research article stimulation of mmp-1 and ccl2 by...
TRANSCRIPT

Hindawi Publishing CorporationMediators of InflammationVolume 2013 Article ID 437123 12 pageshttpdxdoiorg1011552013437123
Research ArticleStimulation of MMP-1 and CCL2 by NAMPT in PDL Cells
Marjan Nokhbehsaim12 Sigrun Eick3 Andressa Vilas Boas Nogueira14
Per Hoffmann56 Stefan Herms56 Holger Froumlhlich7 Soslashren Jepsen28
Andreas Jaumlger29 Joni Augusto Cirelli4 and James Deschner12
1 Experimental Dento-Maxillo-Facial Medicine Center of Dento-Maxillo-Facial Medicine University of Bonn53111 Bonn Germany
2 Clinical Research Unit 208 Center of Dento-Maxillo-Facial Medicine University of Bonn Welschnonnenstraszlige 1753111 Bonn Germany
3Department of Periodontology Laboratory of Oral Microbiology University of Bern 3010 Bern Switzerland4Department of Diagnosis and Surgery School of Dentistry SP UNESP 14801-903 Araraquara Brazil5 Institute of Human Genetics Biomedical Center University of Bonn 53127 Bonn Germany6Division of Medical Genetics University Hospital Basel and Department of Biomedicine University of Basel4058 Basel Switzerland
7 Bonn-Aachen International Center for IT Algorithmic Bioinformatics University of Bonn 53113 Bonn Germany8Department of Periodontology Operative and Preventive Dentistry University of Bonn 53111 Bonn Germany9Department of Orthodontics University of Bonn 53111 Bonn Germany
Correspondence should be addressed to James Deschner jamesdeschneruni-bonnde
Received 14 May 2013 Accepted 18 July 2013
Academic Editor Timo Sorsa
Copyright copy 2013 Marjan Nokhbehsaim et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Periodontitis is an inflammatory disease caused by pathogenic microorganisms and characterized by the destruction of theperiodontium Obese individuals have an increased risk of periodontitis and elevated circulating levels of adipokines such asnicotinamide phosphoribosyltransferase (NAMPT) may be a pathomechanistic link between both diseases The aim of this invitro study was to examine the regulation of periodontal ligament (PDL) cells by NAMPT and its production under inflammatoryand infectious conditions NAMPT caused a significant upregulation of 9 genes and downregulation of 3 genes as analyzed bymicroarray analysis Eight of these genes could be confirmed by real-time PCR NAMPT induced a significant upregulation ofEGR1MMP-1 SYT7 ITPKA CCL2 NTM IGF2BP3 andNRP1 NAMPT also increased significantly theMMP-1 andCCL2 proteinsynthesis NAMPT was significantly induced by interleukin-1120573 and the periodontal microorganism P gingivalis NAMPT maycontribute to periodontitis through upregulation of MMP-1 and CCL2 in PDL cells Increased NAMPT levels as found in obesitymay therefore represent a mechanism whereby obesity could confer an increased risk of periodontitis Furthermore microbial andinflammatory signals may enhance the NAMPT synthesis in PDL cells and thereby contribute to the increased gingival and serumlevels of this adipokine as found in periodontitis
1 Introduction
Periodontitis is a chronic inflammatory disease which ischaracterized by the destruction of the tooth-supportingtissues such as the periodontal ligament (PDL) and causedby pathogenic microorganisms such as Porphyromonas
gingivalis Tannerella forsythia Treponema denticola andAggregatibacter actinomycetemcomitans These periodon-topathogens can trigger local production of proinflamma-tory mediators such as interleukin-1120573 (IL-1120573) cyclooxy-genase 2 (COX2) as well as chemokine cc motif ligand2 (CCL2) and matrix-degrading proteases such as matrix
2 Mediators of Inflammation
metalloproteinase-1 (MMP-1) by infiltrating and residentcells of the periodontium As a consequence of the exag-gerated immunoinflammatory and proteolytic processes theperiodontal tissues are subjected to degradation and resorp-tion which can finally result in tooth loss [1 2] Data from theNationalHealth andNutritionExamination Survey III whichanalyzed the health andnutritional status in theUnited Statesdemonstrate that approximately half of the US populationagedge30 years is afflictedwith periodontitis [3] Periodontitisis also associated with systemic diseases and conditions suchas cardiovascular diseases diabetes mellitus and obesityand has a negative influence on a wide range of physicalpsychological and social aspects of quality of life [4ndash6]
The exact mechanisms underlying the associationbetween periodontitis and obesity are as yet unknownHowever it has been suggested that increased serum levelsof adipokines derived from adipose tissue could make obeseindividuals more susceptible to periodontitis The adiposetissue is not only an energy storage tissue but also acts asan endocrine organ from which adipokines such as nicoti-namide phosphoribosyltransferase (NAMPT also known asvisfatin or pre-B-cell colony-enhancing factor 1) are secretedAdipokines not only regulate insulin sensitivity and energyexpenditure but also inflammatory and healing processes [7]NAMPT is mainly produced by macrophages and adipocytesin the adipose tissue and induces production of inflammatorymediators and nuclear factor-kappaB (NF120581B) activation [8]High serum levels of NAMPT are found in obesity metabolicsyndrome type 2 diabetes atherosclerosis and other diseases[9ndash11] Therefore increased serum levels of NAMPT couldbe at least onemechanism whereby these diseases contributeto periodontitis Recently it has been demonstrated thatNAMPT is also present in high levels in gingival crevicularfluid (GCF) gingival tissues and serum from periodontallydiseased patients indicating that NAMPT might also beproduced locally in the periodontium and play a role inthe etiopathogenesis of periodontitis [12 13] Howeverwhether NAMPT regulates the gene expression and proteinsynthesis of periodontal cells has yet to be elucidatedFurthermore whether NAMPT is induced in these cells byperiodontopathogens and IL-1120573 which has been shown tobe increased in GCF and gingival tissues at inflamed sites isalso largely unknown This study was therefore establishedto examine the actions of NAMPT and its regulation bymicrobial and inflammatory signals in human PDL cells
2 Materials and Methods
21 Culture and Treatment of Cells PDL cells were harvestedfrom periodontally healthy teeth that had to be extractedfor orthodontic reasons Written informed parental consentand approval of the Ethics Committee of the University ofBonn were obtained (04311) The cells were cultured inDulbeccorsquos minimal essential medium (DMEM InvitrogenKarlsruhe Germany) supplemented with 10 fetal bovineserum (FBS Invitrogen) 100 units penicillin and 100 120583gmLstreptomycin (Biochrom Berlin Germany) at 37∘C in ahumidified atmosphere of 5CO
2 PDL cells from passages 3
to 5 were seeded (50000 cellswell) on cell culture plates andgrown to 80 confluence One day prior to the experimentthe FBS concentration was reduced to 1 Medium waschanged every other day
In order to study the actions of NAMPT on PDL cellsvarious concentrations of NAMPT (30ndash300 ngmL BiomolHamburg Germany) were added to cells To mimic an inf-lammatory environment cells were incubated with IL-1120573(02ndash5 ngmL Calbiochem San Diego CA USA) as donein our previous studies [14ndash16] In order to simulate aninfectious environment in vitro cells were stimulated withthe inactivated oral periodontopathogens Porphyromonasgingivalis ATCC 33277 Tannerella forsythia ATCC 43037Treponema denticola ATCC 35405 and Aggregatibacter acti-nomycetemcomitans Y4 (optical density 0025 005 and 01)Bacteria were suspended in PBS (OD
660 nm = 1 equivalentto 12 times 109 bacterial cellsmL) and exposed two timesto ultrasonication (160W for 15min) resulting in a com-plete killing In the present study cells were exposed toNAMPT IL-1120573 or periodontopathogens for up to 3 d Inorder to unravel the intracellular mechanisms underlyingthe effects of NAMPT PDL cells were preincubated withspecific inhibitors against NF120581B (pyrrolidine dithiocarba-mate PDTC 10120583M Calbiochem) JNK (SP600125 10 120583MCalbiochem) p38 (SB203580 10 120583M Calbiochem) MEK12(U0126 10 120583M Calbiochem) or PI3 K (50 120583M Calbiochem)signaling pathways 1 h before experiments
22 Microarray Analysis The regulatory effects of NAMPTon the gene expression of PDL cells were analyzed bya genomewide expression profiling using IlluminarsquosHumanHT-12 v4 Expression BeadArrays (Illumina SanDiego CA USA) The RNA was reverse transcribedamplified and subsequently biotinylated using the IlluminaTotalPrep-96 RNA Amplification Kit (Life TechnologiesCarlsbad CA USA) The resulting cRNA was hybridized tothe arrays according to the manufacturers manual using anautomated liquid handling pipeline and scanned on an IScanSystemThe intensity signals were QC checked and exportedusing Illuminarsquos GenomeStudio 2011 v11 software suite
All array data underwent a rigorous quality controlbefore applying background correction (subtraction of anoffset which is estimated from the control probes) and a sub-sequent variance stabilizing transformation [17] followed byquantile normalization [18] All steps were conducted usingthe lumi R-package [19] After calculation of presentabsentcalls we only kept probes which were present in each patientin each group In order to consider additional confoundingfactors which might potentially influence gene expressionbut are not directly captured by the available patient infor-mation we applied the algorithm by Leek and Storey [20]which estimates a surrogate variable model In additionrelative array quality weights were computed via the limmaR-package [21] using the algorithmbyRitchie et al [22] Bothsurrogate variables and array weights can be consideredwhen fitting a linear model to each gene via limma Inour case the limma model contained two additive factorsnamely a donor and a group factor that is essentiallywewere
Mediators of Inflammation 3
fitting 2-way ANOVAmodel After model fitting differentialgene expression for all clinically relevant differences can beextracted as contrasts from the model and corresponding 119875values can be computed (multiple testing correction) [23]We refer the reader to the limma user guide for an excellentintroduction [24]
23 Real-Time PCR RNA was extracted using an RNAextraction kit (Qiagen Hilden Germany) and a total of1 120583g of RNA was reverse transcribed using iScriptTM SelectcDNA Synthesis Kit (Bio-Rad Laboratories Munich Ger-many) at 42∘C for 90min followed by 85∘C for 5minExpression of early growth response 1 (EGR1) MMP-1 synaptotagmin 7 (SYT7) inositol 145-trisphosphate 3-kinase a (ITPKA) CCL2 neurotrimin (NTM) insulin-like growth factor 2 mRNA-binding protein 3 (IGF2BP3)neuropilin 1 (NRP1) potassium channel tetramerizationdomain-containing protein 12 (KCTD12) heat-shock 27 kDprotein 3 (HSPB3) transmembrane 4 L six family member20 (TM4SF20) regulator of G protein signaling 4 (RGS4)NAMPT COX2 and glyceraldehyde-3-phosphate dehydro-genase (GAPDH) was detected by real-time PCR usingthe iCycler iQ detection system (Bio-Rad Laboratories)SYBR Green (Bio-Rad Laboratories) and specific primers(QuantiTect Primer Assay Qiagen) One 120583L of cDNA wasamplified as a template in a 25120583L reactionmixture containing125 120583L 2x QuantiFast SYBR Green PCR Master Mix (Qia-gen) 25 120583L of primers and deionized water The mixturewas heated initially at 95∘C for 5min and then followed by40 cycles with denaturation at 95∘C for 10 s and combinedannealingextension at 60∘C for 30 s GAPDH was usedas an endogenous control The data were analyzed by thecomparative threshold cycle (CT) method
24 ELISA The levels of NAMPT MMP-1 and CCL2 in thesupernatants of PDL cells were analysed by commerciallyavailable enzyme-linked immunoassay (ELISA) kits (Ray-Biotech Norcross GAUSA) according to themanufacturerrsquosinstructionsThe absorbancewasmeasuredwith amicroplatereader (PowerWave x BioTek Instruments Winooski VTUSA) at 450 nm The data were normalized by the cellnumber which was measured with an automatic cell counter(Moelab Hilden Germany)
25 Immunocytochemistry PDL cells were cultured in thepresence and absence of P gingivalis ATCC 33277 or IL-1120573 on glass coverslips in 24-well plates After 3 days cellswere fixed in 4 paraformaldehyde (Sigma-AldrichMunichGermany) at pH 74 and room temperature (RT) for 15minand then permeabilized in 01TritonX-100 (Sigma-Aldrich)for 5min Non specific antigens were blocked by incubationwith serum block (LSAB System Santa Cruz BiotechnologySanta Cruz CA USA) for 20min Subsequently cells wereincubated at RT for 90min with rabbit polyclonal antibody toNAMPT (Santa Cruz Biotechnology) Afterwards cells werelabeled with goat anti-rabbit IgG-HRP secondary antibody(Cell Signalling Technology Danvers MA USA) for 45min
For staining cells were exposed to DAB chromogen (DakoHamburg Germany) for 3min at RT After each incubationstep cells were washed twice with PBS (Sigma-Aldrich)Counterstaining was performed with Mayerrsquos Haematoxylin(Merck Eurolab Dietikon Switzerland) for 2min Cover-slips were mounted in DePex mounting medium (ServaElectrophoresis Heidelberg Germany) Standardized pho-tomicrographs were taken using an Axioplan 2 imagingmicroscope (Carl Zeiss MicroImaging Jena Germany)
26 Immunofluorescence PDL cells were fixed with 4paraformaldehyde in PBS pH 74 for 10min at RT washedwith PBS and treated with 01 Triton X-100 for 5min atRTThen cells were washed again with PBS and blocked witha blocking buffer (nonfat dry milk Bio-Rad Laboratories)for 1 h at RT After washing the cells were incubated withprimary rabbit antibody NF120581B p65 (1 400 Cell SignallingTechnology) for 90min and with secondary antibody CY3(1 2000 Abcam Cambridge MA USA) for 45min Cellswere observed under a 20x objective using an Axioplan 2imaging microscope (Carl Zeiss MicroImaging) The imageswere captured with a PVCAM camera and the VisiViewcapturing software (Visitron Systems Puchheim Germany)
