retracted: multiple developmental roles of ahnak are suggested by localization to sites of...
TRANSCRIPT

Multiple developmental roles of Ahnak are suggested by localization tosites of placentation and neural plate fusion in the mouse conceptus
Karen M. Downsa,*, Jacalyn McHugha, Andrew J. Coppb, Emma Shtivelmanc
aDepartment of Anatomy, University of Wisconsin–Madison Medical School, 1300 University Avenue, Madison, WI 53706 USAbNeural Development Unit, Institute of Child Health, University College London, 30 Guilford Street, London WC1N 1EH, UK
cCancer Research Institute, University of California San Francisco, Room S333, 2340 Sutter Street, San Francisco CA 94115, USA
Received 14 May 2002; received in revised form 21 August 2002; accepted 4 September 2002
Abstract
Ahnak is a gigantic (700 kD) phosphoprotein with a unique structure whose expression and cellular localization are dynamically regulated
during cell cycle progression. Here, we report that Ahnak is localized to sites of major morphogenesis during mouse placentation and
neurulation. Ahnak was found in: (i) derivatives of trophectoderm, including chorionic ectoderm prior to and during union with the
ectoplacental cone, presumptive syncytiotrophoblast cells in the chorionic labyrinth, and giant cells at the trophoblast-uterine interface;
(ii) the allantois prior to, during, and after union with the chorion; and (iii) the tips of the neural plate during formation of the neural tube. On
the basis of these observations, we suggest that Ahnak may play heretofore unrecognized roles in tissue union during normal mouse
development. q 2002 Elsevier Science B.V. All rights reserved.
Keywords: Ahnak; Allantois; Chorion; Ectoplacental cone; Extraembryonic mesoderm; Gastrulation; Giant cells; Invasion; Neural crest; Neural plate; Neural
tube; Neurulation; Node; Placentation; Surface ectoderm; Trophoblast stem cells; Yolk sac
1. Results and discussion
Ahnak was originally identified as an 18 kb-long messen-
ger RNA subject to transcriptional repression in neuroblas-
toma cell lines (Shtivelman and Bishop, 1992). Later,
Ahnak was recognized as desmoyokin (Hashimoto et al.,
1993), a protein enriched in the region of the desmosomal
plaques of stratified bovine epithelia (Hashimoto et al.,
1993; Hieda et al., 1989).
Changes in phosphorylation promote Ahnak’s shuttling
between the nucleus, cytoplasm and plasma membrane in
adult mammalian cell types (Nie et al., 2000; Shtivelman
and Bishop, 1993; Sussman et al., 2001). For example,
phosphorylation of Ahnak by protein kinase B leads to the
nuclear exclusion of Ahnak via a nuclear export signal
(Sussman et al., 2001) and localizes Ahnak to the inner
surface of the plasma membrane in cultured epithelial
cells that have formed mature cell–cell junctions. Similar
localization has been described for Ahnak in differentiated
keratinocytes in vivo (Masunaga et al., 1995) and in epithe-
lial cells in vitro (Nie et al., 2000). The function of Ahnak is
not known, but Ahnak’s large size and dynamic localization
patterns make it an attractive target for analysis of develop-
mental expression.
Recently, subtractive hybridization between extraem-
bryonic mesoderm and embryonic tissues suggested that
Ahnak might be preferentially expressed in extra-embryonic
mesoderm in the mouse gastrula (Kingsley et al., 2001).
Results of that study anecdotally revealed the presence of
Ahnak in allantoic mesothelium, which mediates chorio-
allantoic union (Downs, 2002; Downs and Gardner, 1995).
Because mesothelium may also promote differentiation of
the allantois (Downs et al., 1998, 2001), localization of
Ahnak to the allantois prior to, during and immediately
after union with the chorion served as a starting point for
this study. ‘KIS’ polyclonal antibodies (see Section 2)
recognized cytoplasmic and plasmalemmal, but not nuclear,
Ahnak in histological sections prepared from mouse
conceptuses between approximately 7.5 and 10.0 days post-
coitum (dpc).
1.1. Ahnak in extraembryonic tissues
The implanting mouse conceptus contains an outer layer
of trophectoderm, extraembryonic endoderm (both parietal
and visceral), and epiblast. Epiblast will form the fetus, as
well as the amnion and extraembryonic mesoderm of the
Gene Expression Patterns 2 (2002) 27–34
1567-133X/02/$ - see front matter q 2002 Elsevier Science B.V. All rights reserved.
PII: S0925-4773(02)00349-0
www.elsevier.com/locate/modgep
* Corresponding author. Tel.: 11-608-265-5411; fax: 11-608-262-7306.
