renal mechanisms - hypertensionhyper.ahajournals.org/content/hypertensionaha/49/1/148.full.pdf ·...
TRANSCRIPT

Role of Renal Medullary Heme Oxygenase in the Regulationof Pressure Natriuresis and Arterial Blood Pressure
Ningjun Li, Fan Yi, Elisabete A. dos Santos, Dustin K. Donley, Pin-Lan Li
Abstract—Recent studies have demonstrated that inhibition of renal medullary heme oxygenase (HO) activity and carbonmonoxide (CO) significantly decreases renal medullary blood flow and sodium excretion. Given the crucial role of renalmedullary blood flow in the control of pressure natriuresis, the present study was designed to determine whether renalmedullary HO activity and resulting CO production participate in the regulation of pressure natriuresis and thereby thelong-term control of arterial blood pressure. In anesthetized Sprague–Dawley rats, increases in renal perfusion pressureinduced significant elevations of CO concentrations in the renal medulla. Renal medullary infusion of chromiummesoporphyrin (CrMP), an inhibitor of HO activity, remarkably inhibited HO activity and the renal perfusionpressure–dependent increases in CO levels in the renal medulla and significantly blunted pressure natriuresis. Inconscious Sprague–Dawley rats, continuous infusion of CrMP into the renal medulla significantly increased meanarterial pressure (129�2.5 mm Hg in CrMP group versus 118�1.6 mm Hg in vehicle group) when animals were fed anormal salt diet (1% NaCl). After rats were switched to a high-salt diet (8% NaCl) for 10 days, CrMP-treated animalsexhibited further increases in mean arterial pressure compared with CrMP-treated animals that were kept on normal saltdiet (152�4.1 versus 130�4.2 mm Hg). These results suggest that renal medullary HO activity plays a crucial role inthe control of pressure natriuresis and arterial blood pressure and that impairment of this HO/CO-mediatedantihypertensive mechanism in the renal medulla may result in the development of hypertension. (Hypertension. 2007;49:148-154.)
Key Words: carbon monoxide � sodium excretion � bilirubin � chromium mesoporphyrin� nitric oxide � high salt
The heme oxygenase (HO) catalyzes the rate-limiting step inheme degradation, producing equimolar quantities of biliv-
erdin (BV), iron, and carbon monoxide (CO). BV is subse-quently converted to bilirubin (BR) by BV reductase.1–3 It hasbeen shown that the products of HO are involved in theregulation of cardiovascular–renal function3 and that inductionof HO-1 by pharmacological/genetic interventions improvesvascular function and ameliorates both genetic and experimentalforms of hypertension.3–5 However, most studies showing thecontribution of HO to the regulation of blood pressure aresystemic interventions and focused on the vasculature. Given thecrucial role of the kidneys in the long-term control of arterialblood pressure, it is imperative to determine whether local HOsin the kidneys participate in the regulation of the renal functionand arterial blood pressure and how HOs and their productswork to alter the blood pressure.
The 2 major functional isoforms of HO are the inducible(HO-1) and the constitutive (HO-2) forms. Numerous tis-sues,1 including the kidneys,6 express HOs. Previous studieshave reported that both HO-1 and HO-2 are more abundantlyexpressed in the renal medulla than in the renal cortex.6,7 It
has been demonstrated that HO participates in the regulationof renal circulation and sodium excretion.6,8–10 Renal medul-lary infusion of HO inhibitor decreased medullary blood flow(MBF) without changing cortical blood flow.6 Because therenal medullary circulation plays a key role in the regulationof sodium excretion and arterial blood pressure,11,12 wehypothesized that renal medullary HO activity, through itsactions of increasing MBF and sodium excretion, therebyproduces an antihypertensive effect. To test this hypothesis,we first determined the effects of HO inhibition on pressurenatriuresis in anesthetized Sprague–Dawley rats. Correspond-ingly, we analyzed HO activity using high-performanceliquid chromatography (HPLC) analysis and CO productionin response to elevations of renal perfusion pressure (RPP)using a microdialysis-oxyhemoglobin CO trapping technique.Then, we addressed the role of renal medullary HO activity inthe long-term control of arterial blood pressure by chronicmedullary infusion of an HO inhibitor. Our data suggest thatthe HO/CO system in the renal medulla plays an importantantihypertensive role by maintenance of pressure natriuresisand reduction of salt sensitivity of arterial blood pressure.
Received January 6, 2006; first decision January 24, 2006; revision accepted October 4, 2006.From the Department of Pharmacology and Toxicology (N.L., F.Y., D.K.D., P.-L.L.), Medical College of Virginia, Virginia Commonwealth
University, Richmond; and the Department of Pharmacology and Toxicology (E.A.d.S.), Medical College of Wisconsin, Milwaukee.Correspondence to Ningjun Li, Department of Pharmacology and Toxicology, Medical College of Virginia, Virginia Commonwealth University, PO
Box 980613, Richmond, VA 23298. E-mail [email protected]© 2006 American Heart Association, Inc.
Hypertension is available at http://www.hypertensionaha.org DOI: 10.1161/01.HYP.0000250086.06137.fb
148
Renal Mechanisms
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from

MethodsAnimalsExperiments were performed on male Sprague–Dawley rats weigh-ing between 300 and 350 g purchased from Harlan. All of theprotocols were approved by the institutional animal care and usecommittee of Virginia Commonwealth University and MedicalCollege of Wisconsin.
HPLC Analysis of HO ActivityRenal tissue HO activities were determined using HPLC analysis ofthe conversion of heme to bilirubin by renal cortical, outer, and innermedullary tissues as described previously.13,14 An expanded Meth-ods section is available online at http://hyper.ahajournals.org.
Effects of Renal Medullary Infusion of HOInhibitor on Pressure—NatriuresisRats were prepared for the study of pressure natriuresis as describedpreviously.15 The left kidney was stabilized, and an interstitialcatheter was placed into the renal medulla (5 mm in depth) forcontinuous renal medullary infusion of CrMP (10 nmol/kg perminute) or vehicle at a rate of 10 �L/min throughout the experi-ment.6 Renal blood flow (RBF) was measured with a transonic flowprobe around the left renal artery (Transonic) as described previ-ously.16 Glomerular filtration rate was measured by a fluoresceinisothiocyanate–inulin (Sigma-Aldrich) clearance rate as describedpreviously by us and others.16,17 Mean arterial pressure (MAP) andRBF were recorded, and urine samples were collected for 30 minutesat each RPP level after a 10-minute equilibration period. GFR, urineflow, and urinary Na� excretion were factored per gram of kidneyweight (kw). For the details, see the expanded Methods section online.
