regulation of in vitro and in vivo differentiation of mouse embryonic stem cells, embryonic germ...
TRANSCRIPT

ISSN 1062-3604, Russian Journal of Developmental Biology, 2009, Vol. 40, No. 6, pp. 325–338. © Pleiades Publishing, Inc., 2009.Original Russian Text © O.F. Gordeeva, T.M. Nikonova, N.V. Lifantseva, 2009, published in Ontogenez, 2009, Vol. 40, No. 6, pp. 403–418.
325
INTRODUCTION
Pluripotent cells of an early embryo originate all celltypes of adult mammals. To study the mechanisms of earlydevelopment, permanent pluripotent stem cell lines werederived from the inner cell mass of a mammalian blasto-cysts and from germ line cells at different developmentalstaged: primordial germ cells, gonocytes, spermatogoniaand parthenogenetically activated oocytes (Evans andKaufman, 1981; Martin, 1981; Matsui et al., 1992; Resnicket al., 1992; Thomson et al., 1998; Shamblott et al., 1998;Cibelli et al., 2002; Kanatsu-Shinohara et al., 2004; Guanet al., 2006; Lin et al., 2007). Pluripotent stem cells—embryonic stem cells (ESCs) and embryonic germ cells(EGCs)—are capable of unlimited self-renewal duringlong-term culturing and retain the ability to differentiateinto different cell types in vitro. Pluripotent stem cell lineswith stable karyotypes retain the ability to develop into allcells of embryo including germ line cells, after the injec-tion of them into a blastocyst (Nagy et al., 1993). Inducedpluripotent stem cells converted experimentally fromadult somatic cells using genetic modifications based onlentiviral vectors transduction have similar features toESC and EGC properties (Takahashi and Yamanaka,2006; Takahashi et al., 2007; Yu et al., 2007).
Embryonic teratocarcinoma cell lines, malignantcounterparts of pluripotent stem cells, derived from mouseand human spontaneous testicular and ovarian tumorshave variable biological features which are partially simi-lar to the pluripotent cell characteristics (for review, seeAndrews, 2002). The available published data indicate thatteratocarcinoma cells in gonads arise out of disturbance inthe mechanism regulating the primordial germ cell differ-entiation from the pluripotent cells (Andrews, 2002;Kimura et al., 2003; Gordeeva, 2007). Chromosomal andgene mutations in teratocarcinoma cells determine theirability to differentiate into different types of cells or com-pletely limit all differentiations, which is observed in thecase of nullipotent cell lines (Blelloch et al., 2004).
The studies of the mechanisms underlying self-renewal and differentiation of different origin pluripotentstem cells originated from progenitor cells with variousontogenetic and epigenetic state indicate that althoughsome of the characteristics of these cell lines are similar,they differ in gene expression, genetic stability, and abilityto differentiate into various cell types (Rohwedel et al.,1996; Allegrucci, Young, 2007; Sharova et al., 2007). Itwas shown that the ontogenetic status of progenitor cellsaffects gene expression in undifferentiated and differenti-ating ESCs and EGCs less significantly than their affilia-tion to different genotypes of mouse strains (Sharova et al.,
Regulation of in vitro and in vivo Differentiation of Mouse Embryonic Stem Cells, Embryonic Germ Cells
and Teratocarcinoma Cells by TGF
b
Family Signaling Factors
O. F. Gordeeva, T. M. Nikonova, and N. V. Lifantseva
Kol’tsov Institute of Developmental Biology, Russian Academy of Sciences, ul. Vavilova 26, Moscow, 119334 Russia e-mail: [email protected]
Received April 27, 2009
Abstract
—The activity of specific signaling and transcription factors determines the cell fate in normal devel-opment and in tumor transformation.The transcriptional profiles of gene-components of different branches ofTGF
β
family signaling pathways were studied in experimental models of initial stages of three-dimensional invitro differentiation of embryonic stem cells, embryonic germ cells and teratocarcinoma cells and in teratomasand teratocarcinomas developed after their transplantation into immunodeficient Nude mice. Gene profile anal-ysis of studied cell systems have revealed that expression patterns of
ActivinA
,
Nodal
,
Lefty
1
,
Lefty
2
,
Tgf
β
1
,
Bmp
4
, and
GDF
3
were identical in pluripotent stem cells whereas the mRNAs of all examined genes with theexception of (Inhibin
Inhibin
β
A
/
ActivinA
) were detected in the teratocarcinoma cells. These results indicate thatdifferential activity of signaling pathways of the TGF
β
family factors regulates the pluripotent state mainte-nance and pluripotent stem cell differentiation into the progenitors of three germ layers and extraembryonicstructures and that normal expression pattern of TGF
β
family factors is rearranged in embryonic teratocarci-noma cells during tumor growth in vitro and in vivo.