27 Statistical Analysis The IBM SPSS Statistics 20 softwarewas used for analysis Mean values and standard errors ofthe mean (SEM) were calculated All experiments were per-formed in triplicate and repeated at least twice For statisticalanalysis parametric (119905 test and ANOVA followed by the post-hoc Dunnettrsquos test) and non parametric tests (Wilcoxon andMann-Whitney 119880 tests) were applied Differences betweengroups were considered significant at 119875 lt 005 Microarraydata were analyzed in the Bonn-Aachen International Centerfor IT Algorithmic Bioinformatics University of BonnGermany (see Section 22)
3 Results
31 Regulation of Gene Expression by NAMPT in PDL CellsFirst we sought to clarify whether NAMPT regulates theexpression of genes in PDL cells by performing amicroarray-based approach As shown in Table 1 NAMPT caused a sig-nificant upregulation of 9 genes and a significant downregula-tion of 3 genes in PDL cells from 3 donors at 1 dTheNAMPT-upregulated genes were the following EGR1 MMP-1 SYT7ITPKA CCL2 NTM IGF2BP3 NRP1 and KCTD12 The 3NAMPT-downregulated genes comprisedHSPB3 TM4SF20and RGS4 (Table 1) In PDL cells from 10 donors the resultsfrom the microarray analysis were then validated by real-time PCRwhich confirmed 8 out of the 12NAMPT-regulatedgenes NAMPT induced a significant upregulation of themRNA expression for EGR1 (24-fold) MMP-1 (38-fold)SYT7 (46-fold) ITPKA (28-fold) CCL2 (23-fold) NTM(21-fold) IGF2BP3 (19-fold) and NRP1 (17-fold) KCTD12was also upregulated by NAMPT but the effect did not reachstatistical significance (Table 2 and Figure 1(a))The effects ofNAMPT on the remaining genes could not be confirmed byPCR (data not shown)
4 Mediators of Inflammation
Table 1 Regulation of gene expression by NAMPT as analyzed bymicroarray
Gene LogFC adj 119875 valueEGR1 2184 0008MMP-1 1545 0010SYT7 1443 0010ITPKA 1292 0010CCL2 1150 0028NTM 1114 0028IGF2BP3 1040 0020NRP1 1024 0010KCTD12 1019 0048HSPB3 minus1035 0030TM4SF20 minus1164 0028RGS4 minus1397 0025Effect of NAMPT on gene expression of PDL cells from 3 donors at 1 d Onlygenes with statistically significant (FDR lt 5) logFC ge 1 (upregulationby at least 2-fold) or logFC le minus1 (downregulation by at least 50)were considered Abbreviations EGR1 early growth response 1 MMP-1matrix metalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12 HSPB3 heat-shock 27-kD protein 3 TM4SF20transmembrane 4 L six family member 20 RGS4 regulator of G proteinsignaling 4
Table 2 Upregulation of gene expression by NAMPT as analyzedby real-time PCR
Gene Fold change(mean) SD SEM 119875 value
EGR1 2422 2306 0421 lt0001MMP-1 3781 3116 0569 lt0001SYT7 4575 7947 1451 lt0001ITPKA 2764 3182 0581 0001CCL2 2285 1346 0246 lt0001NTM 2124 2460 0449 lt0001IGF2BP3 1897 0970 0177 lt0001NRP1 1653 1658 0303 lt0001KCTD12 1713 0958 0175 0095Effect of NAMPT on gene expression of PDL cells from 10 donorsat 1 d Abbreviations EGR1 early growth response 1 MMP-1 matrixmetalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12
32 Effect of NAMPT on MMP-1 and CCL2 Since MMP-1andCCL2 aremolecules known to be strongly associatedwithperiodontitis we further studied their regulation byNAMPTAs observed at 1 d NAMPT increased significantly theMMP-1 (14-fold) and CCL2 (18-fold) expressions at 3 d (Fig-ure 1(b)) Furthermore the stimulatory effects of NAMPTon MMP-1 and CCL2 expressions were dose-dependent atboth time points as shown in Figures 1(c)ndash1(f) In addition
the NAMPT-induced upregulation of the MMP-1 and CCL2expressions was parallelled by increased MMP-1 and CCL2protein levels in the supernatants ofNAMPT-stimulated cellsas compared to those of the control at 1 d and 3 d (Figures 1(g)and 1(h))
33 Exploitation of the JNK Pathway by NAMPT Next westudied how the effects of NAMPT on MMP-1 and CCL2 aremediated intracellularly As evidenced by immunofluores-cence microscopy NAMPT stimulated the nuclear translo-cation of NF120581B and caused a maximal NF120581B accumulationwithin the nucleus at 60min (Figure 2(a)) However pre-incubation of cells with PDTC a specific inhibitor of NF120581Bhad not effect on the NAMPT-stimulated upregulation ofMMP-1 and CCL2 (data not shown) Moreover the upreg-ulation of both molecules by NAMPT was also not affectedby inhibitors against the p38 MEK12 and PI3K signalingpathways However SP600125 an inhibitor of JNK signalingblocked completely the stimulatory effect of NAMPT on theMMP-1 expression at 1 d (Figure 2(b)) In addition SP600125reduced significantly theNAMPT-stimulated upregulation ofCCL2 by 69 at this time point (Figure 2(b))
34 Stimulation of NAMPT Production by P gingivalisWe then wondered whether pathogens that are stronglyassociated with periodontitis are capable of inducing theproduction of NAMPT in PDL cells Whereas T denticolaT forsythia and A actinomycetemcomitans had no significanteffect P gingivalis increased significantly theNAMPTmRNAexpression at 1 d and 3 d (Figures 3(a) and 3(b)) As shown inFigures 3(c) and 3(d) the actions of P gingivalis on NAMPTmRNA expression were dose-dependent Interestingly pre-incubation with a specific inhibitor against NF120581B signalingblocked completely the P gingivalis-induced stimulation ofNAMPT at 1 d Furthermore inhibitors against JNK and p38signaling reduced the stimulatory effects of NAMPT by 87and 37 respectively at this time point
The stimulation of NAMPT expression by P gingivaliswas also observed at protein level as analyzed by ELISA at1 d PDL cells in the presence and absence of P gingivalisproduced 2883 plusmn 029 ng NAMPT protein104 cells and1791 plusmn 058 ng NAMPT protein104 cells respectively At3 d P gingivalis-stimulated cells and control cells synthesized3097 plusmn 309 ng NAMPT protein104 cells and 2165 plusmn294 ng NAMPT protein104 cells respectively The differ-ences between both groups at each time point were sig-nificant The stimulation of NAMPT protein production byP gingivalis was also evidenced by immunocytochemistry(Figure 4(c))
Like NAMPT COX2 was significantly upregulated by Pgingivalis at 1 d and 3 d (Figures 3(e) and 3(f)) By contrast allother periodontopathogens were not capable of stimulatingthe COX2 expression at both time points (Figures 3(e)and 3(f)) The P gingivalis-induced stimulation of COX2expression was dose-dependent (Figures 3(g) and 3(h))
35 Stimulation of NAMPT Production by IL-1120573 Finallywe examined whether NAMPT is also induced by IL-1120573 in
Mediators of Inflammation 5
1d
5
6
7
8
0
2
1
3
4
EGR1 MMP-1 SYT7 ITPKA CCL2 NTM IGF2BP3 NRP1 KCTD12
lowast
lowast
lowast
lowast
mRN
A ex
pres
sion
(fold
of c
ontro
l)
lowast
lowastlowast lowast lowast
(a)
lowast15
20MMP-1 CCL2
lowast 3d
00
05
NAMPT + +minusminus
10
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
lowast
lowast
lowast
2
3
1d4
1
030 100NAMPT
(ngmL)3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast
lowast
2
3
3d4
10
30 100NAMPT(ngmL)
3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
lowast
2
3
1d4
lowast
1
030 100NAMPT
(ngmL)3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(e)
lowast2
3
3d4
lowastlowast
10
30 100NAMPT(ngmL)
3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(f)
125
75100
175150
2001d
lowast
3d
02550
lowast
NAMPT + +minusminus
MM
P-1
prot
ein
(pg10
3ce
lls)
(g)
50
203040
7060
801d
lowast
lowast
3d
010
NAMPT + +minus minus
CCL2
pro
tein
(pg10
4ce
lls)
(h)
Figure 1 Stimulation of gene expression and protein synthesis by NAMPT Upregulation of genes by NAMPT (100 ngmL) in PDL cells from10 donors at 1 d (a) Upregulation ofMMP-1 and CCL2 expression by NAMPT (100 ngmL) in PDL cells from 10 donors at 3 d (b) Stimulationof MMP-1 expression by various concentrations of NAMPT in PDL cells from 3 donors at 1 d (c) and 3 d (d) Stimulation of CCL2 expressionby various concentrations of NAMPT in PDL cells from 3 donors at 1 d (e) and 3 d (f) Stimulation of MMP-1 protein synthesis by NAMPT(100 ngmL) in PDL cells from 6 donors at 1 d and 3 d (g) Stimulation of CCL2 protein synthesis by NAMPT (100 ngmL) in PDL cells from6 donors at 1 d and 3 d (h) All experiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005)different from NAMPT-untreated cells (control)
PDL cells Interestingly IL-1120573 upregulated significantly theNAMPT expression at 1 d and 3 d (Figure 4(a)) Moreoverthe actions of IL-1120573 on NAMPT expression were dose-dependent at 1 d (Figure 1(b)) but not at 3 d (data not shown)When cells were preincubated with an inhibitor against theMEK12 signaling pathway the IL-1120573-induced stimulation ofNAMPT expression was completely suppressed Moreoverinhibitors against JNK NF120581B and p38 signaling reduced the
stimulatory effect of IL-1120573 onNAMPTby 80 78 and 46respectively
The upregulation of NAMPT at transcriptional level wasparalleled by increased NAMPT protein levels in the super-natants of IL-1120573-stimulated cells at 1 d PDL cells in thepresence and absence of IL-1120573 synthesized 3268 plusmn 362 ngNAMPT protein104 cells and 2028 plusmn 436 ng NAMPTprotein104 cells respectively At 3 d IL-1120573-treated cells and
6 Mediators of Inflammation
50 120583m
0 min 30 min
60 min 90 min
(a)
lowast
10
15
20
25
1dCCL2
lowast
MMP-1
00NAMPT
SP600125+ +
+
++
+
05
minus minus
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
Figure 2 Intracellular pathways triggered by NAMPT Stimulation of NF120581B nuclear translocation by NAMPT at 60min (a) Inhibition ofthe NAMPT-stimulated MMP-1 and CCL2 upregulation by preincubation of PDL cells with SP600125 a selective inhibitor of JNK at 1 d (b)Images and data from one representative donor are shown All experiments were performed in triplicate and repeated at least twice Mean plusmnSEM lowastsignificant (119875 lt 005) difference between groups
Mediators of Inflammation 7
1dlowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(a)
3d
lowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(b)
lowast
87
91d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast87
93d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
1dlowast
20
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
25
(e)
3dlowast
20
25
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(f)
lowast
15
20
251d
0025 00500
OD 0100
1
5
10
0000
lowast
lowast
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(g)
lowast
15
20
253d
lowast
00025 0050OD 0100
1
5
10
0000
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(h)
Figure 3 Regulation of NAMPT andCOX2 expressions by periodontopathogens NAMPT expression in PDL cells from 3 donors in responseto various periodontopathogens (OD 01 P gingivalis Pg T denticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (a) and 3 d(b) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of P gingivalis (OD 0025 0050 and 0100)at 1 d (c) and 3 d (d) COX2 expression in PDL cells from 3 donors in response to various periodontopathogens (OD 01 P gingivalis Pg Tdenticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (e) and 3 d (f) Stimulation of COX2 expression in PDL cells from 3 donorsby various concentrations of P gingivalis (OD 0025 0050 and 0100) at 1 d (g) and 3 d (h) All experiments were performed in triplicate andrepeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from P gingivalis-untreated cells (control)
control cells produced 3097 plusmn 382 ng NAMPT protein104
cells and 2043 plusmn 076 ng NAMPT protein104 cells respec-tivelyThedifferences between groupswere significant at bothtime points The stimulation of NAMPT protein synthesisby IL-1120573 was also observed by immunocytochemistry (Fig-ure 4(c))
IL-1120573 also induced a significant upregulation of COX2expression at 1 d and 3 d (Figure 4(d))
4 Discussion
Our study shows for this first time that NAMPT stimulatesthe production ofCCL2 andMMP-1 in humanPDL cells sug-gesting that NAMPT may contribute to periodontal inflam-mation and matrix destruction through the production ofthese molecules Furthermore NAMPT is induced by theperiodontopathogen P gingivalis and the proinflammatorycytokine IL-1120573 in PDL cells which shows that microbial
8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

2 Mediators of Inflammation
metalloproteinase-1 (MMP-1) by infiltrating and residentcells of the periodontium As a consequence of the exag-gerated immunoinflammatory and proteolytic processes theperiodontal tissues are subjected to degradation and resorp-tion which can finally result in tooth loss [1 2] Data from theNationalHealth andNutritionExamination Survey III whichanalyzed the health andnutritional status in theUnited Statesdemonstrate that approximately half of the US populationagedge30 years is afflictedwith periodontitis [3] Periodontitisis also associated with systemic diseases and conditions suchas cardiovascular diseases diabetes mellitus and obesityand has a negative influence on a wide range of physicalpsychological and social aspects of quality of life [4ndash6]
The exact mechanisms underlying the associationbetween periodontitis and obesity are as yet unknownHowever it has been suggested that increased serum levelsof adipokines derived from adipose tissue could make obeseindividuals more susceptible to periodontitis The adiposetissue is not only an energy storage tissue but also acts asan endocrine organ from which adipokines such as nicoti-namide phosphoribosyltransferase (NAMPT also known asvisfatin or pre-B-cell colony-enhancing factor 1) are secretedAdipokines not only regulate insulin sensitivity and energyexpenditure but also inflammatory and healing processes [7]NAMPT is mainly produced by macrophages and adipocytesin the adipose tissue and induces production of inflammatorymediators and nuclear factor-kappaB (NF120581B) activation [8]High serum levels of NAMPT are found in obesity metabolicsyndrome type 2 diabetes atherosclerosis and other diseases[9ndash11] Therefore increased serum levels of NAMPT couldbe at least onemechanism whereby these diseases contributeto periodontitis Recently it has been demonstrated thatNAMPT is also present in high levels in gingival crevicularfluid (GCF) gingival tissues and serum from periodontallydiseased patients indicating that NAMPT might also beproduced locally in the periodontium and play a role inthe etiopathogenesis of periodontitis [12 13] Howeverwhether NAMPT regulates the gene expression and proteinsynthesis of periodontal cells has yet to be elucidatedFurthermore whether NAMPT is induced in these cells byperiodontopathogens and IL-1120573 which has been shown tobe increased in GCF and gingival tissues at inflamed sites isalso largely unknown This study was therefore establishedto examine the actions of NAMPT and its regulation bymicrobial and inflammatory signals in human PDL cells
