respuesta inmune innata pulmonar frente a la infección
TRANSCRIPT

UNIVERSIDAD DE BUENOS AIRES
FACULTAD DE FARMACIA Y BIOQUIMICA
Cátedra de Inmunología e Instituto de Estudios de la Inmunidad Humoral (IDEHU-CONICET)
Respuesta inmune innata
pulmonar frente a la infección
inhalatoria por Brucella abortus
Tesis presentada para optar al título de
Doctor de la Universidad de Buenos Aires
Tesista: Bioq. María Soledad Hielpos
Director: Dr. Pablo César Baldi
Co-directora: Dra. Mariana Cristina Ferrero
2016

Agradecimientos
Al Dr. Pablo Baldi, por haberme aceptado en su laboratorio, por la guía, la confianza y la paciencia. Por todas esas ganas y muchas horas de enseñanza para lograr la superación como profesional
A la Dra Mariana Ferrero, por haber estado incondicionalmente, por las ganas y pujanza que son ejemplo y por todas las horas de BSL3 compartidas
Al Dr. Alberto Fossati, gracias por aceptarme en el grupo y por su amistad.
Al Dr. Diego Comerci, por permitirnos utilizar las instalaciones del Bioterio BLS3 del IIB-INTECH, por las cepas Brucella GFP y Brucella abortus btpAbtpB-. Por las valiosas sugerencias y generosidad.
Al Dr. Sergio Costa, por permitirnos trabajar en su laboratorio, y por sus valiosas sugerencias.
Al Dr. Horacio Salomón y a los integrantes del INBIRS, por permitirnos trabajar en el BSL3.
A la Dra. Silvia Vanzulli, por su gran colaboración en las observaciones microscópicas de los cortes histológicos de pulmón y por su paciencia y buena predisposición.
A los Drs. Jorge Caneva y Alejandro Bertolotti y a Viviana Martinez de Fundación Favaloro por la buena predisposición para colaborar con las muestras pulmonares.
A las instituciones, UBA, CONICET y ANPCyT por brindarme la ayuda económica y la infraestructura necesarias para realizar este trabajo.
A Andrea, y el resto de mis compañeros de laboratorio: Cora, Romi, Florencia, Josefina e Iván. Gracias por su amistad, las discusiones, la valiosa ayuda y los mates. Gracias por hacer del laboratorio un lugar donde querer volver todas las mañanas.
Al resto de mis compañeros del IDEHU, gracias por la buena onda, la ayuda sin condiciones, los mates y la contención. A los que están y a los que siguen su camino por otros lugares.
A mis amigos bioquímicos/farmacéuticos, por la amistad y sostén de tantos años.
A mis amigos músicos, por enseñarme que la vida tiene muchos colores, sonidos, matices y ritmos. Que cada faceta de uno mismo nutre a la otra. No hay belleza más grande que hacer música con ustedes.
A Nico, gracias. Todo resulta más fácil.
A mi Familia: Mamá, Papá y Marian. Gracias por su apoyo incondicional. Por la contención en los buenos y malos momentos. Por las ganas y el motor incansable que son y la felicidad que siento con ustedes.

Parte de los resultados incluídos en este trabajo de Tesis fueron publicados en el siguiente artículo:
CCL20 and beta-defensin 2 production by human lung epithelial cells
and macrophages in response to Brucella abortus infection
M. Soledad Hielpos, Mariana C. Ferrero, Andrea G. Fernández , Josefina
Bonetto , Carlos A. Fossati and Pablo C. Baldi. PLoS ONE 10(10): e0140408.
doi:10.1371/journal.pone.0140408

Resumen.
La brucelosis es una zoonosis causada por bacterias intracelulares gram
negativas del género Brucella, las cuales afectan a animales domésticos y salvajes, y
pueden transmitirse al hombre por diversas vías. Una de estas vías es la inhalación
de aerosoles contaminados. La brucelosis afecta a 500.000 personas en todo el
mundo cada año y en Argentina se producirían anualmente entre 10.000 y 20.000
nuevos casos. La importancia de la vía inhalatoria ha quedado de manifiesto en
brotes epidémicos vinculados a la generación de aerosoles contaminados en
ámbitos laborales, particularmente en laboratorios microbiológicos y en
mataderos. Se estima que se requieren tan sólo 10 a 100 organismos aerosolizados
para generar infección en seres humanos, y es por ello que Brucella se considera
altamente infectiva cuando se adquiere por vía aerógena y es clasificada por el CDC
como agente bioterrorista. A pesar de la importancia de las vías respiratorias para
la entrada de Brucella en el organismo, se desconoce casi por completo la
respuesta inmune pulmonar a la infección inhalatoria por Brucella spp.
El objetivo general de este trabajo de Tesis fue evaluar la interacción de
Brucella abortus con la mucosa respiratoria y sus componentes en las primeras
etapas de la infección para caracterizar la respuesta inmune innata y
antimicrobiana desencadenada en el pulmón, así como evaluar los mecanismos
que utiliza la bacteria para evadir la vigilancia inmunológica pulmonar y
diseminarse sistémicamente. Para ello desarrollamos un modelo murino de
infección intratraqueal por B. abortus que nos permitió evaluar la respuesta
inmune innata durante la primera semana de infección, y analizar un posible
mecanismo de evasión de la inmunidad. También se realizaron estudios in vitro
para evaluar la respuesta de las principales células conformantes de los tejidos
pulmonares.
Los resultados de esta Tesis demuestran que B. abortus genera una
respuesta inmune pulmonar retardada y de baja intensidad, a pesar que es capaz
de infectar y persistir en pulmón murino, pudiéndose diseminar desde el sitio de
infección hacia los órganos periféricos tan tempranamente como 1 día p.i.. Se
detecta en pulmón un leve aumento de las citoquinas IL-1β y MCP-1 a 3 días p.i. y

sólo con cargas bacterianas altas (106 UFC/ ratón), mientras que el resto de las
citoquinas proinflamatorias (IL-12, KC, TNF-α e IFN-γ) no se diferencian del
control hasta 7 días p.i. El análisis histológico demostró que la infección con B.
abortus produce cambios en la composición celular del pulmón afectado,
observándose un leve infiltrado de células mononucleares. Sin embargo, la
citometría no permitió precisar la identidad de las células infiltrantes. Cuando se
infectaron los ratones intratraquealmente con una mutante doble de B. abortus
para las proteínas BtpA y BtpB (btpAbtpB-), involucradas en el bloqueo de la
señalización por TLRs, la histología pulmonar reveló un mayor infiltrado de células
mononucleares que en los pulmones de ratones infectados con la cepa salvaje (wt).
Esto sugiere que las proteínas Btp modulan el reclutamiento de las células
inmunes hacia el pulmón. A diferencia de lo que ocurre en el pulmón, el recuento
de bacterias viables en los órganos periféricos de los ratones infectados por vía
intratraqueal con la doble mutante fue mucho menor que en los ratones infectados
con la cepa wt. Esto podría deberse a que las proteínas Btp modulan los
mecanismos inmunes involucrados en la eliminación de la bacteria en los órganos
periféricos. Los mayores niveles de citoquinas en el bazo de los animales
infectados con la doble mutante avalaron esta hipótesis. Este estudio es el primero
en demostrar in vivo que las proteínas Btp le confieren una ventaja de
supervivencia a B. abortus.
Asimismo se realizaron ensayos para evaluar la capacidad de modulación
de las proteínas Btp frente a agonistas TLR administrados exógenamente. Se halló
que la sobrevida de B. abortus en el ambiente pulmonar previamente inflamado
con LPS de E. coli depende sustancialmente de la expresión de estas proteínas.
Para tener una aproximación a la interacción hospedador-patógeno en
infecciones humanas debe recurrirse a modelos in vitro realizados con células de
origen humano. Para ello se realizaron infecciones de cultivos primarios de
pneumocitos humanos y explantos pulmonares, y se midieron citoquinas y
quemoquinas en sobrenadante de cultivo mediante ELISA. Los resultados indican
que las células epiteliales alveolares son pobres respondedoras frente a la
infección directa por B. abortus en cuanto a secreción de citoquinas

proinflamatorias, al igual que los explantos pulmonares, si bien estos últimos
responden con mayores niveles de citoquinas.
El epitelio respiratorio también responde a las infecciones con la
producción de CCL20 y β-defensinas, los cuales actúan en forma dual como
péptidos antimicrobianos y como quemoquinas. Se evaluó su expresión y secreción
en un modelo de infección in vitro de líneas celulares humanas, THP-1 (monocitos),
Calu-6 (bronquiales) y A549 (alveolares) y en cultivo primario de pneumocitos y
explantos humanos. Además se evaluó in vitro la actividad antimicrobiana de
ambos péptidos frente a B. abortus. Se observó que las células epiteliales humanas
secretan CCL20 y beta-defensina 2 en respuesta a la infección por B. abortus o a las
citoquinas producidas por monocitos infectados. Ambas moléculas son producidas
en concentraciones que no parecen tener actividad antimicrobiana contra B.
abortus, pero que podrían mediar funciones inmunomoduladoras durante la
infección.
En conclusión, la infección inhalatoria por Brucella abortus genera una
respuesta innata pulmonar de poca intensidad que le permite sobrevivir en
pulmón y probablemente facilita su diseminación a otros órganos. La persistencia
pulmonar de la bacteria posiblemente se vea facilitada, entre otros factores, por su
capacidad de supervivencia intracelular y sus mecanismos de modulación de la
respuesta inmune innata.

Indice.
Introducción general 2
1 Historia 2
2 Generalidades de la brucelosis 2
3 Brucella spp 4
3-1 Caracteristicas del género 4
3-2 Estructura antigénica 6
4 Mecanismos patogénicos de Brucella.Factores de virulencia y mecanismo de evasión del sistema inmune
8
4-1 Glucano β1-2 cíclico inhibe la fusión con el lisosoma 10
4-2 El sistema de secreción tipo IV 10
4-3 Brucella TIR containing protein (Btp) 11
4-4 LPS 12
5 Contagio inhalatorio de la brucelosis 13
6 Principales células de las vías respiratorias 14
6-1 Células epiteliales 14
6-2 Macrófagos alveolares 15
6-3 Células dendríticas 17
7 La interacción de los patógenos con la mucosa respiratoria 18
7-1Reconocimiento de patógenos por las células de las vías respiratorias. 18
7-2 La respuesta del epitelio respiratorio a los patógenos 22
7-3 Mecanismos de infección a través de la mucosa respiratoria 23
Hipótesis de trabajo 25
Objetivos 26
Capítulo 1 Respuesta inmune a la infección intratraqueal con B. abortus y efecto modulador de las proteínas Btp
27
Introducción 28
Materiales y Métodos 31
Resultados 41
Conclusiones 70
Capítulo 2 Caracterización de la infección in vitro de pneumocitos tipo II primarios humanos con B. abortus 2308
84
Introducción 85
Materiales y Métodos 88
Resultados 92
Conclusiones 99

Capítulo 3 Respuesta antimicrobiana pulmonar frente a infecciones con B. abortus
103
Introducción 104
Materiales y Métodos 107
Resultados 112
Conclusiones 126
Conclusiones 132
Bibliografía 135

Introducción general

Introducción General
2
1-Historia
Las bacterias del género Brucella fueron aisladas por primera vez en 1887
por David Bruce de los bazos de soldados que morían en la isla de Malta por una
enfermedad conocida en ese entonces como Fiebre de Malta. El reservorio del
agente causal no fue descubierto hasta 1904, cuando se lo aisló de orina y leche de
cabras aparentemente sanas. Cuando se suspendió el consumo de leche cruda de
cabra, la incidencia de la enfermedad descendió en forma brusca. La bacteria se
denominó inicialmente Micrococcus melitensis, y luego fue rebautizada con el
nombre de Brucella melitensis en honor a su descubridor. El segundo
microorganismo del grupo fue aislado por Bang en Dinamarca en 1897 de ganado
que padecía de aborto infeccioso (enfermedad de Bang), y al que inicialmente se
denominó Bacillus abortus. El tercer miembro del grupo fue cultivado en EE.UU. en
1914 del feto de una cerda expulsado prematuramente (Bacillus suis). En 1920
Evans reconoció que los tres microorganismos estaban estrechamente
emparentados y los incluyó en un género aparte, denominado Brucella (Rivero
Puente et al., 1978).
2- Generalidades de brucelosis
La brucelosis es una zoonosis endémica causada por bacterias Gram
negativas del género Brucella, las cuales se comportan como parásitos
intracelulares facultativos. Esta enfermedad afecta al hombre y a distintas especies
animales, incluyendo varias relacionadas con la alimentación y la actividad laboral
ganadera (Serre et al., 1989).
La infección afecta primariamente al aparato genito-urinario del ganado
doméstico como bovinos, ovinos, caprinos y porcinos, como así también de caninos
y de especies salvajes como camellos, bisontes, búfalos, roedores, delfines, etc.
(FAO-WHO Joint Expert Committee on Brucellosis., 1986). La enfermedad del

Introducción General
3
ganado causa disminución de la capacidad reproductora, abortos, parición de
terneros débiles y contaminación de lácteos y carnes. Resulta entonces un
importante problema sanitario y económico.
La incidencia de la brucelosis es muy alta en nuestro país, donde no sólo hay
un importante consumo de lácteos y carnes, sino que además la producción y
elaboración de estos productos es una de las principales actividades económicas,
convirtiendo a la brucelosis en una de las enfermedades laborales más
importantes. Datos obtenidos por el SENASA en el año 2005 muestran que la
prevalencia de la brucelosis en los bovinos a nivel nacional fue del 2.15%. Sin
embargo, una encuesta realizada el mismo año por el INTA Bariloche, la Dirección
de Ganadería de la provincia de Río Negro y el Laboratorio de Brucelosis de la
Fundación Barrera Patagónica revelaron que la prevalencia de la enfermedad en la
ganadería bovina es más elevada que la informada por el SENASA. Los resultados
mostraron un 21.4% de establecimientos positivos a Brucella abortus -con un
rango de 10% al 40%- y una tasa de reaccionantes del 3.7%.
La transmisión de la brucelosis al hombre se produce por contacto con el
animal infectado o sus tejidos o secreciones de animales infectados, y/o por
consumo de lácteos no pasteurizados o carne insuficientemente cocida. Por lo
tanto, constituye una enfermedad laboral y alimentaria, y la población de mayor
riesgo se encuentra conformada por trabajadores de la industria de la carne,
veterinarios, peones de campo, transportistas de ganado o de productos derivados
de éste, y empleados de laboratorios de diagnóstico microbiológico o de
producción de cepas vacunales de Brucella. La infección puede ocurrir también en
personas (residentes o turistas) que tienen contacto con animales infectados en
zonas endémicas o que consumen alimentos derivados del ganado infectado
(especialmente lácteos no pasteurizados) (Crespo León et al., 1993; Moreno-Lafont
et al. 1995). La entrada de las bacterias se puede producir por contacto directo de
la piel lesionada con material infectado, por el ingreso de aerosoles a través de las
vías conjuntival o inhalatoria, o por ingestión de alimentos contaminados. Además,
la entrada por inoculación accidental con cepas vacunales es frecuente en peones y
veterinarios (Serre et al., 1989; Wallach et al. 1997; Bouza et al. 2005).

Introducción General
4
Cuando las bacterias del género Brucella infectan al hombre pueden
provocarle una severa infección aguda, que cursa con bacteriemia. Si la
enfermedad se diagnostica tempranamente y se trata en forma adecuada tiene una
evolución en general favorable. Aunque actualmente constituye una enfermedad
de baja mortalidad, en algunos individuos deja secuelas discapacitantes de
magnitud variable. La brucelosis humana cursa con fiebre, adenopatías,
hepatoesplenomegalia, mialgias, artralgias, astenia y sudores. Se han propuesto
muchas clasificaciones de los estadios clínicos de la brucelosis, pero el más
aceptado es aquel que define enfermedad aguda o crónica en base al tiempo de
evolución de los síntomas (Young et al., 1989). Se considera pacientes agudos a
aquellos que refieren haber padecido síntomas compatibles con brucelosis durante
un lapso de hasta 1 año previo a la consulta. Mientras que se clasifican como
crónicos aquellos pacientes que refieren haber padecido dichos síntomas durante
un lapso superior a 1 año.
En las formas agudas de la enfermedad, la administración precoz y
prolongada de una apropiada combinación de antibióticos provoca la remisión del
cuadro clínico en al menos el 90% de los pacientes. La asociación de doxiciclina y
estreptomicina constituye el tratamiento de elección ya que en estas condiciones
se presenta un menor porcentaje de pacientes con recaídas (Montejo et al. 1993)La
administración de doxiciclina y rifampicina es empleada como tratamiento
alternativo.
Es interesante notar que la virulencia y la tendencia a la cronicidad varían
entre especies de Brucella, siendo B. abortus y B. suis las de mayor tendencia a la
cronicidad. Por el contrario, B. melitensis produce las infecciones más fulminantes,
tanto para humanos como para animales (Young et al., 1989).
3- Brucella spp.
3.1. Características del género
Desde la creación del género Brucella en 1920 se fueron incorporando al
mismo nuevas especies y biotipos. Se caracterizaron tres nuevas especies (B. ovis,
B. neotomae, y B. canis) y las especies de B. abortus y B. suis se subdividieron en

Introducción General
5
ocho y cuatro biotipos respectivamente. En el resto de las especies de Brucella no
se caracterizaron biotipos diferenciados (Rivero Puente et al., 1978). Se ha
detectado la infección por Brucella en mamíferos marinos debida a dos nuevas
especies: B. ceti y B. pinnipedialis (Godfroid et al. 2005). En los últimos años se ha
aislado una nueva especie, B. microti, a partir de topillos de campo (Microtus
arvalis) y de zorros (Scholz et al. 2008; Scholz et al. 2009)y otra especie, B.
inopinata, en una paciente con un implante mamario infectado (Scholz et al. 2010).
Las bacterias del género Brucella pertenecen a la clase -proteobacterias,
orden Rhizobiales, familia Brucellaceae. El orden Rhizobiales incluye también,
entre otros, a los géneros Bartonella, Afipia, Ochrobactrum, Agrobacterium,
Rhizobium, Bradyrhizobium, Mesorhizobium y Sinorhizobium. Según el tipo de
colonias que forman en cultivo las distintas especies del género Brucella se
clasifican en lisas o rugosas. Las especies lisas son: B. abortus, B. melitensis, B.
neotomae, B. suis, B. pinnipedialis y B. ceti; las especies rugosas son: B. canis y B.
ovis.
De las especies del género Brucella identificadas hasta el presente sólo cinco
son comprobadamente patógenas para el hombre: B. melitensis, cuyos reservorios
naturales son cabras y ovejas; B. abortus, que infecta principalmente a bovinos; B.
suis a porcinos, B. canis a caninos, y B. pinnipedialis y B. ceti a mamíferos marinos.
En cambio, B. neotomae y B. ovis, cuyos reservorios naturales son ratas y ovejas,
respectivamente, no provocan enfermedad humana (Ko & Splitter 2003). (Tabla
1). La patogenicidad para el hombre de las nuevas especies (B. microti y B.
inopinata) no ha sido claramente establecida.

Introducción General
6
Tabla 1: Características de las diferentes especies del género Brucella.
Especie
Hospedador
Patogenicidad-
Hombre
B. abortus Bovinos moderada
Especies
lisas
B. melitensis caprinos y ovinos elevada
B. suis Porcinos moderada
B. neotomae Roedores no posee
B. pinnipedialis/
B. ceti Mamíferos marinos moderada
B. ovis Ovinos no posee Especies
rugosas B. canis Caninos moderada
Los miembros del género Brucella son bacilos cocoides, pequeños, Gram
negativos, inmóviles, no esporulantes y que no producen cápsulas ni flagelos. Estas
bacterias son microorganismos intracelulares facultativos capaces de sobrevivir y
multiplicarse dentro de las células del sistema reticuloendotelial y tejidos
asociados; se considera que esta cualidad es el factor que les confiere virulencia.
Son capaces de infectar a fagocitos profesionales (macrófagos, neutrófilos,
linfocitos B y células dendríticas)(Young 1995; Elzer et al. 1996; Porte et al. 1999;
Zwerdling et al. 2009) y no profesionales (células HeLa, células Vero, epitelio
alveolar) (Detilleux et al. 1990a; Ferrero et al. 2009)
3.2. Estructura antigénica
Siendo las bacterias del género Brucella microorganismos Gram negativos,
su envoltura celular está formada por una membrana interna, una membrana
externa y un espacio periplasmático intermedio (Ilustración 1). Este último
contiene algunas enzimas y proteínas relacionadas con el transporte de solutos, y
un gel glucopeptídico denominado peptidoglicano, responsable de la forma e
integridad osmótica de la bacteria.

Introducción General
7
Ilustración 1: Esquema representativo de la membrana de Brucella spp.
La membrana externa contiene distribuidos asimétricamente fosfolípidos,
proteínas y un lipopolisacárido (LPS) considerado el principal antígeno (Ag) de la
bacteria. El LPS consta de una parte glucolipídica (Lípido A) inserta en la
membrana externa, y por tanto no expuesta en la superficie, y otra exclusivamente
polisacarídica dirigida hacia el exterior actuando como soporte o unión del resto
de la molécula a la superficie bacteriana. La parte polisacarídica del LPS se divide
en dos secciones: el núcleo, más interno y la cadena O. El Ag “O” es la porción
antigénicamente dominante de la molécula y está constituido por unidades
repetitivas de oligosacáridos en los cuales suelen residir los determinantes
antigénicos del LPS de las cepas lisas. Este segmento está ausente en el LPS de las
cepas del tipo rugoso.
Asociadas al LPS y formando parte de la membrana se encuentran proteínas
de la membrana externa (Omps), muchas de las cuales han sido caracterizadas
desde el punto de vista genético, inmunológico, estructural y funcional (Cloeckaert
et al. 2002). Estas proteínas se han dividido en tres grupos de acuerdo a su peso
molecular: proteínas de entre 89 y 94 kDa (grupo 1), entre 35 y 40 kDa (grupo 2), y
entre 25 y 30 kDa (grupo 3). Las proteínas de los grupos 2 y 3 se encuentran en

Introducción General
8
mayor cantidad y son conocidas como Omp “mayores”. Las proteínas del grupo 1
junto con otras con pesos moleculares entre 10 y 20 kDa, son denominadas Omp
“menores”. Se conoce que las proteínas del grupo 2 funcionan como porinas
(Douglas et al. 1984; Mobasheri et al. 1997; Marquis & Ficht 1993; Cloeckaert et al.
1996) mientras que las proteínas del grupo 3 son similares a la OmpA de E. coli
(Verstreate et al. 1982). Tibor et al. (1999) han demostrado que las Omp 10, 16 y
19 son lipoproteínas. A través de diversos estudios se ha confirmado la
inmunogenicidad de las Omp y su posible utilidad como vacunas (Bowden et al.
1998; Winter et al. 1983; Pasquevich et al. 2009).
El espacio periplasmático, comprendido entre la membrana externa y la
citoplasmática está constituido por el mucopéptido, formado por peptidoglucano
conteniendo glucosamina, ácido murámico, alanina, ácido glutámico y ácido
-diaminopimélico. En este espacio se han identificado proteínas tales como
p39(de Fays et al. 1999), bp26 (Rossetti et al. 1996) y Cu/Zn superóxido dismutasa
(Bricker et al. 1990). Además se ha informado la asociación de lipoproteínas con el
peptidoglucano (Cloeckaert et al. 1992) y con la membrana externa (Gómez-Miguel
et al. 1987). La fracción citoplasmática contiene ADN, ARN, la fracción ribosomal, y
una gran cantidad de proteínas solubles.
4- Mecanismos patogénicos de Brucella. Factores de
virulencia y mecanismos de evasión inmune.
Las bacterias patogénicas han desarrollado varias estrategias para contrarrestar
los mecanismos de defensa del huésped y crear un nicho de replicación favorable.
Algunas de estas estrategias afectan a mecanismos de la respuesta inmune innata,
tales como la muerte por fagocitos profesionales. Las bacterias pueden evadir la
degradación que ocurre en la vía endocítica inhibiendo la fagocitosis, manipulando
componentes de la célula huésped tales como las Rho GTPasas, modificando el
tráfico de la vía endocítica, etc.
Brucella spp. son bacterias intracelulares facultativas que replican dentro
de la célula huésped. Incluso se ha propuesto considerarlas “parásitos
intracelulares facultativamente extracelulares”, teniendo en cuenta que su forma
de vida habitual es la intracelular y que facultativamente pueden crecer en medios

Introducción General
9
nutritivos (Moreno & Moriyon 2002). Los factores bacterianos implicados en la
supervivencia y replicación intracelular constituyen los principales factores de
virulencia, ya que las mutantes defectivas para éstos han resultado avirulentas
(Hong et al. 2000; Lestrate et al. 2000).
Brucella reside en los tejidos del sistema retículoendotelial y sobrevive
intracelularmente en macrófagos, lo cual es esencial para el establecimiento,
desarrollo y cronicidad de la enfermedad. Ha sido propuesto, y parcialmente
demostrado, que la supervivencia intracelular de Brucella en macrófagos resulta
de su capacidad de manipular la vía endocítica, inhibiendo la fusión entre la
vacuola que contiene a la bacteria y los lisosomas. Se ha observado que las especies
con LPS liso pueden evadir la fusión de la vesícula que las contiene con el lisosoma
mientras que las especies con LPS rugoso (carecen del antígeno O) se unen
rápidamente a los lisosomas y son eliminadas. El mecanismo por el cual la cadena
O del LPS permite a la bacteria evitar la fusión con el lisosoma puede estar
determinado por interacciones con un receptor particular y/o por modificaciones
en las propiedades de la membrana de la vacuola que contiene a la bacteria.
Brucella lisa muerta por calor también puede retardar la fusión con el lisosoma
(Porte et al., 2003). Sin embargo, el retardo dependiente de la cadena O del LPS es
un fenómeno transitorio y no sería suficiente para la superviviencia intracelular
por largo tiempo de la bacteria lisa viva, sugiriendo que es necesaria la presencia
de otros factores para asegurar la supervivencia intracelular de la bacteria.
Brucella reside en un comparimiento acidificado que se fusiona con
componentes de la vía endocítica temprana inmediatamente después de su
entrada en el macrófago (Porte et al. 1999; Celli et al. 2003).. Allí es expuesta al
estallido oxidativo (Jiang et al. 1993). Luego, las cepas virulentas de Brucella llegan
al fagosoma replicativo o “brucelosoma”, donde es inhibida la fusión con el
lisosoma y el pH aumenta (Roop et al. 2004; Köhler et al. 2003). En células HeLa,
Brucella abortus y Brucella melitensis previenen la fusión con los lisosomas,
escapando así de la vía endocítica antes de interaccionar con los endosomas
tardíos. Estas bacterias transitan en una vacuola con características de
autofagosoma y se replican en un compartimiento caracterizado por la presencia
de marcadores de retículo endoplasmático (Pizarro-Cerdá, Méresse, et al. 1998;

Introducción General
10
Pizarro-Cerdá, Moreno, et al. 1998; Delrue et al. 2001). En coincidencia con estos
hallazgos, observaciones en células Vero o trofoblastos de cabras preñadas han
mostrado la presencia de Brucella spp. replicándose en el retículo endoplásmico
(Anderson & Cheville 1986; Detilleux et al. 1990b). Se han identificado dos factores
relacionados con la evasión de la fusión fagosoma-lisosoma: el glucano cíclico y el
aparato de secreción de Tipo IV.
4-1. Glucano 1-2 cíclico inhibe la fusión con el lisosoma.
Un mecanismo usado por Brucella para modular el tráfico intracelular
involucra al Glucano 1-2 cíclico (Arellano-Reynoso et al. 2005). Este componente
periplasmático es producido por organismos de la subdivisión -2 proteobacteria,
en los que está involucrado en la osmoregulación. En el caso de Brucella, se ha
demostrado que las mutantes en el gen codificante para la sintetasa de este
compuesto (cgs) son incapaces de evitar la fusión con los lisosomas o alcanzar la
organela replicativa derivada del retículo endoplasmático, indicando que la
producción de Glucano 1-2 cíclico es necesaria para el apropiado tráfico
intracelular (Arellano-Reynoso et al. 2005; Briones et al. 2001). De acuerdo con
este resultado, el agregado de Glucano 1-2 cíclico exógeno a la mutante cgs
durante el crecimiento previo a la infección restablece la capacidad de la bacteria
de manipular el tráfico intracelular (Arellano-Reynoso et al. 2005).
4-2. El sistema de secreción de Tipo IV.
Aunque las vesículas que contienen Brucella (VCB) experimentan algunas
interacciones con la vía endocítica, estas interacciones son transitorias. En estadios
tempranos de la maduración de la VCB se observa la acidificación de la vacuola, lo
cual coincide con la adquisición de un marcador asociado al estadio endosomal
tardío, la glicoproteína-1 de membrana asociada al lisosoma (LAMP-1) (Porte et al.
1999; Celli et al. 2003). Sin embargo la VCB no adquiere otros marcadores de la vía
endocítica (Pizarro-Cerdá, Méresse, et al. 1998; Celli et al. 2003), indicando que la
vesícula se desvía de la vía endocítica antes de alcanzar el estado de endosoma
tardío. No se conoce aún el mecanismo por el cual la VCB adquiere LAMP-1 y si la
adquisición temprana de este marcador es necesaria para la maduración de la VCB.
En contraste, se sabe que la acidificación es esencial para la supervivencia de

Introducción General
11
Brucella y es necesaria para la expresión intracelular del aparato de secreción de
Tipo IV de Brucella codificado por el operón virB (Boschiroli et al. 2002).
El operón virB, identificado en distintas especies de Brucella (Hong et al.
2000; Lestrate et al. 2000; Delrue et al. 2001; O’Callaghan et al. 1999; Foulongne et
al. 2000; Sieira et al. 2000), es fundamental para la supervivencia intracelular de la
bacteria. Esta región es homóloga al operón virB de A. tumefaciens, el cual incluye
los genes de 12 proteínas (VirB1 a VirB12) que conforman un aparato de secreción
de Tipo IV (O’Callaghan et al. 1999). Estos sistemas se encargan de la secreción o
exportación de proteínas o complejos de nucleoproteínas, y muchos se encuentran
en patógenos animales o vegetales como A. tumefaciens, Bordetella pertussis,
Helicobacter pylori, o el patógeno intracelular Legionella pneumophila (Christie
2001).
Un operón virB funcional es necesario para el establecimiento y
mantenimiento de la infección persistente por Brucella, y se ha observado que
varias mutantes en genes virB son avirulentas en el modelo murino (Hong et al.
2000; Lestrate et al. 2000; Sieira et al. 2000; Delpino et al. 2009). A su vez, este
sistema es necesario para la supervivencia intracelular en macrófagos, células
HeLa y osteoblastos (Delrue et al. 2001; O’Callaghan et al. 1999; Foulongne et al.
2000; Sieira et al. 2000; Delpino et al. 2009). Se ha demostrado que el operón virB
está involucrado en la maduración de la vacuola que contiene a Brucella,
convirtiéndola en una organela que permite la replicación de la bacteria (Delrue et
al. 2001; Comerci et al. 2001). En años recientes se han identificado en Brucella
algunas proteínas que serían secretadas en forma dependiente de la expresión de
genes virB, lo cual hace presumir que son secretadas por el aparato de secreción
tipo IV(Marchesini et al. 2011; Myeni et al. 2013).
4-3. Brucella TIR containing protein (Btp)
Recientemente se ha demostrado que Brucella abortus expresa dos
proteínas con dominios TIR llamadas BtpA y BtpB, las cuales son secretadas por el
sistema de secreción de tipo IV de la bacteria y se localizan en el citosol de las
células infectadas. BtpA y BtpB interfieren en la señalización de TLR2, TLR4 y
TLR5, y están involucradas en la capacidad de B. abortus de reduccir la maduración

Introducción General
12
de las células dendríticas y de inhibir la secreción de citoquinas proinflamatorias
(Salcedo et al 2008 y 2013), por lo que serían un importante factor de regulación
de la inmunidad innata. Además se ha demostrado que BtpA purificada inhibe la
muerte celular inducida por células T CD8+ y de esta manera controla la respuesta
inmune adaptativa (Durward et al. 2012).
En estudios in vivo, se demostró que B. abortus mutante para BtpB se
encuentra atenuada en un modelo de infección intraperitoneal de ratones
inmunocomprometidos carentes del factor regulador de interferón (IFR-1-/-), sin
embargo no se vió atenuación en ratones Balb/c cepa salvaje (Salcedo et al. 2013).
Por lo tanto, a pesar de haberse demostrado su capacidad inmunomoduladora in
vitro, aún se desconoce el rol de las proteínas Btp durante la infección por Brucella
en animales inmunocompetentes.
4-4. LPS
El LPS de Brucella posee multiples propiedades que le otorgan a la bacteria
la capacidad para evadir parcialmente al sistema immune. Exhibe caracteristicas
particulares que le permite evadir la detección por el receptor TLR4. La mayoria de
los LPS de las bacterias entéricas poseen un lípido A con ácidos grasos cortos,
mientras que en Brucella los mismos son largos (Lapaque et al. 2006), lo cual, junto
con patrones de glicosilacion del polisacárido (Gil-Ramírez et al. 2014), hacen que
las caracteristicas de endotoxina se vean disminuídas debido a una activación casi
nula del receptor TLR4. Se ha observado, por ejemplo, que la dosis de LPS de B.
abortus necesaria para inducir la producción de un determinado nivel de TNF-α
por monocitos humanos es 1000 veces mayor que la dosis de LPS de Escherichia
coli(Giambartolomei et al. 2004). Además, el LPS no canónico de Brucella le
confiere resistencia a la activación del complemento por la vía de las lectinas y
resistencia a la actividad microbicida de algunos péptidos antimicrobianos
(Fernandez-Prada et al. 2001; Corbeil et al. 1988).