DOI:
10.1134/S1062360409060010
Key words
: pluripotent stem cells, embryonic stem cells, teratocarcinoma, embryoid body, teratomas, TGF
β
,ActivinA, differentiation.
EXPERIMENTAL EMBRYOLOGY

326
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY
Vol. 40
No. 6
2009
GORDEEVA et al.
2007). There are numerous data regarding the differencesin the transcriptional profiles of human and mouse ESCs,which, nevertheless, have no effect on their general biolog-ical properties (Brandenberger et al., 2004; Calhoun et al.,2004; Adewumi et al., 2007; Sharova et al., 2007). At thesame time, induced pluripotent stem cells derived from var-ious somatic cells demonstrate different oncogenic poten-tial, which apparently depends on the genetic and epigeneticstates of cell line progenitors and may be the result of distur-bance in reprogramming mechanisms (Maherali et al.,2007; Okita et al., 2007; Wernig et al., 2007; Aoi et al.,2008). In addition, the observed variability in the develop-mental potential of embryonic teratocarcinoma lines sug-gests that the cell fate in carcinogenesis is regulated by dif-ferent mechanisms that significantly differ from the mecha-nisms of embryonic development.
The TGF
β
family signaling pathways integrated intothe regulatory cellular networks are involved in variousprocesses including the cell fate determination during nor-mal and pathological development, proliferation and celldeath control (Miura et al., 2008; Siegel and Massague,2008). In humans and other animals, the TGF
β
familyincludes 40 factors which, despite their structural andfunctional similarity, ensure precise regulation of cell pro-cesses (Derynck and Miyazono, 2008). On the other hand,the same factor of this family may be involved in the reg-ulation of various processes in different cells. In a word, allfactors of the TGF
β
family are polyfunctional and con-text-dependent, i.e. they manifest their properties depend-ing on other factors under certain conditions in differenttypes of cells (Katagiri et al., 2008; Shilling et al., 2008;Wiater and Vale, 2008). It is known that member of thisfamily, Nodal, Activin, Lefty, BMP, GDF, and TGF
β
, arethe key regulators of cell determination in three germ lay-ers, germ line and extraembryonic structures and areinvolved in the regulation of morphogenesis and forma-tion of polarity axes in embryo (Beddington and Robert-son, 1999; Vincent et al., 2003; Miura et al., 2008). How-ever, the mechanisms of functioning of signaling systemsactivating specific target genes and interactions of differ-ent signaling pathways in different types of cells remainlargely obscure.
The goal of this study was to investigate differentbranches of TGF
β
family signaling systems during differ-entiation of mouse ESCs, EGCs and embryonic teratocar-cinoma cells (ECCs) in a three-dimensional serum-freeculture system and after their transplantation into immun-odeficient Nude mice. Comparative analysis of growthand differentiation dynamics of ESCs, EGSc, and ECCs invitro and in vivo and their transcriptional profiles includ-ing the gene-components of different branches of TGF
β
family signaling pathways have revealed the commonmechanisms of regulation of pluripotent stem cell differ-entiation, which significantly differ from the regulatorymechanisms in cancer teratocarcinoma cells.