2 Materials and Methods
21 Culture and Treatment of Cells PDL cells were harvestedfrom periodontally healthy teeth that had to be extractedfor orthodontic reasons Written informed parental consentand approval of the Ethics Committee of the University ofBonn were obtained (04311) The cells were cultured inDulbeccorsquos minimal essential medium (DMEM InvitrogenKarlsruhe Germany) supplemented with 10 fetal bovineserum (FBS Invitrogen) 100 units penicillin and 100 120583gmLstreptomycin (Biochrom Berlin Germany) at 37∘C in ahumidified atmosphere of 5CO
2 PDL cells from passages 3
to 5 were seeded (50000 cellswell) on cell culture plates andgrown to 80 confluence One day prior to the experimentthe FBS concentration was reduced to 1 Medium waschanged every other day
In order to study the actions of NAMPT on PDL cellsvarious concentrations of NAMPT (30ndash300 ngmL BiomolHamburg Germany) were added to cells To mimic an inf-lammatory environment cells were incubated with IL-1120573(02ndash5 ngmL Calbiochem San Diego CA USA) as donein our previous studies [14ndash16] In order to simulate aninfectious environment in vitro cells were stimulated withthe inactivated oral periodontopathogens Porphyromonasgingivalis ATCC 33277 Tannerella forsythia ATCC 43037Treponema denticola ATCC 35405 and Aggregatibacter acti-nomycetemcomitans Y4 (optical density 0025 005 and 01)Bacteria were suspended in PBS (OD
660 nm = 1 equivalentto 12 times 109 bacterial cellsmL) and exposed two timesto ultrasonication (160W for 15min) resulting in a com-plete killing In the present study cells were exposed toNAMPT IL-1120573 or periodontopathogens for up to 3 d Inorder to unravel the intracellular mechanisms underlyingthe effects of NAMPT PDL cells were preincubated withspecific inhibitors against NF120581B (pyrrolidine dithiocarba-mate PDTC 10120583M Calbiochem) JNK (SP600125 10 120583MCalbiochem) p38 (SB203580 10 120583M Calbiochem) MEK12(U0126 10 120583M Calbiochem) or PI3 K (50 120583M Calbiochem)signaling pathways 1 h before experiments
22 Microarray Analysis The regulatory effects of NAMPTon the gene expression of PDL cells were analyzed bya genomewide expression profiling using IlluminarsquosHumanHT-12 v4 Expression BeadArrays (Illumina SanDiego CA USA) The RNA was reverse transcribedamplified and subsequently biotinylated using the IlluminaTotalPrep-96 RNA Amplification Kit (Life TechnologiesCarlsbad CA USA) The resulting cRNA was hybridized tothe arrays according to the manufacturers manual using anautomated liquid handling pipeline and scanned on an IScanSystemThe intensity signals were QC checked and exportedusing Illuminarsquos GenomeStudio 2011 v11 software suite
All array data underwent a rigorous quality controlbefore applying background correction (subtraction of anoffset which is estimated from the control probes) and a sub-sequent variance stabilizing transformation [17] followed byquantile normalization [18] All steps were conducted usingthe lumi R-package [19] After calculation of presentabsentcalls we only kept probes which were present in each patientin each group In order to consider additional confoundingfactors which might potentially influence gene expressionbut are not directly captured by the available patient infor-mation we applied the algorithm by Leek and Storey [20]which estimates a surrogate variable model In additionrelative array quality weights were computed via the limmaR-package [21] using the algorithmbyRitchie et al [22] Bothsurrogate variables and array weights can be consideredwhen fitting a linear model to each gene via limma Inour case the limma model contained two additive factorsnamely a donor and a group factor that is essentiallywewere
Mediators of Inflammation 3
fitting 2-way ANOVAmodel After model fitting differentialgene expression for all clinically relevant differences can beextracted as contrasts from the model and corresponding 119875values can be computed (multiple testing correction) [23]We refer the reader to the limma user guide for an excellentintroduction [24]
23 Real-Time PCR RNA was extracted using an RNAextraction kit (Qiagen Hilden Germany) and a total of1 120583g of RNA was reverse transcribed using iScriptTM SelectcDNA Synthesis Kit (Bio-Rad Laboratories Munich Ger-many) at 42∘C for 90min followed by 85∘C for 5minExpression of early growth response 1 (EGR1) MMP-1 synaptotagmin 7 (SYT7) inositol 145-trisphosphate 3-kinase a (ITPKA) CCL2 neurotrimin (NTM) insulin-like growth factor 2 mRNA-binding protein 3 (IGF2BP3)neuropilin 1 (NRP1) potassium channel tetramerizationdomain-containing protein 12 (KCTD12) heat-shock 27 kDprotein 3 (HSPB3) transmembrane 4 L six family member20 (TM4SF20) regulator of G protein signaling 4 (RGS4)NAMPT COX2 and glyceraldehyde-3-phosphate dehydro-genase (GAPDH) was detected by real-time PCR usingthe iCycler iQ detection system (Bio-Rad Laboratories)SYBR Green (Bio-Rad Laboratories) and specific primers(QuantiTect Primer Assay Qiagen) One 120583L of cDNA wasamplified as a template in a 25120583L reactionmixture containing125 120583L 2x QuantiFast SYBR Green PCR Master Mix (Qia-gen) 25 120583L of primers and deionized water The mixturewas heated initially at 95∘C for 5min and then followed by40 cycles with denaturation at 95∘C for 10 s and combinedannealingextension at 60∘C for 30 s GAPDH was usedas an endogenous control The data were analyzed by thecomparative threshold cycle (CT) method
24 ELISA The levels of NAMPT MMP-1 and CCL2 in thesupernatants of PDL cells were analysed by commerciallyavailable enzyme-linked immunoassay (ELISA) kits (Ray-Biotech Norcross GAUSA) according to themanufacturerrsquosinstructionsThe absorbancewasmeasuredwith amicroplatereader (PowerWave x BioTek Instruments Winooski VTUSA) at 450 nm The data were normalized by the cellnumber which was measured with an automatic cell counter(Moelab Hilden Germany)
25 Immunocytochemistry PDL cells were cultured in thepresence and absence of P gingivalis ATCC 33277 or IL-1120573 on glass coverslips in 24-well plates After 3 days cellswere fixed in 4 paraformaldehyde (Sigma-AldrichMunichGermany) at pH 74 and room temperature (RT) for 15minand then permeabilized in 01TritonX-100 (Sigma-Aldrich)for 5min Non specific antigens were blocked by incubationwith serum block (LSAB System Santa Cruz BiotechnologySanta Cruz CA USA) for 20min Subsequently cells wereincubated at RT for 90min with rabbit polyclonal antibody toNAMPT (Santa Cruz Biotechnology) Afterwards cells werelabeled with goat anti-rabbit IgG-HRP secondary antibody(Cell Signalling Technology Danvers MA USA) for 45min
For staining cells were exposed to DAB chromogen (DakoHamburg Germany) for 3min at RT After each incubationstep cells were washed twice with PBS (Sigma-Aldrich)Counterstaining was performed with Mayerrsquos Haematoxylin(Merck Eurolab Dietikon Switzerland) for 2min Cover-slips were mounted in DePex mounting medium (ServaElectrophoresis Heidelberg Germany) Standardized pho-tomicrographs were taken using an Axioplan 2 imagingmicroscope (Carl Zeiss MicroImaging Jena Germany)
26 Immunofluorescence PDL cells were fixed with 4paraformaldehyde in PBS pH 74 for 10min at RT washedwith PBS and treated with 01 Triton X-100 for 5min atRTThen cells were washed again with PBS and blocked witha blocking buffer (nonfat dry milk Bio-Rad Laboratories)for 1 h at RT After washing the cells were incubated withprimary rabbit antibody NF120581B p65 (1 400 Cell SignallingTechnology) for 90min and with secondary antibody CY3(1 2000 Abcam Cambridge MA USA) for 45min Cellswere observed under a 20x objective using an Axioplan 2imaging microscope (Carl Zeiss MicroImaging) The imageswere captured with a PVCAM camera and the VisiViewcapturing software (Visitron Systems Puchheim Germany)
27 Statistical Analysis The IBM SPSS Statistics 20 softwarewas used for analysis Mean values and standard errors ofthe mean (SEM) were calculated All experiments were per-formed in triplicate and repeated at least twice For statisticalanalysis parametric (119905 test and ANOVA followed by the post-hoc Dunnettrsquos test) and non parametric tests (Wilcoxon andMann-Whitney 119880 tests) were applied Differences betweengroups were considered significant at 119875 lt 005 Microarraydata were analyzed in the Bonn-Aachen International Centerfor IT Algorithmic Bioinformatics University of BonnGermany (see Section 22)
3 Results
31 Regulation of Gene Expression by NAMPT in PDL CellsFirst we sought to clarify whether NAMPT regulates theexpression of genes in PDL cells by performing amicroarray-based approach As shown in Table 1 NAMPT caused a sig-nificant upregulation of 9 genes and a significant downregula-tion of 3 genes in PDL cells from 3 donors at 1 dTheNAMPT-upregulated genes were the following EGR1 MMP-1 SYT7ITPKA CCL2 NTM IGF2BP3 NRP1 and KCTD12 The 3NAMPT-downregulated genes comprisedHSPB3 TM4SF20and RGS4 (Table 1) In PDL cells from 10 donors the resultsfrom the microarray analysis were then validated by real-time PCRwhich confirmed 8 out of the 12NAMPT-regulatedgenes NAMPT induced a significant upregulation of themRNA expression for EGR1 (24-fold) MMP-1 (38-fold)SYT7 (46-fold) ITPKA (28-fold) CCL2 (23-fold) NTM(21-fold) IGF2BP3 (19-fold) and NRP1 (17-fold) KCTD12was also upregulated by NAMPT but the effect did not reachstatistical significance (Table 2 and Figure 1(a))The effects ofNAMPT on the remaining genes could not be confirmed byPCR (data not shown)
4 Mediators of Inflammation
Table 1 Regulation of gene expression by NAMPT as analyzed bymicroarray
Gene LogFC adj 119875 valueEGR1 2184 0008MMP-1 1545 0010SYT7 1443 0010ITPKA 1292 0010CCL2 1150 0028NTM 1114 0028IGF2BP3 1040 0020NRP1 1024 0010KCTD12 1019 0048HSPB3 minus1035 0030TM4SF20 minus1164 0028RGS4 minus1397 0025Effect of NAMPT on gene expression of PDL cells from 3 donors at 1 d Onlygenes with statistically significant (FDR lt 5) logFC ge 1 (upregulationby at least 2-fold) or logFC le minus1 (downregulation by at least 50)were considered Abbreviations EGR1 early growth response 1 MMP-1matrix metalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12 HSPB3 heat-shock 27-kD protein 3 TM4SF20transmembrane 4 L six family member 20 RGS4 regulator of G proteinsignaling 4
Table 2 Upregulation of gene expression by NAMPT as analyzedby real-time PCR
Gene Fold change(mean) SD SEM 119875 value
EGR1 2422 2306 0421 lt0001MMP-1 3781 3116 0569 lt0001SYT7 4575 7947 1451 lt0001ITPKA 2764 3182 0581 0001CCL2 2285 1346 0246 lt0001NTM 2124 2460 0449 lt0001IGF2BP3 1897 0970 0177 lt0001NRP1 1653 1658 0303 lt0001KCTD12 1713 0958 0175 0095Effect of NAMPT on gene expression of PDL cells from 10 donorsat 1 d Abbreviations EGR1 early growth response 1 MMP-1 matrixmetalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12
32 Effect of NAMPT on MMP-1 and CCL2 Since MMP-1andCCL2 aremolecules known to be strongly associatedwithperiodontitis we further studied their regulation byNAMPTAs observed at 1 d NAMPT increased significantly theMMP-1 (14-fold) and CCL2 (18-fold) expressions at 3 d (Fig-ure 1(b)) Furthermore the stimulatory effects of NAMPTon MMP-1 and CCL2 expressions were dose-dependent atboth time points as shown in Figures 1(c)ndash1(f) In addition
the NAMPT-induced upregulation of the MMP-1 and CCL2expressions was parallelled by increased MMP-1 and CCL2protein levels in the supernatants ofNAMPT-stimulated cellsas compared to those of the control at 1 d and 3 d (Figures 1(g)and 1(h))
33 Exploitation of the JNK Pathway by NAMPT Next westudied how the effects of NAMPT on MMP-1 and CCL2 aremediated intracellularly As evidenced by immunofluores-cence microscopy NAMPT stimulated the nuclear translo-cation of NF120581B and caused a maximal NF120581B accumulationwithin the nucleus at 60min (Figure 2(a)) However pre-incubation of cells with PDTC a specific inhibitor of NF120581Bhad not effect on the NAMPT-stimulated upregulation ofMMP-1 and CCL2 (data not shown) Moreover the upreg-ulation of both molecules by NAMPT was also not affectedby inhibitors against the p38 MEK12 and PI3K signalingpathways However SP600125 an inhibitor of JNK signalingblocked completely the stimulatory effect of NAMPT on theMMP-1 expression at 1 d (Figure 2(b)) In addition SP600125reduced significantly theNAMPT-stimulated upregulation ofCCL2 by 69 at this time point (Figure 2(b))
34 Stimulation of NAMPT Production by P gingivalisWe then wondered whether pathogens that are stronglyassociated with periodontitis are capable of inducing theproduction of NAMPT in PDL cells Whereas T denticolaT forsythia and A actinomycetemcomitans had no significanteffect P gingivalis increased significantly theNAMPTmRNAexpression at 1 d and 3 d (Figures 3(a) and 3(b)) As shown inFigures 3(c) and 3(d) the actions of P gingivalis on NAMPTmRNA expression were dose-dependent Interestingly pre-incubation with a specific inhibitor against NF120581B signalingblocked completely the P gingivalis-induced stimulation ofNAMPT at 1 d Furthermore inhibitors against JNK and p38signaling reduced the stimulatory effects of NAMPT by 87and 37 respectively at this time point
The stimulation of NAMPT expression by P gingivaliswas also observed at protein level as analyzed by ELISA at1 d PDL cells in the presence and absence of P gingivalisproduced 2883 plusmn 029 ng NAMPT protein104 cells and1791 plusmn 058 ng NAMPT protein104 cells respectively At3 d P gingivalis-stimulated cells and control cells synthesized3097 plusmn 309 ng NAMPT protein104 cells and 2165 plusmn294 ng NAMPT protein104 cells respectively The differ-ences between both groups at each time point were sig-nificant The stimulation of NAMPT protein production byP gingivalis was also evidenced by immunocytochemistry(Figure 4(c))