E-mail address: [email protected] (K.M. Downs).
chorion, yolk sac and all of the allantois. Derivatives of
trophectoderm not only promote attachment of the concep-
tus to the uterus, but they contribute to most of the chorionic
disk of the placenta. Morphogenetic union between the
chorion and the allantois (Downs, 2002; Downs and Gard-
ner, 1995) and between the chorion and the ectoplacental
cone to form the mature chorionic disk are essential for
correct placental ontogeny in the mouse. Chorio-allantoic
union permits penetration of the fetal circulatory system
into the chorionic disk, whilst the mature chorionic disk
mediates essential exchange between the fetal and maternal
bloodstreams.
In accord with a previous report (Kingsley et al., 2001),
we found Ahnak in all mesodermal cells lining the exocoe-
lomic cavity at all stages of development analyzed, includ-
ing the amnion, the yolk sac and its blood islands, the
chorion, and the outer cells of the allantois, whether or not
the latter had epithelialized into presumptive mesothelium
K.M. Downs et al. / Gene Expression Patterns 2 (2002) 27–3428

(Fig. 1A–F). In contrast with Flk-1 and VCAM-1, which
localize with distal-to-proximal directionality to the allan-
tois during differentiation (Downs, 2002; Downs et al.,
1998; Downs and Harmann, 1997), Ahnak exhibited no
obvious polarity (Fig. 1A–C). During fusion with the chor-
ion, Ahnak persisted in the mesothelium of the allantois, and
was prevalent in its loosely organized core (Fig. 1C), with
especially high levels in basal allantoic mesoderm contin-
uous with the amnion (Fig. 1B). By 16–18-somite pairs,
Ahnak was obvious in the endothelium of the nascent allan-
toic vasculature (Fig. 1D). That Ahnak might play a role in
sites of fusion was supported by especially high levels of
Ahnak at the periphery of the chorio-allantoic fusion junc-
tion, where adhesion between allantoic and chorionic
mesothelial cells secures the umbilicus onto the chorionic
disk (Downs, 2002; Fig. 1E). Between 12- and 27-somite
pairs, presumptive syncytiotrophoblast cells found at sites
of association with Flk-1-positive allantoic endothelial cells
also appeared to stain positively for Ahnak (Fig. 1G–J, and
data not shown).
Expression of Ahnak in derivatives of trophectoderm was
observed as early as the neural plate stage (approximately
7.5 dpc). Prior to its union with the relatively negative ecto-
placental cone, chorionic ectoderm contained abundant
cytoplasmic Ahnak (Fig. 2A–C). Then, as medial chorionic
ectoderm began to fuse with the ectoplacental cone, Ahnak
became preferentially localized to the apical portion of the
chorionic cells (Fig. 2B, C). Thereafter, Ahnak was found in
the syncytiotrophoblasts of the labyrinth, as described above
(Fig. 1G-I).
Throughout all stages examined, Ahnak was also loca-
lized to the plasma membrane in many, but not all, tropho-
blast giant cells, whether or not the conceptuses had been
dissected from their implantation sites (Fig. 2E, H). In addi-
tion, high levels of Ahnak were found in the perinuclear
region and cytoplasm of the decidua capsularis which
contains large binucleate cells (Fig. 2F, I), and in the
decidua vascularis (Fig. 2F, J). Very little staining was
found in the decidua basalis (Fig. 2F, K).
1.2. Ahnak in the embryo
Ahnak’s pattern of localization was very dynamic in the
embryonic portion of the conceptus, being apparent first in
anterior mesoderm, embryonic visceral endoderm, and in
the node at the late neural plate stage (Fig. 3A). By the
headfold stage, Ahnak’s expression had spread posteriorly
in cranial mesoderm (Fig. 3B), and was found in the heart
anlage by 3-somite pairs (Fig. 3C, D, F). The primitive
streak, nascent embryonic mesoderm, and somites were
always Ahnak-negative (Fig 3C, E). Although Ahnak was
initially not detectable in the neuroectoderm (Fig. 3A–C),
by 5-somite pairs, however, it became visible at the poster-
ior surface of the trunk (Fig. 3E), in surface ectoderm and/or
neuroectoderm. By 7-somite pairs, some endothelial cells
within the head mesenchyme were positive, as was the
endoderm of the fore- (Fig. 3F) and hindgut (not shown),
and surface ectoderm overlying the neuroectoderm of the
developing hindbrain (e.g. Fig. 3F). Thus, Ahnak may be
involved in ontogeny of the heart, notochord, definitive
endoderm, surface ectoderm and/or neural tube.
By 10–11-somite pairs, the embryos under study were
undergoing axial rotation, which provided frontal views of
the prospective brain region. These revealed Ahnak in the
tips of the dorsal cranial folds prior to, during and after
closure in the putative mid-brain region between 10- and
18-somite pairs (e.g. Fig. 3G–J, and data not shown).