MATERIALS AND METHODS
Cell culture in vitro. For the experiments, we use
d themouse ECS line R1, mouse EGC-10 line kindly providedby Dr. A. McLaren (WTCR Institute of Cancer and Devel-opmental Biology, Cambridge, United Kingdom) andmouse ECC F9 line (the Cell Culture Bank, Institute ofCytology, Russian Academy of Sciences, St. Petersburg).ESCs, EGCs, and ECCs were cultured in the knockoutDMEM supplemented with 1 mM L-glutamine, 0.1 mMnonessential amino acids, 0.1 mM
β
-mercaptoethanol,and 15% Knockout Serum Replacement (Gibco, USA).Undifferentiated ESCs and EGCs were maintained onmouse primary embryonic fibroblast feeder inactivated bymitomycin C (10
μ
g/ml; Sigma, USA). To study thegrowth dynamics, ESCs, EGC and ECCs were cultured ina medium containing or not containing the leukemiainhibitory factor LIF (10 ng/ml, Sigma, United States).The number of cells grown in the presence and absence ofLIF was determined on day 5 of culturing. Statistical anal-ysis of differences was performed using paired Student’s
t
-test; the significance of the hypotheses on the LIF effectwas assessed by ANOVA.
The standard embryoid bodies were generated usingthe hanging drop culture method: drops of suspensions ofpluripotent and teratocarcinoma cells (500 cells per drop)were placed on a Petri dish lids and cultured for 3 days toform embryoid bodies. Developed embryoid bodies(EB1) were collected from drops, transferred to newdishes and cultured in suspension for subsequent 5 days(EB5). Thus, the total duration of cell culturing and differ-entiation under the described conditions was 7 days.
Development of teratomas and teratocarcinomas invivo.
To obtain experimental teratomas and teratocarcino-mas, immunodeficient five- to seven-month-old mice ofthe Nude line (Nu/Nu) were used as recipients. The ani-mals were obtained from the Pushchino nursery for labo-ratory animals, Shemyakin–Ovchinnikov Institute ofBioorganic Chemistry, Pushchino branch, Russian Acad-emy of Sciences; the stock line was obtained from theCharles Rivers nursery, United States. Twenty embryoidbodies (a total of 200–250 thousand cells) formed on day 3in hanging drops were transplanted under the renal cap-sules of experimental animals. Six weeks after ESC andEGS injection, the animals were sacrificed, and the devel-oped teratomas were removed. The teratocarcinomas wereextracted 3 weeks after the cell injection because theygrew more rapidly than the pluripotent stem cells. Thesamples of teratomas and teratocarcinomas were used infurther analysis. For histological studies, tumor sampleswere fixed in 10% formalin, dehydrated by the standardprocedure, and embedded in paraffin to prepare tissue sec-tion. The preparations were stained with hematoxylin andeosin and then studied using the microscopic image anal-ysis system Leica DMRXA2 (Germany).
Histochemical analysis of alkaline phosphatase activ-ity.
Cells were fixed in 3% paraformaldehyde in phos-phate-buffered saline (PBS; Sigma, USA) for 15–20 min.After washing with PBS, the samples were incubated at

RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY
Vol. 40
No. 6
2009
REGULATION OF in vitro AND in vivo DIFFERENTIATION OF MOUSE 327
37°ë
for 60 min in a solution containing 10 ml of 0.02 M
Tris
-HCl buffer (pH 8.6), 1 mg of AS-BI-phosphate naph-thol, and 5 mg of Fast Red-TR dye (Sigma, United States).
Immunohistochemical analysis.
