vol 2 no 4 pp1057ndash1069 2006
[49] A Barman and R Prabhakar ldquoElucidating the catalytic mecha-nism of 120573-secretase (BACE1) a quantum mechanicsmolecularmechanics (QMMM) approachrdquo Journal of Molecular Graphicsand Modelling vol 40 pp 1ndash9 2013
[50] X Lin G Koelsch S Wu D Downs A Dashti and J TangldquoHuman aspartic protease memapsin 2 cleaves the 120573-secretasesite of120573- amyloid precursor proteinrdquoProceedings of the NationalAcademy of Sciences of the United States of America vol 97 no4 pp 1456ndash1460 2000
[51] A Das S Mahale V Prashar S Bihani J-L Ferrer and M VHosur ldquoX-ray snapshot ofHIV-1 protease in action observationof tetrahedral intermediate and short ionic hydrogen bondSIHBwith catalytic aspartaterdquo Journal of the AmericanChemicalSociety vol 132 no 18 pp 6366ndash6373 2010
[52] R Silvestri ldquoBoom in the development of non-peptidic 120573-secretase (BACE1) inhibitors for the treatment of AlzheimersdiseaserdquoMedicinal Research Reviews vol 29 no 2 pp 295ndash3382009
[53] P CMay RADean S L Lowe et al ldquoRobust central reductionof amyloid-120573 in humans with an orally available non-peptidic120573-secretase inhibitorrdquo The Journal of Neuroscience vol 31 no46 pp 16507ndash16516 2011
[54] Y Cheng T C Judd M D Bartberger et al ldquoFrom fragmentscreening to in vivo efficacy optimization of a series of 2-aminoquinolines as potent inhibitors of beta-site amyloid pre-cursor protein cleaving enzyme 1 (bace1)rdquo Journal of MedicinalChemistry vol 54 no 16 pp 5836ndash5857 2011
[55] A K Ghosh S Gemma and J Tang ldquo120573-secretase as atherapeutic target for Alzheimerrsquos diseaserdquo Neurotherapeuticsvol 5 no 3 pp 399ndash408 2008
[56] A K Ghosh N Kumaragurubaran L Hong et al ldquoDesignsynthesis and X-ray structure of potent memapsin 2 (120573-secretase) inhibitors with isophthalamide derivatives as theP2minusP3-ligandsrdquo Journal of Medicinal Chemistry vol 50 no 10
pp 2399ndash2407 2007[57] H Park and S Lee ldquoDetermination of the active site protonation
state of 120573-secretase from molecular dynamics simulation anddocking experiment implications for structure-based inhibitordesignrdquo Journal of the American Chemical Society vol 125 no52 pp 16416ndash16422 2003
[58] A Barman andR Prabhakar ldquoProtonation states of the catalyticdyad of 120573-secretase (BACE1) in the presence of chemicallydiverse inhibitors a molecular docking studyrdquo Journal of Chem-ical Information andModeling vol 52 no 5 pp 1275ndash1287 2012
[59] W Yang W Lu Y Lu et al ldquoAminoethylenes a tetrahedralintermediate isostere yielding potent inhibitors of the aspartylprotease BACE-1rdquo Journal of Medicinal Chemistry vol 49 no 3pp 839ndash842 2006
[60] H A Rajapakse P G Nantermet H G Selnick et al ldquoDiscov-ery of oxadiazoyl tertiary carbinamine inhibitors of 120573-secretase(BACE-1)rdquo Journal of Medicinal Chemistry vol 49 no 25 pp7270ndash7273 2006
[61] C A Coburn S J Stachel K G Jones et al ldquoBACE-1 inhibitionby a series of 120595[CH2NH] reduced amide isosteresrdquo Bioorganicand Medicinal Chemistry Letters vol 16 no 14 pp 3635ndash36382006
[62] N Charrier B Clarke L Cutler et al ldquoSecond generationof BACE-1 inhibitors part 3 towards non hydroxyethylaminetransition state mimeticsrdquo Bioorganic and Medicinal ChemistryLetters vol 19 no 13 pp 3674ndash3678 2009
[63] W Yang R V Fucini B T Fahr et al ldquoFragment-baseddiscovery of nonpeptidic BACE-1 inhibitors using tetheringrdquoBiochemistry vol 48 no 21 pp 4488ndash4496 2009
[64] R Godemann J Madden J Kramer et al ldquoFragment-baseddiscovery of BACE1 inhibitors using functional assaysrdquo Bio-chemistry vol 48 no 45 pp 10743ndash10751 2009
[65] G M Morris D S Goodsell R S Halliday et al ldquoAutomateddocking using a Lamarckian genetic algorithm and an empiricalbinding free energy functionrdquo Journal of Computational Chem-istry vol 19 no 14 pp 1639ndash1662 1998
[66] F Osterberg G M Morris M F Sanner A J Olson andD S Goodsell ldquoAutomated docking to multiple target struc-tures incorporation of protein mobility and structural waterheterogeneity in autodockrdquo Proteins Structure Function andBioinformatics vol 46 no 1 pp 34ndash40 2002
[67] R A Friesner R B Murphy M P Repasky et al ldquoExtraprecision glide docking and scoring incorporating a model ofhydrophobic enclosure for protein-ligand complexesrdquo Journalof Medicinal Chemistry vol 49 no 21 pp 6177ndash6196 2006
[68] R A Friesner J L Banks R B Murphy et al ldquoGlide a newapproach for rapid accurate docking and scoring 1 Methodand assessment of docking accuracyrdquo Journal of MedicinalChemistry vol 47 no 7 pp 1739ndash1749 2004
[69] J L Domınguez T ChristopeitM C Villaverde et al ldquoEffect ofthe protonation state of the titratable residues on the inhibitoraffinity to BACE-1rdquo Biochemistry vol 49 no 34 pp 7255ndash72632010
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology