resistance to adriamycin cytotoxicity among respiratory...
TRANSCRIPT

ANTIMICROBIAL AGENTS AND CHEMOTHERAPY, Mar. 1980, p. 443-4490066-4804/80/03-0443/07$02.00/0
Vol. 17, No. 3
Resistance to Adriamycin Cytotoxicity Among Respiratory-Deficient Mutants in Yeast
SHARON C. HIXON,* ADIL OCAK, JONAH E. THOMAS, AND J. PATRICK DAUGHERTYDepartment ofBiochemistry, University ofAlabama in Birmingham, Birmingham, Alabama 35294
Saccharomyces cell uptake of Adriamycin and the ensuing cytotoxic responsewere found to be dependent upon the ionic strength of the medium used for drugtreatment. A given concentration of Adriamycin which inhibited growth incomplete medium was found to be significantly cytotoxic when administered inwater. Many survivors after Adriamycin treatment in water were found to berespiratory-deficient petite mutants containing mitochondrial deoxyribonucleicacid mutations. Petite mutants arising after Adriamycin treatment were notinduced but selected from the preexisting population of spontaneously derivedpetite mutants (normal frequency, 2%) due to an increased resistance of thesemutants to killing by Adriamycin as compared with normal respiratory-sufficientcells. The response to Adriamycin in mitochondrial deoxyribonucleic acid respi-ratory-deficient mutants (rho-, rho', mitr) with different impaired mitochondrialfunctions was studied. All were similarly more resistant to killing by Adriamycinthan wild-type cells. The common deficiency shared by these mutants, i.e.,nonfunctioning electron transport, may play a role in protecting these mutantsfrom Adriamycin cytotoxicity. In addition, normal cells grown on glycerol, re-quiring aerobic respiration for carbon source utilization were more susceptible tokilling by Adriamycin than cells grown on glucose. These studies suggest that amitochondrial function in yeast may interact with Adriamycin to potentiate a cellcytotoxic mechanism of the drug.
Adriamycin (doxorubicin) has shown a widespectrum of antitumor activity in laboratory an-imals and in humans (4). The mechanism ofaction is generally attributed to deoxyribonu-cleic acid (DNA) binding via the process ofintercalation (5, 21). Adriamycin binding toDNA is thought to be related to the inhibitionof ribonucleic acid (RNA) and DNA synthesis,mitotic inhibition, mutagenesis, and chromo-somal aberrations reported to occur in mam-malian cells treated with Adriamycin (18, 24,26).Attempts to explain a major clinical dose-lim-
iting side effect of the drug, cardiomyopathy,have led to studies of non-nuclear cell targetsaffected by Adriamycin (15). An inhibition ofmitochondrial respiration (10,12,17) and reportsofvisible mitochondrial degeneration from ultra-structural studies of Adriamycin-treated myo-cardial cells (13) have implicated a possible roleof mitochondrial function in Adriamycin-causedcardiotoxicity. Studies with yeast as a modeleucaryote may provide insight into any involve-ment of mitochondrial function in the antitumoror cardiotoxic mechanism(s) of action of Adria-mycin. Saccharomyces cerevisiae, a facultativeanaerobe, is capable of aerobic respiration utiliz-ing mitochondrial function, or growth may occurfrom energy derived only by anaerobic fermen-
tation. This flexibility permits an accumulationof mitochondrial DNA mutants in yeast that arerespiratory deficient but able to grow from en-ergy derived from fermentation (2).
Previous studies with Adriamycin and dau-nomycin in yeast have demonstrated a selectivegrowth inhibition on medium containing drugand a nonfermentable carbon substrate as op-posed to glucose-containing medium (1, 6, 22, 23,27). These experiments have suggested an inhi-bition of mitochondrial function by these an-thracyclines which prevents growth on glycerolor pyruvate medium. We report here additionalexperiments with yeast treated with Adriamycinin water that may indicate a secondary or alter-native interpretation ofAdriamycin mechanismsof action that may depend upon some undefinedmitochondrial function.