The expression of spe-cific proteins was detected using immunofluorescenceanalysis. Cells were fixed in 3% paraformaldehyde inPBS. To inhibit the nonspecific binding and permeabilizethe cells, cell preparations were incubated in the phos-phate buffer supplemented with 0.3% Triton X-100 and4% bovine serum albumin (BSA, fraction V; Sigma, UnitedStates). The expression of
α
-fetoprotein and transcriptionfactors Oct4, GATA4, Brachyury, and Pitx2 was detectedusing antibodies to these proteins (dilution, 1 : 150), as rec-ommended by the manufacturer (Santa Cruz Biotechnol-ogy, USA; Abcam, UK; and Biocompare, USA). Immunof-luorescent detection was performed using respective sec-ondary antibodies against rabbit, goat, and mouseimmunoglobulins, conjugated with fluorochromes Alexa488 and Alexa 596 (dilution, 1 : 900; Molecular Probes,USA). The preparations were post-stained with the fluores-cent dye Hoechst 33342 to visualize cell nuclei. The controlpreparations were treated as the experimental ones, exceptthat the primary antibodies were not used. Primary mouseembryonic fibroblasts were used as a negative cell control.The preparations were specific stained and then photo-graphed using the Leica DMRXA2a image analyzing sys-tem.
Gene expression analysis.
To analyze the expression ofstudied genes total RNA was isolated from undifferenti-ated and differentiated ESCs, EGCs, ECCs, teratomas,and teratocarcinomas using the Trizol mixture (Invitrogen,United States). All DNA samples were treated with DNase(TurboDNA kit, Ambion, USA) according to the protocolrecommended by the manufacturer to prevent the DNAcontamination of samples. The efficiency of DNase treat-ment of each sample was tested in the amplification reac-tion without cDNA synthesis of RNA templates. cDNAwas synthesized on mRNA templates using the M-MuLVreverse transcriptase oligo
(
dT
)
18
primers (Fermentas,Lithuania). cDNA was synthesized using 1
μ
g of totalRNA for each sample. PCR analysis of expression of stud-ied genes was performed in the Eppendorf thermocycler(Germany) under the following conditions: preliminarydenaturation at
94°
C for 5 min, annealing at
58°
C for 45 s,elongation at
72°
C for 45 s, and denaturation at
94°
C for45 s (in total, 35 cycles). Final elongation was performedat
72°
C for 5 min. The specific primers were constructedon the basis of data on the structure of relevant genesextracted from GenBank, MGI, and Ensemble databases(table). PCR products were fractionated in agarose gel.The results of electrophoresis were recorded with a transil-luminator (Bio-Rad, USA).
RESULTS
At the initial stages of pluripotent stem cell and terato-carcinoma cell differentiation cellular spheroids (embry-oid bodies) form in the three-dimensional culture system.The role of various signaling factors of the TGF
β
family
in the regulation of pluripotent stem cell and teratocarci-noma cell differentiation was studied in the standardizedculture system, which implied the use of chemicallydefined media containing no variable components (fetalserum factors). Fetal serum in culture medium wasreplaced with the protein serum substitute (the KnockoutSerum Replacement) containing bovine albumin, aminoacids, lipids, vitamins, insulin, and transferrin (Price et al.,1998; Garcia-Gonzalo and Izpisua Belmonte, 2008). Thisapproach allowed us to correctly study the effects of vari-ous signaling proteins expressed in the cells of interest.The use of the technology of standardized embryoid bod-iesí generation in hanging-drop culture and subsequentsuspension culturing in plates, all obtained embryoid bod-ies of one cell line had nearly equal size and developedsynchronously (Fig. 1). Since the goal of this work was tostudy the mechanisms of three-dimensional differentiationof pluripotent and tumor cells, maximum standardizationof morphological parameters of embryoid bodies was ofprincipal importance for correct interpretation of experi-mental data.
Study of ESC, EGC and ECC differentiation in vitro.
It is known that one of the key factors regulating prolifer-ation and blocking differentiation of mouse ESCs in vitrois the leukemia inhibitory factor LIF. However, althoughthe mechanisms of pluripotency are extensively studied,little is known on the mechanisms in which this factor isinvolved and on their interaction with other signaling path-ways in pluripotent stem cells and teratocarcinoma cells.To elucidate the role of LIF in the regulation of undiffer-entiated ESC, EGC and ECC self-renewal, we studiedtheir growth dynamics in the presence and absence of LIF.
