relationships between bacteria and grain surfaces in intertidal sediments
TRANSCRIPT

Relationships Between Bacteria and Grain Surfaces in Intertidal SedimentsAuthor(s): Mary F. DeFlaun and Lawrence M. MayerSource: Limnology and Oceanography, Vol. 28, No. 5 (Sep., 1983), pp. 873-881Published by: American Society of Limnology and OceanographyStable URL: http://www.jstor.org/stable/2836366 .
Accessed: 14/06/2014 13:22
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
American Society of Limnology and Oceanography is collaborating with JSTOR to digitize, preserve andextend access to Limnology and Oceanography.
http://www.jstor.org
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

Limnol. Oceanogr., 28(5), 1983, 873-881 ? 1983, by the American Society of Limnology and Oceanography, Inc.
Relationships between bacteria and grain surfaces in intertidal sediments' Mary F. DeFlaun2 and Lawrence M. Mayer3 Oceanography Program, Ira C. Darling Center, University of Maine at Orono, Walpole 04573
Abstract Seasonal changes in total bacterial numbers and their associated mucus coatings in surficial
sediments were examined. Bacterial numbers followed the temperature cycle, with highest numbers in summer. The specific surface areas of the sediments were measured rather than inferred from other granulometric properties; bacterial numbers were proportional to surface areas only for sample suites collected at the same time. Bacteria inhabited shallow depres- sions on sand and silt grains; they were not found on grains smaller than about 10 gm or inside smaller pores like those on weathered feldspar grains. Mucus coatings also followed a seasonal cycle, increasing in abundance and coalescence from spring into summer. These coatings accumulated clay grains, suggesting that the relationship of bacteria to surface area may be due to bacterial control of surface area rather than the reverse. Organic carbon con- centrations in grain size separates of these sediments increased with decreasing size until the fine silt fraction, and decreased in the clay fraction; it is not clear, however, whether this trend is a result or a cause of bacterial colonization patterns.
Particle surfaces are important habitats for sedimentary bacteria. Adsorption of nutrients to solid surfaces and an ac- companying increase in bacterial bio- mass was demonstrated in the early lab- oratory work of ZoBell (1943). Subsequent measurements of bacterial numbers or metabolic activities in sediments have exhibited a strong inverse correlation with sediment grain size, implying a surface area dependence (Dale 1974; Hargrave 1972). However, this surface area depen- dence of bacteria has not yet been veri- fied with direct measures of surface area. The difficulties in relating measured sur- face areas to other granulometric param- eters have been pointed out by Mayer and Rossi (1982).
Microtopography of sediment grains has also been implicated as a factor influenc- ing bacterial colonization on sediment grains. In an optical microscope study of sand grains, Meadows and Anderson (1966) found bacteria in patches with large
I This work was supported by NSF ISP 8011448 and NOAA Sea Grant R/LRF-45. Contribution 83- 02 of the Maine Benthic Oceanography Group.
2 Present address: Department of Marine Science, University of South Florida, St. Petersburg 33701.
3To whom reprint requests should be directed.
areas of bare suface between them; these patches were often found in hollows and cracks. Weise and Rheinheimer (1978) found the density of bacterial cells to be determined by microtopography, with the highest densities on subrounded or sub- angular grains. Frankel (1977) found that subangular to subrounded grains of horn- blende supported fewer bacteria than rel- atively smooth biotite grains. The sepa- rated folia of the biotite grains provided a more protected habitat and consequent- ly supported a larger population. The de- gree of protection seems to influence the population size of colonizing microbiota. Surface morphology affected both the to- tal biomass and the community structure of the microbiota (including bacteria, al- gae, and grazers) on silica grains in a run- ning seawater experiment (Nickels et al. 1981). Total microbial biomass was higher on grains with more surface irregulari- ties. Smooth grains of the same size sup- ported a lower population of procaryotes and microalgae, with an increase of mi- croeucaryotic grazers.
Extracellular exudates of bacteria have become subjects of increasing interest due to their potential roles in affecting the sedimentological and trophic status of sediments. There is, however, a limited literature describing mucus coatings on
873
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

874 DeFlaun and Mayer
natural marine sediments (Frankel and Mead 1973; Weise and Rheinheimer 1978).
