research article histological analysis of … · the female reproductive tract in late spring and...
TRANSCRIPT

ARTICLE IN PRESS
Ann Anat 190 (2008) 461—476
0940-9602/$ - sdoi:10.1016/j.
�CorrespondE-mail addr
www.elsevier.de/aanat
RESEARCH ARTICLE
Histological analysis of spermatogenesis and thegerm cell development strategy within the testis ofthe male Western Cottonmouth Snake, Agkistrodonpiscivorus leucostoma
Kevin M. Gribbinsa,�, Justin L. Rheuberta, Matthew H. Colliera,Dustin S. Siegelb, David M. Severc
aDepartment of Biology, Wittenberg University, PO Box 720, Springfield, OH 45501-0720, USAbDepartment of Biology, Saint Louis University, St. Louis, MO 63103, USAcDepartment of Biological Sciences, Southeastern Louisiana University, Hammond, LA 70402, USA
Received 7 December 2007; received in revised form 25 June 2008; accepted 1 July 2008
KEYWORDSCottonmouth;Agkistrodon;Spermatogenesis;Testes;Reproduction;Bimodal
ee front matter & 2008aanat.2008.07.003
ing author.ess: kgribbins@wittenb
SummaryCottonmouth (Agkistrodon piscivorus leucostoma) testes were examined histologi-cally to determine the germ cell development strategy employed duringspermatogenesis. Testicular tissues from Cottonmouths were collected monthlyfrom swamps around Hammond, Louisiana. Pieces of testis were fixed in Trump’sfixative, dehydrated in ethanol, embedded in Spurr’s plastic, sectioned with anultramicrotome, and stained with toluidine blue and basic fuchsin. Spermatogenesiswithin Cottonmouths occurs in two independent events within a single calendar year.The testes are active during the months of March–June and August–October withspermiation most heavily observed during April–May and October. To our knowledge,this is the first study that describes bimodal spermatogenesis occurring in the sameyear within the subfamily Crotalinae. During spermatogenesis, no consistent spatialrelationships are observed between germ cell generations. Typically, either certaincell types were missing (spermatocytes) or the layering of 3–5 spermatids and/orspermatocytes within the same cross-section of seminiferous tubule preventedconsistent spatial stages from occurring. This temporal pattern of sperm develop-ment is different from the spatial development found within birds and mammals,being more reminiscent of that seen in amphibians, and has now been documentedwithin every major clade of reptile (Chelonia, Serpentes, Sauria, Crocodylia). Thisprimitive-like sperm development, within a testis structurally similar to mammals
Elsevier GmbH. All rights reserved.
erg.edu (K.M. Gribbins).

ARTICLE IN PRESS
K.M. Gribbins et al.462
and birds, may represent an intermediate testicular model within the basallypositioned (phylogenetically) reptiles that may be evolutionarily significant.& 2008 Elsevier GmbH. All rights reserved.
Introduction
Two different germ cell development strategiesexist within vertebrates. Anamniotic vertebratetestes are composed of tubules or lobules that arelined with cysts. Germ cells develop as a singlecohort through mitosis, meiosis, spermiogenesis,and upon completion of spermatogenesis maturespermatozoa are released from these cysts into thelumina of the tubules or lobules in a singlespermiation event (Lofts, 1964; Van Oordt andBrands, 1970). Amniotic testes contain seminifer-ous tubules that are lined with seminiferousepithelia (Leblond and Clermont, 1952). Sperma-togenesis occurs in association with Sertoli cells,which make up the seminiferous epithelium ofamniotic testes.
Throughout the spermatogenic cycle in mostamniotes studied to date (limited mostly totemperate and continually breeding birds andmammals), germ cells within the seminiferousepithelium are organized in consistent spatialarrangements as generations of germ cells movefrom the periphery of the epithelium to the lumenof the seminiferous tubules. This type of synchro-nized germ cell development strategy results inconsistent cellular associations that can be seenand predicted. These stages are typically numeri-cally arranged within large segments of seminifer-ous tubules, which lead to waves of sperm releasethroughout the reproductively active months inmammals (Russell et al., 1990).
Current research provides evidence that mosttemperate pitvipers mate, have a period oftesticular quiescence, and then begin spermato-genesis during the late spring/early summer(Srivastava and Thapliyal, 1965; Saint Girons,1982; Aldridge, 2002). Although spermatogenesisgenerally starts during the late spring/early sum-mer, many variations and testicular cycles exist.These cycles differ from species to species and areassociated with internal and external androgeniccues (Shea, 2001; Bertona and Chiaraviglio, 2003).Recent research has also shown that temperatesnakes, like the Black Swamp Snake (Seminatrixpygaea), may rely on a completely different germcell development strategy than that observed inbirds and mammals. Black Swamp Snakes (Gribbinset al., 2005) and other temperate reptiles (Gribbinsand Gist, 2003; Gribbins et al., 2003, 2006) studied
to date have testes with typical amniotic structures(seminiferous tubules, seminiferous epithelium,Sertoli cells). However, their germ cell develop-ment strategy is more reminiscent of that found inanamniotes. This germ cell development may beconserved and considered primitive compared withthe derived spatial spermatogenesis seen in birdsand mammals. The basal position of reptiles withinthe amniotic clade and the reoccurring theme of atemporal germ cell development strategy in everytemperate reptile studied to date suggests that thispleisomorphic-like spermatogenic cycle within astructurally amniotic testis may be evolutionarysignificant.
The present study on Agkistrodon piscivorusleucostoma is a continuation of this long-terminvestigation on germ cell development in reptiles.The Cottonmouth is a viviparous snake that can befound within lowlands of the southeastern UnitedStates (Conant and Collins, 1991). Cottonmouthsare semi-aquatic snakes that in many cases occupypermanent water sources in abundant numberswithin their geographic range. Pairings of males andfemales have been seen in Cottonmouths byWharton (1966) throughout the entire year exceptJanuary with female ovulation occurring in thespring (Zaidan et al., 2003). Spermatogenesisbegins in April and spermiogenesis is completedby late October (Johnson et al., 1982) for popula-tions of Cottonmouths found in Alabama. Theclimax of spermatogenesis, along with increasedtestosterone levels, occurs in mid July in allCottonmouth populations studied to date (Johnsonet al., 1982; Zaidan et al., 2003; Graham, 2006).
The purpose of this study is to provide a detailedhistological evaluation of the major events asso-ciated with spermatogenesis so that the germ celldevelopment strategy of a southeastern LouisianaCottonmouth population (Johnson et al. (1982) didnot provide information about the germ celldevelopment strategy within Alabama male Cotton-mouths) can be compared with that of the SwampSnake and other temperate reptiles. Cottonmouthsare unique to the other temperate species studiedto date in that there is evidence that spermatogen-esis is seasonal, but breeding may occur any timeduring the year (assuming intersexual pairings areindicative of mating activity). Furthermore, withinthe population of Cottonmouths from this studymature sperm can be found in the distal regions of

ARTICLE IN PRESS
Spermatogenesis in Agkistrodon piscivorus leucostoma 463
the female reproductive tract in late spring and fallsuggesting that there may be two major breedingseasons in Louisiana (Siegel and Sever, 2008). Thisinformation leads to the hypothesis that maleCottonmouths in Louisiana either store sperm thatis produced from one seasonal peak of spermato-genesis as observed by Johnson et al. (1982) orthere may be more than one spermatogenic event(i.e. early summer and late fall) that would providesperm for the majority of matings, which areassumed to occur most often during the warmermonths of the year. This type of bimodal sperma-togenesis has not been documented within aspecies of snake belonging to the subfamilyCrotalinae. However, a similar biannual germ celldevelopment in males has been previously de-scribed during the summer months for specieswithin Colubridae (Goldberg, 1995, 1998). If abimodal type of spermatogenesis occurs in thisLouisiana population of Cottonmouths, then thisnew annual pattern of reproduction would be quitedifferent from the postnuptial pattern of sperma-togenesis typical of temperate-zoned snakes thatwas previously described by Johnson et al. (1982)for Alabama Cottonmouths. A biannual pattern ofspermatogenesis within Louisiana Cottonmouthswould provide the opportunity to retest thehypothesis that reproductive strategy, which hasalready been examined in postnuptial (Seminatrixpygaea, Gribbins et al., 2005), prenuptial (Alligatormississippiensis, Gribbins et al., 2006), and mixed(Podarcis muralis, Gribbins and Gist, 2003) repro-ductive cycles, may predetermine the type of germcell development strategy utilized by seasonallybreeding reptiles.