Introducción General
13
5- Contagio inhalatorio de la brucelosis
Como se mencionó anteriormente, el hombre puede adquirir la infección
por Brucella por contacto directo de la piel lesionada con material infectado, pero
las formas más frecuentes de infección son el ingreso de aerosoles a través de las
vías conjuntival o inhalatoria, y la ingestión de alimentos contaminados.
El ingreso por mucosas es también la forma principal de contagio entre los
animales, aunque en este caso se suma la vía venérea y por lo tanto la mucosa
genital (no relevante para la infección humana). Los animales tienen por
costumbre oler y lamer los restos fetales o placentarios, que en el caso del aborto
por Brucella spp. contienen enormes cantidades de estas bacterias.
La transmisión de Brucella por vía aérea en empleados de laboratorios
productores de la cepa vacunal o de laboratorios de análisis microbiológicos ha
sido ampliamente documentada (Memish & Mah 2001; Wallach et al. 1997; Martin-
Mazuelos et al. 1994; Staszkiewicz et al. 1991; Ollé-Goig & Canela-Soler 1987;
Montes et al. 1986; Nelson et al. 1975; SULKIN 1961; TREVER et al. 1959;
Huddleson & Munger 1940). La forma más frecuente de adquisición de la infección
es la inhalación de aerosoles contaminados producidos de forma accidental
durante la manipulación de esta bacteria (Martin-Mazuelos et al. 1994). De hecho
la brucelosis es considerada una de las infecciones más comunes entre las
adquiridas en laboratorios microbiológicos y los aerosoles han sido implicados en
la mayoría de los casos (Yagupsky & Baron 2005). También se han producido
brotes de brucelosis humana en mataderos debidos a aerosoles generados durante
la faena de animales infectados (Kaufmann et al. 1980), y en trabajadores rurales,
entre otras causas, por la manipulación de guano seco de animales infectados que
es empleado como fertilizante (Wallach et al. 1997).
Las numerosas evidencias sobre la transmisión de esta bacteria por
aerosoles han hecho que Brucella sea considerada como un agente potencial de
bioterrorismo (Miller et al. 1987; Davos et al., 1981)En la década de 1950, B. suis
fue el primer agente en un arma biológica en los Estados Unidos, y otros países han
considerado emplear a Brucella para el mismo fin (Bossi et al. 2004; Pappas et al.
2006). Se ha estimado que se requieren tan sólo 10 a 100 organismos

Introducción General
14
aerosolizados para generar infección en seres humanos, y es por ello que Brucella
se considera altamente infectiva cuando se adquiere por vía aerógena (Bossi et al.
2004). Como resultado, B. melitensis, B. abortus y B. suis han sido clasificados como
agentes para bioterrorismo de categoría B por el Centro para el Control y la
Prevención de Enfermedades Infecciosas (CDC) (Bhattacharjee et al. 2006; Pappas
et al. 2006). La ausencia de una vacuna para uso en seres humanos aumenta la
preocupación sobre el posible uso de Brucella como arma biológica.
6 Principales células de las vías respiratorias
6-1. Células epiteliales
La mucosa respiratoria incluye distintos tipos de células epiteliales según el
nivel anatómico que se considere. El epitelio de los conductos aéreos es un epitelio
columnar pseudoestratificado el cual está compuesto por tres tipos celulares:
células ciliadas, células secretorias y células basales. Esta capa de epitelio
pseudoestratificado disminuye gradualmente su grosor desde las vías aéreas más
grandes (tráquea y bronquios) hasta formar una capa muy delgada de epitelio en
los bronquiolos terminales (Ilustración 2). Las células epiteliales bronquiales
constituyen una barrera física estrecha recubierta por una capa de moco que
impide la penetración de agentes patógenos al cuerpo. El moco y los movimientos
ciliares les permiten a las células epiteliales bronquiales atrapar y eliminar los
microorganismos. El epitelio bronquial también produce sustancias
antimicrobianas, incluyendo lisozima, lactoferrina, colectinas, y péptidos
antimicrobianos, que proporcionan una cubierta protectora adicional. Además de
estas funciones, que siempre están activas, las células epiteliales bronquiales
también son capaces de reconocer y responder a los microorganismos, secretando
citoquinas que dan inicio a una respuesta inflamatoria local (Kaiser & Diamond
2000).
El epitelio alveolar forma una capa muy delgada de células que cubren el
99% de la superficie interna del pulmón y está compuesto por dos tipos de células
morfológicamente distintas llamadas neumocitos tipo I y tipo II. Los neumocitos
tipo I tienen un núcleo protruido y extensiones citoplasmáticas extremadamente
finas, cubren más del 90% de la superficie del espacio alveolar. Su principal

Introducción General
15
función es participar del intercambio gaseoso en el alvéolo. Las funciones de los
neumocitos de tipo II son la producción y almacenamiento de surfactante, el
transporte de solutos e iones y la reparación alveolar (Ilustración 2). Además, se
ha demostrado que los neumocitos tipo II juegan un rol muy importante en la
regulación de la respuesta inmune en pulmón. Expresan moléculas de
histocompatibilidad de clase II, moléculas de adhesión (Cunningham et al. 1994) y
secretan citoquinas en respuesta a patógenos (Gentry et al. 2007).
La mayoría de los estudios in vitro sobre interacción del epitelio pulmonar
humano con patógenos han sido realizados con líneas de células bronquiales
(principalmente BEAS-2B y 16HBE14o-) y de células alveolares (línea A549 de
neumocitos tipo II).
6-2. Macrófagos alveolares
Los macrófagos alveolares (MA) constituyen aproximadamente el 93% de la
población de macrófagos pulmonares (Marriott & Dockrell 2007). Los MA están
estratégicamente situados en la luz de los alvéolos y los conductos alveolares y son
por lo tanto las primeras células del sistema inmune en contactar los patógenos y
antígenos inhalados en el tracto respiratorio inferior. Los MA poseen una vida útil
de aproximadamente 3 meses (Thomas et al. 1976). La longevidad de estas células
se debe a mecanismos de resistencia a la apoptosis constitutiva(Liu et al. 2001).
Hay aproximadamente un MA en cada alvéolo. Los monocitos de la sangre al
ingresar en los pulmones se diferencian en MA bajo la influencia de factores locales
(células alveolares de tipo II, epitelio bronquial, citoquinas, surfactante), aunque la
replicación in situ de los MA también contribuye a mantener la población total en
el pulmón (Blussé van Oud Alblas et al. 1981). Los MA, a diferencia de los
macrófagos intersticiales de pulmón, poseen alta capacidad fagocítica y secretan
mayores cantidades de TNF-α en respuesta a patógenos, pero son menos eficientes
en la presentación de antígenos (Franke-Ullmann et al. 1996).

Introducción General
16
Ilustración 2: Ilustración del epitelio de los pulmones en las vías respiratorias
superiores (bronquios) (a), las vías respiratorias inferiores (bronquiolos) (b), y los
alvéolos (c). (A) Células 16HBE14o (1000 x): análisis de microscopía electrónica de células
cultivadas en Transwell durante 1 semana a una densidad de 105 células/ cm2. La capa de
células 16HBE14o se compone de células cuboidales. (B) Células Calu-3 (2600 x): Análisis
de microscopía electrónica de las células cultivadas en Transwell durante 2 semanas con
una densidad de 2x105 células/cm2. La monocapa de células Calu-3 está compuesta por
células con forma de columnas. (C) monocapa hAEpC (1000 x): Análisis de microscopía
electrónica de las células cultivadas por 1 semana en Transwell a una densidad de 6x105
células/cm2. Las células hAEpC poseen una morfología similar al de las células epiteliales
alveolares de tipo I. Reproducido de Forbes et al., 2005.
En condiciones normales (no inflamatorias) los MA se mantienen en un
estado de reposo para evitar una excesiva inflamación, produciendo pocas
citoquinas inflamatorias (Holt 1978). El objetivo principal es evitar daños en las
células epiteliales de tipo I y tipo II en respuesta a antígenos inofensivos. Además,
los MA suprimen activamente la inducción de la inmunidad adaptativa a través de
sus efectos sobre células dendríticas alveolares e intersticiales y células T. Estudios
previos han demostrado que la eliminación in vivo de los MA mediante liposomas
cargados de clodronato da lugar a reacciones inflamatorias fuertes en respuesta a

Introducción General
17
partículas inofensivas y antígenos solubles (Thepen et al. 1989). Los MA se
adhieren estrechamente a las células epiteliales alveolares en la pared alveolar y
están separados por una distancia de sólo 0.2-0.5 mm de las células dendríticas
intersticiales. En ratones donde se han eliminado los MA, las células dendríticas
muestran un aumento en su capacidad presentadora de antígenos (Holt et al.
1993).
En un proceso infeccioso los MA se activan aumentando su capacidad
fagocítica y secretando citoquinas (TNF-α e IL-6) y quemoquinas (IL-8) que
promueven el reclutamiento de neutrófilos (Franke-Ullmann et al. 1996; Marriott
& Dockrell 2007). En la fase de resolución de la inflamación, los MA son también
capaces de fagocitar células inflamatorias apoptóticas (Jennings et al. 2005) e
incluso pueden contribuir a la inducción de la apoptosis en estas células. In vitro,
los MA son capaces de inducir la apoptosis a través de Fas ligando y de otros
ligandos para receptores de muerte en células blanco susceptibles como
monocitos, linfocitos y neutrófilos (Badley et al. 1997; Dockrell et al. 2001).
6-3. Células dendríticas.
Las células dendríticas (CD) comparten el precursor común con MA, pero se
diferencian en su ubicación pulmonar (submucosa de la vía aérea), morfología,
expresión de receptores, procesamiento de antígenos y función. Las CD forman una
malla en la submucosa de la nasofaringe, tráquea, y el árbol bronquial similar al
que forman las células de Langerhans de la piel. Las CD son células presentadoras
de antígenos extremadamente eficientes pero son débiles fagocitos. Luego de
capturar el antígeno, las CD migran a los ganglios linfáticos regionales y presentan
péptidos antigénicos para producir una activación enérgica de las células T (a
distancia de la delicada superficie alveolar).

Introducción General
18
7. La interacción de los patógenos con la mucosa respiratoria.
7-1. Reconocimiento de patógenos por las células de las vías
respiratorias.
El sistema inmune de los mamíferos está compuesto por dos ramas, el
sistema inmune innato y el sistema inmune adaptativo, que trabajan en conjunto
para proporcionar resistencia a la infección. La respuesta inmune innata es la
primera línea de defensa del huésped y es responsable del reconocimiento
inmediato de los agentes agresores y de controlar la invasión microbiana
(Medzhitov & Janeway 1997). Los componentes celulares del sistema inmune
innato son neutrófilos, macrófagos, células asesinas naturales (natural killers o
NK), basófilos, mastocitos, eosinófilos y otros.
En el tracto respiratorio, el epitelio que reviste las vías respiratorias es el
primer punto de contacto para las sustancias inhaladas tales como contaminantes
del medio ambiente, el humo del cigarrillo, alérgenos en el aire, y los
microorganismos (Diamond et al. 2000). Aunque el tracto respiratorio superior
está colonizado por bacterias, el tracto respiratorio inferior suele ser un ambiente
estéril. En los últimos años ha quedado claro que las células epiteliales de las vías
respiratorias no sólo proporcionan una barrera pasiva, sino que también
contribuyen activamente al sistema inmune innato (Diamond et al. 2000; Holgate
et al. 2000). El epitelio de las vías respiratorias reconoce a los patógenos y
responde iniciando una respuesta inflamatoria. Esta respuesta consiste en un
aumento en la liberación de, por ejemplo, péptidos antimicrobianos en el lumen de
las vías respiratorias, y la liberación de citoquinas en la submucosa, que inician una
reacción inflamatoria (Bals et al. 2004). Dicha reacción inflamatoria incluye el
reclutamiento de fagocitos, que sirven para eliminar los microorganismos que no
fueron eliminados por el mismo epitelio, y de células dendríticas y linfocitos que
pueden ayudar a montar una respuesta inmune adaptativa. Por lo tanto, los
mecanismos desencadenados por el epitelio de las vías respiratorias luego de
reconocer a los agentes patógenos son esenciales para montar una respuesta
protectora por el sistema inmune(Bals et al. 2004; Diamond et al. 2000).

Introducción General
19
La respuesta inmune innata depende de receptores codificados por una
línea germinal evolutivamente antigua llamados receptores de reconocimiento de
patrones (pattern-recognition receptors, PRRs), los cuales reconocen estructuras
antigénicas altamente conservadas (Akira et al. 2006; Lee & Kim 2007; Trinchieri
& Sher 2007). Este sistema de reconocimiento codificado genéticamente le permite
al huésped reconocer rápidamente una amplia variedad de patógenos. Los PRRs
reconocen componentes microbianos conocidos como patrones moleculares
asociados a patógenos (PAMPs), los cuales son moléculas esenciales para la
supervivencia de los microorganismos y son relativamente invariantes. Las células
del sistema inmune innato, incluyendo macrófagos y células dendríticas, utilizan
los PRRs para reconocer los PAMPs presentes en los microorganismos.
La inmunidad innata incluye al menos 4 familias de PRRs: receptores tipo
Toll (TLRs), receptores tipo RIG (RLRs), receptores tipo NOD (NLRs) y la familia de
receptores HIN200. Mientras que los TLRs se localizan en membrana plasmática y
endosomal y se especializan en su mayoría en la vigilancia del espacio extracelular,
las otras 3 familias de receptores vigilan el entorno intracelular
Con el descubrimiento de los receptores de tipo toll (Toll-like receptor,
TLR) se produjo un gran avance en la comprensión de la capacidad del sistema
inmune innato para reconocer rápidamente los agentes patógenos. Hasta la fecha,
se han identificado 10 TLR humanos (Akira et al. 2006; Lee & Kim 2007; Trinchieri
& Sher 2007). Los TLR están presentes en casi todas las células del cuerpo y
pueden estar localizados en la membrana plasmática o en vesículas intracelulares
dependiendo del la naturaleza del patógeno que va a ser reconocido (Akira et al.
2006; Lee & Kim 2007; Trinchieri & Sher 2007) (Ilustración 3).
Las células epiteliales de las vías respiratorias expresan una variedad de
TLR que pueden ayudarles a montar una respuesta adecuada a la exposición
microbiana (Brightbill & Modlin 2000; Medzhitov & Janeway 2000). Se ha
demostrado la expresión de diversos TLR en células BEAS-2B, A549, 16HBE14o- y
otras líneas de epitelio respiratorio (Sha et al. 2004; Mayer & Dalpke 2007). La
activación de los TLR en las células epiteliales regula la expresión de una variedad

Introducción General
20
de genes, incluyendo los que codifican citoquinas, quemoquinas y péptidos
antimicrobianos.
Las células epiteliales de las vías respiratorias expresan principalmente
TLR2 a TLR6 (Sha et al. 2004; Homma et al. 2004). TLR4 es un componente central
en la respuesta de las células al LPS y es expresado por las células epiteliales
respiratorias (Sha et al. 2004; Mayer et al. 2007). TLR2 reconoce una amplia gama
de productos microbianos de las bacterias Gram-positivas, Gram-negativas y
hongos. TLR2 dimeriza con TLR1 ó TLR6, y reconoce componentes bacterianos
tales como lipoproteínas, peptidoglicano (PGN), ácido lipoteicoico y algunas
proteínas de la envoltura vírica, como las expresadas por el virus del sarampión y
el citomegalovirus humano. TLR5 reconoce flagelina, que es un componente de los
flagelos de las bacterias. TLR3 es esencial en la respuesta a la doble cadena (dc) de
ácido ribonucleico (ARN) que se produce durante las infecciones virales
(Alexopoulou et al. 2001). TLR3 ha sido implicado en la respuesta de las células
epiteliales respiratorias, por ejemplo a rinovirus, y ARNdc sintéticos (ácido
poliinosínico-policitidílico). Esta respuesta no sólo consiste en un incremento en la
expresión de las quemoquinas (Duits et al. 2003; Gern et al. 2003), sino también de
β-defensinas humanas (Duits et al., 2003). TLR9 media la respuesta al ácido
desoxirribonucleico (ADN) de las bacterias. Un estudio reciente mostró que TLR9
media la respuesta de líneas de células epiteliales bronquiales al CpG, resultando
en la expresión de la interleuquina-8 (IL-8) (Mayer et al., 2007). Los TLR están
estratégicamente posicionados en las células epiteliales y les permiten reconocer
organismos que se encuentran con frecuencia en la superficie de las mucosas.
Recientemente se han identificado receptores citosólicos de reconocimiento
de patógenos (Akira et al. 2006; Lee & Kim 2007; Trinchieri & Sher 2007). Entre
estos receptores se encuentran helicasas de ARN citoplasmáticas que se expresan
en las células epiteliales de las vías respiratorias y detectan ARNdc viral
independientemente de TLR3 (Ilustración 3) (Liu et al. 2007; Opitz et al. 2007).
Además, algunos miembros de la familia de proteínas NACHT-LRR (LNR), que
incluyen tanto las proteínas de dominio de oligomerización de unión a nucleótidos
(NOD) y proteínas que poseen dominios de unión a pirina (NALPs), reconocen
componentes microbianos en el citosol (Ilustración 3) (Akira et al. 2006; Lee &

Introducción General
21
Kim 2007; Fritz et al. 2006). NOD1 y NOD2, reconocen péptidos derivados del PGN,
ácido γ-D-glutamil-mesodiaminopimélico y muramil dipéptido respectivamente, y
se expresan en las células epiteliales de las vías respiratorias (Opitz et al. 2004).
Algunos NLRs y la proteína AIM2 de la familia HIN200 responden a la detección
de infecciones y señales de estrés ensamblando complejos proteicos citosólicos
denominados inflamasomas, cuya función central es la activación autocatalítica de
la caspasa-1 (Lamkanfi & Dixit 2012). La caspasa-1 es producida como zimógeno
inactivo. La consecuencia mejor caracterizada de la activación de caspasa-1 en los
inflamasomas es la secreción de las citoquinas proinflamatorias IL-1β e IL-18. En la
mayoría de los modelos experimentales, los ratones deficientes en caspasa-1 no
producen IL-1β e IL-18 maduras (Lamkanfi & Dixit 2012).
IL-1β es una potente citoquina proinflamatoria, que produce efectos locales
y sistémicos, teniendo como blancos a leucocitos, células endoteliales y epiteliales,
y fibroblastos, y células esplénicas, cerebrales y hepáticas. IL-1β mejora la
capacidad fagocítica de neutrófilos y monocitos e induce la expresión del receptor
C3b de complemento. Además incrementa la producción de especies reactivas del
oxígeno (ROS, por sus siglas en inglés) en neutrófilos ante el reconocimiento de
ligandos de origen microbiano. IL-1β induce además la secreción local de las
quemoquinas MIP-2 y KC, atractantes de neutrófilos, y estimula la expresión de las
moléculas de adhesión ICAM-1 y ELAM en células endoteliales. En el hígado
estimula la producción de proteínas del complemento y proteína C reactiva, y en el
bazo promueve la proliferación linfocitaria. Por su parte, IL-18 actúa en forma
colaborativa con IL-12 para inducir la producción de IFN-γ por células NK y
linfocitos T. También induce en neutrófilos la producción de ROS y la secreción de
IL-6 (Chen & Schroder 2013). Un aspecto notorio de ambas citoquinas es que
carecen de péptido señal y por lo tanto no son secretadas por la vía convencional
de Retículo Endoplásmico-Golgi. En cambio, su secreción, si bien no se conoce el
mecanismo, depende de la activación de inflamasomas y el clivaje autocatalítico de
la procaspasa-1 (Lamkanfi & Dixit 2012)

Introducción General
22
7-2. La respuesta del epitelio respiratorio a los patógenos
Como lo mencionáramos, las células epiteliales se encuentran en los sitios
de contacto con el ambiente externo por lo que son a menudo las primeras células
en interactuar con los patógenos microbianos. De hecho, la adherencia bacteriana a
las células epiteliales es un requisito previo para la colonización y posterior
infección (Beachey & Courtney 1989; Geme 1996), y las células epiteliales a través
de esta interacción tienen la oportunidad de reconocer y responder a los
patógenos. En el epitelio respiratorio esa respuesta incluye la secreción de
citoquinas proinflamatorias (IL-1, IL-6, TNF-α) y quemoquinas (IL-8, MCP-1,
eotaxina, CCL20) que dan inicio a una respuesta inflamatoria (Koyama et al. 2000;
Arnold et al. 1994; Gentry et al. 2007; Rittig et al. 2003; Mayer & Dalpke 2007).
Ilustración 3: Resumen de la influencia de las células epiteliales en la respuesta
inmune innata y adaptativa, así como en procesos anti-inflamatorios en las vías
respiratorias. La parte izquierda de la figura muestra los perfiles de expresión de PRRs en
el epitelio. Los TLR reconocen patógenos en la superficie celular del epitelio respiratorio.
Por el contrario, TLR3, NOD y helicasas a ARN reconocen patógenos intracelulares.
Después de la activación de los PRRs, las células epiteliales producen una amplia gama de
moléculas que aumentan la respuesta inmune innata y adaptativa, el reclutamiento de
células o median respuestas antiinflamatorias. Reproducido de Kato et al., 2007.

Introducción General
23
In vivo, el epitelio respiratorio se ubica muy próximo a las células
endoteliales de la microvasculatura pulmonar, por lo cual las quemoquinas
producidas por el epitelio alveolar infectado pueden inducir la migración de
fagocitos profesionales al sitio de infección. Este fenómeno de atracción de
fagocitos ha sido reproducido en modelos alveolares in vitro (Hurley et al. 2006;
Herold et al. 2006). Entre las quemoquinas secretadas por las células epiteliales en
respuesta a patógenos, IL-8 y MCP-1 juegan un rol muy importante en el
reclutamiento de fagocitos profesionales al sitio de infección. IL-8, y su homólogo
murino CXCL1 o también llamado KC, pertenece a la familia de las quemoquinas
CXC, es quimioatractante de neutrófilos y linfocitos T, y es secretada por una
amplia variedad de células (macrófagos, fibroblastos, células endoteliales, células
epiteliales, etc.) en respuesta a múltiples estímulos (Baggiolini et al. 1989; Bazzoni
et al. 1991; Van Damme et al. 1994). MCP-1 es un miembro de la familia de
quemoquinas CC que desempeña un papel fundamental en el reclutamiento y la
activación de los monocitos durante la inflamación aguda (Tangirala et al. 1997).
En los pulmones humanos MCP-1 se expresa en una variedad de tipos de celulares,
como ser macrófagos, células endoteliales, epiteliales bronquiales, y células del
músculo liso (Antoniades et al. 1992; Iyonaga et al. 1994; Car et al. 1994).
En un sistema biológico intacto, la comunicación entre las células epiteliales
y las células inmunes profesionales es necesaria para la inducción de una
respuesta innata y adaptativa eficiente. Debido a que las células epiteliales están
entre los primeros tipos celulares en interactuar con los patógenos inhalados,
conocer la contribución del epitelio respiratorio durante la invasión de las vías
aéreas por Brucella es crucial para dilucidar el proceso patogénico y puede además
contribuir al desarrollo de estrategias terapéuticas contra la brucelosis.
7-3. Mecanismos de infección a través de la mucosa respiratoria
A pesar de los mecanismos de defensa pulmonares, muchos patógenos
logran atravesar la barrera epitelial y alcanzar la circulación sistémica, con lo cual
se diseminan por el organismo. De hecho, no puede descartarse que la
transmigración de fagocitos hacia el sitio de infección contribuya en algunos casos

Introducción General
24
al pasaje de patógenos a través de la barrera epitelial alveolar, ya sea por alterar la
permeabilidad de la barrera epitelial o por exponer componentes de la membrana
celular basolateral que pueden servir de receptores para la adhesión de los
patógenos.
Distintos patógenos emplean mecanismos diferentes para atravesar la
barrera epitelial alveolar. El alvéolo contiene células epiteliales y macrófagos
alveolares, pero las primeras son mucho más numerosas (aproximadamente
30.000 versus 50). De esta manera, los patógenos podrían interactuar inicialmente
con las células epiteliales que lindan el espacio alveolar. De hecho, B. abortus y
numerosos patógenos (Mycobacterium tuberculosis, Burkholderia cepacia,
Francisella tularensis, Streptococcus pneumoniae) son capaces de invadir y/o
replicarse en células epiteliales alveolares o bronquiales humanas (Ferrero et al.
2009; Gentry et al. 2007; Birkness et al. 1999; Duff et al.). La invasión de células
epiteliales protege a la bacteria de la actividad bactericida de los macrófagos.
Además, algunos patógenos como Streptococcus pneumoniae, Mycobacterium
tuberculosis y Bacillus anthracis pueden pasar a través de estas células (pasaje
transcelular) hacia las células endoteliales adyacentes y así acceder a la circulación
sistémica (Russell et al. 2008; Birkness et al. 1999). Otros patógenos como
Pseudomonas aeruginosa y Burkholderia cepacia dañan la barrera epitelial al
interferir con la integridad de las uniones estrechas (tight junctions) y así logran
traslocar el epitelio (pasaje paracelular) (Duff et al., 2006).
Para algunas infecciones pulmonares causadas por patógenos capaces de
sobrevivir en macrófagos se ha postulado que los macrófagos alveolares infectados
podrían migrar hacia la circulación sistémica. Este mecanismo de traslocación de la
barrera epitelial suele denominarse “caballo de Troya”, ya que el patógeno viaja
recluido en una célula. Se han obtenido resultados que apoyan este mecanismo en
el caso de Chlamydia pneumoniae y B. anthracis. (Gieffers et al. 2004; Passalacqua &
Bergman 2006). Pero la demostración definitiva se obtuvo en un trabajo reciente
que demostró que los macrófagos alveolares murinos migran constitutivamente a
los ganglios drenantes y pueden transportar patógenos desde el pulmón a esos
ganglios incluso antes que las células dendríticas (Kirby et al. 2009).
Recientemente se ha demostrado que Brucella utiliza este mecanismo para

Introducción General
25
atravesar la barrera epitelial pulmonar (Archambaud et al. 2010). Sin embargo,
todavía resta evaluar si junto con éste mecanismo, Brucella es capaz de traslocar el
epitelio respiratorio por algún mecanismo de pasaje transcelular o paracelular.
Hipotesis de trabajo
Luego de ser inhalada B. abortus logra diseminarse sistémicamente y
persistir en el pulmón murino por tiempos prolongados, lo que indicaría que la
respuesta inmune montada por el pulmón es insuficiente para controlar la
infección. Previamente demostramos que B. abortus infecta in vitro macrófagos
alveolares y células epiteliales pulmonares en los que induce la producción de
citoquinas de forma leve y tardía.
La hipótesis de este trabajo es que B. abortus es capaz de infectar y persistir en
el pulmón murino, y de diseminarse sistémicamente desde allí, gracias a su
capacidad de evadir y/o modular la respuesta inmune innata y antimicrobiana
pulmonar, probablemente mediante mecanismos múltiples.

26
OBJETIVOS
El objetivo general de este trabajo de tesis consiste en evaluar la interacción
de Brucella con la mucosa respiratoria en las primeras etapas de la infección para
caracterizar la respuesta inmune innata desencadenada en el pulmón, así como
evaluar los mecanismos que utiliza la bacteria para evadir la vigilancia
inmunológica en el pulmón y diseminarse sistémicamente.
Se plantean los siguientes objetivos particulares:
Evaluar la evolución temporal de la respuesta inmune local (citoquinas,
defensinas, reclutamiento de células inmunes y características histológicas)
y la cinética de diseminación bacteriana en ratones luego de la infección
intratraqueal con B. abortus.
Evaluar la evolución temporal de la respuesta inmune pulmonar a la
infección con una mutante de B. abortus deficiente en las proteínas que
contienen dominios TIR (BtpA y BtpB).
Evaluar in vivo si B. abortus es capaz de modular la respuesta inmune
pulmonar frente a agonistas de TLRs y si ese efecto está mediado por las
proteínas Btp.
Evaluar in vitro la capacidad de los macrófagos y células epiteliales
alveolares murinas infectadas con B. abortus o con mutantes para las
proteínas BtpA y BtpB para modular la respuesta proinflamatoria inducida
por agonistas de TLRs.
Evaluar la capacidad de B. abortus de sobrevivir y de modular la respuesta
inflamatoria pulmonar en animales previamente estimulados con un
agonista TLR por vía intratraqueal.
Determinar si la infección por Brucella abortus, o el estímulo con
sobrenadantes de cultivo de macrófagos infectados, inducen en células
epiteliales alveolares y bronquiales la producción de β-defensinas y CCL20.
Evaluar el efecto antimicrobiano de CCL20, hBD2 y hBD3 sobre B. abortus.

27
Capitulo 1
Respuesta inmune a la infección
intratraqueal con B. abortus y efecto
modulador de las proteínas Btp

28
Introducción

Capítulo 1- Introducción
29
Los modelos animales se han usado ampliamente en el estudio de la Brucelosis.
Si bien cada especie de Brucella tiene su hospedador preferencial, el modelo más
ampliamente usado en investigación es el modelo murino porque aporta
información valiosa para el estudio de virulencia de cepas, ensayos de protección
vacunal o para evaluar la fisiopatología de la enfermedad (Grillá et al. 2012; Ibañez
et al. 2013; Gil-Ramírez et al. 2014).
Como se mencionó anteriormente, son varias las vías plausibles de iniciar una
infección exitosa. En la mayor parte de los ensayos realizados en ratones la vía
preferencial de infección es la intraperitoneal debido a su simplicidad y practicidad
de aplicación, y la reproducibilidad de sus resultados. Sin embargo, esta forma no
representa eficientemente una vía natural de infección.
Brucella posee alta infectividad por vía respiratoria, y hasta el momento se han
realizado pocos estudios acerca de los mecanismos que utiliza la bacteria durante
la infección inhalatoria. Existen tres modelos experimentales para la infección
respiratoria: a) administración del inóculo de forma intranasal, b) administración
intratraqueal, y c) inhalación de aerosoles de suspensiones bacterianas. Sin
embargo, en las infecciones intranasales o por exposición a aerosoles no puede
descartarse que la bacteria, además de contactar a la mucosa respiratoria, entre en
contacto con la mucosa oral. La vía de administración intratraqueal asegura
reproducibilidad del inóculo administrado y localización exclusiva en las vías
respiratorias bajas (Revelli DA, Boylan JA 2012).
Estudios previos con el modelo de infección por aerosoles de B. abortus
revelaron colonización y persistencia de la bacteria en los pulmones de los ratones
infectados hasta 8 semanas p.i. Además, la bacteria puede diseminarse desde
pulmón hacia órganos periféricos, siendo bazo e hígado los órganos blancos
preferenciales, transportada en parte por los macrófagos alveolares residentes en
la luz alveolar que son las células preferencialmente infectadas (Kahl-McDonagh et
al. 2007; Archambaud et al. 2010).
Sin embargo, todavía no se ha caracterizado la respuesta inmune temprana a la
infección inhalatoria con B. abortus, y no se han realizado estudios utilizando la
infección intratraqueal del patógeno. En este capítulo nos enfocaremos en

Capítulo 1- Introducción
30
caracterizar, en un modelo murino de infección intratraqueal con B. abortus, la
cinética de diseminación de la bacteria y la respuesta inmune pulmonar montada
en la primera semana de la infección.

31
Materiales y Métodos

Capítulo 1 – Materiales y Métodos
32
Animales
Se utilizaron ratones Balb/C machos de 8 semanas de edad. Los mismos fueron
albergados en instalaciones de bioseguridad tipo 3, en el Instituto de
Investigaciones Biotecnológicas – Universidad de San Martín.
En los ensayos con animales genéticamente deficientes (knockout, KO) se
utilizaron ratones de 7 a 10 semanas de edad de la cepa C57BL/6 (cepa salvaje,
wild type o wt) o ratones KO para los genes il1r o casp1/casp11 derivados de
C57BL/6. Los experimentos fueron realizados en la Universidad General de Minas
Gerais, Brasil, en el laboratorio del Dr. Sergio Costa Oliveira.
Inoculación intratraqueal
El protocolo de inoculación fue adaptado de Revelli et al. (2012). Los ratones
fueron anestesiados inicialmente con isoflurano en cámara hermética (100 µl/L) y
una vez adormecidos, se inyectaron intraperitonealmente con una mezcla de
ketamina y xilazina (100 mg/kg, 1 mg/kg respectivamente). Luego, se
posicionaron sobre una plancha acrílica inclinada 45°, se sujetaron por las
extremidades, y con una pinza roma se posicionó la lengua de forma tal que la
tráquea fuera fácilmente visible (Figura 1). La aplicación de una fuente de
iluminación externa a la altura de la garganta hizo posible visualizar las cuerdas
vocales, las que constituyeron el punto de referencia para la inoculación. Se
Figura 1. Ratón Balb/c siendo inoculado por vía intratraqueal con 20 µl de Azul de
Evans al 1%. Se utiliza una jeringa Hamilton para dispensar el inóculo.

Capítulo 1 – Materiales y Métodos
33
depositaron 20 µl del inóculo entre las cuerdas vocales mediante una jeringa
Hamilton acoplada a una aguja de extremo romo en ángulo de 45° y se dejó
reposar los animales hasta que se recuperaron de la anestesia.
En la puesta a punto de la técnica, para evaluar su eficacia se inocularon por vía
intratraqueal 20 µl de Azul de Evans al 1% en PBS. A las 2 y 24 h post-
administración se sacrificaron los animales y se colectaron los pulmones,
estómagos e intestinos. Se buscó presencia macroscópica de colorante en los
órganos.
Cepas de B. abortus y condiciones de crecimiento
Las cepas utilizadas en este estudio fueron:
B. abortus 2308 (wt)
B. abortus 2308 GFP (bacteria que expresa proteína fluorescente verde),
B. abortus 2308 btpAbtpB- (doble mutante para los genes btpA y btpB)
Las cepas genéticamente modificadas de B. abortus (GFP y btpAbtpB-) fueron
cedidas gentilmente por el Dr. Diego Comerci (Universidad de Gral. San Martín).
Todas las cepas se crecieron en caldo tripteína de soja a partir de colonias
obtenidas en agar tripteína de soja (TSA), siendo suplementado con kanamicina
(50 µg/ml) y gentamicina (5 µg/ml) para el crecimiento de B. abortus 2308
btpAbtpB-. Se estimó el número de unidades formadoras de colonia (UFC)
midiendo la densidad óptica del cultivo a 600 nm, y se determinó el inóculo exacto
por plaqueo de diluciones seriadas en TSA.
Infección intratraqueal
Los animales fueron infectados como se describió arriba con distintas dosis de
B. abortus 2308 (1x104 y 1x106 UFC por animal) o B. abortus btpAbtpB- (1x106 UFC
por animal) en 20 µl de buffer fosfato salino (PBS) estéril. Como control, otros
animales fueron inoculados con PBS. A distintos días post infección 4 o 5 animales
de cada lote fueron sacrificados y se obtuvieron los pulmones, bazo e hígado. Los
órganos de algunos animales por lote fueron homogeneizados para los ensayos de
recuento de UFC de B. abortus y la medición de citoquinas, mientras que los

Capítulo 1 – Materiales y Métodos
34
órganos de otros animales fueron empleados para histología o para el análisis de
las poblaciones celulares por citometría de flujo. El esquema de infección y
obtención de las muestras para los diferentes estudios se describe abajo:
A 1, 2, 3 y 7 días post-inoculación los animales infectados y del lote control se
sacrificaron por dislocación cervical y se les extrajeron asépticamente pulmones,
hígado y bazo. Los órganos se homogenizaron en 2 ml de PBS estéril empleando un
homogenizador Ultra-Turrax (IKA, Staufen, Alemania). Una parte se alicuotó para
posterior recuento de UFC por plaqueo (en TSA suplementado con bacitracina y
ácido nalidíxico para permitir el recuento diferencial de B. abortus), y el remanente
se clarificó por centrifugación a 8000 rpm durante 15 min, previa adición de
inhibidor de proteasa (c0mplete, Roche) y de gentamicina (100 µg/ml) para matar
las bacterias viables. Los sobrenadantes de los homogenatos se conservaron a -
70ºC para la posterior determinación de citoquinas y quemoquinas por ELISA.
Producción de citoquinas
En los homogenatos de los órganos se cuantificó por ELISAs comerciales la
concentración de IL-1β, TNF-α, IFN-γ, IL-10 (OptEIATM, BD Pharmingen, San Diego,
CA), MCP-1 y KC (DuoSet ELISA, R&D Systems, Minneapolis, USA) siguiendo las
instrucciones del fabricante.
Días p.i.
Infección 1 2 3 4 5 6 7
B. abortus
106 UFC
UFC
Citoquinas
UFC
Citoquinas
Histología
UFC
Citoquinas
UFC
Citoquinas
Histología
Citometría
B. abortus
104 UFC
UFC
Citoquinas
UFC
Citoquinas
UFC
Citoquinas
UFC
Citoquinas
Tabla 1. Esquema de infección y obtención de las muestras para los diferentes estudios

Capítulo 1 – Materiales y Métodos
35
Análisis histológico
Los pulmones obtenidos a los 2 y 7 días p.i. se perfundieron y se fijaron en
paraformaldehido (PFA) al 4% en PBS durante 24 h, luego se incluyeron en
parafina y se realizaron cortes de 5 µm que posteriormente se tiñeron con
hematoxilina-eosina (H&E). Los cortes fueron analizados por una médica patóloga,
la Dra. Silvia Vanzulli perteneciente al equipo profesional de la Academia Nacional
de Medicina.
Análisis de poblaciones celulares por citometría de flujo
Se caracterizaron por citometría de flujo las poblaciones de células inmunes
presentes en el pulmón durante la infección. Para ello los pulmones obtenidos a 7
días p.i. de los lotes de animales infectados y del lote control se disgregaron
mecánicamente a través de una malla de 100 µm y luego la suspensión celular
obtenida se filtró a través de una malla de 40 µm. De esta forma se obtuvo una
suspensión sin agregados celulares.
Las células obtenidas de cada pulmón se contabilizaron por recuento en
cámara de Neubauer y se analizaron por citometría de flujo empleando
anticuerpos contra CD11c, CD11b, CD3, CD4, CD8 (BD Pharmingen, San Diego, CA)
y F4/80 (AbD Serotec, Düsseldorf, Germany) de ratón conjugados a diferentes
fluorocromos. Para ello 1x106 células en 100 µl de BSA 0.1% en PBS se incubaron
con diluciones apropiadas de los respectivos anticuerpos durante 1 h a 4°C, a
continuación las células se lavaron 2 veces con PBS BSA 0.1% y se fijaron con PFA
al 4%. Los análisis de citometría de flujo se realizaron en un equipo PARTEC PAS
III, evaluando 50000 eventos para cada muestra. Las células dendríticas fueron
definidas como CD11c+ y CD11b+, los macrófagos alveolares como F4/80+ CD11c+,
los macrófagos reclutados como F4/80+ CD11b+, y los linfocitos T CD4+ y CD8+ por
marcación con CD3+ y CD4+ y CD8+ respectivamente.
Aislamiento de Macrófagos Alveolares
Los MA se obtuvieron a partir de muestras de lavado broncoalveolar (BAL, por
sus siglas en inglés) de ratones BALB/c. Los ratones fueron sacrificados por
sobredosis de ketamina y xilazina (300 mg/kg, 4 mg/kg respectivamente) y luego

Capítulo 1 – Materiales y Métodos
36
se expuso quirúrgicamente la tráquea. A través de una pequeña incisión en la
tráquea se instiló en los pulmones 1 ml de PBS con 1 mM de EDTA estéril
empleando una pipeta Pasteur de punta capilar. Los lavados se repitieron hasta
recuperar un volumen de 4 ml por animal. Finalizado el BAL, el líquido recuperado
se centrifugó a 200 xg durante 10 min a 4 °C y el sobrenadante se descartó. Las
células obtenidas se resuspendieron en medio de cultivo RPMI suplementado y se
colocaron sobre placa de cultivo de 48 pocillos a razón de 2x105 células/pocillo.
Luego de 2 h de incubación en estufa a 37ºC y 5% CO2, se descartó el sobrenadante
de cultivo y las células se lavaron varias veces con PBS estéril y se adicionó RPMI
suplementado a cada pocillo. Las células que quedan adheridas son MA.
Aislamiento de células epiteliales de pulmón (pneumocitos tipo II)
Las células pulmonares totales se obtuvieron siguiendo el protocolo
previamente descrito por Murphy y col. (1999). Para ello los ratones fueron
sacrificados por sobredosis de ketamina y xilazina como se describió arriba. La
unidad cardio-pulmonar fue extraída quirúrgicamente y los pulmones fueron
posteriormente perfundidos desde la aurícula con Solución A (0.130 M NaCl, 5.2
mM KCl, 10.6 mM HEPES, 2.6 mM Na2HPO4, 10 mM glucosa pH: 7.4). Los pulmones
fueron luego separados del corazón y cortados en pequeñas porciones con tijera.
Las porciones de tejido se lavaron tres veces con Solución A y luego se digirió dos
veces con tripsina (0.5 % en Solución A adicionada con 1.9 mM de CaCl2 y 1.29 mM
de MgSO4) a 37°C durante 15 min. La suspensión resultante fue colectada sobre
SFB con DNAsa (400U/10 ml), luego se filtraron secuencialmente por tamices de
100 µm y 40 µm, se lavaron y se dejaron adherir durante 90 min en placas de
plástico en DMEM/F12 suplementado con 5% SFB para eliminar las células más
adherentes. Las células no adheridas se colectaron y se centrifugaron en un
gradiente de Percoll discontinuo (1.089 g/l y 1.04 g/l). Las células colectadas en la
interfase de los gradientes de Percoll fueron lavadas dos veces con Solución A. Las
células se resuspendieron en DMEM/F12 5 % SFB y se sembraron en placa de 48
pocillos recubiertos previamente con colágeno bovino de tipo I. Las células
epiteliales se cultivaron hasta alcanzar confluencia antes de ser usadas en ensayos
de infección (4-5 días).