Assay of Renal Medullary CO ConcentrationsAn in vivo microdialysis–oxyhemoglobin CO trapping assay wasestablished to determine renal medullary CO concentrations. COreacts with ferrous oxyhemoglobin (OxyHb) to form carboxyhemo-globin (CO-Hb), which can be spectrophotometrically detected tomeasure the endogenous CO formation.9,18–21 In vivo microdialysisexperiments were performed as described in our previous stud-ies.22,23 The animals were prepared as described above for the studiesof pressure natriuresis. At each RPP level, a 30-minute dialysatesample was collected for measurement of CO-Hb and calculation ofCO concentration using the formulas reported previously.18,21,24 Forthe details, see the expanded Methods section online.
Assay of Renal Medullary NO ConcentrationsAn in vivo microdialysis-OxyHb NO trapping assay was used todetermine renal medullary NO concentrations as described previously.23
Effects of Chronic Renal Medullary Infusion ofHO Inhibitor on Arterial Blood PressureThe rats were surgically prepared for chronic renal medullaryinfusion and arterial blood pressure monitoring with a telemetrysystem. There were 3 groups including vehicle�high salt,CrMP�high salt, and CrMP�normal salt. The animals were fed witha normal salt diet for 3 days and then a high-salt diet for 10 days. Forthe details, see the expanded Methods section.
Effects of High Salt Intake and Chronic RenalMedullary Infusion of HO Inhibitor on the mRNAand Protein Expressions of HO-1 and HO-2 in theRenal MedullaAfter the chronic CrMP infusion described above, the expression ofHO-1 and HO-2 mRNA and protein in the renal medulla weredetected using real-time RT- PCR (TaqMan Gene Expression Assayskits, Applied Biosystems) and Western blot analyses using antibod-ies against rat HO-1 and HO-2 (Santa Cruz Biotechnology).
Statistical AnalysisData are presented as mean�1 SE. The significance of differences inmean values within and between multiple groups was evaluatedusing an ANOVA followed by a Duncan’s multiple range test.Student t test was used to evaluate the statistical significance ofdifferences between 2 groups. P�0.05 was considered statisticallysignificant.
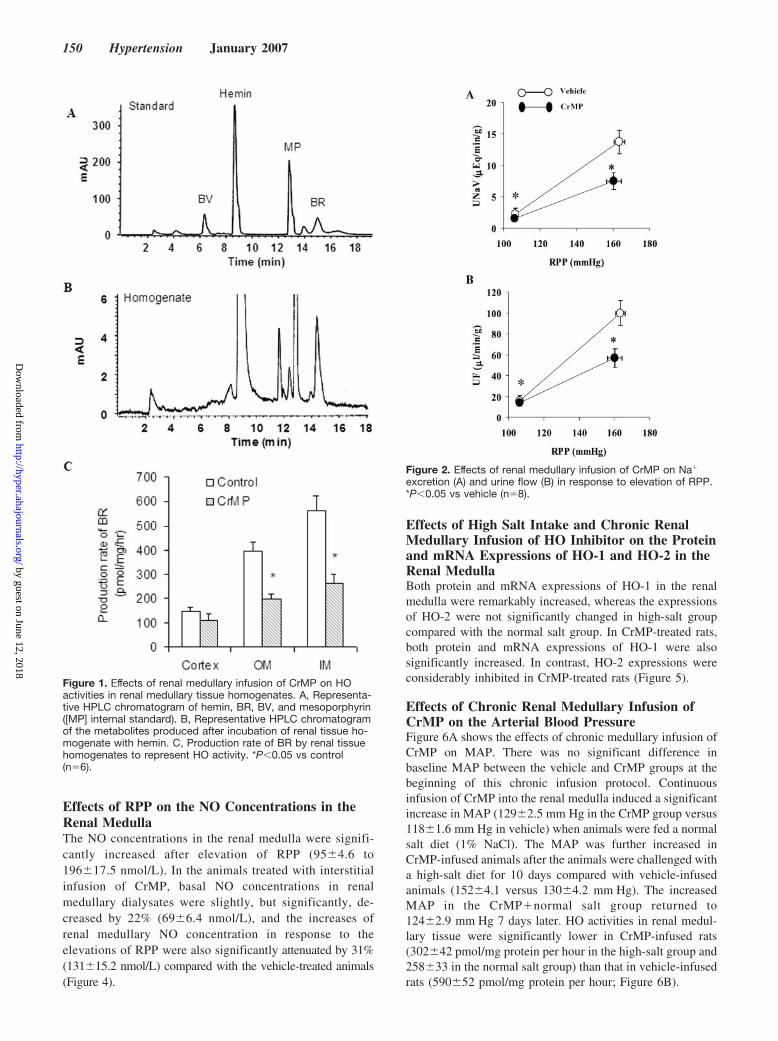
ResultsEffects of Renal Medullary Infusion of CrMP onHO Activity in the KidneysHemin, the substrate of HO, and its products, BV and BR, andinternal standard (mesoporphyrin) were clearly separated byan HPLC chromatogram when mixed in standard solution(Figure 1A). After incubation of renal tissue homogenate withhemin, the production of BR was detected (Figure 1B). HOactivities, as presented by the production rates of BR, weresignificantly decreased by 50% in the outer medulla and innermedulla by renal medullary infusion of CrMP, whereas nosignificant decrease in HO activity was observed in corticaltissue homogenates (Figure 1C).
Effects of Renal Medullary Infusion of CrMP onPressure—NatriuresisAcute renal medullary infusion of CrMP did not changethe baseline MAP. Elevation of RPP in control rats signifi-cantly increased sodium excretion from 2.26�0.28 to13.71�1.87 �mol/min per gram of kw. In CrMP-treatedanimals, the diuretic and natriuretic responses to the eleva-tions of RPP were blunted by 52% (from 1.54�0.32 to6.64�1.57 �mol/min per gram of kw; Figure 2). There wasno significant difference in RBF and GFR between controland CrMP-treated rats (data not shown).
Effects of RPP on the CO Concentrations in theRenal MedullaFigure 3A shows the linear correlation between the measuredCO concentrations and the standard concentrations of CO-saturated solution (r�0.999). The minimal detectable COconcentration was 10 nmol/L. The CO concentrations�40 �mol/L saturated the OxyHb at the OxyHb concentra-tion used in the present experiment. Dialysis efficiency of COthrough microdialysis probe was determined from in vitrodialysis experiments in which concentrations of CO in thedialysates were compared with CO concentrations in theserial dilutions of CO-saturated solution. An 86.9% recoverywas observed in the effluent dialysate solution. Figure 3Bpresents CO concentrations in renal medullary dialysates atdifferent RPP levels with and without renal medullary infu-sion of CrMP. Basal CO concentrations in the renal medullaaveraged 334�58 nmol/L, which were significantly increasedto 873�184 nmol/L after elevation of RPP. In the animalstreated with interstitial infusion of CrMP (10 nmol/kg perminute), basal CO concentrations in renal medullary dialy-sates were significantly decreased by 63% (129�27 nmol/L), and the increases of renal medullary CO concentrationin response to the elevations of RPP were also significantlyattenuated by 54% (399�52 nmol/L) compared with thevehicle-treated animals.