Materials and methods
Animal collection
Twenty-two adult male Cottonmouth snakes,A. piscivorus leucostoma, were collected duringthe months of January 2005 through November2006 (2 snakes were captured per month except: 1specimen per month for September, November, andJanuary and 5 specimens for June) from threelocalities; the Amite River Diversion Canal (North30122.616/West 090168.506, Livingston Parish, LA),Turtle Cove Environmental Research Station on PassManchac (North 30129.426/West 090135.592, Tan-gipahoa Parish, LA), and the private residence ofDr. Clifford Fontenot, 10 km Northwest of NewAlbany (North 30130.871/West 090136.202, Living-ston Parish, LA). The snakes were sacrificed with a
0.2–0.5ml intraperitoneal injection of sodiumpentobarbital (1 g sodium pentobarbital in 10%ethanol/40% propylene glycol solution) and thetestes were removed and fixed in Trump’s fixative(EMS, Hatfield, PA). The testes were then cutinto transverse sections and stored in 70% ethanolat 4 1C.
Tissue preparation for light microscopy
Sections of each testis were cut into approxi-mately 3mm cubes, dehydrated in a graded seriesof ethanol (70%, 85%, 2� 95%, 2� 100%), incubatedin 1:2 Spurr’s plastic (EMS, Hatfield, PA): 100%ethanol for 30min, and then once in 1:1 Spurr’splastic: 100% ethanol for 30min. The tissues werethen infiltrated in pure Spurr’s plastic overnight.New plastic was prepared and the testes wereembedded and cured at 60 1C for 48 h in a Fisherisotemperature vacuum oven (Fisher Scientific,Pittsburg, PA). Sections (2–3 mm) were cut fromthe plastic blocks using a dry glass knife and an LKB-Ultramicrotome III (LKB Produkter AB, Bromma,Sweden). The tissues were visualized using a basicfuchsin and toluidine blue composite stain asdescribed by Hayat (1993).
Histological analysis
The testicular tissues were examined using anOlympus compound microscope (Olympus America,Center Valley, PA) to determine the cytologicalchanges that occurred during spermatogenesis.Sagittal and cross-sectional areas of the seminifer-ous tubules were selected at random and germ cellmorphologies and the presence or absence ofspatial stages were determined. Photographs weretaken with a SPOT digital camera (DiagnosticSystems Laboratories, Webster, TX) and wereviewed and edited using Adobe Photoshop 7.0(Adobe Systems, San Jose, CA).
Data analysis, measurements,and morphometrics
Upon capture, each snake’s snout-to-vent length(SVL) was determined and ranged from 36.0 to73.1 cm. The width and length (in mm) weremeasured for the right and left testes shortly afterdissection and testis volumes were estimated usingthe formula for a prolated spheroid: V ¼ 4/3p(1/2 L)(1/2W)2 (Selby, 1965; Ramirez-Bautistaand Gutierrez-Mayen, 2003). Right and left testi-cular volumes were pooled in order to increasedata collection for each month of the year. Thirty

ARTICLE IN PRESS
K.M. Gribbins et al.464
cross-sections of seminiferous tubules were chosenfor each represented month and the tubulardiameter and germinal epithelial heights weremeasured using an ocular micrometer.
Data analysis was performed using Minitab 15(Minitab Inc., State College, PA) for Windows.Results were deemed significant if Pp0.005.Testicular volume, tubular diameter, and germinalepithelial height data were tested for normalityand homogeneity of variances using the Kolmogor-ov–Smirnov and Bartlett’s tests, respectively,before statistical analyses were performed (Sokaland Rohlf, 1995; Flemming, 1993). These data didnot meet assumptions of normality; thus theKruskal–Wallis analysis of variance was used to testfor significant seasonal variation in testicularvolume, seminiferous tubular diameter, and germi-nal epithelial height.
Results
Germ cell morphology and the cell cycle
The A. piscivorus leucostoma testis containsseminiferous tubules lined with seminiferousepithelia consisting of Sertoli cells and developinggerm cells. Germ cells develop spermatogenicallyin close association with the Sertoli cells of thisepithelium. Histological examination shows thattestes of this southeastern Louisiana population ofCottonmouths are spermatogenically active duringtwo periods within the same year, the months ofMarch–June and again in August–October. Sperma-togenesis commences in March and after a briefquiescence in July starts again in August with theproliferation of spermatogonia A and B near thebasal lamina (basement membrane) of the semi-niferous epithelia. The majority of spermatogoniaundergo meiosis during the months of April andAugust/October and continue through spermiogen-esis in May–June and then again in October.Spermatogonia are present and undergo mitosis inall months of the year; however, divisions areslowed during the months of June/July, November,and January/February. Although there is a decreasein mitotic activity, there are no periods during theyear in which the testes of these Cottonmouths areinactive in terms of proliferation.
