relacion entre infecciones quiescentes … · relacion entre infecciones quiescentes de...
TRANSCRIPT

RELACION ENTRE INFECCIONES QUIESCENTES DE Colletotrichum gloeosporioides (Penz) Y LOS DIFERENTES ESTADOS FENOLÓGICOS
DEL FRUTO DE MANGO (Magnifera indica L) VARIEDAD HILACHA
LORENA PARRA AYA
TRABAJO DE GRADO Presentado como requisito parcial
Para optar al título de
Microbiólogo Agrícola y veterinario
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS BÁSICAS
CARRERA DE MICROBIOLOGÍA AGRÍCOLA Y VETERINARIA BOGOTA
2008

NOTA DE ADVERTENCIA Artículo 23 de la Resolución Nº 13 de julio de 1946 “La Universidad no se hace responsable por los conceptos emitidos por sus alumnos en sus trabajos de tesis. Solo velará porque no se publique nada contrario al dogma y a la moral católica y porque las tesis no contengan ataques personales contra persona alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la justicia".

RELACION ENTRE INFECCIONES QUIESCENTES DE Colletotrichum gloeosporioides (Penz) Y LOS DIFERENTES ESTADOS FENOLÓGICOS
DEL FRUTO DE MANGO (Magnifera indica L) VARIEDAD HILACHA
LORENA PARRA AYA
APROBADO
____________________________
Mª Clemencia Forero de la Rotta Ingeniera Agrònoma M.Sc.
Directora
________________________ _____________________________ José Salvador Montaña Sandra Gómez Biólogo M.Sc. Ingeniera Agrónoma M.Sc.

RELACION ENTRE INFECCIONES QUIESCENTES DE Colletotrichum gloeosporioides (Penz) Y LOS DIFERENTES ESTADOS FENOLÓGICOS
DEL FRUTO DE MANGO (Magnifera indica L) VARIEDAD HILACHA
LORENA PARRA AYA
APROBADO
________________________ ________________________ INGRID SHULER JANETH ARIAS PALACIOS Bióloga Ph.D Bacterióloga M.Sc-M.Ed Decana Académica Directora de Carrera

Agradecimientos
A Dios.
A mis padres Arnulfo Parra y Betty de Parra por darme la oportunidad de
estudiar y ser una persona de bien, a mis hermanas Johana, Natalia y Maria
Camila por el apoyo que me brindaron.
A mi novio Julian por todo el tiempo, colaboración, dedicación y apoyo para
poder realizar con éxito este proyecto.
Maria Clemencia Forero de la Rotta, por la confianza, por su colaboración,
por sus enseñanzas y por guiarme en este proyecto.
A Jairo Osorio por la oportunidad y la confianza de dejarme realizar este
trabajo.
A Juan Clímaco Hío y Erika Patricia Martínez. Laboratorio de Fitopatología.
CORPOICA. Por su apoyo y colaboración.
A Carlos y Dimas. Laboratorio de Fitopatología. CORPOICA Nataima. Por
sus colaboraciones.
A Johana Rozo por su apoyo, colaboración y su amistad.
A mis amigas de la universidad, Silvia Duarte, Irene Chabur, Carmen Pinzon
y Diana Valbuena por estar ahí.

RESUMEN
La antracnosis cuyo agente causal es Colletotrichum gloeosporioides, es
considerada la enfermedad mas limitante en cultivos de mango variedad
hilacha ocasionando numerosas pérdidas en su producción, incitando a la
búsqueda de alternativas de manejo iniciando por un conocimiento pleno del
agente causal y su relación parasítica con el hospedero. Se obtuvieron
aislamientos de Colletotrichum gloeosporioides de frutos aparentemente
sanos de Mango variedad Hilacha, en los diferentes estados fenológicos (E1,
E2, E3 y E4) desde fruto en desarrollo hasta fruto maduro.
Las infecciones quiescentes se presentaron con mayor frecuencia en estados
fenológicos iniciales (E1 y E2), comparados con los estados de desarrollo
avanzado, debido probablemente al efecto del pH del fruto.
Se realizaron pruebas biológicas para determinar la patogenicidad del hongo
y confirmar la especie de Colletotrichum que se presenta en los frutos de
esta especie. Las pruebas de sensibilidad al Benomyl, la prueba cualitativa
de la proteaza y el medio selectivo para Colletotrichum spp. determinaron la
presencia de la especie C. gloeosporioides como el agente causal de la
antracnosis. La patogenicidad de los aislamientos se evaluó inoculando
bloques de agar sobre los frutos. Se demostró que todos los aislamientos
causan infección en los frutos.
Palabras Claves: Antracnosis, pH, Fenología e Incidencia.

ABSTRACT
The anthracnose whose causative agent is Colletotrichum gloeosporioides, is
considered the most limiting disease in mango crops causing numerous
losses in their production, prompting the search for alternative management
by initiating full knowledge of the causative agent and its parasitic relationship
with the accommodation provider. The Insulation of Colletotrichum
gloeosporioides earned of fruits apparently of healthy Mango variety
stringless, at different phenological stages (E1, E2, E3 and E4) from
developing fruit until ripe fruit. The results showed that quiescent infections
were presented with greater frequency in phenological stages initials (E1 and
E2), compared with the advanced stages of development, probably due to the
effect of pH of the fruit.
Biological tests were conducted to determine the pathogenicity of the fungus
and confirm the kind of Colletotrichum occurring in the fruits of this kind.
Evidence of sensitivity to Benomyl, the qualitative evidence from the proteaza
and selective medium for Colletotrichum spp. determined the presence of the
species C. gloeosporioides as the causal agent of anthracnose. The
pathogenicity of the isolates were evaluated by inoculating agar blocks on the
fruits. It showed that all isolates cause infection in the fruits.
Key Words: Anthracnose, pH, Phenology and incidents.

CONTENIDO
Pag
1. INTRODUCCIÓN 14
2. MARCO TEORICO 16
2.1 Mango común o Hilacha (Mangifera indica L.) 16
2.1.1 Botánica y Descripción 16
2.1.2 Condiciones Agroecológicas 18
2.1.3 Plagas y Enfermedades 19
2.2 Agente Causal 19
2.2.1 Taxonomía 20
2.2.1.1 Estado Telemorfo 20
2.2.1.2 Estado Anamorfo 20
2.3 Generalidades 21
2.4 Desarrollo de la Enfermedad 22
2.4.1 Infecciones quiescentes 24
2.5 Síntomas 28
2.6 Condiciones Epidemiológicas 29
2.7 Métodos de Diagnóstico 31
2.8 Control 32
3. FORMULACION DEL PROBLEMA Y JUSTIFICACION 33
4. OBTETIVOS 35
4.1. Objetivo general 35
4.2. Objetivos específicos 35
5. MATERIALES Y METODOS 36
5.1. Localización de la Investigación 36
5.2. Muestreos de frutos. 37
5.3. Detección de infecciones quiescentes 39
5.4. Aislamiento de Colletotrichum sp. 39
5.4.1. Purificación de Aislamientos 40

5.5. Pruebas Biológicas 41
5.5.1. Medio selectivo para Colletotrichum sp. 41
5.5.2 Pruebas de Patogenicidad 42
5.5.3 Prueba de tolerancia al Benomyl 44
5.5.4. Prueba de la Proteasa 44
5.6. Determinación del pH en los diferentes estados del fruto de
Mango. 45
6. RESULTADOS Y DISCUSION 47
6.1 Aislamientos de Colletotrichum sp. 47
6.2 Detección de Infecciones Latentes en frutos asintomáticos de
Mango. 49
6.3 Medio selectivo para Colletotrichum sp. 52
6.4 Prueba de Benomyl 53
6.5 Prueba de la Proteasa. 54
6.6 Pruebas de Patogenicidad 56
6.7 Determinación de pH en Frutos sanos y Enfermos de Mango. 57
7. CONCLUSIONES 63
8 RECOMENDACIONES 65
9. REFERENCIAS 66
ANEXOS 74

TABLA DE FIGURAS
Pag
Figura 1. Ciclo de vida de la enfermedad de Antracnosis causada por
Glomerella cingulata y Colletotrichum gloeosporioides. 22
Figura 2. Proceso de las infecciones quiescentes en Mango. 24
Figura 3. Eventos entre el patógeno y el hospedero dentro de la
infección quiescente en frutos por C. gloeosporioides. 27
Figura 4. Frutos de Mango. 28
Figura 5. Estados Fenológicos de Mango. E1, E2, E3 y E4. 37
Figura 6. Frutos de Mango sanos con y sin bloques de agar con
micelio del microorganismo. 42
Figura 7. Características microscópicas de C. gloeosporioides. 46
Figura 8. Aislamientos de Colletotrichum sp. 47
Figura 9. Porcentaje de infección quiescente de C. gloeosporioides
en los diferentes estados fenológicos de frutos de mango. 49
Figura 10. Prueba de Benomyl. 52
Figura 11. Prueba de la Proteasa. 54
Figura 12. Frutos de Mango con síntomas de antracnosis iniciales y
avanzadas. 55
Figura 13. Efecto de Antracnosis sobre el pH en mesocarpio y
exocarpio y sobre tejido tomado en el fruto. 60

LISTA DE TABLAS
Pag
Tabla 1. . Etapas fenológicas de los frutos de Mango, de acuerdo con
la edad, dimensiones, peso y otras características.
37
Tabla 2. Clasificación de las especies según los medios de
evaluación biológica.
41
Tabla 3. Periodo de Incubación de los Diferentes Estados
Fenológicos de Frutos de Mango.
48
Tabla 4. Valores de pH de Frutos Sanos de Mango. 56
Tabla 5. Valores de pH de Frutos Enfermos de Mango. 57

LISTA DE ANEXOS
Pag
ANEXO 1. Preparación de Agar Agua. 73
ANEXO 2. Preparación de PDA. 74
ANEXO 3. Protocolo de Cámaras Húmedas. 75
ANEXO 4. Protocolo Monospóricos. 77
ANEXO 5. Medio para evaluar la reacción de la proteasa producida por Colletotrichum acutatum.
80
ANEXO 6. Protocolo Medio Suplementado con base Benomyl. 81
ANEXO 7. Protocolo Medio Suplementado con Hidróxido de Cobre y Estreptomicina.
83
ANEXO 8. Incidencia de Infecciones Quiescentes en los estados fenológicos de frutos de Mango.
85
ANEXO 9. Pruebas de Validación, Normalidad y comparación de promedios en la incidencia de infecciones quiescentes en los estados fenológicos de frutos de mango.
88
ANEXO 10. Características Morfológicas de los Aislamientos. 91
ANEXO 11. Prueba de Benomyl con 2 repeticiones. 92
ANEXO 12. Prueba de la Proteasa con 2 repeticiones. 93
ANEXO 13. Prueba de medio selectivo para Colletotrichum sp. 94
ANEXO 14. Presencia de síntomas iniciales de Antracnosis en los tratamientos de Patogenicidad.
96
ANEXO 15. Efecto de la Enfermedad sobre el pH en áreas sanas y enfermas de frutos enfermos y en frutos sanos.
97
ANEXO 16. Pruebas de Validación, Normalidad y comparación de promedios en el efecto de la enfermedad sobre el pH.
111

13
1. INTRODUCCIÓN
El mango es una de las frutas tropicales considerada de gran importancia
socioeconómica en Colombia junto con el banano y los cítricos, ya que
constituye una alternativa productiva de alto potencial en el mercado nacional
y de exportación.
En Colombia existen alrededor de 13.900 hectáreas sembradas con este
frutal, que producen aproximadamente 170.000 toneladas de frutas al año.
Los departamentos de mayor producción son Tolima y Cundinamarca.
Debido a la expansión del mercado interno y a las posibilidades de
exportación se requiere la utilización de tecnología para optimizar y mejorar
los rendimientos. Dentro de la tecnología demandada el mayor interés se
centra en el manejo de la antracnosis, por ser el problema fitosanitario más
limitante de la producción de mango.
La antracnosis es causada por el hongo Colletotrichum gloeosporioides
(Penz), afecta frutos y otras estructuras de la planta. Las pérdidas por esta
enfermedad durante la cosecha se han calculado aproximadamente en un
40%; no obstante, los daños en poscosecha como consecuencia de
infecciones quiescentes hacen de la enfermedad una verdadera amenaza
para la competitividad de los sistemas productivos.
Actualmente en Colombia el manejo de la enfermedad en mango se
fundamenta en la aspersión programada de fungicidas durante la fase
productiva, de esta forma se hacen entre 8 y 12 aspersiones, lo cual resulta
muy costoso y perjudicial para los seres vivos y el medio ambiente; en
muchos casos no se obtienen resultados favorables. Debido a la importancia
económica de esta enfermedad es conveniente conocer la incidencia de las
infecciones quiescentes de C. gloeosporioides (Penz.) a lo largo del

14
desarrollo del fruto y simultáneamente establecer una posible relación entre
ellos, para llegar a enfocar de manera adecuada programas de control
integrado que presenten menor grado de contaminación al ambiente.

15
2. MARCO TEORICO
2.1. Mango común o Hilacha (Mangifera indica L.)
El mango es una de las frutas más importantes en el trópico, se cultiva en
todos los países que se encuentran en esta área. Se conoce hace mas de
4000 años en la India, donde en la actualidad existen mas de 800.000
hectáreas cultivadas con este frutal. (Salazar, 1992).
Es originario de la India, de donde se distribuyó por todo el suroeste de Asia
y el archipiélago malayo. A América llegó por dos vías: de Asia fue llevado
por los portugueses al sur de África y luego a las costas brasileñas, en el
siglo XVI. Los españoles lo introdujeron a México en los siglos XV y XVI,
mediante el comercio que se estableció con Filipinas. (Salazar, 1992).
Colombia cuenta con extensas zonas aptas para su cultivo: la costa Atlántica,
los Llanos Orientales, todo el valle del Magdalena, Cauca (Patía), Guajira,
Tolima y Huila. La gran aceptación de esta fruta tanto en el mercado nacional
como en el internacional, hace que el cultivo a escala comercial en Colombia
sea una empresa rentable y de gran porvenir para el país (Páez, 2006).
2.1.1. Botánica y Descripción El mango Mangifera indica L. pertenece a la familia de las anacardiáceas, el
género Magnifera tiene 62 especies arbóreas, de las cuales 16 producen
fruto comestible. Forman parte del orden de las terebintales. (Salazar, 1992).

16
Es un árbol de gran porte que puede llegar a medir 30 m de altura y de gran
longevidad. El tronco es grueso con una corteza parda oscura, áspera, de
ramas separadas; la forma del árbol depende de varios factores:
propagación, podas, variedad y medio ambiente. El sistema radicular
presenta un amplio desarrollo pudiendo penetrar las raíces principales de 6 a
8 metros, mientras que se extienden en un radio de 10 metros, esta
distribución le permite resistir épocas de baja humedad. (Salazar, 1992).
Las hojas al principio tienen un color rojizo o púrpura, que del cual cambian a
verde claro y luego a verde oscuro. Durante estos estados su actividad
fotosintética cambia dado a la acumulación de clorofila en los tejidos. Las
ramas de un mismo árbol no crecen al tiempo por lo cual es corriente ver el
árbol en diferentes estados de desarrollo y florecido todos los años. (Salazar,
1992).
La flor se presenta en paniculas ramificadas, provenientes de yemas
terminales; el número de flores por panicula puede variar de 300 a 2.000,
dependiendo de la variedad. Las flores tienen un pedúnculo muy corto: el
cáliz se forma de 5 sépalos libres, verdosos, de 2 a 4 mm de largo y de 1 a 2
mm de ancho; los pétalos también libres y caedizos miden de 3 a 6 mm de
largo por 1 a 2 mm de ancho y tienen ápice agudo y curvo. Los árboles
provenientes de semilla florecen de 6 a 8 años después de sembrados,
mientras que los injertados florecen en la mitad del tiempo. La floración en
mango tiende a ser alterna o bianual. (Cartagena, 2001).
El fruto es una drupa con endocarpio duro, del cual salen fibras que se
extienden a través de la pulpa. Varía considerablemente en tamaño, color,
forma, sabor y olor, así como en la cantidad de fibra en la pulpa y en sus
características químicas. El mesocarpio es firme, con jugo dulce y de un

17
exquisito sabor, su color varía de amarillo cremoso a naranja oscuro.
(Cartagena, 2001).
El mango se caracteriza por una elevada caída de flores completas y frutos
jóvenes, llegando a ser hasta del 99%. Algunos autores aseguran que solo el
0.1% de las flores bisexuales llegan a producir fruto maduro. La mayor caída
de frutos se presenta en las tres primeras semanas. (Cartagena. 2001). El
mango común o hilacha tiene un fruto alargado que se caracteriza por tener
la pulpa fibrosa. Es conocido también como Hilaza, brechoso, mango de
puerco, dependiendo de la zona de producción. (ICONTEC, 2002).
2.1.2. Condiciones Agroecológicas
El mango puede prosperar en zonas subtropicales, donde la temperatura
promedio es de 15°C, es muy sensible a bajas temperaturas mostrando una
reducción de crecimiento. Los suelos muy alcalinos dañan el cultivo, los
límites del pH para el cultivo de mango están entre 5.5 y 7.5. Por tener un
sistema radicular profundo, las características del subsuelo y el nivel freático
se deben tener en cuenta. (Salazar, 1992).
El mango fructifica en zonas en donde la precitación varía de 240 a 1500
mm. No es tan importante la cantidad de agua ni el volumen final del año,
sino su distribución durante el año. La humedad relativa es uno de los
factores limitantes para el mango porque favorece el desarrollo de
enfermedades; por esta razón se debe desarrollar en climas secos, donde la
humedad relativa sea baja. Todos estos aspectos de altura, temperatura,
humedad relativa y precipitación indican que el cultivo de mango es exigente
en radiación solar para el buen desarrollo de los frutos, de manera que
presenten excelente calidad y color. (Cartagena, 2001).