Li et al Renal Medullary Heme Oxygenase 149
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
Effects of RPP on the NO Concentrations in theRenal MedullaThe NO concentrations in the renal medulla were signifi-cantly increased after elevation of RPP (95�4.6 to196�17.5 nmol/L). In the animals treated with interstitialinfusion of CrMP, basal NO concentrations in renalmedullary dialysates were slightly, but significantly, de-creased by 22% (69�6.4 nmol/L), and the increases ofrenal medullary NO concentration in response to theelevations of RPP were also significantly attenuated by 31%(131�15.2 nmol/L) compared with the vehicle-treated animals(Figure 4).
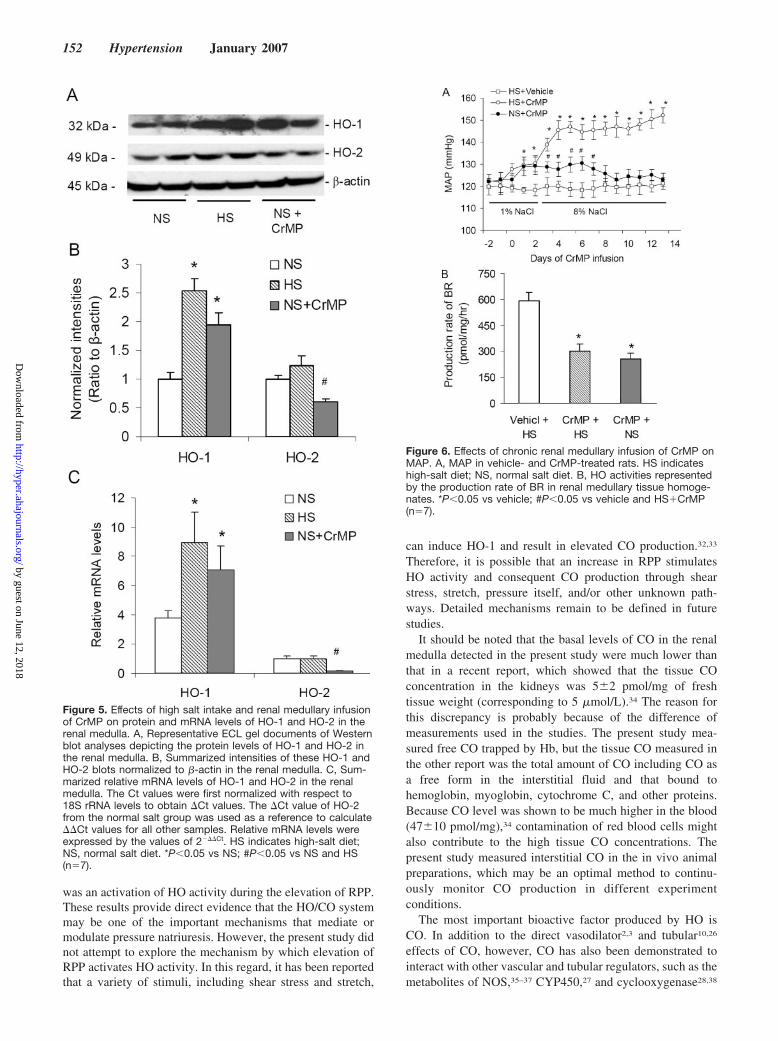
Effects of High Salt Intake and Chronic RenalMedullary Infusion of HO Inhibitor on the Proteinand mRNA Expressions of HO-1 and HO-2 in theRenal MedullaBoth protein and mRNA expressions of HO-1 in the renalmedulla were remarkably increased, whereas the expressionsof HO-2 were not significantly changed in high-salt groupcompared with the normal salt group. In CrMP-treated rats,both protein and mRNA expressions of HO-1 were alsosignificantly increased. In contrast, HO-2 expressions wereconsiderably inhibited in CrMP-treated rats (Figure 5).
Effects of Chronic Renal Medullary Infusion ofCrMP on the Arterial Blood PressureFigure 6A shows the effects of chronic medullary infusion ofCrMP on MAP. There was no significant difference inbaseline MAP between the vehicle and CrMP groups at thebeginning of this chronic infusion protocol. Continuousinfusion of CrMP into the renal medulla induced a significantincrease in MAP (129�2.5 mm Hg in the CrMP group versus118�1.6 mm Hg in vehicle) when animals were fed a normalsalt diet (1% NaCl). The MAP was further increased inCrMP-infused animals after the animals were challenged witha high-salt diet for 10 days compared with vehicle-infusedanimals (152�4.1 versus 130�4.2 mm Hg). The increasedMAP in the CrMP�normal salt group returned to124�2.9 mm Hg 7 days later. HO activities in renal medul-lary tissue were significantly lower in CrMP-infused rats(302�42 pmol/mg protein per hour in the high-salt group and258�33 in the normal salt group) than that in vehicle-infusedrats (590�52 pmol/mg protein per hour; Figure 6B).
Figure 1. Effects of renal medullary infusion of CrMP on HOactivities in renal medullary tissue homogenates. A, Representa-tive HPLC chromatogram of hemin, BR, BV, and mesoporphyrin([MP] internal standard). B, Representative HPLC chromatogramof the metabolites produced after incubation of renal tissue ho-mogenate with hemin. C, Production rate of BR by renal tissuehomogenates to represent HO activity. *P�0.05 vs control(n�6).
Figure 2. Effects of renal medullary infusion of CrMP on Na�
excretion (A) and urine flow (B) in response to elevation of RPP.*P�0.05 vs vehicle (n�8).
150 Hypertension January 2007
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from

DiscussionIn the present study, increases in RPPs elevated CO concen-trations in the renal medulla in anesthetized Sprague–Dawleyrats, whereas renal medullary infusion of CrMP blocked HOactivities and RPP-dependent increases in CO levels in therenal medulla, which was accompanied by a significantblunting of diuretic and natriuretic response to the elevationof RPP. The present study also demonstrated that high saltintake upregulated HO-1 expression, and chronic inhibitionof renal medullary HO activity increased arterial bloodpressure and salt sensitivity of MAP. These data suggest thatHO in the renal medulla plays an important role in the
regulation of renal function and long-term control of bloodpressure.
Inhibition of renal medullary HO activity has been shownto significantly reduce renal MBF.6 Given the important roleof MBF in the control of sodium excretion and arterial bloodpressure,11,12,25 the renal medullary HO/CO system, by alter-ing MBF, may be an important determinant of pressurenatriuresis and thereby the long-term control of arterial bloodpressure. In support of this notion, our data demonstrated thatinhibition of renal medullary HO activity remarkably bluntedthe pressure-natriuretic response to elevations of RPP. Thisalteration of pressure-natriuretic response by HO inhibitionmay be associated with the vasodilator actions of CO in therenal medulla and consequent increases in MBF, as shown ina previous study.6 However, because the HO/CO system hasalso been shown to directly inhibit tubular reabsorption10 andto participate in the regulation of tubular function in the loopof Henle,26 we cannot rule out a possible direct action of COon tubular sodium reabsorption. In addition, the products ofHO, including CO, BV, and BR, have been indicated to affectthe activities of NO synthase (NOS), cyclooxygenase, andcytochrome P450 (CYP450)27,28 and minimize oxidativestress,3,29 which can also mediate the indirect tubular actionsof HO in the renal medulla. As shown for many otherregulators of pressure natriuresis, it is believed that theHO/CO system may participate in the regulation of pressure-natriuretic responses through both medullary hemodynamicand tubular actions.