Capítulo 1 – Materiales y Métodos
37
Inmunofluorescencia para citoqueratina.
Para caracterizar el origen epitelial de las células aisladas de pulmón se
sembraron 1,5 x105 células sobre cubreobjetos de vidrio recubiertos con colágeno
en placa de 24 pocillos y se realizó una inmunofluorescencia para evidenciar la
expresión de citoqueratina 7 (ck-7). Para ello, las células fueron fijadas con PFA al
4% durante 30 min a temperatura ambiente, luego se lavaron secuencialmente con
glicina 0.2% en PBS y PBS Tritón X-100 0.2%. Después del bloqueo con PBS-BSA
2% durante 2 h, las células fueron incubadas con una dilución apropiada del
anticuerpo anti ck-7 (Clon: RCK105, BD Pharmigen) durante toda la noche a 4°C.
Las células se lavaron dos veces y se incubaron con el anticuerpo anti-IgG de ratón
conjugado a rodamina durante 1 h a Tº ambiente. Finalmente las células se
lavaron, y los núcleos celulares se tiñeron con DAPI (1µg/ml) durante 15 minutos y
se observaron en un microscopio de fluorescencia. Las células positivas para
citoqueratina confirman su origen epitelial.
Obtención de células dendríticas derivadas de médula ósea (BMDC):
Para la obtención de BMDC se extrajo medula ósea de fémures y tibias de
ratones Balb/C de 8 semanas de edad. Las células obtenidas se lavaron con RPMI
frío y se plaquearon a razón de 2x106 células en placa de 6 pocillos. Se cultivaron
en RPMI suplementado con 10% SFB Gibco, 0,01M de HEPES, 50 µM de 2-
βmercaptoetanol y 10% de sobrenadante proveniente de células J558 productoras
de GM-CSF. A los 9 días se cosecharon las células y como medida de pureza se
realizó un análisis de la expresión de CD11c.
Infección de cultivos primarios
Macrófagos Alveolares: Los MA fueron infectados con B. abortus WT, o con la
cepa mutante B. abortus btpAbtpB- a una MI de 100, durante 2 horas a 37 ºC.
Finalizada la incubación las células fueron lavadas repetidas veces e incubadas con
medio completo suplementado con 100 ug/ml de gentamicina para eliminar las
bacterias extracelulares. A 24 y 48 h luego de la adición de gentamicina, los
sobrenadantes de cultivo fueron cosechados y conservados a -70° C para luego
determinar TNF-α, IL-1, KC, IL-10 o IL-12 por ELISA, mientras que las células
fueron lavadas y lisadas con TRIzol para extraer RNA.

Capítulo 1 – Materiales y Métodos
38
Pneumocitos: Las células epiteliales respiratorias se infectaron como se
describió arriba para MA, con excepción de la MI utilizada (que en este caso fue
250 bacterias/célula). Los pneumocitos se infectaron con B. abortus WT o
btpAbtpB-. A 24 h p.i., los sobrenadantes de cultivo fueron cosechados y
conservados a -70° C para luego determinar KC por ELISA.
Células dendríticas derivadas de médula ósea: Las células dendríticas se
infectaron como se describió arriba para los macrófagos alveolares modificando la
MI, que fue de 30 bacterias/célula. Los sobrenadantes se colectaron 24 h p.i. para
determinar IL-1β, TNF-α e IL-12 por ELISA.
Inmunofluorescencia para B. abortus.
Para detectar las bacterias intracelulares se sembraron 1,5 x105 células sobre
cubreobjetos de vidrio recubiertos con colágeno en placa de 24 pocillos, se
infectaron con B. abortus wt o btpAbtpB- a una MI de y se realizó una
inmunofluorescencia para evidenciar la expresión de citoqueratina 7
bacterias/células. Luego de 2 h, las células se lavaron y se incubaron con medio de
cultivo suplementado con gentamicina (100µg/ml) para eliminar las bacterias
extracelulares (tiempo 0). A distintos tiempos p.i., las células fueron fijadas con
PFA al 4% durante 30 min a temperatura ambiente, luego se lavaron con PBS
Tritón X-100 0.2%. Después del bloqueo con PBS-BSA 2% durante 2 h, las células
fueron incubadas con un sobrenadante de cultivo del hibridoma BC68 (IgG1(κ) anti-
LPS de Brucella, (Goldbaum et al. 1992)) durante 2 h a temperatura ambiente. Las
células se lavaron dos veces y se incubaron con el anticuerpo anti-IgG de ratón
conjugado a rodamina durante 1 h a temperatura ambiente. Finalmente las células
se lavaron, y los núcleos celulares se tiñeron con DAPI (1µg/ml) durante 15
minutos y se observaron en un microscopio de fluorescencia. Las células positivas
para citoqueratina confirman su origen epitelial.
Retrotranscripción de ARN mensajero y Reacción en Cadena de la
Polimerasa (RT-PCR)
A partir de lisados en TRIzol se extrajo el ARN según recomendaciones del
fabricante y posteriormente se retrotranscribió utilizando la transcriptasa reversa
MMLV y oligodT como primers. Para evaluar los niveles de transcripción del ARN

Capítulo 1 – Materiales y Métodos
39
mensajero de ciertas citoquinas se utilizó PCR y los siguientes primers en
concentraciones de 0.1 mM:
il1b
Primer sentido: 5’ TGCCAGGTTTTGACAGTGATG 3’
Primer antisentido: 5’ AAGGTCCACGGGAAAGACAC 3’
tgb1
Primer sentido: 5’ ACTGGAGTTGTACGGCAGTG 3’
Primer antisentido: 5’ GGATCCACTTCCAACCCAGG 3’
tnf
Primer sentido: 5’ GGCAGGTCTACTTTGGAGTCATTGC 3’
Primer antisentido: 5’ ACATTCGAGGCTCCAGTGAATTCGG 3’
Los productos fueron analizados en geles de agarosa 2% teñidos con SYBRSafe
(Thermo Fisher). El análisis densitométrico se realizó utilizando el software
ImageJ, NIH.
Modulación de la respuesta inmune pulmonar a agonistas TLRs:
Ensayos in vivo
Se evaluó el rol de las proteínas Btp de Brucella en la respuesta inmune
pulmonar a agonistas de receptores TLR. Para ello ratones Balb/c machos fueron
separados en grupos de 5 animales e inoculados intratraquealmente con 106 UFC
de Brucella abortus (wild type; WT), B. abortus btpAbtpB- o PBS (control no
infectado). A 24 h post inoculación se administró intratraquealmente a todos los
grupos 5 µg de LPS de Escherichia coli (Cepa: 055:B5, Sigma). Los animales se
sacrificaron 24 horas más tarde y los pulmones fueron extraídos quirúrgicamente,
perfundidos y fijados para la posterior inclusión en parafina y tinción con
hematoxilina y eosina. Otros pulmones se homogenizaron y el sobrenadante de los

Capítulo 1 – Materiales y Métodos
40
homogenatos se conservó a -70 °C para la posterior cuantificación de citoquinas
por ELISA.
Ensayos in vitro
Los macrófagos alveolares se infectaron como se describió arriba. A las 24 h
post-infección las células infectadas y las células no infectadas (control) se
estimularon con 1 µg de LPS de E. coli. A 24 h post-estimulación se cosecharon los
sobrenadantes de cultivo para medir IL-1β, IL-12 y KC por ELISA.
Supervivencia de B. abortus en pulmones estimulados con agonista
de TLR.
Para evaluar la supervivencia de B. abortus en pulmones que han sido
previamente estimulados con un agonista TLR4 se inoculó intratraquealmente a
ratones Balb/c machos con LPS de E. coli (5 µg/ratón) o PBS (control). Luego de 24
h todos los animales fueron infectados con B. abortus WT o btpAbtpB- , generando
4 grupos experimentales de 5 animales cada uno:
PBS/WT (Administración de PBS y posterior infección con B. abortus WT) PBS/btpAbtpB- (Administración de PBS y posterior infección con B. abortus
btpAbtpB-) LPS/WT (Administración de LPS y posterior infección con B. abortus WT) LPS/btpAbtpB- (Administración de LPS y posterior infección con B. abortus
btpAbtpB-)
A las 24 h post-infección los animales fueron sacrificados y los pulmones
fueron extraídos quirúrgicamente y homogenizados. A partir de este homogenato
se realizó recuento de UFC (por plaqueo en TSA) y medición de citoquinas.
Análisis Estadístico
Los datos se analizaron usando análisis de varianzas (ANOVA). Las
comparaciones entre grupos de datos fueron analizadas con el Test de Tukey’s y
aquellas contra el grupo control fueron analizadas con el test de Dunnett’s. Todos
los análisis estadísticos fueron realizados con el software GraphPad (San Diego,
CA).

41
Resultados

Capítulo 1 - Resultados
42
Figura 2. La tinta se encuentra en los pulmones a 2 h post-administración. El
inóculo de 20 µl de Azul de Evans se distribuye entre ambos pulmones-derecho e
izquierdo-(panel A) y no se encuentran rastros de tinta en estomago o intestinos (panel
C). Panel B. A 24 h pos administración el inóculo se encuentra distribuido
homogéneamente en los pulmones.
Puesta a punto de la técnica de inoculación intratraqueal
Para evaluar la distribución del inóculo administrado con la técnica de
inoculación intratraqueal adoptada, se administró con dicha técnica Azul de Evans
y se buscó macroscópicamente la presencia del colorante en distintos órganos.
Como se muestra en la Figura 2 a) luego de 2 horas de inoculación el colorante
se distribuye con preferencia en el pulmón izquierdo, situación que revierte 24 h
después, donde se encuentra distribuido homogéneamente en ambos pulmones.
No se encontraron rastros de colorante en los estómagos e intestinos, lo que
sugiere que no hay fuga de inóculo hacia el tracto digestivo.
B. abortus infecta y se disemina desde pulmón.
Para evaluar la cinética de supervivencia de Brucella en el modelo murino de
infección intratraqueal, se infectaron ratones Balb/c machos de 8 semanas de edad
con 106 o 104 UFC/animal de B. abortus. A 2 h, 1, 2, 3 y 7 días p.i. los ratones fueron

Capítulo 1 - Resultados
43
sacrificados para la extracción del pulmón, el bazo y el hígado, seguido de posterior
homogenización y recuento de UFC por plaqueo en TSA. En la Figura 3 se muestra
la cinética de replicación y diseminación.
Para ambas condiciones de infección se observa que el inóculo se encuentra en
los pulmones a 2 hs pi, y que el pulmón permanece infectado hasta el último día
evaluado.
Inóculo de 106 UFC/animal
Cuando los ratones son infectados con 106 UFC/ratón, se observan UFC en
pulmón desde las 2 horas p.i. Estas UFC representa un 76% de inoculo instilado. A
los 2 días p.i. el recuento de UFC disminuye un 78 % con respecto a las 2 h p.i. Sin
embargo, a los 3 días p.i. el número de UFC recupera los valores iníciales y se
mantiene hasta el final del estudio.
En hígado y bazo la bacteria se detecta recién a 1 día p.i., siendo el recuento en
bazo aproximadamente 5 veces mayor al del hígado.
Paralelamente a los recuentos en los homogenatos de pulmón, las UFC en los
órganos periféricos disminuyen hasta día 3 p.i. infección, mientras que al día 7 p.i.
se observa un aumento significativo de aproximadamente 1000 veces en el
recuento de UFC.

Capítulo 1 - Resultados
44
Figura 3. Curva de cinética de diseminación de B. abortus 2308. Ratones machos fueron
infectados con 106 o 104 UFC/animal. A distintos tiempos p.i. los pulmones, bazos e hígados
fueron extraídos y homogenizados y las bacterias viables presentes diluidas serialmente y
cultivadas en placas de TSA con bacitracina y ácido nalidíxico.
0 2 4 6 8100
101
102
103
104
105
106
107
Pulmón
Bazo
Hígado
Días pi
UF
C/ó
rgan
o
0 2 4 6 8100
101
102
103
104
105
Pulmón
Bazo
Hígado
Días pi
UF
C/ó
rgan
o
Inóculo de 104 UFC/animal
El recuento de UFC en pulmón de los animales infectados con 104 UFC/ratón
se mantiene sin variaciones significativas desde 2 hs p.i. hasta 7 días p.i. (Figura 3,
panel izquierdo). A diferencia de la infección con mayor inóculo, los recuentos en
órganos periféricos se detectan recién a 7 días p.i., conservando la relación entre
UFC para hígado y bazo observada con 106 UFC/animal, siendo el bazo el órgano
donde se registra el mayor número de UFC.
La respuesta pulmonar de citoquinas y quemoquinas es leve y
retardada en el tiempo.
Para caracterizar la respuesta inmune a la infección durante la primera
semana, se midieron citoquinas y quemoquinas en los homogenatos de pulmón y
bazo de animales infectados. Se analizaron citoquinas pro-inflamatorias (IL-1β,
TNF-α, IL-12 e IFN-γ), quemoquinas (KC y MCP-1) y citoquinas antiinflamatorias
(IL-10)
Como se muestra en la Figura 4, la infección con inóculo de 106 UFC/animal de
B. abortus no produce cambios significativos en los niveles de las citoquinas
analizadas en homogenatos de pulmón durante los primeros dos días p.i. Hacia el
tercer día p.i. se observa un aumento significativo en IL-1β y en MCP-1 que alcanza

Capítulo 1 - Resultados
45
valores máximos a 7 días p.i. Mientras que los niveles de IL-12, KC e IFN-γ
aumentan significativamente a los 7 días p.i.
No se observan aumentos significativos en la producción de TNF-α en pulmón
en respuesta a la infección con 106 UFC/animal durante los tres primeros días de
infección, mientras que a 7 días los niveles de TNF-α aumentan significativamente
en los homogenatos de pulmones de los animales infectados
Al infectar con inóculo bajo no se registran cambios significativos en los niveles
medidos de citoquinas y quemoquinas de los homogenatos de pulmón totales para
ninguna de las citoquinas analizadas, salvo para IL-12, que aumenta
significativamente en los animales no infectados a 7 días p.i. (Figura 5).
B. abortus produce escasos cambios histológicos en pulmón.
Para evaluar los cambios inflamatorios a nivel histológico producidos por la
bacteria en pulmón, se analizaron pulmones fijados en PFA 4% y coloreados con
hematoxilina y eosina 2 y 7 días p.i.
Los pulmones de los ratones infectados muestran a 2 días p.i. pocos sitios de
extravasación hemática alveolar. No existen focos inflamatorios perivasculares o
peribronquiales, sin embargo se observa infiltrado intersticial de aspecto
mononuclear. A 7 días p.i. el infiltrado intersticial es de menor magnitud a pesar
que se observa un aumento en los focos de extravasación hemática (Figura 6).
Citometría de flujo
En base a los resultados obtenidos del análisis histológico, donde se evidenció
un aumento de la celularidad de aspecto monocítico, se quiso caracterizar las
distintas poblaciones en los pulmones a 7 días pi. Para ello, se infectaron animales
por vía intratraqueal con 106 UFC/animal de B. abortus y se realizó un análisis
citométrico de las suspensiones celulares obtenidas mediante disgregación
mecánica del órgano. Se realizó un recuento de células totales para cada
suspensión de cada grupo y el análisis citométrico se ajustó a la cantidad total de
células recuperadas por pulmón.

Capítulo 1 - Resultados
46
Como se observa en la Figura 7 panel izquierdo, se detectó un aumento en el
número de células recuperadas de los pulmones de ratones infectados con B.
abortus en comparación con los de ratones no infectados, aunque no logramos
atribuir la causa a ninguna población celular analizada.
En concordancia con el infiltrado mononuclear observado en los cortes de
hematoxilina-eosina, se observa una tendencia al aumento en los linfocitos T CD4+
y CD8+ en los pulmones de los animales infectados (figura 7, panel derecho), que
no llega a la significancia. Tampoco se observan cambios significativos para las
poblaciones de células dendríticas (CD11b+, CD11c+) o macrófagos alveolares
(F4/80+ CD11c+).

Capítulo 1 - Resultados
47
Figura 4.Panel de citoquinas correspondiente a la infección con 106 UFC/animal. IL-1β, TNF-α, KC, MCP-1, IL-10, IL-12 e IFN-γ se midieron por ELISA en homogenatos de pulmón entero. *p<0.05, ** p<0.01, respecto del grupo no infectado (NI) para cada tiempo
1 2 3 70
200
400
600
800
1000
I
NI
**
**
Días p.i.
IL-1
(p
g/m
l)
1 2 3 70
1000
2000
3000
4000NI
I
****
*
Días p.i.
TN
F (
pg
/ml)
1 2 3 70
100
200
300
400
500NI
I
*
Días p.i.
IL-1
2 (
pg
/ml)
1 2 3 70
100
200
300
400NI
I*
Días p.i.
IFN (
pg
/ml)
1 2 3 70
100
200
300
400NI
I**
Días p.i.
KC
(p
g/m
l)
1 2 3 70
200
400
600
800NI
I
*
**
Días p.i.
MC
P-1
(p
g/m
l)
1 2 3 70
100
200
300NI
I
* *
Días p.i.
IL-1
0 (
pg
/ml)

Capítulo 1 - Resultados
48
Figura 5 Panel de citoquinas correspondiente a la infección con 104 UFC/animal. IL-1β, TNF-α, KC, MCP-1, IL-10, IL-12 e IFNγ se midieron por ELISA en homogenatos de pulmón entero. *p<0.05, respecto del grupo no infectado (NI)
1 2 3 70
200
400
600
800NI
I
Días p.i.
IL-1
(p
g/m
l)
1 2 3 70
100
200
300
400
500NI
I
Días p.i.
KC
(p
g/m
l)
1 2 3 70
500
1000
1500
*
Días p.i.
IL-1
2 (
pg
/ml)
1 2 3 70
50
100
150NI
I
Días p.i.
TN
F (
pg
/ml)
1 2 3 70
500
1000
1500NI
I
Días p.i.
IL-1
0 (
pg
/ml)
1 2 3 70
20
40
60
NI
I
Días p.i.
MC
P-1
(p
g/m
l)
1 2 3 70
200
400
600
800
Días p.i.
IFN (
pg
/ml)

Capítulo 1 - Resultados
49
Figura 6.Histología de ratones no infectados e infectados con 106 UFC de B. abortus.
A 2 y 7 días post-infección los ratones fueron sacrificados y los pulmones fijados en PFA
4% para posterior procesamiento y tinción con hematoxilina y eosina.
Figura 7.Citometría de ratones no infectados e infectados con 106 UFC de B. abortus.
A 7 días post-infección los ratones fueron sacrificados y los pulmones disgregados
mecánicamente. Las células totales fueron contadas (panel izquierdo) y luego marcada
con las diluciones óptimas de los distintos anticuerpos. Se analizaron: Células
dendríticas (CD11b+ CD11c+), Macrófagos alveolares (F4/80+ CD11c+), Macrófagos
reclutados (F4/80+ CD11b+), Linfocitos T CD4+ (CD3+ CD4+) Linfocitos T CD8+ (CD3+
CD8+)(panel derecho)
Control Infectados0
2
4
6
8**
N°
cé
lula
s/p
ulm
ón
(x10
6)
0
1
2
3
4
No infectados
Infectados
Dendríticas Macrófagosalveolares
Linfocitos T
CD4+
Linfocitos T
CD8+
N°
celu
las/p
ulm
ón
(x10
5)
2
7
No infectado Infectado
Día
s p
ost
infe
cció
n
infe
cció
n

Capítulo 1 - Resultados
50
Cinética de supervivencia de B. abortus btpAbtpB- luego de la
infección intratraqueal
Como lo mencionáramos, Brucella expresa las proteínas BtpA y BtpB que
contienen dominios TIR y se secretan al citosol de las células infectadas. Se ha
demostrado que estas proteínas afectan la señalización de TLR2, TLR4 y TLR5,
provocando reducción en la secreción de la IL-12 y TNF-α por células dendríticas
(Salcedo et al 2008 y 2013). Sin embargo, su papel en la modulación de la
respuesta inmune pulmonar a Brucella es aún desconocido, como así también su
rol en la diseminación sistémica de la bacteria desde el pulmón. Los resultados
descriptos arriba demuestran que los pulmones de los ratones infectados con B.
abortus producen una respuesta proinflamatoria retardada y limitada, con muy
pocos signos de inflamación en el análisis histológico. En base a estos antecedentes
nos propusimos evaluar la relevancia de las proteínas Btp en el modelo de
infección intratraqueal murina. Para ello se infectaron ratones con 106 UFC de una
cepa mutante de B. abortus que carece de estas proteínas, denominada B. abortus
btpAbtpB- (en adelante btpAbtpB-), y se colectaron pulmón, bazo e hígado que
posteriormente fueron homogenizados y plaqueados en TSA para recuento de UFC.
El recuento bacteriano en el pulmón a 2 h p.i. correspondió al 33 % del inóculo
inicial. Como se muestra en la Figura 8, los valores de UFC se mantuvieron casi sin
alteraciones desde las 2 h p.i hasta finalizado el estudio. La carga bacteriana viable
a 2 h p.i. fue menor respecto del ensayo realizado con la cepa WT, donde al mismo
tiempo p.i. se obtenía un 77 % del inóculo (1,65 0,52) x105 UFC/pulmón. La
mutante btpAbtpB- persistió en pulmón hasta el final del ensayo, con una tendencia
a la disminución de las UFC, aunque esta diferencia no fue estadísticamente
significativa. No se registró la disminución hacia el día 2 p.i. que se ve en la
infección con la cepa WT. Al día 2 p.i. se obtuvieron recuentos similares para
ambas infecciones, pero a diferencia de la infección con bacteria WT, la doble
mutante no es capaz de replicar y aumentar su recuento hacia el día 3 p.i. Las UFC
en bazo e hígado fueron detectables (límite de detección: 32 UFC/órgano) recién a
7 días p.i., a diferencia de la infección por la cepa WT, donde se detecta bacteria
viable en bazo e hígado 1 día p.i. y replicación a 7 días p.i.

Capítulo 1 - Resultados
51
Figura 8. Curva de cinética de diseminación de B. abortus wt y btpAbtpB-. Ratones
machos fueron infectados con 106 UFC/animal. A distintos tiempos p.i. los pulmones,
bazos e hígados fueron extraídos y homogenizados y las bacterias viables presentes
diluidas serialmente y cultivadas en placas de TSA y para B. abortus wt y btpAbtpB-
suplementadas con gentamicina (5 µg/ml) y kanamicina (50 µg/ml).
0 2 4 6 8100
101
102
103
104
105
106
107
Pulmón btpAbtpB-
Bazo btpAbtpB-
Hígado btpAbtpB-
Pulmón wt
Bazo wt
Hígado wt
Días p.i.
UF
C/ó
rgan
o
La infección con B. abortus btpAbtpB- produce signos moderados de
inflamación en el pulmón
Se evaluó si las proteínas Btp podrían estar involucradas en la regulación de la
respuesta inmune observada luego de la infección intratraqueal con la cepa salvaje.
Para ello se inocularon ratones Balb/c con 106 UFC de btpAbtpB-. Los pulmones se
colectaron a 2 y 7 días p.i., se incluyeron en parafina y se tiñeron con H&E.
En los cortes correspondientes a 2 días p.i. se observaron focos tipo
bronconeumónico, compuestos de un infiltrado celular con predominio
mononuclear, mientras que en la luz alveolar se observaron hematíes y fibrina
(Figura 9). A los 7 días p.i. se observaron los mismos signos, pero con presencia de
hemorragia perivascular. Comparando con la histología de la infección con la cepa
WT (Figura 6) se ve un mayor infiltrado celular de características diferentes. Estos
resultados indican que en ausencia de las proteínas Btp B. abortus provoca un
mayor infiltrado de células en el parénquima pulmonar de los ratones infectados.

Capítulo 1 - Resultados
52
Figura 9 Histología de ratones infectados con B. abortus btpAbtpB- (106 UFC), 2
y 7 días p.i.
Respuesta inmune sistémica a la infección intratraqueal con B.
abortus btpAbtpB-
La Figura 8 muestra diferencias en las cargas bacterianas en bazo e hígado
luego de la infección intratraqueal con la cepa WT y la doble mutante btpAbtpB-.
Esta diferencias podrían deberse a una repuesta inmune diferencial en los órganos
periféricos para ambas cepas. Para probar esta hipótesis evaluamos la respuesta
inmune en bazo e hígado de los ratones infectados con dichas cepas. Para ello se
cuantificaron a los mismos tiempos p.i. las citoquinas IL-1β, TNF-α, IL-12 y la
quemoquina KC en homogenatos de bazos e hígados de los animales infectados con
la cepa WT y con la doble mutante.
Los resultados muestran un aumento en la secreción de IL-1β, TNF-α e IL-12 a
las 24 h p.i. en bazos de animales infectados con la doble mutante respecto del

Capítulo 1 - Resultados
53
grupo control inoculado con PBS (Figura 10). El aumento de IL-12 es moderado,
pero significativamente distinto. Este aumento respecto de los controles no se
registra en los bazos de los animales infectados con la cepa WT. La infección con
ambas cepas produce un aumento de KC a 24 hs.
No se registran diferencias significativas para IL-1β, TNF-α o KC a los días 2, 3 o
7 p.i. para ninguna de las dos cepas. Existe un aumento significativo de IL-12 a día
7 p.i. en bazos de animales infectados con la cepa WT que no se detecta en los
bazos de los ratones infectados con la btpAbtpB-. No se detectaron aumentos en la
secreción de ninguna de las citoquinas estudiadas en los homogenatos de hígados
de los animales infectados para ninguna de las cepas respecto de los controles sin
infectar (datos no mostrados)
B. abortus btpAbtpB- modula la respuesta en citoquinas de los MA
Los resultados histológicos obtenidos de la infección in vivo con btpAbtpB-
sugieren que las proteínas Btp están involucradas en la modulación de la respuesta
inmune innata en el pulmón. Teniendo en cuenta que los macrófagos alveolares
(MA) constituyen la primera línea de defensa frente a la entrada de un potencial
patógeno, evaluamos si las proteínas Btp son capaces de modificar la respuesta
inmune de estas células. Para ello, se estudió la secreción de quemoquinas y
citoquinas por MA infectados in vitro con cepa WT y con la cepa btpAbtpB-, así
como la capacidad de estas bacterias para replicar intracelularmente en dichas
células.
Se evaluó la expresión de citoquinas proinflamatorias y antiinflamatorias por
MA infectados con las cepas WT y btpAbtpB-. Para ello se midieron los niveles de
transcriptos de mRNA para IL-1β y TGF-β en MA a 24 h p.i. mediante PCR y
posterior densitometría de geles de agarosa.
Como se muestra en la Figura 11, la transcripción del ARNm de IL-1β fue mayor
en MA infectados con btpAbtpB- que en aquellos infectados con la cepa WT
(p<0.05). A su vez, ambas infecciones resultaron en un mayor nivel de
transcripción respecto de los MA no infectados.

Capítulo 1 - Resultados
54
Figura 10. Panel de citoquinas correspondiente a la infección con 106 UFC/animal de B. abortus WT (izquierda) o B. abortus btpAbtpB-. IL-1β, TNF-α, KC, e IL-12 se midieron por ELISA en homogenatos de bazos enteros. *p<0.05, ** p<0.01,*** p<0.001 respecto del grupo no infectado (NI) Para cada tiempo
1 2 3 70
100
200
300NI
I
Días p.i.
IL-1
(p
g/m
l)
1 2 3 70
200
400
600
800NI
I
*
Días p.i.
IL-1
(p
g/m
l)
1 2 3 70
50
100
150NI
I
Días p.i.
TN
F-
(p
g/m
l)
1 2 3 70
100
200
300NI
I
*
Días p.i.
TN
F-
(p
g/m
l)
1 2 3 70
20
40
60NI
I
**
Días p.i.
KC
(p
g/m
l)
1 2 3 70
50
100
150NI
I*
Días p.i.
KC
(p
g/m
l)
1 2 3 70
200
400
600
800
NI
I
**
*
Días p.i.
IL-1
2 (
pg
/ml)
1 2 3 70
100
200
300
400
500NI
I
****
Días p.i.
IL-1
2 (
pg
/ml)

Capítulo 1 - Resultados
55
Figura 11. Expresión de il1b y tgfb en macrófagos alveolares infectados con B. abortus WT y
mutante btpAbtpB-. Las células fueron infectadas con 100 bacterias/célula y luego de 24 h
lisadas, y su ARNm retrotranscripto y medido por PCR semicuantitativa. En el panel superior se
muestra una imagen representativa de las bandas obtenidas en gel de agarosa. En el panel
inferior el resultado de la semicuantificación por densitometría realizada con el programa
ImageJ. *p<0.05, respecto de no infectado (NI)
La transcripción de TGF-β se encuentra aumentada en ambas infecciones, sin
evidenciar diferencias entre la inducción por la infección con la cepa WT o con
btpAbtpB-. Al evaluar la producción de ARN mensajero para TNF-α no se
detectaron diferencias significativas en ninguna de las tres condiciones ensayadas
(no mostrado).
NI wt btpAbtpB-0.0
0.2
0.4
0.6
0.8
1.0
**
p<0.05
IL1B
/GA
DP
H
NI wt btpAbtpB-
0.0
0.2
0.4
0.6
0.8
1.0
* *
TG
FB
/GA
DP
H
ns
Se midieron también mediante ELISA comerciales los niveles de secreción de
IL-1β, IFN-γ y KC a 24 y 48 hs p.i. en los sobrenadantes de macrófagos alveolares
infectados. La infecciones con las cepas WT y btpAbtpB- aumentaron
significativamente la secreción de todas las citoquinas ensayadas cuando las
comparamos con la secreción basal de las células sin infectar tanto a 24 h como a
48 h p.i. (Figura 12). Sin embargo, la infección con btpAbtpB- produjo una
secreción significativamente más alta que la infección con la cepa WT a las 24 h p.i.
para todas las citoquinas ensayadas. Además, a las 48 h p.i. es mayor la secreción

Capítulo 1 - Resultados
56
Figura 12. Secreción de IL1β, IFN-γ y KC en macrófagos alveolares. Las células fueron infectadas
con 100 bacterias/célula y luego de 24 h o 48 h el sobrenadante fue colectado y las citoquinas
medidas por ELISA. *p<0.05, ** p<0.01,*** p<0.001 respecto de no infectado (NI)
NI btpAbtpB- wt NI btpAbtpB- wt0
200
400
600
800
24h 48h
***
p<0.01
**
mIF
N (
pg
/ml)
NI btpAbtpB- wt NI btpAbtpB- wt0
500
1000
1500
24h 48h
***
***
* *
p<0.05
IL-1
(p
g/m
l)
p<0.05
NI btpAbtpB- wt NI btpAbtpB- wt0
1000
2000
3000
4000
5000
6000
30000
40000
P<0.01
***
***
ns
24 h 48 h
*** ***
KC
(p
g/m
l)

Capítulo 1 - Resultados
57
Figura 13. Curva de replicación de B. abortus wt y btpAbtpB- en macrófagos alveolares
primarios murinos. Las células fueron infectadas con 100 bacterias/célula y luego de 24 h las
células fueron lisadas selectivamente y las bacterias intracelulares viables cuantificadas
mediante el crecimiento en TSA.
de IL-1β por los MA infectados con btpAbtpB- que por los infectados con la cepa
WT. La secreción de IFN-γ no resulta significativamente diferente para ambas
infecciones, pero muestra una tendencia cercana a la significancia.
Estas diferencias observadas entre las cepas respecto de la inducción de
citoquinas no se deben a diferencias en la capacidad de replicación ni a la carga de
bacterias intracelulares dado que como se ve en la Figura 13, ambas bacterias son
capaces de ingresar a las células en la misma magnitud, y replican eficientemente,
sin registrarse cambios significativos en la tasa de incremento.
0 1 2104
105
106
107
108
wt
btpAbtpB-
Días p.i.
UF
C/m
l

Capítulo 1 - Resultados
58
Supervivencia de B. abortus luego de la infección intratraqueal en
ratones KO para caspasa-1 e IL-1R
Los resultados presentados anteriormente demuestran que la expresión de IL-
1β es modulada por las proteínas Btp durante la infección por B. abortus. Cabe
preguntarse entonces cuál es la importancia de esa modulación para el éxito de la
infección pulmonar por esta bacteria. Diversos estudios en modelos murinos de
infección respiratoria por otros patógenos indican que la respuesta local de IL-1β
es fundamental para el control efectivo de dichas infecciones en el pulmón. Por ello
nos propusimos determinar si IL-1β tienen un rol protectivo en el pulmón durante
la infección inhalatoria in vivo por B. abortus. Para ello se infectaron ratones
C57BL/6 (cepa WT) o genéticamente deficientes (KO) para caspasa-1 (enzima
indispensable para el clivaje del precursor pro-IL-1β) o para el receptor de IL-1
(IL-R) como se describió en M&M. A distintos tiempos p.i., los ratones fueron
sacrificados, sus pulmones colectados y homogenizados y este homogenato fue
plaqueado para obtener recuento de UFC en pulmones enteros.
Como muestra la Figura 14, el recuento de UFC en los pulmones de los ratones
KO para IL-1R se incrementa al día 2 y 7 p.i., mientras que el recuento de UFC en
pulmones de ratones WT disminuye a 2 días p.i. y recupera aproximadamente el
mismo número de UFC hacia día 7 p.i. Los recuentos pulmonares para los ratones
KO para caspasa-1/11 disminuyen también hacía día 2 p.i., pero aumentan
significativamente hacia día 7 respecto de los ratones WT. Estos resultados
sugieren que IL-1 tiene un rol protectivo en el control de la infección intratraqueal
por B. abortus

Capítulo 1 - Resultados
59
Figura 14. Curva de cinética de diseminación de B. abortus wt en ratones KO para
caspasa1/11 e IL-1R. Ratones machos fueron infectados con 106 UFC/animal. A distintos
tiempos p.i. los pulmones, bazos e hígados fueron extraídos y homogenizados y las
bacterias viables presentes diluidas serialmente y cultivadas en placas de TSA. * marca
p<0.05 respecto del grupo wt para cada tiempo ensayado.
0 2 4 6 8103
104
105
106
107
108
KO caspasa-1/11
KO IL-1R
wt
*
*
Días p.i.
UF
C/p
ulm
ón
KO cas1/11 KO IL-1R wt100
101
102
103
104
105 *
UF
C/h
ígad
o
KO cas1/11 KO IL-1R wt100
101
102
103
104
105
ns
UF
C/b
azo
7 días p.i.
B. abortus WT y btpAbtpB- infectan y replican en pneumocitos tipo
II murinos
Trabajos previos del grupo han demostrado la capacidad de B. abortus de
invadir y replicar en líneas de células epiteliales respiratorias humanas, así como
también la respuesta de estos a la infección (Ferrero et al. 2009; Ferrero et al.
2010). Sin embargo hasta el momento no se ha descripto la interacción de Brucella
abortus con las células alveolares del epitelio respiratorio murino. Considerando la
importancia de dichas células epiteliales en la fisiología pulmonar, y en vista de su
capacidad de participar en la respuesta inmune innata decidimos evaluar el efecto
de la infección sobre un cultivo primario de pneumocitos tipo II murinos.
Se evaluó mediante microscopía de fluorescencia la capacidad de Brucella de
infectar pneumocitos murinos. Para ello se infectó un cultivo primario de
pneumocitos con B. abortus expresando GFP y a las 24 h p.i. se fijó y se realizó una
marcación con un anticuerpo contra citoqueratina-7.