MATERIALS AND METHODSS. cerevisiae N123 (a hisl rho') was used in all
experiments to determine the typical response of cellsto Adriamycin in water. N123 (a hisl rho') is a petitemutant with no mitochondrial DNA obtained by treat-ment with 10 uM ethidium bromide for 8 h (8). Anabsence of mitochondrial DNA was verified by radio-active labeling of nucleic acids in the presence ofcycloheximide (11). Isolation of total cell DNA andfractionation on a preparative cesium chloride gra-dient revealed an absence of mitochondrial DNA.
443
on June 4, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
ANTIMICROB. AGENTS CHEMOTHER.
Strains used to study the response of mit- mutants toAdriamycin treatment were kindly donated by P. Perl-man of Ohio State University. These strains were: theparent wild-type 161 (a ade lys rho') and the petitemutant 14-9 (a ade lys rho-). Mitochondrial mitCmutants derived from the parent strain were: PZ1(cob) intron, PZ5 (cob) exon, M44 (oxi3), E3 (oxi2),and E65 (maps between oxil and cap) (16). Cellgrowth medium was YPD (1% yeast extract, 2% pep-tone, and 2 or 8% glucose) or YPG (3% glycerol).
Cells were grown to exponential phase (3 x 107/ml)in either YPD or YPG on a shaker at 30°C, washed indistilled water, sonicated to remove any cell clumps,and diluted to 5 x 106 cells/ml in a given concentrationof drug in a selected medium. Cells were then incu-bated with shaking at 30°C in the dark. At given timeintervals cells were withdrawn, diluted in water, andspread onto YPD 2% agar plates in triplicate andallowed to grow into visible colonies (5 days). Celldilutions were adjusted so that each set of agar platestotaled approximately 600 colonies. Actual survivingcells per milliliter could be calculated from the knowndilution factors and numbers of surviving colonies onagar plates as compared with the control plates fromcells suspended in medium without Adriamycin. Petiteand mixed petite mutant colonies (sectored) were iden-tified by a tetrazolium overlay in 2% agar (20). Eachexperiment represented in the figures is one of two ormore repeated experiments.The cell uptake of Adriamycin in different media
was determined by measuring the absorbance of thesupernatants from cell suspensions that were pelletedby centrifugation. The absorbance was measured at479 nm, the A. for Adriamycin (3).Adriamycin was supplied by the National Cancer
Institute, Bethesda, Md., and by John Durant, Com-prehensive Cancer Center, University of AlabamaMedical Center, Birmingham.
too. X- _ =-=
(a)
10.-
-J
10a.V
RESULTSThe previously reported growth inhibition of
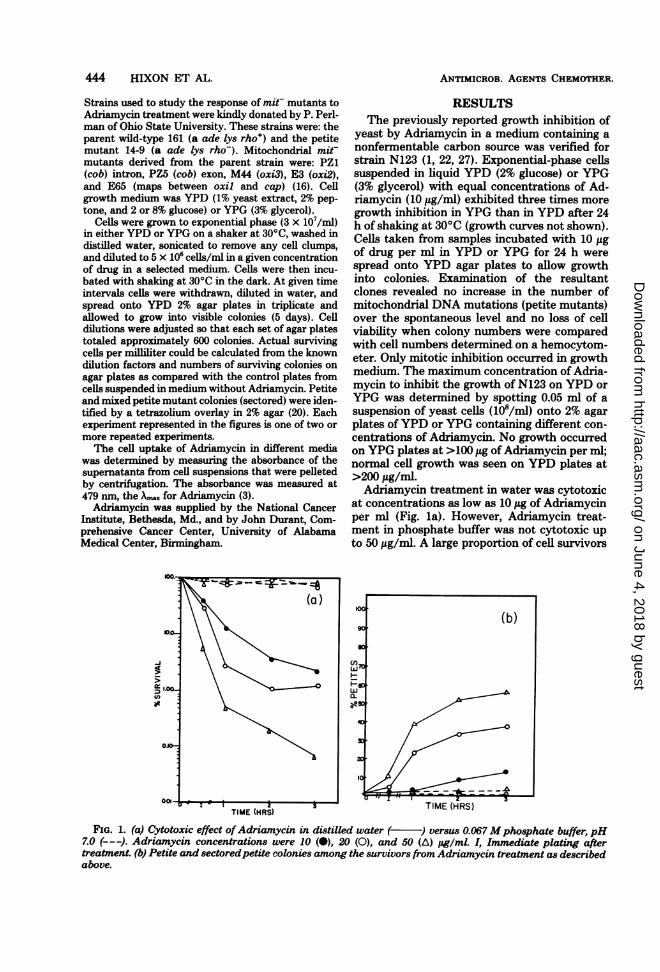
yeast by Adriamycin in a medium containing anonfermentable carbon source was verified forstrain N123 (1, 22, 27). Exponential-phase cellssuspended in liquid YPD (2% glucose) or YPG(3% glycerol) with equal concentrations of Ad-riamycin (10 ,Ag/ml) exhibited three times moregrowth inhibition in YPG than in YPD after 24h of shaking at 30°C (growth curves not shown).Cells taken from samples incubated with 10 ,ugof drug per ml in YPD or YPG for 24 h werespread onto YPD agar plates to allow growthinto colonies. Examination of the resultantclones revealed no increase in the number ofmitochondrial DNA mutations (petite mutants)over the spontaneous level and no loss of cellviability when colony numbers were comparedwith cell numbers determined, on a hemocytom-eter. Only mitotic inhibition occurred in growthmedium. The maximum concentration of Adria-mycin to inhibit the growth of N123 on YPD orYPG was determined by spotting 0.05 ml of asuspension of yeast cells (108/ml) onto 2% agarplates ofYPD or YPG containing different con-centrations of Adriamycin. No growth occurredon YPG plates at >100 ,ug of Adriamycin per ml;normal cell growth was seen on YPD plates at>200 jig/ml.Adriamycin treatment in water was cytotoxic