To further clarify the role of the renal medullary HO/COsystem in the regulation of pressure natriuresis, we measuredthe CO levels in the renal medulla at different levels of RPPby spectrophotometric analyses of the formation of CO-Hb inthe renal medullary microdialysates. In previous studies, thiscombination of in vivo microdialysis and hemoglobin-trapping techniques has been successfully used to examineNO levels in the kidneys and brain.23,30,31 The adequatesensitivity of CO-Hb assay in determining CO concentrationsand the high dialysis efficiency of CO as shown in the presentstudy make this method a useful tool in monitoring the COlevels in the renal medulla. We found that the elevation ofRPP significantly increased the CO concentration, and inhi-bition of HO activity blunted the RPP-dependent increases ofCO concentration in the renal medulla, indicating that there
Figure 4. Effects of elevation of RPP and renal medullary infu-sion of CrMP on NO concentrations in the renal medulla. Renalmedullary interstitial NO concentrations in response to elevationof RPP with and without interstitial infusion of CrMP. *P�0.05vs vehicle; #P�0.05 vs values at lower RPP (n�6).
Figure 3. Effects of elevation of RPP and renal medullary infu-sion of CrMP on CO concentrations in the renal medulla. A,Standard curve of CO-saturated solution measured by spectro-photometric assay of CO-Hb. Insert, Low range of CO-saturatedsolution. B, Renal medullay interstitial CO concentrations inresponse to elevation of RPP with and without interstitial infu-sion of CrMP. *P�0.05 vs vehicle; #P�0.05 vs values at lowerRPP (n�6).
Li et al Renal Medullary Heme Oxygenase 151
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from
was an activation of HO activity during the elevation of RPP.These results provide direct evidence that the HO/CO systemmay be one of the important mechanisms that mediate ormodulate pressure natriuresis. However, the present study didnot attempt to explore the mechanism by which elevation ofRPP activates HO activity. In this regard, it has been reportedthat a variety of stimuli, including shear stress and stretch,
can induce HO-1 and result in elevated CO production.32,33
Therefore, it is possible that an increase in RPP stimulatesHO activity and consequent CO production through shearstress, stretch, pressure itself, and/or other unknown path-ways. Detailed mechanisms remain to be defined in futurestudies.
It should be noted that the basal levels of CO in the renalmedulla detected in the present study were much lower thanthat in a recent report, which showed that the tissue COconcentration in the kidneys was 5�2 pmol/mg of freshtissue weight (corresponding to 5 �mol/L).34 The reason forthis discrepancy is probably because of the difference ofmeasurements used in the studies. The present study mea-sured free CO trapped by Hb, but the tissue CO measured inthe other report was the total amount of CO including CO asa free form in the interstitial fluid and that bound tohemoglobin, myoglobin, cytochrome C, and other proteins.Because CO level was shown to be much higher in the blood(47�10 pmol/mg),34 contamination of red blood cells mightalso contribute to the high tissue CO concentrations. Thepresent study measured interstitial CO in the in vivo animalpreparations, which may be an optimal method to continu-ously monitor CO production in different experimentconditions.
The most important bioactive factor produced by HO isCO. In addition to the direct vasodilator2,3 and tubular10,26
effects of CO, however, CO has also been demonstrated tointeract with other vascular and tubular regulators, such as themetabolites of NOS,35–37 CYP450,27 and cyclooxygenase28,38
Figure 5. Effects of high salt intake and renal medullary infusionof CrMP on protein and mRNA levels of HO-1 and HO-2 in therenal medulla. A, Representative ECL gel documents of Westernblot analyses depicting the protein levels of HO-1 and HO-2 inthe renal medulla. B, Summarized intensities of these HO-1 andHO-2 blots normalized to �-actin in the renal medulla. C, Sum-marized relative mRNA levels of HO-1 and HO-2 in the renalmedulla. The Ct values were first normalized with respect to18S rRNA levels to obtain �Ct values. The �Ct value of HO-2from the normal salt group was used as a reference to calculate��Ct values for all other samples. Relative mRNA levels wereexpressed by the values of 2���Ct. HS indicates high-salt diet;NS, normal salt diet. *P�0.05 vs NS; #P�0.05 vs NS and HS(n�7).
Figure 6. Effects of chronic renal medullary infusion of CrMP onMAP. A, MAP in vehicle- and CrMP-treated rats. HS indicateshigh-salt diet; NS, normal salt diet. B, HO activities representedby the production rate of BR in renal medullary tissue homoge-nates. *P�0.05 vs vehicle; #P�0.05 vs vehicle and HS�CrMP(n�7).
152 Hypertension January 2007
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from

to exert related vascular and tubular regulatory action. In thisregard, the evidence for the interaction of HO/CO and NO isconflicting. It has been reported that low level of CO inducesNO release, whereas a high level of CO suppresses NOSactivity and decreases NO production.36 To address whetherinhibition or activation of HO in the present study wouldincrease or decrease the NO production, we also measured theNO concentrations in the renal medulla during activation ofHO by elevated RPP and the inhibition of HO by CrMP. Ourresults demonstrated that elevation in RPP increased NOproduction in the renal medulla, which is consistent withprevious studies.39 Inhibition of HO caused small but signif-icant decreases in both basal and RPP-induced production ofNO in the renal medulla. Because concentration ranges of COreported to stimulate NO release are at nanomoles per literand to inhibit NO production at micromoles per liter, thenanomoles per liter range of CO detected in the renal medullain the present study falls into the “low” level of CO, whichacts to stimulate NO release. Therefore, inhibition of HOblocked the stimulation of NO production by CO and de-creased the NO concentration in the renal medulla. These datafurther support the view that there is an interaction betweenHO/CO and NO in the renal medulla and that both of them areinvolved in the pressure natriuresis process.
There is a concern regarding the role of CO-mediatedregulation of CYP450 in the pressure natriuresis. CO inhibitsthe activity of CYP450 and thereby blocks the generation ofvasoconstrictive substances, such as 20-HETE.3,27,40 It hasbeen shown that 20-HETE has potent natriuretic effects, anddecreased 20-HETE levels are responsible for the bluntedpressure natriuresis and salt-sensitive hypertension in theDahl salt-sensitive rat.15,41 Therefore, inhibition of HO in thepresent study could be speculated to increase the productionof 20-HETE, which could result in natriuretic effects. How-ever, 20-HETE possesses both natriuretic tubular actions andantinatriuretic vascular actions.40,41 Its natriuretic tubularactions mainly target proximal tubules,1541 and inhibition of20-HETE has been shown to increase renal MBF.40 Withrespect to the possible increase of 20-HETE by CrMP in thepresent study, blunt of pressure natriuresis by renal medullaryinhibition of HO could be partially attributed to the vasocon-strictor effect of 20-HETE in the renal medulla, becauseCrMP was administered locally.