Like NAMPT COX2 was significantly upregulated by Pgingivalis at 1 d and 3 d (Figures 3(e) and 3(f)) By contrast allother periodontopathogens were not capable of stimulatingthe COX2 expression at both time points (Figures 3(e)and 3(f)) The P gingivalis-induced stimulation of COX2expression was dose-dependent (Figures 3(g) and 3(h))
35 Stimulation of NAMPT Production by IL-1120573 Finallywe examined whether NAMPT is also induced by IL-1120573 in
Mediators of Inflammation 5
1d
5
6
7
8
0
2
1
3
4
EGR1 MMP-1 SYT7 ITPKA CCL2 NTM IGF2BP3 NRP1 KCTD12
lowast
lowast
lowast
lowast
mRN
A ex
pres
sion
(fold
of c
ontro
l)
lowast
lowastlowast lowast lowast
(a)
lowast15
20MMP-1 CCL2
lowast 3d
00
05
NAMPT + +minusminus
10
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
lowast
lowast
lowast
2
3
1d4
1
030 100NAMPT
(ngmL)3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast
lowast
2
3
3d4
10
30 100NAMPT(ngmL)
3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
lowast
2
3
1d4
lowast
1
030 100NAMPT
(ngmL)3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(e)
lowast2
3
3d4
lowastlowast
10
30 100NAMPT(ngmL)
3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(f)
125
75100
175150
2001d
lowast
3d
02550
lowast
NAMPT + +minusminus
MM
P-1
prot
ein
(pg10
3ce
lls)
(g)
50
203040
7060
801d
lowast
lowast
3d
010
NAMPT + +minus minus
CCL2
pro
tein
(pg10
4ce
lls)
(h)
Figure 1 Stimulation of gene expression and protein synthesis by NAMPT Upregulation of genes by NAMPT (100 ngmL) in PDL cells from10 donors at 1 d (a) Upregulation ofMMP-1 and CCL2 expression by NAMPT (100 ngmL) in PDL cells from 10 donors at 3 d (b) Stimulationof MMP-1 expression by various concentrations of NAMPT in PDL cells from 3 donors at 1 d (c) and 3 d (d) Stimulation of CCL2 expressionby various concentrations of NAMPT in PDL cells from 3 donors at 1 d (e) and 3 d (f) Stimulation of MMP-1 protein synthesis by NAMPT(100 ngmL) in PDL cells from 6 donors at 1 d and 3 d (g) Stimulation of CCL2 protein synthesis by NAMPT (100 ngmL) in PDL cells from6 donors at 1 d and 3 d (h) All experiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005)different from NAMPT-untreated cells (control)
PDL cells Interestingly IL-1120573 upregulated significantly theNAMPT expression at 1 d and 3 d (Figure 4(a)) Moreoverthe actions of IL-1120573 on NAMPT expression were dose-dependent at 1 d (Figure 1(b)) but not at 3 d (data not shown)When cells were preincubated with an inhibitor against theMEK12 signaling pathway the IL-1120573-induced stimulation ofNAMPT expression was completely suppressed Moreoverinhibitors against JNK NF120581B and p38 signaling reduced the
stimulatory effect of IL-1120573 onNAMPTby 80 78 and 46respectively
The upregulation of NAMPT at transcriptional level wasparalleled by increased NAMPT protein levels in the super-natants of IL-1120573-stimulated cells at 1 d PDL cells in thepresence and absence of IL-1120573 synthesized 3268 plusmn 362 ngNAMPT protein104 cells and 2028 plusmn 436 ng NAMPTprotein104 cells respectively At 3 d IL-1120573-treated cells and
6 Mediators of Inflammation
50 120583m
0 min 30 min
60 min 90 min
(a)
lowast
10
15
20
25
1dCCL2
lowast
MMP-1
00NAMPT
SP600125+ +
+
++
+
05
minus minus
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
Figure 2 Intracellular pathways triggered by NAMPT Stimulation of NF120581B nuclear translocation by NAMPT at 60min (a) Inhibition ofthe NAMPT-stimulated MMP-1 and CCL2 upregulation by preincubation of PDL cells with SP600125 a selective inhibitor of JNK at 1 d (b)Images and data from one representative donor are shown All experiments were performed in triplicate and repeated at least twice Mean plusmnSEM lowastsignificant (119875 lt 005) difference between groups
Mediators of Inflammation 7
1dlowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(a)
3d
lowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(b)
lowast
87
91d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast87
93d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
1dlowast
20
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
25
(e)
3dlowast
20
25
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(f)
lowast
15
20
251d
0025 00500
OD 0100
1
5
10
0000
lowast
lowast
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(g)
lowast
15
20
253d
lowast
00025 0050OD 0100
1
5
10
0000
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(h)
Figure 3 Regulation of NAMPT andCOX2 expressions by periodontopathogens NAMPT expression in PDL cells from 3 donors in responseto various periodontopathogens (OD 01 P gingivalis Pg T denticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (a) and 3 d(b) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of P gingivalis (OD 0025 0050 and 0100)at 1 d (c) and 3 d (d) COX2 expression in PDL cells from 3 donors in response to various periodontopathogens (OD 01 P gingivalis Pg Tdenticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (e) and 3 d (f) Stimulation of COX2 expression in PDL cells from 3 donorsby various concentrations of P gingivalis (OD 0025 0050 and 0100) at 1 d (g) and 3 d (h) All experiments were performed in triplicate andrepeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from P gingivalis-untreated cells (control)
control cells produced 3097 plusmn 382 ng NAMPT protein104
cells and 2043 plusmn 076 ng NAMPT protein104 cells respec-tivelyThedifferences between groupswere significant at bothtime points The stimulation of NAMPT protein synthesisby IL-1120573 was also observed by immunocytochemistry (Fig-ure 4(c))
IL-1120573 also induced a significant upregulation of COX2expression at 1 d and 3 d (Figure 4(d))
4 Discussion
Our study shows for this first time that NAMPT stimulatesthe production ofCCL2 andMMP-1 in humanPDL cells sug-gesting that NAMPT may contribute to periodontal inflam-mation and matrix destruction through the production ofthese molecules Furthermore NAMPT is induced by theperiodontopathogen P gingivalis and the proinflammatorycytokine IL-1120573 in PDL cells which shows that microbial
8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 3
fitting 2-way ANOVAmodel After model fitting differentialgene expression for all clinically relevant differences can beextracted as contrasts from the model and corresponding 119875values can be computed (multiple testing correction) [23]We refer the reader to the limma user guide for an excellentintroduction [24]
23 Real-Time PCR RNA was extracted using an RNAextraction kit (Qiagen Hilden Germany) and a total of1 120583g of RNA was reverse transcribed using iScriptTM SelectcDNA Synthesis Kit (Bio-Rad Laboratories Munich Ger-many) at 42∘C for 90min followed by 85∘C for 5minExpression of early growth response 1 (EGR1) MMP-1 synaptotagmin 7 (SYT7) inositol 145-trisphosphate 3-kinase a (ITPKA) CCL2 neurotrimin (NTM) insulin-like growth factor 2 mRNA-binding protein 3 (IGF2BP3)neuropilin 1 (NRP1) potassium channel tetramerizationdomain-containing protein 12 (KCTD12) heat-shock 27 kDprotein 3 (HSPB3) transmembrane 4 L six family member20 (TM4SF20) regulator of G protein signaling 4 (RGS4)NAMPT COX2 and glyceraldehyde-3-phosphate dehydro-genase (GAPDH) was detected by real-time PCR usingthe iCycler iQ detection system (Bio-Rad Laboratories)SYBR Green (Bio-Rad Laboratories) and specific primers(QuantiTect Primer Assay Qiagen) One 120583L of cDNA wasamplified as a template in a 25120583L reactionmixture containing125 120583L 2x QuantiFast SYBR Green PCR Master Mix (Qia-gen) 25 120583L of primers and deionized water The mixturewas heated initially at 95∘C for 5min and then followed by40 cycles with denaturation at 95∘C for 10 s and combinedannealingextension at 60∘C for 30 s GAPDH was usedas an endogenous control The data were analyzed by thecomparative threshold cycle (CT) method
24 ELISA The levels of NAMPT MMP-1 and CCL2 in thesupernatants of PDL cells were analysed by commerciallyavailable enzyme-linked immunoassay (ELISA) kits (Ray-Biotech Norcross GAUSA) according to themanufacturerrsquosinstructionsThe absorbancewasmeasuredwith amicroplatereader (PowerWave x BioTek Instruments Winooski VTUSA) at 450 nm The data were normalized by the cellnumber which was measured with an automatic cell counter(Moelab Hilden Germany)
25 Immunocytochemistry PDL cells were cultured in thepresence and absence of P gingivalis ATCC 33277 or IL-1120573 on glass coverslips in 24-well plates After 3 days cellswere fixed in 4 paraformaldehyde (Sigma-AldrichMunichGermany) at pH 74 and room temperature (RT) for 15minand then permeabilized in 01TritonX-100 (Sigma-Aldrich)for 5min Non specific antigens were blocked by incubationwith serum block (LSAB System Santa Cruz BiotechnologySanta Cruz CA USA) for 20min Subsequently cells wereincubated at RT for 90min with rabbit polyclonal antibody toNAMPT (Santa Cruz Biotechnology) Afterwards cells werelabeled with goat anti-rabbit IgG-HRP secondary antibody(Cell Signalling Technology Danvers MA USA) for 45min
For staining cells were exposed to DAB chromogen (DakoHamburg Germany) for 3min at RT After each incubationstep cells were washed twice with PBS (Sigma-Aldrich)Counterstaining was performed with Mayerrsquos Haematoxylin(Merck Eurolab Dietikon Switzerland) for 2min Cover-slips were mounted in DePex mounting medium (ServaElectrophoresis Heidelberg Germany) Standardized pho-tomicrographs were taken using an Axioplan 2 imagingmicroscope (Carl Zeiss MicroImaging Jena Germany)
26 Immunofluorescence PDL cells were fixed with 4paraformaldehyde in PBS pH 74 for 10min at RT washedwith PBS and treated with 01 Triton X-100 for 5min atRTThen cells were washed again with PBS and blocked witha blocking buffer (nonfat dry milk Bio-Rad Laboratories)for 1 h at RT After washing the cells were incubated withprimary rabbit antibody NF120581B p65 (1 400 Cell SignallingTechnology) for 90min and with secondary antibody CY3(1 2000 Abcam Cambridge MA USA) for 45min Cellswere observed under a 20x objective using an Axioplan 2imaging microscope (Carl Zeiss MicroImaging) The imageswere captured with a PVCAM camera and the VisiViewcapturing software (Visitron Systems Puchheim Germany)
27 Statistical Analysis The IBM SPSS Statistics 20 softwarewas used for analysis Mean values and standard errors ofthe mean (SEM) were calculated All experiments were per-formed in triplicate and repeated at least twice For statisticalanalysis parametric (119905 test and ANOVA followed by the post-hoc Dunnettrsquos test) and non parametric tests (Wilcoxon andMann-Whitney 119880 tests) were applied Differences betweengroups were considered significant at 119875 lt 005 Microarraydata were analyzed in the Bonn-Aachen International Centerfor IT Algorithmic Bioinformatics University of BonnGermany (see Section 22)
3 Results
31 Regulation of Gene Expression by NAMPT in PDL CellsFirst we sought to clarify whether NAMPT regulates theexpression of genes in PDL cells by performing amicroarray-based approach As shown in Table 1 NAMPT caused a sig-nificant upregulation of 9 genes and a significant downregula-tion of 3 genes in PDL cells from 3 donors at 1 dTheNAMPT-upregulated genes were the following EGR1 MMP-1 SYT7ITPKA CCL2 NTM IGF2BP3 NRP1 and KCTD12 The 3NAMPT-downregulated genes comprisedHSPB3 TM4SF20and RGS4 (Table 1) In PDL cells from 10 donors the resultsfrom the microarray analysis were then validated by real-time PCRwhich confirmed 8 out of the 12NAMPT-regulatedgenes NAMPT induced a significant upregulation of themRNA expression for EGR1 (24-fold) MMP-1 (38-fold)SYT7 (46-fold) ITPKA (28-fold) CCL2 (23-fold) NTM(21-fold) IGF2BP3 (19-fold) and NRP1 (17-fold) KCTD12was also upregulated by NAMPT but the effect did not reachstatistical significance (Table 2 and Figure 1(a))The effects ofNAMPT on the remaining genes could not be confirmed byPCR (data not shown)
4 Mediators of Inflammation
Table 1 Regulation of gene expression by NAMPT as analyzed bymicroarray
Gene LogFC adj 119875 valueEGR1 2184 0008MMP-1 1545 0010SYT7 1443 0010ITPKA 1292 0010CCL2 1150 0028NTM 1114 0028IGF2BP3 1040 0020NRP1 1024 0010KCTD12 1019 0048HSPB3 minus1035 0030TM4SF20 minus1164 0028RGS4 minus1397 0025Effect of NAMPT on gene expression of PDL cells from 3 donors at 1 d Onlygenes with statistically significant (FDR lt 5) logFC ge 1 (upregulationby at least 2-fold) or logFC le minus1 (downregulation by at least 50)were considered Abbreviations EGR1 early growth response 1 MMP-1matrix metalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12 HSPB3 heat-shock 27-kD protein 3 TM4SF20transmembrane 4 L six family member 20 RGS4 regulator of G proteinsignaling 4
Table 2 Upregulation of gene expression by NAMPT as analyzedby real-time PCR
Gene Fold change(mean) SD SEM 119875 value
EGR1 2422 2306 0421 lt0001MMP-1 3781 3116 0569 lt0001SYT7 4575 7947 1451 lt0001ITPKA 2764 3182 0581 0001CCL2 2285 1346 0246 lt0001NTM 2124 2460 0449 lt0001IGF2BP3 1897 0970 0177 lt0001NRP1 1653 1658 0303 lt0001KCTD12 1713 0958 0175 0095Effect of NAMPT on gene expression of PDL cells from 10 donorsat 1 d Abbreviations EGR1 early growth response 1 MMP-1 matrixmetalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12
32 Effect of NAMPT on MMP-1 and CCL2 Since MMP-1andCCL2 aremolecules known to be strongly associatedwithperiodontitis we further studied their regulation byNAMPTAs observed at 1 d NAMPT increased significantly theMMP-1 (14-fold) and CCL2 (18-fold) expressions at 3 d (Fig-ure 1(b)) Furthermore the stimulatory effects of NAMPTon MMP-1 and CCL2 expressions were dose-dependent atboth time points as shown in Figures 1(c)ndash1(f) In addition
the NAMPT-induced upregulation of the MMP-1 and CCL2expressions was parallelled by increased MMP-1 and CCL2protein levels in the supernatants ofNAMPT-stimulated cellsas compared to those of the control at 1 d and 3 d (Figures 1(g)and 1(h))