To determine more accurately the whereabouts of Ahnak
during formation of the central nervous system, subsequent
efforts were focused on a systematic analysis of the neural
tube during primary and secondary neurulation at the level
of the posterior neuropore (Shum and Copp, 1996). Primary
K.M. Downs et al. / Gene Expression Patterns 2 (2002) 27–34 29
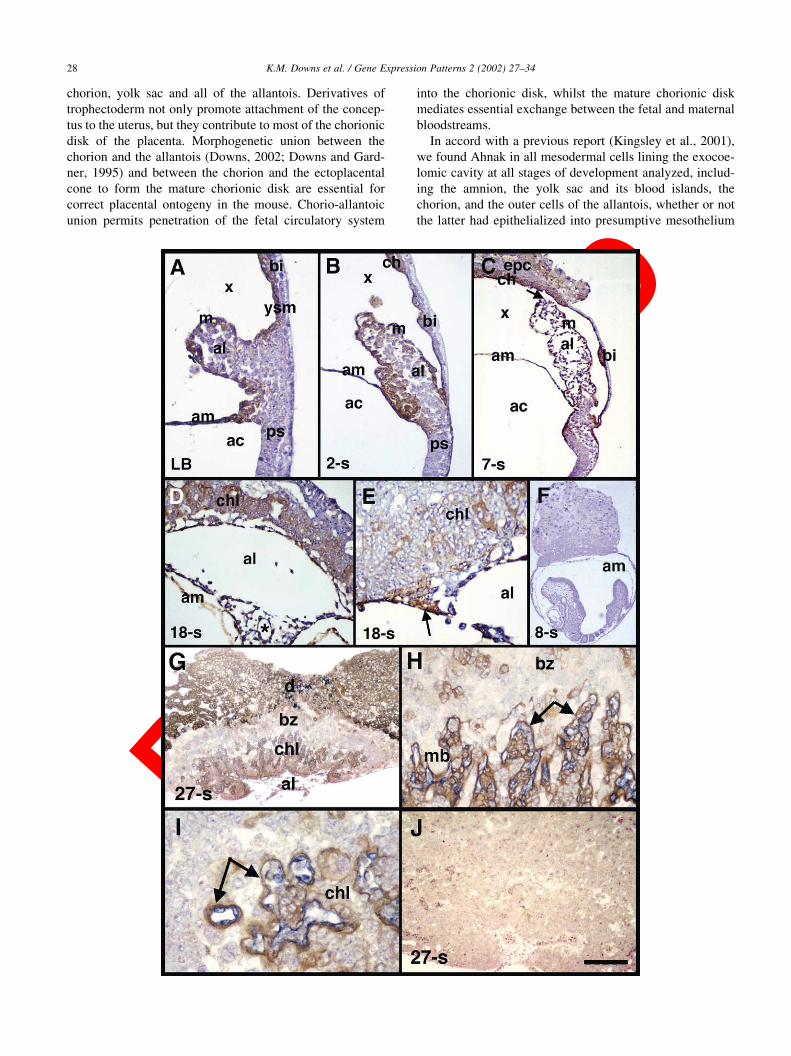
Fig. 1. Ahnak in the murine allantois, components of the exocoelomic cavity, and the chorionic labyrinth. Brightfield photomicrographs localizing Ahnak alone
(panels A–E; brown color) or with Flk-1 (panels G–I; blue color) to sagittal 6 mm thick histological sections counterstained with hematoxylin (A–F) or nuclear
fast red (G–J). (A–F): Posterior, right; anterior, left. (A) Neural plate/late allantoic bud (LB) stage, approximately 7.5 dpc. Ahnak is present in the outer cells of
the allantois (al), some distal allantoic core cells, the yolk sac mesothelium (ysm), yolk sac blood islands (bi), and the mesodermal layer of the amnion (am). ac,
amniotic cavity; m, nascent allantoic mesothelium; ps, primitive streak; x, exocoelomic cavity. (B) 2-somite pair stage (approximately 8.0 dpc). Ahnak persists
in the outer cells of the allantois, some distal allantoic core cells, as well as in basal allantoic core cells continuous with amniotic mesoderm, and is visible in the
chorionic ectoderm and mesoderm (both indicated collectively by ‘ch’). (C) 7-somite pair stage (approximately 8.5 dpc) during chorio-allantoic union. Arrow
indicates where the allantois has retracted from its point of fusion with the chorion during fixation (Downs, 2002). epc, ectoplacental cone. (D) 18-somite stage
allantois (approximately 9.5 dpc) after spreading onto the chorion. Ahnak is seen clearly in a nascent blood vessel (surrounding the asterisk), other core
allantoic cells (e.g. possibly mesenchyme and/or nascent smooth muscle), and mesothelium. chl, chorionic labyrinth. (E) Chorio-allantoic fusion junction at 18-
somite pairs showing high levels of Ahnak in peripheral intermingled chorio-allantoic mesoderm (arrow) which is thought to secure the umbilicus to the
chorionic disk (Downs, 2002), as well as in the distal allantoic region (al), including allantoic endothelium, and the nascent chorionic labyrinth (chl). (F) 8-
somite pair conceptus used as a negative control with pre-immune serum. Although the allantois is not visible in this panel, it was always negative. For this and
all experiments shown in subsequent panels, additional negative controls were the omission of primary antibodies. (G–J): 27-somite pair conceptus in which
chorionic disks were snipped away from the fetus at the level of the base of the allantois. (G) Low-magnification sagittal profile of the allantois (al), chorionic
labyrinth (chl), basal zone (bz), and decidual cells (d). (H) High magnification view of Flk-1-positive allantoic vasculature surrounded by Ahnak-positive
syncytiotrophoblast cells (arrows) in the labyrinth. Some trophoblast derivatives of the basal zone exhibited Ahnak staining. mb, maternal blood. (I) Example
of a random cross section through the labyrinth showing the relationship between Flk-1-positive allantoic blood vessels and syncytiotrophoblast-stained Ahnak
(arrows). (J) Pre-immune serum negative control of a similarly staged chorionic disk. Scale bar in (J): 200 mm (A,E,I); 150 mm (B,C); 100 mm (D,H); 30 mm
(F); 50 mm (J); 20 mm (G).