Testicular volume has been used in reproductivestudies performed on reptiles as an indicator ofspermatogenic activity (Srivastava and Thapliyal,1965; Flemming, 1993; Ramirez-Bautista andGutierrez-Mayen, 2003). However, previous studies(see Brown and Bomberger Brown, 2003) have
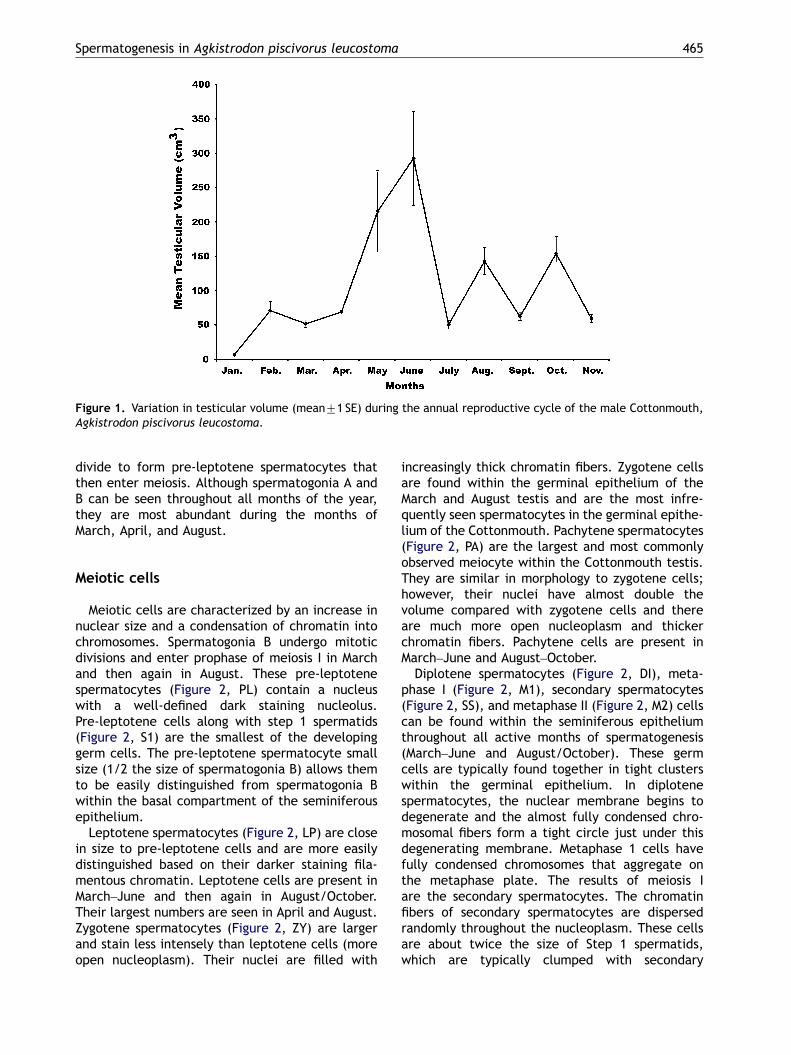
shown that SVL can bias testicular volume data;therefore, linear regression was first used todetermine whether mean testicular volumes weresignificantly affected by variations in male meanSVL. Linear regression analysis indicated thattesticular volumes were not significantly correlatedwith variations in mean male SVL (right testicularvolumes: R2 ¼ 0.217, Pp0.05; left testicularvolumes: R2 ¼ 0.261, Pp0.05). Conversely, meantesticular volumes in these Cottonmouths doindeed show a significant annual trend (Kruskal–Wallis; H ¼ 27.7, df ¼ 10, P ¼ 0.002, Figure 1).Testis volumes were at a minimum during themonths of quiescence (November, 59.00 cm3; Jan-uary, 16.6 cm3; February, 70.6 cm3; July, 50.15 cm3)and during the month of September (61.77 cm3).The specimen caught during September shouldhave a high testis volume and also show spermio-genic activity similar to the August and Septembertestis. However, there are only spermatogoniapresent in the seminiferous epithelium and thetestis volume is low. It also should be noted thatthis Cottonmouth was caught 2 weeks afterhurricane Katrina and outside of its normal habitat.
Testicular volumes (Figure 1) increase steadily asspermatogenic activity increases (April, 68.42 cm3;May, 215.30 cm3) and volumes peak during theclimax of spermiogenesis and spermiation in June(292.60 cm3). Testicular volume then drops signifi-cantly during July (50.15 cm3) when the testis isquiescent and then increases again as spermato-genesis and spermiogenesis increases during August(142.60 cm3) and October (153.60 cm3). Testisvolume is much larger during the spring spermato-genic climax (292.60 cm3) when compared with thefall peak of sperm development (153.60 cm3).
Pre-meiotic cells
The seminiferous epithelium contains twomorphologies of pre-meiotic cells (SpermatogoniaA and B) (Figure 2; SpA and SpB) during all monthsof the year. These cells are characterized by nucleiwith random clumps of heterochromatin. Themajor morphological differences between the twotypes of spermatogonia are that the A type is ovoidin shape with one large nucleolus and B type ismore round in shape and usually lacks a prominentnucleolus. Both spermatogonial types are generallyfound near the basement membrane of the epithe-lium away from the lumen and associated with thebasal compartments formed by Sertoli cells. Duringthe spermatogenic cycle both types of spermato-gonia undergo mitosis to maintain the spermatogo-nial population and many of the B spermatogonia
ARTICLE IN PRESS
Figure 1. Variation in testicular volume (mean71 SE) during the annual reproductive cycle of the male Cottonmouth,Agkistrodon piscivorus leucostoma.
Spermatogenesis in Agkistrodon piscivorus leucostoma 465
divide to form pre-leptotene spermatocytes thatthen enter meiosis. Although spermatogonia A andB can be seen throughout all months of the year,they are most abundant during the months ofMarch, April, and August.
Meiotic cells
Meiotic cells are characterized by an increase innuclear size and a condensation of chromatin intochromosomes. Spermatogonia B undergo mitoticdivisions and enter prophase of meiosis I in Marchand then again in August. These pre-leptotenespermatocytes (Figure 2, PL) contain a nucleuswith a well-defined dark staining nucleolus.Pre-leptotene cells along with step 1 spermatids(Figure 2, S1) are the smallest of the developinggerm cells. The pre-leptotene spermatocyte smallsize (1/2 the size of spermatogonia B) allows themto be easily distinguished from spermatogonia Bwithin the basal compartment of the seminiferousepithelium.
Leptotene spermatocytes (Figure 2, LP) are closein size to pre-leptotene cells and are more easilydistinguished based on their darker staining fila-mentous chromatin. Leptotene cells are present inMarch–June and then again in August/October.Their largest numbers are seen in April and August.Zygotene spermatocytes (Figure 2, ZY) are largerand stain less intensely than leptotene cells (moreopen nucleoplasm). Their nuclei are filled with
increasingly thick chromatin fibers. Zygotene cellsare found within the germinal epithelium of theMarch and August testis and are the most infre-quently seen spermatocytes in the germinal epithe-lium of the Cottonmouth. Pachytene spermatocytes(Figure 2, PA) are the largest and most commonlyobserved meiocyte within the Cottonmouth testis.They are similar in morphology to zygotene cells;however, their nuclei have almost double thevolume compared with zygotene cells and thereare much more open nucleoplasm and thickerchromatin fibers. Pachytene cells are present inMarch–June and August–October.
Diplotene spermatocytes (Figure 2, DI), meta-phase I (Figure 2, M1), secondary spermatocytes(Figure 2, SS), and metaphase II (Figure 2, M2) cellscan be found within the seminiferous epitheliumthroughout all active months of spermatogenesis(March–June and August/October). These germcells are typically found together in tight clusterswithin the germinal epithelium. In diplotenespermatocytes, the nuclear membrane begins todegenerate and the almost fully condensed chro-mosomal fibers form a tight circle just under thisdegenerating membrane. Metaphase 1 cells havefully condensed chromosomes that aggregate onthe metaphase plate. The results of meiosis Iare the secondary spermatocytes. The chromatinfibers of secondary spermatocytes are dispersedrandomly throughout the nucleoplasm. These cellsare about twice the size of Step 1 spermatids,which are typically clumped with secondary

ARTICLE IN PRESS
Figure 2. Germ cell types found within the seminiferous epithelium Agkistrodon piscivorus leucostoma. Bar ¼ 15 mm.SpA, type A spermatogonia; SpB, type B spermatogonia; PL, pre-leptotene spermatocyte; LP, leptotene spermatocyte;ZY, zygotene spermatocyte; PA, pachytene spermatocyte; DI, diplotene spermatocyte; M1, meiosis I; SS, secondaryspermatocyte; M2, meiosis II; S1, step 1 spermatid; S2, step 2 spermatid; S3, step 3 spermatid; S4, step 4 spermatid; S5,step 5 spermatid; S6, step 6 spermatid; S7, step 7 spermatid; MS, mature spermatozoa.
K.M. Gribbins et al.466
spermatocytes within the seminiferous epithelium.During metaphase 2, chromosomes aggregatearound the metaphase plate again. The onlydifferential factor between metaphase 1 and
metaphase 2 is the germ cell size and the amountof chromatin present. The metaphase 2 cells areslightly smaller and contain about half the amountof chromatin found in metaphase 1 cells.