18
Los suelos de Tolima y Huila sembrados con mango son arenosos con un
pH que oscila entre 6.5 y 7.2 y su fertilidad es medianamente alta. Estas
regiones se encuentran ubicadas entre los 380 y 600 m.s.n.m., la
temperatura promedio es de 28°C y la humedad relativa promedio es de
68%, el promedio de lluvias está entre 1000 y 1300 mm anuales. (Cartagena,
2001).
2.1.3. Plagas y Enfermedades En la actualidad el cultivo de mango se ve afectado por innumerables plagas
y enfermedades, las plagas mas importantes que afectan el cultivo de mango
son: Mosca de la fruta Anastrepha sp., el falso piojo blanco Aulacaspis
tubercularis, la escama articulada Selenaspidus articulatus y los comedores
de hoja Megalopyge lanata. Entre las enfermedades fungosas se encuentra
la antracnosis que es la principal enfermedad que ataca los mangos en
Colombia y el agente causal es C. gloeosporioides (ICA, 1996).
2.2. Agente Causal de la Antracnosis
Colletotrichum sp., es considerando el principal agente causal del deterioro
en precosecha y poscosecha de muchos frutales tropicales y subtropicales
(Bernstein et al., 1995); de acuerdo con numerosas investigaciones, la gran
mayoría de estas enfermedades se atribuyen a ataques del hongo C.
gloeosporioides (Zulfiqar et al, 1996), sin desconocer que también causa
daños en otros tejidos de la planta; sin embargo es durante la poscosecha
donde mayores pérdidas económicas se generan en todo el mundo (Ploetz, y
Prakash, 1997).

19
2.2.1. Taxonomía
Según Arauz, (2000) la antracnosis en mango es causada por el hongo
Colletotrichum gloeosporioides (Penz.) Penz. & Sacc (anaformo),
Glomerella cingulata (Telemorfo) Spauld. & H. Schrenk.
2.2.1.1. Estado Telemorfo
REINO Fungi
SUBDIVISION Ascomicotyna
CLASE Pyrenomicetos
ORDEN Sphaeriales
GENERO Glomerella
ESPECIE cingulata
2.2.1.2. Estado Anamorfo
REINO Fungi
SUBDIVISION Deuteromicotina
CLASE Deuteromycetes
ORDEN Melanconiales
GENERO Colletotrichum
ESPECIE gloeosporioides

20
2.3. Generalidades
Colletotrichum es un habitante muy común en cultivos y con frecuencia no
ocasiona síntomas de enfermedad en los órganos de la planta; en los frutos
de algunas especies frutales solo se manifiesta cuando el pericarpio es
debilitado, o cuando ocurren aumentos en los niveles de etileno como
resultado de procesos de maduración de los frutos. (Adoskaveg y and Hartin,
1997).
Este hongo presenta un micelio septado de coloración hialina o castaña
clara, los acérvulos separados tienen forma de disco o cojín, de textura
cerosa y puede ubicarse en forma subepidermal, epidermal o subcuticular.
Típicamente presenta setas o espinas negras en los bordes o entre el
conidióforo; los acérvulos están formados por pseudoparénquima con
paredes delgadas o gruesas, conidióforos simples, elongados con
numerosas conidias. Las conidias son hialinas, curvadas y fusiformes, y por
lo general un acérvulo setoso. Las conidias son producidas en estructuras
especializadas llamadas acérvulos, que se forman en la superficie de los
tejidos infectados. (Barnett ,1998 y Holliday, 1995). El género Colletotrichum
tiene enorme variación ecológica, morfológica y patológica, las diferentes
formas en la naturaleza varían desde saprofito a cepas parasíticas con un
estrecho rango de hospederos (Mena, 1999).
Las conidias son producidas en masas mucilaginosas típicamente hundidas,
de color rosado y con un contorno irregular en las lesiones necróticas, este
tipo de lesiones que con frecuencia se observa sobre las hojas, frutos y
ramas son las conocidas como antracnosis; sin embargo en algunos frutos
también es frecuente observar lesiones como chancros, cicatrices, verrugas
o costras con relieve. (Mena, 1999).

21
Una característica de algunas especies de Colletotrichum, es que pueden
causar infecciones latentes o quiescentes sobre los frutos, que
posteriormente se desarrollan durante la fase de maduración. Las diferentes
especies sobreviven por largos periodos sobre los desechos vegetales o en
el suelo; el patógeno mas común en los trópicos es Colletorichum
gloeosporioides. (Holliday, 1995).
Las infecciones quiescentes en el contexto de enfermedades de poscosecha
involucra la inhibición del desarrollo del patógeno a través de condiciones
fisiológicas impuestas por el hospedero hasta que se lleva a cabo el estado
de maduración. Una vez el hongo penetra la capa exterior del fruto, este
permanece allí en estado de quiescencia o dormancia hasta que ocurren
cambios en la superficie del fruto los cuales proporcionan las condiciones
adecuadas para permitir una infección (Contreras, 2006).
2.4. Desarrollo de la enfermedad
El inóculo del microorganismo que ha sobrevivido en hojas, frutos y ramas
afectadas causa las infecciones de los frutos por la dispersión de las conidias
transportadas en el agua. (Adoskaveg y Hartin, 1997). Una vez dispersas las
conidias se adhieren a la superficie del hospedero y germinan en un periodo
de 12 a 24 horas y luego produce tubo germinal que penetra la cutícula
directamente. Las fuentes de inóculo son las conidias producidas en
acérvulos o las ascosporas producidas y liberadas en el peritecio. (Figura 1.).
La hifa infectiva penetra directamente la cutícula colonizando la pared celular
de las células de hospedero (Kuo, 1999). Existen varias formas de
penetración, una de ellas es a través de aberturas naturales como estomas,
lenticelas y otras por penetración directa o a través de pequeñas heridas.

22
Figura 1. Ciclo de vida de la enfermedad de Antracnosis causada por Glomerella cingulata y Colletotrichum gloeosporioides. Tomado de Agrios 2002.
La infección se puede desarrollar casi en cualquier tejido de la planta. Una
vez el microorganismo penetra la cutícula, se pueden presentar dos
estrategias de infección: la primera es conocida hemibiotrofía intracelular y la
segunda necrotrofía subcuticular e intramural. (Agrios, 2002).
Muchas especies de Colletotrichum presentan procesos infectivos en dos
fases: la fase asintomática inicial, donde el patógeno se establece en los
tejidos sin matar las células y son consideradas especies biotróficas
(hemibiotrofia intracelular) y la fase destructiva visible (necrotrofía

23
subcuticular e intramural), donde el patógeno crece bajo la cutícula y disuelve
extensamente la matriz péptica de las células epidermales (Bailey, 1992). El
desarrollo intramular se asocia a la hinchazón y disolución de las paredes
celulares (O´Connell et al., 2000).
En condiciones favorables, el hongo puede crecer rápidamente en la planta y
causar diversos síntomas muy rápidamente, pero en otras circunstancias el
hongo puede permanecer quiescente dentro de los tejidos por un largo
periodo, que en algunos casos solo se evidencia después de la cosecha
(Figura 2.). Pero también durante periodos de condiciones ambientales
extremas, puede permanecer en latencia por algún tiempo en el suelo o
permanecer por largos periodos sobre tejido vegetal muerto (CABI-EPPO,
1996).
2.4.1. Infecciones quiescentes
Una de las características destacadas de Colletotrichum sp., es su capacidad
de sobrevivir en estado de dormancia o quiescencia cuando las condiciones
ambientales o fisiológicas del hospedero le impiden desarrollarse (Figura 2.).
Los cambios bioquímicos que ocurren durante la maduración el fruto, se
creen que son los que permiten que la infección continué su curso. En
aguacate, el cambio en los compuestos antifúngicos llamados dienes, son
importantes en la regulación de la quiescencia de la antracnosis. (Prusky,
1996).
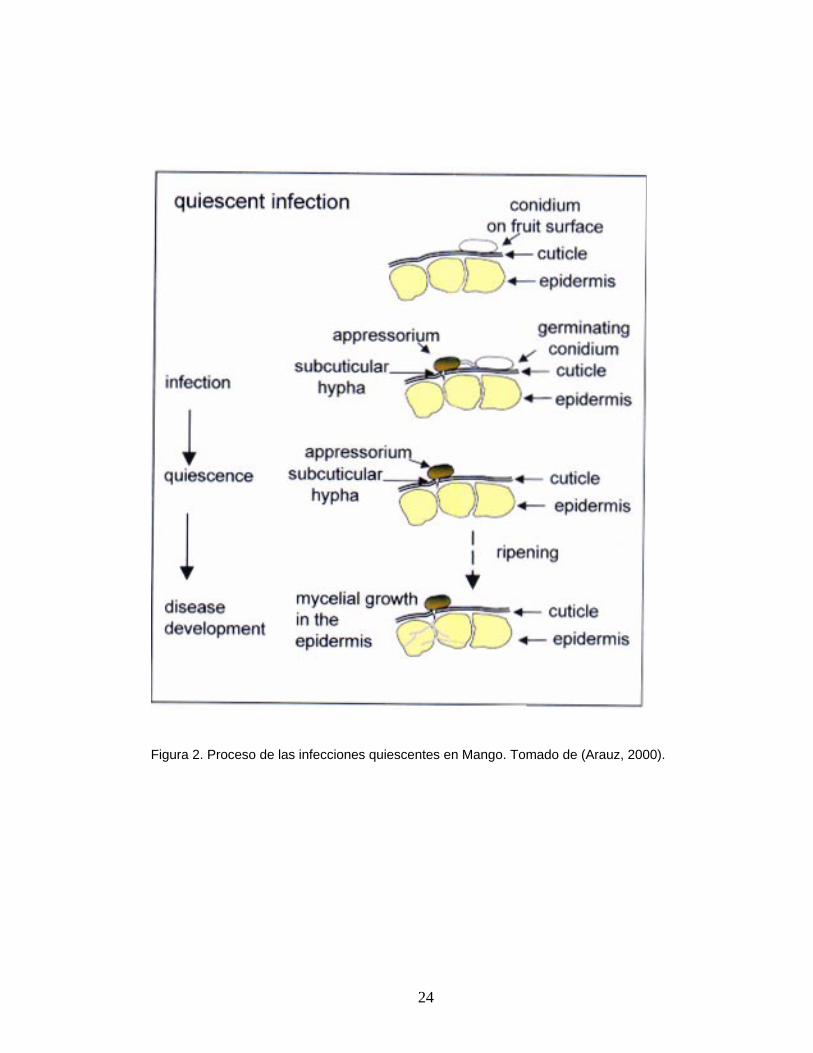
24
Figura 2. Proceso de las infecciones quiescentes en Mango. Tomado de (Arauz, 2000).

25
Según Yerhoeff (1974), una relación parasítica latente o quiescente, se
define como una condición en la cual el patógeno reduce su actividad
metabólica, deteniendo el proceso de infección por un periodo considerable
durante la vida del hospedero, que se reactiva cuando las circunstancias
fisiológicas o ambientales específicas lo permitan. De Lapeyre (2000), define
el estado quiescente del hongo como la circunstancia en la cual una espora
que entra en contacto con la superficie del fruto, germina y forma un
apresorio que se melaniza y permanece inactivo hasta la maduración del
fruto.
Según Swimburne (1983), un organismo se puede tornar quiescente en
cualquiera de las siguientes etapas de su vida: en germinación, elongación
del tubo germinativo, formación del apresorio y penetración. Un punto de
infección se desarrolla cuando la hifa penetra la cutícula y la pared celular del
hospedero, con la ayuda de enzimas cutinasas y celulolíticas. La fase latente
puede ser corta como ocurre en las enfermedades de floración, y de dos o
tres semanas a varios meses, como ocurre en frutos. (Bailey, 1992). En la
permanencia de la latencia del patógeno dentro del hospedero hay una
dinámica de equilibrio entre el hospedero, el patógeno y el ambiente; de tal
manera que los cambios fenológicos y fisiológicos en el hospedero, en el
ambiente o en ambos, conllevan a la pérdida del equilibrio establecido y
permiten que el patógeno continué su ataque. (Jarvis, 1994).
Flaishman y Kolattukudy (1994) indican que la participación del etileno en la
terminación de la quiescencia, se debe a la habilidad de Colletotrichum sp.
para desarrollar un mecanismo donde usa la hormona de la maduración del
hospedero como señal para reactivar el proceso de infección. Este
mecanismo evita el contacto del patógeno con tejidos del hospedero que
tienen altos niveles de compuestos antifúngicos (Figura 3.)

26
INFECCIONES QUIESCENTES
PATÓGENO INTERACCIÓN HOSPEDERO Infección Desarrollo del Invasión RESISTENCIA fruto Quiescencia Fruto en maduración Estimulo Periodo Reactivación del Climatérico Patógeno Fruto maduro Invasión ENFERMEDAD Figura 3. Eventos entre el patógeno y el hospedero dentro de la infección quiescente en frutos por C. gloeosporioides. Adaptación de Arauz, 2000. Existen varias condiciones que promueven la quiescencia como condiciones
físicas adversas como el inapropiado almacenamiento de frutos, daños
físicos, mecánicos, plagas, estrés y cambios climáticos bruscos en campo; y
condiciones fisiológicas como deficiencia nutricional y cambios fisiológicos en
la maduración del fruto. (Swinburne, 1983). Las infecciones latentes han sido
reportadas en frutas como cítricos (Zulfigar et al, 1996), en tomate de árbol
(Rondón, 2003) y en mora de castilla (Forero de la Rotta, 2001). C.
gloeosporioides, ha sido asociado con infecciones quiescentes y de
poscosecha sobre frutos como mango, aguacate, papaya, maracuyá,
guayaba, cítricos, uva, granadilla, manzana y mora. (ICA, 1996).
Reducción en Compuestos Antifungoso Etileno Factores Nutricionales

27
2.5. Síntomas Colletotrichum sp. causa numerosas enfermedades en un amplio rango de
plantas a nivel mundial, tanto en precosecha como en poscosecha. En
poscosecha predomina en los trópicos, donde es un factor limitante en la
calidad de los frutos ocasionando grandes pérdidas en la comercialización
interna y en los mercados internacionales. El efecto del hongo en el
rendimiento radica en el hecho que afecta frutos en todos los estados de
desarrollo, los cuales se desechan en el momento de comercialización
(Cartagena & Vega, 2001).
En plantas de mango (M. indica), se presentan primero en las panículas
florales, con pequeñas manchas de color negras o cafés, que se van
agrandando y pueden causar ennegrecimiento total de la inflorescencia y
marchitamiento antes que el fruto inicie su formación. Los síntomas en la
superficie del fruto son manchas hundidas de color negro (Figura 4.),
presentando un alto grado de severidad después de periodos de clima
húmedo (Allende-Molar y Cols, 2003).

28
Figura 4. Frutos de Mango. A: Fruto sano, B: Fruto con lesiones de antracnosis. (Foto L.Parra).
En los frutos jóvenes se producen pequeñas lesiones hundidas, mientras en
los grandes (4-5 cm), por lo general no hay desarrollo de la enfermedad, pero
el hongo permanece en estado de quiescencia hasta que los frutos maduran
y aparecen las lesiones oscuras, necróticas, angulares o de forma irregular.
En todos los casos el desarrollo de la enfermedad va acompañado de masas
de conidias de color salmón como consecuencia de la maduración del
acérvulo.
2.6. Condiciones epidemiológicas
Aunque se han realizado varias investigaciones acerca de la epidemiología
de esta enfermedad en mango, dichos estudios se han concentrado en las
relaciones entre variables del clima y la cantidad de la enfermedad en el
cultivo. La infección ocurre cuando el fruto se desarrolla o durante los
periodos de cosecha, pero por lo general permanece en estado de
quiescencia hasta la maduración del fruto. En el cultivo, el desarrollo del
hongo se ve favorecido por condiciones ambientales como humedad relativa
A B

29
del 96% con temperaturas entre 10-30°C, ocasionando lesiones sobre hojas,
ramas e inflorescencia momificadas. (Bailey, 1992).
Existe una correlación importante entre la precipitación en todo el año y el
desarrollo de la enfermedad, demostrando que el hongo es dependiente de la
precipitación y la humedad relativa para su germinación, desarrollo y
dispersión. (Álvarez, 1996). Para el desarrollo de la enfermedad se requiere
agua durante todas las etapas, de esta manera la esporulación no solo
necesita altos niveles de humedad (95%), sino que la liberación y dispersión
de las esporas es dependiente del agua libre y se requiere de
aproximadamente cuatro horas mas de humedad a temperaturas óptimas de
20-25°C; la ubicuidad de la fuente de inóculo tiende a sobrepasar cualquier
restricción en el desarrollo de la enfermedad que pueda imponer el agua.
(Álvarez, 1996).
La variabilidad en la latencia del patógeno representa una adaptación muy
importante a las condiciones fisiológicas de los tejidos y los patrones
climáticos que ocurren durante el ciclo de cultivos perennes. Debido a esta
variabilidad el impedimento para tomar medidas de control exitosas se
dificulta. (Waller, 1992).
Los estudios realizados en tomate de árbol por Rondón (1998), determinaron
que las hojas, los pedúnculos, las ramas jóvenes, los racimos, botones,
flores, sépalos de la flor y los frutos en cualquier estado, son tejidos
potencialmente susceptibles al hongo.

30
2.7. Métodos de Diagnóstico
Las herramientas moleculares han ganado gran aceptación en los últimos
años, siendo empleados para diferenciar entre poblaciones de Colletotrichum
en diferentes hospederos y también para diferencias entre especies y
subespecies que no se permite con las técnicas tradicionales. Las técnicas
moleculares más utilizadas son: análisis de las regiones codificadoras del
DNA ribosomal (ITS), realizando amplificaciones vía PCR (Polymerase Chain
Reaction) con cebadores o primers específicos. Análisis de polimorfismo en
DNA nuclear con marcadores moleculares RAPD (Random Amplified
Polymorphic DNAs) y AFLP (Amplified Fragment Length Polymorphism) al
interior de la especie, para determinar la posible existencia y los niveles
aparentes de variabilidad genética en la población, además del análisis de
enriquecimiento de A+T-rich DNA asociado con (mt) DNA (Freeman et al.,
1998).
Los métodos tradicionales son empleados aunque en algunos casos no
permiten diferenciar entre especies y subespecies de Colletotrichum. Para
las especies C. acutatum y C. gloeosporioides, algunas características como
la velocidad de crecimiento y color predominante de la colonia en medios
selectivos, la susceptibilidad o la tolerancia que presenta el patógeno en el
medio suplementado con el fungicida benomyl y la prueba cualitativa de la
proteasa (Martín & García-Figueres, 1999), por la producción de enzimas
extracelulares que hidrolizan la caseina de la leche generando halos
transparentes de 3 a 4 mm sobre el medio de cultivo, permitiendo diferenciar
entre C. acutatum y C. gloeosporioides (Freeman et al., 1998; Peres, et al;
2002 y Abang et al., 2003).