Because of a major role of pressure natriuresis in thelong-term control of arterial blood pressure and sodiumbalance,42 the regulation of pressure natriuresis by the renalmedullary HO/CO system may represent an important intra-renal antihypertensive mechanism. It was assumed that in-creased production of CO in response to elevations of RPPimportantly contributes to the long-term control of sodiumbalance and arterial blood pressure. To test this hypothesis,we further examined the effects of chronic inhibition of renalmedullary HO activity on MAP and the salt sensitivity ofMAP. Our results showed that high salt intake significantlyupregulated HO-1 expression, which may be mediated byhigh-salt–induced activation of hypoxia-inducible factor 1�in the renal medulla.43–45 It was also found that chronicallycontinuous infusion of CrMP into the renal medulla inhibitedHO activity and resulted in hypertension and increased salt
sensitivity of MAP. Interestingly, in the animals kept onnormal salt diet, CrMP initially induced an increase of MAP,but the MAP returned to normal 7 days later. This is probablybecause of other adaptive mechanisms to compensate for theinhibitory effect of CrMP on sodium excretion. However, thecompensative mechanisms could not overcome the antinatri-uretic effect induced by CrMP under high-salt diet, and theMAP continuously increased, indicating that the HO/COsystem is an important pathway in renal adaptation tohigh-salt intake. These results provide direct evidence thatrenal medullary HO activity possesses important antihyper-tensive action and thereby contributes to the long-termcontrol of arterial blood pressure and the determination of saltsensitivity of arterial blood pressure. As discussed above, thisrenal medullar antihypertensive action of the HO/CO systemmay be attributed to its action on both vascular bed andtubules in the renal medulla.
PerspectivesThe present study showed that elevation in RPP activated HOactivity in the renal medulla and that locally inhibition of HOactivity in this kidney region significantly blunted natriureticresponse to the elevation of RPP, and chronic inhibition ofrenal medullary HO activity induced hypertension and in-creased salt sensitivity of arterial blood pressure. Theseresults suggest a contributing role of renal medullary HOactivity to the renal adaptation to high-salt intake and thelong-term control of arterial blood pressure. Therefore, im-pairment in the HO/CO system in the renal medulla withconsequent blunting of pressure natriuresis may be one of theimportant renal mechanisms leading to the development ofhypertension.
Sources of FundingThis study was supported by National Institutes of Health grantsHL-70726 and DK-54927.
DisclosuresNone.
References1. Maines MD. The heme oxygenase system. A regulator of second mes-
senger gases. Ann Rev Pharmacol Toxicol. 1997;37:517–554.2. Ndisang JF, Tabien HE, Wang R. Carbon monoxide and hypertension.
J Hypertens. 2004;22:1057–1074.3. Abraham NG, Kappas A. Heme oxygenase and the cardiovascular-renal
system. Free Rad Biol Med. 2005;39:1–25.4. Yang L, Quan S, Nasjletti A, Laniado-Schwartzman M, Abraham NG.
Heme Oxygenase-1 gene expression modulates angiotensin II-inducedincrease in blood pressure. Hypertension. 2004;43:1221–1226.
5. Botros FT, Schwartzman ML, Stier CT, Goodman AI, Abraham NG.Increase in heme oxygenase-1 levels ameliorates renovascular hyper-tension. Kidney Int. 2005;68:2745–2755.
6. Zou A-P, Billington H, Su N, Cowley AW Jr. Expression and actions ofheme oxygenase in the renal medulla of rats. Hypertension. 2000;35:342–347.
7. Hu Y, Ma N, Yang M, Semba R. Expression and distribution of hemeoxygenase-2 mRNA and protein in rat kidney. J Histochem Cytochem.1998;46:249–256.
8. Mustafa MR, Johns EJ. The role of haem oxygenase in renal vascularreactivity in normotensive and hypertensive rats. J Hypertens. 2001;19:1105–1111.
9. Arregui B, Lopez B, Salom MG, Valero F, Navarro C, Fenoy FJ. Acuterenal hemodynamic effects of dimanganese decacarbonyl and cobaltprotoporphyrin. Kidney Int. 2004;65:564–574.
Li et al Renal Medullary Heme Oxygenase 153
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from

10. Rodriguez F, Kemp R, Balazy M, Nasjletti A. Effects of exogenous hemeon renal function: role of heme oxygenase and cyclooxygenase. Hyper-tension. 2003;42:680–684.
11. Evans RG, Majid DS, Eppel GA. Mechanisms mediating pressure natri-uresis: what we know and what we need to find out. Clin Exp PharmacolPhysiol. 2005;32:400–409.
12. Mattson DL. Importance of the renal medullary circulation in the controlof sodium excretion and blood pressure. Am J Physiol Regul Integr CompPhysiol. 2003;284:R13–R27.
13. Ryter S, Kvam E, Richman L, Hartmann F, Tyrrell RM. A chromato-graphic assay for heme oxygenase activity in cultured human cells:application to artificial heme oxygenase overexpression. Free Rad BiolMed. 1998;24:959–971.
14. Ryter S, Kvam E, Tyrrell RM. Heme oxygenase activity determination byhigh-performance liquid chromatography. Methods Enzymol. 1999;300:322–336.
15. Dos Santos EA, Dahly-Vernon AJ, Hoagland KM, Roman RJ. Inhibitionof the formation of EETs and 20-HETE with 1-aminobenzotriazoleattenuates pressure natriuresis. Am J Physiol Regul Integr Comp Physiol.2004;287:R58–R68.
16. Zhang AY, Chen Y-F, Zhang DX, Yi F-X, Qi J, Andrade-Gordon P, deGaravilla L, Li P-L, Zou A-P. Urotensin II is a nitric oxide-dependentvasodilator and natriuretic peptide in the rat kidney. Am J Physiol RenalPhysiol. 2003;285:F792–F798.
17. Lorenz JN, Gruenstein E. A simple, nonradioactive method for evaluatingsingle-nephron filtration rate using FITC-inulin. Am J Physiol RenalPhysiol. 1999;276:F172–F177.
18. Beutler E, West C. Simplified determination of carboxyhemoglobin. ClinChem. 1984;30:871–874.
19. Chalmers A. Simple, sensitive measurement of carbon monoxide inplasma. Clin Chem. 1991;37:1442–1445.
20. Marks GS, Vreman HJ, McLaughlin BE, Brien JF, Nakatsu K. Mea-surement of endogenous carbon monoxide formation in biologicalsystems. Antioxid Redox Signal. 2002;4:271–277.
21. Stonek F, Dietrich W, Schneeberger C, Vycudilik W, Tschugguel W.Quantitative determination of carbon monoxide in cell culture super-natants by spectrophotometric analysis. J Biochem Biophys Methods.2004;58:49–58.