ARTICLE IN PRESS
Figure 3. Top: variation in seminiferous tubule diameter (mean71 SE) and bottom: variation in germinal epithelialheight (mean71 SE) during the annual reproductive cycle of the male Cottonmouth, Agkistrodon piscivorusleucostoma.
Spermatogenesis in Agkistrodon piscivorus leucostoma 467
Spermiogenic cells
Spermiogenesis can be divided into seven steps inthe Cottonmouth germinal epithelium based on theterminology of Russell et al. (1990) for mammalianspecies. Steps of spermiogenesis are defined basedon acrosomal formation, nuclear elongation, andchromosomal condensation. The presence of step 1spermatids (Figure 2, S1) in April and August marksthe beginning of spermiogenesis. Step 1 spermatidsare small in size, have a well-defined nuclearmembrane, and no definable acrosomal vesicle.A well-defined acrosomal vesicle in contact withthe nuclear membrane characterizes step 2 sper-
matids (Figure 2, S2). These germ cells havechromatin that is interspersed throughout thenucleoplasm. Step 2 spermatids are present inlarge numbers in May and October.
Step 3 spermatids (Figure 2, S3) and Step 4spermatids (Figure 2, S4) are often present at thesame time as step 2 spermatids. As step 3 and 4spermatids continue to develop, the acrosomebegins to widen and envelope the nuclearhead, which flattens the apex of the nucleus. Acentrally located acrosome granule is oftenpresent during this stage of spermatid develop-ment. Step 5 spermatids (Figure 2, S5) markthe transition between round and elongating
ARTICLE IN PRESS
K.M. Gribbins et al.468
spermatids. Elongation begins at the opposite endof the nucleus away from the acrosome creating anucleus that is stretched in its dorsoventral plane.As elongating spermatids undergo development,they also begin to accumulate near the apicalsurfaces of the Sertoli cells with their tailsstretching out into the lumen and their nuclearheads facing the basement membrane.
The nuclei of step 6 spermatids (Figure 2, S6) arelonger than they are wide. The acrosomal vesiclecontinues to extend over the head of the nucleuswhen visible. Step 7 spermatids (Figure 2, S7)represent the climax of elongation. They haveundergone nuclear condensation and cytoplasmicelimination, resulting in nuclei more intenselystained and thinner in diameter than any of the
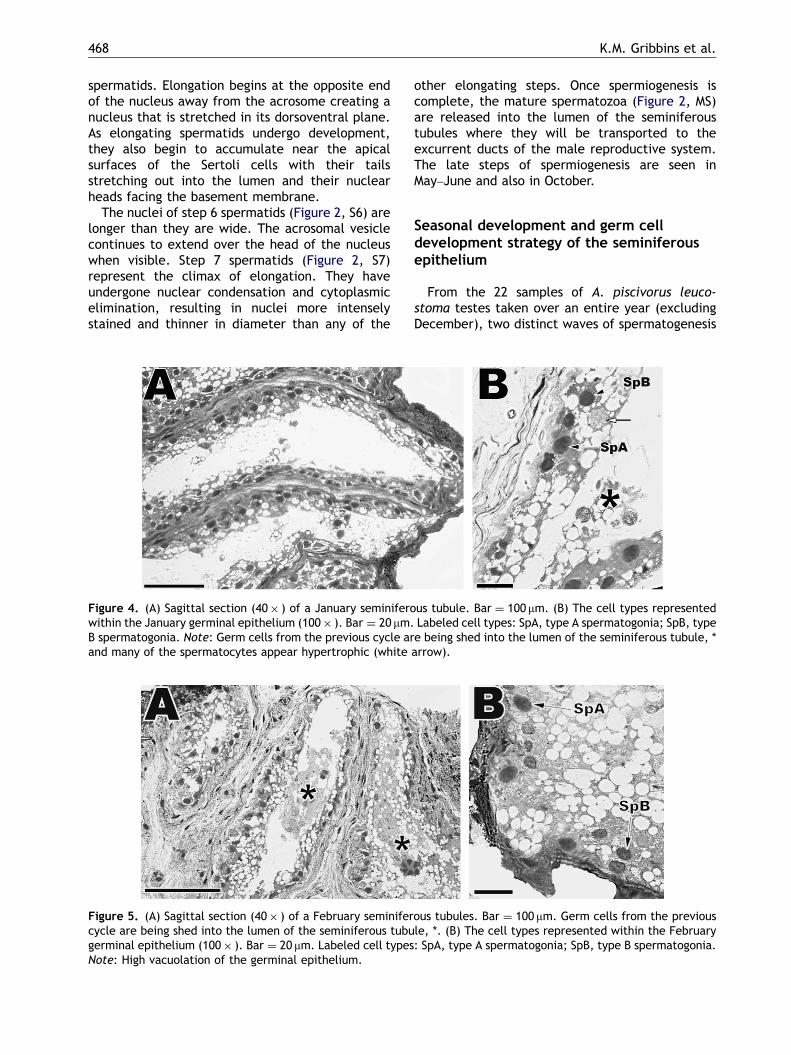
Figure 4. (A) Sagittal section (40� ) of a January seminiferowithin the January germinal epithelium (100� ). Bar ¼ 20 mm.B spermatogonia. Note: Germ cells from the previous cycle arand many of the spermatocytes appear hypertrophic (white
Figure 5. (A) Sagittal section (40� ) of a February seminifercycle are being shed into the lumen of the seminiferous tubugerminal epithelium (100� ). Bar ¼ 20 mm. Labeled cell typesNote: High vacuolation of the germinal epithelium.
other elongating steps. Once spermiogenesis iscomplete, the mature spermatozoa (Figure 2, MS)are released into the lumen of the seminiferoustubules where they will be transported to theexcurrent ducts of the male reproductive system.The late steps of spermiogenesis are seen inMay–June and also in October.
Seasonal development and germ celldevelopment strategy of the seminiferousepithelium
From the 22 samples of A. piscivorus leuco-stoma testes taken over an entire year (excludingDecember), two distinct waves of spermatogenesis
us tubule. Bar ¼ 100 mm. (B) The cell types representedLabeled cell types: SpA, type A spermatogonia; SpB, typee being shed into the lumen of the seminiferous tubule, *arrow).
ous tubules. Bar ¼ 100 mm. Germ cells from the previousle, *. (B) The cell types represented within the February: SpA, type A spermatogonia; SpB, type B spermatogonia.

ARTICLE IN PRESS
Spermatogenesis in Agkistrodon piscivorus leucostoma 469
are observed histologically within the seminiferousepithelium. This biannual type of spermatogenesisis supported by seminiferous tubule diameter andgerminal epithelial height data. Seasonal variationin seminiferous tubule diameter (Kruskal–Wallis;H ¼ 287.22, df ¼ 10, P ¼ 0.000, Figure 3) andgerminal epithelial height (Kruskal–Wallis;H ¼ 270.93, df ¼ 10, P ¼ 0.000, Figure 3) paralleleach other and show a significant monthly trend.
Prior to March the seminiferous tubules are in aquiescent phase of development (January andFebruary, Figures 4 and 5) and spermatogonia Aand B are the only major germ cell types presentwithin the seminiferous epithelium. Seminiferoustubule diameters (January, 101.25 mm; February,99.80 mm) and germinal epithelial heights (January,21.55 mm; February, 21.65 mm) are also close totheir smallest values during these two months. It isnot uncommon to observe large portions of the
Figure 6. (A) Sagittal section (40� ) of a March seminiferowithin the March germinal epithelium (100� ). Bar ¼ 20 mm. Lspermatogonia; LP, leptotene spermatocytes; ZY, zygotene spespermatocytes; S1, step 1 spermatid.