31
2.9. Control
El control químico es el más común debido a la importancia económica del
cultivo. Se disponen de muchos químicos, entre los más utilizados se
encuentran compuestos como: dithiocarbamato, benzamidazoles y triazoles;
y otros fungicidas como chlorothalonil, imazalil y prochloraz. Los fungicidas
sistémicos son útiles y ampliamente utilizados, pero es mejor alternarlos con
aplicaciones de productos de acción protectante (Páez, 2006).
Es necesario planificar las aspersiones con los fungicidas que por lo general
inician antes de la floración y continua hasta que el fruto este en procesos de
maduración. Las aplicaciones se hacen con intervalos de 7, 14, 21 o 30 días
dependiendo de la frecuencia de la lluvia. Los productos comerciales mas
empleados son benlate 07 (1g/L), dithane M45 y oxicloruro de cobre (2-3g/L),
mezclados con un adherente (sulfatante HA o superstiker), para asegurar la
cobertura apropiada de la superficie cerosa del follaje y fruto. (Sena, 2006).
En los controles culturales se emplean podas que ayudan con el rápido
secado de la copa del árbol. La reducción de daños mecánicos o fisiológicos
por el ataque de otras plagas, disminuyen las infecciones secundarias por el
hongo. (Páez, 2006).

32
3. FORMULACION DEL PROBLEMA Y JUSTIFICACION
El área dedicada al cultivo del mango, es mayor año tras año a nivel mundial
y nacional es así como en el año 2005, según cifras del Ministerio de
Agricultura, existen aproximadamente 13.900 has de mango cultivado en 14
departamentos, las cuales producían alrededor de 170.000 toneladas
anuales de diferentes variedades. Los departamentos en los que se
concentra la mayor producción de mango son Tolima y Cundinamarca,
seguidos de Antioquia, Bolívar y Magdalena. En el periodo de 1992 al 2003
en conjunto, estos 4 departamentos, han cultivado entre el 70 y el 80% del
área cultivada a nivel nacional; mientras que la producción en el mismo
periodo fue entre el 62 y el 83%. (Ministerio de Agricultura 2005).
El cultivo de mango presenta múltiples problemas fitosanitarios, la
antracnosis es una de las enfermedades más importantes y la mas frecuente,
sobre todo en regiones con temperaturas y humedades relativas elevadas.
(Fitzell y Peak, 1984).
En Colombia la antracnosis se encuentra diseminada en todas las zonas
productoras de mango, afectando la mayoría de variedades de mango
cultivadas. En la actualidad las variedades criollas, azúcar e hilacha, son las
que ofrecen grandes ventajas en comparación con variedades mejoradas y
que si son bien utilizadas por los cultivadores, pueden liderar el mercado, la
comercialización y el precio del mango a nivel nacional; ya que poseen
resistencia a plagas y enfermedades, no exigen un manejo agronómico
estricto y presentan características óptimas para la obtención de pulpa
(Cartagena & Vega, 2001).
El estudio de infecciones quiescentes causadas por Colletotrichum sp. es de
gran importancia, ya que conocer su dinámica e incidencia de acuerdo con

33
los diferentes estados de desarrollo del fruto, permite desarrollar las
estrategias de control. A pesar de que estas infecciones se inician en el
campo y solo se desarrollan cuando el fruto madura, ocasionan grandes
pérdidas en el momento de su comercialización.

34
4. OBJETIVOS 4.1. OBJETIVO GENERAL Determinar la relación entre el estado de desarrollo de frutos de mango
variedad hilacha y la incidencia de infecciones quiescentes de Colletotrichum
gloeosporioides.
4.1 OBJETIVOS ESPECIFICOS 4.2.1. Determinar la incidencia de infecciones quiescentes de Colletotrichum
gloeosporioides en cada uno de los estados fenológicos del fruto.
4.2.2. Confirmar mediante pruebas biológicas la patogenicidad del agente
causal de las infecciones quiescentes encontradas en los frutos recolectados.
4.2.3. Determinar las variaciones del pH de los frutos enfermos durante los
diferentes estados de maduración.

35
5. MATERIALES Y METODOS
5.1. Localización de la investigación
El estudio se adelantó entre los meses de noviembre 2007 y febrero 2008,
analizando la presencia y cantidad de infecciones quiescentes en frutos de
mango variedad hilacha en los estados fenológicos, muestreados cada 25
días en los lotes del Centro de Investigación Nataima de la Corporación
Colombiana de Investigación, CORPOICA que se encuentra ubicado en el
departamento del Tolima, municipio de Espinal, donde la temperatura
promedio anual está en 28 ºC, con una precipitación de 1300 mm al año, una
humedad relativa del 70%, 430 m.s.n.m de altitud y presenta una ubicación
de 4° 12´ de latitud norte y 74° 56´ longitud oeste. (IDEAM, 2006).
La investigación se desarrolló en las instalaciones de Fitopatología del
Programa Manejo Integrado de Plagas de CORPOICA, Tibaitatá, ubicado en
el municipio de Mosquera (Cundinamarca), km 14 de Bogotá, D. C. (N 4° 42',
W 74° 12'; 2543 msnm). Los ensayos realizados permitieron detectar y
cuantificar las infecciones quiescentes y mediante pruebas biológicas se
determinó el agente causal de la antracnosis. Después de haber establecido
el comportamiento de las infecciones quiescentes en los fruto de mango, se
desarrollaron pruebas para determinar el pH en frutos y así establecer una
posible relación entre las infecciones quiescentes y los valores de pH.

36
5.2. Muestreos de frutos
Los frutos se seleccionaron en un lote de árboles de mango variedad hilacha,
de 15 años de edad, al los que generalmente se les realiza podas de
formación y un control periódico de malezas.
El número de frutos que fueron muestreados corresponden al tamaño mínimo
de muestra que permite un análisis estadístico adecuado con un error
experimental de 0.05. Se realizaron 4 muestreos entre los meses de
Noviembre 2007 y Febrero 2008, con intervalos de 25 días para facilitar la
selección de los frutos en el campo, y de acuerdo con la metodología
sugerida por Medilicott et al (1986), con algunas modificaciones, en cada
muestreo se colectaron al azar 112 frutos (aparentemente sanos) en
diferentes estados de desarrollo, (Tabla 1.); para el muestreo se tomaron
frutos de los estados E1, E2, E3 y E4. (Figura 5.).
Los frutos colectados fueron empacados en papel dentro de bolsas plásticas
una vez removidos del árbol y se colocaron en una nevera de icopor, para su
transporte al Laboratorio de Fitopatología del Centro de Investigación de
Tibaitatá.
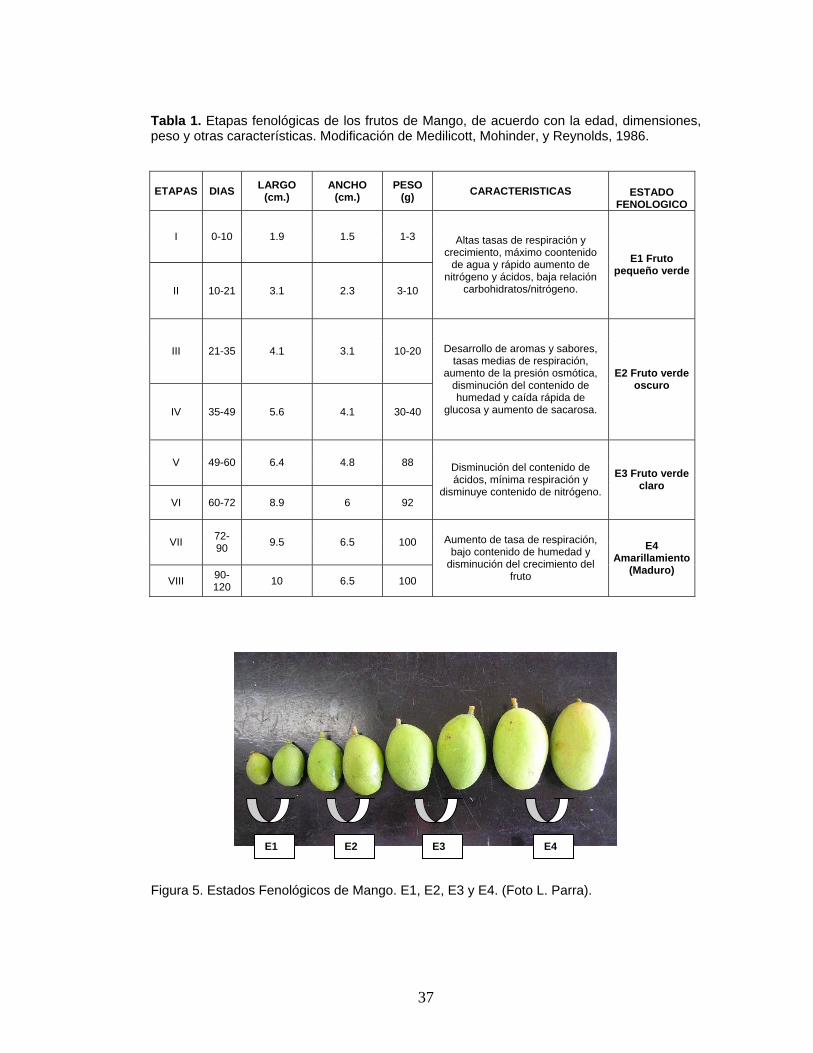
37
Tabla 1. Etapas fenológicas de los frutos de Mango, de acuerdo con la edad, dimensiones, peso y otras características. Modificación de Medilicott, Mohinder, y Reynolds, 1986.
ETAPAS DIAS LARGO (cm.)
ANCHO (cm.)
PESO (g) CARACTERISTICAS ESTADO
FENOLOGICO
I 0-10 1.9 1.5 1-3
II 10-21 3.1 2.3 3-10
Altas tasas de respiración y crecimiento, máximo coontenido
de agua y rápido aumento de nitrógeno y ácidos, baja relación
carbohidratos/nitrógeno.
E1 Fruto pequeño verde
III 21-35 4.1 3.1 10-20
IV 35-49 5.6 4.1 30-40
Desarrollo de aromas y sabores, tasas medias de respiración,
aumento de la presión osmótica, disminución del contenido de humedad y caída rápida de
glucosa y aumento de sacarosa.
E2 Fruto verde oscuro
V 49-60 6.4 4.8 88
VI 60-72 8.9 6 92
Disminución del contenido de ácidos, mínima respiración y
disminuye contenido de nitrógeno.
E3 Fruto verde claro
VII 72- 90 9.5 6.5 100
VIII 90-120 10 6.5 100
Aumento de tasa de respiración, bajo contenido de humedad y
disminución del crecimiento del fruto
E4 Amarillamiento
(Maduro)
Figura 5. Estados Fenológicos de Mango. E1, E2, E3 y E4. (Foto L. Parra).
E1 E2 E3 E4

38
5.3 Detección de infecciones quiescentes Los frutos recolectados fueron lavados en tres oportunidades con agua
corriente, luego se desinfestaron por un minuto con hipoclorito de sodio al
2.6% y se lavaron dos veces con agua destilada; por último se trataron con
alcohol al 70% y se lavaron nuevamente con agua destilada. Una vez secos
se llevaron a incubar en cámaras húmedas a temperatura ambiente.
(Cedeño, et al., 1993; Dhingra, et al., 1995).
Para el montaje de las cámaras se utilizaron contenedores plásticos
transparentes de 30 cm de ancho, 45 cm de largo y 17 cm de alto, con rejillas
plásticas previamente lavadas con detergente y agua, desinfestadas con
hipoclorito de sodio al 2.6% y asperjadas con alcohol antiséptico. (Anexo 3).
La presencia de las infecciones quiescentes se evaluó diariamente,
observando el tiempo de aparición de los primeros síntomas de la
enfermedad (Reyes, A,. 2007).
El diseño experimental que se utilizó en el ensayo fue un diseño de bloques
completos al azar BCA, con cuatro repeticiones y cuatro tratamientos
correspondientes a los estados fenológico. La unidad experimental estuvo
representada por una cámara húmeda con 7 frutos. Por cada tratamiento se
colectaron 28 frutos, para un total de 112 frutos.
5.4. Aislamientos de Colletotrichum sp.
A partir de los frutos que presentaron las lesiones características de la
enfermedad, con la ayuda de un bisturí estéril se tomaron fragmentos del
tejido enfermo de aproximadamente 5X5 mm, tratando de involucrar tejido
infectado (20%) y sano (80%); que posteriormente se sembraron en cajas de

39
Petri con (PDA) papa-dextrosa-agar (Anexo 2); se realizaron 2 repeticiones
por fruto, colocando cuatro fragmentos de tejido en la placa de agar.
(Cedeño, et al., 1993; Dhingra, et al., 1995).
Las cajas sembradas se incubaron a temperatura ambiente (± 20°C), durante
6 días, luego cada uno de los aislamientos obtenidos se le asignó un número
consecutivo y con el fin de purificar el hongo, se resembraron por separado
en placas de PDA; simultáneamente se realizaron observaciones
microscópicas coloreadas con azul de lactofenol para verificar las
características del hongo (Contreras, 2006). La taxonomía del hongo se
realizó con base a la clave para la identificación de hongos imperfectos de
Barnett (2003) y las características registradas por Bailey (1992).
5.4.1 Purificación de Aislamientos
Luego de confirmar que los aislamientos obtenidos pertenecían al hongo en
estudio, se realizaron cultivos monospóricos de acuerdo a una modificación
de la metodología propuesta por Wang-Ching y Wen-Huising (1997), para
obtener colonias puras (Anexo 4). Para este fin se llenaron con 1 ml de agua
destilada estéril 160 tubos de vidrio de 6x1.5 cm previamente lavados y
esterilizados. Con un asa estéril se tomó una pequeña porción de cada uno
de los aislamientos y se incorporó en los tubos, en donde con ayuda del
vortex se homogenizó la solución. Para calcular la concentración de 1
esporas/ml, se tomaron 10 µl de la solución y se sirvió en una cámara de
Nuebauer, en donde se contó el número de esporas localizadas en los 64
cuadros y posteriormente se estableció el volumen inicial para obtener una
concentración final de 1 espora/µl (Manual de procedimientos CORPOICA,
2001).

40
Se tomaron cajas con agar agua (Anexo 1) y en la base con un marcador de
vidrio se dividieron en 18 círculos aproximadamente de 7mm de diámetro
cada uno, para que una vez obtenidas las soluciones para cada aislamiento,
sembrar 1.7 µl de la solución en cada uno de los pozos. Las placas con agar
agua fueron incubadas por 24 horas a 27°C en penumbra. Transcurridas las
24 horas se observó bajo el microscopio (40X) la superficie de la placa de
agar agua buscando una única espora germinada, la cual se sembró en PDA
y se incubó a 27°C en penumbra, hasta obtener la colonia pura (Manual de
procedimientos CORPOICA, 2001).
5.5. Pruebas biológicas Las pruebas biológicas se realizaron con el objeto de establecer la etiología
del agente causante de las infecciones quiescentes observadas en los frutos
de Mango y su patogenicidad sobre frutos sanos.
5.5.1. Medio selectivo para Colletotrichum sp. El medio selectivo para Colletotrichum sp., se compone de Hidróxido de
Cobre (CuOH2) como ingrediente activo (42mg /L Cu) y 300 mg/L de
estreptomicina (Anexo 7). El CuOH2 es un fungistático que actúa
desnaturalizando las proteínas, a través de la reactivación de los grupos
sulfhídricos (Ware y Whitaker, 2000). Este medio permite una aproximación
para clasificar los aislamientos obtenidos y visualizar algunas características
que pueden diferenciar estas dos especies de Colletotrichum sp., dentro de
las mas evidentes se tiene la coloración predominante de la colonia y la tasa
de crecimiento (Abang, et al., 2003; Alvis, 2007).

41
Algunos investigadores plantean que la velocidad de crecimiento de la
colonia que se evalúa en este medio, puede ser rápida (fast) o lenta (slow) y
los colores predominantes de las cepas pueden ser naranja (orange), verde
(olive), gris (gray) y salmón (salmón). A partir de estos criterios de
evaluación se pueden obtener las clasificaciones: FGG (Fast growing gray),
SGG (Slow growing gray), FGS (Fast growing salmon), SGO (Slow growing
orange) y FGO (Fast growing olive) (Abang, et al; 2003).
Estas características se utilizan para diferenciar al patógeno, entre las
especies C. acutatum y C. gloeosporioides. Los criterios que permiten
clasificar a las cepas como C. gloeosporioides son: SGG, FGG, FGS y FGO
y la clasificación SGO orienta hacia la especie C. acutatum (Tabla 2).
Tabla 2. Clasificación de las especies según los medios de evaluación biológica. (Abang et al., 2003).
Medio Selectivo Medio Benomyl Clasificación
FGG Susceptible C. gloeosporioides
FGO Susceptible C. gloeosporioides
SGO Tolerante C. acutatum
SGG Susceptible C. gloeosporioides
FGS Susceptible C. gloeosporioides
5.5.2. Pruebas de Patogenicidad
Este ensayo tuvo como objetivo conocer la capacidad patogénica que tienen
los diferentes aislamientos, para generar síntomas de la enfermedad sobre
frutos sanos. Se emplearon dos frutos por aislamiento y dos se dejaron como

42
controles; en todos los casos se realizó el proceso de desinfestación de
frutos descrito anteriormente.
Posteriormente, sobre la parte media y superior de cada uno de los frutos se
les coloco un disco de agar con el aislamiento (Figura 6.), ya que mediante
las observaciones preliminares es la zona donde con mayor presencia se
presentan las infecciones quiescentes. Sobre los frutos usados como
controles se colocaron discos de agar estéril, luego de la inoculación se
colocaron en cámaras húmedas estas se mantuvieron a una temperatura de
± 18 C° por un periodo de 10 días y diariamente se revisaron para observar
el momento en que se evidenciaran los primeros síntomas de la enfermedad.
A partir de las primeras lesiones se realizaron los aislamientos del
microorganismo para comprobar los postulados de Koch. (Cedeño et al..
1993).
Figura 6. Frutos de Mango sanos. A. Fruto sin bloques de agar y B, Fruto con bloques de agar con micelio del microorganismo. (Foto, L. Parra).
A B

43
5.5.3. Prueba de susceptibilidad al Benomyl De acuerdo a las metodología que usaron (Péres, et al., 2005) para
identificar y conocer la susceptibilidad o la tolerancia que presentan las
especies de Colletotrichum sp. a este producto. Se utilizó la concentración
del fungicida Benomyl (PDA + Benomyl 2µg/ml), ya que las mínimas
concentraciones del fungicida fueron efectivas. Con base en la
susceptibilidad que presenta C. gloeosporiodes y la tolerancia de C.
acutatum. (Anexo 6) se diferenciaron las especies del patógeno. Para cada
aislamiento se realizaron dos repeticiones, una vez inoculadas las cajas se
incubaron por 72 horas en penumbra a una temperatura de 27°C.
En este medio se evaluó el diámetro de crecimiento de cada colonia
tomando como parámetro de comparación el diámetro de crecimiento que
presentó el hongo en el medio PDA. Esto se hace con el fin de determinar el
porcentaje de inhibición del microorganismo, al crecer en contacto con este
fungicida, respecto al control en PDA (Alvis, 2007).
5.5.4 Prueba de la Proteasa
Esta prueba ha sido utilizada para diferenciar las especies de de C. acutatum
y C. gloeosporioides. El medio hidrólisis de la caseína (MHC) (Anexo 5),
permite establecer dos tipos de reacción una positiva (+) y otra negativa (-).
El principio de esta prueba se basa en la producción extracelular de la
enzima proteasa (prueba +), característica de la especie C. acutatum, que
hidroliza la caseína de la leche, en aminoácidos y péptidos. Esta reacción
ocasiona que en el agar, normalmente opaco por la leche se observen halos
transparentes alrededor del crecimiento de la colonia del microorganismo
(Paterson y Bridge, 1994).