at concentrations as low as 10 ,ug of Adriamycinper ml (Fig. la). However, Adriamycin treat-ment in phosphate buffer was not cytotoxic upto 50 jg/ml. A large proportion of cell survivors
TIME (HRS)
FIG. 1. (a) Cytotoxic effect ofAdriamycin in distilled water ( ) versus 0.067Mphosphate buffer, pH7.0 (---J. Adriamycin concentrations were 10 (0), 20 (0), and 50 (A) pg/ml. I, Immediate plating aftertreatment. (b) Petite and sectoredpetite colonies among the survivors from Adriamycin treatment as describedabove.
444 HIXON ET AL.
on June 4, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

ADRIAMYCIN CYTOTOXICITY RESISTANCE 445
after Adriamycin treatment in water were res-piratory-deficient mutants identified from thetetrazolium overlay of colonies on each petridish (Fig. lb). Greater than 90% of the petitemutants observed after Adriamycin treatmentwere whole-colony nonsectored mutants.The difference in cytotoxic response to Adria-
mycin in water versus buffer appeared to berelated to a decreased cell affinity for the drugwith the increasing ionic strength of the mediumused for treatment. Cells were suspended invarious dilutions of phosphate buffer (pH 7.0)containing 20 ug of Adriamycin per ml. Cellsuspensions were withdrawn at increasing timeintervals and centrifuged to pellet the cells. Theabsorbance of the supernatants was read at 479nm. By using the absorbance of an Adriamycinsolution at 20 ,Ag/ml as a standard, the percent-age of the drug bound to the cells could becalculated from the decrease in absorbance fromthe cell supernatants. The drug uptake wasgreatly reduced by increasing the ionic strengthof the medium (Fig. 2). After 5 h of incubation,cells in buffer continued to be blocked for drug
100
A
70 -/
50 -
30~
uptake. Although the percentage of drug uptakedoes not discriminate between outside cell bind-ing and drug entry into the cell, the inability toproduce a cytotoxic effect with Adriamycin inbuffer suggests that reduced uptake does relateto reduced cell penetration.The increase in petite mutants among the cell
survivors after Adriamycin treatment in watercould be due to petite mutant induction or aselective killing of normally respiring cells,which would favor an enrichment of the popu-lation by petite mutants. Figure 3 provides theanswer to this question. A mixed population ofselected spontaneous petite mutants (rho-) ofN123 was treated with 50 jig of Adriamycin perml in water. A parallel experiment was per-formed on a normal cell population containingapproximately 2% petite mutants and 98% nor-mal cells (grandes). The percentages of total cellsurvival were determined for the pure sponta-neous petite population and the normal mixed-cell population. In addition, the percentages ofcell survival of those petites and normal cells
TIME(HRS) TIME (HRS)FIG. 3. Survival of a population of spontaneous
FIG. 2. Percentage of Adriamycin taken up by rho- petite mutants (0) and N123 rho' (O) afteryeast from an initial concentration of20 pg/ml in (A) treatment with 50 pg ofAdriamycin per ml in water.distilled water, (B) 0.0067 M phosphate buffer, (C) Symbols: (0) Overall survival ofa normalpopulation0.067M buffer, and (D) 0.67M buffer, pH 7. Percent- of cells treated as described above; (x) calculatedage uptake was calculated from the optical density of survival ofspontaneous petite mutants; and (A) nor-cell supernatants at 479 nm at increasing time inter- mal cells among the petite mutants and normal cells,vals. respectively, in the mixed population.