We describe here the seasonal changes in total bacterial numbers and their as- sociated extracellular mucus in a temper- ate intertidal sediment. These descrip- tions are compared with actual surface area measurements of the sediments. Lastly, we describe the preferential col- onization of bacteria on grains of differ- ent size and surface porosity and com- pare these colonization patterns to the organic matter content of grains of differ- ent sizes.
We thank V. Westcott for the SEM used in this study. Valuable advice and com- ments were contributed by L. Watling, J. Sieburth, G. Taghon, and D. Pratt. H. Holman and C. Brower provided techni- cal assistance and P. McCall reviewed the manuscript.
Methods Samples of the top 4-5 mm of sediment
were taken from February to October 1981 at low tide at three stations in Lowes Cove, an intertidal mudflat on the Dam- ariscotta River estuary in Maine. The samples were divided into four parts: two for SEM and epifluorescence examina- tion were fixed in a glutaraldehyde-buff- er mixture adjusted to the osmolality of seawater with sucrose, a third was dried to obtain a wet-to-dry-weight ratio, and the fourth was frozen for later surface-area analysis.
The SEM samples were postfixed in 1% osmium tetroxide with sodium cacodyl- ate buffer after the unbound glutaralde- hyde had been removed by a series of washes. The sediments were then dehy- drated, dried in a critical-point dryer with liquid CO2, and sputter coated with gold- palladium before observation in a Coates and Welter Quikscan scanning electron microscope.
DAPI (4'-6-diamidino-2-phenylindole dihydrochloride) has been used by mi- crobiologists as a very specific DNA binding fluorochrome (Coleman 1980). Its use in staining for direct counts of bac- teria has been explored and it has been
shown to be a much more specific stain than acridine orange, with greater fluo- rescence intensity and a much slower rate of fading. In view of the advantages of DAPI for staining bacteria in direct counting procedures, we modified the method of Porter and Feig (1980) for use with marine sediments.
DAPI is photodegradable; therefore all samples were processed in a lightless hood. The DAPI (Polysciences) was di- luted to 0.05 mg* ml-' of stock solution with 0.2-,um-filtered Nanopure (Sybron-Barn- stead) water and stored in 50-,ul portions at - 10?C. Glassware and plasticware were prewashed with 1:1 H2O:HNO3 and rinsed five times with Nanopure water; all reagents were filtered before use through a Whatman GFF underlain by a 0.2-,um Nuclepore filter.
To prepare slides, we shook the sedi- ment sample thoroughly in the fixative and pipetted 1.0 ml into a 50-ml beaker containing 35 ml of 0.2-,um-filtered sea- water. The sample was then sonicated with a Fisher sonic dismembranator with a 25-mm titanium tip at 75 W for 1 min. Sonication breaks up clumps of algae and sediment which could prevent a thin, even spread of the material over the filter and thus affect counting accuracy, but does not destroy the bacteria.
While the beaker was being stirred, 5.0 ml of the sonicated sample were pipetted into a 15-ml conical-bottom centrifuge tube. One 50-,l aliquot of DAPI was added to the tube, the tube was spun on a Vortex mixer, and then placed in com- plete darkness for at least 5 min. The final concentration of DAPI in solution was 0.5 ,ug ml-,.
Two milliliters of sample were filtered through a 0.2-,um Irgalan black-stained Nuclepore filter. Two filters were pre- pared from each sample so that replicates could be counted. The slides were stored in the dark at 40C and counted within 24 h. Blanks were prepared by staining a se- rial dilution of seawater and fixative with- out sediment and counted for each set of slides.
Two microscopes were used for direct counts. Most bacterial preparations were
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

Bacteria in intertidal sediments 875
examined under a Nikon Fluophot Series V equipped with an epifluorescent illu- mination system including a mercury lamp and a x 100 Planachromat objective. The filter combination consisted of violet light excitation (385-425 nm) with a 445- nm dichroic mirror and 470K barrier fil- ter. The remaining preparations were ex- amined with a Leitz Laborlux 12 equipped with the Leitz Ploempak epi- fluorescence unit, with a mercury lamp (50 W) and a x100 EF objective. An "A" filter cube for ultraviolet light excitation was found most suitable for DAPI counts (340-380 excitation filter, 400-nm di- chroic mirror, and 430 barrier filter). Ten fields or 200 cells were counted, spaced across the width of the slide as delineat- ed by an eyepiece micrometer (total di- mensions 70 x 70 gim). The mean num- ber of bacteria per field was then converted to number of bacteria per gram dry weight of sediment.