Capítulo 1 - Resultados
60
Figura 15 Infección de pneumocitos tipo II primarios murinos por Brucella abortus
GFP. Panel izquierdo. Pneumoctos tipo II marcados con citoqueratina-7 (Ck-7 rojo) y
DAPI (azul). Panel derecho. Pneumocitos tipo II marcados con citoqueratina-7 (rojo) y
DAPI (azul) e infectados con B. abortus GFP a una MI de 250 (verde).
Figura 16 Infección de pneumocitos tipo II primarios murinos por Brucella abortus
wt o btpAbtpB-. Las células se infectaron con una MI de 250 bacterias/células de Brucella
abortus wt o btpAbtpB-. Luego de 2 h p.i. las células fueron lisadas selectivamente y las
bacterias intracelulares viables cuantificadas mediante el crecimiento en TSA.
Como muestra la Figura 15, B. abortus puede invadir células positivas para
marcación de citoqueratina 7, la bacteria se observa en el citoplasma y en posición
perinuclear como se describió previamente para otras infecciones por Brucella en
líneas celulares epiteliales (Ferrero et al. 2009).
De manera similar se comparó la capacidad de infección y replicación intracelular
de la cepa B. abortus WT y la mutante btpAbtpB- en neumocitos primarios. No se
observan diferencias en el número de bacterias intracelulares para las cepas a 2 h
p.i. (Figura16). En la Figura 17, se observa un aumento leve en la carga intracelular
de ambas cepas entre las 2 h y las 48 h p.i.
wt btpAbtpB-0
20000
40000
60000ns
UF
C/m
l
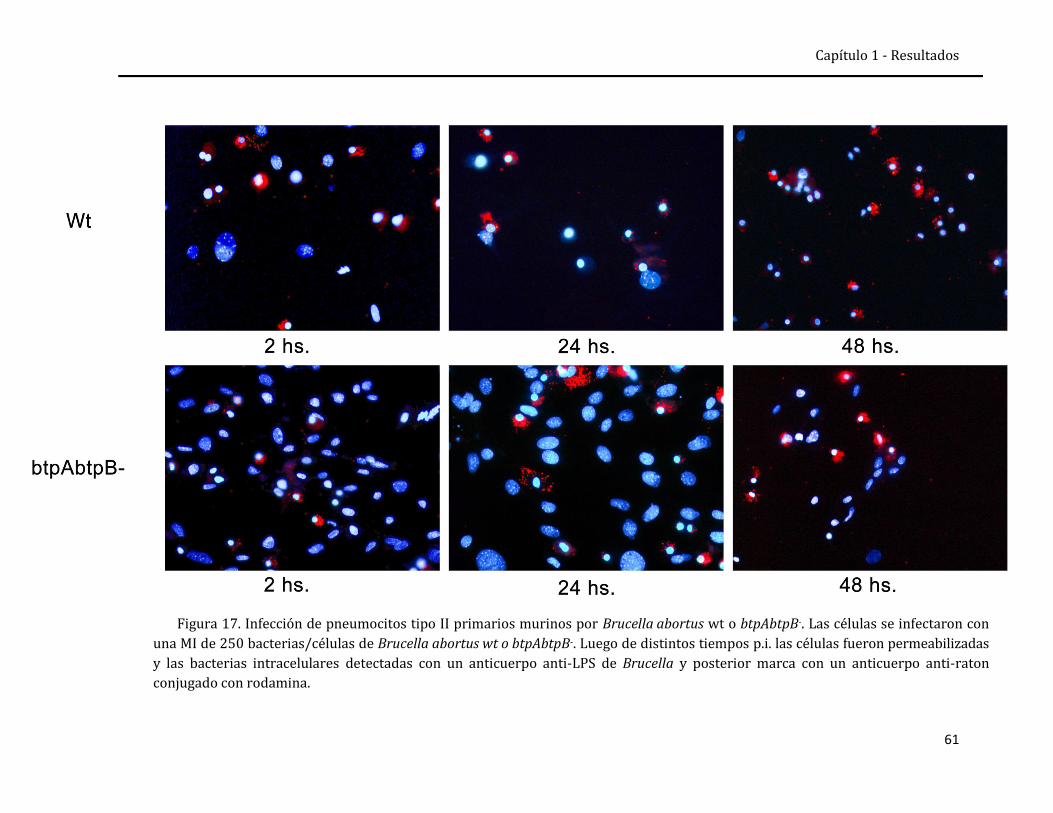
Capítulo 1 - Resultados
61
Figura 17. Infección de pneumocitos tipo II primarios murinos por Brucella abortus wt o btpAbtpB-. Las células se infectaron con
una MI de 250 bacterias/células de Brucella abortus wt o btpAbtpB-. Luego de distintos tiempos p.i. las células fueron permeabilizadas
y las bacterias intracelulares detectadas con un anticuerpo anti-LPS de Brucella y posterior marca con un anticuerpo anti-raton
conjugado con rodamina.
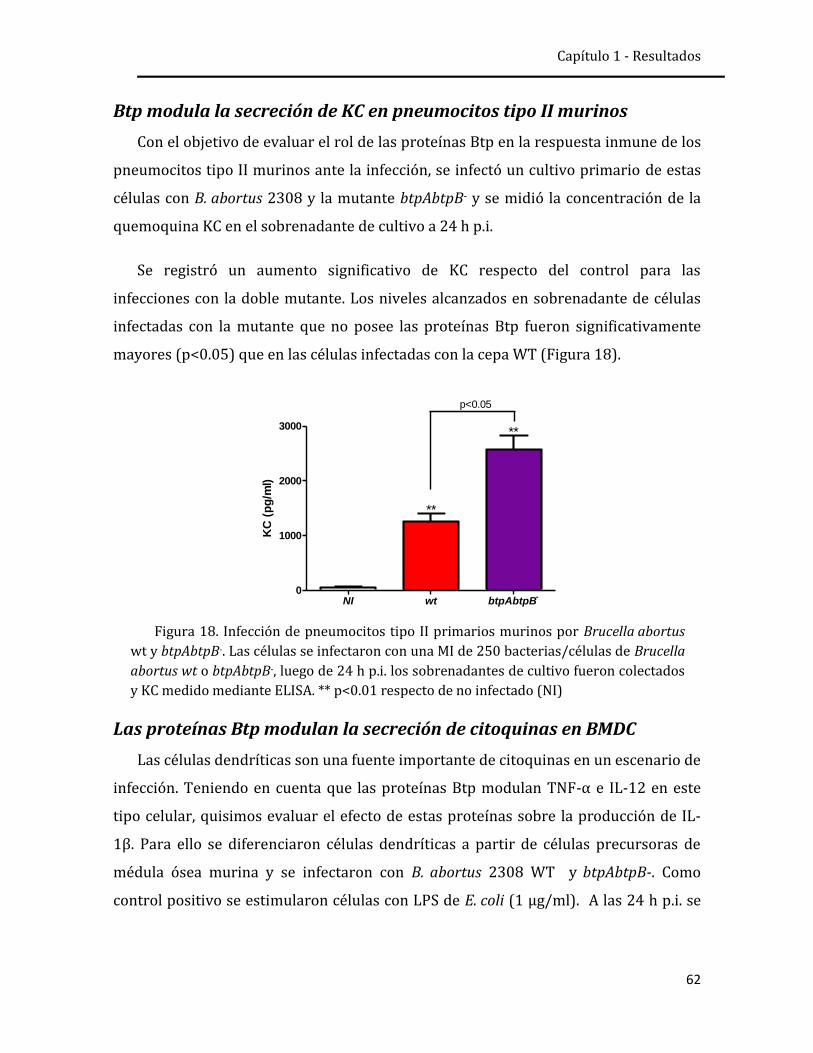
Capítulo 1 - Resultados
62
Figura 18. Infección de pneumocitos tipo II primarios murinos por Brucella abortus
wt y btpAbtpB-. Las células se infectaron con una MI de 250 bacterias/células de Brucella
abortus wt o btpAbtpB-, luego de 24 h p.i. los sobrenadantes de cultivo fueron colectados
y KC medido mediante ELISA. ** p<0.01 respecto de no infectado (NI)
Btp modula la secreción de KC en pneumocitos tipo II murinos
Con el objetivo de evaluar el rol de las proteínas Btp en la respuesta inmune de los
pneumocitos tipo II murinos ante la infección, se infectó un cultivo primario de estas
células con B. abortus 2308 y la mutante btpAbtpB- y se midió la concentración de la
quemoquina KC en el sobrenadante de cultivo a 24 h p.i.
Se registró un aumento significativo de KC respecto del control para las
infecciones con la doble mutante. Los niveles alcanzados en sobrenadante de células
infectadas con la mutante que no posee las proteínas Btp fueron significativamente
mayores (p<0.05) que en las células infectadas con la cepa WT (Figura 18).
NI wt btpAbtpB-
0
1000
2000
3000
**
**
p<0.05
KC
(p
g/m
l)
Las proteínas Btp modulan la secreción de citoquinas en BMDC
Las células dendríticas son una fuente importante de citoquinas en un escenario de
infección. Teniendo en cuenta que las proteínas Btp modulan TNF-α e IL-12 en este
tipo celular, quisimos evaluar el efecto de estas proteínas sobre la producción de IL-
1β. Para ello se diferenciaron células dendríticas a partir de células precursoras de
médula ósea murina y se infectaron con B. abortus 2308 WT y btpAbtpB-. Como
control positivo se estimularon células con LPS de E. coli (1 µg/ml). A las 24 h p.i. se

Capítulo 1 - Resultados
63
Figura 19 Infección de BMDC murinas por Brucella abortus wt btpAbtpB-. Las células se
infectaron con una MI de 30 bacterias/células de Brucella abortus wt o btpAbtpB-. Luego de
24 h p.i. los sobrenadantes de cultivo fueron colectados y las citoquinas medidas mediante
ELISA. *p<0.05, ** p<0.01,*** p<0.001 respecto de no infectado (NI)
colectó el sobrenadante y se midieron las citoquinas IL1β, TNF-α e IL-12. Los
resultados se muestran en la Figura 19.
NI btpAbtpB- wt LPS0
500
1000
1500
20006000
8000
10000
12000
* *
***
p<0.001
IL-1
(p
g/m
l)
NI btpAbtpB- wt LPS0
1000
2000
3000
4000
8000
10000
12000
***
*
***
p<0.001
TN
F-
(p
g/m
l)
NI btpAbtpB- wt LPS0
5000
10000
15000
20000
p<0.01
**
*
***
IL-1
2 (
pg
/ml)

Capítulo 1 - Resultados
64
En todas las citoquinas analizadas se detectaron niveles menores en la infección
con B. abortus WT con respecto a la infección con la mutante btpAbtpB-. La citoquina
con mayor modulación fue IL-1β, con un aumento de aproximadamente 15 veces en la
infección con btpAbtpB- respecto de la infección con WT. TNF-α aumenta
aproximadamente 10 veces respecto del control. IL-12 duplica el valor en
sobrenadantes de células dendríticas infectadas con btpAbtpB- respecto de las
infectadas con wt. Estos resultados, junto con los obtenidos de TNF-α, se encuentran
en concordancia con lo reportado en bibliografía (Salcedo et al. 2013).
Las proteínas Btp modulan la respuesta inmune pulmonar frente a
agonistas TLR
Los ensayos realizados hasta este momento indicaban que las proteínas Btp de B.
abortus pueden modular la respuesta inflamatoria a esta bacteria in vivo y por
distintos tipos celulares in vitro. Para evaluar la potencia inhibitoria de las proteínas
Btp se diseñó un ensayo en el que los ratones fueron inoculados intratraquealmente
con PBS, B. abortus WT o la mutante btpAbtpB- 24 h antes de la administración
intratraqueal de LPS de E. coli. Al día siguiente se sacrificaron los animales y se
realizaron cortes histológicos de los pulmones colectados.
Como muestra en la Figura 20, la administración del agonista TLR4 a ratones que
habían recibido PBS (PBS/LPS) genera inflamación y produce un infiltrado con
predominio de polimorfonucleares, aunque también se encuentran presentes células
mononucleares. Cuando se pre-infecta con B. abortus WT y luego se estimula con LPS
(WT/LPS) se observa una histología casi comparable con la de los pulmones controles
(PBS/PBS, no mostrado), con signos débiles de vasocongestión. Sin embargo, cuando
se infecta con btpAbtpB- y luego se estimula con LPS se observan signos inflamatorios
con infiltrado de células polimorfonucleares, células mononucleares y vasocongestión.
Si bien este infiltrado inflamatorio es menor que el del grupo PBS/LPS, es también
definitivamente mayor al del grupo WT/LPS.
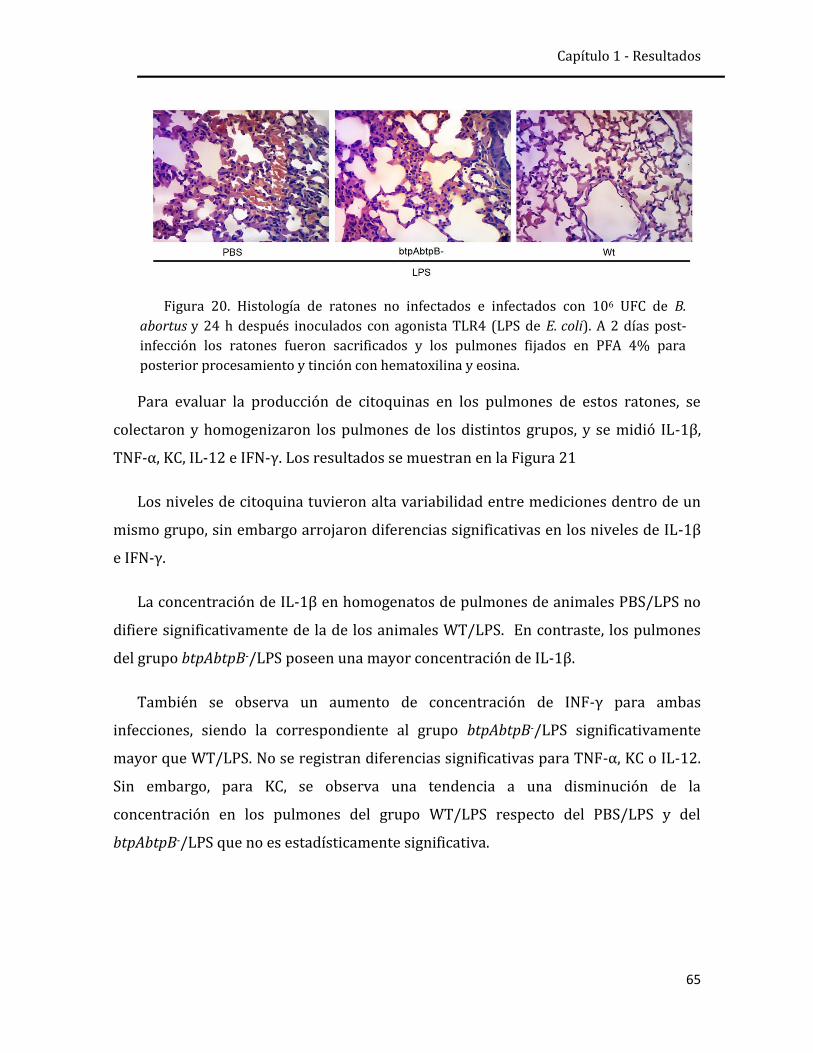
Capítulo 1 - Resultados
65
Figura 20. Histología de ratones no infectados e infectados con 106 UFC de B.
abortus y 24 h después inoculados con agonista TLR4 (LPS de E. coli). A 2 días post-
infección los ratones fueron sacrificados y los pulmones fijados en PFA 4% para
posterior procesamiento y tinción con hematoxilina y eosina.
Para evaluar la producción de citoquinas en los pulmones de estos ratones, se
colectaron y homogenizaron los pulmones de los distintos grupos, y se midió IL-1β,
TNF-α, KC, IL-12 e IFN-γ. Los resultados se muestran en la Figura 21
Los niveles de citoquina tuvieron alta variabilidad entre mediciones dentro de un
mismo grupo, sin embargo arrojaron diferencias significativas en los niveles de IL-1β
e IFN-γ.
La concentración de IL-1β en homogenatos de pulmones de animales PBS/LPS no
difiere significativamente de la de los animales WT/LPS. En contraste, los pulmones
del grupo btpAbtpB-/LPS poseen una mayor concentración de IL-1β.
También se observa un aumento de concentración de INF-γ para ambas
infecciones, siendo la correspondiente al grupo btpAbtpB-/LPS significativamente
mayor que WT/LPS. No se registran diferencias significativas para TNF-α, KC o IL-12.
Sin embargo, para KC, se observa una tendencia a una disminución de la
concentración en los pulmones del grupo WT/LPS respecto del PBS/LPS y del
btpAbtpB-/LPS que no es estadísticamente significativa.

Capítulo 1 - Resultados
66
Figura 21. Panel de citoquinas correspondiente a la infección con 106 UFC/animal
de B. abortus wt o btpAbtpB- y 24 h después, inoculados con agonista TLR4 (LPS de E.
coli). A 2 días post-infección los ratones fueron sacrificados y los pulmones extraídos y
homogenizados. IL-1β, TNF-α, KC, IFN-γ e IL-12 se midieron por ELISA en los
homogenatos. *p<0.05, ** p<0.01,*** p<0.001 respecto de no infectado (NI)
PBS wt btpAbtpB-0
2000
4000
6000
8000p=0.08
*
LPS 24h p.i.
IL-1
(p
g/m
l)
PBS wt btpAbtpB-0
200
400
600
LPS 24h p.i.
TN
F (
pg
/ml)
PBS wt btpAbtpB-0
1000
2000
3000
4000
5000
LPS 24h p.i.
KC
(p
g/m
l)
PBS wt btpAbtpB-0
2000
4000
6000
8000
LPS 24h p.i.
IL-1
2 (
pg
/ml)
NI wt btpAbtpB-0
50
100
150
200
250
LPS 24h p.i.
*
***
p<0.01
IFN
- (
pg
/ml)

Capítulo 1 - Resultados
67
Las proteinas Btp modulan la respuesta de MA infectados frente a un
agonista TLR4
Teniendo en cuenta la capacidad de las proteínas Btp de modular la respuesta
inflamatoria pulmonar total, como se mostró en la sección anterior, resultó de interés
evaluar este efecto a nivel de macrófagos alveolares (MA). Para evaluar la capacidad
moduladora de las proteínas Btp, se infectaron MA con B. abortus WT o btpAbtpB-.
Luego de 24 h p.i. las células se estimularon con LPS de E. coli (1µg/mL) como
agonista TLR4 y al día siguiente se colectó sobrenadante para analizar los niveles de
IL-1β, IL-12 y KC por ELISA. Loa resultados se muestran en la Figura 22.
La infección con la cepa WT seguida de estimulación con LPS induce un aumento
significativo de los niveles de IL-1β e IFN-γ respecto del control no infectado
estimulado con LPS, mientras que no se registran diferencias para IL-12 o KC.
Si bien no existen modulaciones negativas respecto de los MA sin infectar
estimulados con LPS, si se midió un aumento significativo de los niveles de IL-12, IFN-
γ e IL-1β en los sobrenadantes de los macrófagos infectados con la doble mutante
respecto de los infectados con la cepa WT.
B. abortus WT sobrevive en pulmones estimulados con LPS
Brucella abortus btpAbtpB- produce signos de inflamación moderada en los
pulmones de los ratones infectados. Sin embargo, este reclutamiento de células y
ambiente proinflamatorio no afecta finalmente el recuento de UFC en pulmón. Una
posible explicación es que el grado de inflamación generada, aunque mayor que el
inducido por la cepa WT, sea insuficiente para controlar la infección. Si esto fuera así,
cabe la posibilidad de que una respuesta inflamatoria pulmonar más marcada pueda
controlar a la bacteria. Nos propusimos entonces evaluar la capacidad de la bacteria
de sobrevivir en el ambiente inflamatorio producido por un agonista TLR4. Para ello
administramos intratraquealmente LPS de E. coli a ratones Balb/c y a las 24 h post-
administración se infectaron por vía intratraqueal con B. abortus WT y con la mutante

Capítulo 1 - Resultados
68
Figura 22. Secreción de IL1β, IL-12, IFN-γ y KC en macrófagos alveolares. Las
células fueron infectadas con MI de 100 bacterias/célula y luego de 24 h estimulados
con LPS de E. coli. Al día siguiente el sobrenadante fue colectado y las citoquinas
medidas por ELISA. *p<0.05, ** p<0.01,*** p<0.001 respecto de no infectado (NI)
NI btpAbtpB- wt0
1000
2000
3000
4000
5000
6000 ***
**
LPS 24h p.i.
p<0.001
mIL
-1
(p
g/m
l)
NI btpAbtpB-
wt0
500
1000
1500
2000
2500
*
p<0.05
LPS 24h p.i.
IL-1
2 (
pg
/ml)
NI btpAbtpB- wt0
5000
10000
15000
20000
25000
30000
35000
LPS 24h p.i.
ns
KC
(p
g/m
l)
NI btpAbtpB- wt0
2000
4000
6000
8000
LPS 24h p.i.
***
p<0.05
IFN
- (
pg
/ml)
***
btpAbtpB-. Los pulmones de los animales fueron extraídos a las 24 h p.i. y
homogenizados, y las UFC se midieron por plaqueo en TSA y las citoquinas por ELISA.
Cuando los animales fueron estimulados intratraquealmente con LPS de E. coli y
posteriormente infectados con B. abortus WT, no se observaron cambios significativos
en los recuentos de UFC en los pulmones respecto de aquellos ratones controles que
recibieron PBS previo a la infección (Figura 23).
Sin embargo, cuando se administró LPS y posteriormente se infectó con la mutante
btpAbtpB- se observó una disminución de un 97% del inóculo respecto del grupo
pretratado con PBS e infectado con la misma cepa, con uno de los recuentos por
debajo del límite de detección de la técnica. Estos resultados sugieren que en ausencia

Capítulo 1 - Resultados
69
de Btp B. abortus es sensible a la repuesta inmune pulmonar desencadenada por
agonistas de TLR4.
Al analizar los niveles de citoquinas en los homogenatos de los pulmones se
observa que no existen diferencias significativas en los niveles de las citoquinas
analizadas para los grupos administrados con PBS o LPS y posteriormente infectados
con la cepa WT (PBS/WT, LPS/WT respectivamente), sugiriendo una modulación del
efecto del LPS en el grupo LPS/WT.
Esta modulación de la respuesta en la secreción no se observa en los grupos
infectados con la mutante btpAbtpB-, donde se registra un aumento de IL-1β para el
grupo que fue administrado con LPS y posteriormente infectado con la doble mutante
(LPS/btpAbtpB-) respecto de los animales administrados con PBS e infectados con la
mutante (PBS/btpAbtpB-). Los niveles de secreción de IL-1β del grupo LPS/btpAbtpB-
son mayores a los medidos para el grupo LPS/WT, indicando una modulación por
parte de la bacteria WT aún una vez establecida la respuesta al agonista TLR, que no
puede ser realizada por la doble mutante. Sin embargo, no se registran diferencias en
los pulmones de los grupos PBS/btpAbtpB- versus PBS/WT.
Un perfil similar de secreción se registra para KC e IL-12, donde los niveles de
secreción de la quemoquina y la citoquina aumentan en los pulmones del grupo
LPS/btpAbtpB, siendo significativamente mayor que los del grupo LPS/WT. Aquí sí se
registra una diferencia en la secreción de los grupos PBS/btpAbtpB y PBS/WT, siendo
el primero significativamente superior al segundo.

Capítulo 1 - Resultados
70
Figura 23.Supervivencia de B. abortus wt y btpAbtpB- en pulmones estimulados con LPS.
(A) Recuento de UFC en homogenatos de pulmones totales 24 h p.i. inoculados previamente
con PBS o LPS de E. coli. (B) Secreción de IL1β, TNF-α, IFN-γ y KC en homogenatos de
pulmón enteros, medido por ELISA. *p<0.05, ** p<0.01 respecto del grupo PBS/btpAbtpB-
PBS LPS PBS LPS100
101
102
103
104
105
106
wt btpAbtpB-
*ns
UF
C/ó
rgan
o
PBS LPS PBS LPS0
200
400
600
800
wt btpAbtpB-
TN
F (
pg
/ml)
PBS LPS PBS LPS0
1000
2000
3000
4000
5000
wt btpAbtpB-
p<0.05
**
IL-1
(p
g/m
l)
PBS LPS PBS LPS0
500
1000
1500
2000
wt btpAbtpB-
p<0.05
p<0.001
**
KC
(p
g/m
l)
PBS LPS PBS LPS0
200
400
600
800
1000
p<0.001
wt btpAbtpB -
*
p<0.05
IL-1
2 (
pg
/ml)
A
B

71
Discusión

Capítulo 1 - Discusión
72
Existen muchos patógenos que infectan y colonizan las vías aéreas. Varios de ellos
generan inflamación y sintomatología pulmonar característica, desplegando toda su
capacidad patogénica en un florido cuadro localizado predominantemente en el
pulmón (Scharf et al. 2010a; de Vries et al. 2009). Otros patógenos como Brucella,
usan el pulmón como puerta de entrada para luego diseminarse hacia otros órganos,
produciendo escasas manifestaciones pulmonares. En el caso de Brucella dichas
manifestaciones se presentan en un bajo porcentaje de pacientes, e incluye: neumonía
lobular, neumonía intersticial o derrame pleural (Pappas et al. 2003)
Estudios in vitro realizados previamente por nuestro grupo indican que Brucella
puede infectar varios tipos celulares constituyentes del pulmón y, que estas células
responden a la infección con un perfil levemente proinflamatorio ya sea directamente
o indirectamente por interacción con otras células infectadas (Ferrero et al. 2009;
Ferrero et al. 2010; Ferrero et al. 2014)
El objetivo de este trabajo de Tesis fue evaluar la infección por Brucella de ratones
Balb/c en las primeras etapas para caracterizar la respuesta inmune innata
desencadenada en el pulmón, así como evaluar los mecanismos que utiliza la bacteria
para evadir la vigilancia inmunológica en el pulmón y diseminarse sistémicamente.
Los resultados obtenidos indican que B. abortus es capaz de infectar y persistir en
pulmón murino, pudiéndose diseminar desde el sitio de infección hacia los órganos
periféricos. Si bien esto fue descripto en un trabajo previo, (Kahl-McDonagh, Arenas-
Gamboa, and Ficht 2007), en aquella oportunidad se puso el foco en tiempos más
tardíos de la infección, abarcando desde la primera a la octava semana p.i. En este
trabajo de Tesis evaluamos la respuesta inmune innata y la cinética de diseminación
durante la primera semana p.i. Nuestros resultados indican que Brucella es capaz de
diseminarse a otros órganos tan tempranamente como 1 día p.i., lo que concuerda con
otro trabajo donde encontraron diseminación desde pulmón ya a las 18 hs p.i.
(Archambaud et al. 2010). Mientras que en ese estudio los animales se infectaron por
inhalación de aerosoles, en esta Tesis se utilizó la inoculación intratraqueal. Si bien
ésta no replica exactamente la inhalación, tiene como ventaja la certeza de cantidad de

Capítulo 1 - Discusión
73
inóculo que se deposita y que asegura la entrada por vías aéreas inferiores, lo cual, a
diferencia de la infección por inhalación, permite descartar el ingreso contaminante
por vía digestiva o conjuntival (Watson et al. 1969; Driscoll et al. 2000; Munder et al.
2002)
Brucella abortus exhibe una rápida diseminación desde pulmón hacia órganos
periféricos y una persistencia prolongada en pulmón(Archambaud et al. 2010; Kahl-
McDonagh et al. 2007). Esto podría deberse, al menos en parte, a una escasa respuesta
inmune pulmonar, posiblemente modulada por la bacteria en el momento de la
infección para lograr una diseminación más efectiva. Además se han reportado
manifestaciones pulmonares en pacientes infectados con B. abortus en una incidencia
muy baja (5-7%) (Pappas et al. 2003) Por ello decidimos evaluar la respuesta inmune
innata que se monta en el pulmón frente a Brucella en los primeros días de infección.
Para ello evaluamos los niveles de citoquinas en los pulmones de los animales
infectados.
Nuestros resultados muestran un leve aumento de las citoquinas IL-1β y MCP-1 a 3
días p.i. y sólo con cargas bacterianas altas (106 UFC/ ratón), mientras que el resto de
las citoquinas proinflamatorias (IL-12, KC, TNF-α e IFN-γ) no se diferencian del
control hasta 7 días p.i. A diferencia de B. abortus, patógenos respiratorios como
Legionella pneumophila, Moraxella catarrhalis o Klebsiella pneumoniae aumentan la
concentración de citoquinas o quemoquinas proinflamatorias en el pulmón o en el
líquido de lavado broncoalveolar entre las 6 y las 24 hs p.i. (Brieland et al. 1994;
Gutbier et al. 2015; Rosen et al. 2015). En contraste, la respuesta a B. abortus parece
débil y retardada en el tiempo. En relación con esta escasa capacidad proinflamatoria
de B. abortus cabe mencionar que en infecciones intraperitoneales realizadas en
paralelo con B. abortus y Salmonella en modelos murinos se encontró un aumento en
la concentración sérica de las citoquinas IL-10, IL-1β, IL-6 y TNF-α en los animales
infectados con Salmonella a diferencia de los infectados con Brucella (Barquero-Calvo
et al. 2007).
Las citoquinas TNF-α, IL-12 e IFN-γ son necesarias para mejorar la capacidad
bactericida de los macrófagos contra B. abortus (Murphy et al. 2001). En nuestro

Capítulo 1 - Discusión
74
modelo de infección intratraqueal estas citoquinas recién aumentan en los animales
infectados hacia el día 7 p.i. y para inóculos elevados de 106 UFC/animal. La ausencia
de una respuesta innata temprana podría favorecer la diseminación sistémica de la
bacteria desde el sitio inicial de infección, debido a que a los 7 días postinfección,
cuando la secreción de citoquinas en pulmón es mayor, la bacteria ya colonizó y
replicó en bazo e hígado. Además, los niveles de TNF-α en pulmones de animales
infectados en los dos primeros días p.i. son menores que en los controles. Se han
reportado mecanismos de evasión de la bacteria que disminuyen la secreción de TNF-
α en infecciones in vitro, que también podrían estar actuando in vivo. Trabajos previos
del grupo mostraron que vesículas de membrana externa de B. abortus, así como la
proteína VirB7 de esta bacteria, inhiben la secreción de TNF-α en monocitos de la
línea THP-1 en respuesta a agonistas TLR (Pollak et al. 2012). Otros estudios han
demostrado que una proteína de membrana externa de 25 kDa de Brucella produce el
mismo efecto (Jubier-Maurin et al. 2001).
Se puede concluir que la respuesta inmune innata contra Brucella en los primeros
días p.i. es de carácter levemente proinflamatorio, y se monta cuando ya han sido
colonizados los principales órganos periféricos.
El análisis histológico demostró que la infección con B. abortus produce cambios
en la composición celular del pulmón afectado, observándose un leve infiltrado de
células mononucleares. Sin embargo, la citometría no permitió precisar la identidad
de las células infiltrantes. No se observaron cambios en células dendríticas (CD11b+,
CD11c+, F4/80-,) o macrófagos alveolares (F4/80+, CD11c+, CD11b-) pero hubo una
tendencia a un mayor número de linfocitos CD4+ o CD8+ en los pulmones de los
animales infectados. Este infiltrado leve concuerda con los niveles bajos y retardados
de quemoquinas obtenidos en los homogenatos. Sin embargo, se observa un mayor
número de células totales en los homogenatos de los ratones infectados que en los
controles, lo que podría deberse a un reclutamiento de células que todavía no hemos
tipificado. En concordancia con nuestros resultados, trabajos publicados describen
que ratones infectados con B. melitensis aerosolizada presentan cambios histológicos
leves durante el transcurso de la infección (Mense et al. 2001). Asimismo, en un
modelo de infección intranasal de Macaca mulatta con B. melitensis se detectaron

Capítulo 1 - Discusión
75
alteraciones histológicas en los pulmones infectados correspondiente a una mínima
neumonía en una pequeña porción de los animales (Mense et al. 2004).
El reconocimiento antigénico por TLR induce una cascada de señalización que
desencadena la transcripción de citoquinas proinflamatorias. Teniendo en cuenta que
durante la infección intratraqueal con B. abortus en el modelo murino se genera una
escasa y retarda secreción de las mismas, decidimos evaluar el efecto de las proteínas
BtpA y BtpB de Brucella sobre la respuesta inmune desencadenada por la infección
intratraqueal. In vitro, estas proteínas son potentes inhibidores de TLR2, TLR5 y TLR9
en células dendríticas derivadas de médula ósea murina (Salcedo et al. 2013).
Cuando se infectaron los ratones intratraquealmente con la doble mutante
btpAbtpB- de B. abortus la histología pulmonar reveló un mayor infiltrado de células
mononucleares que los pulmones de ratones infectados con la cepa wt. Esto sugiere
que las proteínas Btp modulan el reclutamiento de las células inmunes hacia el
pulmón. Aun así, el infiltrado inflamatorio generado por la doble mutante no fue
sustancial y tampoco fue comparable al descripto para bacterias que son patógenos
pulmonares, como Legionella pneumophila, donde se evidencia una clara neumonía
con un importante infiltrado neutrofílico (Brieland et al. 1994)
Este aumento en el infiltrado de pulmones infectados con la doble mutante podría
haber supuesto un aumento en la eliminación de la bacteria, asumiendo que más
células inmunes en el sitio de infección lograrían controlar la replicación o
persistencia. Sin embargo, al realizar el recuento de UFC de la mutante en los
homogenatos de pulmón no se registraron cambios en su número desde las dos horas
p.i. hasta el día 7 p.i. Esto resulta interesante, ya que a pesar del mayor infiltrado
inflamatorio pulmonar no se logra eliminar más eficientemente a la bacteria. Resta
caracterizar por citometría el infiltrado producido en respuesta a la mutante Btp. A
diferencia de lo que ocurre en el pulmón, el recuento de bacterias viables en los
órganos periféricos de los ratones infectados por vía intratraqueal con la doble
mutante fue mucho menor que en los órganos de los ratones infectados con la cepa wt,
cercano al límite de detección de la técnica. Este menor recuento en órganos
periféricos sugiere dos posibilidades: a) que las proteínas Btp participan en la