VOL. 17, 1980
on June 4, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

446 HIXON ET AL.
within the petite and nornal cell populationwere calculated separately. There was a largedifference in cell viability of petite mutants ver-sus grandes (aerobic-respiring cells) whentreated with Adriamycin in water (Fig. 3). Thepetite mutants were more resistant to killing byAdriamycin by more than a factor of 10. Inaddition, the survival of the petites within thenormal population was almost identical to thatof the pure culture of spontaneous petite mu-tants. These data suggest that the increase inpetite mutants among the survivors after Adria-mycin treatment is due primarily to their selec-tive advantage, for viability after Adriamycintreatment and not to a significant conversion ofnormal cells to petite mutants by the drug. Spon-taneously derived petite mutants (rho-) in yeastare known to retain mitochondrial DNA withdeletions and repetitions (2). Normal cells orpetite mutants may be treated with ethidiumbromide to cause a complete degradation andloss of mitochondrial DNA from the cells (rho')(8). These mutants (rho- and rho') have non-functioning mitochondria with regard to elec-tron transport, adenosine 5'-triphosphate syn-thesis, and protein synthesis. Both rho- and
DUtr1e kR \
DRUG( pg/ml)FIG. 4. Overall survival of cells grown on YPD
with 2% glucose (A), YPD with 8% glucose (-), orYPG with 3% glycerol (0) before Adriamycin treat-ment in water for 2 h. Open symbols refer to thesurvival ofnormal non-petite cells within the normalcellpopulation.
ANTIMICROB. AGENTS CHEMOTHER.
rho' petite mutants were found to be moreresistant to killing by Adriamycin in water thannormally respiring cells (Fig. 3).To determine the influence of mitochondrial
aerobic respiration on Adriamycin cytotoxicity,the response to Adriamycin in water was ob-served in cells grown in either YPD (2% glucose),YPD (8% glucose), or YPG (3% glycerol) (Fig.4). Cells grown in glycerol before Adriamycintreatment were more susceptible to cell killingby drug than cells grown on 2 or 8% glucose. Noconsistent differences in cell killing could bedetected in cells grown in different concentra-tions of glucose. Measurements of drug bindingas previously described (Fig. 2) did not indicatesignificant differences in drug uptake in thethree categories of cells. The enrichment of pe-tite mutants within the population of cells pre-viously grown in YPD (2 and 8% glucose) andYPG (3% glycerol) and treated with Adriamycinis shown in Fig. 5. Cells grown in glucose showeda higher proportion of petite mutants among thesurvivors than those grown in glycerol at equalsurvival fractions. These results may be ex-plained by the higher spontaneous number ofpetite mutants in cells grown in glucose. Sincepetite mutants are less vulnerable to cell killingby Adriamycin, they are more quickly enrichedin glucose-grown cells due to their higher spon-taneous number within the starting population.Since spontaneous petite mutants cannot grow
% SURVIVALFIG. 5. Proportion of petite mutants among cell
survivors after Adriamycin treatment of cells grownon YPD with 2% glucose (A), YPD with 8% glucose(), or YPG with 3% glycerol (0). Points shown arethose from several independent experiments.