neurulation involves three consecutive modes of neural tube
formation, the major difference between them being varia-
tions in bending of the neural plate (Fig. 4). Expression of
Ahnak was especially robust during Mode 1 neurulation,
being found in surface ectoderm overlying the neural
folds, in putative neural crest lying between it and the
surface ectoderm, and in the dorsal tips of the neural plate
itself (Fig. 4A, B). As the neural folds approached each
K.M. Downs et al. / Gene Expression Patterns 2 (2002) 27–3430
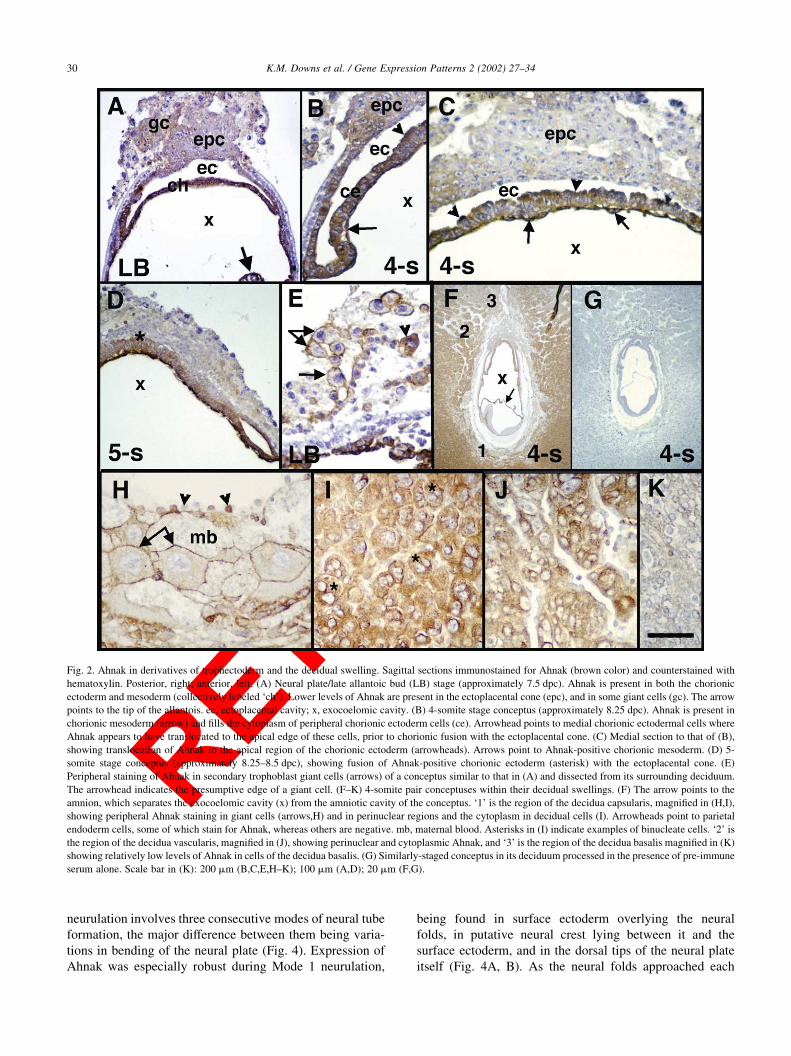
Fig. 2. Ahnak in derivatives of trophectoderm and the decidual swelling. Sagittal sections immunostained for Ahnak (brown color) and counterstained with
hematoxylin. Posterior, right; anterior, left. (A) Neural plate/late allantoic bud (LB) stage (approximately 7.5 dpc). Ahnak is present in both the chorionic
ectoderm and mesoderm (collectively labeled ‘ch’). Lower levels of Ahnak are present in the ectoplacental cone (epc), and in some giant cells (gc). The arrow
points to the tip of the allantois. ec, ectoplacental cavity; x, exocoelomic cavity. (B) 4-somite stage conceptus (approximately 8.25 dpc). Ahnak is present in
chorionic mesoderm (arrow) and fills the cytoplasm of peripheral chorionic ectoderm cells (ce). Arrowhead points to medial chorionic ectodermal cells where
Ahnak appears to have translocated to the apical edge of these cells, prior to chorionic fusion with the ectoplacental cone. (C) Medial section to that of (B),
showing translocation of Ahnak to the apical region of the chorionic ectoderm (arrowheads). Arrows point to Ahnak-positive chorionic mesoderm. (D) 5-
somite stage conceptus (approximately 8.25–8.5 dpc), showing fusion of Ahnak-positive chorionic ectoderm (asterisk) with the ectoplacental cone. (E)
Peripheral staining of Ahnak in secondary trophoblast giant cells (arrows) of a conceptus similar to that in (A) and dissected from its surrounding deciduum.