Capítulo 1 - Discusión
76
diseminación desde el pulmón hacia los órganos periféricos, con lo cual la mutante
tiene una menor capacidad de diseminación, o b) que las proteínas Btp modulan los
mecanismos inmunes involucrados en la eliminación de la bacteria en los órganos
periféricos. Para abordar la segunda posibilidad evaluamos el perfil de citoquinas en
los bazos de animales infectados con la cepa salvaje o con la doble mutante para
evaluar diferencias que podrían indicar una mayor activación inmune del bazo en los
ratones infectados con la mutante. Los resultados mostraron que los valores de IL-1β,
IL-12, TNF-α y KC a 1 día p.i. aumentan significativamente en los ratones infectados
con la cepa doble mutante a diferencia de lo que ocurre en los bazos de los animales
infectados con la wt, donde no se detectaron aumentos en la producción de citoquinas
respecto de los animales no infectados para ningún tiempo analizado, con la excepción
de KC a 1 día p.i., que coincide con la primera detección de bacteria en el órgano. Esto
sugiere que la menor carga bacteriana de la doble mutante en órganos periféricos
luego de la infección intratraqueal se debería, al menos en parte, a su menor
capacidad de modular la respuesta inmune antibacteriana en esos órganos. Cabe
destacar que, hasta donde sabemos, este estudio es el primero en demostrar in vivo
que las proteínas Btp le confieren una ventaja de supervivencia a B. abortus, ya que
hasta el momento esto sólo se había explorado en estudios in vitro.
El alvéolo pulmonar está formado por diversos tipos celulares, incluyendo
pneumocitos tipo I y II, células endoteliales y macrófagos alveolares (MA). Los MA son
las principales células fagocíticas ubicadas en la luz alveolar, y son capaces de
responder a los patógenos inhalados iniciando la respuesta inmune pulmonar. En el
caso de B. abortus estudios previos indican que la bacteria establece su nicho
replicativo en MA, los cuales responden a la infección con una secreción retardada y
disminuida de citoquinas proinflamatorias en comparación con la estimulación por
agonistas TLR4 como el LPS de E. coli (Ferrero et al., 2014). Estos datos sugieren que
B. abortus posee algún mecanismo inhibitorio de la secreción de citoquinas o de la
activación de MA. En concordancia con esta hipótesis, los estudios realizados en este
trabajo de Tesis sugieren que las proteínas Btp tendrían un rol en el control de la
secreción de citoquinas por los MA, indicado tanto por la menor transcripción del gen
de IL-1β (il1b) como por la menor secreción de varias citoquinas (IL-1β, IFN-γ y KC)

Capítulo 1 - Discusión
77
en los MA infectados con la cepa wt respecto de los infectados con la mutante
btpAbtpB-. Trabajos previos demostraron resultados similares para la transcripción
de il1b y tnf en esplenocitos de ratones IRF-1-/- infectados con B. melitensis y su
mutante knock-out para BtpB (TcpB), proteína homóloga a su homónima en B. abortus
(Radhakrishnan et al. 2009). En nuestras investigaciones, la transcripción de tnf no
resultó modificada respecto de los MA sin infectar a 24 h p.i. Existen reportes donde el
aumento de la transcripción para tnf en macrófagos alveolares ocurre a tiempos más
tempranos, como 4h, (Becker et al. 2005), y cabe la posibilidad que el momento de
transcripción diferencial quede por fuera de nuestro tiempo de medida.
Además de la modulación de citoquinas, pueden existir otros mecanismos de
evasión del sistema inmune. Francisella tularensis evade el sistema inmune mediante
la inducción de la secreción de TGF-β en bazo y pulmón (Bosio et al. 2007). En
nuestro caso la infección de MA con B. abortus también generó un aumento de la
transcripción de tgfb, no siendo afectado por la presencia de las proteínas Btp. Si este
aumento en la transcripción de ARNm se refleja efectivamente en un aumento en la
concentración de la citoquina extracelular, es posible que sea este otro mecanismo de
control de la respuesta inmune.
Estudiamos también la capacidad de las proteínas Btp de modular la secreción
de citoquinas: IL-1β, IFN-γ y KC en MA infectados con B. abortus. Para IFN-γ y KC
encontramos que las proteínas Btp podían modular la secreción a 24 h p.i.,
bloqueando la secreción de IFN-γ. Sin embargo, a 48 h p.i. ya no se observa la
modulación.
Al analizar la concentración de IL-1β (el producto clivado, secretado y con
actividad biológica del gen il1b) en sobrenadante de cultivo de MA infectados, se
detectó un aumento para ambas infecciones, siendo el mayor para la infección con la
doble mutante. Esto concuerda con las diferencias en la expresión a nivel de ARNm.
Para la secreción de IL-1β es necesaria la conjunción de dos señales: 1) la activación
de un receptor TLR y consecuente incremento en la producción del precursor pro-IL-
1β, 2) la activación de los inflamasomas, lo cual conduce a la activación de caspasa-1
que madura por clivaje la pro-IL-1β (forma no secretada) a IL-1β (forma secretada)
(Bauernfeind & Hornung 2013). Con los estudios que hemos realizado hasta el

Capítulo 1 - Discusión
78
momento no se puede determinar a qué nivel estarían actuando las proteínas Btp para
disminuir la secreción de IL-1β en MA infectados, pero dada su conocida actividad
inhibitoria sobre la señalización de TLRs es posible que el mecanismo de acción sea
mediante la inhibición de la “señal 1”.
La IL-1β es una citoquina crítica para la mayoría de los procesos infecciosos.
Estudios realizados en ratones knockout mostraron que los inflamasomas
dependientes de la proteína adaptadora ASC (necesarios para la maduración y
secreción de IL-1β e IL-18) contribuyen al control de la infección con B. abortus por
vía intraperitoneal (Gomes et al. 2013). Sin embargo, en estudios realizados en
ratones KO para IL-1β no se encontraron diferencias en las UFC de B. abortus
recuperadas a 14 días p.i. de bazos de animales infectados intraperitonealmente
(Weiss et al. 2005). El rol de los inflamasomas en la respuesta pulmonar a Brucella es
todavía desconocido. El rol de protector de IL-1β en la respuesta inmune pulmonar
depende del agente infeccioso, habiéndose reportado casos donde juega un papel
protector, como en Mycobacterium tuberculosis (Yamada et al. 2000), y otros casos
donde el patógeno se beneficia de la inflamación producida, como el caso de
Streptococcus pneumonie (Lemon et al. 2015).
Para evaluar la importancia de IL-1β en la defensa contra la infección intratraqueal
por B. abortus se infectaron ratones C57BL/6 wt y knock-out para el receptor de IL-1β
(IL-1R) y para caspasa-1, y luego se midieron las UFC en los homogenatos
pulmonares. Se debió recurrir a la cepa C57BL/6 ya que los ratones knock-out
disponibles fueron derivados de esa cepa. La cinética de replicación de B. abortus en
los ratones C57BL/6 wt fue similar a la obtenida en ratones Balb/c en los tiempos
analizados (2 y 7 días p.i.) mostrando que, al menos en lo que respecta a la
permisividad a la replicación bacteriana, ambas cepas de ratones se comportan igual.
Hallamos que en ausencia de receptores para IL-1 la bacteria aumenta su recuento a
los 7 días p.i. respecto del inóculo inicial. Sin embargo, el recuento de bacterias a los 2
y los 7 días p.i. es mayor en los ratones KO para IL-1R. El aumento en el recuento de
bacterias viables en los pulmones de los ratones deficientes en IL-1R muestra la
importancia de IL-1 para el control pulmonar de la infección por B. abortus.

Capítulo 1 - Discusión
79
Los epitelios son parte importante de la respuesta inmune innata. No sólo forman
una barrera física al ingreso de microorganismos y partículas, sino que también
pueden secretar citoquinas y quemoquinas en respuesta a antígenos microbianos y a
factores solubles producidos por células inmunes. Además de los MA, los pneumocitos
son un tipo celular inmunológicamente activo en el alveólo. Para evaluar la
contribución de este tipo celular en la infección murina aislamos pneumocitos tipo II a
partir de pulmón y los infectamos con B. abortus wt y las mutantes en proteínas Btp.
En este estudio demostramos que Brucella puede infectar pneumocitos murinos,
los que a su vez responden secretando KC y MCP-1 de manera dependiente de la
multiplicidad de infección. Sin embargo, la cantidad de KC secretada luego de la
infección con la cepa wt fue mucho menor que para la infección con mutantes simples
o dobles para las proteínas Btp, donde los valores específicos de secreción de KC se
duplican respecto de la infección con wt. Esto indica que si bien Brucella es capaz de
infectar pneumocitos y generar una respuesta inmune, la expresión de las proteínas
Btp en la bacteria atenúa en parte dicha respuesta.
En los trabajos donde se describieron por primera vez las mutantes para las
proteínas Btp se demostró que estas proteínas pueden modular la secreción de TNF-α
e IL-12 en células dendríticas infectadas con B. abortus (Salcedo et al. 2008; Salcedo et
al. 2013). Las células dendríticas (CD) tienen un importante papel en la respuesta
inmune pulmonar ya que por su localización inmediatamente debajo de los
pneumocitos pueden captar antígenos luminales (a través de sus proyecciones
interdigitantes) o antígenos que traslocan el epitelio. En respuesta a la detección de
antígenos foráneos las CD se activan, incrementando la expresión de moléculas del
CMH pero también la secreción de diversas citoquinas, incluyendo IL-12 e IL-1β.
Además, en algunas infecciones pulmonares las células dendríticas pueden actuar
como transportadores (“caballos de Troya”) de bacterias viables hacia los ganglios
drenantes, contribuyendo a la diseminación del patógeno. Este fenómeno se verifica
en el caso de la infección inhalatoria por Brucella, ya que tanto los MA como las células
dendríticas pulmonares migran a los ganglios drenantes portando bacteria viable
(Archambaud et al. 2010). Teniendo en cuenta estos antecedentes y nuestros
resultados que indican la importancia de IL-1β en el control pulmonar de la infección

Capítulo 1 - Discusión
80
por B. abortus, nos pareció importante determinar si las proteínas Btp también
pueden modular la expresión de IL-1β en CD. Los resultados obtenidos indicaron que
las proteínas Btp inhiben la secreción de IL-1β en CD infectadas con B. abortus, (junto
con una inhibición de TNF-α e IL-12 como ya estaba descripto).
En conjunto los resultados obtenidos indican la capacidad de B. abortus de
modular la respuesta inmune de células pulmonares fagocíticas y no fagocíticas, lo
cual podría contribuir a su supervivencia prolongada en el pulmón. Además, teniendo
en cuenta que en el contexto fisiológico del alvéolo los distintos tipos celulares están
en estrecha proximidad y se estimulan o inhiben recíprocamente a través de factores
solubles, la modulación de las citoquinas producidas por un tipo celular podría
influenciar la secreción de las producidas por otro.
Diversos patógenos suprimen activamente la respuesta inmune para asegurar su
supervivencia en el hospedador. En muchos casos la inmunosupresión no es específica
para antígenos del patógeno sino que afecta a la respuesta inmune a estímulos
antigénicos generales (ej. PAMPs) aplicados con posterioridad a la infección (Bosio et
al. 2007). Teniendo en cuenta la falta de activación inmune observada en los
pulmones de los ratones durante los primeros días luego de la infección con B. abortus
y la modulación que ejercen las proteínas Btp de la bacteria sobre la secreción de
citoquinas en macrófagos alveolares, pneumocitos y células dendríticas quisimos
evaluar la capacidad de B. abortus de suprimir activamente la respuesta inflamatoria a
un agonista TLR. Para estos estudios elegimos el LPS de E. coli, un conocido agonista
TLR4.
Como se esperaba, los pulmones de los ratones estimulados con LPS de E. coli
luego del pretratamiento con PBS mostraron altos grados de infiltrado intersticial. En
cambio, aquellos provenientes de ratones previamente infectados con B. abortus wt y
luego estimulados con LPS (grupo wt/LPS) exhibieron un infiltrado escaso o nulo. Por
otra parte, cuando la preinfección se realizó con la doble mutante Btp se obtuvo un
infiltrado intermedio entre el observado en el grupo PBS/LPS y el observado en el
grupo wt/LPS. Estos resultados indicarían que la infección por B. abortus modula la

Capítulo 1 - Discusión
81
respuesta inmune pulmonar a un agonista TLR4 (LPS de E. coli) y que las proteínas
Btp son sólo parcialmente responsables de ese fenómeno.
El efecto de la preinfección con B. abortus sobre la respuesta a LPS de E. coli se
evaluó también a nivel de la producción de citoquinas en homogenatos de pulmón. Los
niveles de la quemoquina KC fueron menores (aunque las diferencias no fueron
estadísticamente significativas) en el grupo wt/LPS respecto de los grupos PBS/LPS y
btpAbtpB-/LPS. Esto podría explicar la diferencia en los infiltrados pulmonares
observados en los estudios histológicos, ya que KC es un quimioatractante de
neutrófilos. Resta por evaluar si la preinfección con la cepa wt puede modular también
otras quemoquinas inducidas por LPS en el pulmón. Es sabido que KC es secretada por
células epiteliales en respuesta a agonistas TLR y citoquinas proinflamatorias como
IL-1β. A su vez, es secretada por MA también en respuesta a PAMPs. Notablemente,
hallamos que KC no es modulada por la preinfección con B. abortus en MA estimulados
con LPS, con lo que es posible que las diferencias de KC entre grupos observada en los
homogenatos pulmonares se deban a la modulación de citoquinas que estimulan su
secreción, como IL-1β (ver luego).
En homogenatos pulmonares de los 3 grupos experimentales se evaluaron
también los niveles de IL-1β e IFN-γ. A diferencia de lo observado con KC, en el caso
de estas citoquinas no se observaron diferencias entre el grupo wt/LPS y el grupo
PBS/LPS, lo cual en principio sugeriría que la preinfección con B. abortus no modula la
producción de IL-1β inducida por el estímulo con LPS. Sin embargo, cuando la
preinfección se realizó con la doble mutante (grupo btpAbtpB-/LPS) se obtuvieron
niveles de IL-1β significativamente mayores a los del grupo wt/LPS. Esto demuestra
que las proteínas Btp contribuyen a disminuir la respuesta de IL-1β en pulmones
estimulados con LPS, a pesar de que la preinfección con B. abortus wt no pareció tener
un efecto modulador. Una posible explicación a estos resultados aparentemente
contradictorios es que, en el ensayo de preinfección con la cepa wt el efecto
antiinflamatorio (a nivel de IL-1β) producido por las proteínas Btp de la bacteria se
vea contrarrestado por un efecto proinflamatorio producido por los propios antígenos
de B. abortus, generando un efecto neto neutro que no difiere del producido por el
PBS. Es decir que en ambos grupos (PBS/LPS y wt/LPS) la secreción de IL-1β

Capítulo 1 - Discusión
82
observada se debería sólo al efecto del LPS de E. coli. En el caso de la preinfección con
la mutante, en cambio, al desaparecer el efecto antiinflamatorio de las proteínas Btp
se pondría en evidencia el efecto proinflamatorio sumado del LPS y de los antígenos
de B. abortus, dando como resultado una mayor secreción de IL-1β.
Los resultados obtenidos en experimentos similares realizados con MA aislados
siguieron el mismo perfil: se halló muy disminuida la concentración de IL-1β e IFN-γ
en células preinfectadas con la cepa wt y luego estimuladas con LPS (wt/LPS) respecto
de las infectadas con la doble mutante (btpAbtpB-/LPS), demostrando el rol
modulador de las proteínas Btp. Para ambas citoquinas, no obstante, los niveles
producidos por MA del grupo wt/LPS fueron algo mayores que los de las células
pretratadas con PBS (PBS/LPS). Sólo en el caso de IL-12 los niveles de los grupos
wt/PBS y PBS/LPS fueron similares.
Conjuntamente estos resultados indican que B. abortus, a través de sus proteínas
Btp, ejerce algún grado de modulación inmune disminuyendo el grado de
reclutamiento de células y la producción de KC inducidos por el LPS de E. coli en
pulmón. Por otra parte, si bien en las condiciones experimentales evaluadas no se
logró demostrar que B. abortus module otras citoquinas proinflamatorias en
comparación con los ratones o los MA no preinfectados, los resultados sugieren
fuertemente la capacidad de las proteínas Btp de modular la producción de tales
citoquinas ante el estímulo con LPS.
La activación previa de los receptores TLR pulmonares por agonistas específicos
puede incrementar los mecanismos efectores de la inmunidad innata frente al ingreso
posterior de un patógeno. A este fenómeno se lo conoce como resistencia innata
estimulada. La protección contra bacterias conferida por este mecanismo se ha
probado con diferentes patógenos respiratorios, como Bordetella pertussis,
Streptococcus pneumoniae y Pseudomonas aeruginosa (Errea et al. 2010; Evans et al.
2010). Para probar la capacidad de la respuesta inmune innata pulmonar estimulada
por LPS para eliminar B. abortus, infectamos intratraquealmente ratones
condicionados previamente con LPS de E. coli (también administrado por vía
intratraqueal).

Capítulo 1 - Discusión
83
Nuestros resultados muestran que B. abortus resiste la inmunidad innata inducida
por LPS ya que no se observan diferencias significativas en los recuentos de bacterias
viables en los pulmones de los animales pre-condicionados con LPS respecto de los
controles pre-condicionados con PBS. Estos resultados contrastan de alguna manera
con un estudio previo que demostró que los macrófagos murinos estimulados con LPS
de E. coli tienen mayor poder bactericida frente a B. abortus (Barquero-Calvo et al.
2007). No obstante, debe tenerse en cuenta que en un caso se trata de una infección in
vivo por vía mucosa y en el otro caso se trata de una infección in vitro sobre un único
tipo celular.
La capacidad de sobrevida de B. abortus en el ambiente pulmonar previamente
inflamado con LPS de E. coli parece depender sustancialmente de las proteínas Btp. A
diferencia de lo observado con la cepa wt, la mutante carente de estas proteínas
exhibe un recuento disminuido en los pulmones de animales precondicionados con
LPS en comparación con los de animales administrados con PBS. A su vez, este grupo
(LPS/btpAbtpB-) produce altos niveles de citoquinas proinflamatorias, condición que
podría ser causal del recuento disminuído. En conjunto los resultados sugieren que B.
abortus puede resistir los fenómenos inflamatorios pulmonares desencadenados por
el LPS sólo si puede comenzar a modularlos (gracias a la expresión de proteínas Btp),
mientras que si no puede hacerlo (por carecer de estas proteínas) su supervivencia
pulmonar se ve afectada.
Aunque no sabemos cómo opera exactamente esa modulación, un escenario
posible es que al momento de administrar B. abortus wt (24 h post-LPS) todavía
quede LPS de E. coli en las vías aéreas, de forma tal que las proteínas Btp detienen la
estimulación de TLR4 por el LPS remanente (y la estimulación de otros TLR por
antígenos de B. abortus). Por otra parte, aunque ya no quedara LPS en las vías
respiratorias, la preadministración de este PAMP habrá inducido el reclutamiento y
preactivación de células fagocíticas como ha sido indicado por estudios previos (Le
Feuvre et al. 2002; Deng et al. 2013). En ese caso habrá una mayor densidad de
receptores TLR, incrementando la capacidad de detectar antígenos de B. abortus y de
desencadenar mecanismos microbicidas. La capacidad de las proteínas Btp de inhibir

Capítulo 1 - Discusión
84
la señalización por TLRs podría ser crucial para proteger a la bacteria de esos
mecanismos.
En conjunto, los resultados obtenidos indicarían que las proteínas Btp conferirían
a B. abortus características que le permitirían modular la respuesta inmune en el
pulmón favoreciendo su supervivencia y permitiendo su diseminación hacia órganos
periféricos.
Hasta donde sabemos, estos resultados muestran por primera vez in vivo el rol
inmunomodulador de las proteínas Btp en pulmón. Por otra parte, es el primer
estudio en demostrar la participación de estas proteínas en la supervivencia de las
bacterias en órganos blanco. Estos resultados podrían contribuir no solo a entender la
infección inhalatoria por Brucella y al diseño de estrategias de profilaxis, sino que
además sienta precedentes sobre mecanismos de inmunoevasión que podrían ser
compartidos por otros patógenos respiratorios.

85
Capítulo 2
Caracterización de la infección in vitro de pneumocitos
tipo II primarios humanos con B. abortus 2308

86
Introducción

Capítulo 2 - Introducción
87
El epitelio respiratorio cumple no sólo una función de barrera anatómica, sino
que además expresa receptores capaces de reconocer moléculas microbianas y puede
responder a la presencia de patógenos secretando citoquinas (IL-1β, IL-6, TNF-α) y
quemoquinas (IL-8, MCP-1, eotaxina, CCL20) que dan inicio a una respuesta
inflamatoria (Sha et al. 2004; Mayer & Dalpke 2007; Koyama et al. 2000; Starner et al.
2003).
En el primer capitulo de esta Tesis evaluamos in vivo en ratones la respuesta
inmune inntata pulmonar a Brucella abortus y analizamos in vitro la respuesta de los
tipos celulares pulmonares más importantes (neumocitos y macrófagos alveolares)
obtenidos de pulmones murinos. Aunque el modelo de ratón es imprescindible para
poder evaluar distintos aspectos de la infección y para caracterizar el rol de distintas
moléculas del sistema inmune, cabe recordar que el ratón no es un hospedador
natural de la infección por Brucella. Para tener una aproximación a la interacción
hospedador-patógeno en infecciones humanas debe recurrise a modelos in vitro
realizados con células de origen humano.
Recientemente nuestro grupo de trabajo ha demostrado que el epitelio
pulmonar humano responde a la infección por Brucella secretando citoquinas, aunque
a niveles bajos. Sin embargo, puede también responder a la infección de manera
indirecta, luego del estímulo con citoquinas proinflamatorias producidas por
macrófagos infectados, secretando IL-8, MCP-1 y GM-CSF (Ferrero et al., 2010). En el
sentido inverso, citoquinas secretadas por las células epiteliales infectadas son
capaces de inducir en los macrófagos la secreción de MCP-1. Estos resultados sugieren
que durante la infección por Brucella el epitelio respiratorio humano podría montar
una leve respuesta inflamatoria en forma directa o indirecta. No obstante, debe
tenerse en cuenta que tales estudios se realizaron con líneas celulares alveolares
(A549) y bronquiales (Calu-6) establecidas a partir de tumores pulmonares. Por lo
tanto, dado que no se trata de células normales, nos interesó validar tales resultados
utilizando células pulmonares primarias.

Capítulo 2 - Introducción
88
Otro aspecto de la interacción entre Brucella y el epitelio alveolar que puede ser
mejor evaluada con un cultivo primario es la capacidad de la bacteria de traslocar la
barrera epitelial. Como se mencionó en la Introducción general, no se conocen con
precisión los mecanismos por los cuales la bacteria logra pasar del lumen alveolar a la
circulación sistémica, aunque un estudio hecho en ratones sugiere que la bacteria
podría ser transportada a los ganglios linfáticos por macrófagos alveolares infectados
(mecanismo que se conoce como “caballo de Troya” dado que Brucella puede
sobrevivir dentro de los macrófagos) (Archambaud et al 2003). No obstante, existen
otros posibles mecanismos (descriptos para patógenos de ingreso inhalatorio) que no
han sido explorados hasta el momento y que también podrían contribuir a la
traslocación epitelial de Brucella. En particular nos interesó evaluar el pasaje
transcelular y el pasaje paracelular. En el primer caso el patógeno ingresa por el polo
apical del epitelio y egresa por la cara basolateral, con lo cual no altera las uniones
intercelulares y las propiedades de barrera del epitelio se mantienen intactas. En el
segundo caso el patógeno atraviesa el espacio entre células (espacio paracelular) en
sentido apical-basal, para lo cual necesita alterar las uniones intercelulares, lo cual se
traduce en un aumento de la permeabilidad de la barrera epitelial. Para realizar
estudios in vitro sobre estos mecanismos es imprescindible contar con un cultivo de
células epiteliales confluente y polarizado, con formación de uniones estrechas entre
células que redunden en una verdadera barrera epitelial. Este tipo de cultivos no se
logra con la línea celular A549, que es la única línea humana de neumocitos tipo II,
dado que no se establecen buenas uniones intercelulares (Steimer et al 2005). En
cambio, si se logran con cultivos primarios de pneumocitos.

89
Materiales y Métodos

Capítulo 2 – Materiales y Métodos
90
Crecimiento de B. abortus
B. abortus 2308 se creció en caldo tripteína de soja a partir de colonias obtenidas
en agar tripteína de soja (TSA). Se estimó el número de unidades formadoras de
colonia (UFC) midiendo la densidad óptica del cultivo a 600 nm, y se determinó el
inóculo exacto por plaqueo de diluciones seriadas en TSA.
Aislamiento y purificación de células epiteliales
Los pneumocitos tipo II fueron aislados de tejido pulmonar siguiendo los
procedimientos publicados (Murphy et al. 1999). Las muestras fueron cedidas por la
Unidad de Trasplante Pulmonar Hospital Favaloro, a través de un convenio de
colaboración entre esta institución y el CONICET, protocolo N° 3099. Las muestras de
tejido humano pulmonar fueron obtenidas de zonas saludables y normales de los
límites de seguridad que se extraen junto a los tumores pulmones durante las cirugías.
Brevemente, los tejidos fueron lavados con Solución A (0.130 M NaCl,5.2 mM KCl, 10.6
mM HEPES, 2.6 mM Na2HPO4, 10 mM D-glucosa, pH 7.4) para remover el exceso de
sangre y contaminaciones. Luego el tejido fue picado en piezas de 1 mm de grosor. El
tejido fue digerido con tripsina (0.5% en solución A adicionado con 1.9 mM CaCl2 y
1.29 mM de MgSO4) y luego incubado a 37°C durante 20 minutos en agitación. El
digesto fue filtrado a través de una malla de 100 µm y colectado sobre SFB
conteniendo DNAsa (400 U/ml, SIGMA). Esta suspensión se filtró por una malla de 40
µm. El filtrado fue centrifugado y el pellet enriquecido con pneumocitos tipo II fue
resuspendido en medio de cultivo y dispensado sobre placas de 6 pocillos para
permitir la adhesión de leucocitos y células adherentes contaminantes. Las placas
fueron incubadas en la estufa a 37º C durante 2 h. Finalizada la incubación las células
no adherentes fueron colectadas y separadas en un gradiente de densidad discontinuo
de Percoll (1.089 g/ml y 1.040 g/ml). La capa celular interfacial enriquecida en
pneumocitos tipo II fue colectada y lavada dos veces con Solución A. Las células fueron
contadas por exclusión con azul Trypan y sembradas en placas de 48 pocillos
previamente tratadas con colágeno de tipo I bovino, seroalbúmina bovina y
fibronectina humana. Las células se dejaron adherir durante 36 h, luego el medio de
cultivo fue cambiado cada 48 h hasta llegar a confluencia (5 a 6 días).

Capítulo 2 – Materiales y Métodos
91
Explantos de pulmón humanos.
Para obtener los explantos el tejido pulmonar fue cortado en fragmentos de
aproximadamente 1 cm3, los que fueron cultivados en 1 ml de RPMI 1640 en placa de
24 pocillos. Para la infección se incubaron con 1 ml de una suspensión 1x108 UFC/ml
de B. abortus.
Infección de cultivos celulares
Las células epiteliales respiratorias se infectaron con B. abortus 2308 a MI de 100,
250, 500 o 1000 bacteria/célula durante 2 horas a 37 ºC. Finalizada la incubación las
células fueron lavadas repetidas veces e incubadas con medio completo suplementado
con 100 ug/ml de gentamicina para eliminar las bacterias extracelulares. A diferentes
tiempos luego de la adición de gentamicina (24 y 48 h), los sobrenadantes de cultivo
fueron cosechados y conservados a -80° C para luego determinar TNF-α, IL-1β, IL-8 o
KC, CCl-20 o MCP-1 por ELISAs comerciales, mientras que las células fueron lavadas,
lisadas con Tritón X-100 al 0.2% en agua destilada estéril, y plaqueadas para
determinar el número de UFC.
Infección de explantos
Los explantos de pulmón se infectaron con 108 UFC de B. abortus 2308 durante 24
horas a 37 ºC. A 24 hs se agregó gentamicina a los sobrenadantes e inmediatamente
fueron colectados y conservados a -80° C para luego determinar TNF-α, IL-1β, IL-6,
CCL5, CCL20 o MCP-1 por ELISAs comerciales.
Ensayos de traslocación
Pneumocitos aislados de pulmón humano como se describió anteriormente fueron
cultivados sobre insertos Transwell de 0.4 µm de poro y superficie de 0.33 cm2
previamente recubiertos con colágeno, fibronectina y seroalbúmina bovina, a una
densidad de 150.000 células por filtro durante 11 días. Periódicamente se midió la
resistencia eléctrica transmembrana (TEER) y una vez alcanzado un valor >700 Ω.cm2
(condición donde se establece la monocapa confluente) se realizaron los ensayos de
traslocación.

Capítulo 2 – Materiales y Métodos
92
Pasaje paracelular.
Se adicionó B. abortus en el compartimiento superior (cara apical) del cultivo de
pneumocitos (MI 200 bacterias/célula) junto con dextrán de 40 kDa conjugado a FITC
(200µg/ml) en medio de cultivo (medio SAGM suplementado, sin antibióticos). Luego
de 2 o 4 h de incubación se colectó una alícuota de medio del compartimento inferior
(cara basolateral del cultivo) y se plaqueó para realizar recuento de bacterias viables,
mientras que otra alícuota se conservó para medir fluorescencia. Como control se
usaron monocapas celulares tratadas sólo con Dextrán-FITC (sin bacterias) y
Transwell sin monocapa celular (como control de pasaje máximo).
Pasaje transcelular.
Se infectaron monocapas confluentes de pneumocitos cultivados en Transwell con
B. abortus a una MI de 200 bacterias/células y luego de dos horas de incubación se
eliminaron las bacterias extracelulares incubando 40 min a 37°C con medio SAGM
suplementado con gentamicina y estreptomicina. Finalizada la incubación se lavaron
ambos compartimientos, apical y basal, y se adicionó medio sin antibióticos en el basal
y Dextrán-FITC en el superior. Para evaluar el pasaje transcelular se colectó medio
del compartimiento basal a las 24 h p.i. y se plaqueó en TSA para recuento de
bacterias viables. A su vez se midió fluorescencia en el medio colectado para
comprobar la integridad de la monocapa celular.
La presencia de bacteria y ausencia de fluorescencia indica que la bacteria puede
traslocar a través del epitelio o endotelio polarizado sin dañar la integridad de la
barrera celular, lo que sugeriría una ruta transcelular de traslocación. Mientras que la
presencia de bacteria y de Dextrán-FITC en el compartimiento inferior indicaría
traslocación paracelular (con alteración de la permeabilidad de membrana).

93
Resultados

Capítulo 2 - Resultados
94
B. abortus induce la secreción de citoquinas y quemoquinas en cultivo primario
alveolar humano
Trabajos previos del grupo demuestran que B. abortus es incapaz de estimular la
secreción de citoquinas y quemoquinas en la línea de células alveolares tumorales
A549 mediante la infección directa, aún cuando es capaz de replicar hasta 48 hs p.i.
(Ferrero et al. 2009). Para corroborar la respuesta a B. abortus por parte del epitelio
alveolar humano normal se desarrolló un modelo de cultivo primario de pneumocitos
tipo II humanos provenientes de donantes sanos de pulmón. El cultivo se dejó llegar a
confluencia (aproximadamente 7 días de cultivo) y posteriormente se infectó con B.
abortus 2308 a distintas MI. Para evaluar la respuesta inmune se midió IL-6, IL-8,
CCL20, CCL5 y MCP-1 en sobrenadante de cultivo, y para evaluar la replicación
intracelular de la bacteria las células se lisaron y se realizó recuento de UFC en placa.
Como muestra la Figura 24, B. abortus fue capaz de invadir y replicar
intracelularmente, alcanzando el máximo de UFC intracelulares hacia las 48 h p.i. y la
mayor tasa de replicación durante las primeras 24 hs p.i. A 72 hs p.i. existe un
descenso marcado en el recuento de bacterias intracelulares, llegando hasta valores
menores que los iniciales.
Las células respondieron a la infección por B. abortus secretando quemoquinas
como CCL-20, IL-8 y MCP-1 de manera MI dependiente. IL-8 aumentó en respuesta a la
infección de manera significativa para todas las MI ensayadas a 24 y 48 h p.i., con
mayores niveles a las 48 h p.i. Por otro lado, CCL20 sólo registró un aumento
significativo a 24 h p.i. cuando se infecta con una MI de 1000 bacterias/células. A 48 h
los niveles se vuelven significativos para todas las MI. Los niveles de MCP-1 e IL-6
permanecen no significativos respecto del control a 24 h p.i. y a 48 h p.i. sólo las
células infectadas con MI de 1000 bacterias/células aumentan la secreción respecto
de los controles. No se detectaron niveles significativos de IL-1β o de TNF-α (no
mostrado)

Capítulo 2 - Resultados
95
0 20 40 60 80100
101
102
103
104
h p.i.
UF
C/m
l
NI
250
500
1000
IL-1 N
I25
050
0
1000
IL-1
0
1000
2000
3000
4000
5000
24 48
*
** ****
**
**
IL-8
(p
g/m
l)
NI
250
500
1000
IL-1 N
I25
050
010
00
IL-1
0
2000
4000
6000
24 48
*** ****
***
***
CC
L20 (
pg
/ml)
NI
250
500
1000
IL-1 N
I25
050
0
1000
IL-1
0
5000
10000
15000
24 48
***
*
MC
P-1
(p
g/m
l)
NI
500
1000
IL-1 N
I25
050
0
1000
IL-1
0
1000
2000
3000
4000
5000
*
24 48
*** ***
IL-6
(p
g/m
l)
A
B
Figura 24. A) Replicación de B. abortus 2308 Las células fueron infectadas a una
MI de 250 y a distintos tiempos p.i. lisadas y diluidas para realizar el recuento de
bacterias viables intracelulares por plaqueo en TSA. B) Nivel de expresión de
citoquinas y quemoquinas en respuesta a la infección por cultivo primario de
pneumocitos humanos. Las células fueron infectadas a MI de 250, 500 y 1000 o
estimuladas con 1 ng/ml de IL-1β y 24 o 48 h p.i. los sobrenadantes colectados y las
proteínas medidas por ELISA comerciales. *p<0.05, ** p<0.01,*** p<0.001 respecto de
no infectado (NI) para cada tiempo.

