relaxation of a shear-induced lamellar phase measured with time-resolved small-angle neutron...
TRANSCRIPT

ARTICLE IN PRESS
Physica B 350 (2004) e963–e966
*Corresp
921-9847.
E-mail a
0921-4526/$
doi:10.1016
Relaxation of a shear-induced lamellar phase measured withtime-resolved small-angle neutron scattering
L. Porcara,*, W.A. Hamiltonb, P.D. Butlerb, G.G. Warrc
aNIST Center For Neutron Research, Gaithersburg, MD 20899-8562, USAbCenter for Neutron Scattering, Condensed Matter Sciences Division, Oak Ridge National Laboratory, Oak Ridge, TN 37831-6393, USA
cSchool of Chemistry, University of Sydney, Sydney NSW 2006, Australia
Abstract
We have measured the relaxation of Couette shear-induced La lamellar states to their isotropic L3 ‘‘sponge’’
equilibrium phases in the cetylpryridinium–hexanol/dextrose–brine system by (cycled) time-resolved small-angle
neutron scattering. Although diffusive motions of adjacent membrane sheets may be estimated to bring them into
contact with frequencies B10 kHz, we observe structural relaxation times on the order of seconds. This indicates a
significant activation energy against the re-establishment of the passages characterizing the convoluted sponge
structure.
r 2004 Elsevier B.V. All rights reserved.
PACS: 82.70.Dd; 87.16.Dg; 47.20.Hw; 61.12.Ex
Keywords: SANS; Shear; Relaxation; Sponge phase; Lamellar phase
The isotropic L3 ‘‘sponge’’ phase consists of asingle self-assembled surfactant bilayer spanningsolution volume in a convoluted multi-connectednetwork and separating two equivalent sub-vo-lumes, which interpenetrate through randomlydirected passages [1]. Despite this extended struc-ture, these phases are typically highly fluid andshow little or no response to applied shear, as thepassages can rapidly reorient to relieve stresses onthe membrane and allow free flow of the solvent[2]. When they occur in a chemical system L3
onding author. Tel.:+1-301-975-5049; fax: +1-301-
ddress: [email protected] (L. Porcar).
- see front matter r 2004 Elsevier B.V. All rights reserve
/j.physb.2004.03.305
phases are found in narrow regions of stabilityadjacent to those of the very different La lamellarphases. In the La morphology surfactant mem-branes are stacked in regular smectic order. La
phases are anisotropic and strongly birefringent,and while viscous at low shear rates shear-thin asthe membranes align in the flow field. This latterresponse is easily observed in shear-cell scatteringmeasurements.Dilute sponge phases do exhibit one interesting
dynamical response—transient birefringence upongentle stirring. This immediately suggests a shear-induced sponge to lamellar transition. Cates andMilner [3], treating shear as quasi-thermodynamicparameter in the free energy for isotropic phases,
d.

ARTICLE IN PRESS
Fig. 1. Cycled time-resolved SANS data acquisition.