33 Exploitation of the JNK Pathway by NAMPT Next westudied how the effects of NAMPT on MMP-1 and CCL2 aremediated intracellularly As evidenced by immunofluores-cence microscopy NAMPT stimulated the nuclear translo-cation of NF120581B and caused a maximal NF120581B accumulationwithin the nucleus at 60min (Figure 2(a)) However pre-incubation of cells with PDTC a specific inhibitor of NF120581Bhad not effect on the NAMPT-stimulated upregulation ofMMP-1 and CCL2 (data not shown) Moreover the upreg-ulation of both molecules by NAMPT was also not affectedby inhibitors against the p38 MEK12 and PI3K signalingpathways However SP600125 an inhibitor of JNK signalingblocked completely the stimulatory effect of NAMPT on theMMP-1 expression at 1 d (Figure 2(b)) In addition SP600125reduced significantly theNAMPT-stimulated upregulation ofCCL2 by 69 at this time point (Figure 2(b))
34 Stimulation of NAMPT Production by P gingivalisWe then wondered whether pathogens that are stronglyassociated with periodontitis are capable of inducing theproduction of NAMPT in PDL cells Whereas T denticolaT forsythia and A actinomycetemcomitans had no significanteffect P gingivalis increased significantly theNAMPTmRNAexpression at 1 d and 3 d (Figures 3(a) and 3(b)) As shown inFigures 3(c) and 3(d) the actions of P gingivalis on NAMPTmRNA expression were dose-dependent Interestingly pre-incubation with a specific inhibitor against NF120581B signalingblocked completely the P gingivalis-induced stimulation ofNAMPT at 1 d Furthermore inhibitors against JNK and p38signaling reduced the stimulatory effects of NAMPT by 87and 37 respectively at this time point
The stimulation of NAMPT expression by P gingivaliswas also observed at protein level as analyzed by ELISA at1 d PDL cells in the presence and absence of P gingivalisproduced 2883 plusmn 029 ng NAMPT protein104 cells and1791 plusmn 058 ng NAMPT protein104 cells respectively At3 d P gingivalis-stimulated cells and control cells synthesized3097 plusmn 309 ng NAMPT protein104 cells and 2165 plusmn294 ng NAMPT protein104 cells respectively The differ-ences between both groups at each time point were sig-nificant The stimulation of NAMPT protein production byP gingivalis was also evidenced by immunocytochemistry(Figure 4(c))
Like NAMPT COX2 was significantly upregulated by Pgingivalis at 1 d and 3 d (Figures 3(e) and 3(f)) By contrast allother periodontopathogens were not capable of stimulatingthe COX2 expression at both time points (Figures 3(e)and 3(f)) The P gingivalis-induced stimulation of COX2expression was dose-dependent (Figures 3(g) and 3(h))
35 Stimulation of NAMPT Production by IL-1120573 Finallywe examined whether NAMPT is also induced by IL-1120573 in
Mediators of Inflammation 5
1d
5
6
7
8
0
2
1
3
4
EGR1 MMP-1 SYT7 ITPKA CCL2 NTM IGF2BP3 NRP1 KCTD12
lowast
lowast
lowast
lowast
mRN
A ex
pres
sion
(fold
of c
ontro
l)
lowast
lowastlowast lowast lowast
(a)
lowast15
20MMP-1 CCL2
lowast 3d
00
05
NAMPT + +minusminus
10
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
lowast
lowast
lowast
2
3
1d4
1
030 100NAMPT
(ngmL)3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast
lowast
2
3
3d4
10
30 100NAMPT(ngmL)
3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
lowast
2
3
1d4
lowast
1
030 100NAMPT
(ngmL)3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(e)
lowast2
3
3d4
lowastlowast
10
30 100NAMPT(ngmL)
3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(f)
125
75100
175150
2001d
lowast
3d
02550
lowast
NAMPT + +minusminus
MM
P-1
prot
ein
(pg10
3ce
lls)
(g)
50
203040
7060
801d
lowast
lowast
3d
010
NAMPT + +minus minus
CCL2
pro
tein
(pg10
4ce
lls)
(h)
Figure 1 Stimulation of gene expression and protein synthesis by NAMPT Upregulation of genes by NAMPT (100 ngmL) in PDL cells from10 donors at 1 d (a) Upregulation ofMMP-1 and CCL2 expression by NAMPT (100 ngmL) in PDL cells from 10 donors at 3 d (b) Stimulationof MMP-1 expression by various concentrations of NAMPT in PDL cells from 3 donors at 1 d (c) and 3 d (d) Stimulation of CCL2 expressionby various concentrations of NAMPT in PDL cells from 3 donors at 1 d (e) and 3 d (f) Stimulation of MMP-1 protein synthesis by NAMPT(100 ngmL) in PDL cells from 6 donors at 1 d and 3 d (g) Stimulation of CCL2 protein synthesis by NAMPT (100 ngmL) in PDL cells from6 donors at 1 d and 3 d (h) All experiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005)different from NAMPT-untreated cells (control)
PDL cells Interestingly IL-1120573 upregulated significantly theNAMPT expression at 1 d and 3 d (Figure 4(a)) Moreoverthe actions of IL-1120573 on NAMPT expression were dose-dependent at 1 d (Figure 1(b)) but not at 3 d (data not shown)When cells were preincubated with an inhibitor against theMEK12 signaling pathway the IL-1120573-induced stimulation ofNAMPT expression was completely suppressed Moreoverinhibitors against JNK NF120581B and p38 signaling reduced the
stimulatory effect of IL-1120573 onNAMPTby 80 78 and 46respectively
The upregulation of NAMPT at transcriptional level wasparalleled by increased NAMPT protein levels in the super-natants of IL-1120573-stimulated cells at 1 d PDL cells in thepresence and absence of IL-1120573 synthesized 3268 plusmn 362 ngNAMPT protein104 cells and 2028 plusmn 436 ng NAMPTprotein104 cells respectively At 3 d IL-1120573-treated cells and
6 Mediators of Inflammation
50 120583m
0 min 30 min
60 min 90 min
(a)
lowast
10
15
20
25
1dCCL2
lowast
MMP-1
00NAMPT
SP600125+ +
+
++
+
05
minus minus
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
Figure 2 Intracellular pathways triggered by NAMPT Stimulation of NF120581B nuclear translocation by NAMPT at 60min (a) Inhibition ofthe NAMPT-stimulated MMP-1 and CCL2 upregulation by preincubation of PDL cells with SP600125 a selective inhibitor of JNK at 1 d (b)Images and data from one representative donor are shown All experiments were performed in triplicate and repeated at least twice Mean plusmnSEM lowastsignificant (119875 lt 005) difference between groups
Mediators of Inflammation 7
1dlowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(a)
3d
lowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(b)
lowast
87
91d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast87
93d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
1dlowast
20
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
25
(e)
3dlowast
20
25
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(f)
lowast
15
20
251d
0025 00500
OD 0100
1
5
10
0000
lowast
lowast
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(g)
lowast
15
20
253d
lowast
00025 0050OD 0100
1
5
10
0000
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(h)
Figure 3 Regulation of NAMPT andCOX2 expressions by periodontopathogens NAMPT expression in PDL cells from 3 donors in responseto various periodontopathogens (OD 01 P gingivalis Pg T denticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (a) and 3 d(b) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of P gingivalis (OD 0025 0050 and 0100)at 1 d (c) and 3 d (d) COX2 expression in PDL cells from 3 donors in response to various periodontopathogens (OD 01 P gingivalis Pg Tdenticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (e) and 3 d (f) Stimulation of COX2 expression in PDL cells from 3 donorsby various concentrations of P gingivalis (OD 0025 0050 and 0100) at 1 d (g) and 3 d (h) All experiments were performed in triplicate andrepeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from P gingivalis-untreated cells (control)
control cells produced 3097 plusmn 382 ng NAMPT protein104
cells and 2043 plusmn 076 ng NAMPT protein104 cells respec-tivelyThedifferences between groupswere significant at bothtime points The stimulation of NAMPT protein synthesisby IL-1120573 was also observed by immunocytochemistry (Fig-ure 4(c))
IL-1120573 also induced a significant upregulation of COX2expression at 1 d and 3 d (Figure 4(d))
4 Discussion
Our study shows for this first time that NAMPT stimulatesthe production ofCCL2 andMMP-1 in humanPDL cells sug-gesting that NAMPT may contribute to periodontal inflam-mation and matrix destruction through the production ofthese molecules Furthermore NAMPT is induced by theperiodontopathogen P gingivalis and the proinflammatorycytokine IL-1120573 in PDL cells which shows that microbial
8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

4 Mediators of Inflammation
Table 1 Regulation of gene expression by NAMPT as analyzed bymicroarray
Gene LogFC adj 119875 valueEGR1 2184 0008MMP-1 1545 0010SYT7 1443 0010ITPKA 1292 0010CCL2 1150 0028NTM 1114 0028IGF2BP3 1040 0020NRP1 1024 0010KCTD12 1019 0048HSPB3 minus1035 0030TM4SF20 minus1164 0028RGS4 minus1397 0025Effect of NAMPT on gene expression of PDL cells from 3 donors at 1 d Onlygenes with statistically significant (FDR lt 5) logFC ge 1 (upregulationby at least 2-fold) or logFC le minus1 (downregulation by at least 50)were considered Abbreviations EGR1 early growth response 1 MMP-1matrix metalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12 HSPB3 heat-shock 27-kD protein 3 TM4SF20transmembrane 4 L six family member 20 RGS4 regulator of G proteinsignaling 4
Table 2 Upregulation of gene expression by NAMPT as analyzedby real-time PCR
Gene Fold change(mean) SD SEM 119875 value
EGR1 2422 2306 0421 lt0001MMP-1 3781 3116 0569 lt0001SYT7 4575 7947 1451 lt0001ITPKA 2764 3182 0581 0001CCL2 2285 1346 0246 lt0001NTM 2124 2460 0449 lt0001IGF2BP3 1897 0970 0177 lt0001NRP1 1653 1658 0303 lt0001KCTD12 1713 0958 0175 0095Effect of NAMPT on gene expression of PDL cells from 10 donorsat 1 d Abbreviations EGR1 early growth response 1 MMP-1 matrixmetalloproteinase-1 SYT7 synaptotagmin 7 ITPKA inositol 145-trisphosphate 3-kinase a CCL2 chemokine cc motif ligand 2 NTMneurotrimin IGF2BP3 insulin-like growth factor 2 mRNA-binding protein3 NRP1 neuropilin 1 KCTD12 potassium channel tetramerization domain-containing protein 12
32 Effect of NAMPT on MMP-1 and CCL2 Since MMP-1andCCL2 aremolecules known to be strongly associatedwithperiodontitis we further studied their regulation byNAMPTAs observed at 1 d NAMPT increased significantly theMMP-1 (14-fold) and CCL2 (18-fold) expressions at 3 d (Fig-ure 1(b)) Furthermore the stimulatory effects of NAMPTon MMP-1 and CCL2 expressions were dose-dependent atboth time points as shown in Figures 1(c)ndash1(f) In addition
the NAMPT-induced upregulation of the MMP-1 and CCL2expressions was parallelled by increased MMP-1 and CCL2protein levels in the supernatants ofNAMPT-stimulated cellsas compared to those of the control at 1 d and 3 d (Figures 1(g)and 1(h))
33 Exploitation of the JNK Pathway by NAMPT Next westudied how the effects of NAMPT on MMP-1 and CCL2 aremediated intracellularly As evidenced by immunofluores-cence microscopy NAMPT stimulated the nuclear translo-cation of NF120581B and caused a maximal NF120581B accumulationwithin the nucleus at 60min (Figure 2(a)) However pre-incubation of cells with PDTC a specific inhibitor of NF120581Bhad not effect on the NAMPT-stimulated upregulation ofMMP-1 and CCL2 (data not shown) Moreover the upreg-ulation of both molecules by NAMPT was also not affectedby inhibitors against the p38 MEK12 and PI3K signalingpathways However SP600125 an inhibitor of JNK signalingblocked completely the stimulatory effect of NAMPT on theMMP-1 expression at 1 d (Figure 2(b)) In addition SP600125reduced significantly theNAMPT-stimulated upregulation ofCCL2 by 69 at this time point (Figure 2(b))
34 Stimulation of NAMPT Production by P gingivalisWe then wondered whether pathogens that are stronglyassociated with periodontitis are capable of inducing theproduction of NAMPT in PDL cells Whereas T denticolaT forsythia and A actinomycetemcomitans had no significanteffect P gingivalis increased significantly theNAMPTmRNAexpression at 1 d and 3 d (Figures 3(a) and 3(b)) As shown inFigures 3(c) and 3(d) the actions of P gingivalis on NAMPTmRNA expression were dose-dependent Interestingly pre-incubation with a specific inhibitor against NF120581B signalingblocked completely the P gingivalis-induced stimulation ofNAMPT at 1 d Furthermore inhibitors against JNK and p38signaling reduced the stimulatory effects of NAMPT by 87and 37 respectively at this time point
The stimulation of NAMPT expression by P gingivaliswas also observed at protein level as analyzed by ELISA at1 d PDL cells in the presence and absence of P gingivalisproduced 2883 plusmn 029 ng NAMPT protein104 cells and1791 plusmn 058 ng NAMPT protein104 cells respectively At3 d P gingivalis-stimulated cells and control cells synthesized3097 plusmn 309 ng NAMPT protein104 cells and 2165 plusmn294 ng NAMPT protein104 cells respectively The differ-ences between both groups at each time point were sig-nificant The stimulation of NAMPT protein production byP gingivalis was also evidenced by immunocytochemistry(Figure 4(c))
Like NAMPT COX2 was significantly upregulated by Pgingivalis at 1 d and 3 d (Figures 3(e) and 3(f)) By contrast allother periodontopathogens were not capable of stimulatingthe COX2 expression at both time points (Figures 3(e)and 3(f)) The P gingivalis-induced stimulation of COX2expression was dose-dependent (Figures 3(g) and 3(h))
35 Stimulation of NAMPT Production by IL-1120573 Finallywe examined whether NAMPT is also induced by IL-1120573 in
Mediators of Inflammation 5
1d
5
6
7
8
0
2
1
3
4
EGR1 MMP-1 SYT7 ITPKA CCL2 NTM IGF2BP3 NRP1 KCTD12
lowast
lowast
lowast
lowast
mRN
A ex
pres
sion
(fold
of c
ontro
l)
lowast
lowastlowast lowast lowast
(a)
lowast15
20MMP-1 CCL2
lowast 3d
00
05
NAMPT + +minusminus
10
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
lowast
lowast
lowast
2
3
1d4
1
030 100NAMPT
(ngmL)3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast
lowast
2
3
3d4
10
30 100NAMPT(ngmL)
3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
lowast
2
3
1d4
lowast
1
030 100NAMPT
(ngmL)3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(e)
lowast2
3
3d4
lowastlowast
10
30 100NAMPT(ngmL)
3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(f)
125
75100
175150
2001d
lowast
3d
02550
lowast
NAMPT + +minusminus
MM
P-1
prot
ein
(pg10
3ce
lls)
(g)
50
203040
7060
801d
lowast
lowast
3d
010
NAMPT + +minus minus
CCL2
pro
tein
(pg10
4ce
lls)
(h)