on June 4, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

ADRIAMYCIN CYTOTOXICITY RESISTANCE 447
in glycerol medium, fewer mutants were avail-able for selection by Adriamycin.The selective resistance of petite mutants to
killing by Adriamycin as compared with normalcells could be explained by a difference in someaspect of mitochondrial metabolism in thesecells. To ascertain which factor may be influenc-ing this response, three types of cells with differ-ent mitochondrial DNA mutations were ob-served: rho-, rho', and mit-. Table 1 indicatesthe different mitochondrial functions that areinoperative in these types of mutants. Both rho-and rho' petite mutants responded similarly toAdriamycin treatment (Fig. 3). These resultseliminate the role of any mitochondrial RNAtranscription product which might influence Ad-riamycin cell entry or metabolism. The behaviorof mit- mutants was investigated to determinethe presence of a mitochondrial protein whichmight influence the response to Adriamycin.mit- mutants have been shown to have pointmutations on mitochondrial DNA in regionscoding for proteins involved in aerobic respira-tion (2). mit- mutants do, however, have anintact mitochondrial protein-synthesizing ma-chinery, in contrast to petite mutants. Five mit-mutants were treated with Adriamycin in wateralong with the parent wild type and a petitemutant. The mit- mutants were more resistantto killing by Adriamycin than the wild type.Three separate experiments revealed no consist-ent orderly trend among the rho& and mit-mutants except for the fact that as a group theywere >10-fold more resistant to the drug thannormal wild-type cells. The similar response toAdriamycin of all three mitochondrial DNA mu-tants indicates that the differences in mitochon-drial function among them probably does notplay a role in their selective resistance to Adria-mycin. The only known common denominator,no electron transport or oxidative phosphoryla-tion, may be the deciding factor which influencestheir selective survival in the presence of Adria-mycin.
TABLE 1. Mitochondrial functions in variousmitochondrial genotypes
a Normal Petite mutantFunction0 No' mitCrh~ rho- rhoo
DNA synthesis + + - +RNA synthesis + + - +Protein synthesis + - - +Electron transport +and oxidativephosphorylationaSee Borst and Grivell (2) for discussion of the
functions.
DISCUSSION
These studies on Adriamycin treatment ofyeast in growth medium or buffer versus waterexplain why other investigators have not ob-served a significant enrichment ofpetite mutantsamong cells treated with Adriamycin in growthmedium. Inhibition of cell growth rather thancell death did not provide a sufficient selectivepressure to reveal the resistance of petite mu-
tants to the cytotoxic effects of the drug.The similar resistance of both petite and mit-
mutants to cell killing does not support a mech-anism whereby a mitochondrial transporter pro-tein or RNA transcript influences the cell entryof Adriamycin. Uptake studies on petite andmit- mutants and cells grown in glucose or glyc-erol did not indicate any observable differencesin uptake in various genotypes or when cells
were grown under different conditions.Although differences in drug transport may
not be conclusively excluded from these data, a
more plausible explanation of the experimentalresults may relate to the absence of electrontransport in petite and mit- mutants. This ideais further supported by the increased suscepti-bility of normal cells to killing by Adriamycinwhen grown on glycerol rather than glucose. Thenecessity of aerobic respiration required for glyc-erol utilization as a carbon source could explainthe increased cytotoxic effect of Adriamycin forcells grown in glycerol if electron transport isresponsible for potentiating the cytotoxicity ofAdriamycin.The experiments reported here suggest that
the absence of mitochondrial electron transportand oxidative phosphorylation may serve to pro-tect cells from death by Adriamycin. Therefore,in normal cells the cytotoxic nature of Adria-mycin may be enhanced by a mitochondrialfunction.
It was previously suggested that Adriamycininhibits mitochondrial function in yeast, whichtherefore prevents growth on glycerol (1, 6, 22,23, 27). The experiments reported here suggest
TABLE 2. Survival of mit- mutants in 50 pg ofAdriamycin per ml in water
% Survival
StrainImeitSnImmegte 2 h 4 hplating
161 wild type 0.55 0.093 0.03414-9 rho- 100 83.5 27.6PZ1 cob intron 89.0 58.0 5.10PZ5 cob exon 100 50.0 58.6E65 oxil and cap 52.8 22.3 4.30E3 oxi2 59.5 16.2 7.60M44 oxi3 69.0 86.0 3.15
VOL. 17, 1980
on June 4, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