22. Li N, Zhang G, Yi F-X, Zou A-P, Li P-L. Activation of NAD(P)H oxidaseby outward movements of H� ions in renal medullary thick ascendinglimb of Henle. Am J Physiol Renal Physiol. 2005;289:F1048–F1056.
23. Zou A-P, Cowley AW Jr. Nitric oxide in renal cortex and medulla: an invivo microdialysis study. Hypertension. 1997;29:194–198.
24. Vreman HJ, Wong RJ, Stevenson DK, Engel RR, Shi Y, Li H, Shen C.Role of carbon monoxide and nitric oxide in newborn infants withpostasphyxial hypoxic–ischemic encephalopathy. Pediatrics. 2002;109:715–716.
25. Bergstrom G, Evans RG. Mechanisms underlying the antihyperten-sive functions of the renal medulla. Acta Physiol Scand. 2004;181:475– 486.
26. Wang T, Sterling H, Shao WA, Yan Q, Bailey MA, Giebisch G, WangW-H. Inhibition of heme oxygenase decreases sodium and fluidabsorption in the loop of Henle. Am J Physiol Renal Physiol. 2003;285:F484–F490.
27. Botros FT, Laniado-Schwartzman M, Abraham NG. Regulation of cy-clooxygenase- and cytochrome P450-derived eicosanoids by heme oxy-genase in the rat kidney. Hypertension. 2002;39:639–644.
28. Li Volti G, Seta F, Schwartzman ML, Nasjletti A, Abraham NG. Hemeoxygenase attenuates angiotensin II-mediated increase in cyclooxygen-ase-2 activity in human femoral endothelial cells. Hypertension. 2003;41:715–719.
29. Quan S, Yang L, Shnouda S, Schwartzman ML, Nasjletti A, GoodmanAI, Abraham NG. Expression of human heme oxygenase-1 in the thickascending limb attenuates angiotensin II-mediated increase in oxidativeinjury 1. 2004;65:1628–1639.
30. Szentivanyi M Jr, Zou A-P, Mattson DL, Soares P, Moreno C, Roman RJ,Cowley AW Jr. Renal medullary nitric oxide deficit of Dahl S ratsenhances hypertensive actions of angiotensin II. Am J Physiol RegulIntegr Comp Physiol. 2002;283:R266–R272.
31. Balcioglu A, Watkins CJ, Maher TJ. Use of a hemoglobin-trappingapproach in the determination of nitric oxide in in vitro and in vivosystems. Neurochem Res. 1998;23:815–820.
32. Silacci P, Desgeorges A, Mazzolai L, Chambaz C, Hayoz D. Flowpulsatility is a critical determinant of oxidative stress in endothelial cells.Hypertension. 2001;38:1162–1166.
33. Wagner CT, Durante W, Christodoulides N, Hellums JD, Schafer AI.Hemodynamic forces induce the expression of heme oxygenase incultured vascular smooth muscle cells. J Clin Invest. 1997;100:589 –596.
34. Vreman HJ, Wong RJ, Kadotani T, Stevenson DK. Determination ofcarbon monoxide (CO) in rodent tissue: effect of heme administration andenvironmental CO exposure. Anal Biochem. 2005;341:280–289.
35. Botros FT, Navar LG. Interaction between endogenously-producedcarbon monoxide and nitric oxide in regulation of renal afferent arterioles.Am J Physiol Heart Circ Physiol. In press.
36. Thorup C, Jones CL, Gross SS, Moore LC, Goligorsky MS. Carbonmonoxide induces vasodilation and nitric oxide release but suppressesendothelial NOS. Am J Physiol Renal Physiol. 1999;277:F882–F889.
37. Xue L, Farrugia G, Miller SM, Ferris CD, Snyder SH, Szurszewski JH.Carbon monoxide and nitric oxide as coneurotransmitters in the entericnervous system: Evidence from genomic deletion of biosyntheticenzymes. Proc Natl Acad Sci U S A. 2000;97:1851–1855.
38. Haider A, Olszanecki R, Gryglewski R, Schwartzman ML, Llanos E,Kappas A, Nasjletti A, Abraham NG. Regulation of cyclooxygenase bythe heme-heme oxygenase system in microvessel endothelial cells.J Pharmacol Exp Ther. 2002;300:188–194.
39. Majid DSA, Said KE, Omoro SA. Responses to acute changes in arterialpressure on renal medullary nitric oxide activity in dogs. Hypertension.1999;34:832–836.
40. Roman RJ. P-450 metabolites of arachidonic acid in the control ofcardiovascular function. Physiol Rev. 2002;82:131–185.
41. Imig JD. 20-HETE or EETs: which arachidonic acid metabolite regulatesproximal tubule transporters and contributes to pressure natriuresis? Am JPhysiol Regul Integr Comp Physiol. 2004;287:R3–R5.
42. Hall JE. The kidney, hypertension, and obesity. Hypertension. 2003;41:625–633.
43. Yang ZZ, Zou AP. Transcriptional regulation of heme oxygenases byHIF-1alpha in renal medullary interstitial cells. Am J Physiol RenalPhysiol. 2001;281:F900–F908.
44. Zou A-P, Yang Z-Z, Li P-L, Cowley AW Jr. Oxygen-dependentexpression of hypoxia-inducible factor-1�alpha in renal medullary cellsof rats. Physiol Genomics. 2001;6:159–168.
45. Liang M, Yuan B, Rute E, Greene AS, Zou A-P, Soares P, MCQuestionGD, Slocum GR, Jacob HJ, Cowley AW Jr. Renal medullary genes insalt-sensitive hypertension: a chromosomal substitution and cDNAmicroarray study. Physiol Genomics. 2002;8:139–149.
154 Hypertension January 2007
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from

Ningjun Li, Fan Yi, Elisabete A. dos Santos, Dustin K. Donley and Pin-Lan LiArterial Blood Pressure
Role of Renal Medullary Heme Oxygenase in the Regulation of Pressure Natriuresis and
Print ISSN: 0194-911X. Online ISSN: 1524-4563 Copyright © 2006 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Hypertension doi: 10.1161/01.HYP.0000250086.06137.fb
2007;49:148-154; originally published online October 30, 2006;Hypertension.