Capítulo 2 - Resultados
96
B. abortus induce la secreción de quemoquinas en Explantos de pulmón humano
Para lograr una mejor aproximación al parénquima pulmonar se evaluó la
respuesta inmune en un modelo de explantos de pulmones humanos infectados con B.
abortus. Se cosecharon los sobrenadantes de los explantos pulmonares a las 24 h
luego de ser infectados con B. abortus 2308 para la posterior determinación de
citoquinas por ELISA. Los resultados obtenidos mostraron que la infección con B.
Figura 25 Secreción de citoquinas y quemoquinas en modelo de explanto de pulmón
humano. Los sobrenadantes de los explantos de pulmón humano fueron colectados 24 h p.i.,
y las citoquinas medidas por ELISA comerciales. *p<0.05, ** p<0.01,respecto de no infectado
(NI)
NI B. abortus IL-10
1000
2000
3000
4000
*
***
CC
L20 (
pg
/ml)
NI B. abortus IL-10
500
1000
1500
2000
MC
P-1
(p
g/m
l)
NI B. abortus IL-10
200
400
600
800
*
CC
L-5
(p
g/m
l)
NI B. abortus IL-10
100000
200000
300000
400000
**
**
IL-6
(p
g/m
l)
NI B. abortus IL-10
100
200
300*
IL-1
(p
g/m
l)
NI B. abortus IL-10
200
400
600
800
**
**
TN
F-
(p
g/m
l)

Capítulo 2 - Resultados
97
abortus induce la secreción de CCL20, CCL5, IL-1β y TNF-α, mientras que no se
observan aumentos significativos respecto de control (NI) para MCP-1 (Figura 25). En
el tiempo p.i. evaluado, los niveles de IL-6 se redujeron en los sobrenadantes de
explantos infectados respecto del control y del estímulo con IL-1β.
Ensayo de traslocación de B. abortus en una monocapa confluente de
pneumocitos tipo II humanos.
Como se demostró arriba (figura 24 A), B. abortus puede infectar y replicar en
pneumocitos humanos tipo II. Es posible que estas células puedan actuar como
mediadores del paso de B. abortus hacía el torrente sanguíneo, debido a su
proximidad y su extensa superficie de contacto con los capilares que conforman el
pulmón. Para corroborar esta hipótesis, cultivamos pneumocitos tipo II humanos
primarios en Transwells y, una vez alcanzada la formación de una monocapa
polarizada (determinada por un TEER de más de 1000 Ω.cm2, (Elberts et al. 1999;
Fuchs et al. 2003)), aproximadamente a los 11 días de cultivo, se evaluó el pasaje
paracelular (pasaje por el espacio adyacente a las células) y el transcelular (pasaje
transitando por el interior de las células).
Pasaje paracelular
Para evaluar el pasaje paracelular, se depositó en el lado apical de las células una
suspensión de B. abortus equivalente a una MI de 100 bacterias/células junto con una
solución de dextrán marcado con FITC. A las 2 y 4 h post administración se colectó
medio del compartimiento inferior (cara basolateral), se diluyó y se plaqueó en placas
de TSA para determinar el número de bacterias viables.
Los resultados mostrados en la figura 26 y en la tabla 2 indican que Brucella
abortus puede traslocar las monocopas de pneumocitos humanos. Luego de 2 y 4 h
p.i., se recuperan UFC en el compartimiento basal que es acompañado de un aumento
en la permeabilidad al dextrán-FITC, lo que sugiere disrupción de la monocapa. La
bacteria pasa eficientemente a través de los poros del inserto Transwell, como se ve
en los recuentos del filtro sin células, usado como control de pasaje máximo.

Capítulo 2 - Resultados
98
Fluorescencia Dextran-FITC
0
1000
2000
3000
400010000
20000
30000
40000NI
I 2h
I 4h
S/C 2 h
S/C 4 h*
Un
idad
es a
rbit
rari
as
UFC/ml en compartimiento basolateral
0
100
200
300
400
500
8000
10000
12000
14000 2 h
4 h
S/C 2 h
S/C 4 h
UF
C/m
l
A B
2 h p.i. 4 h p.i.
Unidades arbitrarias de fluorescencia
UFC Unidades
arbitrarias de fluorescencia
UFC
No infectado 734 534 B. abortus 2018 208 225 106 3171 303 363 124 Sin células 36751 10000 31375 11250
Pasaje transcelular.
Para evaluar el pasaje transcelular, monocapas de pneumocitos humanos se
infectaron con B. abortus a una MI de 100 bacterias/células y luego de 2 h se eliminó
las bacterias extracelulares con gentamicina durante 40 minutos, de manera que la
únicas bacterias viables remanentes fuesen aquellas que se encontraban en el
compartimiento intracelular. Luego de 24 h, se colectó medio del compartimiento
basal y se plaqueó sobre placas de TSA. También se realizó recuento de las bacterias
intracelulares como control.
TABLA 2 Fluorescencia y recuento de bacterias viables en medio del compartimiento basal a 2 y 4 hs.
Figura 26 A) Fluorescencia en medio del compartimiento basal. NI, no infectado, I,
infectado, S/C Sin células. B) Recuento de bacterias viables en medio del compartimiento
basal a 2 y 4 hs. *p<0.05, respecto de no infectado (NI)

Capítulo 2 - Resultados
99
Como muestra la tabla 3, la infección de la monocapa fue exitosa, pero no se
detectó recuento de bacteria viable en el compartimiento basal. Estos resultados
indican que B. abortus no tiene la capacidad de translocar el epitelio por la vía
transcelular.
Unidades
arbitrarias de
fluorescencia
UFC medio
basal
UFC
intracelulares
Basal 2714 778 - -
B. abortus 5048 2060 0 150 106
Sin células 105371 3,1x105 -
TABLA 3 Fluorescencia en medio del compartimiento basal.y recuento de bacterias
viables a 24 hs.

100
Discusión

Capítulo 2 - Discusión
101
Las células epiteliales de las vías aéreas son una de las primeras barreras con las
que se encuentran los patógenos transmitidos por aire. No solo actúan como barrera
física, si no que pueden iniciar la respuesta inmune mediante la detección del
patógeno y la consecuente secreción de mediadores solubles como las citoquinas y
quemoquinas(Sha et al. 2004; Mayer & Dalpke 2007).
Trabajos previos del grupo demostraron que la línea celular A549 (pneumocitos
tipo II de origen tumoral humano) no secreta las quemoquinas IL-8 o MCP-1 en
respuesta a la infección con B. abortus, a pesar de que la bacteria es capaz de replicar
eficientemente dentro de ellas (Ferrero et al. 2010; Ferrero et al. 2009). En este
capítulo demostramos que el cultivo primario de pneumocitos tipo II también es
permisivo a la replicación de B. abortus, pero es capaz de responder con aumento de
secreción de IL-8 a diferencia de la línea A549. En cambio, los niveles de MCP-1 y de
otras cuatro citoquinas analizadas (CCL20, IL-6, IL-1β y TNF-α) no se diferenciaron
significativamente de los controles no infectados a 24 h p.i. aunque si se
incrementaron en respuesta al estímulo con IL-1β. Sólo se registaron algunos
aumentos significativos de estas citoquinas a las 48 h p.i. y para multiplicidades de
infección elevadas. Estos resultados indican que las células epiteliales alveolares son
pobres respondedoras frente a la infección por B. abortus, en concordancia con los
estudios in vitro realizados previamente por el grupo con células A549.
El ambiente pulmonar se constituye de numerosas células que interactúan entre sí
y con la matriz extracelular, generando redes de complejas dependencias e
interacciones que determinan finalmente el resultado de la respuesta inmune. Para
abordar la problemática de la interacción de Brucella con el pulmón humano
decidimos utilizar un modelo de explantos de tejido pulmonar humano. Estos modelos
se han utilizado para el estudio de otros patógenos pulmonares(Jäger et al. 2014;
Nicholas et al. 2015). Para ello analizamos la secreción de citoquinas y quemoquinas
en sobrenadante de cultivo de explantos. Similarmente a los resultados obtenidos en
infecciones de pneumocitos tipo II, existe un aumento leve de las quemoquinas CCL20
y CCL5 en respuesta a la infección, mientras que MCP-1 no se modula. Además,
aumentan moderadamente los niveles de citoquinas proinflamatorias como IL-1β o

Capítulo 2 - Discusión
102
TNF-α. Esto concuerda con un perfil proinflamatorio leve. Nuestros resultados
muestran una disminución en los niveles de IL-6 en los explantos infectados con B.
abortus. La implicancia de esta disminución es difícil de predecir, debido a su rol
controvertido en inflamación pulmonar (Yu 2002)
La diferencia entre los perfiles de secreción de citoquinas entre el modelo de
explanto y el cultivo celular es evidente, y es importante el subsecuente estudio de las
otras células que componen pulmón como macrófagos alveolares y células
endoteliales y la relevancia de los factores secretados en la respuesta inmune final.
Trabajos previos del grupo han demostrado que factores solubles secretados por
monocitos de la línea humana THP-1 pueden inducir la secreción de citoquinas y
quemoquinas en líneas de células epiteliales humanas (Ferrero et al. 2010). Si bien
existen diferencias fenotípicas y funcionales entre estos monocitos y los macrófagos
alveolares, es un fuerte indicio predictivo de lo que puede ocurrir.
Estos resultados representan la continuación del estudio de la interacción entre
Brucella y el epitelio respiratorio humano, demostrando que la bacteria produce una
respuesta inmune levemente proinflamatoria en tejido y cultivo de células humanas.
Brucella puede colonizar distintos órganos desde su entrada por pulmón. Una de
las preguntas fundamentales respecto de este fenómeno es cómo hace la bacteria para
atravesar la barrera epitelial pulmonar desde su sitio de ingreso inicial (el lumen
alveolar o bronquial). Existen estudios que demuestran que puede ser transportada
por macrófagos alveolares en primera instancia y luego por células dendríticas hacia
los ganglios mesentéricos, mecanismo denominado “Caballo de Troya”(Archambaud
et al. 2010). Sin embargo, es posible que existan otros mecanismos de traslocación
actuando en simultáneo. Debido a la anatomía de los alvéolos pulmonares, la distancia
que separa la luz alveolar de los capilares sanguíneos es muy poca (0,4 µm), con lo que
una pequeña disrupción implicaría inmediato acceso a la circulación sistémica. Es por
esto que se evaluó la posibilidad que B. abortus pueda atravesar una monocapa
confluente de pneumocitos humanos, ya sea de manera paracelular, alterando las
uniones intercelulares, o transcelular, ingresando por el lado apical de las células,

Capítulo 2 - Discusión
103
transitándolas y egresando por el lado basal. Estudios previos del grupo demostraron
que Brucella no es capaz de traslocar paracelular o transcelularmente una monocapa
polarizada de células 16HBE14o-, que se consideran como el mejor modelo de epitelio
bronquial respiratorio humano (Ferrero, Tesis Doctoral, 2010), pero restaba realizar
los estudios con monocapas de pneumocitos primarios.
Nuestros resultados muestran que Brucella es capaz de atravesar
paracelularmente la monocapa de pneumocitos humanos, ya que se detectan bacterias
viables en el medio basal luego de 2 y 4 h de incubación con la bacteria en el lado
apical. Además acompaña a este pasaje un aumento en la permeabilidad a dextrán,
macromolécula usada como control de integridad de epitelio. No detectamos pasaje
transcelular a 24 h p.i., a pesar de que las células se infectaron exitosamente. Es
posible que el pasaje paracelular ocurra en las infecciones in vivo, ya que en la
histología de la infección intratraqueal murina se observó extravasación de hematíes
hacia la luz alveolar, sugiriendo incremento en la permeabilidad epitelio alveolar.
Ambos resultados sugieren la posibilidad de que haya un mecanismo de disrupción de
las uniones estrechas del epitelio alveolar pulmonar activo por Brucella durante la
infección, que deberá seguir siendo investigado. Los resultados obtenidos sugieren
que además del mecanismo de caballo de Troya, ya demostrado por Archambaud et al,
podrían existir otros mecanismos para la diseminación sistémica de B. abortus.

104
Capítulo 3
Respuesta antimicrobiana pulmonar frente a infecciones con
B. abortus

105
Introducción

Capítulo 3 - Introducción
106
Las células epiteliales de las vías aéreas son las primeras células que entran en
contacto con los microorganismos inhalados, y por lo tanto están preparadas para
montar respuestas inmunes rápidas. Como mencionamos previamente, el epitelio
respiratorio responde a la presencia de patógenos con una respuesta inflamatoria, (Li
et al. 2012; Sha et al. 2004), la cual puede ser aumentada por la acción estimulatoria
de las citoquinas secretadas por los macrófagos alveolares(Marriott et al. 2012; Ishii
et al. 2005; Korpi-Steiner et al. 2010).
Los factores producidos por el epitelio respiratorio en respuesta a la infección
incluyen beta-defensinas, péptidos antimicrobianos pequeños que pueden ser
encontrados en los fluidos respiratorios junto con otros componentes
antimicrobianos, como la lisozima y la catelicidina. La beta-defensina 2 (hBD2) es la
defensina más expresada en pulmón, y su expresión es estimulada durante las
infecciones o inflamación (Doss et al. 2010). Todas las defensinas son péptidos
catiónicos pequeños con propiedades microbicidas que contienen 6 residuos de
cisteína altamente conservados que forman tres puentes disulfuro intramoleculares.
Se ha postulado que estos péptidos interaccionan electrostáticamente con las cargas
negativas de la superficie de las membranas celulares, de las cuales son responsables
el LPS y algunos componentes de bacterias gram positivas. Esto lleva a una alteración
de la estructura de la membrana y a la creación de un poro que genera la filtración del
contenido celular (Lai & Gallo 2009). La hBD2 es efectiva contra varios patógenos in
vivo incluidos Escherichia coli, Pseudomona aeruginosa, Klebsiella pneumonie, entre
otros (Pazgier et al. 2006).
El epitelio respiratorio también responde a las infecciones con la producción de
varias quemoquinas incluida CCL20. Es un potente quimioatractante de células
dendríticas inmaduras, linfocitos T de memoria y linfocitos B. CCL20 y su receptor
específico CCR6 median el reclutamiento in vivo de células dendríticas y linfocitos en
varios tejidos, incluyendo el pulmón (Osterholzer et al. 2005; Phadke et al. 2007;
Thomas et al. 2007). El repertorio de células con receptor CCR6 también incluyen a
células Th17 (Hirota et al. 2007), hecho relevante para la respuesta inmune a agentes
infecciosos.

Capítulo 3 - Introducción
107
hBD2 y CCL20 comparten ciertas características. Ambos factores interactúan con
el receptor CCR6 (Ghannam et al. 2011; Röhrl et al. 2010; Yang et al. 1999; Barabas et
al. 2013) y como consecuencia hBD2 puede quimioatraer a linfocitos T de memoria,
células dendríticas inmaduras, mastocitos y neutrófilos (Yang et al. 1999; Niyonsaba
et al. 2002; Niyonsaba et al. 2004). En contraste, aunque CCL20 fue descripta
inicialmente como quemoquina, estudios recientes demostraron que esta molécula
puede tener actividad antimicrobiana frente a bacterias Gram positivas y Gram
negativas (Starner et al. 2003; Yang et al. 2003; Chan et al. 2008).
Debido a sus propiedades quimiotácticas y antimicrobianas, se postula que ambas
tienen un rol importante en la respuesta inmune innata pulmonar a patógenos
inhalados. (Ito et al. 2011; Schutte & McCray 2002; Tecle et al. 2010) Varios estudios
han demostrado la producción de hBD2 en varios tejidos en respuesta a infecciones,
sin embargo nada se sabe de la producción de hBD2 o CCL20 en la infección de células
epiteliales de pulmón en respuesta a la infección por Brucella, ni de las actividades
antimicrobianas de estas sustancias frente a este patógeno. Decidimos investigar estos
aspectos para ampliar la caracterización de la respuesta innata pulmonar a la
infección por B. abortus.

108
Materiales y métodos

Capítulo 3 –Materiales y Métodos
109
Cultivo celular.
La línea celular A549 (pneumocitos tipo II humanos) fue crecida en DMEM
suplementado con 10% SFB, 2mM de L-glutamina, 100 U/ml de penicilina 100 ug/ml
de estreptomicina. Las células Calu-6 fueron crecidas en RPMI 1640 suplementado de
la misma forma.
Para los ensayos de infección todas las células epiteliales fueron sembradas a una
densidad de 2x105 células en placas de 24 pocillos y cultivadas en atmosfera con 5%
de CO2 a 37°C en medio suplementado sin antibióticos.
Aislamiento de monocitos humanos de sangre periférica.
La sangre de donantes anónimos fue diluída al medio y separada mediante
gradiente estándar de Fycoll-Hipaque. Luego de obtenida la capa enriquecida en
células mononucleares, se contaron en cámara de Neubauer y se sembraron en placas
de plástico de 48 pocillos a razón de 1x106 células/pocillo. Luego de 2 h de incubación
a 37°C y 5% de CO2 se lavaron las células no adheridas múltiples veces con PBS. Las
células remanentes fueron mayoritariamente monocitos.
Explantos de pulmón humanos.
Las muestras de tejido humano pulmonar fueron obtenidas de zonas saludables y
normales del tejido marginal de seguridad que se extrae junto a los tumores pulmones
durante las cirugías. Las muestras fueron cedidas por la Unidad de Trasplante
Pulmonar del Hospital Favaloro, a través de un convenio de colaboración entre esta
institución y el CONICET (protocolo N°3099). Para obtener los explantos el tejido
pulmar fue cortado en fragmentos de aproximadamente 1 cm3 que fueron cultivados
en 1 ml RPMI 1640 en placa de 24 pocillos. Para la infección se incubaron con 1 ml de
una suspensión 1x108 UFC/ml de B. abortus.
Aislamiento de células primarias de pulmón
Los pneumocitos tipo II fueron aislados de tejido pulmonar obtenido como se
describió arriba, siguiendo procedimientos publicados (Murphy et al. 1999).

Capítulo 3 –Materiales y Métodos
110
Brevemente, los tejidos fueron lavados con Solución A (0.130 M NaCl, 5.2 mM KCl,
10.6 mM HEPES, 2.6 mM Na2HPO4, 10 mM D-glucosa, pH 7.4) para remover el exceso
de sangre y contaminaciones. Luego el tejido fue picado en piezas de 1 mm de grosor.
El tejido fue digerido con tripsina (0.5% en solución A adicionado con 1.9 mM CaCl2 y
1.29 mM de MgSO4) y luego incubado a 37°C durante 20 minutos en agitación. El
digesto fue filtrado a través de una malla de 100 µm y colectado sobre SFB
conteniendo DNAsa (400 U/ml, Sigma). Esta suspensión se filtró por una malla de 40
µm. El filtrado fue centrifugado y el pellet enriquecido con pneumocitos tipo II fue
resuspendido en medio de cultivo y dispensado sobre placas de 6 pocillos para
permitir la adhesión de leucocitos y células adherentes contaminantes. Las placas
fueron incubadas en la estufa a 37º C durante 2 h. Finalizada la incubación las células
no adherentes fueron colectadas y separadas en un gradiente de densidad discontinuo
de Percoll (1.089 g/ml y 1.040 g/ml). La capa celular interfacial enriquecida en
pneumocitos tipo II fue colectada y lavada dos veces con Solución A. Las células fueron
contadas por exclusión con azul Trypan y sembradas en placas de 48 pocillos
previamente tratadas con colágeno de tipo I bovino, seroalbúmina bovina y
fibronectina humana. Las células se dejaron adherir durante 36 h, luego el medio de
cultivo fue cambiado cada 48 h hasta llegar a confluencia (5 a 6 días).
Infecciones celulares.
Las infecciones de las células respiratorias con Brucella fueron realizadas a
distintas multiplicidades de infección (MI). Luego de dispensar la suspensión
bacteriana, las placas fueron centrifugadas y posteriormente incubadas durante 2
horas a 37 °C. Al finalizar la incubación, cada pocillo fue lavado tres veces con PBS
estéril. Para eliminar las bacterias intracelulares, las monocapas infectadas se
incubaron con medio suplementado con gentamicina y estreptomicina (100 µg/ml y
50 µg/ml respectivamente). A diferentes tiempos p.i. los sobrenadantes de cultivo
fueron colectados para medir péptidos antimicrobianos por ELISA comerciales.

Capítulo 3 –Materiales y Métodos
111
Análisis de la producción de péptidos antimicrobianos en respuesta a
antígenos
Las células fueron cultivadas en placas de 24 pocillos a una densidad de 5 x 105
células/pocillo y luego estimuladas con distintos antígenos de B. abortus (HKBA, LPS,
L-Omp19) o con agonistas TLR (LPS de E. coli y Pam3CSK4). Los cultivos fueron
incubados durante 24 h. Luego, los sobrenadantes fueron cosechados y conservados a
-70°C hasta ser analizados.
Estimulación con medio condicionado.
La línea premonocítica humana THP-1 fue infectada con B. abortus a una MI de
100 bacterias/célula durante 2 h y luego lavada con PBS para eliminar bacterias
extracelulares. El medio condicionado de monocitos fue cosechado a 24 h p.i.,
esterilizado mediante el filtrado a través de filtros de nitrocelulosa de poro 0.22 µm y
usado para estimular células epiteliales. El medio condicionado de THP-1 no
infectadas fue usado como control.
Para la estimulación, el medio condicionado fue usado en diluciones 1/2, 1/5 o
1/10. Luego de 24 h post-estimulación los sobrenadantes de los cultivos fueron
cosechados para medir péptidos antimicrobianos. Para calcular la secreción específica
por parte de los cultivos, los niveles ya presentes en los medios condicionados
utilizados para estimular fueron sustraídos. Los experimentos de neutralización
fueron realizados con el antagonista del receptor de IL-1 (IL-1Ra) o con un anticuerpo
neutralizante anti- TNF-α humano. Las células epiteliales fueron pre-incubadas por 1
h con IL-1Ra antes de la adición del medio condicionado. Para el ensayo de inhibición
de TNF-α, el medio condicionado de células THP-1 fue preincubado con el anticuerpo
1 h previa a su adición a las células epiteliales.
PCR semicuantitativa.
Los niveles de transcripción de ARNm para hBD2 fueron determinados mediante
PCR semicuantitativa usando primers descriptos previamente. Las células epiteliales
fueron lavadas con PBS y el RNA total fue aislado usando TRIzol. La transcripción

Capítulo 3 –Materiales y Métodos
112
reversa de 2 µg ARN total se realizó con reactivos y métodos estándard. El ADN copia
correspondiente a 200 ng de ARN fue amplificado mediante PCR conteniendo 1 mM de
primers específicos para hBD2 con un indicador de copia, gliceraldehido-3-fosfato
deshidrogenasa (GAPDH). Los productos fueron visualizados mediante una
electroforesis en gel de agarosa 2% y SYBRsafe. La especificidad del fragmento
amplificado fue corroborada mediante secuenciación.
Ensayos de actividad antimicrobiana
Para realizar los ensayos antimicrobianos se utilizaron 104 UFC en crecimiento
exponencial de B. abortus 2308, B. abortus RB51 o E. coli 25922 en buffer fosfato 10
mM pH 7.4. Las bacterias fueron incubadas con CCL20, hBD2 o hBD3 recombinantes
(PeproTech, Rocky Hill, NJ) a distintas concentraciones finales (0.1 a 50 μg/ml)
durante 2h a 37°C. Luego, fueron diluidas y plaqueadas en TSA para realizar el
recuento de UFC.

113
Resultados

Capítulo 3 - Resultados
114
B. abortus induce secreción de CCL20 pero no de hBD2 en células
epiteliales respiratorias y macrófagos.
Dado que varios patógenos estimulan la producción de hBD2 y CCL20 por parte de
las células epiteliales en respuesta a infecciones (Rivas-Santiago et al. 2005; Scharf et
al. 2010b; Harder et al. 2000), quisimos determinar si la infección con B. abortus
estimula la expresión de estos factores.
Los análisis por PCR y ELISA mostraron que B. abortus no induce la expresión ni la
secreción de hBD2 en la línea A549 (Figura 27) o Calu-6 (no mostrado), aunque ambas
líneas fueron capaces de responder a IL-1β con un aumento en la expresión del ARNm
para hBD2.
En contraste, la infección con B. abortus aumentó la secreción de CCL20 de forma
dependiente de MI en ambas líneas celulares, aunque los niveles fueron más bajos en
A549. Los niveles de CCL20 producidos por cultivos primarios de pneumocitos
humanos dieron niveles similares a los producidos por Calu-6 (Figura 28).
La producción de hBD2 y CCL20 en respuesta a la infección con B. abortus también
fue evaluada en la línea celular de monocitos THP-1 (Figura 28). La infección de
células THP-1 con B. abortus a una MI de 10 o 100 bacterias/célula no produjo
inducción de la expresión de ARNm de hBD2 a ninguno de los tiempos estudiados. Por
lo contrario, la infección indujo un aumento significativo en la secreción de CCL20 por
parte de monocitos THP-1 de una manera dependiente de la MI. En concordancia con
estos resultados, los monocitos aislados de sangre periférica y los macrófagos
derivados de monocitos produjeron cantidades significativas de CCL20 en respuesta a
la infección con B. abortus.
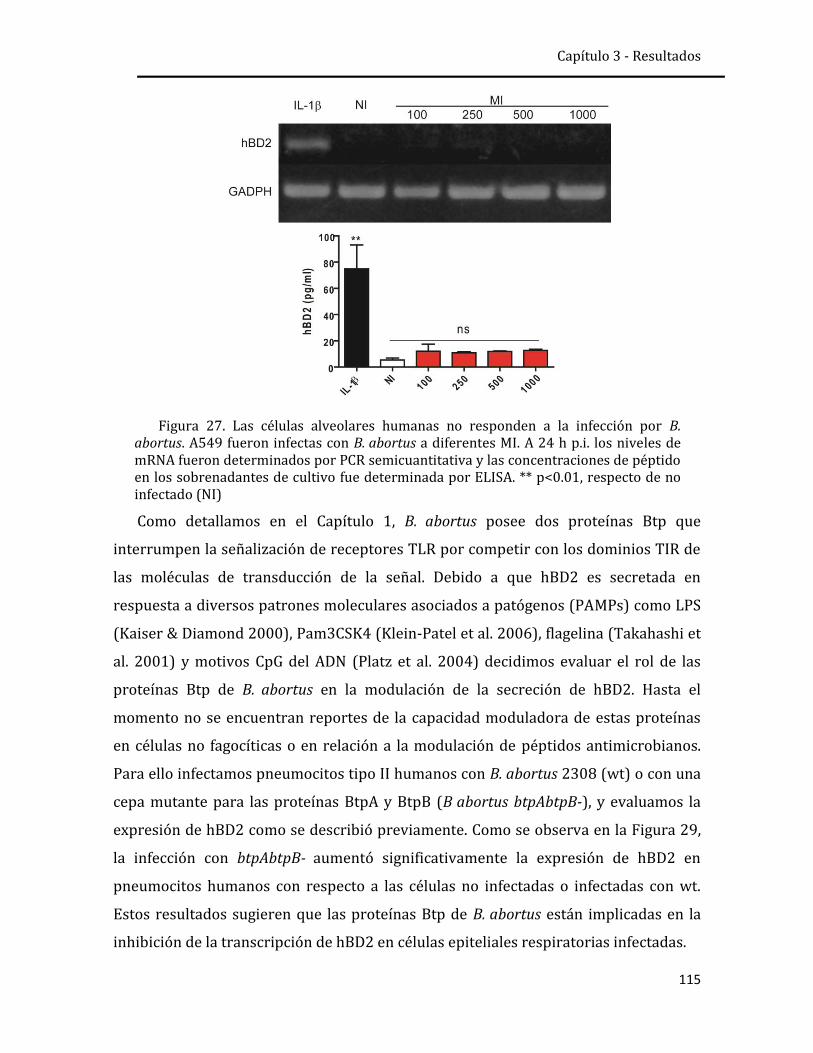
Capítulo 3 - Resultados
115
Como detallamos en el Capítulo 1, B. abortus posee dos proteínas Btp que
interrumpen la señalización de receptores TLR por competir con los dominios TIR de
las moléculas de transducción de la señal. Debido a que hBD2 es secretada en
respuesta a diversos patrones moleculares asociados a patógenos (PAMPs) como LPS
(Kaiser & Diamond 2000), Pam3CSK4 (Klein-Patel et al. 2006), flagelina (Takahashi et
al. 2001) y motivos CpG del ADN (Platz et al. 2004) decidimos evaluar el rol de las
proteínas Btp de B. abortus en la modulación de la secreción de hBD2. Hasta el
momento no se encuentran reportes de la capacidad moduladora de estas proteínas
en células no fagocíticas o en relación a la modulación de péptidos antimicrobianos.
Para ello infectamos pneumocitos tipo II humanos con B. abortus 2308 (wt) o con una
cepa mutante para las proteínas BtpA y BtpB (B abortus btpAbtpB-), y evaluamos la
expresión de hBD2 como se describió previamente. Como se observa en la Figura 29,
la infección con btpAbtpB- aumentó significativamente la expresión de hBD2 en
pneumocitos humanos con respecto a las células no infectadas o infectadas con wt.
Estos resultados sugieren que las proteínas Btp de B. abortus están implicadas en la
inhibición de la transcripción de hBD2 en células epiteliales respiratorias infectadas.
Figura 27. Las células alveolares humanas no responden a la infección por B. abortus. A549 fueron infectas con B. abortus a diferentes MI. A 24 h p.i. los niveles de mRNA fueron determinados por PCR semicuantitativa y las concentraciones de péptido en los sobrenadantes de cultivo fue determinada por ELISA. ** p<0.01, respecto de no infectado (NI)

Capítulo 3 - Resultados
116
Calu-6
NI
50 200
500
1000
IL-1
0
200
400
600
800
******
**
MI
**
CC
L20(p
g/m
l)
A549
NI
50 200
500
1000
IL-1
0
30
60
90
3000
6000
MI
***
CC
L20 (
pg
/ml)
NI
200
500
1000
IL-1
0
500
10002000
2500
3000
3500
4000
***
MI
***
Pneumocitos tipo II primarios
CC
L20 (
pg
/ml)
Monocitos
NI
10 50 100
0
100
200
300
MI
***
*****
CC
L20 (
pg
/ml)
THP-1
NI
10 50 100
0
500
1000
1500
2000
MI
***
***
***
CC
L20 (
pg
/ml)
Macrófagos
NI
10 50 100
0
200
400
600
800
1000
MI
***
**
ns
CC
L20 (
pg
/ml)
A B
Figura 28. B. abortus induce la secreción de CCL20 por las células epiteliales de las vías aéreas y monocitos humanos. (A) Calu-6, A549 y pneumocitos tipo II primarios fueron infectados con B. abortus a diferentes MI o estimulados con IL-1β (1ng/ml). (B) THP-1, monocitos humanos derivados de sangre periférica y macrófagos derivados de monocitos fueron infectados con B. abortus a MI de 10 a 100 o no infectados (NI). Los sobrenadantes fueron colectados a 48 h, y los niveles de CCl20 medidos por ELISA. *p<0.05, ** p<0.01,*** p<0.001 respecto de no infectado (NI)

Capítulo 3 - Resultados
117
NI btpAbtpB- wt0.0
0.1
0.2
0.3
0.4p<0.05
*
hB
D2/G
AD
PH
Como indicamos más arriba, las células epiteliales pulmonares respondieron a la
infección por B. abortus con producción de CCL20. Para ahondar en la importancia
fisiológica de dicha secreción, los niveles de esta quemoquina fueron evaluados en un
modelo de explantos humanos pulmonares. Como se muestra en la Figura 30 a las 24
h p.i. los niveles de CCL20 en los sobrenadantes de cultivo de los explantos infectados
fueron significativamente más altos que aquellos producidos por explantos no
infectados.
NI I0
1000
2000
3000
*
Pulmón 1
CC
L20 (
pg
/ml)
NI I0
1000
2000
3000
*
Pulmón 2
CC
L20 (
pg
/ml)
Figura 30. B. abortus induce la secreción de CCL20 en explantos de tejido de pulmón humano. Muestras de pulmón humano de aproximadamente 1 cm3 de dos donantes distintos fueron incubadas con 108 B. abortus o medio por 24 hs. Los niveles de CCL20 fueron medidos por ELISA en sobrenadantes de cultivo de explantos infectados (I) o no infectados (NI). *p<0.05, respecto de no infectado
.
Figura 29. Las proteínas Btp inhiben el aumento de la transcripción de hBD2.
Pneumocitos primarios humanos fueron infectados con B. abortus 2308 (wt) o con B.
abortus btpAbtpB- (btpAbtpB-). 24 h p.i. se lisaron las células y se midió la expresión de
ARNm específico mediante PCR semicuantitativa. Los valores fueron normalizados
respecto de GAPDH. NI: células no infectadas. *p<0.05, respecto de no infectado (NI)
.

Capítulo 3 - Resultados
118
La inducción de CCL20 por la infección es mediada por L-Omp19 y no por LPS de B.
abortus
Se evaluó si la viabilidad de B. abortus es necesaria para inducir la producción de
CCL20 por células epiteliales de pulmón y macrófagos. Para ello, se evaluó la
capacidad de Brucella muerta por calor (Heat killed Brucella abortus-HKBA) (Figura
31) para estimular la producción de CCL20. El LPS de E. coli induce la expresión e
CCL20 en distintos tipos celulares, incluidos los pneumocitos tipo II (Thorley et al.
2005; Hosokawa et al. 2005), por ello lo cual se lo empleó como control positivo. La
secreción de CCL20 fue significativamente mayor que los controles en los
sobrenadantes de células A549, THP-1 y Calu-6 estimuladas con 1 x 109 bacterias/ml
de HKBA. Estos resultados sugieren que la secreción de CCL20 puede ser inducida por
un componente estructural de B. abortus. Se ha descripto que las lipoproteínas de
Brucella inducen la producción de citoquinas por parte de distintos tipos
celulares(Giambartolomei et al. 2004; Ferrero et al. 2011). Nuestra hipótesis es que
las lipoproteínas podrían estar involucradas en el aumento de la producción de
CCL20. Para investigar esta hipótesis, estimulamos células A549, Calu-6 y THP-1 con
L-Omp19, lipoproteína modelo de B. abortus, y colectamos los sobrenadantes a las 24
h p.i. para medir CCL20.
Como se muestra en la Figura 31, L-Omp19 induce un aumento significativo en la
secreción de CCL20 de manera dependiente de la MI en los tres tipos celulares
estudiados. La producción de CCL20 también fue evaluada luego de la incubación de
las células con un hexapéptido sintético (Pam3SCK4) que mimetiza la estructura del
dominio lipídico de las lipoproteínas bacterianas. En contraposición, el LPS de B.
abortus no indujo producción de CCL20 incluso cuando se lo utilizó a alta
concentración (5 µg/ml). En conjunto, estos resultados indican que las lipoproteínas
de B. abortus están involucradas en la inducción de la secreción de CCL20 por parte de
células epiteliales pulmonares y monocitos en contacto con la bacteria

Capítulo 3 - Resultados
119
TLR2 media la secreción de CCL20 por las células bronquiales y monocitos
infectados con B. abortus
Nuestros resultados indican que las lipoproteínas de Brucella pero no su LPS
inducen la secreción de CCL20 por células epiteliales pulmonares y monocitos, por lo
cual hipotetizamos que TLR2 podría llegar a estar involucrado en la red de
señalización que lleva a la secreción de CCL20. Para evaluar esta hipótesis, monocitos
THP-1 y células epiteliales Calu-6 fueron incubadas con un anticuerpo bloqueante
anti-TLR2 antes de la infección con B. abortus. El bloqueo de TLR2 redujo
significativamente la secreción de CCL20 en las células infectadas (67.5% de
inhibición para Calu-6 y 77.9% para THP-1) mientras que la incubación con el control
de isotipo no modificó la secreción de CCL20 en ningún tipo celular (Figura 32).
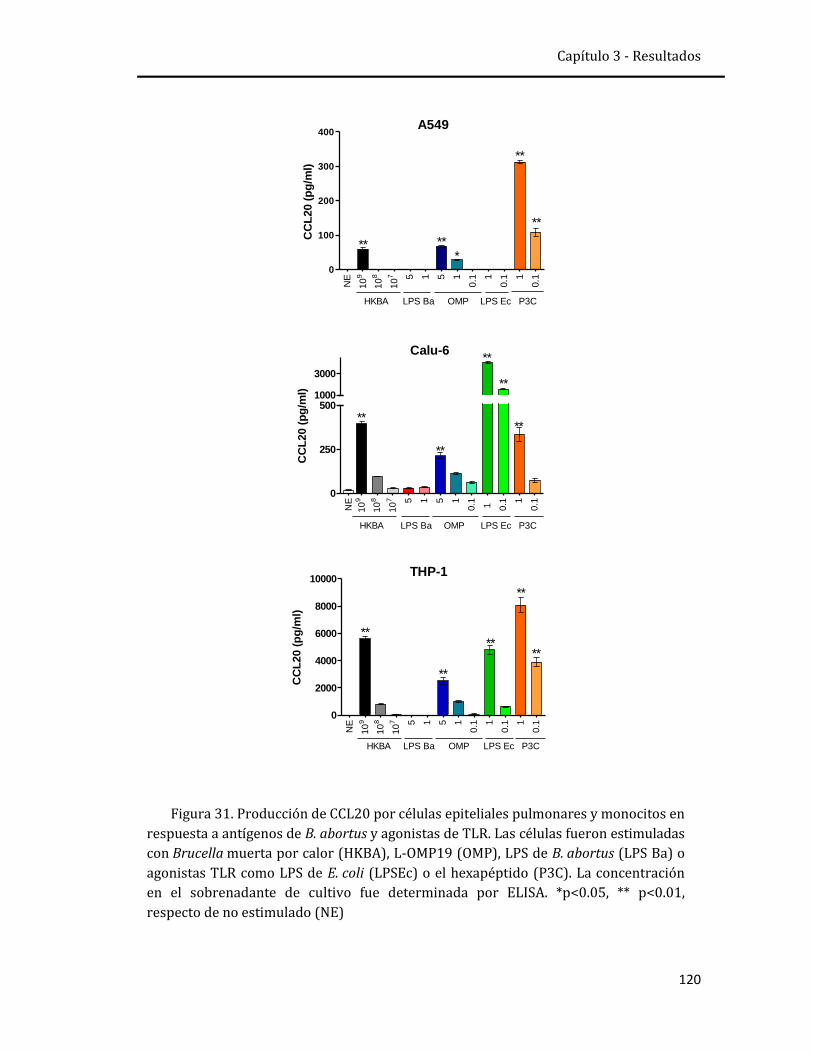
Capítulo 3 - Resultados
120
A549
NE 9
10
810
710 5 1 5 1
0.1 1
0.1 1
0.1
0
100
200
300
400
HKBA P3CLPS Ba LPS Ec
**
**
**
**
OMP
*
CC
L20 (
pg
/ml)
Calu-6N
E 91
0
81
0
71
0 5 1 5 1
0.1 1
0.1 1
0.1
0
250
5001000
3000
HKBA P3CLPS Ba LPS Ec
**
**
**
**
OMP
**
CC
L20
(p
g/m
l)
THP-1
NE 9
10
81
0
71
0 5 1 5 1
0.1 1
0.1 1
0.1
0
2000
4000
6000
8000
10000
**
**
**
****
HKBA P3CLPS Ba LPS EcOMP
CC
L20
(p
g/m
l)
Figura 31. Producción de CCL20 por células epiteliales pulmonares y monocitos en
respuesta a antígenos de B. abortus y agonistas de TLR. Las células fueron estimuladas
con Brucella muerta por calor (HKBA), L-OMP19 (OMP), LPS de B. abortus (LPS Ba) o
agonistas TLR como LPS de E. coli (LPSEc) o el hexapéptido (P3C). La concentración
en el sobrenadante de cultivo fue determinada por ELISA. *p<0.05, ** p<0.01,
respecto de no estimulado (NE)

Capítulo 3 - Resultados
121
NI anti TLR-2 Isotipo0
200
400
600
Calu-6
B. abortus
ns
****** ***
*
CC
L20 (
pg
/ml)
NI anti TLR-2 Isotipo0
500
1000
1500
2000 *** ***
*
ns
***
THP-1
B. abortus
CC
L20 (
pg
/ml)
Figura 32. TLR2 media la secreción de CCL20 por las células bronquiales y monocitos infectados con B. abortus. Calu-6 y THP-1 fueron preincubadas con el anticuerpo neutralizante contra TLR-2 o con un control de isotipo durante 1 h a 37°C previo a la infección con B. abortus. Los sobrenadantes de cultivo fueron colectados 24 h p.i. y medidos por ELISA. *p<0.05, *** p<0.001 respecto de no infectado (NI)
.