Specific surface area analysis was per- formed on sediments which had their or- ganic coatings removed with boiling hy- drogen peroxide. Directly measured surface area values by nitrogen sorption were obtained with a Quantachrome Monosorb.
One sample of Lowes Cove surface sediment was separated into its compo- nent size fractions by sieving and gravity settling (Folk 1974; Krumbein and Pet- tijohn 1938). The individual fractions were analyzed for organic carbon, after acidification to remove carbonates, in a Hewlett-Packard 185B CHN analyzer.
The colonization patterns of bacteria on sediment were examined in culture by incubating intertidal sediment, pretreat- ed with hydrogen peroxide, in 0.1 mg ml-, peptone solution in seawater at room temperature for several days. These sed- iments were then prepared for SEM ex- amination as above.
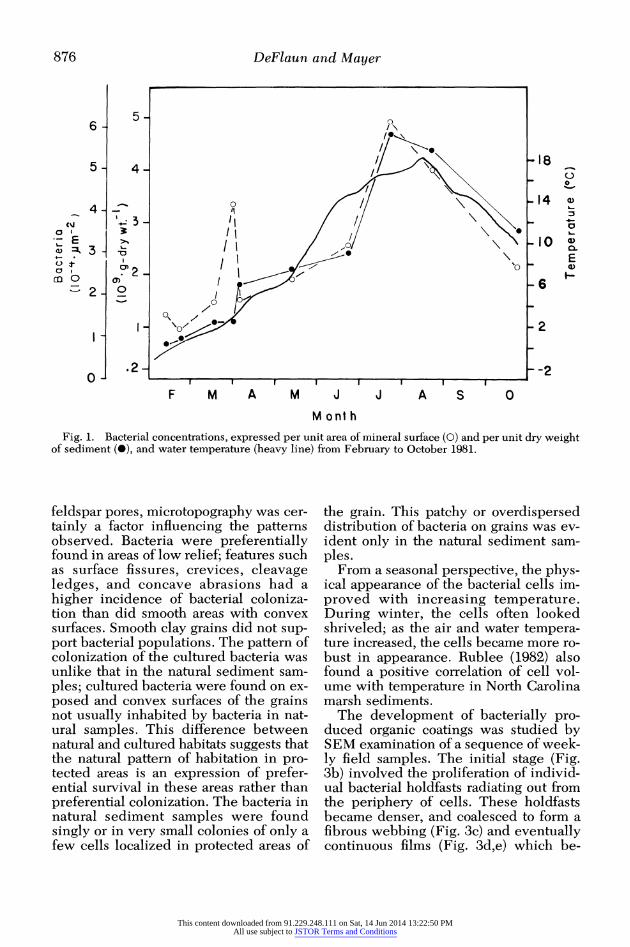
Results Figure 1 shows the concentrations of
bacterial cells in the sediments during the study period, along with water tempera- tures in the adjacent estuary. The preci- sion of these counts ranged from +10%
for high bacterial numbers to ?25% for low bacterial numbers. There was a grad- ual increase from February, when the ice began to break up in the cove, to July. The August sample marks the beginning of a decline in bacterial numbers, which is associated with a decline in water tem- perature. The most rapid increase oc- curred from June (2.36 x 109 cells g-I dry wt) to July (4.68 x 109 cells g-I dry wt). This period immediately follows the most abrupt increase in air and water temper- atures. The same trend was observed with bacterial numbers normalized to in situ sediment volume.
Sediment specific surface area (Fig. 2) is lowest during winter, the sample at the end of March having the lowest value. The specific surface area peaked sharply in early April, followed by a gradual de- cline throughout spring and early sum- mer and a rise into fall. These results are consistent with the seasonal trend in sur- face area found in an earlier seasonal sur- vey on the same mudflat (unpubl. data). This sequence follows the pattern of ero- sion and deposition in Lowes Cove ob- served by Anderson et al. (1981b). They found the greatest period of erosion in Lowes Cove to be from 3 February to 9 March, with the maximum erosion occur- ring with the melting of the ice cover. The beginning of April was found to be a period of rapid deposition, with pellet production contributing to a fining of the sediments by stabilizing the clay and silt fractions (Anderson et al. 1981a).