44
Una vez preparado el medio de la hidrólisis de la caseína (MHC) (Anexo 5) y
servido en las cajas de Petri estériles, se tomaron bloques de PDA con el
crecimiento micelial de cada aislamiento y se colocaron sobre la superficie
del medio MHC. Se realizaron 4 repeticiones por cada aislamiento (Martín &
García-Figueres, 1999); las lecturas se realizaron 24 horas después de
iniciada la prueba. Para asegurar que el medio funcionaba se tomo como
control positivo una cepa de C. acutatum (c-852) y un control negativo con
una cepa de C. gloeosporioides (c-1042) caracterizadas molecularmente, que
se encuentran en la Colección Nacional de Colletotrichum del Laboratorio de
Fitopatología de CORPOICA
5.6. Determinación del pH en frutos sanos y enfermos de Mango.
De acuerdo con Prusky, Keen (1993), la transición de una etapa biotrófica a
una necrotrófica en este grupo de microorganismos, parece estar relacionada
con factores que son modulados intracelularmente y son afectados por los
contenidos de nutrientes y el pH presente en el sustrato; por esta razón es
necesario conocer las variaciones de los niveles de pH en los diferentes
estados fenológicos de los frutos, para de esta manera comprender la
dinámica de las infecciones quiescentes en la medida que se va
desarrollando el fruto.
En este trabajo, se estudiaron las variaciones del pH, con el fin de comprobar
que este es un factor estrechamente relacionado con el establecimiento de
una relación parasítica patógeno-hospedero. (Prusky, 2001).

45
Para esta prueba se colectaron para cada estado fenológico, 4 frutos de
mango sanos y 4 enfermos, se lavaron tres veces con agua destilada (pH 7),
durante 10 minutos, luego se colocaron sobre papel periódico y se dejaron
secar a temperatura ambiente por un periodo de 30 minutos, para garantizar
un completo secado.
Luego con ayuda de un bisturí, se removieron 0.5 mm. del exocarpio de los
frutos, en los enfermos por antracnosis se separó el exocarpio y mesocarpio
enfermo del sano; para evitar alteraciones de las lecturas, se dejo una
distancia de 3 cm. entre el tejido sano del afectado. El exocarpio se maceró
en un mortero y una vez se obtuvo una pasta pura se realizó la lectura
directa del pH con ayuda de un potenciómetro de (Beckam).
Para determinar el pH del mesocarpio tanto de área enferma como sana, se
realizó un orificio de 2 cm de diámetro, en el cual se maceró el tejido
circunvecino con ayuda de un cuchillo estéril y una vez macerado el tejido se
introdujo el electrodo para la lectura del pH. En este caso los frutos
evaluados presentaron lesiones aceitosas de color negro, con bordes
definidos y centro deprimido.
Para establecer los valores de pH en frutos sanos de cada estado fenológico,
se realizaron cuatro lecturas del exocarpio y cuatro lecturas del mesocarpio;
en los frutos enfermos se hicieron cuatro lecturas tanto del exocarpio sano y
enfermo, como del mesocarpio sano y enfermo. Para en análisis estadístico
se empleo un diseño completamente al azar, evaluando un factorial de
4x3x2.

46
6. RESULTADOS Y DISCUSIÓN 6.1 Aislamientos de Colletotrichum sp.
En este estudio se obtuvieron varios aislamientos del hongo y se agruparon
según su color y crecimiento, obteniendo un total de 20 aislamientos (Anexo
10), que fueron clasificados dentro del género Colletotrichum de acuerdo a
las observaciones microscópicas realizadas, donde se identificaron las
estructuras del hongo (Figura 7). Los aislamientos se obtuvieron de
diferentes partes del fruto, confirmando lo encontrado en otros cultivos como
aguacate, guanábana, tomate de árbol, en donde el hongo fue aislado de
diferentes tejidos de la planta (Agostin, et al., 1992). Todos los aislamientos
tuvieron un buen crecimiento en PDA (Figura 8), lo cual concuerda con lo
propuesto por Agostin (1992), quien manifiesta que las especies de
Colletotrichum sp. no requieren de medios selectivos para su crecimiento.
Figura 7. Características microscópicas de C. gloeosporioides. A: Micelio y conidias del hongo formadas en PDA, B: Conidias típicas del microorganismo. (Foto L. Parra).
A B

47
El crecimiento de las colonias se presentó en forma radial y de aspecto
algodonoso, el color predominante fue el gris y el oliva lo que indica según
Contreras (2006) que pertenecen al género Colletotrichum sp. y a la especie
C. gloeosporioides (Figura 8); sin embargo se han demostrado incoherencias
entre las descripciones morfológicas y los estudios moleculares para la
identificación de especies de Colletotrichum sp. (Afanador, et al., 2003).
Figura 8. Aislamientos de Colletotrichum sp. En PDA. A y B Colonias obtenidas directamente a partir de frutos enfermos, C y D colonias obtenidas de cultivos monospóricos. (Foto L. Parra).
A
B
C
D

48
6.2 Detección de Infecciones Latentes en frutos asintomáticos de Mango
Transcurrido el periodo de incubación para cada estado fenológico, se
observó sobre la superficie de los frutos, la presencia de lesiones de color
negro y hundidas, características de antracnosis tal como lo afirma (Allende-
Molar y Cols, 2003) (Tabla 3.).
Tabla 3. Días Transcurridos hasta que aparecieron las lesiones en los Diferentes Estados Fenológicos de Frutos de Mango.
Estado Fenológico Tamaño de Muestra Periodo de Incubación (Días)
E1 28 10 E2 28 10
E3 28 15 E4 28 15
Se observaron infecciones quiescentes en todos los estados fenológicos del
fruto de mango; en los estado fenológico E1 y E2, el porcentaje de incidencia
fue superior al 75%, es decir que de un total de 112 frutos incubados en todo
el estudio, 84 frutos mostraron lesiones de infecciones quiescentes, mientras
que para los estados fenológicos E3 y E4 la incidencia de las infecciones
estuvo por encima del 60%. (Figura 9). Estos resultados ratifican la presencia
de infecciones quiescentes de C. gloeosporioides en frutos aparentemente
sanos, lo cual coincide con los resultados obtenidos por Arauz, (2000).
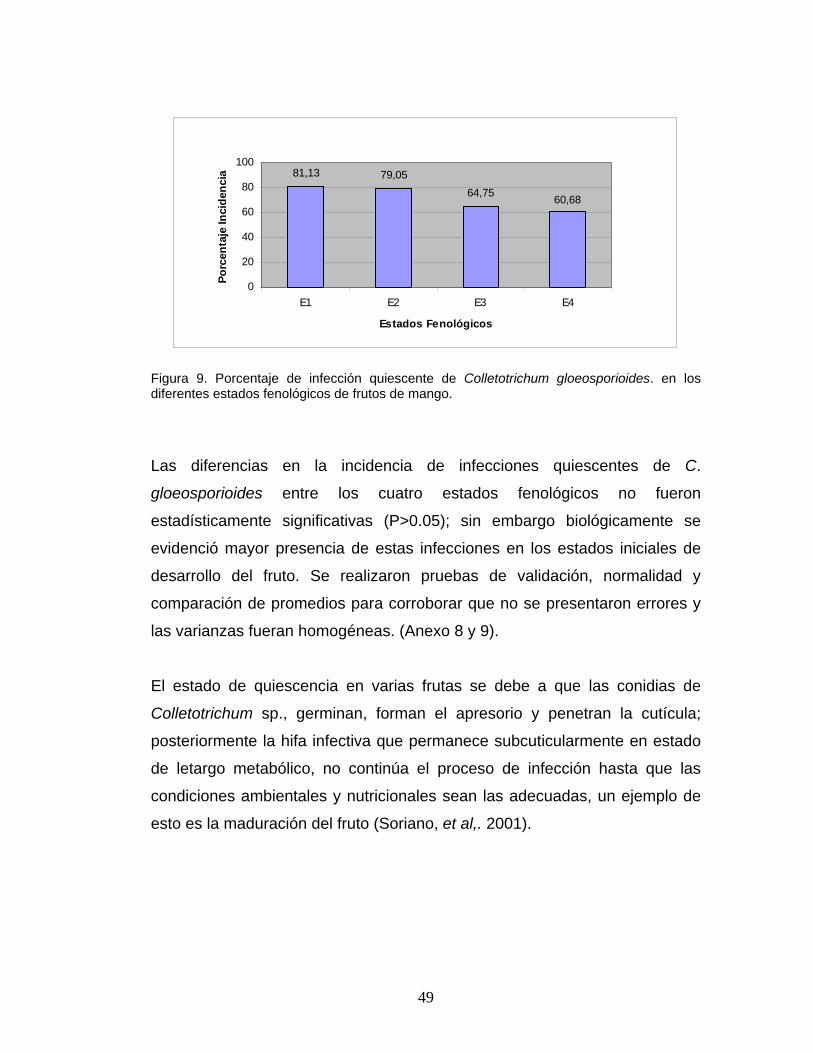
49
81,13 79,0564,75 60,68
0
20
40
60
80
100
E1 E2 E3 E4
Estados Fenológicos
Porc
enta
je In
cide
ncia
Figura 9. Porcentaje de infección quiescente de Colletotrichum gloeosporioides. en los diferentes estados fenológicos de frutos de mango.
Las diferencias en la incidencia de infecciones quiescentes de C.
gloeosporioides entre los cuatro estados fenológicos no fueron
estadísticamente significativas (P>0.05); sin embargo biológicamente se
evidenció mayor presencia de estas infecciones en los estados iniciales de
desarrollo del fruto. Se realizaron pruebas de validación, normalidad y
comparación de promedios para corroborar que no se presentaron errores y
las varianzas fueran homogéneas. (Anexo 8 y 9).
El estado de quiescencia en varias frutas se debe a que las conidias de
Colletotrichum sp., germinan, forman el apresorio y penetran la cutícula;
posteriormente la hifa infectiva que permanece subcuticularmente en estado
de letargo metabólico, no continúa el proceso de infección hasta que las
condiciones ambientales y nutricionales sean las adecuadas, un ejemplo de
esto es la maduración del fruto (Soriano, et al,. 2001).

50
Se podría sugerir que la fenología del fruto tiene un efecto en la incidencia de
las infecciones quiescentes, siendo los frutos maduros menos susceptibles a
adquirir estas infecciones. Cuatro mecanismos han sido postulados para
explicar la presencia de Colletotrichum sp. como infección quiescente en
frutos inmaduros: a) Requerimientos nutricionales del patógeno, b)
Compuestos antifungales preformados presentes en frutos verdes que
disminuyen a medida que el fruto madura por un aumento en la actividad de
la enzima lipoxigenasa, cuya actividad es regulada por la presencia de
flavan-3-ol, epicatequin (compuesto fenológico). Estos compuestos son el 1-
acetoxi-2-hidroxi-4-oxo-henicosa-12, 15-dieno y el 1-acetoxi-2,4-dihidroxi-n-
heptadeca-16-eno., c) Presencia de fitoalexinas y compuestos preformados y
d) Factores de patogenicidad que pueden ser activados solamente en frutos
maduros. (Prusky, et al,. 1996).
Para que el hongo pueda penetrar la cutícula del fruto requiere de un
complejo enzimático que le permita macerar la cutícula y penetrar en
exocarpio. Según Wattad y Prusky (1994), la secreción de enzimas
pectolíticas responsables de la maceración de la pared celular durante el
ataque de C. gloeosporioides, es un factor limitante durante la patogénesis y
las condiciones que afectan la secreción de estas enzimas, pueden evitar la
activación del Colletotrichum sp. en la colonización de frutos inmaduros.
Otro factor que determina la relación parasítica hospedero-patogeno es el pH
del hospedero, que puede ser concluyente en la dinámica de las infecciones
quiescentes en los diferentes estados fenológicos de los frutos de mango.
Yakoby et al,. (2000), sugieren que la secreción de pectinasas están
influenciadas por el pH, indicando que la susceptibilidad de los frutos está
regulada por decrecimientos de compuestos antifungales e incrementos en el
pH del pericarpio que modula la secreción de pectinasas y otras enzimas. La

51
variación en los niveles de pH explica el comportamiento de las infecciones
quiescentes en frutos de mango. (Reyes, 2007).
Las pectinasas de C. gloeosporioides son secretadas con valores de pH
mayores a 5.8, aunque también puede ser producida con valores de 5.1
(Prusky, et al,. 2001), lo que indica que cuando los niveles de pH del
hospedero son adecuados se puede iniciar el proceso de infección. A medida
que maduran los frutos de mango se van reduciendo los niveles de pH, lo
que sugiere que los inicios del proceso de infección ocurren en los primeros
estados de desarrollo del fruto.
La temperatura promedio semanal durante los muestreos fluctuó entre 27°C
y 28°C, las cuales fueron favorables para el establecimiento de las
infecciones quiescentes, según Arauz, (2000), temperaturas entre 20°C y
30°C, han sido reportadas como óptimas para la infección de C.
gloeosporioides. El hongo requiere una humedad relativa superior a 95%
para que las conidias germinen y se presente la formación del apresorio
(Arauz, 2000). En este estudio la humedad relativa durante los muestreos,
estuvo entre 67% y 79%.
6.3 Medio Selectivo para Colletotrichum sp. En esta prueba se obtuvieron colonias con crecimiento rápido y coloración
gris y verde oliva (Anexo 13). Este medio permite clasificar los diferentes
aislamientos, entre estas dos especies de Colletotrichum sp., por medio de la
observación de la coloración que predomina en la colonia y la tasa de
crecimiento (Abang, et al., 2003; Alvis, 2007). El CuOH2 es un fungistático
que actúa desnaturalizando las proteínas, a través de la reactivación de los
grupos sulfhídricos (Ware y Whitaker, 2000).
C
D
CD

52
Las diferentes clasificaciones se utilizan para diferenciar al patógeno entre
las especies C. acutatum y C. gloeosporioides. Los criterios que permiten
clasificar a las cepas como C. gloeosporioides son: SGG, FGG, FGS y FGO
y la clasificación SGO orienta hacia la especie C. acutatum (Figura 10).
Figura 10. Desarrollo de las colonias de Colletotrichum en los diferentes medios de cultivo: A, Crecimiento sobre PDA, B, Medio selectivo para Colletotrichum sp.(CuOH2) y C, Medio Benomyl (2µl/ml) (Foto L. Parra).
6.4 Prueba de Benomyl En esta prueba que permite diferenciar especies de C. acutatum y C.
gloeosporioides (Freeman, et al,.1998; Bernstein, et al,. 1995), se encontró
que todos los aislamientos fueron susceptibles al Benomyl (Anexo 11), por lo
tanto se considera que pertenecen a la especie C. gloeosporioides (Figura
10).
Estos resultados son similares a encontrados por Cerón et al (2006) en
trabajos de investigación realizados con aislamientos de las dos especies de
Colletotrichum que afectan los cultivos de lulo (Solanum quitoense L.).
A B C

53
El mecanismo de acción de los benzimidasoles consiste en inhibir el proceso
mitótico fungoso, bloqueando al mismo tiempo la síntesis de ácido
desoxirribonucleico (De la Isla, 1994). Los fungicidas sistémicos como el
benomyl son convertidos en las plantas al compuesto metil benzimidazol
carbamato, el cual interfiere en la división nuclear de los hongos que son
sensibles a sus componentes (Agrios, 2002).
En nuestro país dentro de las prácticas de manejo de la antracnosis, se
utiliza con frecuencia el producto, el cual sólo es efectivo en el control de la
enfermedad cuando el agente causal es C. gloeosporioides (Páez, 2006), por
lo cual es necesario que en el país se inicie su manejo mediante diferentes
alternativas de control. 6.5 Prueba de la Proteasa
En las lecturas realizadas en el medio MHC se observó que todos los
aislamientos presentaron una reacción negativa (-), que corresponden a C.
glooesporioides (Anexo 12). Esta prueba ha mostrado ser una forma rápida y
útil para diferenciar entre especies de C. gloeosporioides y C. acutatum
(Martín & García-Figueres, 1999). La respuesta positiva consistió en que en
el medio se observó la formación de un halo transparente alrededor del sitio
de contacto con el aislamiento del hongo y cuando no se observó algún
cambio en el medio, la reacción se tomo como negativa (Figura 11).