Figure 7. (A) Sagittal section (40� ) of an April seminiferowithin the April germinal epithelium (100� ). Bar ¼ 20 mm. Laspermatogonia; PL, pre-leptotene spermatocytes; LP, leptomeiosis 1; M2, meiosis 2; SS, secondary spermatocytes; S1, ste
seminiferous epithelium sloughed off into thelumen (Figures 4B and 5A, *) at this time of theyear.
The first wave of spermatogenesis begins inMarch and April (Figures 6 and 7) with an increasein spermatogonial proliferation and the earlyevents of meiosis I dominating the seminiferousepithelia. The increase in mitosis and meiosis leadsto larger seminiferous tubule diameters (March,126.05 mm; April, 130.75 mm) and germinal epithe-lial heights (March, 28.20 mm; April, 29.15 mm). Noconsistent cellular associations are observed bet-ween early and late developing generations of germcells because the later events of spermiogenesisare missing from the seminiferous tubules ofJanuary–April Cottonmouth testes.
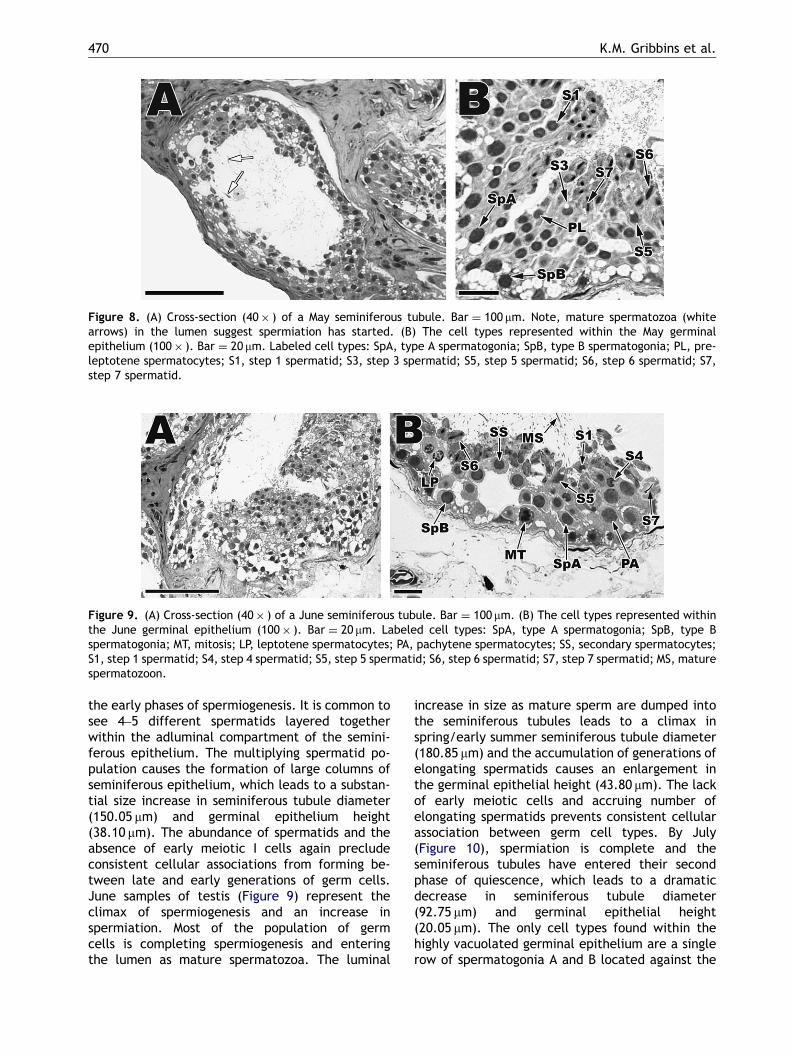
Spermatogenesis continues to advance in the Maytestis (Figure 8). The majority of the population ofgerm cells has completed meiosis II and has entered
us tubule. Bar ¼ 100 mm. (B) The cell types representedabeled cell types: SpA, type A spermatogonia; SpB, type Brmatocytes; PA, Pachytene spermatocytes; SS, secondary
us tubule. Bar ¼ 100 mm. (B) The cell types representedbeled cell types: SpA, type A spermatogonia; SpB, type Btene spermatocytes; PA, Pachytene spermatocytes; M1,p 1 spermatid; S3, step 3 spermatid; S4, step 4 spermatid.
ARTICLE IN PRESS
Figure 8. (A) Cross-section (40� ) of a May seminiferous tubule. Bar ¼ 100 mm. Note, mature spermatozoa (whitearrows) in the lumen suggest spermiation has started. (B) The cell types represented within the May germinalepithelium (100� ). Bar ¼ 20 mm. Labeled cell types: SpA, type A spermatogonia; SpB, type B spermatogonia; PL, pre-leptotene spermatocytes; S1, step 1 spermatid; S3, step 3 spermatid; S5, step 5 spermatid; S6, step 6 spermatid; S7,step 7 spermatid.
Figure 9. (A) Cross-section (40� ) of a June seminiferous tubule. Bar ¼ 100 mm. (B) The cell types represented withinthe June germinal epithelium (100� ). Bar ¼ 20 mm. Labeled cell types: SpA, type A spermatogonia; SpB, type Bspermatogonia; MT, mitosis; LP, leptotene spermatocytes; PA, pachytene spermatocytes; SS, secondary spermatocytes;S1, step 1 spermatid; S4, step 4 spermatid; S5, step 5 spermatid; S6, step 6 spermatid; S7, step 7 spermatid; MS, maturespermatozoon.
K.M. Gribbins et al.470
the early phases of spermiogenesis. It is common tosee 4–5 different spermatids layered togetherwithin the adluminal compartment of the semini-ferous epithelium. The multiplying spermatid po-pulation causes the formation of large columns ofseminiferous epithelium, which leads to a substan-tial size increase in seminiferous tubule diameter(150.05 mm) and germinal epithelium height(38.10 mm). The abundance of spermatids and theabsence of early meiotic I cells again precludeconsistent cellular associations from forming be-tween late and early generations of germ cells.June samples of testis (Figure 9) represent theclimax of spermiogenesis and an increase inspermiation. Most of the population of germcells is completing spermiogenesis and enteringthe lumen as mature spermatozoa. The luminal
increase in size as mature sperm are dumped intothe seminiferous tubules leads to a climax inspring/early summer seminiferous tubule diameter(180.85 mm) and the accumulation of generations ofelongating spermatids causes an enlargement inthe germinal epithelial height (43.80 mm). The lackof early meiotic cells and accruing number ofelongating spermatids prevents consistent cellularassociation between germ cell types. By July(Figure 10), spermiation is complete and theseminiferous tubules have entered their secondphase of quiescence, which leads to a dramaticdecrease in seminiferous tubule diameter(92.75 mm) and germinal epithelial height(20.05 mm). The only cell types found within thehighly vacuolated germinal epithelium are a singlerow of spermatogonia A and B located against the

ARTICLE IN PRESS
Figure 10. (A) Sagittal section (40� ) of a July seminiferous tubule. Bar ¼ 100 mm. (B) The cell types representedwithin the July germinal epithelium (100� ). Bar ¼ 20 mm. Labeled cell types: SpA, type A spermatogonia; SpB, type Bspermatogonia. Note: Germ cells from the previous cycle are being shed into the lumen of the seminiferous tubule,white arrow.