Figure 1 Stimulation of gene expression and protein synthesis by NAMPT Upregulation of genes by NAMPT (100 ngmL) in PDL cells from10 donors at 1 d (a) Upregulation ofMMP-1 and CCL2 expression by NAMPT (100 ngmL) in PDL cells from 10 donors at 3 d (b) Stimulationof MMP-1 expression by various concentrations of NAMPT in PDL cells from 3 donors at 1 d (c) and 3 d (d) Stimulation of CCL2 expressionby various concentrations of NAMPT in PDL cells from 3 donors at 1 d (e) and 3 d (f) Stimulation of MMP-1 protein synthesis by NAMPT(100 ngmL) in PDL cells from 6 donors at 1 d and 3 d (g) Stimulation of CCL2 protein synthesis by NAMPT (100 ngmL) in PDL cells from6 donors at 1 d and 3 d (h) All experiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005)different from NAMPT-untreated cells (control)
PDL cells Interestingly IL-1120573 upregulated significantly theNAMPT expression at 1 d and 3 d (Figure 4(a)) Moreoverthe actions of IL-1120573 on NAMPT expression were dose-dependent at 1 d (Figure 1(b)) but not at 3 d (data not shown)When cells were preincubated with an inhibitor against theMEK12 signaling pathway the IL-1120573-induced stimulation ofNAMPT expression was completely suppressed Moreoverinhibitors against JNK NF120581B and p38 signaling reduced the
stimulatory effect of IL-1120573 onNAMPTby 80 78 and 46respectively
The upregulation of NAMPT at transcriptional level wasparalleled by increased NAMPT protein levels in the super-natants of IL-1120573-stimulated cells at 1 d PDL cells in thepresence and absence of IL-1120573 synthesized 3268 plusmn 362 ngNAMPT protein104 cells and 2028 plusmn 436 ng NAMPTprotein104 cells respectively At 3 d IL-1120573-treated cells and
6 Mediators of Inflammation
50 120583m
0 min 30 min
60 min 90 min
(a)
lowast
10
15
20
25
1dCCL2
lowast
MMP-1
00NAMPT
SP600125+ +
+
++
+
05
minus minus
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
Figure 2 Intracellular pathways triggered by NAMPT Stimulation of NF120581B nuclear translocation by NAMPT at 60min (a) Inhibition ofthe NAMPT-stimulated MMP-1 and CCL2 upregulation by preincubation of PDL cells with SP600125 a selective inhibitor of JNK at 1 d (b)Images and data from one representative donor are shown All experiments were performed in triplicate and repeated at least twice Mean plusmnSEM lowastsignificant (119875 lt 005) difference between groups
Mediators of Inflammation 7
1dlowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(a)
3d
lowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(b)
lowast
87
91d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast87
93d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
1dlowast
20
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
25
(e)
3dlowast
20
25
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(f)
lowast
15
20
251d
0025 00500
OD 0100
1
5
10
0000
lowast
lowast
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(g)
lowast
15
20
253d
lowast
00025 0050OD 0100
1
5
10
0000
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(h)
Figure 3 Regulation of NAMPT andCOX2 expressions by periodontopathogens NAMPT expression in PDL cells from 3 donors in responseto various periodontopathogens (OD 01 P gingivalis Pg T denticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (a) and 3 d(b) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of P gingivalis (OD 0025 0050 and 0100)at 1 d (c) and 3 d (d) COX2 expression in PDL cells from 3 donors in response to various periodontopathogens (OD 01 P gingivalis Pg Tdenticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (e) and 3 d (f) Stimulation of COX2 expression in PDL cells from 3 donorsby various concentrations of P gingivalis (OD 0025 0050 and 0100) at 1 d (g) and 3 d (h) All experiments were performed in triplicate andrepeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from P gingivalis-untreated cells (control)
control cells produced 3097 plusmn 382 ng NAMPT protein104
cells and 2043 plusmn 076 ng NAMPT protein104 cells respec-tivelyThedifferences between groupswere significant at bothtime points The stimulation of NAMPT protein synthesisby IL-1120573 was also observed by immunocytochemistry (Fig-ure 4(c))
IL-1120573 also induced a significant upregulation of COX2expression at 1 d and 3 d (Figure 4(d))
4 Discussion
Our study shows for this first time that NAMPT stimulatesthe production ofCCL2 andMMP-1 in humanPDL cells sug-gesting that NAMPT may contribute to periodontal inflam-mation and matrix destruction through the production ofthese molecules Furthermore NAMPT is induced by theperiodontopathogen P gingivalis and the proinflammatorycytokine IL-1120573 in PDL cells which shows that microbial
8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 5
1d
5
6
7
8
0
2
1
3
4
EGR1 MMP-1 SYT7 ITPKA CCL2 NTM IGF2BP3 NRP1 KCTD12
lowast
lowast
lowast
lowast
mRN
A ex
pres
sion
(fold
of c
ontro
l)
lowast
lowastlowast lowast lowast
(a)
lowast15
20MMP-1 CCL2
lowast 3d
00
05
NAMPT + +minusminus
10
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
lowast
lowast
lowast
2
3
1d4
1
030 100NAMPT
(ngmL)3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast
lowast
2
3
3d4
10
30 100NAMPT(ngmL)
3000
MM
P-1
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
lowast
2
3
1d4
lowast
1
030 100NAMPT
(ngmL)3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(e)
lowast2
3
3d4
lowastlowast
10
30 100NAMPT(ngmL)
3000
CCL2
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(f)
125
75100
175150
2001d
lowast
3d
02550
lowast
NAMPT + +minusminus
MM
P-1
prot
ein
(pg10
3ce
lls)
(g)
50
203040
7060
801d
lowast
lowast
3d
010
NAMPT + +minus minus
CCL2
pro
tein
(pg10
4ce
lls)
(h)
Figure 1 Stimulation of gene expression and protein synthesis by NAMPT Upregulation of genes by NAMPT (100 ngmL) in PDL cells from10 donors at 1 d (a) Upregulation ofMMP-1 and CCL2 expression by NAMPT (100 ngmL) in PDL cells from 10 donors at 3 d (b) Stimulationof MMP-1 expression by various concentrations of NAMPT in PDL cells from 3 donors at 1 d (c) and 3 d (d) Stimulation of CCL2 expressionby various concentrations of NAMPT in PDL cells from 3 donors at 1 d (e) and 3 d (f) Stimulation of MMP-1 protein synthesis by NAMPT(100 ngmL) in PDL cells from 6 donors at 1 d and 3 d (g) Stimulation of CCL2 protein synthesis by NAMPT (100 ngmL) in PDL cells from6 donors at 1 d and 3 d (h) All experiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005)different from NAMPT-untreated cells (control)
PDL cells Interestingly IL-1120573 upregulated significantly theNAMPT expression at 1 d and 3 d (Figure 4(a)) Moreoverthe actions of IL-1120573 on NAMPT expression were dose-dependent at 1 d (Figure 1(b)) but not at 3 d (data not shown)When cells were preincubated with an inhibitor against theMEK12 signaling pathway the IL-1120573-induced stimulation ofNAMPT expression was completely suppressed Moreoverinhibitors against JNK NF120581B and p38 signaling reduced the
stimulatory effect of IL-1120573 onNAMPTby 80 78 and 46respectively
The upregulation of NAMPT at transcriptional level wasparalleled by increased NAMPT protein levels in the super-natants of IL-1120573-stimulated cells at 1 d PDL cells in thepresence and absence of IL-1120573 synthesized 3268 plusmn 362 ngNAMPT protein104 cells and 2028 plusmn 436 ng NAMPTprotein104 cells respectively At 3 d IL-1120573-treated cells and
6 Mediators of Inflammation
50 120583m
0 min 30 min
60 min 90 min
(a)
lowast
10
15
20
25
1dCCL2
lowast
MMP-1
00NAMPT
SP600125+ +
+
++
+
05
minus minus
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
Figure 2 Intracellular pathways triggered by NAMPT Stimulation of NF120581B nuclear translocation by NAMPT at 60min (a) Inhibition ofthe NAMPT-stimulated MMP-1 and CCL2 upregulation by preincubation of PDL cells with SP600125 a selective inhibitor of JNK at 1 d (b)Images and data from one representative donor are shown All experiments were performed in triplicate and repeated at least twice Mean plusmnSEM lowastsignificant (119875 lt 005) difference between groups
Mediators of Inflammation 7
1dlowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(a)
3d
lowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(b)
lowast
87
91d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast87
93d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
1dlowast
20
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
25
(e)
3dlowast
20
25
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(f)
lowast
15
20
251d
0025 00500
OD 0100
1
5
10
0000
lowast
lowast
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(g)
lowast
15
20
253d
lowast
00025 0050OD 0100
1
5
10
0000
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(h)
Figure 3 Regulation of NAMPT andCOX2 expressions by periodontopathogens NAMPT expression in PDL cells from 3 donors in responseto various periodontopathogens (OD 01 P gingivalis Pg T denticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (a) and 3 d(b) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of P gingivalis (OD 0025 0050 and 0100)at 1 d (c) and 3 d (d) COX2 expression in PDL cells from 3 donors in response to various periodontopathogens (OD 01 P gingivalis Pg Tdenticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (e) and 3 d (f) Stimulation of COX2 expression in PDL cells from 3 donorsby various concentrations of P gingivalis (OD 0025 0050 and 0100) at 1 d (g) and 3 d (h) All experiments were performed in triplicate andrepeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from P gingivalis-untreated cells (control)
control cells produced 3097 plusmn 382 ng NAMPT protein104
cells and 2043 plusmn 076 ng NAMPT protein104 cells respec-tivelyThedifferences between groupswere significant at bothtime points The stimulation of NAMPT protein synthesisby IL-1120573 was also observed by immunocytochemistry (Fig-ure 4(c))
IL-1120573 also induced a significant upregulation of COX2expression at 1 d and 3 d (Figure 4(d))
4 Discussion
Our study shows for this first time that NAMPT stimulatesthe production ofCCL2 andMMP-1 in humanPDL cells sug-gesting that NAMPT may contribute to periodontal inflam-mation and matrix destruction through the production ofthese molecules Furthermore NAMPT is induced by theperiodontopathogen P gingivalis and the proinflammatorycytokine IL-1120573 in PDL cells which shows that microbial
8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

6 Mediators of Inflammation
50 120583m
0 min 30 min
60 min 90 min
(a)
lowast
10
15
20
25
1dCCL2
lowast
MMP-1
00NAMPT
SP600125+ +
+
++
+
05
minus minus
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(b)
Figure 2 Intracellular pathways triggered by NAMPT Stimulation of NF120581B nuclear translocation by NAMPT at 60min (a) Inhibition ofthe NAMPT-stimulated MMP-1 and CCL2 upregulation by preincubation of PDL cells with SP600125 a selective inhibitor of JNK at 1 d (b)Images and data from one representative donor are shown All experiments were performed in triplicate and repeated at least twice Mean plusmnSEM lowastsignificant (119875 lt 005) difference between groups
Mediators of Inflammation 7
1dlowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(a)
3d
lowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(b)
lowast
87
91d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast87
93d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
1dlowast
20
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
25
(e)
3dlowast
20
25
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(f)
lowast
15
20
251d
0025 00500
OD 0100
1
5
10
0000
lowast
lowast
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(g)
lowast
15
20
253d
lowast
00025 0050OD 0100
1
5
10
0000
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(h)
Figure 3 Regulation of NAMPT andCOX2 expressions by periodontopathogens NAMPT expression in PDL cells from 3 donors in responseto various periodontopathogens (OD 01 P gingivalis Pg T denticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (a) and 3 d(b) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of P gingivalis (OD 0025 0050 and 0100)at 1 d (c) and 3 d (d) COX2 expression in PDL cells from 3 donors in response to various periodontopathogens (OD 01 P gingivalis Pg Tdenticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (e) and 3 d (f) Stimulation of COX2 expression in PDL cells from 3 donorsby various concentrations of P gingivalis (OD 0025 0050 and 0100) at 1 d (g) and 3 d (h) All experiments were performed in triplicate andrepeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from P gingivalis-untreated cells (control)
control cells produced 3097 plusmn 382 ng NAMPT protein104
cells and 2043 plusmn 076 ng NAMPT protein104 cells respec-tivelyThedifferences between groupswere significant at bothtime points The stimulation of NAMPT protein synthesisby IL-1120573 was also observed by immunocytochemistry (Fig-ure 4(c))
IL-1120573 also induced a significant upregulation of COX2expression at 1 d and 3 d (Figure 4(d))
4 Discussion
Our study shows for this first time that NAMPT stimulatesthe production ofCCL2 andMMP-1 in humanPDL cells sug-gesting that NAMPT may contribute to periodontal inflam-mation and matrix destruction through the production ofthese molecules Furthermore NAMPT is induced by theperiodontopathogen P gingivalis and the proinflammatorycytokine IL-1120573 in PDL cells which shows that microbial
8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
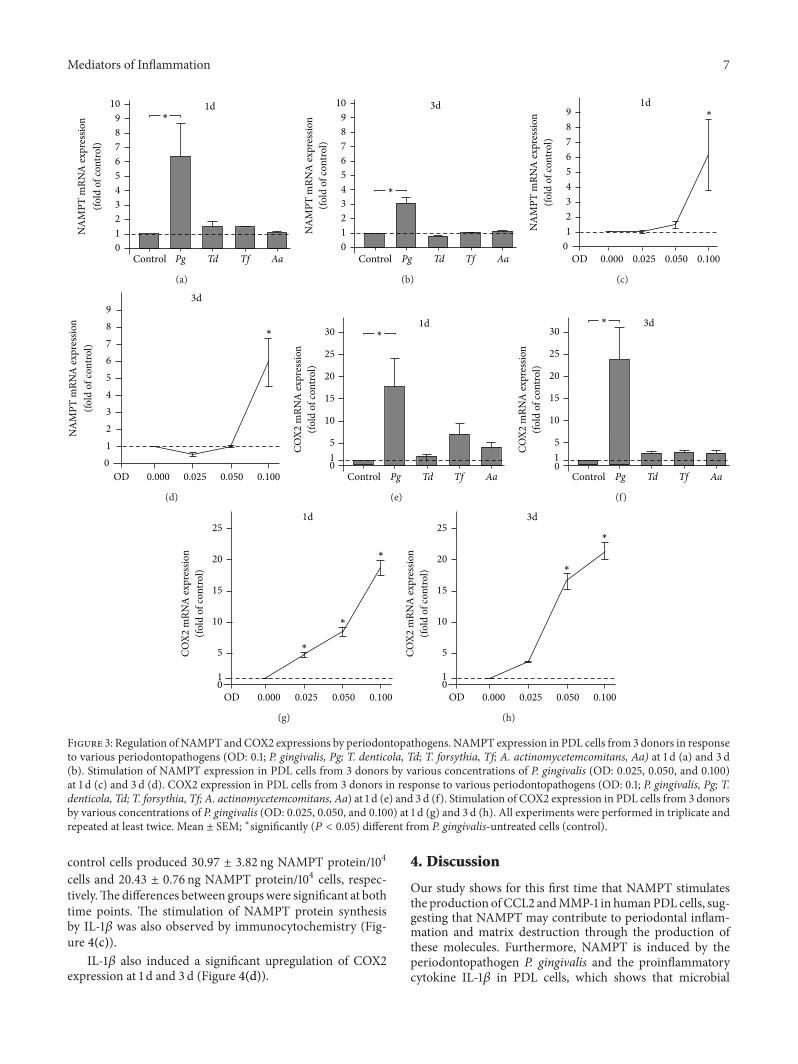
Mediators of Inflammation 7
1dlowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(a)
3d
lowast
0
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
21
3456789
10
Control Pg Td Tf Aa
(b)
lowast
87
91d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(c)
lowast87
93d
0
5
1234
6
0025 0050OD 01000000
NA
MPT
mRN
A ex
pres
sion
(fold
of c
ontro
l)
(d)
1dlowast
20
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
25
(e)
3dlowast
20
25
30
0
51
10
15
Control Pg Td Tf Aa
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(f)
lowast
15
20
251d
0025 00500
OD 0100
1
5
10
0000
lowast
lowast
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(g)
lowast
15
20
253d
lowast
00025 0050OD 0100
1
5
10
0000
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
(h)