54
Figura 11. Prueba de la Proteasa. A. Respuesta negativa sin halo transparente característica, de la especie gloeosporiodes. B. Respuesta con halo transparente, típica de acutatum. (Foto, L. Parra).
Los hongos patógenos de plantas utilizan diferentes enzimas capaces de
destruir los componentes estructurales de los tejidos vegetales ocasionando
la muerte celular. Estas enzimas degradan carbohidratos que disuelven la
pared celular e hidrolizan la cutícula. Existen otras enzimas que degradan la
pared celular como las pectinliasas y proteasas que son consideradas
indispensables para la colonización de las plantas por parte del patógeno
(Bailey, et al,. 1992). En el caso de las proteasas, cuando el patógeno
secreta estas enzimas, inactiva los componentes proteínicos de respuesta de
defensa de las plantas tales como las quitinas y la ß- 1,3 glucanasa. (Cerón,
2004).
A B

55
6.6 Pruebas de Patogenicidad
De los frutos evaluados cuando se emplearon discos de agar con el
crecimiento del microorganismo, el 100% mostraron los síntomas
característicos de antracnosis. Los primeros síntomas se identificaron como
puntos superficiales necróticos de color café oscuro y negro, que
posteriormente ocasionaron el hundimiento en la zona central, como se
puede observar en la Figura 12 (A y B). El reaislamiento del patógeno a
partir de los frutos con síntomas confirmó la presencia del mismo
comprobando los postulados de Koch.
En los aislamientos caracterizados como C. gloeosporioides según las
pruebas de Benomyl, proteasa y medio selectivo para Colletotrichum spp.
causaron en los frutos los síntomas inicialies de antracnosis a los 4 días en
su gran mayoría y algunos con 7, 5 y 9 días. (Anexo 14)
La humedad de los tejidos permite que la conidia germine, forme apresorio y
se desarrolle la enfermedad, condición que al igual que en otras especies de
frutos favorece el desarrollo del microorganismo. (Arauz, L. F. 2000).
Figura 12. Frutos de Mango con lesiones de antracnosis. A, Lesiones primarias y B, Lesiones avanzadas. (Foto, L. Parra).
A B

56
6.7 Determinación de pH en frutos sanos y enfermos de Mango
Según Lakshminarayana et al (1970) los frutos de mango en sus primeros
estados son ácidos y altamente astringentes, con mayor desarrollo presentan
un contenido alto de polifenol y conservan la astringencia, sin embargo
alrededor de las ocho semanas presenta una reducción de polifenoles y
luego permanece estable. El ácido ascórbico aumenta rápidamente
alcanzando su máximo a las cinco semanas, declina a las ocho semanas y
permanece estable hasta cosecha, pero al colocar en condiciones de
maduración frutos cosechados con seis, ocho, once y trece semanas de
desarrollo, se encontró que con mayor tiempo se tornaban altamente ácidos.
(Lakshminarayana et al 1970).
Coincidiendo con los resultados obtenidos en este estudio, ya que mostraron
que a medida que el fruto madura, los niveles de pH tanto en el exocarpio
como en el mesocarpio, disminuyen haciéndose mas ácidos (Tabla 4.). Al
comparar estos resultados, con los obtenidos por Lagos et al (1989), en un
estudio sobre mango de la variedad Kent, encontramos que son similares,
indicando que los valores de pH disminuyen en los frutos maduros.
Tabla 4. Valores de pH de Frutos Sanos de Mango en los diferentes Estados Fenológicos, en Exocarpio y en Mesocarpio.
FRUTOS SANOS pH Exocarpio pH Mesocarpio FRUTOS E1 E2 E3 E4 E1 E2 E3 E4
1 5,20 5,00 5,10 4,20 5,12 4,95 4,32 4,12 2 5,40 5,20 4,60 3,50 4,91 4,63 4,45 3,95 3 5,80 5,00 4,50 3,70 5,11 4,78 4,23 3,74 4 6,00 5,50 5,00 3,30 4,52 4,45 4,12 3,35
Media 5,6 5,2 4,8 3,7 4,9 4,7 4,3 3,8
57
Algunos factores como el pH juegan un papel importante en el desarrollo de
la enfermedad, ya que sus variaciones de acuerdo con la fenología del fruto
sirve como una barrera y un mecanismo de defensa contra patógenos. De
esta manera el patógeno supera la barrera del exocarpio y las esporas
germinadas melanizan el apresorio, no solo por la reducción del pH que se
observa en el mesocarpio (Tabla 4), sino por la alta concentración de
compuestos antifungosos presentes en los frutos inmaduros (Prusky, et al
2001). Esta condición de quiescencia permanece hasta la maduración del
fruto, donde los contenidos nutricionales y su fisiología le permiten al
patógeno activar sus mecanismos de patogenicidad.
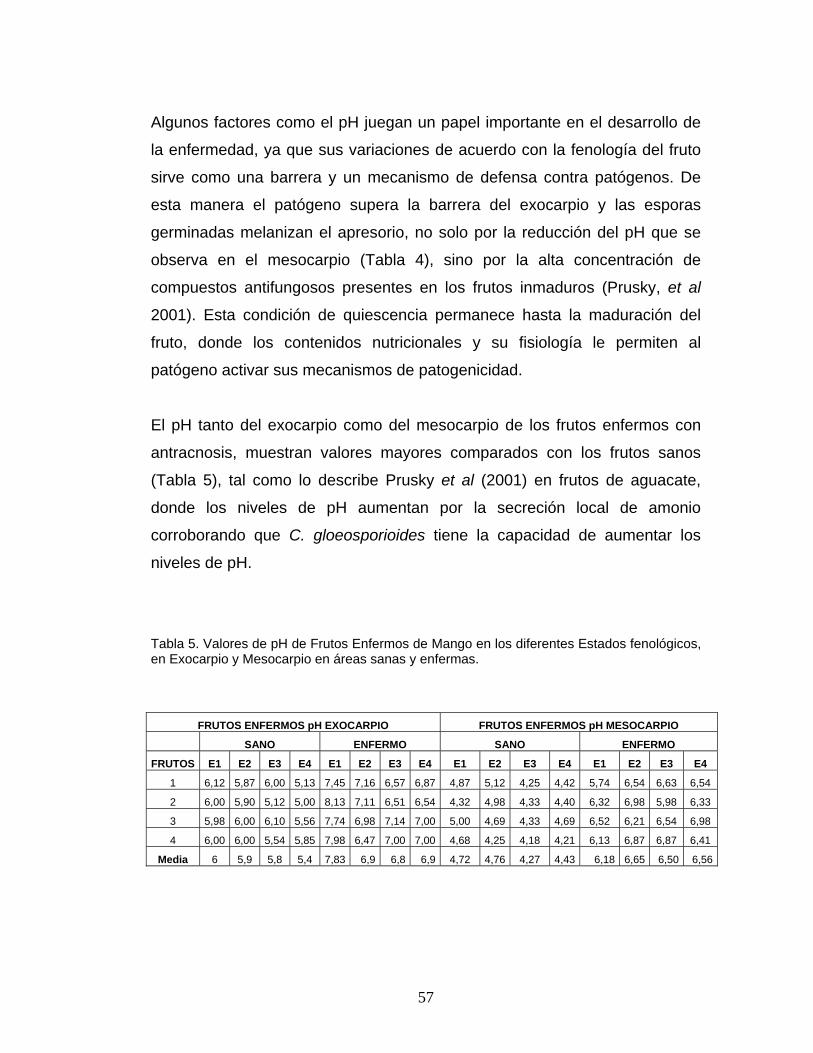
El pH tanto del exocarpio como del mesocarpio de los frutos enfermos con
antracnosis, muestran valores mayores comparados con los frutos sanos
(Tabla 5), tal como lo describe Prusky et al (2001) en frutos de aguacate,
donde los niveles de pH aumentan por la secreción local de amonio
corroborando que C. gloeosporioides tiene la capacidad de aumentar los
niveles de pH.
Tabla 5. Valores de pH de Frutos Enfermos de Mango en los diferentes Estados fenológicos, en Exocarpio y Mesocarpio en áreas sanas y enfermas.
FRUTOS ENFERMOS pH EXOCARPIO FRUTOS ENFERMOS pH MESOCARPIO
SANO ENFERMO SANO ENFERMO
FRUTOS E1 E2 E3 E4 E1 E2 E3 E4 E1 E2 E3 E4 E1 E2 E3 E4
1 6,12 5,87 6,00 5,13 7,45 7,16 6,57 6,87 4,87 5,12 4,25 4,42 5,74 6,54 6,63 6,54
2 6,00 5,90 5,12 5,00 8,13 7,11 6,51 6,54 4,32 4,98 4,33 4,40 6,32 6,98 5,98 6,33
3 5,98 6,00 6,10 5,56 7,74 6,98 7,14 7,00 5,00 4,69 4,33 4,69 6,52 6,21 6,54 6,98
4 6,00 6,00 5,54 5,85 7,98 6,47 7,00 7,00 4,68 4,25 4,18 4,21 6,13 6,87 6,87 6,41
Media 6 5,9 5,8 5,4 7,83 6,9 6,8 6,9 4,72 4,76 4,27 4,43 6,18 6,65 6,50 6,56

58
Con los resultados obtenidos tanto para el diseño factorial como en el diseño
de bloques al azar, hubo diferencia significativa mínima (Anexo 15 y 16),
entre el estado fenológico y el área donde se midió el pH (p<0.05) y
diferencia significativa entre el área donde se midió el pH y el tejido
(Mesocarpio y Exocarpio). En la figura 13 A. se observa el comportamiento
de los valores de pH en frutos sanos y enfermos en áreas sanas y enfermas,
tanto en exocarpio como en mesocarpio de frutos enfermos.
Los resultados muestran que el pH de frutos enfermos es superior a 6.0, y el
valor mínimo para que se active la secreción de pectinasas y por ende la
degradación de la cutícula, es de mínimo 5.1 (Yakoby et al 1999) situación
que puede estar influyendo en la actividad del microorganismo. También se
puede observar que los valores de pH del exocarpio en áreas sanas de frutos
enfermos, se ve alterada por la presencia de C. gloeosporioides aumentando
el valor de pH con respecto al de un fruto completamente sano sin haber
diferencia estadísticamente significativa (p>0.05); en el mesocarpio esta
tendencia no se ve tan marcada. Las variaciones en los valores del pH en
áreas sanas con respecto a enfermas, indican que el patógeno necesita
alterar el pH para poder superar barreras fisiológicas impuestas por el
hospedero (Prusky, et al. 2001).
En la figura 13 B-C. se observa que el pH aumenta a medida que el fruto se
enferma y que dependiendo del estado de desarrollo del fruto, existe una
interacción entre el valor del pH y el área donde se realiza la evaluación; es
decir que en frutos sanos los valores de pH del exocarpio y mesocarpio son
diferentes (p<0.05) y cuando el fruto está enfermo los valores de pH
cambian, siendo iguales en mesocarpio y exocarpio (p>0.05). En áreas
sanas de frutos enfermos los valores de pH se comportan igual que en frutos
sanos.

59
B
A
A

60
Figura 13. Efecto de Antracnosis sobre el pH. A, Te: tejido enfermo, ts: tejido sano. Ts-fe: tejido sano en fruto enfermo, exo: exocarpio y meso: mesocarpio. B, efecto sobre el pH en mesocarpio y exocarpio. C, efecto sobre tejido tomado en el fruto. Te: tejido enfermo, ts-fe: tejido sano en fruto enfermo y ts: tejido sano.
Con lo resultados obtenidos en este estudio se puede decir que a medida
que el fruto se desarrolla los valores de pH disminuyen dificultando la
penetración del patógeno durante la maduración del fruto; por lo tanto se
sugiere que los síntomas observados en los estados fenológicos E3 y E4 son
el resultado de infecciones quiescentes que se formaron cuando el fruto se
encontraba en los estados fenológicos E1 y E2.
C

61
La baja presencia de infecciones quiescentes en estados de desarrollo
avanzado del fruto, puede atribuirse a factores como, uso de fungicidas
sistémicos que pueden afectar al hongo, y el lavado natural del inóculo por
encontrarse expuesto mayor tiempo a condiciones de campo. Además como
en frutos maduros los niveles de pH son bajos, el patógeno no puede iniciar
la enfermedad y su actividad se ve limitada a las infecciones quiescentes o
latentes que persistieron a las condiciones adversas.

62
7. CONCLUSIONES
La incidencia de infecciones quiescentes en frutos de mango variedad
hilacha fue mayor en estados de desarrollo iniciales, y el periodo de
incubación de las infecciones quiescentes fue en los estados E1 y E2 de 10
días y en los estados E3 y E4 fue de 15 días.
El color de las colonias obtenidas de los aislamientos en PDA fue de un
aspecto característico reportados para el género Colletotrichum.
Los sistemas tradicionales de clasificación con base en las características
morfológicas, deben complementarse con otras pruebas para poder
determinar la especie dentro del género.
Las pruebas de la proteasa, sensibilidad al benomyl y crecimiento en medio
selectivo para Colletotrichum sp. permitieron establecer que los aislamientos
obtenidos de las infecciones quiescentes en los diferentes estados
fenológicos correspondieron a la especie C. gloeosporioides.
Mediante las pruebas de patogenicidad se comprobó que los 20 aislamientos
fueron patogénicos en frutos de mango variedad hilacha y su presencia se
determinó mediante la observación de la expresión de síntomas sobre los
frutos.
A medida que el fruto se va desarrollando se disminuyen los niveles de pH,
dificultando la penetración del patógeno, razón por la cual se observa una
abundante presencia de infecciones quiescentes en frutos pocos
desarrollados, comparados con frutos de fenología avanzada.

63
El efecto que ejerce C. gloeosporioides sobre el pH es un factor de
patogenicidad que debe ser tenido en cuenta a la hora de realizar un control
y manejo sobre la antracnosis.

64
8. RECOMENDACIONES
Se debe investigar la incidencia de infecciones quiescentes sobre otros
tejidos o estructuras de la planta para saber si el microorganismo sobrevive
de forma quiescente ocasionando infecciones tempranas en frutos. También
se debe evaluar durante los doce meses del año para observar el tiempo en
donde ocurren los mayores porcentajes de enfermedad.
Realizar investigaciones relacionadas con el pH, ya que puede ser un factor
de patogenicidad importante para el control de la antracnosis.
Realizar investigaciones donde se conozcan las propiedades químicas y
nutricionales de los frutos de mango a medida que se desarrollan, para
identificar factores que puedan controlar el desarrollo de infecciones por
parte de C. gloeosporioides.

65
9. REFERENCIAS Abang, M., Winter, S., Mignouna, H., Green, K., Asiedu, R, 2003. Molecular
taxonomic, epidemiological and population genetic approaches to
understanding yam anthracnose disease. African Journal of Biothechnology
Vol. 2 (12): 486 – 496. Adaskaveg, J & Hartin, R. 1997. Characterization of Colletotrichum acutatum
causing annthracnose of almond and peach in California. Phytopatology 87
(9): 979-987. Afanador, K. L., Minz, D., Maymon, M & Freeman, S. 2003. Characterization
of Colletotrichum isolates from Tamarillo, Passiflora, and Mango in Colombia
and Identification of a Unique Species from de Genus. The American
Phytopathologycal Society. 93 (5): 579-587. Agostin, J. P, Timer, L. W., y Michell, D. J. 1992. Morphological and
pathological characteristics of strains of Colletotrichum gloesporioides from
citrus. Phytopathology 82: 1377-1382.
Agrios, G.N. 2002. Fitopatología. Editorial Limusa S.A de C.V. México D.F.,
México. 838p. Allende-Molar, R., García, R. S. y Carrillo, CIAD-Culiacán. 2003. Antracnosis
causada por el hongo Colletotrichum gloesporioides. Álvarez, J.M. 1996. Manejo integrado de la antracnosis (Colletotrichum
gloesporioides Penz.) en el cultivo de tomate de árbol (Cyphomandra
betacea) en el municipio de Manizales. Tesis Facultad de Ciencias
Agropecuarias, Universidad de Caldas. Pp 15-25.

66
Alvis, J. 2007. Evaluación de pruebas biológicas para la identificación de
especies de Colletotrichum sp. aislados de diferentes especies frutales. Tesis
de pregrado. Facultad de Ciencias de la Salud. Programa de Bacteriología.
Universidad Colegio Mayor de Cundinamarca. Bogotá, Colombia. Pp. 40. Arauz, L.F. 2000. Mango Antrhacnose: Economyc Impact and current options
for interfrated management. Plant Dis. (84): 600-611. Bailey J. A, Jeger M. J, 1992. Colletotrichum: Biology, pathology and control.
Wallingford, UK: CAB International and control of Colletotrichum species on
tropical fruit crops. Barnet, H.L y Hunter, B.B. 1998. Illustrated genera of imperfect fungi. Cuarta
edición. Minesota. APS Press. 218p. Bernstein, B., Zehr, E. I., Dean, R. A., and Shabi, E. 1995. Characteristics of
Colletotrichum from peach, apple, pecan, and others host. Plant Dis. (79):
478-482. CABI-EPPO (1996). Data Sheets on Quarantine Pest. Colletotrichum
acutatum . Bulletin CABI-EPPO 90-399003. Cartagena, J., Vega, D.2001. Mango. Manual de asistencia técnica Nº 43.
ICA. Bogotá. Colombia. Cedeño, L., Mohali, S., y Palacios Prü, E. 1993. Antracnosis causada por dos
cepas de Glomerella congulata en frutos de parchita. Fitopatol. Venez. (6):
30-33.

67
Cerón, S. 2004. Detección de Colletotrichum (Penz) Sacc en estructuras
vegetativas y reproductivas de Lulo Solanum quitoense Lam. Trabajo de
grado presentado como requisito para la maestría en Ciencias Agrarias con
énfasis en Fitopatología. Universidad Nacional de Colombia. Contreras, C., 2006. Caracterización y pruebas de patogenicidad cruzada
entre aislamientos de Colletotrichum spp. Obtenidos de frutos de lulo
(Solanum quitoense Lam), tomate de árbol (Solanum betacea Sendt),
granadilla (Passiflora ligularis Juss), mango (Magnifera indica L) y tallos de
mora (rubís glaucus Benth) con síntomas de antracnosis. Trabajo de grado
presentado como requisito para la carrera de Microbiología Agrícola y
Veterinaria. Pontificia Universidad Javeriana. De Lapyere, L.; Chillet, M.; Dubois, C.; Mourichon, X. 2000. Importance of
different spurces of inoculum and dispersal methods of conidia of
Colletotrichum musae, the causal agent of banana anthracnose, for fruit
contamination. Plant Pathology. (49): 782-790.
De la Isla, L. 1994. Fitopatología. Segunda reimpresión. Noriega Editores.
México, D.F. 384p. Dhyngra, O.D., Sinclair. B.J. 1995. Basic Plant Pathology Methods. CRC.
Press. 434 p. Fitzel, R.D., Peak, C.M. 1984. The epidemiology of anthracnose disease of
mango: inoculum source, spore production and dispersal. Ann. Appl. Biol.
(104): 53-59.

68
Flaishman, M. A., and Kolattukudy, P. E. 1994. Timing of fungal invasion
using host’s ripening hormone as a signal. Proc. Natl. Acad. Sci. USA
91:6579-6583. Forero de la Rotta, M. 2001. Enfermedades de la mora de castilla. Boletín de
Sanidad Vegetal. ICA, Bogotá. Pp. 34-36.
Freeman, S. and Shabi, E. 1996. Croos-infection of subtropical and
temperate fruit by Colletotrichum species from various hosts. Physiol. Mol.
Plant patho. (49): 395-404. Freeman, S., Katan, T and Shabi, E. 1998. Characterization of Colletotrichum
spp. Species responsible for antrachnose disease of various fruit. Plant Dis.
(82): 596-605. Holliday, P. 1995. Fungus diseases of tropical fruits. Dovel Publication, Inc.
New York. 624p. ICA. 1996. Plagas y enfermedades en frutas tropicales. División de Sanidad
Vegetal. Boletín de Sanidad Vegetal Nº 11. Santa fe de Bogotá. ICONTEC. 2002. Norma técnica colombiana (NTC 5139): Frutas frescas.
Mango criollos. Especificaciones. Instituto de Hidrología, Meteorología y Estudios Ambientales IDEAM. 2006.
Valores medios diarios de temperatura año 2004. Jarvis, W. 1994. Latent infections in the pre and postharvest environment.
Hort Science 29: 749 – 750.