Figure 11. (A) Cross-section (40� ) of an August seminiferous tubule. Bar ¼ 100 mm. (B) The cell types representedwithin the August germinal epithelium (100� ). Bar ¼ 20 mm. Labeled cell types: SpA, type A spermatogonia; SpB, typeB spermatogonia; PL, pre-leptotene; PA, pachytene spermatocytes; SS, secondary spermatocytes; S1, step 1 spermatid;S2, step 2 spermatid; S3, step 3 spermatid; S6, step 6 spermatid; S7, step 7 spermatid; MS, mature spermatozoon.
Spermatogenesis in Agkistrodon piscivorus leucostoma 471
basement membrane. The lumina of these tubulesare void of most spermatozoa and have pieces ofthe seminiferous epithelium with left over germcells (Figure 10B, white arrow) from the springcycle of spermatogenesis.
The second wave of spermatogenesis has begunin the August Cottonmouth testis (Figure 11). Theearly stages of proliferation and meiosis are similarto the March and April samples; however, spermio-genic cells are in more advanced stages than Maytestes. August is the only month in which prolif-erative, meiotic, and spermiogenic cells are foundtogether regularly in the seminiferous epithelium,leading to the largest increase in seminiferoustubule diameter (227.70 mm) and germinal epithe-lial height (46.10 mm). Although all three stages of
spermatogenesis are observed in August, no con-sistent cellular association are formed becauseof the 4–5 different spermatids occupying theapical portion of the seminiferous epithelium. TheSeptember testis (Figure 12) represents an aberra-tion in the sequence of events that are occurringduring spermatogenesis. The seminiferous tubuleswithin this testis are shriveled and only A and Btype spermatogonia are present in the seminiferousepithelium that is very thin in comparison to theother represented months (seminiferous tubulediameter, 89.35 mm; germinal epithelial height,19.30 mm). Large numbers of immature spermatidshave been sloughed into the seminiferous tubules inthe September samples (Figure 12B, Sp). Sperma-togenesis in the October seminiferous epithelium

ARTICLE IN PRESS
Figure 12. (A) Cross-section (40� ) of a September seminiferous tubule. Bar ¼ 100 mm. Note: Many shed generations ofgerm cells are found in the lumen of the seminiferous tubules (black arrows). (B) The cell types represented within theSeptember germinal epithelium (100� ). Bar ¼ 20 mm. Labeled cell types: SpA, type A spermatogonia; SpB, type Bspermatogonia. Note: Many shed generations of spermatids are found in the lumen of the seminiferous tubules (Sp).
Figure 13. (A) Cross-section (40� ) of an October seminiferous tubule. Bar ¼ 100 mm. (B) The cell types representedwithin the October germinal epithelium (100� ). Bar ¼ 20 mm. Labeled cell types: SC, Sertoli cell nucleus; SpA, type Aspermatogonia; SpB, type B spermatogonia; PL, pre-leptotene; S1, step 1 spermatid; S3, step 3 spermatid; S4, step 4spermatid; S5, step 5 spermatid; S6, step 6 spermatid; S7, step 7 spermatid; MS, mature spermatozoon.
Figure 14. (A) Sagittal section (40� ) of a November seminiferous tubule. Bar ¼ 100 mm. (B) The cell types representedwithin the November germinal epithelium (100� ). Bar ¼ 20 mm. Labeled cell types: SpA, type A spermatogonia; SpB,type B spermatogonia; PL, pre-leptotene spermatocyte. Note: Remnant deteriorating germ cells are being shed into thelumen of the seminiferous tubule (white arrows: spermatocytes; white arrowhead: elongating spermatid).
K.M. Gribbins et al.472
(Figure 13) has advanced into spermiogenesiswith round and elongating spermatids represented.Spermatocytes have been exhausted and sperma-
togonia A and B are found near the base-ment membrane of the seminiferous tubules. Theloss of meiotic cells has slightly decreased the

ARTICLE IN PRESS
Spermatogenesis in Agkistrodon piscivorus leucostoma 473
seminiferous tubule diameter (215.40 mm) andgerminal epithelial (41.55 mm) height when com-pared with August seminiferous tubules. Many ofthe developing spermatids have completed sper-miogenesis and are being shed to the lumina of theseminiferous tubules as mature spermatozoa. Likethe July sample, the November seminiferoustubules (Figure 14) are in a state of quiescencewith spermatogonia A and B making up the majorityof germ cells and lipid-rich vacuoles dominating theseminiferous epithelium. The seminiferous tubulediameter (94.25 mm) and germinal epithelial height(20.15 mm) in November also mirror that of Julytestes.
Discussion
The testes of the A. piscivorus leucostomaconsist of seminiferous tubules lined by seminifer-ous epithelia. Sertoli cells are present at all timesof the year and are associated with a continuoussupply of spermatogonia. This overall testicularstructure is consistent with nonmammalian amnio-tic testes (Pudney, 1995) and the testes of BlackSwamp Snakes (Gribbins et al., 2005).
Spermatogenesis takes place during two inde-pendent events between the months of March–Juneand August–October in this Louisiana population ofA. piscivorus leucostoma. Monthly morphometricdata, testicular volume measurements, and histo-logical analysis of the Cottonmouth testis allprovide strong support for a biannual/bimodal typeof spermatogenesis. However, it should be notedthat the September testis does not fall intosequence as far as spermatogenesis with that ofthe other fall samples. In September, only sperma-togonia are seen within the seminiferous epithe-lium and this snake has the smallest seminiferoustubule diameter (89.35 mm) and germinal epithelialheight (19.30 mm) measured during this entirestudy. August testes show the early events ofspermatogenesis and October seminiferous tubulesshow the later events of spermiogenesis andspermiation. September seminiferous epitheliashould be dominated by spermatocytes and earlyround spermatids, similar to the September testisdescribed for populations of Cottonmouths inAlabama (Johnson et al., 1982). This particularsnake was caught outside of its normal habitat and2 weeks after hurricane Katrina. Thus, this snakewas exposed to extreme stress and the spermato-genic cycle may have been shut down in response tothis environmental catastrophe due to conflictbetween the hypothalamic–pituitary–gonadal axisand the hypothalamic–pituitary–adrenal axis as
demonstrated in stress-induced Cottonmouths(Graham, 2006) and supported by the large numberof detached spermatids found in the Septemberseminiferous tubular lumen (Figure 12B, Sp). TheAugust, October, and November samples all show asimilar histological sequence to the fall spermato-genic cycle described by Johnson et al. (1982) forA. piscivorus in Alabama.
The two spermatogenic events within this Cot-tonmouth population are separated by a quiescentperiod (July) in which the testes are inactive andonly spermatogonia type A and B are present andthe seminiferous tubule diameter, germinal epithe-lial height and testicular volume are smaller thanall other measurements except for the months ofSeptember and January (testicular volume only). Toour knowledge, this is the first evidence of bimodalspermatogenesis described for a temperate specieswithin Crotalinae. Like other crotalids, mosttemperate snakes have a single annual spermato-genic cycle that typically follows a postnuptialpattern where spermatogenesis commences afterspring mating (Licht, 1984; Saint Girons, 1982).Other studies (Johnson et al., 1982) on A. pisci-vorus have shown that spermatogenesis starts inthe spring, peaks in late summer, and terminates inthe fall. The populations of Cottonmouths studiedby Johnson et al. (1982) were found in Alabama(at a more northern latitude versus southernLouisiana). The further north a population of snakesresides, the shorter the number of warm months,which may limit energy sources and the metabolismneeded to maintain sperm development. This maybe the reason why the early (spring) spermatogeniccycle is absent in the Alabama Cottonmouthpopulation.