Figure 3 Regulation of NAMPT andCOX2 expressions by periodontopathogens NAMPT expression in PDL cells from 3 donors in responseto various periodontopathogens (OD 01 P gingivalis Pg T denticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (a) and 3 d(b) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of P gingivalis (OD 0025 0050 and 0100)at 1 d (c) and 3 d (d) COX2 expression in PDL cells from 3 donors in response to various periodontopathogens (OD 01 P gingivalis Pg Tdenticola Td T forsythia Tf A actinomycetemcomitans Aa) at 1 d (e) and 3 d (f) Stimulation of COX2 expression in PDL cells from 3 donorsby various concentrations of P gingivalis (OD 0025 0050 and 0100) at 1 d (g) and 3 d (h) All experiments were performed in triplicate andrepeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from P gingivalis-untreated cells (control)
control cells produced 3097 plusmn 382 ng NAMPT protein104
cells and 2043 plusmn 076 ng NAMPT protein104 cells respec-tivelyThedifferences between groupswere significant at bothtime points The stimulation of NAMPT protein synthesisby IL-1120573 was also observed by immunocytochemistry (Fig-ure 4(c))
IL-1120573 also induced a significant upregulation of COX2expression at 1 d and 3 d (Figure 4(d))
4 Discussion
Our study shows for this first time that NAMPT stimulatesthe production ofCCL2 andMMP-1 in humanPDL cells sug-gesting that NAMPT may contribute to periodontal inflam-mation and matrix destruction through the production ofthese molecules Furthermore NAMPT is induced by theperiodontopathogen P gingivalis and the proinflammatorycytokine IL-1120573 in PDL cells which shows that microbial
8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

8 Mediators of Inflammation
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
1
2
3
4
1d 3dlowast
0++minus minusIL-1120573
(a)
lowast
1d
15
5
10
25
NA
MPT
mRN
A ex
pres
sion
(fold
of co
ntro
l) 20
lowast
002 10
(ngmL)50
1
00IL-1120573
(b)
Control P gingivalis
3d
IL-1120573
(c)
1d 3d
COX2
mRN
A ex
pres
sion
(fold
of co
ntro
l)
lowast
0
1
2
3
4
lowast
+ +minus minusIL-1120573
(d)
Figure 4 Regulation of NAMPT and COX2 expressions by IL-1120573 NAMPT expression in PDL cells from 6 donors in response to IL-1120573(1 ngmL) at 1 d and 3 d (a) Stimulation of NAMPT expression in PDL cells from 3 donors by various concentrations of IL-1120573 (02 10and 50 ngmL) at 1 d (b) NAMPT protein synthesis in PDL cells in the presence and absence of IL-1120573 and P gingivalis at 3 d Images fromone representative donor are shown (c) COX2 expression in PDL cells from 6 donors in response to IL-1120573 (1 ngmL) at 1 d and 3 d (d) Allexperiments were performed in triplicate and repeated at least twice Mean plusmn SEM lowastsignificantly (119875 lt 005) different from IL-1120573-untreatedcells (control)
and inflammatory signals may use this adipokine for theirdetrimental effects on the periodontium
MMP-1 degrades specifically type I collagen and addi-tionally types II III V IX and X collagen thereby playinga critical role for modeling and remodeling of the extracel-lular matrix [25] Several studies have revealed that MMP-1 levels are increased in GCF and human gingiva fromperiodontitis patients [26 27] Furthermore MMP-1 levels
could be significantly reduced by periodontal treatment [27]CCL2 is synthesized by a variety of cell types and regulatesthe migration and infiltration of monocytes memory Tlymphocytes and natural killer cells In addition CCL2 seemto influence T-cell immunity [28] Higher levels of CCL2 inGCF and gingiva from inflamed sites have been reported [20ndash31] Our experiments provide original evidence that NAMPTinduces increased expression and synthesis of MMP-1 and
Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 9
CCL2 in human PDL cells as evidenced by several assaysand in a high number of donors These findings are inaccordance with previous observations in nonperiodontalcells and underline the proinflammatory and catabolic roleof NAMPT in the pathophysiology of periodontitis [32ndash34]Since increased NAMPT levels are found in obesity our datasuggest at least one mechanism whereby obesity could conferan increased risk of periodontitis in obese individuals [5 9]
Surprisingly NAMPT also upregulated a number ofgenes most of which have mainly been reported in oncol-ogy and neurosciences The transcription factor EGR1 is aregulator of multiple tumor suppressors but paradoxicallycan also promote tumor progression [35 36] EGR1 is alteredby hypoxia [37] stimulates angiogenesis [35] and regulatesthe immune system [38] Synaptotagmins are transmem-brane proteins with two Ca2+-binding C(2) domains in theircytosolic region Syt7 regulates the exocytosis of lysosomesand thereby plays a role in bone resorption [39] cell migra-tion [40] CTL responses [41] and neurodegeneration [42]ITPKA phosphorylates inositol 145-trisphosphate therebymodulating the calcium (Ca2+) level within the cell and thelevels of a large number of inositol polyphosphates [43]ITPKA promotes cell motility and the metastatic potential oftumor cells [43 44] Furthermore ITPKA is critical for thespatial and temporal regulation of spine actin remodelingsynaptic plasticity and learning and memory [45] Little isknown about neurotrimin a neural cell adhesion protein Ithas been shown to be an estrogen-regulated determinant ofperipheral sympathetic innervation [46] and to play a rolein axonal fasciculation of specific cerebellar systems and theformation of excitatory synapses and their stabilization [47]The IGF2BPs bind to the 51015840-untranslated region of IGF-2mRNA and regulate a number of important aspects of cellfunction such as cell polarization morphology migrationproliferation differentiation and invasion [48 49] NRP1plays a critical part in neuronal development in angiogenesisand tumor invasion [50 51] NRP1 is a coreceptor formembers of the vascular endothelial growth factor familyand the class 3 semaphorins which are polypeptides withroles in axonal guidance [52] Furthermore NRP1 mediatesinteraction of regulatory T cells with dendritic cells andmodulates the immune response [53] KCTDs act as auxiliarysubunits of GABA(B) receptors and generate desensitizingreceptor responses [54] Further studies should clarify thephysiological and pathological role of these genes in theperiodontium
We also sought to examine the intracellular mechanismwhereby the stimulatory effects of NAMPT on MMP-1 andCCL2 are accomplished Although NAMPT stimulated theNF120581B nuclear translocation in PDL cells which concurs withobservations in other cells [32 55 56] pre-incubation of cellswith a specific NF120581B inhibitor did not abrogate the NAMPTactions on MMP-1 and CCL2 in our cells However theNF120581B signaling pathway could be involved in the NAMPT-induced upregulation of some of the other genes which wereidentified by microarray and PCR analyses Interestingly ourexperiments revealed that theNAMPT-induced upregulation
ofMMP-1 and CCL2was JNK dependent which is supportedby findings from others [32]
Recently increased NAMPT levels have been found inGCF gingival tissues and serum from periodontally diseasedpatients as compared to periodontally healthy individualsThis suggests that NAMPT might also be produced locallyin the periodontium and regulated by periodontopathogensandor inflammatory mediators [12 13] Interestingly wefound that P gingivalis a key pathogen associated withperiodontitis and the proinflammatory cytokine IL-1120573which is increased at inflamed periodontal sites can induceNAMPT in PDL cells thereby supporting the assumptionthat NAMPT is locally produced in the presence of an infec-tious andor inflammatory environment By upregulationof MMP-1 and CCL2 NAMPT may mediate and amplifyproinflammatory and proteolytic actions of P gingivalis andIL-1120573 on the PDL The finding that NAMPT is induced by Pgingivalis is in linewith our previous experiments which havedemonstrated that NAMPT is upregulated by F nucleatuma gram-negative anaerobic microorganism which acts as abridge bacterium between early and late colonizers duringplaque development [57] Interestingly T forsythia T den-ticola and A actinomycetemcomitans did not regulate theNAMPT expression at least at the concentration tested in thepresent study Our observation that NAMPT is increased byIL-1120573 in PDL cells concurs with findings in other cells whichhave also been shown to produce increased NAMPT levelsin response to IL-1120573 [58ndash60] Our data provide evidencethat PDL cells produce increased levels of NAMPT underinfectious and inflammatory conditions which suggests thatlocal production of NAMPT in the inflamed periodontiumcould contribute to the increased gingival and serum levels ofNAMPT as observed in patients with periodontitis NAMPTcould therefore also represent a pathomechanistic link howperiodontitis affects systemic diseases such as diabetes mel-litus and cardiovascular diseases
Interestingly the upregulation of NAMPT by P gingivaliscould be completely blocked by an inhibitor against NF120581Band reduced by inhibitors against the JNK and p38 signalingpathways Recently other investigators have also shown thatP gingivalis induces NF120581B activation and p38 signaling forits actions [61 62] An inhibitor against MEK12 signalingpathway suppressed completely the IL-1120573-induced stimula-tion of NAMPT expression JNK NF120581B and p38 signalingwere also found to be involved in the stimulatory effects ofIL-1120573 onNAMPTThat IL-1120573 exploits theNF120581B andmitogen-activated protein kinase pathways for its actions is well knownand supported by the present findings [63 64]
In order to simulate an infectious environment PDLcells were incubated with a suspension of various periodon-topathogens Since the suspensions were exposed to intensiveultrasonication the suspensions contained disrupted cell wallparticles with a high amount of lipopolysaccharide Howeverother bacterial components may also have been presentin the suspension The concentrations used in this studywere determined by dose-response experiments However asignificant upregulation of NAMPT was only observed forP gingivalis P gingivalis is a gram-negative bacterium andstrongly associated with periodontitis For its detrimental
10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

10 Mediators of Inflammation
effects P gingivalis possesses a number of virulence factorssuch as gingipains and fimbriae P gingivalis can invade hostcells and also evade the host defense system [65ndash67] Asit has been shown in several cells that COX2 an enzymewhich is responsible for the formation of prostanoids isupregulated by P gingivalis and IL-1120573 we also analyzed theCOX2 expression as a positive control in our study [61 62 6869] As expected P gingivalis and IL-1120573 increased the COX2expression in a dose- and time-dependent manner in PDLcells
In summary the present study demonstrates for the firsttime that NAMPT stimulates the production of MMP-1 andCCL2 in human PDL cells which suggests that NAMPTmay contribute to periodontal inflammation and matrixdestruction through the production of these moleculesTherefore increased NAMPT levels as found in obesity mayrepresent at least one mechanism whereby obesity couldconfer an increased risk of periodontitis in obese individ-uals In addition our study provides original evidence thatNAMPT is induced by the periodontopathogen P gingivalisand the proinflammatory cytokine IL-1120573 in PDL cells whichshows that microbial and inflammatory signals may use thisadipokine for their detrimental effects on the periodontiumThese findings suggest that local production of NAMPT inthe inflamed PDL could contribute to the increased gingivaland serum levels of NAMPT as observed in patients withperiodontitis and therefore represent a pathomechanisticlink whereby periodontitis affects systemic diseases such asdiabetes mellitus and cardiovascular diseases
Conflict of Interests
The authors declare that they have no conflict of interests
Acknowledgments
The authors would like to thank Ms Ramona HomigProfessor Werner Gotz and Professor Stephan Baader fortheir supportThis studywas funded by theGermanResearchFoundation (KFO208TP4) and the University of BonnProfessorHolger Frohlich is amember of the ExzellenzclusterImmunoSensation
References
[1] B L Pihlstrom B S Michalowicz and N W Johnson ldquoPeri-odontal diseasesrdquoThe Lancet vol 366 no 9499 pp 1809ndash18202005
[2] L Sbordone and C Bortolaia ldquoOral microbial biofilms andplaque-related diseases microbial communities and their rolein the shift from oral health to diseaserdquo Clinical Oral Investiga-tions vol 7 no 4 pp 181ndash188 2003
[3] J M Albandar ldquoCommentary underestimation of periodonti-tis in nhanes surveysrdquo Journal of Periodontology vol 82 no 3pp 337ndash341 2011
[4] T Beikler and T F Flemmig ldquoOral biofilm-associated diseasestrends and implications for quality of life systemic health andexpendituresrdquo Periodontology 2000 vol 55 no 1 pp 87ndash1032011
[5] J Suvan F DrsquoAiuto D R Moles A Petrie and N DonosldquoAssociation between overweightobesity and periodontitis inadults A systematic reviewrdquo Obesity Reviews vol 12 no 501pp e381ndashe404 2011
[6] G J Seymour P J Ford M P Cullinan S Leishman and KYamazaki ldquoRelationship between periodontal infections andsystemic diseaserdquoClinicalMicrobiology and Infection vol 13 no4 pp 3ndash10 2007
[7] J Conde M Scotece R Gomez et al ldquoAdipokines biofactorsfrom white adipose tissue A complex hub among inflamma-tion metabolism and immunityrdquo BioFactors vol 37 no 6 pp413ndash420 2011
[8] A R Moschen R R Gerner and H Tilg ldquoPre-b cell colonyenhancing factornamptvisfatin in inflammation and obesi-tyrelated disordersrdquo Current Pharmaceutical Design vol 16 no17 pp 1913ndash1920 2010
[9] Y-H Chang D-M Chang K-C Lin S-J Shin and Y-J LeeldquoVisfatin in overweightobesity type 2 diabetesmellitus insulinresistance metabolic syndrome and cardiovascular diseasesa meta-analysis and systemic reviewrdquo DiabetesMetabolismResearch and Reviews vol 27 no 6 pp 515ndash527 2011