69
Lagos, R. E y Kairus de Civetta, L. A. 1989. Crecimiento del fruto y momento
óptimo de cosecha en mangos Tommy Atkins y Kent. Trabajo de
investigación llevado a cabo como requisito para optar al título de Magíster
en Fitotécnica. Universidad Nacional de Colombia, sede Medellín y Bogotá.
Agricultura Tropical 28: 51-68.
Lakshminarayana, S., Subhadra, N. U. and Subramanyam, H. 1970. Some
aspects of development physiology of mango fruit. J. Hort. Sci., 48: 133-142. León, G. 1968. Fundamentos botánicos de los cultivos tropicales. Instituto
Interamericano de Ciencias Agrícolas de la OEA. San José, Costa Rica.
Manual de Procedimientos, CORPOICA. 2001. Obtención de Cultivos
monospóricos (Adaptada de la metodología propuesta por HO, W.C y KO,
W.H., 1996). Martín MP, García-Figueres F, 1999. Colletotrichu acutatum and
Colletotrichum gloesporioides cause anthracnose on olives. European
Journal of Plant Pathology 105 (8): 733-41.
Mena, G. 1999. Epidemiología y control de la antracnosis (Colletotrichum
gloesporioides Penz) en mango. México.
Medlicott, A.P., Mohinder, B. and Reynolds, S.B. 1986. Changes in peel
pigmentation during ripening of mango fruit. Am. Appl. Biol. 109: 651-656. Ministerio de Agricultura y Desarrollo Rural. 2005. Área, producción y
rendimiento del Mango. Colombia.

70
O´connell, S., Perfect, S., Hughes, B., Carzaniga, R., Bailey, J. & Green, J.
2000. Dissecting the Cell Biology of Colletotrichum Infecting Processes. pp
57-77 In: Colletotrichum : Host Specificity, Pathology, and Host-Pathogen
Interaction. D. Prusky. S. Freeman and M.B. Dickman, eds. APS Press. St.
Paul, Minnesota.
Páez, A. 2006. Tecnologías sostenibles para el manejo de la antracnosis en
papaya y mango. Boletín Técnico Nº 8. CORPOICA. Valledupar. Paterson RRM, Bridge PD, 1994. Biochemical Tecniques for Filamentous
Fungi. IMI Technical Handbooks Nº. 1. Wallingford, UK: CAB International. Peres, N. A. R., Timmer, L. W., Adaskaveg, J. E., Correll, J. C, 2005.
Lifestyles of Colletotrichum acutatum. Plant disease. 89 (8): 784 – 795. Ploetz, R.C. and Prakash, O. 1997. Foliar, Floral and Soliborne disease. In:
The Mango: Botany, Production and uses. Litz, R.E. (Ed.), CAB International.
Oxon, UK. 281-325p. Prusky, D. 1996. Pathogen quiescence in post-harvest disease. Annu. Rev.
Phytopatology. (34): 413-434. Prusky, D; and N. T. Keen. 1993. Involvement of preformed antifungal
compounds in the resistance of subtropical fruit decay. Plant Dis. (77): 114-
119.
Prusky, D; McEvoy, J.L; Conway, WS. 2001. Local modulation of Host pH by
Colletotrichum species as a mechanism to increase virulence. MPMI Vol. 14.
No. 19. 1105-1112.

71
Reyes, A,. 2007. Estudio de la relación fenológica reproductiva y el
comportamiento de las infecciones latentes de Colletotrichum acutatum en
tomate de árbol (Solanum betacea) (Cav) Sendt. Trabajo de grado
presentado para obtener el titulo de Ingeniera Agrónoma. Universidad
Nacional de Colombia. Rondón, G.J. 1998. Aspectos fisiológicos del tomate de árbol, epidemiología
de la antracnosis y su relación con el manejo del problema. En 2: Seminario
Frutales de Clima Frío Moderado. Memorias.
Rondón, G.J. 2003. Estudio Biológico y epidemiológico de la antracnosis
(Colletotrichum gloesporioides penz), del tomate de árbol (Solanum betacea,
(cav) sendt), y generación de alternativas para su manejo integrado en
Colombia. Informe técnico. MIP. CORPOICA. Saavedra, L., Fuentes, G., Martínez, C., López, E., Duveiller, R., Henry, M. y
Garcia, A. (2005). Guía práctica para la identificación de algunas
enfermedades de trigo y cebada. Segunda edición. Centro Internacional de
Mejoramiento de Maíz y Trigo (CIMMYT).Pág. 10- 11. Salazar, C., R. 1982. Propagación de frutales. Fruticultura tropical.
Federación Nacional de Cafeteros. 78-90. SENA, 2006. Boletín sobre el cultivo de mango. pp. 21-25.
Soriano, E.C; Colinas, M.T; Zamora, T y Cajuste, J.F. 2001.Anatomia del
daño por rozamiento y por Colletotrichum gloesporioides PENZ en frutos de
aguacate "HASS". Agrociencia Vol. 35 Nº. 002. 237-244.

72
Swimburne, T. R. 1983. Quiescent infections in post-harvest diseases. In:
Dennis, C. (ed.), Post-harvest Pathology of fruits and vegetables, Academic
Press, London, pp. 1-21. Waller, J.M. 1992. Colletotrichum diseases of perennial and other cash crops.
En: Colletotrichum: biology, pathology and control. Bailey, Jy Jeger, M., (eds)
CBA internacional pp. 167-170, 176-9. Wang, C.H. and Wen, H.K. 1997. A simple method for obtaining single-spore
isolates of fungi. Bot. Bull. Acad. Sin. (38): 41-44. Ware, Whitaker, G. 2000. Pesticide book. Freeman and company. Wattad, C; A y Prusky, D. 1994. Purification of pectate lyase produced by
Colletotrichum gloesporioides and its inhibition by epicatechin: A possible
faxtor involved in the resistance of unripe avocado fruits to anthracnose. Mol.
Plant Microbre Interact. 7: 293-297.
Yakoby, N; Kobiler, I; Dinoor, A; Prusky, D. 2000. pH regulation of pectate
lyase secretion modulates the attack of Colletotrichum gloesporioides on
avocado fruit. Apple. Environ. Microbial. 66: 1026-1030. Yerhoeff, K. 1974. Latent infections by fungi. Phytopathol. (12): 99-110. Zulfiqar, M., Bransky, R.H, and Timmer, L.W. 1996. Infection of flower and
vegetative tissues of citrus by Colletotrichum acutatum and Colletotrichum
gloesporioides. Mycologia. (88): 121-128.

73
ANEXOS
ANEXO 1. Preparación del AGAR AGUA
Agar-Agar 15g
Agua Destilada 1000 ml
Esterilizar a 121°C a 15 lb por 15 minutos

74
ANEXO 2. Preparación de agar PDA
*Infusión de Papa 200g
Dextrosa 10g
Agar-Agar 20g
Ajustar el pH a 5.6
*Infusión de papa: preparada a partir de 200g de papa por litro.

75
ANEXO 3. Protocolo de Cámaras Húmedas
1. Objetivo
Crear condiciones favorables de humedad para el rápido desarrollo de
hongos que pueden estar involucrados en la producción de síntomas de
enfermedad, pero cuya presencia no es detectada en el momento de la
primera observación.
2. Alcance:
Este procedimiento se aplica para obtener crecimiento tanto vegetativo como
reproductivo de los microorganismos para su posterior análisis
3. Materiales y Equipos:
− Material vegetal
− Hipoclorito 1%
− Alcohol antiséptico 1%
− Bisturí
− Portaobjeto estéril
− Cinta scotch
− Toalla de papel
− Cajas de petri
− Recipiente plástico desinfectado
− Agua estéril
− Parafilm
− Cuarto de crecimiento

76
4. Contenido
Seleccionar y tomar los frutos a estudiar, esterilizar los frutos con hipoclorito
al 1% durante 30 a 60 segundos; lavarlos tres veces con agua destilada
estéril; sumergirlos en alcohol durante 30 a 60 segundos y lavarlos tres veces
con agua destilada estéril para eliminar el exceso de hipoclorito y secar el
material vegetal sobre toalla de papel. Se recomienda escoger tejido que no
esté demasiado dañado o con lesiones muy avanzadas para evitar la
presencia de saprófitos.
Agregar al recipiente 600ml de agua destilada estéril y colocar una toalla de
papel de modo tal que no quede en contacto directo con el agua para evitar
pudriciones de tejido.
Colocar el material vegetal sobre la toalla y tapar la cámara sellándola con
vinipel para permeabilizar la cámara.
Incubar en un cuarto de crecimiento esterila 25+/- 2 °C.
Al completar el 3 y 5 día de incubación, abrir la caja y observar la presencia o
ausencia y tamaño de la lesión para pruebas de patogenicidad.

77
ANEXO 4. Protocolo Monospóricos.
1. Objetivos
Obtener aislamientos puros generados a partir de una sola espora de
determinado hongo con el fin de garantizar la pureza y estabilidad genética
de las colecciones.
2. Alcance
Va dirigido a todo el personal que labora en el laboratorio de fitopatología
3. Documentos relacionados
Preparación de agar agua
4. Materiales y métodos
- Agua estéril
- Tubos tapa rosca con 1 ml de agua estéril
- Tubos tapa rosca secos, estériles
- Agar agua
- Pipetas automáticas de 0.5 a 10 µl, 40 a 200 µl y de 100 a1000 µl
- Puntas estériles
- Cámara de Nuebauer
- Laminillas
- Vórtex
- Microscopio
- Asas
- Mecheros

78
5. Procedimiento
A partir del cultivo inicial con el asa estéril tomar un inoculo.
Suspenderlo en 1 ml de agua estéril.
Agitarlo en el vórtex.
A la cámara de Nuebauer colocarle una laminilla.
Tomar 10 ul de la suspensión y ponerlo entre la cámara y la laminilla.
Realizar el conteo de esporas en los cuadrantes a, b, c y d.
Realizar el cálculo para obtener una espora en 1 mL de agua con la siguiente
formula:
Nº Esporas _________________ X 160.000 = esporas / ml 64 Nº Esporas _________________ X 160 = esporas / µl 64
Para obtener una suspensión de 1 espora / ml o 1 espora / µl, se aplica la
fórmula siguiente:
V1 C1 = V2 C2
Donde:
C1 = Concentración inicial (conocida en el conteo)
V1 = Volumen final (desconocido)
C2 = Concentración final deseada (según el estudio a realizar)
V2 = Volumen inicial (establecido arbitrariamente al preparar el inóculo)
Con los datos que se obtengan con la primera, se calcula la cantidad de agua
que se debe agregar al volumen necesario de suspensión de esporas
preparada, para obtener una concentración de 1 espora por ml o por µl.

79
Ej. V1 x C1 = V2 x C2
10 esporas 1mL 1mL
= 0.1 mL X 1.000 = 100 UL completar con 900 UL de agua, para obtener
1 espora en 1 mL.
Realizar círculos en al caja de agar agua.
Sembrar en cada pozo 2 µl de la suspensión final
Dejar a 25º C por 24 horas
Al completar las 24 horas se realiza la lectura en el microscopio. Observando
si hay germinación de las esporas.
Retirar con el asa estéril la espora que germino y colocarla en agar PDA.

80
ANEXO 5. Medio para evaluar la reacción de la proteasa producida por
Colletotrichum acutatum.
KH2PO4 1.0 g
KCL 0.5 g
CaCl2.2H2O 0.2 g Glucosa 10.0 g
Agar-Agar 12.0 g
Agua Destilada 975 ml
*Leche Desnaturalizada 15% 25 ml
Ajustar el pH al 5.4 con ácido clorhídrico y posteriormente autoclavar a
121°C, 20 min y 15 lb de presión.
*Preparar una solución de leche al 15% (Pesar 15 g de leche
desnaturalizada y diluirlo en 100 ml de agua), someter a agitación y tomar
el volumen requerido para completar el litro (25 ml).
Recomendación: El volumen de agua (975 ml) debe estar bajo agitación
constante y luego comenzar a agregar cada compuesto en el orden
establecido.

81
ANEXO 6. Protocolo Medio Suplementado con Base Benomyl.
1. Objetivo
Diferenciar especies de Colletotrichum susceptibles y tolerantes a benomyl
2. Alcance
Evaluación de pruebas biológicas para la identificación de especies de
Colletotrichum sp. aislados de mango.
3. Definiciones
El benomyl es un fungicida sistémico que pertenece al grupo de los
benzimidazoles y que interfiere con la división nuclear de los hongos que son
sensibles a su efecto (Delp y Kloping, 1968, citado por Tabares en el 2002).
Tiene acción protectora y erradicante, es efectivo contra numerosos hongos
fitopatógenos y también posee propiedades acaricidas (Tabares, 2002).
Este producto es absorbido por las plantas, luego de los cual se acumula
principalmente en las venas y en las hojas. Una vez penetra en el tejido
vegetal, el benomyl es convertido a butil isocianato y carbendazim, el cual se
une a las unidades de tubulina del patógeno, lo que afecta la formación de
microtúbulos. Esto interfiere directamente con ciertas funciones celulares, ya
que afecta el aparato mitótico, inhibe la división nuclear y el transporte
intracelular (Torres y Osorio, 2005).
4. Materiales y Equipos
- Fungicida Benomyl
- Agua destilada
- Agar PDA

82
- Autoclave
- Enlermeyer
- Cajas petri
- Tiras de pH
5. Contenido
5.1 Prepara el agar PDA
5.2 Autoclavar a 121 °C y 15 Lb de presión (15 mint)
5.3 Dejar atemperar el agar PDA
5.4 Agragar el fungicida benomyl hasta obtener una concentración de 2ug/ml
(Peres et al; 2002)
5.5 Servir en cajas petri y conservar las cajas en refrigeración
5.6 Sembrar las cepas de Colletotrichum que se desean evaluar
5.7 Incubar durante 72 horas
5.8 Medir los halos de crecimiento y observar el color predominante de la
colonia.
5.9 Interpretación: en base a los resultados del crecimiento de las especies
de Colletotrichum se debe determinar la susceptibilidad o la tolerancia del
fungicida, estos parámetros deben ir a la mano con las otras evaluaciones
biológicas para identificar la especie en estudio.

83
ANEXO 7. Protocolo Medio Suplementado con Hidróxido de Cobre y
Estreptomicina
1. Objetivo
Diferenciar especies de Colletotrichum susceptibles y tolerantes a CuOH2
2. Alcance
Este procedimiento se aplica para obtener una clasificación preliminar de
especies de Colletotrichum sp. aislados de diversos frutales.
3. Definiciones
CuOH2 es el medio selectivo para Colletotrichum, se compone de Hidroxido
de Cobre como ingrediente ativo, y del antibiótico Estreptomicina. El CuOH2
es un fungistático que actúa desnaturalizando las proteínas, a través de la
reactivación de los grupos sulfihídricos (Ware y Whitaker, 2000). Este medio
permite clasificar los diferentes aislamientos para la diferenciación entre
estas dos especies de Colletotrichum por medio de la observación de la
coloración que predomina en la colonia y la taza de crecimiento (Abang, et al;
2003).
4. Materiales y Equipos
- Fungicida CuOH2
- Antibiótico estreptomicina
- Agua destilada
- Agar PDA
- Autoclave enlermeyer
- Cajas de petri
- Tiras de pH

84
5. Contenido
5.1 preparar el agar PDA
5.2 autoclavar a 121°C y 15Lb (15min)
5.3 dejar atemperar el agar PDA
5.4 Agregar el fungicida CuPH2 hasta obtener una concentración de 42mg/L.
5.5 Agregar 300 mg/L de estreptomicina
5.6 Servir en cajas de petri y conservar las cajas en refrigeración
5.7 Sembrar las cepas de Colletotrichum que se deseen evalaur
5.8 Incubar durane 72 horas
5.9 Medir los halos de crecimiento y observar el color predominante de la
colonia
5.10 Interpretación: en base a los resultados del crecimiento de las especies
de Colletotrichum, se debe determinar la susceptibilidad o la tolerancia del
fungicida, estos parámetros deben ir a la mano con las otras evaluaciones
biológicas para identificar la especie en estudio.