In the presence of LIF in the culture medium, ESCsand EGCs are maintained in undifferentiated state andalkaline phosphatase activity and expression of the Oct4transcription factor are detected in all cells (Fig. 2). Theresults of counting ESCs, EGCs, and ECCs grown in thepresence and absence of LIF showed that, after LIF with-drawal for 5 days the growth rate of ESCs and EGCsdiminished by 26 and 47%, respectively, compared to thecontrol cells grown in the LIF presence (Fig. 3). Differen-tiating cells, in which alkaline phosphatase activity wasnot detected, appeared in the populations of ESCs andEGCs growing in the LIF-free medium (Fig. 4). At thesame time, LIF had no effect on the growth of ECCs invitro, because the number of cells in the populations grow-ing in the presence and absence of LIF did not differ sig-nificantly (Fig. 3). These data indicate that a characteristicfeature of teratocarcinoma cells of the studied subline F9is that the mechanisms of regulation of their self-renewaland differentiation do not depend on LIF. On the otherhand, the comparison of the growth dynamics of ESCsand EGCs suggests that after LIF withdrawal the EGCgrowth rate have decreased in considerably higher degreethan the ESC growth. Note that the total number of ESCsand EGCs cultured for 5 days in serum-free medium con-taining LIF was significantly higher than the number ofECCs (142.3
±
2.4
×
10
4
; 127.9
±
2.9
×
10
4
, and
68.0
±
5.6
×
10
4
, respectively). Thus, it can be postulated that the
328
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY
Vol. 40
No. 6
2009
GORDEEVA et al.
Structure of primers used in gene expression analysis
Gene Sequence no. Forward and reverse primers Size, bp.
Oct-4/Pou5f1
NM_013633 5'tggagactttgcagcctgag3' 6215'catactcttctcgttgggatta3'
Nanog
NM_028016 5'caagcggtggcagaaaaac3' 7025'tggataagagcacccgactg3'
Gata6
NM_010258 5'gccaactgtcacaccacaac3' 6485'ctgcaaaagcccatctcttc3'
Gata4
NM_008092 5'ctgtgccaactgccagacta3' 4465'gcgatgtctgagtgacagga3'
Afp
NM_007423 5'agctcagcgaggagaaatgg3' 3505'caaaaggcccgagaaatctg3'
BryT
NM_009309 5'atgaaggcgcctgtgtcttt3' 4965'ttggagagctgttccgatga3'
Pitx2
NM_001042504NM_001042502NM_011098
5'gcggggaaaacatactctga35'tcgagttcacggactctcc3'
537
Nestin/Nes
NM_016701 5'ggagagtcgcttagaggtgcag3' 2785'gagagttctcagcctccagcag3'
Pax6
NM_013627 5'cagaagatcgtagagctagc3' 324/2955'gaagaactctgtttattgatgac3'
ActivinA/Inhba
NM_008380 5'acgacttttgctgccaggat3' 4265'cagtgtcttcctggctgtgc3'
Nodal
NM_013611 5'actctccaagtggctaaagg3' 4515'ttggtatcgtttcagcaggc3'
Lefty1
NM_010094 5'gctgatgtggaagggatggt3' 2825'gtgtggggacagcctctttt3'
Lefty2
NM_177099 5'ctggacctcaaggactacgg3' 2445'tctctgaggcaacacactgc3'
Tdgf1
NM_011562 5'ttgggaccagaaagaacctg3' 2425'tgcacagggaacacttcttg3'
Tgfb1
NM_011577 5'taccttggtaaccggctgct3' 4355'gggtcccagacagaagttgg3'
Bmp4
NM_007554 5'gggaaccgggcttgagta3' 4695'tcggctgattctgacatgct3'
Gdf3
NM_008108 5'gccttcacctcacaggttcc3' 4065'agggacagaccgcagaaaaa3'
Acvr1b
NM_007395 5'agacgctccaggatctcgtc3' 3025'ctgggtccaggtgccattat3'
Acvr2b
NM_007397 5'gcttcaggaggtggttgtcc3' 2415'gagcaggtccacattggtga3'
Tgfbr1
NM_009370 5'aaatggcggggagaagaagt3' 3265'tgggcaatagctggttttcc3'
Bmpr1a
NM_009758 5'attcaccgaaagcccagcta3' 3855'acctgccgaaccatctgaat3'
Smad2
NM_010754 5'tgagctcaaggcaatcgaaa3' 2505'tccaggtggtggtgtttctg3'
Smad4
NM_008540 5'atgagcgggttgtctcacct3' 2835'atactggccggctgacttgt3'
Smad5
NM_008541 5'gttggggaagcttttcatgc3' 2885'gcagctgctggggatcttac3'
Hprt
NM_013556 5'-gctggtgaaaaggacctct-3' 249 5'-cacaggactagaacacctgc-3'
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY
Vol. 40
No. 6
2009
REGULATION OF in vitro AND in vivo DIFFERENTIATION OF MOUSE 329
mechanisms of regulation of self-renewal and initialstages of differentiation of ESCs, EGCs and ECCs signif-icantly differ.