The arrowhead indicates the presumptive edge of a giant cell. (F–K) 4-somite pair conceptuses within their decidual swellings. (F) The arrow points to the
amnion, which separates the exocoelomic cavity (x) from the amniotic cavity of the conceptus. ‘1’ is the region of the decidua capsularis, magnified in (H,I),
showing peripheral Ahnak staining in giant cells (arrows,H) and in perinuclear regions and the cytoplasm in decidual cells (I). Arrowheads point to parietal
endoderm cells, some of which stain for Ahnak, whereas others are negative. mb, maternal blood. Asterisks in (I) indicate examples of binucleate cells. ‘2’ is
the region of the decidua vascularis, magnified in (J), showing perinuclear and cytoplasmic Ahnak, and ‘3’ is the region of the decidua basalis magnified in (K)
showing relatively low levels of Ahnak in cells of the decidua basalis. (G) Similarly-staged conceptus in its deciduum processed in the presence of pre-immune
serum alone. Scale bar in (K): 200 mm (B,C,E,H–K); 100 mm (A,D); 20 mm (F,G).
other, they became intensely stained for Ahnak (Fig. 4C, D).
Following closure of the neural tube, Ahnak was found in
surface ectoderm overlying the neural tube (Fig. 4E), as well
as within the ‘seam’ at the site of dorsal neuroepithelial
fusion (not shown, but similar to that in Fig. 3J). Similarly,
in both Modes 2 and 3, Ahnak was present in the surface
ectoderm and the dorsal tips of the neuroepithelium prior to
(Fig. 4F–I, K–M ), and during fusion of the neural folds
(Fig. 4J, N). Thereafter, Ahnak was subsequently found in
the surface ectoderm overlying the dorsal neural tube (Fig.
K.M. Downs et al. / Gene Expression Patterns 2 (2002) 27–34 31
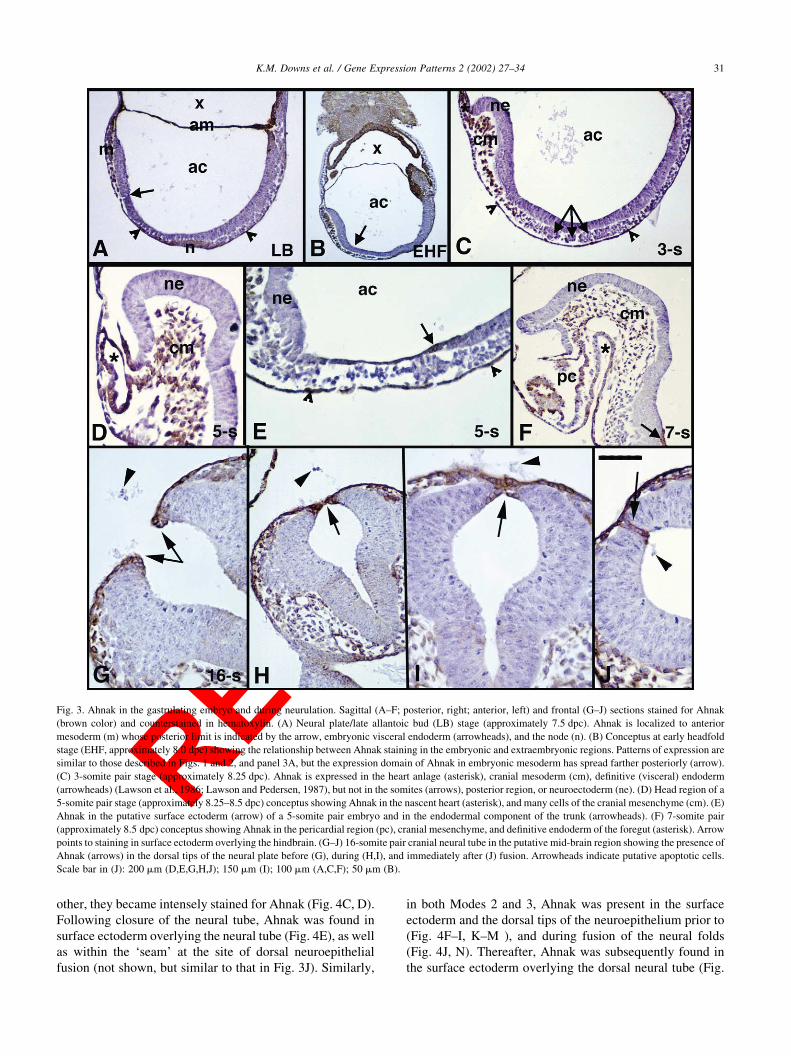
Fig. 3. Ahnak in the gastrulating embryo and during neurulation. Sagittal (A–F; posterior, right; anterior, left) and frontal (G–J) sections stained for Ahnak
(brown color) and counterstained in hematoxylin. (A) Neural plate/late allantoic bud (LB) stage (approximately 7.5 dpc). Ahnak is localized to anterior
mesoderm (m) whose posterior limit is indicated by the arrow, embryonic visceral endoderm (arrowheads), and the node (n). (B) Conceptus at early headfold
stage (EHF, approximately 8.0 dpc) showing the relationship between Ahnak staining in the embryonic and extraembryonic regions. Patterns of expression are
similar to those described in Figs. 1 and 2, and panel 3A, but the expression domain of Ahnak in embryonic mesoderm has spread farther posteriorly (arrow).