http://hyper.ahajournals.org/content/49/1/148World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://hyper.ahajournals.org/content/suppl/2008/12/19/01.HYP.0000250086.06137.fb.DC1Data Supplement (unedited) at:
http://hyper.ahajournals.org//subscriptions/
is online at: Hypertension Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialHypertensionin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 12, 2018http://hyper.ahajournals.org/
Dow
nloaded from

1
ONLINE SUPPLEMENT
ROLE OF RENAL MEDULLARY HEME OXYGENASE IN THE REGULATION OF
PRESSURE NATRIURESIS AND ARTERIAL BLOOD PRESSURE
Ningjun Li1, Fan Yi1, Elisabete A. dos Santos2, Dustin K. Donley1, Pin-Lan Li1
HPLC analysis of HO activity. Renal tissue HO activities were determined by HPLC analysis
as described previously 1, 2. Briefly, renal cortical, outer and inner medullary (OM and IM)
tissues were homogenized with a glass homogenizer in ice-cold HEPES buffer containing (in
mmol/L) 25 Na-HEPES, 1 EDTA, 255 sucrose, and 0.1 phenylmethylsulfonyl fluoride, and then
centrifuged at 6,000g for 30 minutes at 4°C. The HO reaction was performed in a reaction buffer
containing (in mmol/L): 500 µg fresh homogenate protein suspension, 1 β-NADPH, 2 glucose 6-
phosphate, 1 U glucose-6-phosphate dehydrogenase, 0.25 hemin, 250 sucrose, 20 Tris (pH 7.4)
in a final volume of 100 µl. The reaction mixture was incubated at 37ºC for 60 min and the
reaction was then terminated by addition of 100 µl stop solution (95:5 ethanol:DMSO with 0.4
µmol/L mesoporphyrin). The sample was centrifuged at 15,000 g at room temperature and the
supernatant (50 µl) was transferred into auto sampler vials, and then injected and
chromatographed on a 1090 Series II Liquid Chromatograph (Hewlett-Packard Co., Palo Alto,
CA). All procedures were carried out under dark condition. Metabolites were separated by using
a Waters Novapak reverse phase C18 steel cartridge column with a linear gradient from 100%
buffer A to 100% buffer B over 20 min. Buffer A contains 100 mmol/L ammonium acetate and
70% methanol (v/v), pH 5.2; and buffer B contains 100% methanol. The flow rate was 1.5
ml/min. The visible detector was set for continuous absorbance monitoring at 405 nm. The
concentrations of BR were calculated based on their integrated peak areas from the
chromatograms of each sample and standard curve. HO activity is presented as production rate of
BR at pmol/mg protein/hr.

2
Effect of renal medullary infusion of HO inhibitor on pressure-natriuresis. Rats were
anesthetized with ketamine (30mg/kg body wt i.m.) and Inactin (50 mg/kg body wt i.p.) and
placed on a thermostatically controlled warming table to maintain body temperature at 37°C and
prepared for the study of pressure-natriuresis as previously described 3. Catheters were inserted
in the right femoral vein for i.v. infusions and in the femoral artery for monitoring renal
perfusion pressure (RPP). An abdominal incision was made, the left kidney was placed in a
stainless steel cup to stabilize the organ, and an interstitial catheter (5 mm in depth) was placed at
the renal medulla and anchored in the kidney surface with Vetbond Tissue Adhesive (3M).
Continuous infusion of CrMP (10 nmol/kg/min) or vehicle at a rate of 0.6ml/h (10 µl/min) was
maintained throughout the experiment. Precise placement of the catheter was confirmed at the
end of the experiment. Left ureter was isolated and catheterized for collection of urine during
experiments. After surgery, the animals received a continuous infusion of 0.9% NaCl solution
containing 2% albumin at a rate of 100 µl/min throughout the experiment. For renal blood flow
(RBF) measurement, a transonic flow probe (2 mm) was placed around the left renal artery to
measure RBF with a flowmeter (Transonic) as described previously 4. For pressure natriuresis,
vasopressin (52 pg/min), aldosterone (20 ng/min), norepinephrine (100 ng/min), and
hydrocortisol (20 µg/min) were included in the intravenous infusion solution to fix the
circulating levels of these hormones as previously described 5. An adjustable clamp was placed
on the aorta above renal arteries so that RPP could be controlled. After a 1-h equilibration period,
control MAP and RBF were recorded and urine samples were collected for 30 min. Then, RPP
was acutely increased to 160 mmHg by tying off the celiac and mesenteric arteries. After a 10-
min equilibration period, urine samples were collected during a 30-min clearance period. To
measure GFR, a 0.5-ml bolus of FITC-inulin (8.0 mg/ml) was given, and a steady intravenous
infusion of FITC-inulin (4.0 mg/ml) in the infusion solution continued throughout the
experiment 4, 6, 7. At each RPP levels, blood samples (100µl) were also collected in heparinized
tubes. At the end of each experiment, blood samples were centrifuged, and 20 µl of plasma and
1:50 diluted urine samples were pipetted into a microtiter plate and mixed with 200 µl HEPES
buffer (10 mM) for FITC-inulin fluorescence measurement with excitation and emission
wavelengths of 480 and 530, respectively, using an automatic microplate reader (KC4; Bio-Tek
Instruments, Winooski, VT). The urine flow rate was determined gravimetrically, and sodium
(Na+) concentrations of urine samples were measured using a flame photometer (Buck Scientific,

3
East Norwalk, CT). GFR was calculated as the product of urine flow and the ratio of urine-to-
plasma FITC-inulin fluorescence intensity. GFR, urine flow, and urinary Na+ excretion were
factored per gram kidney weight (kw). At the end of experiment, kidneys were removed,
weighed, dissected into cortical and medullary tissues, and rapidly frozen in liquid nitrogen for
later measurement of HO activity as described above. The animals were then euthanized with an
excess intravenous dose of pentobarbital sodium (150 mg/kg).
Assay of renal medullary CO concentrations. An in vivo microdialysis-oxyhemoglobin CO
trapping assay was established to determine renal medullary CO concentrations.
Spectrophotometric measurement of carboxyhemoglobin (CO-Hb) has been extensively used to
measure the endogenous CO formation 8-12. CO reacts with ferrous oxyhemoglobin (OxyHb) to
form CO-Hb. Sodium hydrosulfite (sodium dithionite) reduces OxyHb and methemoglobin but
not CO-Hb, thereby forming a mixture of two pigments of hemoglobin, i.e. reduced Hb and
COHb.
Absorbencies of these two forms of Hb are 420 nm for CO-Hb and 432 nm for reduced Hb.
A ratio of absorbance at 420 and 432 nm represents the CO-Hb level. CO concentration was
calculated using the absorbance at 420 and 432 nm and the parameters obtained from 100%
reduced Hb and 100% CO-Hb (pure CO gas-saturated Hb) 8, 9, 12.
For in vivo microdialysis, the animals were prepared as described above for the studies of
pressure natriuresis. In vivo microdialysis experiments were performed as described in our
previous studies 13, 14. In brief, a microdialysis probe (Bioanalytical Systems, West Lafayette, IN)
with a 0.5-mm tip diameter, 2-mm dialysis length, and 20-kDa transmembrane diffusion cutoff
was gently implanted into the outer medulla (5–5.5 mm in depth) vertically from the dorsal
surface. The dialysis probe was perfused with OxyHb (0.5mg/ml, Sigma-Aldrich) diluted in PBS
containing (in mmol/L) 205 NaCl, 40.5 Na2HPO4, and 9.5 NaH2PO4
(pH 7.4, 550 mOsm/L) at a rate of 2.0 µl/min throughout the experiment. This microdialysis
probe was also constructed with an incorporated infusion line, which was used for renal
medullary interstitial infusion. At each RPP level, a 30-min dialysate sample was collected. At
the end of experiment, 50 µl of dialysate samples were mixed with 50 µl of sodium dithionite (5

4
mg/ml, Sigma-Aldrich). The absorbance at 420 and 432 nm was measured with a Cary 100
spectrophotometer using microcells (Varian, Palo Alto, CA). The CO-Hb and consequent CO
concentration were calculated using the formulas reported previously 8, 12, 15. A calibration curve
was constructed in vitro with the use of standard solutions of CO-saturated solution (817 µmol/L 15) . The dialysis efficiency was determined in vitro using serial diluted CO-saturated solutions.