448 HIXON ET AL.
an altemative interpretation: low levels of celldamage in glycerol medium may result in mitoticinhibition. It is also possible that these twomechanisms operate independently or in con-cert, depending upon the conditions used for celltreatment by Adriamycin.These experiments are of particular interest
because ofthe unexplained vulnerability ofhearttissue to Adriamycin. The high aerobic energyrequirements of heart cells may pose specialrisks if heart mitochondrial function is inhibitedby the drug or is responsible for convertingAdriamycin to a more cytotoxic agent. Studieswith animal heart mitochondria have indicatedthat Adriamycin inhibits overall aerobic respi-ration and specifically the coenzyme Qlo en-zymes succinoxidase and reduced nicotinamideadenine dinucleotide oxidase (12). Subsequently,coenzyme Qlo has been used to prevent theinhibition by Adriamycin of coenzyme Qlo en-zymes and to partially prevent the cardiotoxicside effect of the drug in animal studies (7, 14).The yeast experiments described here suggestthat an inhibition ofelectron transport and aero-bic respiration per se may not completely de-scribe the consequences of Adriamycin interac-tion with electron transport. Since yeast cellsmay survive without aerobic respiration, an in-hibition of electron transport does not fully ac-count for the enhanced cytotoxic effect of thedrug in normal yeast cells. A proliferation of freeradicals as a consequence of Adriamycin inter-action with electron transport may explain thecytotoxic effect of the drug in normal yeast cells.The quinone moiety ofAdriamycin may interactwith mitochondrial electron transport to gener-ate free radicals by redox cycling like that dem-onstrated by Goodman and Hockstein (9), usingrat liver microsomes and Adriamycin. Free-rad-ical production by Adriamycin in animal cellsand ensuing lipid peroxidation have been sug-gested as possible contributing factors for car-diac damage by Adriamycin (25). The reportedvitamin E protection of cardiac damage by Ad-riamycin serves to emphasize the relevance ofthis mechanism (19). Our ongoing studies arepresently focused on identifying the molecularmechanism in yeast whereby a mitochondrialfunction may serve to enhance the cytotoxicproperties of the drug.
ACKNOWLEGMENTSThese experiments were supported by grants from Adria
Laboratories and the American Heart AssOciation AlabamaAffiliate.We thank Sevim Ocak, Major Bledsole, Harold Franks, and
Neal Ellis for their technical assistance.
LITERATURE CITED
1. Andreini, G., C. K. Beretta, and 0. Sonzogni. 1977.Antimitochondrial activity of adriamycin and dauno-
ANTIMICROB. AGENTS CHEMOTHER.
mycin on Saccharomyces cerevisiae. Pharmacol. Res.Commun. 9:155-164.
2. Borst, P., and L. A. Grivell. 1978. The mitochondrialgenome of yeast. Cell 15:705-723.
3. Daugherty, J. Patrick, S. C. Hixon, and K L Yield-ing. 1979. Direct in vitro photoaffinity labeling ofDNAwith daunoribicin, adriamycin, and rubidazone.Biochim. Biophys. Acta 565:13-21.
4. Di Marco, A. 1975. Adriamycin (NSC-123127) mode andmechanism of action. Cancer Chemother. Rep. 6:91-106.
5. Di Marco, A., F. Zunino, and Ri Silverstrini. 1971.Interaction of some daunomycin derivatives with de-oxyribonucleic acid and their biological activity. Bio-chem. Pharmacol. 20:1323-1328.
6. Evans, I., D. Linstead, P. K Rhodes, and D. Wilkie.1973. Inhibition of RNA synthesis in mitochondria bydaunomycin. Biochim. Biophys. Acta 312:323-336.
7. Folkers, K., J. Y. Choe, and A. B. Combs. 1978. Rescueby coenzyme Qlo from electrocardiographic abnormali-ties caused by the toxicity of adriamycin in the rat.Proc. Natl. Acad. Sci. U.S.A. 75:5178-5180.
&. Goldring, E., L. Grossman, D. .Krupnick, D. Cryer,and J. Marmur. 1970. The petite mutation in yeast.Loss of mitochondrial DNA during induction of petiteswith ethidium bromide. J. Mol. Biol. 52:323-335.
9. Goodman, J., and P. Hochstein. 1977. Generation offree radicals and lipid peroxidation by redox cycling ofadriamycin and daunomycin. Biochem. Biophys. Res.Commun. 77:797-03.