Scanning electron microscopy showed that the characteristic deep pores of weathered feldspar (Fig. 3a), which we originally thought would provide ideal bacterial niches, did not harbor bacteria, but instead were usually filled with small clay particles. These pores were large enough for bacteria, but were apparently not suitable habitats despite their ob- vious advantages of providing protection against grazers and physical abrasion. The etched pits of weathered feldspars may be too narrow and deep to allow a suffi- cient flow of nutrient-containing seawa- ter to supply bacterial needs. Although there was no preferential colonization of
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions
876 DeFlaun and Mayer
6 \
18 0
0 1 4~~~~~~~~~~~~~~~~~~
0'~~~~~~~~~~~~~~~~~~~~~~~~~~~ E~~~~~~~~~~~~~~~~~~~ =;
e :3
o~~~~~~~~~~~~~~~~~ 2 2 -
F M A M J J A S 0 M ont h
Fig. 1. Bacterial concentrations, expressed per unit area of mineral surface (0) and per unit dry weight of sediment (0), and water temperature (heavy line) from February to October 1981.
feldspar pores, microtopography was cer- tainly a factor influencing the patterns observed. Bacteria were preferentially found in areas of low relief; features such as surface fissures, crevices, cleavage ledges, and concave abrasions had a higher incidence of bacterial coloniza- tion than did smooth areas with convex surfaces. Smooth clay grains did not sup- port bacterial populations. The pattern of colonization of the cultured bacteria was unlike that in the natural sediment sam- ples; cultured bacteria were found on ex- posed and convex surfaces of the grains not usually inhabited by bacteria in nat- ural samples. This difference between natural and cultured habitats suggests that the natural pattern of habitation in pro- tected areas is an expression of prefer- ential survival in these areas rather than preferential colonization. The bacteria in natural sediment samples were found singly or in very small colonies of only a few cells localized in protected areas of
the grain. This patchy or overdispersed distribution of bacteria on grains was ev- ident only in the natural sediment sam- ples.
From a seasonal perspective, the phys- ical appearance of the bacterial cells im- proved with increasing temperature. During winter, the cells often looked shriveled; as the air and water tempera- ture increased, the cells became more ro- bust in appearance. Rublee (1982) also found a positive correlation of cell vol- ume with temperature in North Carolina marsh sediments.
The development of bacterially pro- duced organic coatings was studied by SEM examination of a sequence of week- ly field samples. The initial stage (Fig. 3b) involved the proliferation of individ- ual bacterial holdfasts radiating out from the periphery of cells. These holdfasts became denser, and coalesced to form a fibrous webbing (Fig. 3c) and eventually continuous films (Fig. 3d,e) which be-
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

Bacteria in intertidal sediments 877
16
14
-12
to
8
4
T - _ o~~~~~~- F' A J 'A '0
MONTH
Fig. 2. Sediment specific surface area from Feb- ruary to October 1981.
came thicker and more prevalent as sum- mer progressed, often covering a substan- tial portion of the grain. At high magnifications, the films had a "cauli- flower" texture (Fig. 3D. This temporal sequence has been observed before in studies of fouling on artificial substrates (Dempsey 1981a,b). Diatoms also con- tributed to this surficial organic matrix by exuding mucus around the periphery of their frustules and by producing mucus trails from the raphe during travel over grain surfaces.
These bacterial webs, or glycocalyx, and the mucus produced by diatoms tended to collect fine clay particles. Dempsey (1981b) observed a similar phenomenon on antifouling paint sub- strates. In fact, most of the clay particles that we observed in SEM samples were
tied up in bacterial fibrous webs or embedded in the mucus surrounding dia- tom frustules. Bacteria were never found attached to clay particles. Although clay- sized particles contribute significantly to the surface area of the sediment (Mayer and Rossi 1982), these particles are ap- parently too small or too smooth to pro- vide microenvironments for bacterial at- tachment.