85
ANEXO 8. Incidencia de Infecciones Quiescentes en los estados fenológicos de frutos de Mango. INCIDENCIA DE LA ENFERMEDAD 99 19:42 Wednesday, April 14, 1998 The GLM Procedure Class Level Information Class Levels Values rep 4 1 2 3 4 est 4 E1 E2 E3 E4 Number of observations 16
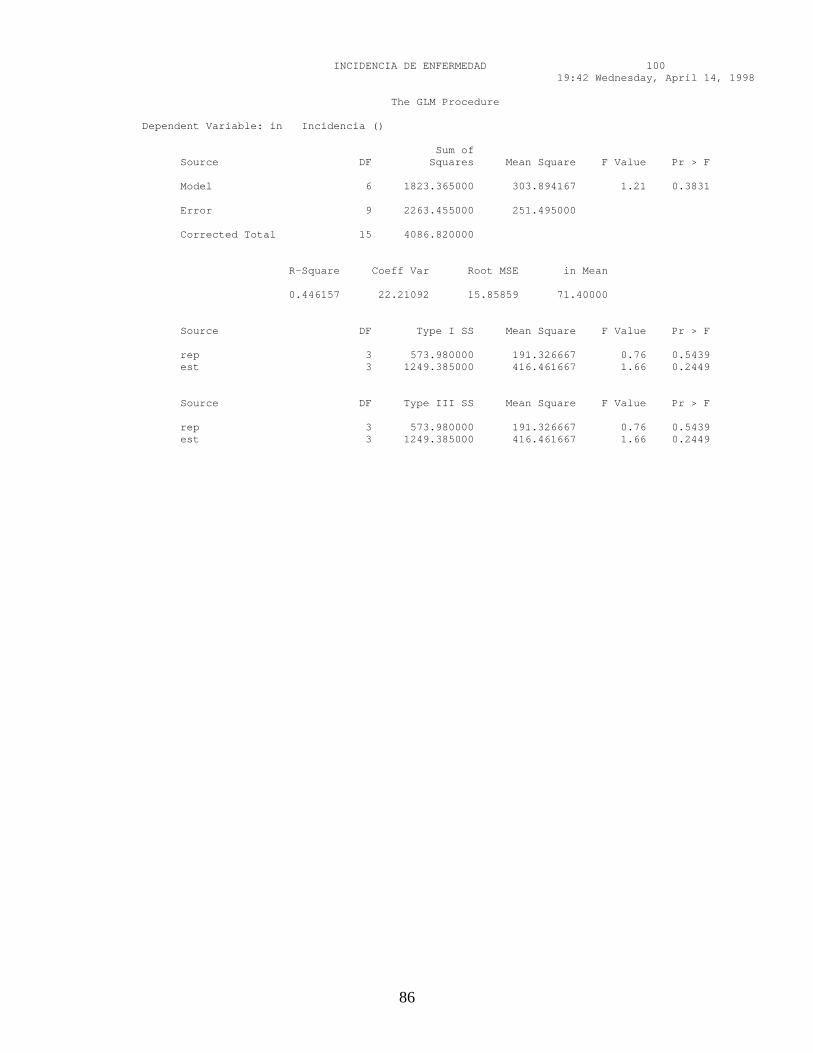
86
INCIDENCIA DE ENFERMEDAD 100 19:42 Wednesday, April 14, 1998 The GLM Procedure Dependent Variable: in Incidencia () Sum of Source DF Squares Mean Square F Value Pr > F Model 6 1823.365000 303.894167 1.21 0.3831 Error 9 2263.455000 251.495000 Corrected Total 15 4086.820000 R-Square Coeff Var Root MSE in Mean 0.446157 22.21092 15.85859 71.40000 Source DF Type I SS Mean Square F Value Pr > F rep 3 573.980000 191.326667 0.76 0.5439 est 3 1249.385000 416.461667 1.66 0.2449 Source DF Type III SS Mean Square F Value Pr > F rep 3 573.980000 191.326667 0.76 0.5439 est 3 1249.385000 416.461667 1.66 0.2449

87
INCIDENCIA DE LA ENFERMEDAD 101 19:42 Wednesday, April 14, 1998 The GLM Procedure t Tests (LSD) for in NOTE: This test controls the type I comparisonwise error rate not the experimentwise error rate. Alpha 0.05 Error Degrees of Freedom 9 Error Mean Square 251.495 Critical Value of t 2.26216 Least Significant Difference 25.367 Means with the same letter are not significantly different. t Grouping Mean N est A 81.13 4 E1 A A 79.05 4 E2 A A 64.75 4 E3 A A 60.68 4 E4

88
ANEXO 9. Pruebas de Validación, Normalidad y comparación de promedios en la
incidencia de infecciones quiescentes en los estados fenológicos de frutos de mango. INCIDENCIA DE ENFERMEDAD 102 19:42 Wednesday, April 14, 1998 The UNIVARIATE Procedure Variable: r Moments N 16 Sum Weights 16 Mean 0 Sum Observations 0 Std Deviation 12.284014 Variance 150.897 Skewness 0.05635386 Kurtosis -0.484169 Uncorrected SS 2263.455 Corrected SS 2263.455 Coeff Variation . Std Error Mean 3.0710035 Basic Statistical Measures Location Variability Mean 0.000000 Std Deviation 12.28401 Median 0.212500 Variance 150.89700 Mode . Range 44.90000 Interquartile Range 16.58750 Tests for Location: Mu0=0 Test -Statistic- -----p Value------ Student's t t 0 Pr > |t| 1.0000 Sign M 0 Pr >= |M| 1.0000 Signed Rank S -1 Pr >= |S| 0.9799 Tests for Normality Test --Statistic--- -----p Value------ Shapiro-Wilk W 0.985528 Pr < W 0.9926 Kolmogorov-Smirnov D 0.121483 Pr > D >0.1500 Cramer-von Mises W-Sq 0.02133 Pr > W-Sq >0.2500 Anderson-Darling A-Sq 0.139598 Pr > A-Sq >0.2500 Quantiles (Definition 5) Quantile Estimate 100% Max 21.9500 99% 21.9500 95% 21.9500 90% 17.8000

89
INCIDENCIA DE LA ENFERMEDAD 103 19:42 Wednesday, April 14, 1998 The UNIVARIATE Procedure Variable: r Quantiles (Definition 5) Quantile Estimate 75% Q3 8.1500 50% Median 0.2125 25% Q1 -8.4375 10% -15.3000 5% -22.9500 1% -22.9500 0% Min -22.9500 Extreme Observations -----Lowest----- -----Highest---- Value Obs Value Obs -22.950 14 6.075 9 -15.300 3 10.225 12 -10.575 5 13.300 2 -8.725 8 17.800 6 -8.150 10 21.950 15 Stem Leaf # Boxplot 2 2 1 | 1 8 1 | 1 03 2 | 0 66 2 +-----+ 0 23 2 *--+--* -0 1 1 | | -0 9885 4 +-----+ -1 1 1 | -1 5 1 | -2 3 1 | ----+----+----+----+ Multiply Stem.Leaf by 10**+1

90
INCIDENCIA DE LA ENFERMEDAD 104 19:42 Wednesday, April 14, 1998 The UNIVARIATE Procedure Variable: r Normal Probability Plot 22.5+ +*++ | *+++ | *+*++ 7.5+ *+*++ | +**++ | ++** -7.5+ *+*+* | ++*+ | +++* -22.5+ ++*+ +----+----+----+----+----+----+----+----+----+----+ -2 -1 0 +1 +2

91
ANEXO 10. Características Morfológicas de los Aislamientos.
ESTADO AISLAMIENTO COLOR CRECIMIENTO 1 Gris Radial 2 Gris oscuro Radial 3 Gris Oliva Radial 4 Blanco Radial 5 Salmón Radial
E1
6 Oliva Radial 1 Blanco Radial 2 Oliva Radial 3 Gris Oscuro Radial 4 Blanco Salmón Radial 5 Gris Radial 6 Salmón Radial
E2
7 Gris Oliva Radial 1 Gris Radial 2 Oliva Radial 3 Gris Oscuro Radial
E3
4 Blanco Radial 1 Oliva Radial 2 Gris Radial 3 Gris Oscuro Radial 4 Salmón Radial
E4
5 Blanco Radial

92
ANEXO 11. Prueba de Benomyl en 2 repeticiones.
ESTADO AISLAMIENTO CONTROL 1 21 + - - 2 + - - 3 + - - 4 + - - 5 + - -
E1
6 + - - 1 + - - 2 + - - 3 + - - 4 + - - 5 + - - 6 + - -
E2
7 + - - 1 + - - 2 + - - 3 + - -
E3
4 + - - 1 + - - 2 + - - 3 + - - 4 + - -
E4
5 + - -
-: Sensibilidad e inhibición del crecimiento micelial.
+: Tolerancia y crecimiento micelial.

93
ANEXO 12. Prueba de la proteasa en 2 repeticiones.
ESTADO AISLAMIENTO 1 2 1 - - 2 - - 3 - - 4 - - 5 - -
E1
6 - - 1 - - 2 - - 3 - - 4 - - 5 - - 6 - -
E2
7 - - 1 - - 2 - - 3 - -
E3
4 - - 1 - - 2 - - 3 - - 4 - -
E4
5 - -
C. acutatum (852) C. gloeosporioides (1022)
+ - + - + - + -
La respuesta positiva es una característica que presenta C. acutatum.
La respuesta negativa es una característica que presenta C. gloeosporioides.

94
ANEXO 13. Prueba de Medio Selectivo para Colletotrichum spp.
ESTADOS AISLAMIENTO REPETICIÓN DIAMETRO (cm) CLASIFICACIÓN
PDA COBRE COBRE Crecimiento Especie
1 2.7 3.0 Salmón FGS C.
gloeosporioides
1 2 2.7 2.8 Salmón FGS C.
gloesoporioides
1 2.6 2.8 Salmón FGS C.
gloeosporioides
2 2 2.5 2.6 Salmón FGS C.
gloeosporioides
1 3.0 3.5 Gris FGG C.
gloeosporioides
3 2 3.4 3.3 Gris FGG C.
gloeosporioides
1 3.4 3.1 Gris FGG C.
gloeosporioides
4 2 3.2 3.0 Gris FGG C.
gloeosporioides
1 3.2 2.8 Oliva FGO C.
gloeosporioides
5 2 3.2 2.6 Oliva FGO C.
gloeosporioides
1 3.4 2.4 Gris SGG C.
gloeosporioides
E1
6 2 3.4 2.3 Gris SGG C.
gloeosporioides
1 3.3 3.6 Salmón FGS C.
gloeosporioides
1 2 3.1 2.7 Salmón FGS C.
gloeosporioides
1 2.7 3.2 Gris FGG C.
gloeosporioides
2 2 3.4 3.4 Gris FGG C.
gloeosporioides
1 3.3 3.6 Oliva FGO C.
gloeosporioides
3 2 3.3 3.0 Oliva FGO C.
gloeosporioides
1 3.0 2.6 Gris FGG C.
gloeosporioides
4 2 2.9 2.8 Gris FGG C.
gloeosporioides
1 3.4 3.2 Salmón FGS C.
gloeosporioides
5 2 3.6 3.3 Salmón FGS C.
gloeosporioides
1 2.9 3.4 Gris FGG C.
gloeosporioides
6 2 2.7 3.3 Gris FGG C.
gloeosporioides
1 2.9 3.1 Oliva FGO C.
gloeosporioides
E2
7 2 3.2 2.9 Oliva FGO C.
gloeosporioides
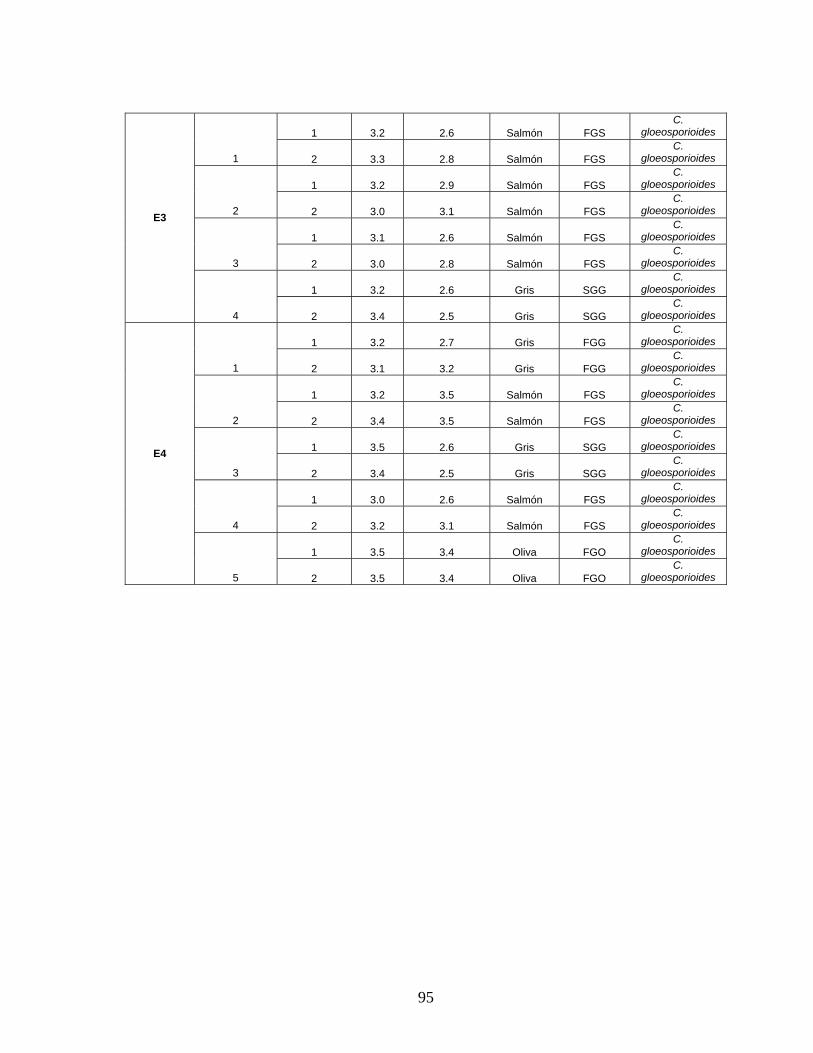
95
1 3.2 2.6 Salmón FGS C.
gloeosporioides
1 2 3.3 2.8 Salmón FGS C.
gloeosporioides
1 3.2 2.9 Salmón FGS C.
gloeosporioides
2 2 3.0 3.1 Salmón FGS C.
gloeosporioides
1 3.1 2.6 Salmón FGS C.
gloeosporioides
3 2 3.0 2.8 Salmón FGS C.
gloeosporioides
1 3.2 2.6 Gris SGG C.
gloeosporioides
E3
4 2 3.4 2.5 Gris SGG C.
gloeosporioides
1 3.2 2.7 Gris FGG C.
gloeosporioides
1 2 3.1 3.2 Gris FGG C.
gloeosporioides
1 3.2 3.5 Salmón FGS C.
gloeosporioides
2 2 3.4 3.5 Salmón FGS C.
gloeosporioides
1 3.5 2.6 Gris SGG C.
gloeosporioides
3 2 3.4 2.5 Gris SGG C.
gloeosporioides
1 3.0 2.6 Salmón FGS C.
gloeosporioides
4 2 3.2 3.1 Salmón FGS C.
gloeosporioides
1 3.5 3.4 Oliva FGO C.
gloeosporioides
E4
5 2 3.5 3.4 Oliva FGO C.
gloeosporioides

96
ANEXO 14. Presencia de síntomas iniciales de Antracnosis en los
tratamientos de Patogenicidad.
ESTADO AISLAMIENTO MANGO1 4 Días 2 4 Días 3 4 Días 4 4 Días 5 7 Días
E1
6 4 Días 1 9 Días 2 4 Días 3 4 Días 4 4 Días 5 4 Días 6 4 Días
E2
7 4 Días 1 4 Días 2 4 Días 3 4 Días
E3
4 4 Días 1 4 Días 2 4 Días 3 4 Días 4 4 Días
E4
5 5 Días

97
ANEXO 15. Efecto de la Enfermedad sobre el pH en áreas sanas y enfermas de frutos
enfermos y en frutos sanos.
EFECTO DE LA ENFERMEDAD SOBRE EL pH 83 19:42 Wednesday, April 14, 1998 The GLM Procedure Class Level Information Class Levels Values est 4 e1 e2 e3 e4 enf 3 te ts ts_fe tej 2 exo meso Number of observations 96

98
EFECTO DE LA ENFERMEDAD SOBRE EL pH 84 19:42 Wednesday, April 14, 1998 The GLM Procedure Dependent Variable: pH Sum of Source DF Squares Mean Square F Value Pr > F Model 23 93.45663333 4.06333188 48.04 <.0001 Error 72 6.09015000 0.08458542 Corrected Total 95 99.54678333 R-Square Coeff Var Root MSE ph Mean 0.938821 5.341744 0.290836 5.444583 Source DF Type I SS Mean Square F Value Pr > F est 3 4.55715833 1.51905278 17.96 <.0001 enf 2 64.71193958 32.35596979 382.52 <.0001 est*enf 6 9.62831042 1.60471840 18.97 <.0001 tej 1 8.49660000 8.49660000 100.45 <.0001 est*tej 3 0.44172500 0.14724167 1.74 0.1663 enf*tej 2 4.80166875 2.40083437 28.38 <.0001 est*enf*tej 6 0.81923125 0.13653854 1.61 0.1556 Source DF Type III SS Mean Square F Value Pr > F est 3 4.55715833 1.51905278 17.96 <.0001 enf 2 64.71193958 32.35596979 382.52 <.0001 est*enf 6 9.62831042 1.60471840 18.97 <.0001 tej 1 8.49660000 8.49660000 100.45 <.0001 est*tej 3 0.44172500 0.14724167 1.74 0.1663 enf*tej 2 4.80166875 2.40083437 28.38 <.0001 est*enf*tej 6 0.81923125 0.13653854 1.61 0.1556

99
EFECTO DE LA ENFERMEDAD SOBRE EL pH 85
19:42 Wednesday, April 14, 1998
The GLM Procedure t Tests (LSD) for ph NOTE: This test controls the type I comparison wise error rate not the experiment wise error rate. Alpha 0.05 Error Degrees of Freedom 72 Error Mean Square 0.084585 Critical Value of t 1.99346 Least Significant Difference 0.1674 Means with the same letter are not significantly different. t Grouping Mean N est A 5.69333 24 e2 A A 5.57667 24 e1 B 5.39208 24 e3 C 5.11625 24 e4

100
EFECTO DE LA ENFERMEDAD SOBRE EL pH 86 19:42 Wednesday, April 14, 1998 The GLM Procedure t Tests (LSD) for ph NOTE: This test controls the type I comparisonwise error rate not the experimentwise error rate. Alpha 0.05 Error Degrees of Freedom 72 Error Mean Square 0.084585 Critical Value of t 1.99346 Least Significant Difference 0.1449 Means with the same letter are not significantly different. t Grouping Mean N enf A 6.56375 32 te B 5.15281 32 ts_fe C 4.61719 32 ts