10. Gosalvez, M., M. Blanco, J. Hunter, K. Miko, and B.Chance. 1974. Effects of anticancer agents on the res-piration of isolated mitochondria and tumor cells. Eur.J. Cancer 10:567-574.
11. Grossman, L. I., E. S. Goldring, and J. Marmur. 1969.Preferential synthesis of yeast mitochondrial DNA inthe absence of protein synthesis. J. Mol. Biol. 46:367-376.
12. Iwamoto, Y., I. L Hansen, T. H. Porter, and K.Folkers. 1974. Inhibition of coenzyme Qlo-enzymes,succinoxidase and NADH-oxidase by adriamycin andother quinones having antitumor activity. Biochem.Biophys. Res. Commun. 58:633-638.
13. Jaenke, aL S. 1974. An anthracycline antibiotic-inducedcardiomyopathy in rabbits. Lab. Invest. 30:292-304.
14. Kishi, T., T. Watanabe, and K. Folkers. 1976. Bioener-getics in clinical medicine: prevention by forms of coen-zyme Q of the inhibition by adriamycin of coenzymeQjo-enzymes in mitochondria of the myocardiium. Proc.Natl. Acad. Sci. U.S.A. 73:4653-4656.
15. Lenaz, L, and J. A. Page. 1976. Cardiotoxicity of adria-mycin and related anthracyclines. Cancer Treat. Rev.3:111-120.
16. Mahler, HI R., D. Hanson, D. Miller, C. C. Li, N. J.Alexander. R. D. Vincent, and P. S. Perlman. 1978.Regulatory aspects of mitochondrial biogenesis, p. 513-547. In M. Bacila, B. L. Horecker, and A. 0. M. Stop-pani, (ed.), Biochemistry and genetics of yeast. Aca-demic Press Inc., New York.
17. Mailer, K., and D. H. Petering. 1976. Inhibition ofoxidative phosphorylation in tumor cells and mitochon-dria by daunomycin and adriamycin. Biochem. Phar-macol. 25:2085-2089.
18. Marquardt, H., and H. Marquardt. 1977. Induction ofmalignant transfonnation and mutagenesis in cell cul-tures by cancer chemotherapeutic agents. Cancer 40:1930-1934.
19. Myers, C. E., W. P. McGuire, R. HIL Lis, K. Grotain-ger, and R. C. Young. 1977. Adriamycin: the rote oflipid peroxidation in cardiac toxicity and tumor re-sponse. Science 197:165-167.
20. Ogur, M., R. St. John, and S. NagaL 1957. Tetrazoliumoverlay technique for population studies of respirationdeficiency in yeast. Science 125:928-929.
on June 4, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

ADRIAMYCIN CYTOTOXICITY RESISTANCE 449
21. Pigram, W. J., W. Fuller, and L. D. Hamilton. 1972.Stereochemistry of intercalation: interaction of dauno-mycin with DNA. Nature (London) New Biol. 235:17-19.
22. Rhodes, P. M., and D. Wilkie. 1971. Inhibition effect ofdaunorubicin on mitochondrial synthesis in Saccharo-myces cerevisiae. Heredity 26:347.
23. Roberts, H., W. M. Choo, S. C. Smith, S. Marzuki, A.W. Linnane, T. H. Porter, and K. Folkers. 1978. Thesite of inhibition of mitochondrial electron transfer bycoenzyme Q analogues. Arch. Biochem. Biophys. 191:-
306-315.24. Rusconi, A., and A. DiMarco. 1969. Inhibition of nucleic
acid synthesis by daunomycin and its relationship tothe uptake of the drug in Hela cells. Cancer Res. 29:1507-1511.
25. Sato, S., M. Iwaizumi, K. Handa, and Y. Tamura.1977. Electron spin resonance study on the mode ofgeneration of free radicals of daunomycin, adriamycinand carboquone in NAD (P)H-microsome system. Gann68:603-608.
26. Vig, B. K. 1971. Chromosome aberrations induced inhuman leukocytes by the antileukemia antibiotic adria-mycin. Cancer Res. 31:32-38.
27. Wilkie, D. 1972. The yeast cell in anti-mitochondrialactivity of drugs. Med. Biol. Illus. 22:119-124.
VOL. 17, 1980
on June 4, 2018 by guesthttp://aac.asm
.org/D
ownloaded from