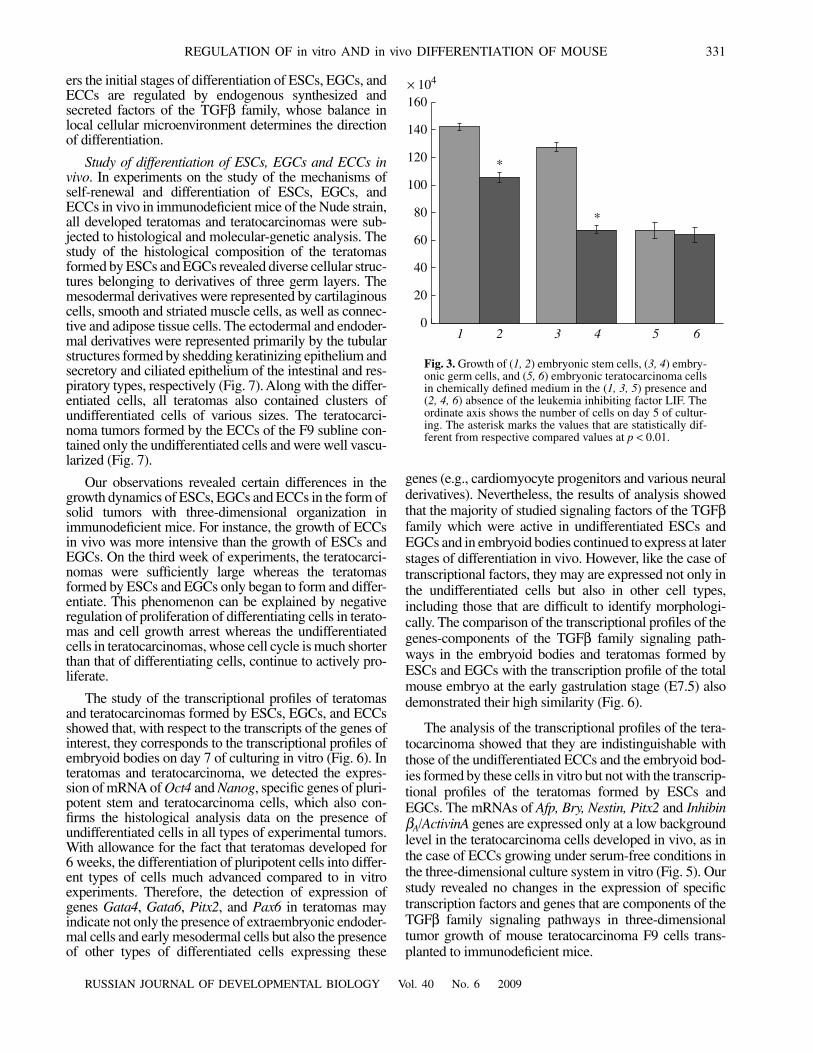
Then, to initiate three-dimensional differentiation ofESCs, EGCs, and ECCs, we cultured them in hangingdrops for 3 days in the absence of LIF. Morphologicalanalysis showed that all formed embryoid bodies (EB1)consisted of uniform cells in which a high alkaline phos-phatase activity was detected. On day 5 of culturing, theouter surface of the embryoid bodies (EB5) formed byESCs and EGCs contained a small number of extraembry-onic endodermal cells which do not express alkaline phos-phatase but express the transcription factor Gata4 and
α
-fetoprotein (Fig. 5). The appearance of these factors inthe surface cells of embryoid bodies is indicative of differ-entiation of parietal and visceral extraembryonic endoder-mal cells. Note that the maximum number of extraembry-onic endodermal cells was revealed in ESC culture whilein EGC and ECC cultures extraembryonic endodermalcell number was lower and moreover,
α
-fetoprotein-posi-tive cells were not detected in EGCs and ECCs at all. Theresults of histochemical and immunofluorescent analysesalso showed that, on day 5 of culturing, the majority of