Figure 4 shows the concentration of or- ganic carbon in the different size frac- tions of Lowes Cove sediment. Organic carbon increases with decreasing grain size to the fine silt fraction, but falls off slightly in the clay fraction. Similar trends have been observed in soil samples (Tur- chenek and Oades 1979) and deep-sea sediments (Tanoue and Handa 1979). Our higher carbon values in the sand fractions than in the silt fractions are due to our separation technique; sieving the sands caused retention of Spartina fragments in these fractions while the coarser silts sep- arated by gravity consisted primarily of clean quartz and feldspar grains.
Discussion Bacterial numbers in surface intertidal
sediments fluctuated seasonally. A warm- ing trend in both air and water tempera- ture during the study period was reflect- ed in an increase in bacterial numbers. Increases in temperature increase bacte- rial metabolic rates, reproductive rates and consequently the standing stock of cells. Such temperature-dependent growth rates and consequent biomass in- creases have been observed for bacteria in the water by Meyer-Reil (1977) and for intertidal bacteria in culture by Schroder and van Es (1980). Although rates of graz- ing on bacteria by animals would also be expected to rise with increasing temper- ature, apparently the response in bacte- rial reproductive rate is somewhat great- er. The low temperatures in winter would slow metabolism and induce dormancy in the bacterial population, explaining both the low numbers and the lack of colonies and dividing cells in our winter samples. Our seasonal trend is somewhat different than that noted by Rublee (1982), who
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

878 DeFlaun and Mayer
4.~~~~0
4p
Fig. 3. a. Etched pits of naturally- weathered feldspar (scale bar-20 1&). b. Bacterium with radiating holdfasts (scale bar-0.5 u). c. Bacterially produced mucopolysaccharide strands (scale bar-i 1L). d. Hold- fasts coalescing to form fibrous webs (scale bar-0.2 tt). e. Bacterial colony covered by mucopolysaccharide sheath (scale bar-I tL). f. Organic coating on the surface of the sediment grain (scale bar-0.2 u).
found an autumnal maximum in North Carolina salt marsh sediments.
Bacterial populations in sediments are strongly correlated with sediment specif-
ic surface area (e.g. Dale 1974). Plotting the bacterial numbers vs. surface area (Fig. 5) shows that this relationship may indeed be valid for different samples
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

Bacteria in intertidal sediments 879
z 0
crI
0
10 100 PARTICLE SIZE (pjm)
Fig. 4. Organic carbon concentration in differ- ent size separations of surficial sediment.
measured at any one time as indicated by the rough proportionality seen for the data from three stations from any one sam- pling date, but the relationship is not as strong if data from different sampling times are considered together. Either temperature or some parameter related to it markedly affects the ratio of bacterial number to surface area.
Several workers have calculated the percentage of coverage of grain area by bacteria (see Rublee and Domseif 1978), but all estimates to date have been made by using surface areas calculated from grain diameter measurements. Such cal- culated surface areas are generally low by one to two orders of magnitude be- cause of the high porosity of sediment grains (Weiler and Mills 1965; Slabaugh and Stump 1964; Mayer and Rossi 1982). Our measured surface areas allow us to calculate percentage of cover using ac- tual surface areas; these values range from 0.01 to 0.05% in this study. However, our nitrogen sorption technique measures surface areas that are certainly not avail- able to bacterial colonization, such as nanometer-sized pores. Our percentages should, therefore, be regarded as an un- derestimate of the cover of colonizable area, whereas other studies have provid- ed overestimates. Nevertheless, the con-
6-
~5- 3.' ~ ~~A 0
23 ~~~~~~~~~~~~~~~00 0 B
0 a~~~~~~
0 2 4 6 8 10 12 14 Surface area
(im2 g-1 )
Fig. 5. Bacteria concentration vs. sediment spe- cific surface area. Lines connect points representing samples collected on same date. A-July; B-June; C-April.
clusion from these estimates, that bacte- rial cells utilize only a small proportion of available area (Hargrave 1972) and are therefore not area-limited, remains valid.