Interestingly, Johnson et al. (1982) and Severet al. (2008) both provide data for hypertrophicrenal sexual segments (RSSs) during spring and latesummer/early fall even though there is only onelate summer testosterone peak (Johnson et al.,1982; Graham, 2006). Though testosterone levelswere not taken during the present study, it is worthnoting that the literature to date suggests thathypertrophic RSSs in the Cottomouth do notparallel the timing of peak testosterone as pre-cisely as those found in other squamates (Bishop,1959; Misra and Prasad, 1965; Prasad and Sanyal,1969; Krohmer, 1986). Nevertheless, RSSs docorrelate strongly with the timing of spermatogen-esis and when breeding may occur. HypertrophicRSSs are often good indicators of breeding becausesexual segments in reptiles are presumed toproduce the seminal fluid added to spermatozoaduring ejaculation (Prasad and Reddy, 1972).Furthermore, sperm aggregates found in the

ARTICLE IN PRESS
K.M. Gribbins et al.474
posterior uterus, alleged artifacts of recent breed-ing (Saint Girons, 1957, 1962a, b), of femaleCottonmouths, also occur during the spring andfall in Louisiana (Siegel and Sever, 2008).
In light of the breeding/endocrine/RSSs data,we suggest that the two spermatogenic cyclesobserved in A. piscivorus leucostoma from south-eastern Louisiana provide mature spermatozoa forthe two major breeding periods suggested by Siegeland Sever (2008) and supported by previous studies(Beyer, 1898; Martin, 1984). The majority of femaleA. piscivorus practice biennial breeding, wherecopulation initially begins in late summer andcontinues through early fall (Burkett, 1966; Whar-ton, 1966; Ford, 2002; Ford et al., 2004). Also,during this late summer period, vitellogenesis (yolkaccumulation in the developing oocyte) presumablybegins and is subsequently halted during hiberna-tion (Aldridge and Duvall, 2002). The first sperma-togenic cycle ending in June would providespermatozoa for these late summer/fall breedingevents. Vitellogenesis in females resumes again inthe following early spring, when the second matingseason starts, and is completed by late spring/earlysummer when mature ovarian follicles are ready forovulation (Burkett, 1966). The second (fall) sper-matogenic cycle is completed before hibernationand sperm is stored in the excurrent duct systemuntil emergence from hibernation in the followingspring, which is when the second mating seasonbegins (Johnson et al., 1982).
Recent studies on temperate squamates(Gribbins and Gist, 2003; Gribbins et al., 2005)have shown a similar temporal germ cell develop-ment strategy described here for the Cottonmouth,which is different from the spatial germ celldevelopment seen in seasonally and continuallybreeding birds and mammals (Yamamoto et al.,1967; Rossen-Runge, 1977; Tait and Johnson, 1982;Tsubota and Kanagawa, 1989; Tiba and Kita, 1990;Foreman, 1997). This episodic germ cell develop-ment has also been observed within the otherreptilian orders (Gribbins et al., 2003: Trachemysscripta, Chelonia; Gribbins et al., 2006: Alligatormississippiensis, Crocodylia) and is very similar tothe temporal germ cell development strategy ofderived amphibians such as anurans (Lofts, 1964;Van Oordt and Brands, 1970). Furthermore, anuranamphibians have been described as a transitionaltaxon between the anamniotes and amniotes interms of testicular organization (Van Oordt, 1955).Yet, the seminiferous tubules in anurans are linedwith seasonal cysts and not a continuous epitheliumlike that of other amniotes. Extant reptiles andpresumably their ancestors are considered themost primitive amniotes phylogenetically and have
testes that are structurally similar to that ofderived amniotic lineages (birds and mammals).Thus, reptiles might represent a better transitionalintermediary in terms of testicular organizationbetween anurans and the derived avian andmammalian taxa.
Temperate reproductive strategies seem to haveno effect on the type of germ cell developmentstrategy employed by the reptiles studied to date.Previously studied prenuptial (Gribbins et al.,2006), postnuptial (Gribbins et al., 2003, 2005),mixed (Gribbins and Gist, 2003), and the presentresults on bimodal spermatogenesis all express thesame temporal type of germ cell development. Itwould be interesting to test whether continuallyreproducing populations of reptiles, such as thosein the tropics, would show this same temporal germcell development or if they have a more spatialgerm cell development strategy as observed incontinually breeding mammals and birds. Unfortu-nately, data on the details of spermatogenesis fortropical and temperate reptilian species are lackingand need to be addressed in order for suchcomparative models to be tested. These types ofcomparative, phylogenetic, and anatomical ques-tions on spermatogenesis and the morphology ofthe testis cannot be answered conclusively untilfurther information is collected from a variety oftropical and temperate species representing themajor taxa within Reptilia.
Acknowledgments
We thank Robert Aldridge for his helpful com-ments on this manuscript. This study is funded inpart by competitive research grants from Witten-berg University.
References
Aldridge, R.D., 2002. The link between mating seasonand male reproductive anatomy in the rattlesnakesCrotalus viridis oreganus and Crotalus viridis helleri.J. Herpetol. 36, 295–300.
Aldridge, R.D., Duvall, D., 2002. Evolution of the matingseason in the pitvipers of North America. Herpetol.Monogr. 16, 1–25.
Bertona, M., Chiaraviglio, M., 2003. Reproductive biol-ogy, mating aggregations, and sexual dimorphismof the Argentine Boa Constrictor (Boa constrictoroccidentalis). J. Herpetol. 37, 510–516.
Beyer, G.E., 1898. Contribution on the life histories ofcertain snakes. Am. Nat. 32, 17–24.
Bishop, J.E., 1959. A histological and histochemical studyof the kidney tubule of the common Garter Snake,

ARTICLE IN PRESS
Spermatogenesis in Agkistrodon piscivorus leucostoma 475
Thamnophis sirtalis, with special reference to thesexual segment in the male. J. Morphol. 104, 307–357.
Brown, C.R., Bomberger Brown, M., 2003. Testis sizeincreases with colony size in Cliff Swallows. Behav.Ecol. 14, 569–575.
Burkett, R.D., 1966. Natural history of the CottonmouthMoccasin, Agkistrodon piscivorus (Reptilia). Nat. Hist.17, 435–491.
Conant, R., Collins, J.T., 1991. Reptiles and Amphibians.Houghton Mifflin Company, MA, MA, Eastern/CentralNorth America, p. 450.
Flemming, A.F., 1993. The male reproductive cycle of theLizard Pseudocordylus m. melanotus (Sauria: Cordyli-dae). J. Herpetol. 27, 473–478.
Ford, N.B., 2002. Ecology of the western Cottonmouth(Agkistrodon piscivorus leucostoma) in northeasternTexas. In: Schuett, G.W., Hoggren, M., Douglas, M.E.,Greene, H.W. (Eds.), Biology of Vipers. Eagle MountainPublishing, Utah, pp. 167–177.
Ford, N.B., Brischoux, F., Lancaster, D., 2004. Reproduc-tion in the western cottonmouth, Agkistrodon pisci-vorus leucostoma, in a floodplain forest. Southwest.Nat. 49, 465–471.
Foreman, D., 1997. Seminiferous tubule stages in theprairie dog (Cynomys ludovicianus) during the annualbreeding cycle. Anat. Rec. 247, 355–367.
Goldberg, S.R., 1995. Reproduction in the Lyre Snake,Trimophodon biscutatus (Colubridae), from Arizona.Southwest. Nat. 40, 334–335.