cells in EBs of all three lines still expressed alkaline phos-phatase and the transcription factor Oct4. Our results dem-onstrated that, even on day 7 of cell differentiation in theserum-free medium in the absence of LIF (EB5), themajor part of both ESC and EGC populations remainsundifferentiated. These data indicate that, in addition tothe LIF/Stat3 signaling cascade, other signaling pathwayseffectively inhibiting early differentiation stages areinvolved in the regulation of pluripotent stem cell self-renewal.
To establish the role of TGF
β
family factors in the reg-ulation of self-renewal and differentiation of ESCs, EGCsand ECCs we studied the expression profiles of the genesthat are components of signaling pathways—factorsActivinA, Nodal, Lefty1, 2, GDF3, TGF
β
1
, BMP4 andcofactor Tdgf1/—ripto; their receptors Actr1, Actr2,Bmpr1, and Tdgf1/Cripto; signal transducing proteinsSmad2, Smad4, and Smad5; as well as specific and tran-scription factors Oct4, Nanog, Gata4, Gata6, Afp, Bry,Pitx2, Nestin, and Pax6, which are differentially expressedin the progenitor cells of three germ layers (Fig. 6). A com-parative analysis of transcriptional profiles showed thatdifferentiation of ESCs, EGCs, and ECCs in the three-
c d
a b
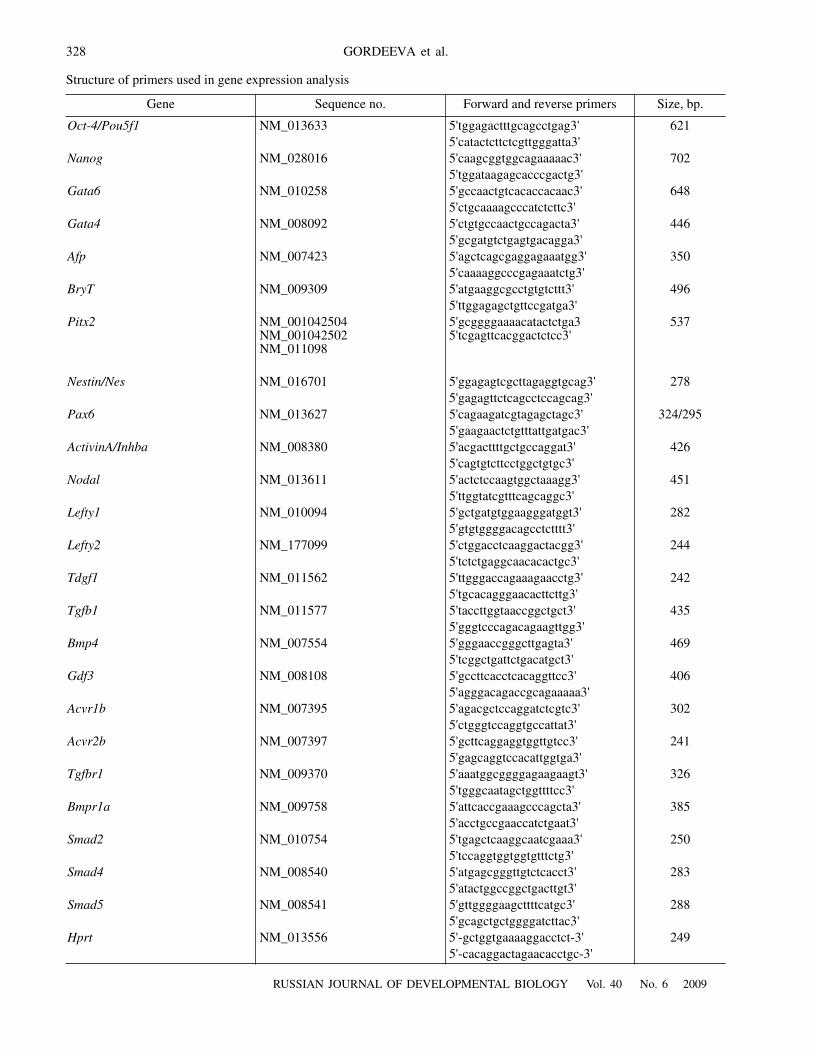
Fig. 1.