This conclusion of nonutilization of available area can be further modified by our observations that bacteria do not ap- pear to colonize clay grains. Mayer and Rossi (1982) have shown that the clay fraction contributes a significant portion of the surface area of estuarine muds, yet we did not find bacteria colonizing any grains smaller than about 10 ,m. We sug- gest three hypotheses to explain this pat- tern. First, unless the particle is appre- ciably larger than the bacterium it will not provide protection against resuspen- sion by fluid shear forces in a high energy environment. Second, there will be little room for colony formation on small grains. Third, clay grains in these sediments have very smooth surfaces, with virtually no shallow depressions apparently so fa- vored by bacteria.
The strong association of clay grains with microbial exudates observed in our SEM study suggests that the relationship of bacteria to surface area may be due in part to the bacteria rather than to the sur- face area. Because clay accumulation is so important to the total sediment specif- ic surface area and because the accumu-
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

880 DeFlaun and Mayer
lation of clays in these intertidal sedi- ments evidently requires biological incorporation (Anderson et al. 198 la; Cole 1979; Dempsey 1981b), then the propor- tionality of bacteria to surface area is per- haps caused by the extent of bacterial col- onization (controlled by temperature or a correlative) controlling clay incorpora- tion, rather than vice versa.
Nevertheless, bacteria may colonize sand and silt grains to an extent propor- tional to grain surface areas, assuming that the fraction of desirable area in shallow depressions is constant with decreasing grain size while surface area is inversely proportional to grain size. An important implication of such a trend, however, is that hypotheses suggesting that selection of particles by deposit feeders is posi- tively related to their surface area (e.g. Taghon et al. 1978) should apply only to grains coarser than fine silts.
The similarity of bacterial colonization and organic matter accurmulation on grains of different sizes presents another cause and effect problem. Higher bacterial con- centrations on fine silt than on clay grains may result in higher organic matter ac- cumulations in fine silts, as a result of processes such as mucus formation. Con- versely the bacteria may have simply re- sponded to higher organic matter levels on fine silts than on clays, which accu- mulated there for some other reason.
The extensive development of mucus coatings in spring and summer lends sup- port to the recent hypothesis of Hobbie and Lee (1980) that these coatings may be a more significant source of nutrition for deposit feeders than the bacteria themselves. We were not able to quantify the accumulation of mucus coatings, but our visual impressions were that they formed a volume several times that of the bacterial cells. Epipelic diatonms also con- tributed mucus to the grains, but benthic microalgae on this mudflat exhibit a sea- sonal colonization sequence quite oppo- site to that of the bacteria (unpubl. data), with abundances highest in winter.
The seasonality of these mucus coat- ings is similar to the increases in critical shear stresses (that stress necessary to
initiate grain movement at the sediment- water interface) found by Grant et al. (1982). They, also working in New En- gland intertidal sediments, found that critical shear stresses in summer and par- ticularly in fall were higher than in win- ter; they suggested that this was due to mucus adhesion of grains in the warmer months. Although our SEM results did not show whether mucus abundance was maximal in summer or fall, our data would seem to be very supportive of their sug- gestion.
References ANDERSON, F. E., L. BLACK, L. M. MAYER, AND L.
WATLING. 1981a. A temporal and spatial study of mudflat texture. Northeast. Geol. 3: 184-192.
) L. WATLING, W. MOOK, AND L. M. MAYER. 1981b. A temporal and spatial study of mudflat erosion and deposition. J. Sediment. Petrol. 51: 729-736.
COLE, S. M. 1979. Benthic microalgae popula- tions on intertidal sediments and their role as precursors to salt marsh development, p. 25-42. In R. L. Jeffries and A. J. Davy [eds.], Ecolog- ical processes in coastal environments. Black- well.
COLEMAN, A. W. 1980. Enhanced detection of bacteria in natural environments by fluoro- chrome staining of DNA. Limnol. Oceanogr. 25: 948-951.
DALE, N. G. 1974. Bacteria in intertidal sedi- ments: Factors related to their distribution. Limnol. Oceanogr. 19: 509-518.
DEMPSEY, M. J. 1981a. Marine bacterial fouling: A scanning electron microscope study. Mar. Biol. 61: 305-315.
1981b. Colonization of antifouling paints by marine bacteria. Bot. Mar. 24: 185-191.