L. Porcar et al. / Physica B 350 (2004) e963–e966e964
predicted that shear should induce an L3 to La
transition by suppressing the fluctuations thatstabilize the phase, with critical shear rates varyingas the cube of the membrane volume fraction fand inversely as the solvent viscosity Zs. At thetypical concentrations convenient to structuraland rheological measurement such a transitionoccurs at very high shear rates. However, inmeasurements on hyperswollen C12E5 spongesunder oscillatory shear in thin film cells Yamamo-to and Tanaka [4] were able to observe an opticaldiffraction signal consistent with a transition to La
membrane ordering.Recently, we directly addressed the question of
shear-induced sponge to lamellar transitions andin particular the predicted f3/Zs scaling of thisresponse. Rather than working with very dilutesamples we pursued an alternative strategy ofadding a thickener to the solvent, which if inert(excluded from the membranes) should have littleeffect on phase behavior while slowing itsdynamics. We found that dextrose can play thisrole in the well-known cetylpyridinium (CPCl)–hexanol/brine system. Small-angle scattering(SANS) measurements showed that La structureis maintained to dextrose concentrations of up to40% volume fraction while the viscosity of thesolvent by increases by over an order of magnitude[5].Slowed diffusive motion of the membranes in
these ‘‘sweetened’’ sponges causes strong structur-al and rheological responses to applied shear ateasily accessible shear rates (o2000 s�1). Inrheological measurements an initial Newtonianconstant viscosity response is followed by shear-thinning region, and a final quasi-Newtonianregion. Consistent with the predicted scaling,appropriately normalized viscosities for a rangeof samples (volume fraction f of: 3–10%, Zs: 1.1–16.3� 10�3 Pa s) for which the onset of shear-thinning varied by about two orders of magnitudefell to a master curve plotted against a rescaledshear rate (dg/dt)Zs/f
3. Parallel Couette shearSANS showed that the shear-thinning responsecorresponds to a shear-induced L3 to La transition,with the appearance of strong anisotropy in thescattering patterns and the eventual appearance ofdefinite Bragg peaks in tangential Couette SANS
data (beam along the flow direction) indicating‘‘c’’ lamellar stacking (normals along the sheargradient). In region III this signal disappearsrapidly, presumably indicating a collapse of theshear-induced La phase and strong scatteringemerging at small scattering vectors indicates someas yet unidentified larger scale ordering [6].The shear-induced La signal saturates indicating
full alignment of the membranes over about afactor of two in (dg/dt)Zs/f
3 and is easily accessiblefor lower f and higher Zs (dextrose) samples. Wehave recently been successful in measuring therelaxation from this ‘‘passage free’’ La state to theequilibrium L3 phase upon cessation of shear. Thisre-establishment of the L3 structure disrupted byshear offers a strong clear signal of the fusion ofmembranes to (re)form passages. A process whichusually occurs in dynamic equilibrium (creation/destruction) against a background of other phe-nomena.Even with the slowed dynamics of our system
relaxation takes only a few seconds. These shorttimes (for SANS) required that we time multiplexthe NIST NG3 2D SANS detector output intoseveral memory modules, rather than measuresequential runs. To achieve adequate statistics wecycled through the process of shearing to a steadyLa state, stopping, and relaxing fully to L3 severalthousand times (see Fig. 1). Each data acquisitioncycle was synchronized at a delay to a stop signalto our ORNL Couette shear cell [7,8]. Repeatedcycling did not degrade these samples and datawas completely reproducible. Finally, in order toobtain sufficient points on the relaxation curve wehardware rebinned the detector output from128� 128 to a 32� 32 array by dropping the
ARTICLE IN PRESS
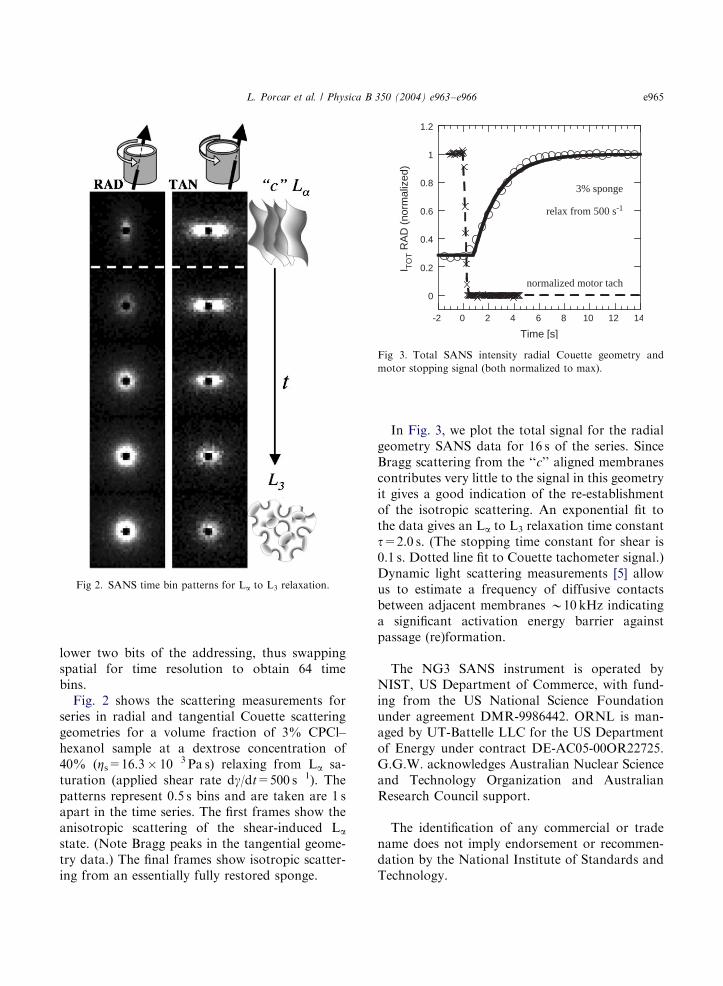
Fig 2. SANS time bin patterns for La to L3 relaxation.