Capítulo 3 - Resultados
122
Factores secretados por monocitos infectados con B. abortus aumentan la
secreción de CCL20 y hBD2 por las células epiteliales alveolares.
Se ha demostrado que las células epiteliales secretan CCL20 y hBD2 en respuesta a
citoquinas inflamatorias como TNF-α e IL-1β (Harder et al. 2000; Reibman et al.
2003). En el pulmón los macrófagos alveolares residentes y los monocitos reclutados
a los focos de infección pueden responder a B. abortus con citoquinas proinflamatorias
que, a su vez, podrían estimular la producción de CCL20 y hBD2 por las células
epiteliales. Para evaluar esta hipótesis, células A549 fueron estimuladas con distintas
diluciones de medio condicionado de monocitos THP-1 infectados con B. abortus
(MCI). Luego de 24 h post estimulación se midió CCL20 y hBD2 en los sobrenadantes
de las células estimuladas. En paralelo, otras células fueron estimuladas con las
mismas diluciones de sobrenadante de monocitos no infectados. Los niveles de CCL20
y de hBD2 presentes en los MCI fueron restados para determinar la secreción
específica.
Como se muestra en la Figura 33, las células A549 produjeron CCL20 en respuesta
al estímulo con MCI en forma dependiente de la dosis (dilución) de este medio
condicionado. En cambio, el estímulo con medio condicionado de monocitos THP-1 no
infectados (MCNI) no produjo cambios significativos en la secreción de CCL20.
Mientras que no se había detectado transcripción de ARNm de hBD2 en las células
A549 infectadas directamente con B. abortus, la estimulación de estas células con MCI
logró inducir un aumento de la transcripción. Para correlacionar los niveles de
transcripción con la concentración de proteína, se midió hBD2 en sobrenadantes de
cultivo de las células epiteliales. Como se muestra en la Figura 33, la adición de medio
condicionado de monocitos THP-1 no infectados (MCNI) no estimula la secreción de
hBD2 respecto del basal, mientras que la estimulación con MCI aumentó la secreción
de esta defensina. Estos datos sugieren que la secreción de hBD2 por células
estimuladas con MCI es mayor que la observada cuando estas células son infectadas
directamente con B. abortus.

Capítulo 3 - Resultados
123
Figura 33. Factores secretados por monocitos infectados con B. abortus pueden aumentar la secreción de CCL20 y hBD2 por parte de las células alveolares epiteliales, Las células fueron expuestas a diferentes diluciones de MCI y luego de 24 h los niveles de transcripción y la concentración de CCL20 y hBD2 fueron medidos en los sobrenadantes de cultivo. *p<0.05, ** p<0.01 respecto de no infectado (NI)
.

Capítulo 3 - Resultados
124
Teniendo en cuenta que TNF-α e IL-1β inducen la secreción de CCL20 y hBD2 en
células alveolares pulmonares (Harder et al. 2000; Reibman et al. 2003; Tsutsumi-
Ishii & Nagaoka 2003), y que ambas citoquinas son secretadas por macrófagos
infectados con B. abortus (Ferrero et al. 2010; Ferrero et al. 2014), estas citoquinas
fueron consideradas candidatos para mediar los efectos del MCI. Para evaluarlo, se
utilizó un anticuerpo neutralizante contra TNF-α y el antagonista del receptor de IL-1,
IL-1Ra. El pretratamiento del MCI con el anticuerpo anti-TNF-α no reduce la secreción
de CCL20 o de hBD2 en las células pulmonares (Figura 34). Sin embargo, el
tratamiento de las células A549 con IL-1Ra antes de la estimulación con MCI redujo la
secreción de CCL20 en un 48.3% y 86% para las diluciones 1/5 y 1/2
respectivamente. La secreción de hBD2 por las células estimuladas con MCI también
fue reducida en un 63.6% luego del tratamiento con IL-1Ra, mientras que los niveles
de ARNm bajaron hasta niveles no detectables. Estos resultados sugieren que IL-1β es
esencial para la secreción de CCL20 y hBD2 en células A549 estimuladas con MCI.
Actividad antimicrobiana
Se probó la actividad antimicrobiana de los péptidos CCL20 y hBD2 frente a B.
abortus usando un ensayo de recuento de UFC luego de la incubación durante dos
horas de 104 UFC en buffer fosfato 10 mM con distintas concentraciones de los
péptidos. La dosis letal 50 (DL50, aquella que elimina la mitad de las UFC) de CCL20
para B. abortus fue mayor a 50 µg/ml (Figura 35). La capacidad de CCL20 de eliminar
E. coli 25922 también fue evaluada como control positivo. La DL50 para E. coli fue 1.5
µg/ml. Dada la resistencia de B. abortus a CCL20 evaluamos si la presencia del
lipopolisacárido O del LPS podría tener un efecto protectivo. Para probar esto,
evaluamos la actividad antimicrobiana de CCL20 contra B. abortus RB51, una cepa
vacunal que carece el lipopolisacárido O. Como se ve en la Figura 35, CCL20 tuvo una
DL50 de 8.7 µg/ml para B. abortus RB51, sugiriendo que el lipopolisacárido O está
involucrado en la resistencia de B. abortus a los efectos de CCL20.
También se estudió la actividad antimicrobiana de hBD2 y hBD3 contra las
distintas cepas de B. abortus. En el caso de B. abortus 2308 ninguno de los péptidos

Capítulo 3 - Resultados
125
logro disminuir la cantidad de UFC a las concentraciones ensayadas (hasta 50 µg/ml),
mientras que cuando se coincubó B. abortus RB51 con hBD3 se observó una
disminución de las UFC a la mayor concentración ensayada.
Figura 34. IL-1β media el aumento de CCL20 y hBD2 en células alveolares estimuladas con MCI. A549 fueron preincubadas con IL-1Ra por 1 h a 37°C antes de la estimulación con el medio condicionado de THP-1 infectadas con B. abortus. 24 h post-estimulación se midieron los niveles de CCL20 y hBD2 en los sobrenadantes de cultivo y los niveles de expresión de hBD2 en las células. NE: sobrenadantes de células no estimuladas ** p<0.01,*** p<0.001 respecto de no estimulado (NI)
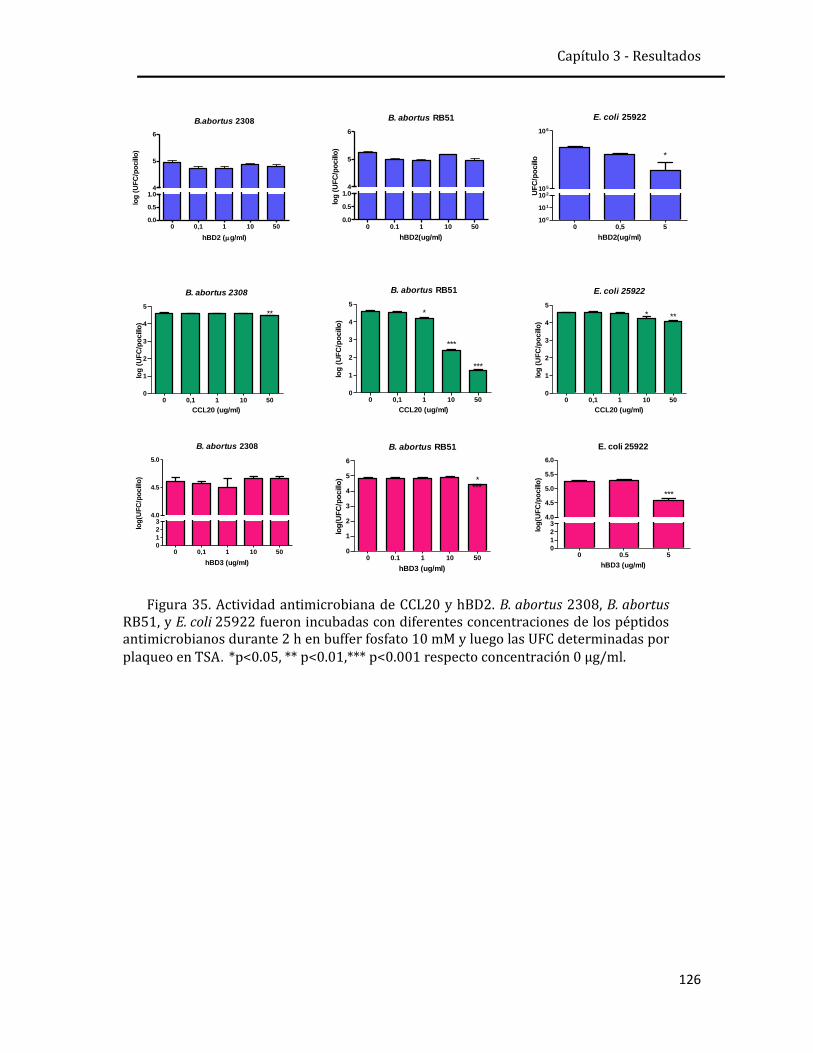
Capítulo 3 - Resultados
126
B.abortus 2308
0 0,1 1 10 500.0
0.5
1.04
5
6
hBD2 (g/ml)
log
(U
FC
/po
cill
o)
B. abortus RB51
0 0.1 1 10 500.0
0.5
1.04
5
6
hBD2(ug/ml)
log
(U
FC
/po
cill
o)
E. coli 25922
0 0,5 5100
101
102105
106
*
hBD2(ug/ml)
UF
C/p
ocill
o
B. abortus 2308
0 0,1 1 10 500
1
2
3
4
5**
CCL20 (ug/ml)
log
(U
FC
/po
cill
o)
B. abortus RB51
0 0,1 1 10 500
1
2
3
4
5
*
***
***
CCL20 (ug/ml)
log
(U
FC
/po
cill
o)
E. coli 25922
0 0,1 1 10 500
1
2
3
4
5
* **
CCL20 (ug/ml)
log
(U
FC
/po
cill
o)
B. abortus 2308
0 0,1 1 10 500
1
2
34.0
4.5
5.0
hBD3 (ug/ml)
log
(UF
C/p
ocill
o)
B. abortus RB51
0 0.1 1 10 500
1
2
3
4
5
6
****
hBD3 (ug/ml)
log
(UF
C/p
ocillo
)
E. coli 25922
0 0.5 50
1
2
34.0
4.5
5.0
5.5
6.0
***
hBD3 (ug/ml)
log
(UF
C/p
ocill
o)
Figura 35. Actividad antimicrobiana de CCL20 y hBD2. B. abortus 2308, B. abortus RB51, y E. coli 25922 fueron incubadas con diferentes concentraciones de los péptidos antimicrobianos durante 2 h en buffer fosfato 10 mM y luego las UFC determinadas por plaqueo en TSA. *p<0.05, ** p<0.01,*** p<0.001 respecto concentración 0 µg/ml.
.

127
Discusión

Capítulo 3 - Discusión
128
Los péptidos antimicrobianos son considerados un componente de defensa
importante del húesped en las superficies mucosas, incluido el pulmón. Las beta-
defensinas producidas por las células epiteliales respiratorias y los macrófagos
alveolares son secretadas al líquido de la superficie del tubo respiratorio, y aumentan
sus niveles durante los procesos inflamatorios e infecciosos. Además de su actividad
antimicrobiana directa, las beta-defensinas tienen múltiples roles como mediadores
de la inflamación(Schutte & McCray 2002; Tecle et al. 2010; Bals 2000). Asimismo, se
ha demostrado que CCL20 es producida por las células epiteliales de pulmón y puede
ser inducida por algunas infecciones.(Starner et al. 2003; Sim et al. 2009; Thorley et al.
2005) Diversos estudios han sugerido que en el pulmón, CCL20 no sólo puede ejercer
actividades quimiotácticas sobre linfocitos y células dendríticas(Grahnert et al. 2014),
sino que también puede exhibir actividad antimicrobiana(Starner et al. 2003). A pesar
de los roles conocidos de CCL20 y hBD2 en la defensa pulmonar contra infecciones y
la habilidad de Brucella spp. para infectar hospedadores por vía inhalatoria, nada se
sabe de la expresión de estas moléculas en células epiteliales de pulmón infectadas y
su actividad antimicrobiana contra este patógeno.
Los resultados indican que B. abortus induce la expresión de CCL20 en líneas
celulares alveolares humanas (A549) o bronquiales (Calu-6) y también en células
alveolares primarias obtenidas de pulmones humanos. Estos resultados están en
concordancia con otros reportados para células epiteliales de las vías aéreas
infectadas por otros patógenos (Sim et al. 2009; Park et al. 2010; Yamamoto et al.
2012) o estimuladas con antígenos microbianos (Pichavant et al. 2006; Van Maele et
al. 2014). CCL20 también es producida por monocitos/macrófagos en respuesta a
infecciones o antígenos microbianos. Esto puede ser relevante durante infecciones
pulmonares dada la presencia de macrófagos alveolares residentes y el reclutamiento
de monocitos por quemoquinas inflamatorias. En particular, estudios previos han
mostrado que células epiteliales pulmonares producen MCP-1, un quimioatractante
para monocitos(Ferrero et al. 2010). En este capítulo demostramos que la línea
monocítica THP-1, así como también monocitos y macrófagos derivados de monocitos
primarios responden a la infección por B. abortus con un incremento en la producción

Capítulo 3 - Discusión
129
de CCL20. Además, la misma respuesta se vió en explantos de tejido humano infectado
con B. abortus ex vivo, sugiriendo fuertemente que el aumento en la secreción de
CCL20 puede ocurrir en pulmones de pacientes infectados.
Conjuntamente estos resultados sugieren que los distintos tipo celulares pueden
responder a la infección con B. abortus con producción de CCL20, contribuyendo
potencialmente al reclutamiento de linfocitos y células dendríticas hacia el sitio de
infección. Los niveles alcanzados por las células epiteliales, monocitos, macrófagos y
explantos son compatibles con los reportados para la quimiotaxis de células
dendríticas y linfocitos (Thorley et al. 2005; Miao et al. 2012; Baggetta et al. 2010;
Hirota et al. 2007)
La inducción de CCl20 por B. abortus parece ser independiente de la viabilidad
celular, debido a que también fue inducida por HKBA en A549, Calu-6 y THP-1. Estos
resultados concuerdan con estudios previos donde muestran la inducción de
diferentes citoquinas y quemoquinas por HKBA en distintos tipos celulares
(Giambartolomei et al. 2004; Ferrero et al. 2011). Además, esos estudios demostraron
que las respuestas inducidas por HKBA fueron mediadas por lipoproteínas de B.
abortus y no por su LPS. Nuestros resultados concuerdan con esos estudios debido a
que encontramos que CCL20 se secreta en respuesta a la estimulación con L-Omp19
pero no al LPS de Brucella. También en concordancia con esos estudios, la respuesta
es dependiente de TLR2. La incapacidad del LPS de B. abortus de inducir secreción de
CCL20 concuerda con la baja actividad proinflamatoria descripta para esta molécula,
debida principalmente a su inusual lípido A (Giambartolomei et al. 2004; Goldstein et
al. 1992; Qureshi et al. 1994).
Contrariamente a los resultados obtenidos con CCL20, no se observa inducción de
hBD2 a nivel de ARNm o proteína en las células humanas o monocitos infectados con
B. abortus aunque el ARNm sí se incrementó en respuesta a IL-1β que fue utilizado
como control positivo. Estos resultados contrastan con aquellos encontrados en
infecciones de células epiteliales por otros patógenos cono Mycobacterium

Capítulo 3 - Discusión
130
tuberculosis, Pseudomonas aeruginosa o Aspergillus fumigatus (Alekseeva et al. 2009;
Rivas-Santiago et al. 2005; Harder et al. 2000; Moranta et al. 2010).
Las BD contribuyen a la protección de las mucosas contra los patógenos, por lo que
es razonable postular que los patógenos han desarrollado estrategias de
supervivencia contra ellas. En este sentido, se ha demostrado que la cápsula de
polisacárido de Klebsiella pneumoniae, le confiere un rol protectivo dual confiriéndole
resistencia a la acción microbicida de las β-defensinas a la vez que inhibe su
transcripción en células alveolares respiratorias humanas (Moranta et al. 2010).
Shigella flexneri también suprime la transcripción de péptidos antimicrobianos a los
que es sensible en células epiteliales colónicas humanas. Nuestros resultados
muestran que Brucella también es capaz de inhibir la transcripción de hBD2 en
pneumocitos primarios murinos. Se ha demostrado que los receptores TLR están
involucrados en la transcripción y secreción de hBD en células epiteliales de
diferentes orígenes (Takahashi et al. 2001; Duits et al. 2003; Kaiser & Diamond 2000;
Platz et al. 2004) En los últimos años se han descripto dos proteínas de Brucella, BtpA
y BtpB, capaces de interferir en las vías de señalización de TLR2, TLR4 y como
consecuencia de ello, controlan la activación de células dendríticas y macrófagos. Sin
embargo el rol de estas proteínas en la producción de citoquinas y defensinas por las
células epiteliales respiratorias no ha sido evaluado. Nosotros demostramos que los
pneumocitos primarios expresan significativamente más hBD2 cuando son infectados
con una cepa mutante de B. abortus para los genes que codifican para btpA y btpB.
Estos resultados sugieren que las proteínas Btp de Brucella son capaces de interferir
en la transcripción de hBD2. Se requerirán otros estudios para evaluar el rol de estas
proteínas en la regulación de la transcripción de otros péptidos antimicrobianos
No existen estudios que comparen en paralelo la secreción de hBD2 y de CCL20 en
células pulmonares epiteliales en respuesta a patógenos o a antígenos microbianos.
En células colónicas epiteliales (Caco-2) ambas moléculas fueron inducidas a nivel de
ARNm por la infección con la cepa salvaje de E. coli enteropatogénica, mientras que
sólo CCL20 fue inducida por la mutante carente de flagelina (Khan et al. 2008). Esto
sugiere que, al menos en este modelo, la inducción de vías de señalización que

Capítulo 3 - Discusión
131
desembocan en la producción de CCL20 difiere de aquellas involucradas en la
expresión de hBD2.
Aunque las células epiteliales no produjeron hBD2 en respuesta a la infección con
B. abortus, hay que tener en cuenta que en el contexto pulmonar existen interacciones
entre estas células con los fagocitos residentes o reclutados. Estas interacciones
podrían amplifica la respuesta inmune a patógenos. Como modelo de este tipo de
interacciones, estimulamos las células epiteliales pulmonares con medio condicionado
de monocitos infectados con B. abortus (MCI). Notablemente encontramos que la
expresión de hBD2 fue significativamente inducida en A549 por MCI y un efecto
similar fue observado para CCL20. Los experimentos usando bloqueantes revelaron
que IL-1β, pero no TNF-α, participa en la inducción de hBD2 y la secreción de CCL20
por MCI. Estos resultados son parecidos a los previamente reportados por nuestro
grupo para la respuesta de quemoquinas por las A549 y células bronquiales humanas
a MCI (Ferrero et al. 2010). En ese trabajo las células epiteliales tenían una pobre o
nula respuesta proinflamatoria a la infección directa con B. abortus pero exhibían un
marcado incremento en la producción de quemoquinas (IL-8 y MCP-1) luego de la
estimulación con MCI. IL-1β fue la responsable de estas inducciones en A549 y TNF-α
en las células bronquiales.
Además de su función inmunomoduladora, ambas CCL20 y hBD2 han demostrado
tener actividad antimicrobiana contra ciertas bacterias Gram positivas y negativas
(Yang et al. 2003; Harder et al. 2000). Sin embargo, la actividad antimicrobiana contra
B. abortus nunca fue testeada. Encontramos que CCL20 tiene actividad microbicida in
vitro con una DL50 mayor a 50 µg, siendo ampliamente mayor que la DL50 para E. coli
(1.5 µg/ml). La DL para B. abortus fue significativamente superior que aquella
reportada para distintas bacterias incluyendo: Pseudomonas aeruginosa, Moraxella
catarrhalis, Enterococcus faecium, Staphylococcus aureus o Streptococcus pyogenes, las
cuales se encuentran en un rango de 0.2 a 10 µg/ml(Yang et al. 2003). Por lo tanto, B.
abortus parece resistente a la acción antimicrobiana de CCL20, comparativamente a la
sensibilidad de otras bacterias.

Capítulo 3 - Discusión
132
Como las células epiteliales pulmonares estimuladas con MCI produjeron hBD2,
fue de interés probar la acción antimicrobiana de esta defensina frente a B. abortus.
Mientras que las DL50 para P. aeruginosa y E. coli se encuentran alrededor de los 5
µg/ml (Harder et al. 2000), B. abortus resistió hasta 50 µg/ml de hBD2 y hBD3. Esto
concuerda con otros reportes donde se probaron otros péptidos antimicrobianos de
diferentes familias, como la polimixina B, la melitina (abejas), magainina (piel de
sapo) y cecropina (cerdo)(Halling 1996).
Los niveles de hBD2 y CCL20 secretados por las células epiteliales pulmonares en
respuesta a la infección por B. abortus, ya sea por infección directa o indirectamente a
través de la estimulación con MCI, son mucho más bajos que la DL50 de ambos
péptidos antimicrobianos para esta bacteria. Estos resultados sugieren que el rol de
estas moléculas durante la infección sería el de quimioatractantes y no el de
antimicrobianos.
La resistencia de ambas cepas lisas de Brucella a estos péptidos antimicrobianos
ha sido atribuida al lipopolisacárido O en el LPS de la bacteria, debido a que las cepas
lisas son más resistentes a los péptidos catiónicos (polimixina, cecropina, melitina)
que las cepas rugosas (De Tejada et al. 1995). Cuando se ensayó hBD2, no se observo
sensibilidad diferencial para la cepa rugosa, mientras que hBD3 logró ejercer un
efecto microbicida a concentraciones de 50 µg/ml.
En conjunto, estos resultados muestran que las células epiteliales humanas
secretan CCL20 y hBD2 en respuesta a la infección por B. abortus o a las citoquinas
producidas por monocitos infectados. Mientras que estas moléculas son producidas en
concentraciones que no parecen tener actividad antimicrobiana, podrían estar
involucradas en funciones inmunomoduladoras durante la infección con Brucella
abortus, como el reclutamiento y la activación de monocitos y otras células inmunes.

133
Conclusiones

Conclusiones
134
Capitulo 1:
La respuesta inmune innata contra Brucella en los primeros días p.i. es de
carácter levemente proinflamatorio, y se monta cuando ya han sido
colonizados los principales órganos periféricos
Las proteínas Btp de B. abortus modulan el reclutamiento de las células
inmunes hacia el pulmón y contribuyen a la colonización de órganos
periféricos (bazo e hígado) desde pulmón.
B. abortus puede modular la respuesta inmune de células pulmonares
fagocíticas y no fagocíticas (células epiteliales alveolares, macrófagos
alveolares y células dendríticas)
B. abortus, a través de sus proteínas Btp, ejerce algún grado de modulación
inmune disminuyendo el reclutamiento de células y la producción de KC
inducidos por el LPS de E. coli en pulmón
La capacidad de sobrevida de B. abortus en el ambiente pulmonar previamente
inflamado con LPS de E. coli depende de las proteínas Btp.
Capítulo 2:
Las células epiteliales alveolares humanas son pobres respondedoras frente a
la infección directa por B. abortus en cuanto a secreción de citoquinas
proinflamatorias, al igual que los explantos pulmonares, si bien estos últimos
responden con mayores niveles de citoquinas.
Brucella es capaz de atravesar por vía paracelular una monocapa de
pneumocitos humanos polarizados

Conclusiones
135
Capítulo 3:
Las células epiteliales humanas secretan CCL20 y beta-defensina 2 en
respuesta a la infección por B. abortus o a las citoquinas producidas por
monocitos infectados.
B. abortus es resistente a la acción antimicrobiana in vitro de CCL20, hBD2 y
hBD3 a dosis mucho mayores que las producidas por el epitelio infectado o
estimulado.
Conclusión general
En conclusión, la infección inhalatoria por Brucella abortus genera una respuesta
innata pulmonar de poca intensidad que le permite sobrevivir en pulmón y
probablemente facilita su diseminación a otros órganos. Es probable que el pasaje de
la bacteria a la circulación sistémica ocurra, al menos en parte, por pasaje paracelular
a través del epitelio alveolar. La persistencia pulmonar de la bacteria posiblemente se
vea facilitada, entre otros factores, por su capacidad de supervivencia intracelular y
sus mecanismos de modulación de la respuesta inmune innata

136
Bibliografía

Bibliografía
137
Akira, S., Uematsu, S. & Takeuchi, O., 2006. Pathogen recognition and innate immunity. Cell, 124(4), pp.783–801.
Alekseeva, L. et al., 2009. Inducible expression of beta defensins by human respiratory epithelial cells exposed to Aspergillus fumigatus organisms. BMC microbiology, 9, p.33.
Alexopoulou, L. et al., 2001. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature, 413(6857), pp.732–8.
Anderson, T.D. & Cheville, N.F., 1986. Ultrastructural morphometric analysis of Brucella abortus-infected trophoblasts in experimental placentitis. Bacterial replication occurs in rough endoplasmic reticulum. The American journal of pathology, 124(2), pp.226–37.
Antoniades, H.N. et al., 1992. Expression of monocyte chemoattractant protein 1 mRNA in human idiopathic pulmonary fibrosis. Proceedings of the National Academy of Sciences of the United States of America, 89(12), pp.5371–5.
Archambaud, C. et al., 2010. Contrasting roles of macrophages and dendritic cells in controlling initial pulmonary Brucella infection. European Journal of Immunology, 40(12), pp.3458–3471.
Arellano-Reynoso, B. et al., 2005. Cyclic beta-1,2-glucan is a Brucella virulence factor required for intracellular survival. Nature immunology, 6(6), pp.618–25.
Arnold, R. et al., 1994. Interleukin-8, interleukin-6, and soluble tumour necrosis factor receptor type I release from a human pulmonary epithelial cell line (A549) exposed to respiratory syncytial virus. Immunology, 82(1), pp.126–33.
Badley, A.D. et al., 1997. Macrophage-dependent apoptosis of CD4+ T lymphocytes from HIV-infected individuals is mediated by FasL and tumor necrosis factor. The Journal of experimental medicine, 185(1), pp.55–64.
Baggetta, R. et al., 2010. The interferon-inducible gene IFI16 secretome of endothelial cells drives the early steps of the inflammatory response. European journal of immunology, 40(8), pp.2182–9.
Baggiolini, M., Walz, A. & Kunkel, S.L., 1989. Neutrophil-activating peptide-1/interleukin 8, a novel cytokine that activates neutrophils. The Journal of clinical investigation, 84(4), pp.1045–9.
Bals, R., 2000. Epithelial antimicrobial peptides in host defense against infection. Respiratory research, 1(3), pp.141–150.
Bals, R. et al., 2004. Isolation and air-liquid interface culture of human large airway and bronchiolar epithelial cells. Journal of cystic fibrosis : official journal of the European Cystic Fibrosis Society, 3 Suppl 2, pp.49–51.
Barabas, N. et al., 2013. Beta-defensins activate macrophages and synergize in pro-inflammatory cytokine expression induced by TLR ligands. Immunobiology, 218(7), pp.1005–11.
Barquero-Calvo, E. et al., 2007. Brucella abortus uses a stealthy strategy to avoid activation of the innate immune system during the onset of infection. PloS one, 2(7), p.e631.
Bauernfeind, F. & Hornung, V., 2013. Of inflammasomes and pathogens--sensing of microbes by the inflammasome. EMBO molecular medicine, 5(6), pp.814–26.
Bazzoni, F. et al., 1991. Phagocytosing neutrophils produce and release high amounts of the neutrophil-activating peptide 1/interleukin 8. The Journal of experimental medicine, 173(3), pp.771–4.

Bibliografía
138
Beachey, E.H. & Courtney, H.S., 1989. Bacterial adherence of group A streptococci to mucosal surfaces. Respiration; international review of thoracic diseases, 55 Suppl 1, pp.33–40.
Becker, S. et al., 2005. Regulation of cytokine production in human alveolar macrophages and airway epithelial cells in response to ambient air pollution particles: Further mechanistic studies. Toxicology and Applied Pharmacology, 207(2 SUPPL.), pp.269–275.
Bhattacharjee, A.K. et al., 2006. Comparison of protective efficacy of subcutaneous versus intranasal immunization of mice with a Brucella melitensis lipopolysaccharide subunit vaccine. Infection and immunity, 74(10), pp.5820–5.
Birkness, K.A. et al., 1999. An in vitro tissue culture bilayer model to examine early events in Mycobacterium tuberculosis infection. Infection and immunity, 67(2), pp.653–8.
Blussé van Oud Alblas, A., van der Linden-Schrever, B. & van Furth, R., 1981. Origin and kinetics of pulmonary macrophages during an inflammatory reaction induced by intravenous administration of heat-killed bacillus Calmette-Guérin. The Journal of experimental medicine, 154(2), pp.235–52.
Boschiroli, M.L. et al., 2002. Type IV secretion and Brucella virulence. Veterinary microbiology, 90(1-4), pp.341–8.
Bosio, C.M., Bielefeldt-Ohmann, H. & Belisle, J.T., 2007. Active suppression of the pulmonary immune response by Francisella tularensis Schu4. Journal of immunology (Baltimore, Md. : 1950), 178(7), pp.4538–4547.
Bossi, P. et al., 2004. Bichat clinical guidelines for bioterrorist agents. Euro sur eillance : bulletin Europ en sur les maladies transmissibles = European communicable disease bulletin, 9(12), pp.E1–2.
Bouza, E. et al., 2005. Laboratory-acquired brucellosis: a Spanish national survey. The Journal of hospital infection, 61(1), pp.80–83.
Bowden, R.A. et al., 1998. Evaluation of immunogenicity and protective activity in BALB/c mice of the 25-kDa major outer-membrane protein of Brucella melitensis (Omp25) expressed in Escherichia coli. Journal of medical microbiology, 47(1), pp.39–48.
Bricker, B.J. et al., 1990. Cloning, expression, and occurrence of the Brucella Cu-Zn superoxide dismutase. Infection and immunity, 58(9), pp.2935–9.
Brieland, J. et al., 1994. Replicative Legionella pneumophila lung infection in intratracheally inoculated A/J mice. A murine model of human Legionnaires’ disease. The American journal of pathology, 145(6), pp.1537–1546.
Brightbill, H.D. & Modlin, R.L., 2000. Toll-like receptors: molecular mechanisms of the mammalian immune response. Immunology, 101(1), pp.1–10.
Briones, G. et al., 2001. Brucella abortus cyclic beta-1,2-glucan mutants have reduced virulence in mice and are defective in intracellular replication in HeLa cells. Infection and immunity, 69(7), pp.4528–35.
Car, B.D. et al., 1994. Elevated IL-8 and MCP-1 in the bronchoalveolar lavage fluid of patients with idiopathic pulmonary fibrosis and pulmonary sarcoidosis. American journal of respiratory and critical care medicine, 149(3 Pt 1), pp.655–9.

Bibliografía
139
Celli, J. et al., 2003. Brucella evades macrophage killing via VirB-dependent sustained interactions with the endoplasmic reticulum. The Journal of experimental medicine, 198(4), pp.545–56.
Chan, D.I. et al., 2008. Human macrophage inflammatory protein 3alpha: protein and peptide nuclear magnetic resonance solution structures, dimerization, dynamics, and anti-infective properties. Antimicrobial agents and chemotherapy, 52(3), pp.883–94.
Chen, K.W. & Schroder, K., 2013. Antimicrobial functions of inflammasomes. Current opinion in microbiology, 16(3), pp.311–8.
Christie, P.J., 2001. Type IV secretion: intercellular transfer of macromolecules by systems ancestrally related to conjugation machines. Molecular microbiology, 40(2), pp.294–305.
Cloeckaert, A. et al., 1992. Demonstration of peptidoglycan-associated Brucella outer-membrane proteins by use of monoclonal antibodies. Journal of general microbiology, 138(7), pp.1543–50.
Cloeckaert, A. et al., 2002. Major outer membrane proteins of Brucella spp.: past, present and future. Veterinary microbiology, 90(1-4), pp.229–47.
Cloeckaert, A. et al., 1996. Molecular and immunological characterization of the major outer membrane proteins of Brucella. FEMS microbiology letters, 145(1), pp.1–8.
Comerci, D.J. et al., 2001. Essential role of the VirB machinery in the maturation of the Brucella abortus-containing vacuole. Cellular microbiology, 3(3), pp.159–68.
Corbeil, L.B. et al., 1988. Killing of Brucella abortus by bovine serum. Infection and immunity, 56(12), pp.3251–61.
Crespo León F. 1993. Brucelosis ovina y caprina. O.I.E.
Cunningham, A.C. et al., 1994. Constitutive expression of MHC and adhesion molecules by alveolar epithelial cells (type II pneumocytes) isolated from human lung and comparison with immunocytochemical findings. Journal of cell science, 107 ( Pt 2, pp.443–9.
Van Damme, J. et al., 1994. Induction of monocyte chemotactic proteins MCP-1 and MCP-2 in human fibroblasts and leukocytes by cytokines and cytokine inducers. Chemical synthesis of MCP-2 and development of a specific RIA. Journal of immunology (Baltimore, Md. : 1950), 152(11), pp.5495–502.
Davos, D.E. et al., 1981. Outbreak of brucellosis at a South-Australian abattoir. 2. Epidemiological investigations. The Medical journal of Australia, 2(12-13), pp.657–60.
Delpino, M.V., Fossati, C.A. & Baldi, P.C., 2009. Proinflammatory response of human osteoblastic cell lines and osteoblast-monocyte interaction upon infection with Brucella spp. Infection and immunity, 77(3), pp.984–95.
Delrue, R.M. et al., 2001. Identification of Brucella spp. genes involved in intracellular trafficking. Cellular microbiology, 3(7), pp.487–97.
Deng, H. et al., 2013. Molecular mechanism responsible for the priming of macrophage activation. The Journal of biological chemistry, 288(6), pp.3897–906.
Detilleux, P.G., Deyoe, B.L. & Cheville, N.F., 1990a. Entry and intracellular localization of Brucella spp. in Vero cells: fluorescence and electron microscopy. Veterinary pathology, 27(5), pp.317–28.