Effects of chronic renal medullary infusion of HO inhibitor on arterial blood pressure. The
rats were first uninephrectomized to remove the right kidney after anesthetized with an
intramuscular injection of ketamine (60 mg/kg) and xylazine (5 mg/kg). One week after
uninephrectomy, surgeries were performed to implant arterial and renal medullary interstitial
catheters described in detail previously 16. Briefly, for chronically monitoring mean arterial blood
pressure (MAP), a catheter connected to a telemetry transmitter was inserted into femoral artery.
The transmitter was placed subcutaneously, and blood pressure signals was recorded through a
remote receiver, which allowed rats to be housed unrestrainedly as usual. To implant the renal
medullary infusion catheter, the left kidney was exposed by a flank incision (1-1.5 cm), and a
medullary interstitial catheter (tapped tip) was implanted into the kidney. The catheter was made
with a number of circular “pig-tail” bends, which prevented the catheter from being pulled out of
the kidney during normal movement of the animal. The catheter was anchored into place on the
kidney surface with cyanoacrylate adhesive and a small piece of abdominal fat. These catheters
were tunneled to the back of neck and passed through a flexible spring that was attached to the
rat with a skin button. This spring was attached to a swivel above the cage and allowed the
animal full range of movement in the home cage but protected the catheters. After surgery, the
catheters were continuously infused with saline (8 µl/min) to maintain the potency of the
catheter.
After 7 days of recovery from surgery, mean arterial pressure (MAP) were recorded for 3
hours/day from 09:00 AM and 12:00 PM. The rats were fed a normal salt diet (1% NaCl) and
had free access to food and water throughout the study. After baseline MAP was recorded on 2
consecutive control days while the rats remained on the 1% salt diet, and the animals were then
divided into 2 groups and received renal medullary infusion of either saline (n=7) or chromium
mesoporphyrin (CrMP 10 µmol/kg/day, n=14), heme oxygenase inhibitor. Three days after

5
starting infusion of CrMP, animals were switched to high salt diet (8% NaCl) except that half of
the CrMP-treated rats were kept on normal salt diet, and MAP was recorded for additional 10
days. There were three groups in total, including Vehicle + high salt (HS), CrMP + HS and
CrMP + normal salt (NS). At the end of experiment, kidneys were removed and rapidly
dissected into the renal cortex, OM and IM and then frozen in liquid N2, and the precise location
of interstitial infusion catheter was determined when dissecting kidney tissue.
Reference
1. Ryter S, Kvam E, Richman L, Hartmann F, Tyrrell RM. A Chromatographic Assay for
Heme Oxygenase Activity in Cultured Human Cells: Application to Artificial Heme
Oxygenase Overexpression. Free Radical Biology and Medicine. 1998; 24:959-971.
2. Ryter S, Kvam E, Tyrrell RM. Heme oxygenase activity determination by high-performance
liquid chromatography. Methods Enzymol. 1999; 300:322-336.
3. Zou A-P, Billington H, Su N, Cowley AW, Jr. Expression and Actions of Heme Oxygenase
in the Renal Medulla of Rats. Hypertension. 2000; 35:342-347.
4. Zhang AY, Chen Y-F, Zhang DX, Yi F-X, Qi J, Andrade-Gordon P, de Garavilla L, Li P-L,
Zou A-P. Urotensin II is a nitric oxide-dependent vasodilator and natriuretic peptide in the
rat kidney. Am J Physiol Renal Physiol. 2003; 285:F792-798.
5. Dos Santos EA, Dahly-Vernon AJ, Hoagland KM, Roman RJ. Inhibition of the formation of
EETs and 20-HETE with 1-aminobenzotriazole attenuates pressure natriuresis. Am J Physiol
Regul Integr Comp Physiol. 2004; 287:R58-68.
6. Lorenz JN, Gruenstein E. A simple, nonradioactive method for evaluating single-nephron
filtration rate using FITC-inulin. Am J Physiol Renal Physiol. 1999; 276:F172-177.
7. Wang W, Poole B, Mitra A, Falk S, Fantuzzi G, Lucia S, Schrier R. Role of Leptin
Deficiency in Early Acute Renal Failure during Endotoxemia in ob/ob Mice. J Am Soc
Nephrol. 2004; 15:645-649.
8. Beutler E, West C. Simplified determination of carboxyhemoglobin. Clin Chem. 1984;
30:871-874.
9. Arregui B, Lopez B, Salom MG, Valero F, Navarro C, Fenoy FJ. Acute renal hemodynamic
effects of dimanganese decacarbonyl and cobalt protoporphyrin. Kidney International.
2004; 65:564-574.

6
10. Chalmers A. Simple, sensitive measurement of carbon monoxide in plasma. Clin Chem.
1991; 37:1442-1445.
11. Marks GS, Vreman HJ, McLaughlin BE, Brien JF, Nakatsu K. Measurement of Endogenous
Carbon Monoxide Formation in Biological Systems. Antioxidants & Redox Signaling. 2002;
4:271-277.
12. Stonek F, Dietrich W, Schneeberger C, Vycudilik W, Tschugguel W. Quantitative
determination of carbon monoxide in cell culture supernatants by spectrophotometric
analysis. Journal of Biochemical and Biophysical Methods. 2004; 58:49-58.
13. Li N, Zhang G, Yi F-X, Zou A-P, Li P-L. Activation of NAD(P)H oxidase by outward
movements of H+ ions in renal medullary thick ascending limb of Henle. Am J Physiol
Renal Physiol. 2005; 289:F1048-1056.
14. Zou A-P, Cowley AW, Jr. Nitric Oxide in Renal Cortex and Medulla: An In Vivo
Microdialysis Study. Hypertension. 1997; 29:194-198.
15. Vreman HJ, Wong RJ, Stevenson DK, Engel RR, Shi Y, Li H, Shen C. Role of Carbon
Monoxide and Nitric Oxide in Newborn Infants With Postasphyxial Hypoxic-Ischemic
Encephalopathy. Pediatrics. 2002; 109:715-716.
16. Makino A, Skelton MM, Zou A-P, Roman RJ, Cowley AW, Jr. Increased Renal Medullary
Oxidative Stress Produces Hypertension. Hypertension. 2002; 39:667-672.