(C) 3-somite pair stage (approximately 8.25 dpc). Ahnak is expressed in the heart anlage (asterisk), cranial mesoderm (cm), definitive (visceral) endoderm
(arrowheads) (Lawson et al., 1986; Lawson and Pedersen, 1987), but not in the somites (arrows), posterior region, or neuroectoderm (ne). (D) Head region of a
5-somite pair stage (approximately 8.25–8.5 dpc) conceptus showing Ahnak in the nascent heart (asterisk), and many cells of the cranial mesenchyme (cm). (E)
Ahnak in the putative surface ectoderm (arrow) of a 5-somite pair embryo and in the endodermal component of the trunk (arrowheads). (F) 7-somite pair
(approximately 8.5 dpc) conceptus showing Ahnak in the pericardial region (pc), cranial mesenchyme, and definitive endoderm of the foregut (asterisk). Arrow
points to staining in surface ectoderm overlying the hindbrain. (G–J) 16-somite pair cranial neural tube in the putative mid-brain region showing the presence of
Ahnak (arrows) in the dorsal tips of the neural plate before (G), during (H,I), and immediately after (J) fusion. Arrowheads indicate putative apoptotic cells.
Scale bar in (J): 200 mm (D,E,G,H,J); 150 mm (I); 100 mm (A,C,F); 50 mm (B).
4J, O) and fused seam in some sections of the nascent neural
tube (not shown).
Whilst the brain and upper spinal cord develop by
folding of the neural plate in primary neurulation, second-
ary neurulation involves cavitation of mesoderm to
produce the lower spinal cord (Copp et al., 1990). Ahnak
was not found in the neuroectoderm during secondary
neurulation (Fig. 5A–D), but anti-Ahnak antibodies did
K.M. Downs et al. / Gene Expression Patterns 2 (2002) 27–3432
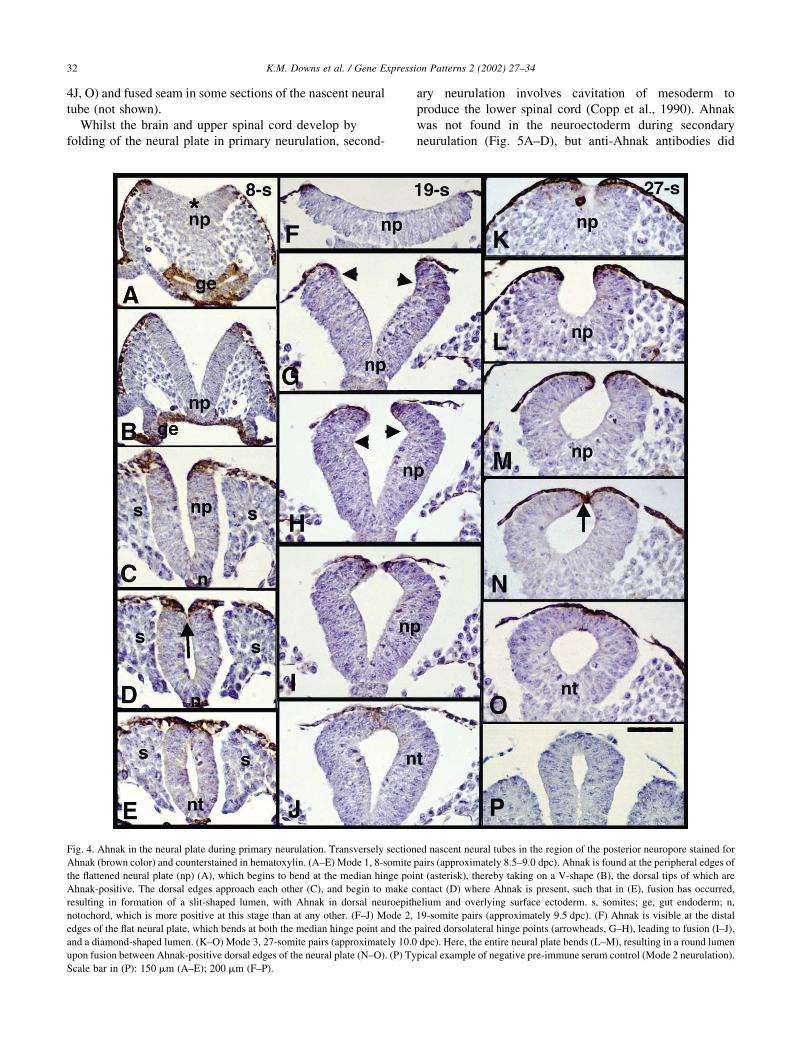
Fig. 4. Ahnak in the neural plate during primary neurulation. Transversely sectioned nascent neural tubes in the region of the posterior neuropore stained for
Ahnak (brown color) and counterstained in hematoxylin. (A–E) Mode 1, 8-somite pairs (approximately 8.5–9.0 dpc). Ahnak is found at the peripheral edges of
the flattened neural plate (np) (A), which begins to bend at the median hinge point (asterisk), thereby taking on a V-shape (B), the dorsal tips of which are
Ahnak-positive. The dorsal edges approach each other (C), and begin to make contact (D) where Ahnak is present, such that in (E), fusion has occurred,