FOLK, R. L. 1974. Petrology of sedimentary rocks. Hemphill.
FRANKEL, L. 1977. Microorganism induced weathering of biotite and hornblende grains in estuarine sands. J. Sediment. Petrol. 47: 849- 854.
, AND D. J. MEAD. 1973. Mucilagenous ma- trix of some estuarine sands in Connecticut. J. Sediment. Petrol. 43: 1090-1095.
GRANT, W. D., L. F. BOYER, AND L. P. SANFORD. 1982. The effects of bioturbation on the initi- ation of motion of intertidal sands. J. Mar. Res. 40: 659-677.
HARGRAVE, B. T. 1972. Aerobic decomposition of sediment and detritus as a function of particle surface area and organic content. Limnol. Oceanogr. 17: 583-596.
HOBBIE, J. E., AND C. LEE. 1980. Microbial pro- duction of extracellular material: Importance in benthic ecology, p. 341-346. In Marine benthic
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions

Bacteria in intertidal sediments 881
dynamics. Belle W. Baruch Symp. Mar. Sci. (llth). Univ. S. Carolina.
KRUMBEIN, W. C., AND F. J. PETTIJOHN. 1938. Manual of sedimentary petrography. Appleton- Century.
MAYER, L. M., AND P. M. Rossi. 1982. Specific surface areas in coastal sediments: Relation- ships with other textural factors. Mar. Geol. 45: 241-252.
MEADOWS, P. S., AND J. G. ANDERSON. 1966. Mi- croorganisms attached to marine and freshwa- ter sand grains. Nature 212: 1059-1060.
MEYER-REIL, L. A. 1977. Bacterial growth rates and biomass production, p. 223-236. In G. Rheinheimer [ed.], Microbial ecology of a brackish water environment. Springer.
NICKELS, J. S., AND OTHERS. 1981. Effect of sili- cate grain shape, structure and location on the biomass and community structure of colonizing marine microbiota. Appl. Environ. Microbiol. 41: 1262-1268.
PORTER, K. G., AND Y. S. FEIG. 1980. The use of DAPI for identifying and counting aquatic mi- croflora. Limnol. Oceanogr. 25: 943-948.
RUBLEE, P. A. 1982. Seasonal distribution of bac- teria in salt marsh sediments in North Carolina. Estuarine Coastal Shelf Sci. 15: 67-74.
, AND B. E. DORNSEIF. 1978. Direct counts of bacteria in the sediments of a North Carolina salt marsh. Estuaries 1: 188-191.
SCHR6DER, H. G. J., AND F. B. VAN Es. 1980. Dis-
tribution of bacteria in intertidal sediments of the Ems-Dollard estuary, Netherlands. Neth. J. Sea Res. 14: 268-287.
SLABAUGH, W. H., AND A. D. STUMP. 1964. Sur- face areas and porosity of marine sediments. J. Geophys. Res. 69: 4773-4778.
TAGHON, G. L., R. F. SELF, AND P. A. JUMARS. 1978. Predicting particle selection by deposit feeders: A model and its implications. Limnol. Oceanogr. 23: 752-759.
TANOUE, E., AND N. HANDA. 1979. Differential sorption of organic matter by various sized sed- iment particles in recent sediment from the Bering Sea. J. Oceanogr. Soc. Jap. 35: 199-208.
TURCHENEK, L. W., AND J. M. OADES. 1979. Frac- tionation of organo-mineral complexes by sedi- mentation and density techniques. Geoderma 21: 311-343.
WEILER, R. R., AND A. A. MILLS. 1965. Surface properties and pore structure of marine sedi- ment. Deep-Sea Res. 12: 511-529.
WIESE, W., AND G. RHEINHEIMER. 1978. Scanning electron microscopy and epifluorescence in- vestigation of bacterial colonization of marine sand sediments. Microb. Ecol. 4: 175-188.
ZoBELL, C. E. 1943. The effect of solid surfaces upon bacterial activity. J. Bact. 46: 39-56.
Submitted: 3 June 1982 Accepted: 14 March 1983
This content downloaded from 91.229.248.111 on Sat, 14 Jun 2014 13:22:50 PMAll use subject to JSTOR Terms and Conditions