Bibliografía
140
Detilleux, P.G., Deyoe, B.L. & Cheville, N.F., 1990b. Penetration and intracellular growth of Brucella abortus in nonphagocytic cells in vitro. Infection and immunity, 58(7), pp.2320–8.
Diamond, G., Legarda, D. & Ryan, L.K., 2000. The innate immune response of the respiratory epithelium. Immunological reviews, 173, pp.27–38.
Dockrell, D.H. et al., 2001. Immune-mediated phagocytosis and killing of Streptococcus pneumoniae are associated with direct and bystander macrophage apoptosis. The Journal of infectious diseases, 184(6), pp.713–22.
Doss, M. et al., 2010. Human defensins and LL-37 in mucosal immunity. Journal of leukocyte biology, 87(1), pp.79–92.
Douglas, J.T. et al., 1984. Porins of Brucella species. Infection and immunity, 44(1), pp.16–21.
Driscoll, K.E. et al., 2000. Intratracheal instillation as an exposure technique for the evaluation of respiratory tract toxicity: uses and limitations. Toxicological sciences : an official journal of the Society of Toxicology, 55(1), pp.24–35.
Duff, C. et al., Differences in invasion and translocation of Burkholderia cepacia complex species in polarised lung epithelial cells in vitro. Microbial pathogenesis, 41(4-5), pp.183–92.
Duits, L. a. et al., 2003. Rhinovirus increases human ??-defensin-2 and -3 mRNA expression in cultured bronchial epithelial cells. FEMS Immunology and Medical Microbiology, 38(1), pp.59–64.
Durward, M. et al., 2012. Active evasion of CTL mediated killing and low quality responding CD8+ T cells contribute to persistence of brucellosis. PloS one, 7(4), p.e34925.
Elberts, K. et al., 1999. Monolayers of Human Alveolar Epithelial Cells in Primary Culture for Pulmonary Absorption and Transport Studies. Pharmaceutical Research, 16(5), pp.601–608.
Elzer, P.H. et al., 1996. The HtrA stress response protease contributes to resistance of Brucella abortus to killing by murine phagocytes. Infection and immunity, 64(11), pp.4838–41.
Errea, a. et al., 2010. Mucosal innate response stimulation induced by lipopolysaccharide protects against Bordetella pertussis colonization. Medical Microbiology and Immunology, 199(2), pp.103–108.
Evans, S.E. et al., 2010. Stimulated innate resistance of lung epithelium protects mice broadly against bacteria and fungi. American journal of respiratory cell and molecular biology, 42(1), pp.40–50.
de Fays, K. et al., 1999. Structure and function prediction of the Brucella abortus P39 protein by comparative modeling with marginal sequence similarities. Protein engineering, 12(3), pp.217–23.
Fernandez-Prada, C.M. et al., 2001. Deletion of wboA enhances activation of the lectin pathway of complement in Brucella abortus and Brucella melitensis. Infection and immunity, 69(7), pp.4407–16.
Ferrero, M.C. et al., 2014. Key role of toll-like receptor 2 in the inflammatory response and major histocompatibility complex class ii downregulation in brucella abortus-infected alveolar macrophages. Infection and Immunity, 82(2), pp.626–639.
Ferrero, M.C. et al., 2011. Proinflammatory response of human endothelial cells to Brucella infection. Microbes and infection / Institut Pasteur, 13(10), pp.852–61.

Bibliografía
141
Ferrero, M.C., Fossati, C. a. & Baldi, P.C., 2010. Direct and monocyte-induced innate immune response of human lung epithelial cells to Brucella abortus infection. Microbes and Infection, 12(10), pp.736–747.
Ferrero, M.C., Fossati, C. a. & Baldi, P.C., 2009. Smooth Brucella strains invade and replicate in human lung epithelial cells without inducing cell death. Microbes and Infection, 11(4), pp.476–483.
Le Feuvre, R.A. et al., 2002. Priming of macrophages with lipopolysaccharide potentiates P2X7-mediated cell death via a caspase-1-dependent mechanism, independently of cytokine production. The Journal of biological chemistry, 277(5), pp.3210–8.
Foulongne, V. et al., 2000. Identification of Brucella suis genes affecting intracellular survival in an in vitro human macrophage infection model by signature-tagged transposon mutagenesis. Infection and immunity, 68(3), pp.1297–303.
Franke-Ullmann, G. et al., 1996. Characterization of murine lung interstitial macrophages in comparison with alveolar macrophages in vitro. Journal of immunology (Baltimore, Md. : 1950), 157(7), pp.3097–104.
Fritz, J.H. et al., 2006. Nod-like proteins in immunity, inflammation and disease. Nature immunology, 7(12), pp.1250–7.
Fuchs, S. et al., 2003. Differentiation of human alveolar epithelial cells in primary culture: Morphological characterization and synthesis of caveolin-1 and surfactant protein-C. Cell and Tissue Research, 311(1), pp.31–45.
Geme, J.W., 1996. Molecular determinants of the interaction between Haemophilus influenzae and human cells. American journal of respiratory and critical care medicine, 154(4 Pt 2), pp.S192–6.
Gentry, M. et al., 2007. Role of primary human alveolar epithelial cells in host defense against Francisella tularensis infection. Infection and immunity, 75(8), pp.3969–78.
Gern, J.E. et al., 2003. Double-stranded RNA induces the synthesis of specific chemokines by bronchial epithelial cells. American journal of respiratory cell and molecular biology, 28(6), pp.731–7.
Ghannam, S. et al., 2011. CCL20 and β-defensin-2 induce arrest of human Th17 cells on inflamed endothelium in vitro under flow conditions. Journal of immunology (Baltimore, Md. : 1950), 186(3), pp.1411–20.
Giambartolomei, G.H. et al., 2004. Lipoproteins, not lipopolysaccharide, are the key mediators of the proinflammatory response elicited by heat-killed Brucella abortus. Journal of immunology (Baltimore, Md. : 1950), 173(7), pp.4635–4642.
Gieffers, J. et al., 2004. Phagocytes transmit Chlamydia pneumoniae from the lungs to the vasculature. The European respiratory journal, 23(4), pp.506–10.
Gil-Ramírez, Y. et al., 2014. The identification of wadB, a new glycosyltransferase gene, confirms the branched structure and the role in virulence of the lipopolysaccharide core of Brucella abortus. Microbial Pathogenesis, 73, pp.53–59.
Godfroid, J. et al., 2005. From the discovery of the Malta fever’s agent to the discovery of a marine mammal reservoir, brucellosis has continuously been a re-emerging zoonosis. Veterinary research, 36(3), pp.313–26.
Goldbaum, F.A. et al., 1992. Differentiation between active and inactive human brucellosis by measuring

Bibliografía
142
antiprotein humoral immune responses. Journal of clinical microbiology, 30(3), pp.604–7.
Goldstein, J. et al., 1992. Lipopolysaccharide (LPS) from Brucella abortus is less toxic than that from Escherichia coli, suggesting the possible use of B. abortus or LPS from B. abortus as a carrier in vaccines. Infection and immunity, 60(4), pp.1385–9.
Gomes, M.T.R. et al., 2013. Critical role of ASC inflammasomes and bacterial type IV secretion system in caspase-1 activation and host innate resistance to Brucella abortus infection. Journal of immunology (Baltimore, Md. : 1950), 190(7), pp.3629–38.
Gómez-Miguel, M.J., Moriyón, I. & López, J., 1987. Brucella outer membrane lipoprotein shares antigenic determinants with Escherichia coli Braun lipoprotein and is exposed on the cell surface. Infection and immunity, 55(1), pp.258–62.
Grahnert, A. et al., 2014. IL-4 receptor-alpha-dependent control of Cryptococcus neoformans in the early phase of pulmonary infection. PloS one, 9(1), p.e87341.
Grillá, M.J. et al., 2012. What have we learned from brucellosis in the mouse model? Veterinary Research, 43(1), pp.1–35.
Gutbier, B. et al., 2015. Moraxella catarrhalis induces an immune response in the murine lung which is independent of human CEACAM5 expression and long-term smoke exposure. American Journal of Physiology - Lung Cellular and Molecular Physiology, p.ajplung.00265.2014.
Halling, S.M., 1996. The effects of magainin 2, cecropin, mastoparan and melittin on Brucella abortus. Veterinary Microbiology, 51(1-2), pp.187–192.
Harder, J. et al., 2000. Mucoid Pseudomonas aeruginosa, TNF-alpha, and IL-1beta, but not IL-6, induce human beta-defensin-2 in respiratory epithelia. American journal of respiratory cell and molecular biology, 22(6), pp.714–21.
Herold, S. et al., 2006. Alveolar epithelial cells direct monocyte transepithelial migration upon influenza virus infection: impact of chemokines and adhesion molecules. Journal of immunology (Baltimore, Md. : 1950), 177(3), pp.1817–24.
Hirota, K. et al., 2007. Preferential recruitment of CCR6-expressing Th17 cells to inflamed joints via CCL20 in rheumatoid arthritis and its animal model. The Journal of experimental medicine, 204(12), pp.2803–12.
Holgate, S.T. et al., 2000. Bronchial epithelium as a key regulator of airway allergen sensitization and remodeling in asthma. American journal of respiratory and critical care medicine, 162(3 Pt 2), pp.S113–7.
Holt, P.G. et al., 1993. Downregulation of the antigen presenting cell function(s) of pulmonary dendritic cells in vivo by resident alveolar macrophages. The Journal of experimental medicine, 177(2), pp.397–407.
Holt, P.G., 1978. Inhibitory activity of unstimulated alveolar macrophages on T-lymphocyte blastogenic response. The American review of respiratory disease, 118(4), pp.791–3.
Homma, T. et al., 2004. Corticosteroid and cytokines synergistically enhance toll-like receptor 2 expression in respiratory epithelial cells. American journal of respiratory cell and molecular biology, 31(4), pp.463–9.
Hong, P.C., Tsolis, R.M. & Ficht, T.A., 2000. Identification of genes required for chronic persistence of

Bibliografía
143
Brucella abortus in mice. Infection and immunity, 68(7), pp.4102–7.
Hosokawa, Y. et al., 2005. Increase of CCL20 expression by human gingival fibroblasts upon stimulation with cytokines and bacterial endotoxin. Clinical and experimental immunology, 142(2), pp.285–91.
Huddleson, I.F. & Munger, M., 1940. A Study of an Epidemic of Brucellosis Due to Brucella melitensis. American journal of public health and the nation’s health, 30(8), pp.944–54.
Hurley, B.P., Williams, N.L. & McCormick, B.A., 2006. Involvement of phospholipase A2 in Pseudomonas aeruginosa-mediated PMN transepithelial migration. American journal of physiology. Lung cellular and molecular physiology, 290(4), pp.L703–L709.
Ibañez, A.E. et al., 2013. Unlipidated Outer Membrane Protein Omp16 (U-Omp16) from Brucella spp. as Nasal Adjuvant Induces a Th1 Immune Response and Modulates the Th2 Allergic Response to Cow’s Milk Proteins. PLoS ONE, 8(7), pp.1–13.
Ishii, H. et al., 2005. Alveolar macrophage-epithelial cell interaction following exposure to atmospheric particles induces the release of mediators involved in monocyte mobilization and recruitment. Respiratory research, 6, p.87.
Ito, T. et al., 2011. CCR6 as a mediator of immunity in the lung and gut. Experimental cell research, 317(5), pp.613–9.
Iyonaga, K. et al., 1994. Monocyte chemoattractant protein-1 in idiopathic pulmonary fibrosis and other interstitial lung diseases. Human pathology, 25(5), pp.455–63.
Jäger, J. et al., 2014. Human lung tissue explants reveal novel interactions during Legionella pneumophila infections. Infection and Immunity, 82(1), pp.275–285.
Jennings, J.H. et al., 2005. Monocytes recruited to the lungs of mice during immune inflammation ingest apoptotic cells poorly. American journal of respiratory cell and molecular biology, 32(2), pp.108–17.
Jiang, X. et al., 1993. Macrophage control of Brucella abortus: role of reactive oxygen intermediates and nitric oxide. Cellular immunology, 151(2), pp.309–19.
Jubier-Maurin, V. et al., 2001. Major outer membrane protein Omp25 of Brucella suis is involved in inhibition of tumor necrosis factor alpha production during infection of human macrophages. Infection and immunity, 69(8), pp.4823–30.
Kahl-McDonagh, M.M., Arenas-Gamboa, a. M. & Ficht, T. a., 2007. Aerosol infection of BALB/c mice with Brucella melitensis and Brucella abortus and protective efficacy against aerosol challenge. Infection and Immunity, 75(10), pp.4923–4932.
Kaiser, V. & Diamond, G., 2000. Expression of mammalian defensin genes. Journal of leukocyte biology, 68(6), pp.779–784.
Kaufmann, a F. et al., 1980. Airborne spread of brucellosis. Annals of the New York Academy of Sciences, 353, pp.105–114.
Khan, M.A. et al., 2008. Flagellin-dependent and -independent inflammatory responses following infection by enteropathogenic Escherichia coli and Citrobacter rodentium. Infection and immunity, 76(4), pp.1410–22.
Kirby, A.C., Coles, M.C. & Kaye, P.M., 2009. Alveolar macrophages transport pathogens to lung draining

Bibliografía
144
lymph nodes. Journal of immunology (Baltimore, Md. : 1950), 183(3), pp.1983–1989.
Klein-Patel, M.E. et al., 2006. Inhibition of beta-defensin gene expression in airway epithelial cells by low doses of residual oil fly ash is mediated by vanadium. Toxicological sciences : an official journal of the Society of Toxicology, 92(1), pp.115–25.
Ko, J. & Splitter, G.A., 2003. Molecular host-pathogen interaction in brucellosis: current understanding and future approaches to vaccine development for mice and humans. Clinical microbiology reviews, 16(1), pp.65–78.
Köhler, S. et al., 2003. What is the nature of the replicative niche of a stealthy bug named Brucella? Trends in microbiology, 11(5), pp.215–9.
Korpi-Steiner, N.L. et al., 2010. Human monocytic cells direct the robust release of CXCL10 by bronchial epithelial cells during rhinovirus infection. Clinical and experimental allergy : journal of the British Society for Allergy and Clinical Immunology, 40(8), pp.1203–13.
Koyama, S. et al., 2000. The potential of various lipopolysaccharides to release IL-8 and G-CSF. American journal of physiology. Lung cellular and molecular physiology, 278(4), pp.L658–66.
Lai, Y. & Gallo, R.L., 2009. AMPed up immunity: how antimicrobial peptides have multiple roles in immune defense. Trends in Immunology, 30(3), pp.131–141.
Lamkanfi, M. & Dixit, V.M., 2012. Inflammasomes and their roles in health and disease. Annual review of cell and developmental biology, 28, pp.137–61.
Lapaque, N. et al., 2006. Differential inductions of TNF-?? and IGTP, IIGP by structurally diverse classic and non-classic lipopolysaccharides. Cellular Microbiology, 8(3), pp.401–413.
Lee, M.S. & Kim, Y.-J., 2007. Pattern-recognition receptor signaling initiated from extracellular, membrane, and cytoplasmic space. Molecules and cells, 23(1), pp.1–10.
Lemon, J.K., Miller, M.R. & Weiser, J.N., 2015. Sensing of IL-1 cytokines during Streptococcus pneumoniae colonization contributes to macrophage recruitment and bacterial clearance. Infection and Immunity, (June), pp.IAI.00224–15.
Lestrate, P. et al., 2000. Identification and characterization of in vivo attenuated mutants of Brucella melitensis. Molecular microbiology, 38(3), pp.543–51.
Li, Y., Wang, Y. & Liu, X., 2012. The Role of Airway Epithelial Cells in Response to Mycobacteria Infection. Clinical and Developmental Immunology, 2012, pp.1–11.
Liu, H. et al., 2001. Constitutively activated Akt-1 is vital for the survival of human monocyte-differentiated macrophages. Role of Mcl-1, independent of nuclear factor (NF)-kappaB, Bad, or caspase activation. The Journal of experimental medicine, 194(2), pp.113–26.
Liu, P. et al., 2007. Retinoic acid-inducible gene I mediates early antiviral response and Toll-like receptor 3 expression in respiratory syncytial virus-infected airway epithelial cells. Journal of virology, 81(3), pp.1401–11.
Van Maele, L. et al., 2014. Airway structural cells regulate TLR5-mediated mucosal adjuvant activity. Mucosal immunology, 7(3), pp.489–500.
Marchesini, M.I. et al., 2011. In search of Brucella abortus type iv secretion substrates: Screening and identification of four proteins translocated into host cells through virb system. Cellular

Bibliografía
145
Microbiology, 13(8), pp.1261–1274.
Marquis, H. & Ficht, T.A., 1993. The omp2 gene locus of Brucella abortus encodes two homologous outer membrane proteins with properties characteristic of bacterial porins. Infection and immunity, 61(9), pp.3785–90.
Marriott, H.M. et al., 2012. Interleukin-1β regulates CXCL8 release and influences disease outcome in response to Streptococcus pneumoniae, defining intercellular cooperation between pulmonary epithelial cells and macrophages. Infection and immunity, 80(3), pp.1140–9.
Marriott, H.M. & Dockrell, D.H., 2007. The role of the macrophage in lung disease mediated by bacteria. Experimental lung research, 33(10), pp.493–505.
Martin-Mazuelos, E. et al., 1994. Outbreak of Brucella melitensis among microbiology laboratory workers. Journal of clinical microbiology, 32(8), pp.2035–6.
Mayer, A.K. et al., 2007. Differential recognition of TLR-dependent microbial ligands in human bronchial epithelial cells. Journal of immunology (Baltimore, Md. : 1950), 178(5), pp.3134–42.
Mayer, A.K. & Dalpke, A.H., 2007. Regulation of local immunity by airway epithelial cells. Archivum Immunologiae et Therapiae Experimentalis, 55(6), pp.353–362.
Medzhitov, R. & Janeway, C., 2000. Innate immune recognition: mechanisms and pathways. Immunological reviews, 173, pp.89–97.
Medzhitov, R. & Janeway, C.A., 1997. Innate immunity: impact on the adaptive immune response. Current opinion in immunology, 9(1), pp.4–9.
Memish, Z.A. & Mah, M.W., 2001. Brucellosis in laboratory workers at a Saudi Arabian hospital. American journal of infection control, 29(1), pp.48–52.
Mense, M.G. et al., 2001. Bacteriologic and histologic features in mice after intranasal inoculation of Brucella melitensis. American journal of veterinary research, 62(3), pp.398–405.
Mense, M.G. et al., 2004. Pathologic changes associated with brucellosis experimentally induced by aerosol exposure in rhesus macaques (Macaca mulatta). American Journal of Veterinary Research, 65(5), pp.644–652.
Miao, H. et al., 2012. FOXO1 increases CCL20 to promote NF-κB-dependent lymphocyte chemotaxis. Molecular endocrinology (Baltimore, Md.), 26(3), pp.423–37.
Miller, C.D., Songer, J.R. & Sullivan, J.F., 1987. A twenty-five year review of laboratory-acquired human infections at the National Animal Disease Center. American Industrial Hygiene Association journal, 48(3), pp.271–5.
Mobasheri, H. et al., 1997. Brucella Omp2a and Omp2b porins: single channel measurements and topology prediction. FEMS microbiology letters, 155(1), pp.23–30.
Montejo, J.M. et al., 1993. Open, randomized therapeutic trial of six antimicrobial regimens in the treatment of human brucellosis. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America, 16(5), pp.671–6.
Montes, J. et al., 1986. Laboratory-acquired meningitis caused by Brucella abortus strain 19. The Journal of infectious diseases, 154(5), pp.915–6.

Bibliografía
146
Moranta, D. et al., 2010. Klebsiella pneumoniae capsule polysaccharide impedes the expression of β-defensins by airway epithelial cells. Infection and Immunity, 78(3), pp.1135–1146.
Moreno, E. & Moriyon, I., 2002. Brucella melitensis: a nasty bug with hidden credentials for virulence. Proceedings of the National Academy of Sciences of the United States of America, 99(1), pp.1–3.
Moreno-Lafont, M.C., López-Merino, A. & López-Santiago, R., 1995. Cell response to a salt-extractable and sonicated Brucella melitensis 16M antigen in human brucellosis. Clinical and diagnostic laboratory immunology, 2(3), pp.377–80.
Munder, A. et al., 2002. Pulmonary microbial infection in mice: comparison of different application methods and correlation of bacterial numbers and histopathology. Experimental and toxicologic pathology : official journal of the Gesellschaft für Toxikologische Pathologie, 54(2), pp.127–33.
Murphy, E. a. et al., 2001. Immune control of Brucella abortus 2308 infections in BALB/c mice. FEMS Immunology and Medical Microbiology, 32(1), pp.85–88.
Murphy, S. a. et al., 1999. A comparative study of the isolation of type II epithelial cells from rat, hamster, pig and human lung tissue. Methods in Cell Science, 21(1), pp.31–38.
Myeni, S. et al., 2013. Brucella modulates secretory trafficking via multiple type IV secretion effector proteins. PLoS pathogens, 9(8), p.e1003556.
Nelson, K.E., Ruben, F.L. & Andersen, B., 1975. An unusual outbreak of brucellosis. Archives of internal medicine, 135(5), pp.691–5.
Nicholas, B. et al., 2015. A Novel Lung Explant Model for the Ex Vivo Study of Efficacy and Mechanisms of Anti-Influenza Drugs. The Journal of Immunology, 194(12), pp.6144–6154.
Niyonsaba, F. et al., 2002. Epithelial cell-derived human beta-defensin-2 acts as a chemotaxin for mast cells through a pertussis toxin-sensitive and phospholipase C-dependent pathway. International immunology, 14(4), pp.421–6.
Niyonsaba, F., Ogawa, H. & Nagaoka, I., 2004. Human beta-defensin-2 functions as a chemotactic agent for tumour necrosis factor-alpha-treated human neutrophils. Immunology, 111(3), pp.273–81.
O’Callaghan, D. et al., 1999. A homologue of the Agrobacterium tumefaciens VirB and Bordetella pertussis Ptl type IV secretion systems is essential for intracellular survival of Brucella suis. Molecular microbiology, 33(6), pp.1210–20.
Ollé-Goig, J.E. & Canela-Soler, J., 1987. An outbreak of Brucella melitensis infection by airborne transmission among laboratory workers. American journal of public health, 77(3), pp.335–8.
Opitz, B. et al., 2007. IFNbeta induction by influenza A virus is mediated by RIG-I which is regulated by the viral NS1 protein. Cellular microbiology, 9(4), pp.930–8.
Opitz, B. et al., 2004. Nucleotide-binding oligomerization domain proteins are innate immune receptors for internalized Streptococcus pneumoniae. The Journal of biological chemistry, 279(35), pp.36426–32.
Osterholzer, J.J. et al., 2005. CCR2 and CCR6, but not endothelial selectins, mediate the accumulation of immature dendritic cells within the lungs of mice in response to particulate antigen. Journal of immunology (Baltimore, Md. : 1950), 175(2), pp.874–83.
Pappas, G. et al., 2006. Brucella as a biological weapon. Cellular and molecular life sciences : CMLS,

Bibliografía
147
63(19-20), pp.2229–36.
Pappas, G. et al., 2003. Brucellosis and the respiratory system. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America, 37(7), pp.e95–e99.
Park, C.-S. et al., 2010. Chlamydophila pneumoniae enhances secretion of VEGF, TGF-beta and TIMP-1 from human bronchial epithelial cells under Th2 dominant microenvironment. Allergy, asthma & immunology research, 2(1), pp.41–7.
Pasquevich, K.A. et al., 2009. Immunization with recombinant Brucella species outer membrane protein Omp16 or Omp19 in adjuvant induces specific CD4+ and CD8+ T cells as well as systemic and oral protection against Brucella abortus infection. Infection and immunity, 77(1), pp.436–45.
Passalacqua, K.D. & Bergman, N.H., 2006. Bacillus anthracis: interactions with the host and establishment of inhalational anthrax. Future microbiology, 1(4), pp.397–415.
Pazgier, M. et al., 2006. Human beta-defensins. Cellular and molecular life sciences : CMLS, 63(11), pp.1294–313.
Phadke, A.P. et al., 2007. The role of CC chemokine receptor 6 in host defense in a model of invasive pulmonary aspergillosis. American journal of respiratory and critical care medicine, 175(11), pp.1165–72.
Pichavant, M. et al., 2006. Impact of bronchial epithelium on dendritic cell migration and function: modulation by the bacterial motif KpOmpA. Journal of immunology (Baltimore, Md. : 1950), 177(9), pp.5912–9.
Pizarro-Cerdá, J., Méresse, S., et al., 1998. Brucella abortus transits through the autophagic pathway and replicates in the endoplasmic reticulum of nonprofessional phagocytes. Infection and immunity, 66(12), pp.5711–24.
Pizarro-Cerdá, J., Moreno, E., et al., 1998. Virulent Brucella abortus prevents lysosome fusion and is distributed within autophagosome-like compartments. Infection and immunity, 66(5), pp.2387–92.
Platz, J. et al., 2004. Microbial DNA Induces a Host Defense Reaction of Human Respiratory Epithelial Cells. The Journal of Immunology, 173(2), pp.1219–1223.
Pollak, C.N. et al., 2012. Outer Membrane Vesicles from Brucella abortus Promote Bacterial Internalization by Human Monocytes and Modulate Their Innate Immune Response. PLoS ONE, 7(11), p.e50214.
Porte, F., Liautard, J.P. & Köhler, S., 1999. Early acidification of phagosomes containing Brucella suis is essential for intracellular survival in murine macrophages. Infection and immunity, 67(8), pp.4041–7.
Qureshi, N. et al., 1994. Structural analysis of the lipid A derived from the lipopolysaccharide of Brucella abortus. Innate Immunity, 1(3), pp.137–148.
Radhakrishnan, G.K. et al., 2009. Brucella TIR domain-containing protein mimics properties of the toll-like receptor adaptor protein TIRAP. Journal of Biological Chemistry, 284(15), pp.9892–9898.
Reibman, J. et al., 2003. Airway epithelial cells release MIP-3alpha/CCL20 in response to cytokines and ambient particulate matter. American journal of respiratory cell and molecular biology, 28(6), pp.648–54.

Bibliografía
148
Revelli, D. a, Boylan, J. a & Gherardini, F.C., 2012. A non-invasive intratracheal inoculation method for the study of pulmonary melioidosis. Frontiers in cellular and infection microbiology, 2(December), p.164.
Revelli DA, Boylan JA, G.F., 2012. A non-invasive intratracheal inoculation method for the study of pulmonary melioidosis. Front Cell Infect Microbiol, 2(December), p.164.
Rittig, M.G. et al., 2003. Smooth and rough lipopolysaccharide phenotypes of Brucella induce different intracellular trafficking and cytokine/chemokine release in human monocytes. Journal of leukocyte biology, 74(6), pp.1045–1055.
Rivas-Santiago, B. et al., 2005. Human {beta}-defensin 2 is expressed and associated with Mycobacterium tuberculosis during infection of human alveolar epithelial cells. Infection and immunity, 73(8), pp.4505–11.
Rivero Puente, Maravi-Poma E, Burusco M, Díaz R. 1978 Brucelosis.
Röhrl, J. et al., 2010. Specific binding and chemotactic activity of mBD4 and its functional orthologue hBD2 to CCR6-expressing cells. The Journal of biological chemistry, 285(10), pp.7028–34.
Roop, R.M. et al., 2004. Adaptation of the Brucellae to their intracellular niche. Molecular microbiology, 52(3), pp.621–30.
Rosen, D. et al., 2015. Klebsiella pneumoniae FimK Promotes Virulence in Murine Pneumonia. Journal of Infectious Diseases.
Rossetti, O.L. et al., 1996. Cloning of Brucella abortus gene and characterization of expressed 26-kilodalton periplasmic protein: potential use for diagnosis. Journal of clinical microbiology, 34(1), pp.165–9.
Russell, B.H. et al., 2008. Potential dissemination of Bacillus anthracis utilizing human lung epithelial cells. Cellular microbiology, 10(4), pp.945–57.
Salcedo, S.P. et al., 2008. Brucella control of dendritic cell maturation is dependent on the TIR-containing protein Btp1. PLoS Pathogens, 4(2).
Salcedo, S.P. et al., 2013. BtpB, a novel Brucella TIR-containing effector protein with immune modulatory functions. Frontiers in cellular and infection microbiology, 3(July), p.28.
Scharf, S. et al., 2010a. Legionella pneumophila induces human beta defensin-3 in pulmonary cells. Respiratory research, 11, p.93.
Scharf, S. et al., 2010b. Legionella pneumophila induces human beta defensin-3 in pulmonary cells.,
Scholz, H.C. et al., 2010. Brucella inopinata sp. nov., isolated from a breast implant infection. International journal of systematic and evolutionary microbiology, 60(Pt 4), pp.801–8.
Scholz, H.C. et al., 2008. Brucella microti sp. nov., isolated from the common vole Microtus arvalis. International journal of systematic and evolutionary microbiology, 58(Pt 2), pp.375–82.
Scholz, H.C. et al., 2009. Isolation of Brucella microti from mandibular lymph nodes of red foxes, Vulpes vulpes, in lower Austria. Vector borne and zoonotic diseases (Larchmont, N.Y.), 9(2), pp.153–6.
Schutte, B.C. & McCray, P.B., 2002. [Beta]-Defensins in Lung Host Defense. Annual review of physiology, 64, pp.709–748.

Bibliografía
149
Sha, Q. et al., 2004. Activation of airway epithelial cells by toll-like receptor agonists. American journal of respiratory cell and molecular biology, 31(3), pp.358–64.
Serre A. 1989. Immunology and pathophysiology of human brucellosis. In:Brucellosis Clinical and Laboratory Aspects. Ed Young EJ y Corbel MJ, 85. CRC Press
Sieira, R. et al., 2000. A homologue of an operon required for DNA transfer in Agrobacterium is required in Brucella abortus for virulence and intracellular multiplication. Journal of bacteriology, 182(17), pp.4849–55.
Sim, S.H. et al., 2009. Innate immune responses of pulmonary epithelial cells to Burkholderia pseudomallei infection. PloS one, 4(10), p.e7308.
Starner, T.D. et al., 2003. CCL20 is an inducible product of human airway epithelia with innate immune properties. American journal of respiratory cell and molecular biology, 29(5), pp.627–33.
Staszkiewicz, J. et al., 1991. Outbreak of Brucella melitensis among microbiology laboratory workers in a community hospital. Journal of clinical microbiology, 29(2), pp.287–90.
SULKIN, S.E., 1961. Laboratory-acquired infections. Bacteriological reviews, 25, pp.203–9.
Takahashi, A. et al., 2001. Production of β-defensin-2 by human colonic epithelial cells induced by Salmonella enteritidis flagella filament structural protein. FEBS Letters, 508(3), pp.484–488.
Tangirala, R.K., Murao, K. & Quehenberger, O., 1997. Regulation of expression of the human monocyte chemotactic protein-1 receptor (hCCR2) by cytokines. The Journal of biological chemistry, 272(12), pp.8050–6.
Tecle, T., Tripathi, S. & Hartshorn, K.L., 2010. Review: Defensins and cathelicidins in lung immunity. Innate immunity, 16(3), pp.151–159.
De Tejada, G.M. et al., 1995. The outer membranes of Brucella spp. are resistant to bactericidal cationic peptides. Infection and Immunity, 63(8), pp.3054–3061.
Thepen, T., Van Rooijen, N. & Kraal, G., 1989. Alveolar macrophage elimination in vivo is associated with an increase in pulmonary immune response in mice. The Journal of experimental medicine, 170(2), pp.499–509.
Thomas, E.D. et al., 1976. Direct evidence for a bone marrow origin of the alveolar macrophage in man. Science (New York, N.Y.), 192(4243), pp.1016–8.
Thomas, S.Y. et al., 2007. Multiple Chemokine Receptors, Including CCR6 and CXCR3, Regulate Antigen-Induced T Cell Homing to the Human Asthmatic Airway. The Journal of Immunology, 179(3), pp.1901–1912.
Thorley, A.J. et al., 2005. Primary human alveolar type II epithelial cell CCL20 (macrophage inflammatory protein-3alpha)-induced dendritic cell migration. American journal of respiratory cell and molecular biology, 32(4), pp.262–7.
Tibor, A., Decelle, B. & Letesson, J.J., 1999. Outer membrane proteins Omp10, Omp16, and Omp19 of Brucella spp. are lipoproteins. Infection and immunity, 67(9), pp.4960–2.
TREVER, R.W. et al., 1959. Brucellosis. I. Laboratory-acquired acute infection. A.M.A. archives of internal medicine, 103(3), pp.381–97.

Bibliografía
150
Trinchieri, G. & Sher, A., 2007. Cooperation of Toll-like receptor signals in innate immune defence. Nature reviews. Immunology, 7(3), pp.179–90.
Tsutsumi-Ishii, Y. & Nagaoka, I., 2003. Modulation of human beta-defensin-2 transcription in pulmonary epithelial cells by lipopolysaccharide-stimulated mononuclear phagocytes via proinflammatory cytokine production. Journal of immunology (Baltimore, Md. : 1950), 170(8), pp.4226–36.
Verstreate, D.R. et al., 1982. Outer membrane proteins of Brucella abortus: isolation and characterization. Infection and immunity, 35(3), pp.979–89.
de Vries, S.P.W. et al., 2009. Molecular aspects of Moraxella catarrhalis pathogenesis. Microbiology and molecular biology re iews : MMBR, 73(3), pp.389–406, Table of Contents.
Wallach, J.C. et al., 1997. Human infection by Brucella melitensis: An outbreak attributed to contact with infected goats. FEMS Immunology and Medical Microbiology, 19(4), pp.315–321.
Watson, J.A. et al., 1969. Deposition and clearance following inhalation and intratracheal injection of particles. Archives of environmental health, 19(1), pp.51–8.
Weiss, D.S. et al., 2005. MyD88, but not toll-like receptors 4 and 2, is required for efficient clearance of Brucella abortus. Infection and immunity, 73(8), pp.5137–5143.
Winter, A.J. et al., 1983. Immune response to porin in cattle immunized with whole cell, outer membrane, and outer membrane protein antigens of Brucella abortus combined with trehalose dimycolate and muramyl dipeptide adjuvants. Infection and immunity, 42(3), pp.1159–67.
Yagupsky, P. & Baron, E.J., 2005. Laboratory exposures to brucellae and implications for bioterrorism. Emerging infectious diseases, 11(8), pp.1180–5.
Yamada, H. et al., 2000. Protective role of interleukin-1 in mycobacterial infection in IL-1 alpha/beta double-knockout mice. Laboratory investigation; a journal of technical methods and pathology, 80(5), pp.759–767.
Yamamoto, K. et al., 2012. Type I alveolar epithelial cells mount innate immune responses during pneumococcal pneumonia. Journal of immunology (Baltimore, Md. : 1950), 189(5), pp.2450–9.
Yang, D. et al., 1999. Beta-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science (New York, N.Y.), 286(5439), pp.525–8.
Yang, D. et al., 2003. Many chemokines including CCL20/MIP-3alpha display antimicrobial activity. Journal of leukocyte biology, 74(3), pp.448–55.
Young EJ, Corbel MJ eds. 1989. CRC Press. Inc. Boca Ratón; chap. 5, pp. 53-72.
Young, E.J., 1995. An overview of human brucellosis. Clinical infectious diseases : an official publication of the Infectious Diseases Society of America, 21(2), pp.283–9; quiz 290.
Yu, M., 2002. The Role of Interleukin-6 in Pulmonary Inflammation and Injury Induced by Exposure to Environmental Air Pollutants. Toxicological Sciences, 68(2), pp.488–497.
Zwerdling, A. et al., 2009. Brucella abortus activates human neutrophils. Microbes and infection / Institut Pasteur, 11(6-7), pp.689–97.