resulting in formation of a slit-shaped lumen, with Ahnak in dorsal neuroepithelium and overlying surface ectoderm. s, somites; ge, gut endoderm; n,
notochord, which is more positive at this stage than at any other. (F–J) Mode 2, 19-somite pairs (approximately 9.5 dpc). (F) Ahnak is visible at the distal
edges of the flat neural plate, which bends at both the median hinge point and the paired dorsolateral hinge points (arrowheads, G–H), leading to fusion (I–J),
and a diamond-shaped lumen. (K–O) Mode 3, 27-somite pairs (approximately 10.0 dpc). Here, the entire neural plate bends (L–M), resulting in a round lumen
upon fusion between Ahnak-positive dorsal edges of the neural plate (N–O). (P) Typical example of negative pre-immune serum control (Mode 2 neurulation).
Scale bar in (P): 150 mm (A–E); 200 mm (F–P).

bind to median dorsal cells of the closed neural tube (Fig.
5E–H).
2. Materials and methods
Polyclonal antibodies (KIS) raised against human
AHNAK peptides have been described previously (Shtivel-
man and Bishop, 1993; Sussman et al., 2001) and are reac-
tive with murine Ahnak as expected based on the identity of
the immunizing peptide sequences between human and
murine Ahnak. Mouse matings, collection and staging of
conceptuses, histology, Bouin’s based immunostaining
and isolation of neural tubes have been previously described
(Downs, 2002; Downs and Davies, 1993; Downs and Gard-
ner, 1995; Shum and Copp, 1996). All materials used here
were fixed in Bouin’s fluid for 2 h at 4 8C, followed by
subsequent rinses in phosphate-buffered saline (PBS;
Sigma) and standard methods of dehydration, clearing and
embedding in paraffin wax. All materials were sectioned at a
thickness of 6 mm. Immunostaining in Bouin’s-fixed mate-
rial was compared within the same experiments with mate-
rial fixed in 4% paraformaldehyde- and 10% formalin-fixed
material. Results revealed Ahnak only in extraembryonic
mesoderm (data not shown), as previously described
(Kingsley et al., 2001) in the latter two fixatives. For each
stage, experiments were carried out minimally in triplicate,
and included negative controls, both pre-immune serum and
omission of antibodies, with no false positive results. Care
and use of laboratory animals for this study were in accor-
dance with the University of Wisconsin Institutional Animal
Care and Use Committee and the Guide for the Care and
Use of Laboratory Animals (National Institute of Health
publication 85-23, revised 1985).