Goldberg, S.R., 1998. Reproduction in the SonoranWhipsnake, Masticophis bilineatus (Serpentes: Colu-bridae). Southwest. Nat. 43, 412–414.
Graham, S., 2006. An integrative analysis of reproductionand stress in free-living male Cottonmouths, Agkis-trodon piscivorus. Science, Thesis, Georgia StateUniversity, pp. 1–85.
Gribbins, K.M., Gist, D.H., 2003. Cytological evaluationof spermatogenesis within the germinal epitheliumof the male European Wall Lizard, Podarcis muralis.J. Morphol. 258, 296–306.
Gribbins, K., Gist, D., Congdon, J., 2003. Cytologicalevaluation of spermatogenesis in the Red-eared Slider,Trachemys scripta. J. Morphol. 255, 337–346.
Gribbins, K.M., Happ, C.S., Sever, D.M., 2005. Ultra-structure of the reproductive system of the BlackSwamp Snake (Seminatrix pygaea). V. The temporalgerm cell development strategy of the testis. ActaZool. 86, 223–230.
Gribbins, K.M., Elsey, R.M., Gist, D.H., 2006. Cytologicalevaluation of the germ cell development strategywithin the testis of the American alligator, Alligatormississippiensis. Acta Zool. 87, 59–69.
Hayat, M.A., 1993. Stains and Cytochemical Methods.Plenum Press, New York and London, p. 455.
Johnson, L.F., Jacob, J.S., Torrance, P., 1982. Annualtesticular and androgenic cycles of the Cottonmouth(Agkistrodon piscivorus) in Alabama. Herpetologica38, 16–25.
Krohmer, R.W., 1986. Effects of mammalian gonadotro-pins (FSH and LH) on testicular development in the
immature water snake, Nerodia sipedon. Gen. Comp.Endocrinol. 64, 330–338.
Leblond, C.P., Clermont, Y., 1952. Spermiogenesis of rat,mouse, hamster, guinea pig as revealed by theperiodic acid-fuchsin sulfurous acid technique. Am.J. Anat. 90, 167–215.
Licht, P., 1984. Reptiles. In: Lamming, G.E. (Ed.),Marshall’s Physiology of Reproduction, ReproductiveCycle of Vertebrates, vol. 1. Churchill Livingstone,New York, pp. 206–282.
Lofts, B., 1964. Seasonal changes in the functionalactivity of the interstitial and spermatogenetic tissuesof the green frog, Rana esculenta. Gen. Comp.Endocrinol. 4, 550–562.
Martin, D.L., 1984. An instance of sexual defense in theCottonmouth, Agkistrodon piscivorus. Copeia 1984,772–774.
Misra, UKS., Prasad, M.R.N., 1965. Phospholipids of thesexual segment of the kidney of the Indian HouseLizard, Hemidactylus flaviviridis (Ruppell). Life Sci. 4,159–166.
Prasad, M.R.N., Reddy, P.R.K., 1972. Physiology of thesexual segment of the kidney in reptiles. Gen. Comp.Endocrinol. 3, 649–662.
Prasad, M.R.N., Sanyal, M.K., 1969. Effect of sexhormone on the RSS of kidney and other accessoryreproductive organs of the Indian House LizardHemidactylus flaviviridis (Ruppell). Gen. Comp.Endocrinol. 12, 110–118.
Pudney, J., 1995. Spermatogenesis in nonmammalianvertebrates. Microsc. Res. Tech. 32, 459–497.
Ramirez-Bautista, A., Gutierrez-Mayen, G., 2003. Repro-ductive ecology of Sceloporus utiformis (Sauria:Phrynosomatidae) from a tropical dry forest of Mexico.J. Herpetol. 37, 1–10.
Rossen-Runge, E.C., 1977. The Process of Spermatogen-esis in Animals. Cambridge University Press, Cam-bridge, UK.
Russell, L.D., Hikim, S.A.P., Ettlin, R.A., Legg, E.D.,1990. Histological and Histopathological Evaluation ofthe Testis. Cache River Press, Florida.
Saint Girons, H., 1957. Le cycle sexuel chez Vipera aspis(L.) dans l’oues de la France. Bull. Biol. FranceBelg. 91, 284–350.
Saint Girons, H., 1962a. Le cycle reproducteur de lavipere a cornes, Cerastes cereastes (L). dans la natureet en captivite. Bull. Zool. France 87, 41–51.
Saint Girons, H., 1962b. Presence de receptacle seminauxchez les cameleons. Beaufortia 9, 165–172.
Saint Girons, H., 1982. Reproductive cycles ofmale snakes and their relationships with climateand female reproductive cycles. Herpetologica 16,1–25.
Selby, S.M., 1965. Standard Math Tables, fourteenth ed.Chemical Rubber Co., Cleveland, OH.
Sever, D.M., Siegel, D.S., Bagwill, A., Eckstut, M.E.,Alexander, L., Camus, A., Morgan, C., 2008. Renalsexual segment of the Cottonmouth snake, Agkistro-don piscivorus (Reptilia, Squamata, Viperidae).J. Morphol. (Available on early web view).

ARTICLE IN PRESS
K.M. Gribbins et al.476
Shea, G.M., 2001. Spermatogenic cycle, sperm storage,and Sertoli cell size in a Scloecophidian (Ramphotyph-lops nigrescens) from Australia. J. Herpetol. 35, 85–91.
Siegel, D.S., Sever, D.M., 2008. Sperm aggregations infemale Agkistrodon piscivorus (Reptilia: Squamata):a histological and ultrastructural investigation.J. Morphol. 269, 189–206.
Sokal, R.R., Rohlf, F.J., 1995. Biometry, third ed. W.H.Freeman, San Francisco, CA.
Srivastava, P.C., Thapliyal, J.P., 1965. The male sexualcycle of the Chequered Water Snake, Natrix psicator.Copeia 1965, 410–415.
Tait, A.J., Johnson, E., 1982. Spermatogenesis in theGrey Squirrel (Sciurus carolinensis) and changes duringsexual regression. J. Reprod. Fertil. 65, 53–58.
Tiba, T., Kita, I., 1990. Undifferentiated spermato-gonia and their role in the seasonally fluctuatingspermatogenesis in the ferret, Mustela putorius furo(Mammalia). Zool. Anz. 224, 140–155.
Tsubota, T., Kanagawa, H., 1989. Annual changes inserum testosterone levels and spermatogenesis in the
Hokkaido Brown Bear, Ursus arctos yesoensis.J. Mammal. Soc. Japan 14, 11–17.
Van Oordt, P.G.W.J., 1955. Regulation of the spermato-genetic cycle in the frog. Mem. Soc. Endocrinol. 4,25–38.
Van Oordt, P.G.W.J., Brands, F., 1970. The Sertoli cell inthe testis of the common frog, Rana temporaria. In:Proceedings of the Society of Endocrinology 119thMeeting, J. Endocrinol., 48, Abs 100.
Wharton, C.H., 1966. Reproduction and growth inCottonmouths, Agkistrodon piscivorus (Lacepede) ofCedar Keys, Florida. Copeia 1966, 149–161.
Yamamoto, S., Tamate, H., Itikawa, O., 1967. Morpholo-gical studies on the sexual maturation in the maleJapanese Quail (Coturnix coturnix jaonica). TohukuJ. Agric. Response 18, 27–37.
Zaidan III, F., Kreider, D.L., Beaupre, S.J., 2003.Testosterone cyles and reproductive energetics: im-plications for northern range limits of the Cotton-mouth (Agkistrodon piscivorus leucostoma). Copeia2003, 231–240.