101
EFECTO DE LA ENFERMEDAD SOBRE EL pH 87 19:42 Wednesday, April 14, 1998 The GLM Procedure t Tests (LSD) for ph NOTE: This test controls the type I comparison wise error rate not the experiment wise error rate. Alpha 0.05 Error Degrees of Freedom 72 Error Mean Square 0.084585 Critical Value of t 1.99346 Least Significant Difference 0.1183 Means with the same letter are not significantly different. t Grouping Mean N tej A 5.74208 48 exo B 5.14708 48 meso
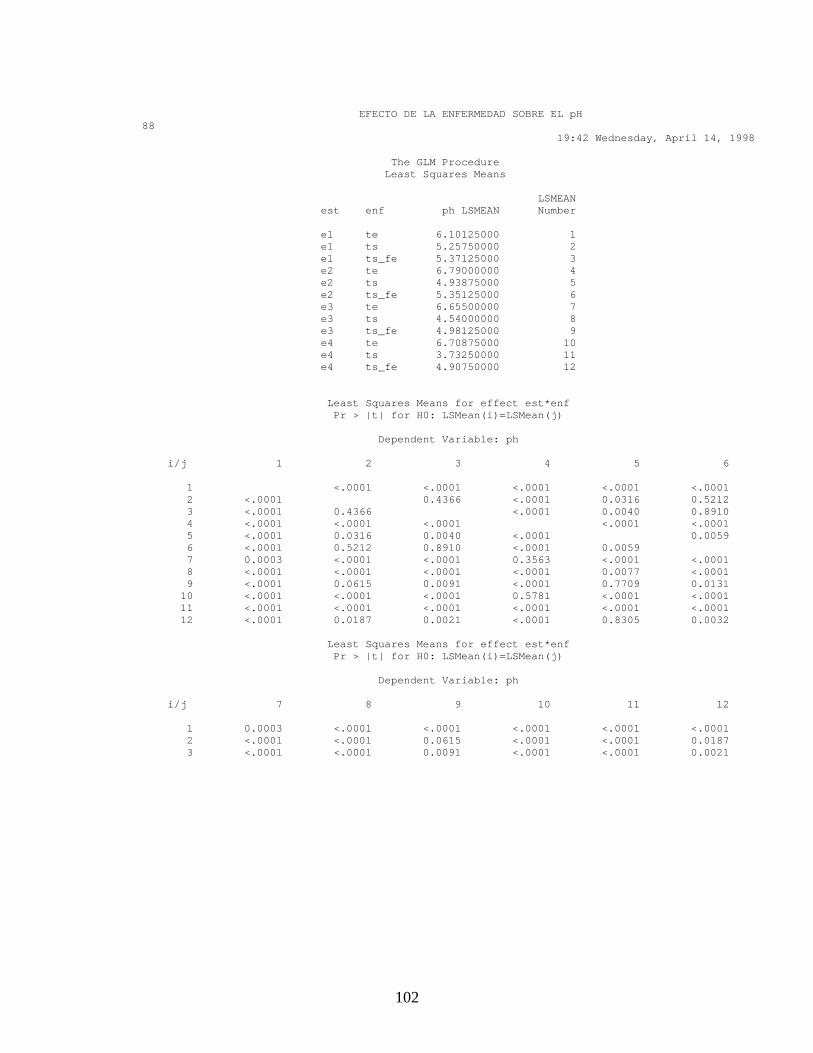
102
EFECTO DE LA ENFERMEDAD SOBRE EL pH 88 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means LSMEAN est enf ph LSMEAN Number e1 te 6.10125000 1 e1 ts 5.25750000 2 e1 ts_fe 5.37125000 3 e2 te 6.79000000 4 e2 ts 4.93875000 5 e2 ts_fe 5.35125000 6 e3 te 6.65500000 7 e3 ts 4.54000000 8 e3 ts_fe 4.98125000 9 e4 te 6.70875000 10 e4 ts 3.73250000 11 e4 ts_fe 4.90750000 12 Least Squares Means for effect est*enf Pr > |t| for H0: LSMean(i)=LSMean(j) Dependent Variable: ph i/j 1 2 3 4 5 6 1 <.0001 <.0001 <.0001 <.0001 <.0001 2 <.0001 0.4366 <.0001 0.0316 0.5212 3 <.0001 0.4366 <.0001 0.0040 0.8910 4 <.0001 <.0001 <.0001 <.0001 <.0001 5 <.0001 0.0316 0.0040 <.0001 0.0059 6 <.0001 0.5212 0.8910 <.0001 0.0059 7 0.0003 <.0001 <.0001 0.3563 <.0001 <.0001 8 <.0001 <.0001 <.0001 <.0001 0.0077 <.0001 9 <.0001 0.0615 0.0091 <.0001 0.7709 0.0131 10 <.0001 <.0001 <.0001 0.5781 <.0001 <.0001 11 <.0001 <.0001 <.0001 <.0001 <.0001 <.0001 12 <.0001 0.0187 0.0021 <.0001 0.8305 0.0032 Least Squares Means for effect est*enf Pr > |t| for H0: LSMean(i)=LSMean(j) Dependent Variable: ph i/j 7 8 9 10 11 12 1 0.0003 <.0001 <.0001 <.0001 <.0001 <.0001 2 <.0001 <.0001 0.0615 <.0001 <.0001 0.0187 3 <.0001 <.0001 0.0091 <.0001 <.0001 0.0021
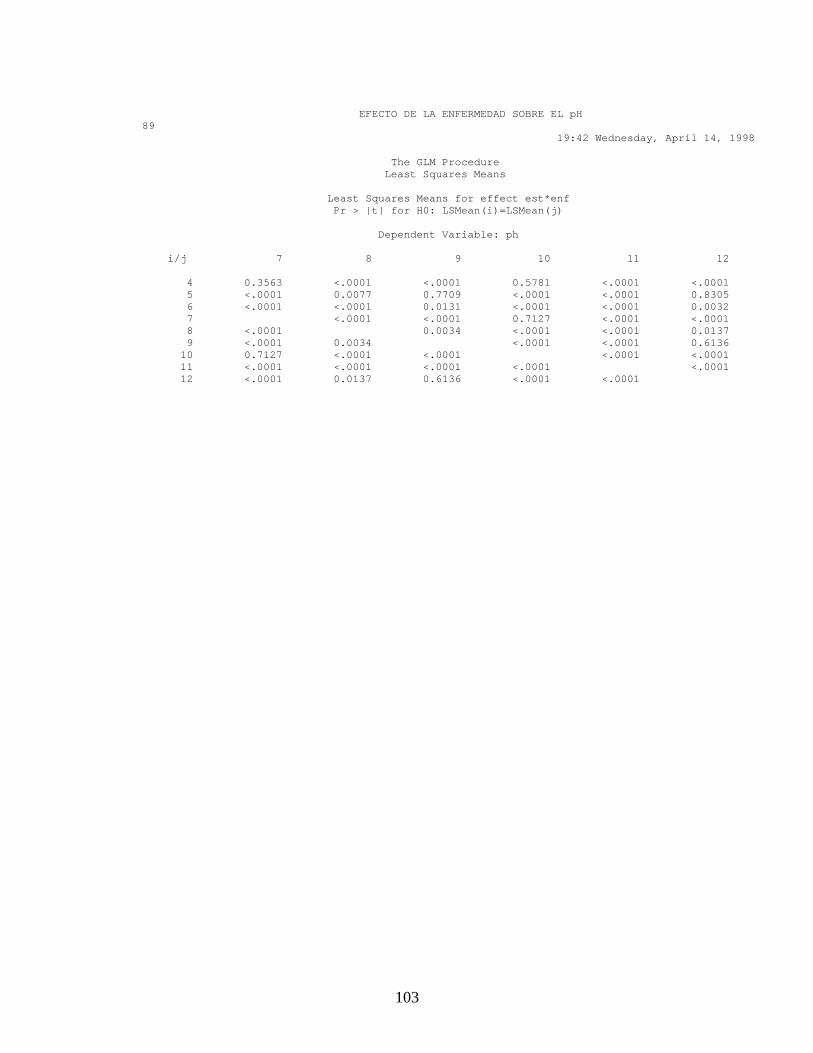
103
EFECTO DE LA ENFERMEDAD SOBRE EL pH 89 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means Least Squares Means for effect est*enf Pr > |t| for H0: LSMean(i)=LSMean(j) Dependent Variable: ph i/j 7 8 9 10 11 12 4 0.3563 <.0001 <.0001 0.5781 <.0001 <.0001 5 <.0001 0.0077 0.7709 <.0001 <.0001 0.8305 6 <.0001 <.0001 0.0131 <.0001 <.0001 0.0032 7 <.0001 <.0001 0.7127 <.0001 <.0001 8 <.0001 0.0034 <.0001 <.0001 0.0137 9 <.0001 0.0034 <.0001 <.0001 0.6136 10 0.7127 <.0001 <.0001 <.0001 <.0001 11 <.0001 <.0001 <.0001 <.0001 <.0001 12 <.0001 0.0137 0.6136 <.0001 <.0001
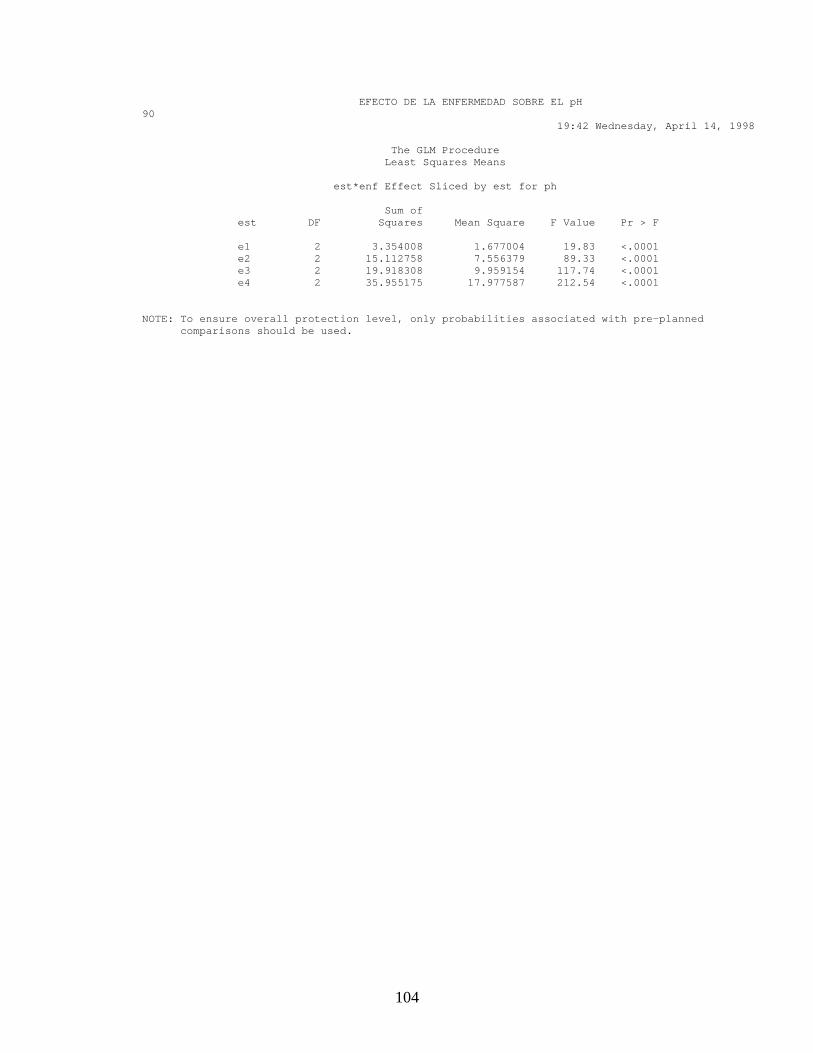
104
EFECTO DE LA ENFERMEDAD SOBRE EL pH 90 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means est*enf Effect Sliced by est for ph Sum of est DF Squares Mean Square F Value Pr > F e1 2 3.354008 1.677004 19.83 <.0001 e2 2 15.112758 7.556379 89.33 <.0001 e3 2 19.918308 9.959154 117.74 <.0001 e4 2 35.955175 17.977587 212.54 <.0001 NOTE: To ensure overall protection level, only probabilities associated with pre-planned comparisons should be used.

105
EFECTO DE LA ENFERMEDAD SOBRE EL pH 91 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means est enf ph LSMEAN e1 te 6.10125000 e1 ts 5.25750000 e1 ts_fe 5.37125000 e2 te 6.79000000 e2 ts 4.93875000 e2 ts_fe 5.35125000 e3 te 6.65500000 e3 ts 4.54000000 e3 ts_fe 4.98125000 e4 te 6.70875000 e4 ts 3.73250000 e4 ts_fe 4.90750000

106
EFECTO DE LA ENFERMEDAD SOBRE EL pH 92 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means est*enf Effect Sliced by enf for ph Sum of enf DF Squares Mean Square F Value Pr > F te 3 2.355575 0.785192 9.28 <.0001 ts 3 10.416259 3.472086 41.05 <.0001 ts_fe 3 1.413634 0.471211 5.57 0.0017
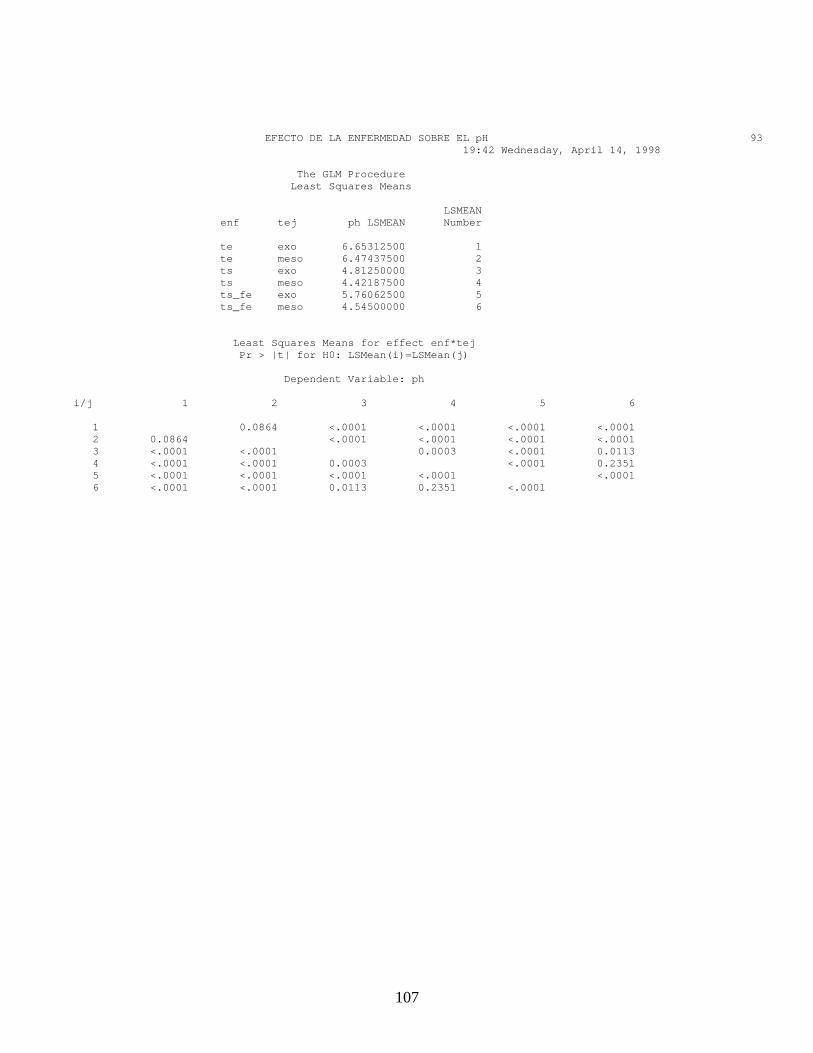
107
EFECTO DE LA ENFERMEDAD SOBRE EL pH 93 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means LSMEAN enf tej ph LSMEAN Number te exo 6.65312500 1 te meso 6.47437500 2 ts exo 4.81250000 3 ts meso 4.42187500 4 ts_fe exo 5.76062500 5 ts_fe meso 4.54500000 6 Least Squares Means for effect enf*tej Pr > |t| for H0: LSMean(i)=LSMean(j) Dependent Variable: ph i/j 1 2 3 4 5 6 1 0.0864 <.0001 <.0001 <.0001 <.0001 2 0.0864 <.0001 <.0001 <.0001 <.0001 3 <.0001 <.0001 0.0003 <.0001 0.0113 4 <.0001 <.0001 0.0003 <.0001 0.2351 5 <.0001 <.0001 <.0001 <.0001 <.0001 6 <.0001 <.0001 0.0113 0.2351 <.0001

108
EFECTO DE LA ENFERMEDAD SOBRE EL pH 94 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means enf*tej Effect Sliced by enf for ph Sum of enf DF Squares Mean Square F Value Pr > F te 1 0.255613 0.255613 3.02 0.0864 ts 1 1.220703 1.220703 14.43 0.0003 ts_fe 1 11.821953 11.821953 139.76 <.0001 NOTE: To ensure overall protection level, only probabilities associated with pre-planned comparisons should be used.

109
EFECTO DE LA ENFERMEDAD SOBRE EL pH 95 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means enf tej ph LSMEAN te exo 6.65312500 te meso 6.47437500 ts exo 4.81250000 ts meso 4.42187500 ts_fe exo 5.76062500 ts_fe meso 4.54500000

110
EFECTO DE LA ENFERMEDAD SOBRE EL pH 96 19:42 Wednesday, April 14, 1998 The GLM Procedure Least Squares Means enf*tej Effect Sliced by tej for ph Sum of tej DF Squares Mean Square F Value Pr > F exo 2 27.111454 13.555727 160.26 <.0001 meso 2 42.402154 21.201077 250.65 <.0001

111
ANEXO 16. Pruebas de Validación, Normalidad y comparación de promedios en el
efecto de la enfermedad sobre el pH.
EFECTO DE LA ENFEMEDAD SOBRE pH 97 19:42 Wednesday, April 14, 1998 The UNIVARIATE Procedure Variable: r Moments N 96 Sum Weights 96 Mean 0 Sum Observations 0 Std Deviation 0.25319329 Variance 0.06410684 Skewness -0.2383586 Kurtosis -0.566571 Uncorrected SS 6.09015 Corrected SS 6.09015 Coeff Variation . Std Error Mean 0.02584143 Basic Statistical Measures Location Variability Mean 0.00000 Std Deviation 0.25319 Median -0.00750 Variance 0.06411 Mode -0.17500 Range 1.09500 Interquartile Range 0.37000 NOTE: The mode displayed is the smallest of 8 modes with a count of 2. Tests for Location: Mu0=0 Test -Statistic- -----p Value------ Student's t t 0 Pr > |t| 1.0000 Sign M -1 Pr >= |M| 0.9188 Signed Rank S 52 Pr >= |S| 0.8504 Tests for Normality Test --Statistic--- -----p Value------ Shapiro-Wilk W 0.981149 Pr < W 0.1830 Kolmogorov-Smirnov D 0.077977 Pr > D >0.1500 Cramer-von Mises W-Sq 0.069137 Pr > W-Sq >0.2500 Anderson-Darling A-Sq 0.480666 Pr > A-Sq 0.2340 Quantiles (Definition 5) Quantile Estimate 100% Max 0.5250 99% 0.5250

112
EFECTO DE LA ENFERMEDAD SOBRE EL pH 98 19:42 Wednesday, April 14, 1998 The UNIVARIATE Procedure Variable: r Quantiles (Definition 5) Quantile Estimate 95% 0.4000 90% 0.3300 75% Q3 0.1950 50% Median -0.0075 25% Q1 -0.1750 10% -0.3950 5% -0.4400 1% -0.5700 0% Min -0.5700 Extreme Observations -----Lowest---- ----Highest---- Value Obs Value Obs -0.570 66 0.400 4 -0.525 62 0.410 67 -0.510 48 0.415 87 -0.460 36 0.465 92 -0.440 80 0.525 73