[10] D Taskesen B Kirel and T Us ldquoSerum visfatin levels adiposityand glucose metabolism in obese adolescentsrdquo Journal ofClinical Research in Pediatric Endocrinology vol 4 pp 76ndash812012
[11] L Q Zhang D P Heruth and S Q Ye ldquoNicotinamide phos-phoribosyltransferase in human diseasesrdquo Journal of Bioanalysisand Biomedicine vol 3 no 1 pp 13ndash25 2011
[12] A R Pradeep R N M Raghavendra NM M V R PrasadR Kathariya S P Patel and A Sharma ldquoGingival crevicularfluid and serum visfatin concentration their relationship inperiodontal health and diseaserdquo Journal of Periodontology vol82 no 9 pp 1314ndash1319 2011
[13] A R Pradeep N M Raghavendra A Sharma et al ldquoAsso-ciation of serum and crevicular visfatin levels in periodontalhealth and disease with type 2 diabetes mellitusrdquo Journal ofPeriodontology vol 83 no 5 pp 629ndash634 2012
[14] M Nokhbehsaim B Deschner J Winter et al ldquoInteractions ofregenerative inflammatory and biomechanical signals on bonemorphogenetic protein-2 in periodontal ligament cellsrdquo Journalof Periodontal Research vol 46 no 3 pp 374ndash381 2011
[15] M Nokhbehsaim J Winter B Rath A Jager S Jepsen and JDeschner ldquoEffects of enamel matrix derivative on periodontalwound healing in an inflammatory environment in vitrordquoJournal of Clinical Periodontology vol 38 no 5 pp 479ndash4902011
[16] M Nokhbehsaim B Deschner J Winter et al ldquoAnti-inflammatory effects of EMD in the presence of biomechanicalloading and interleukin-1120573 in vitrordquoClinical Oral Investigationsvol 16 no 1 pp 275ndash283 2012
[17] S M Lin P Du W Huber and W A Kibbe ldquoModel-basedvariance-stabilizing transformation for Illumina microarraydatardquo Nucleic Acids Research vol 36 no 2 article e11 2008
[18] B M Bolstad R A Irizarry M Astrand and T P Speed ldquoAcomparison of normalizationmethods for high density oligonu-cleotide array data based on variance and biasrdquo Bioinformaticsvol 19 no 2 pp 185ndash193 2003
[19] P Du W A Kibbe and S M Lin ldquolumi a pipeline forprocessing Illumina microarrayrdquo Bioinformatics vol 24 no 13pp 1547ndash1548 2008
[20] J T Leek and J D Storey ldquoA general framework for multipletesting dependencerdquo Proceedings of the National Academy of
Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

Mediators of Inflammation 11
Sciences of the United States of America vol 105 no 48 pp18718ndash18723 2008
[21] G K Smyth ldquoLinear models and empirical bayes methods forassessing differential expression in microarray experimentsrdquoStatistical Applications in Genetics and Molecular Biology vol3 no 1 article 3 2004
[22] M E Ritchie D Diyagama J Neilson et al ldquoEmpiricalarray quality weights in the analysis of microarray datardquo BMCBioinformatics vol 7 article 261 2006
[23] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society B vol 57 pp 89ndash300 1995
[24] G K Smyth ldquoLimma linear models for microarray datardquo inBioinformatics andComputational Biology SolutionsUsing R andBioconductor R Gentleman V Carey W Huber R Irizarryand S Dudoit Eds pp 397ndash420 Springer New York NY USA2005
[25] P A Arakaki M R Marques and M C L G Santos ldquoMMP-1 polymorphism and its relationship to pathological processesrdquoJournal of Biosciences vol 34 no 2 pp 313ndash320 2009
[26] A-L Ejeil S Igondjo-Tchen S Ghomrasseni B Pellat GGodeau and B Gogly ldquoExpression of matrix metallopro-teinases (MMPs) and tissue inhibitors of metalloproteinases(TIMPs) in healthy and diseased human gingivardquo Journal ofPeriodontology vol 74 no 2 pp 188ndash195 2003
[27] G Tuter B Kurtis and M Serdar ldquoEffects of phase I peri-odontal treatment on gingival crevicular fluid levels of matrixmetalloproteinase-1 and tissue inhibitor of metalloproteinase-1rdquo Journal of Periodontology vol 73 no 5 pp 487ndash493 2002
[28] S L Deshmane S Kremlev S Amini and B E SawayaldquoMonocyte chemoattractant protein-1 (MCP-1) an overviewrdquoJournal of Interferon and Cytokine Research vol 29 no 6 pp313ndash326 2009
[29] G P Garlet W Martins Jr B R Ferreira C M Milanezi andJ S Silva ldquoPatterns of chemokines and chemokine receptorsexpression in different forms of human periodontal diseaserdquoJournal of Periodontal Research vol 38 no 2 pp 210ndash217 2003
[30] M Gupta R Chaturvedi and A Jain ldquoRole of monocytechemoattractant protein-1 (MCP-1) as an immune-diagnosticbiomarker in the pathogenesis of chronic periodontal diseaserdquoCytokine vol 61 pp 892ndash897 2013
[31] B Kurtis G Tuter M Serdar et al ldquoGingival crevicularfluid levels of monocyte chemoattractant protein-1 and tumornecrosis factor-alpha in patients with chronic and aggressiveperiodontitisrdquo Journal of Periodontology vol 76 no 11 pp1849ndash1855 2005
[32] F Brentano O Schorr C Ospelt et al ldquoPre-B cell colony-enhancing factorvisfatin a new marker of inflammationin rheumatoid arthritis with proinflammatory and matrix-degrading activitiesrdquo Arthritis and Rheumatism vol 56 no 9pp 2829ndash2839 2007
[33] S W Liu S B Qiao J S Yuan and D Q Liu ldquoVisfatinstimulates production of monocyte chemotactic protein-1 andinterleukin-6 in human vein umbilical endothelial cellsrdquo Hor-mone and Metabolic Research vol 41 no 4 pp 281ndash286 2009
[34] G Sommer S Kralisch N Kloting et al ldquoVisfatin is a positiveregulator of MCP-1 in human adipocytes in vitro and inmice invivordquo Obesity vol 18 no 8 pp 1486ndash1492 2010
[35] E D Adamson and D Mercola ldquoEgr1 transcription factormultiple roles in prostate tumor cell growth and survivalrdquoTumor Biology vol 23 no 2 pp 93ndash102 2002
[36] V Baron E D Adamson A Calogero G Ragona and DMercola ldquoThe transcription factor Egr1 is a direct regulator ofmultiple tumor suppressors including TGF1205731 PTEN p53 andfibronectinrdquo Cancer Gene Therapy vol 13 no 2 pp 115ndash1242006
[37] V S Ten and D J Pinsky ldquoEndothelial response to hypoxiaphysiologic adaptation and pathologic dysfunctionrdquo CurrentOpinion in Critical Care vol 8 no 3 pp 242ndash250 2002
[38] S B McMahon and J G Monroe ldquoThe role of early growthresponse gene 1 (egr-1) in regulation of the immune responserdquoJournal of Leukocyte Biology vol 60 no 2 pp 159ndash166 1996
[39] S L Teitelbaum ldquoThe osteoclast and its unique cytoskeletonrdquoAnnals of the New York Academy of Sciences vol 1240 no 1 pp14ndash17 2011
[40] R A Colvin T K Means T J Diefenbach et alldquoSynaptotagmin-mediated vesicle fusion regulates cellmigrationrdquo Nature Immunology vol 11 no 6 pp 495ndash5022010
[41] K T Fowler NW Andrews and JW Huleatt ldquoExpression andfunction of synaptotagmin VII in CTLsrdquo Journal of Immunol-ogy vol 178 no 3 pp 1498ndash1504 2007
[42] G Glavan R Schliebs and M Zivin ldquoSynaptotagmins inneurodegenerationrdquo Anatomical Record vol 292 no 12 pp1849ndash1862 2009
[43] H Kato K Uzawa T Onda et al ldquoDown-regulation of 1D-myo-inositol 145-trisphosphate 3-kinase A protein expressionin oral squamous cell carcinomardquo International Journal ofOncology vol 28 no 4 pp 873ndash881 2006
[44] S Windhorst R Fliegert C Blechner et al ldquoInositol 145-trisphosphate 3-kinase-A is a new cell motility-promotingprotein that increases the metastatic potential of tumor cells bytwo functional activitiesrdquo Journal of Biological Chemistry vol285 no 8 pp 5541ndash5554 2010
[45] HK Il K P Soon TH Soon et al ldquoInositol 145-trisphosphate3-kinase A functions as a scaffold for synaptic rac signalingrdquoJournal of Neuroscience vol 29 no 44 pp 14039ndash14049 2009
[46] D Krizsan-Agbas T Pedchenko and P G Smith ldquoNeurotriminis an estrogen-regulated determinant of peripheral sympatheticinnervationrdquo Journal of Neuroscience Research vol 86 no 14pp 3086ndash3095 2008
[47] S Chen O Gil Y Q Ren G Zanazzi J L Salzer and D EHillman ldquoNeurotrimin expression during cerebellar develop-ment suggests roles in axon fasciculation and synaptogenesisrdquoJournal of Neurocytology vol 30 no 11 pp 927ndash937 2002
[48] J L Bell KWachter BMuhleck et al ldquoInsulin-like growth fac-tor 2 mRNA-binding proteins (IGF2BPs) post-transcriptionaldrivers of cancer progressionrdquo Cellular and Molecular LifeSciences vol 70 no 15 pp 2657ndash2675 2013
[49] R Suvasini B Shruti B Thota et al ldquoInsulin growth factor-2binding protein 3 (IGF2BP3) is a glioblastoma-specific markerthat activates phosphatidylinositol 3-kinasemitogen-activatedprotein kinase (PI3KMAPK) pathways by modulating IGF-2rdquoJournal of Biological Chemistry vol 286 no 29 pp 25882ndash25890 2011
[50] C A Staton I Kumar M W R Reed and N J BrownldquoNeuropilins in physiological and pathological angiogenesisrdquoJournal of Pathology vol 212 no 3 pp 237ndash248 2007
[51] J R L Wild C A Staton K Chapple and B M Corfe ldquoNeu-ropilins expression and roles in the epitheliumrdquo InternationalJournal of Experimental Pathology vol 93 no 2 pp 81ndash1032012
12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom

12 Mediators of Inflammation
[52] I C Zachary ldquoHow neuropilin-1 regulates receptor tyrosinekinase signalling the knowns and known unknownsrdquo Biochem-ical Society Transactions vol 39 no 6 pp 1583ndash1591 2011
[53] M Mizui and H Kikutani ldquoNeuropilin-1 the glue betweenregulatory T cells and dendritic cellsrdquo Immunity vol 28 no3 pp 302ndash303 2008
[54] J Schwenk M Metz G Zolles et al ldquoNative GABAB receptorsare heteromultimerswith a family of auxiliary subunitsrdquoNaturevol 465 no 7295 pp 231ndash235 2010
[55] R Adya B K Tan J Chen and H S Randeva ldquoNuclear factor-120581b induction by visfatin in human vascular endothelial cellsrdquoDiabetes Care vol 31 no 4 pp 758ndash760 2008
[56] Y Fan S Meng Y Wang J Cao and C Wang ldquoVis-fatinPBEFNampt induces EMMPRIN and MMP-9 produc-tion in macrophages via the NAMPT-MAPK (p38 ERK12)-NF-120581B signaling pathwayrdquo International Journal of MolecularMedicine vol 27 no 4 pp 607ndash615 2011
[57] A V Nogueira M Nokhbehsaim S Eick et al ldquoRegulation ofvisfatin by microbial and biomechanical signals in PDLcellsrdquoClinical Oral Investigations In press
[58] M Gosset F Berenbaum C Salvat et al ldquoCrucial role ofvisfatinpre-B cell colony-enhancing factor in matrix degrada-tion and prostaglandin E2 synthesis in chondrocytes possibleinfluence on osteoarthritisrdquo Arthritis and Rheumatism vol 58no 5 pp 1399ndash1409 2008
[59] C E Kendal and G D Bryant-Greenwood ldquoPre-B-cell colony-enhancing factor (PBEFVisfatin) gene expression ismodulatedby NF-120581B and AP-1 in human amniotic epithelial cellsrdquo Pla-centa vol 28 no 4 pp 305ndash314 2007
[60] M R Williams N Kataoka Y Sakurai C M Powers S GEskin and L V McIntire ldquoGene expression of endothelialcells due to interleukin-1 beta stimulation and neutrophiltransmigrationrdquo Endothelium vol 15 no 1-2 pp 73ndash84 2008
[61] Y Murakami A Kawata Y Seki et al ldquoComparative inhibitoryeffects of magnolol honokiol eugenol and bis-eugenol oncyclooxygenase-2 expression and nuclear factor-kappa B acti-vation in RAW264 7 macrophage-like cells stimulated withfimbriae of Porphyromonas gingivalisrdquo In Vivo vol 26 pp 941ndash950 2012
[62] D Reddi S J Brown and G N Belibasakis ldquoPorphyromonasgingivalis induces RANKL in bone marrow stromal cellsinvolvement of the p38 MAPKrdquoMicrobial Pathogenesis vol 51no 6 pp 415ndash420 2011
[63] A Dunne and L A J OrsquoNeill ldquoThe interleukin-1 receptorToll-like receptor superfamily signal transduction during inflam-mation and host defenserdquo Sciencerdquos STKE vol 2003 no 171article re3 2003
[64] J-K Lee S-H Kim E C Lewis T Azam L L Reznikov andC A Dinarello ldquoDifferences in signaling pathways by IL-1120573 andIL-18rdquo Proceedings of the National Academy of Sciences of theUnited States of America vol 101 no 23 pp 8815ndash8820 2004
[65] S C Holt and J L Ebersole ldquoPorphyromonas gingivalis Tre-ponema denticola and Tannerella forsythia the ldquored complexrdquoa prototype polybacterial pathogenic consortium in periodon-titisrdquo Periodontology 2000 vol 38 pp 72ndash122 2005
[66] K Nakayama ldquoMolecular genetics of Porphyromonas gingi-valis gingipains and other virulence factorsrdquo Current Proteinand Peptide Science vol 4 no 6 pp 389ndash395 2003
[67] J Potempa A Sroka T Imamura and J Travis ldquoGingipainsthe major cysteine proteinases and virulence factors of Por-phyromonas gingivalis structure function and assembly of
multidomain protein complexesrdquo Current Protein and PeptideScience vol 4 no 6 pp 397ndash407 2003
[68] Y-C Chang F-M Huang S-F Yang et al ldquoInduction ofcyclooxygenase-2 mRNA and protein expression in humanpulp cells stimulated with black-pigmented bacteroidesrdquo Jour-nal of Endodontics vol 29 no 4 pp 240ndash243 2003
[69] Y Hayashi M Kobayashi H Kuwata et al ldquoInterferon-120574 andinterleukin 4 inhibit interleukin 1120573-induced delayed prostagi-landin E2 generation through suppression of cyclooxugenase-2 expression in human fibroblastsrdquo Cytokine vol 12 no 6 pp603ndash612 2000
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom
Submit your manuscripts athttpwwwhindawicom
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
MEDIATORSINFLAMMATION
of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Behavioural Neurology
EndocrinologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Disease Markers
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
OncologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Oxidative Medicine and Cellular Longevity
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
PPAR Research
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Immunology ResearchHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Journal of
ObesityJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Computational and Mathematical Methods in Medicine
OphthalmologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Diabetes ResearchJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Research and TreatmentAIDS
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Gastroenterology Research and Practice
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Parkinsonrsquos Disease
Evidence-Based Complementary and Alternative Medicine
Volume 2014Hindawi Publishing Corporationhttpwwwhindawicom