0
0.2
0.4
0.6
0.8
1
1.2
-2 0 2 4 6 8 10 12 14
I TO
T R
AD
(no
rmal
ized
)
Time [s]
3% sponge
relax from 500 s-1
normalized motor tach
Fig 3. Total SANS intensity radial Couette geometry and
motor stopping signal (both normalized to max).
L. Porcar et al. / Physica B 350 (2004) e963–e966 e965
lower two bits of the addressing, thus swappingspatial for time resolution to obtain 64 timebins.Fig. 2 shows the scattering measurements for
series in radial and tangential Couette scatteringgeometries for a volume fraction of 3% CPCl–hexanol sample at a dextrose concentration of40% (Zs=16.3� 10�3 Pa s) relaxing from La sa-turation (applied shear rate dg/dt=500 s�1). Thepatterns represent 0.5 s bins and are taken are 1 sapart in the time series. The first frames show theanisotropic scattering of the shear-induced La
state. (Note Bragg peaks in the tangential geome-try data.) The final frames show isotropic scatter-ing from an essentially fully restored sponge.
In Fig. 3, we plot the total signal for the radialgeometry SANS data for 16 s of the series. SinceBragg scattering from the ‘‘c’’ aligned membranescontributes very little to the signal in this geometryit gives a good indication of the re-establishmentof the isotropic scattering. An exponential fit tothe data gives an La to L3 relaxation time constantt=2.0 s. (The stopping time constant for shear is0.1 s. Dotted line fit to Couette tachometer signal.)Dynamic light scattering measurements [5] allowus to estimate a frequency of diffusive contactsbetween adjacent membranes B10 kHz indicatinga significant activation energy barrier againstpassage (re)formation.
The NG3 SANS instrument is operated byNIST, US Department of Commerce, with fund-ing from the US National Science Foundationunder agreement DMR-9986442. ORNL is man-aged by UT-Battelle LLC for the US Departmentof Energy under contract DE-AC05-00OR22725.G.G.W. acknowledges Australian Nuclear Scienceand Technology Organization and AustralianResearch Council support.
The identification of any commercial or tradename does not imply endorsement or recommen-dation by the National Institute of Standards andTechnology.

ARTICLE IN PRESS
L. Porcar et al. / Physica B 350 (2004) e963–e966e966
References
[1] G. Porte, Curr. Opin. Colloid Interface Sci. 1 (1998) 345
and references therein.
[2] P. Snabre, G. Porte, Europhys. Lett. 13 (1990) 641.
[3] M.E. Cates, S.T. Milner, Phys. Rev. Lett. 62 (1989) 1856.
[4] J. Yamamoto, H. Tanaka, Phys. Rev. Lett. 77 (1996)
4390.
[5] L. Porcar, W.A. Hamilton, P.D. Butler, G.G. Warr,
Langmuir 19 (2003) 10779.
[6] L. Porcar, W.A. Hamilton, P.D. Butler, G.G. Warr, Phys.
Rev. Lett. 89 (2003) 168301.
[7] L. Porcar, W.A. Hamilton, P.D. Butler, G.G. Warr, Rev.
Sci. Instrum. 73 (2002) 2345.
[8] W.A. Hamilton, P.D. Butler, L.J. Magid, Z. Han, T.M.
Slawecki, Phys. Rev. E 60 (1999) R1146.