Acknowledgements
K.M.D. gratefully acknowledges funding from the
National Institutes of Health (Grants HD36847 and
HD42706), the Wellcome Trust Programme Grant 051690
to A. J. C., and the institutional support from the Cancer
Research Institute of the University of California-San Fran-
cisco to E. S.
References
Copp, A.J., Brook, F.A., Estibeiro, J.P., Shum, A.S.W., Cockroft, D.L.,
1990. The embryonic development of mammalian neural tube defects.
Prog. Neurobiol. 35, 363–403.
Downs, K.M., 2002. Early placentation in the mouse. Placenta 23, 116–131.
Downs, K.M., Davies, T., 1993. Staging of gastrulation in mouse embryos
by morphological landmarks in the dissection microscope. Develop-
ment 118, 1255–1266.
Downs, K.M., Gardner, R.L., 1995. An investigation into early placental
ontogeny: allantoic attachment to the chorion is selective and develop-
mentally regulated. Development 121, 407–416.
Downs, K.M., Gifford, S., Blahnik, M., Gardner, R.L., 1998. The murine
allantois undergoes vasculogenesis that is not accompanied by erythro-
poiesis. Development 125, 4507–4521.
Downs, K.M., Harmann, C., 1997. Developmental potency of the murine
allantois. Development 124, 2769–2780.
Downs, K.M., Temkin, R., Gifford, S., McHugh, J., 2001. Study of the
murine allantois by allantoic explants. Dev. Biol. 233, 347–364.
Hashimoto, T., Amagai, M., Parry, D.A.D., Dixon, T.W., Tsukita, S.,
Tsukita, S., Miki, K., Sakai, K., Inokuchi, Y., Kudoh, J., Shimizu, N.,
Nishikawa, T., 1993. Desmoyokin, a 680 kDa keratinocyte plasma
membrane-associated protein, is homologous to the protein encoded
by human gene AHNAK. J. Cell Sci. 105, 275–286.
Hieda, Y., Tsukita, S., Tsukita, S., 1989. A new high molecular mass
protein showing unique localization in desmosomal plaque. J. Cell
Biol. 109, 1511–1518.
Kingsley, P.D., McGrath, K.E., Maltby, K.M., Koniski, A.D., Ramchan-
dran, R., Palis, J., 2001. Subtractive hybridization reveals tissue-speci-
fic expression of ahnak during embryonic development. Dev. Growth
Differ. 43, 133–143.
Lawson, K.A., Meneses, J.J., Pedersen, R.A., 1986. Cell fate and cell line-
age in the endoderm of the presomite mouse embryo, studied with an
intracellular tracer. Dev. Biol. 115, 325–339.
Lawson, K.A., Pedersen, R.A., 1987. Cell fate, morphogenetic movement
and population kinetics of embryonic endoderm at the time of germ
layer formation in the mouse. Development 101, 627–652.
Masunaga, T., Shimizu, H., Ishiko, A., Fujiwara, T., Hasimoto, T., Nishi-
kawa, T., 1995. Desmoyokin/AHNAK protein localizes to the non-
desmosomal keratinocyte cell surface of human epidermis. J. Invest.
Dermatol. 104, 941–945.
Nie, Z., Ning, W., Amagai, M., Hashimoto, K., 2000. C-terminus of desmoyo-
kin/AHNAK protein is responsible for its translocation between the
nucleus and cytoplasm. J. Invest. Dermatol. 114, 1044–1049.
K.M. Downs et al. / Gene Expression Patterns 2 (2002) 27–34 33
Fig. 5. Ahnak during secondary neurulation. Transversely-sectioned neural
tube in the region of the posterior neuropore at approximately 31-somite
pairs showing cavitation of mesoderm in the tail bud (asterisk, A) during
secondary neurulation, leading to neural tube formation (B–D). After the
neural tube has formed, Ahnak-positive cells (brown color) appear in the
dorso-medial region (arrowheads, E–G) in some, but not all (H), sections.
Scale bar in (H): 100 mm.

Shtivelman, E., Bishop, J.M., 1992. A human gene (AHNAK) encoding an
unusually large protein with a 1.2 mm polyionic rod structure. Proc.
Natl. Acad. Sci. USA 89, 5472–5476.
Shtivelman, E., Bishop, J.M., 1993. The human gene AHNAK encodes a
large phosphoprotein located primarily in the nucleus. J. Cell Biol. 120,
625–630.
Shum, A.S.W., Copp, A.J., 1996. Regional differences in morphogenesis of
the neuropeithelium suggest multiple mechanisms of spinal neurulation
in the mouse. Anat. Embryol. 194, 65–73.
Sussman, J., Stokoe, D., Ossina, N., Shtivelman, E., 2001. Protein kinase B
phosphorylates AHNAK and regulates its subcellular localization. J.
Cell Biol. 154, 1019–1030.
K.M. Downs et al. / Gene Expression Patterns 2 (2002) 27–3434