Formation of standardized embryoid bodies from embryonic stem cells (ESCs), embryonic germ cells (EGCs), and embryoicteratocarcinoma cells (ECCs): (
a
) formation of embryoid bodies from cell suspension in a hanging drop culture; (b–d) the embryoidbodies formed by ESCs, EGCs, and ECCs on the third day of culturing. Scale, 200 μm.

330
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
GORDEEVA et al.
dimensional culture grown in the serum-free medium isaccompanied by slight changes in the expression levels ofOct4 and Nanog genes, which encode specific transcrip-tion factors for pluripotent cells. This agrees with the mor-phological observations. On the other hand, it was estab-lished that the expression of mRNA of Gata4, Gata6 andAfp, specific genes of extraembryonic visceral and parietalendoderm, is detected in embryoid bodies formed byESCs; however, the level of transcripts of these genes inthe embryoid bodies formed by EGCs and ECCs was verylow. A low background level of transcription of Bry 2 andPitx2, specific genes of early mesodermal progenitors,was also detected in all embryoid bodies; however, immu-nohistochemical analysis did not reveal the expression ofBry and Pitx2 proteins in any line at any stage. At the sametime, differentiation of ESCs and EGCs was accompaniedby an increase in the level of expression of mRNA of Nes-tin and Pax6, characteristic genes of early neuroendoder-mal cells. Interestingly, the expression of two transcriptsof the Pax6 gene corresponding to two splicing variantswas detected in ECCs whereas mRNA of the Nestin genewas expressed at a very low background level.
Analysis of transcriptional profiles revealed no signifi-cant changes in the levels of mRNA expression of themajority of genes encoding signaling ligands, their recep-
tors, and transducers, except for the Inhibin βA gene,encoding the factor ActivinA subunits. The expression ofthe Inhibin βA/ActivinA gene drastically decreased in theembryoid bodies formed by ESCs and EGCs compared tothe undifferentiated cells and was completely absent in theteratocarcinoma cells at all stages studied. It should benoted that the changes in the expression of the majority ofsignaling factors and specific transcription factors werestrongly correlated. In view of this, it can be assumed thata considerable change in the Inhibin βA/ActivinA geneexpression is an early initiating event in the course ofspontaneous three-dimensional differentiation of pluripo-tent stem cells. This assumption is based on the fact that,in this case, a direct correlation between the expression ofmRNA of the Inhibin β /ActivinA gene in the undifferenti-ated ESCs and EGCs and the presence of the LIF factor inthe culture medium are observed. Conversely, the mecha-nisms of regulation of self-renewal of teratocarcinomacells depended neither on the LIF nor on the ActivinA fac-tor. At the same time, the results of our study showed that,when ECCs were cultured in a serum-free medium, theydifferentiated only into the cells of the primary extraem-bryonic endoderm and were unable to differentiate into theprogenitors of three germ layers. Thus, our data provideevidence that in the absence of exogenous signaling induc-
d e f
a b c
Fig. 2. (a–c) Alkaline phosphatase activity and (d–f) expression of the specific transcription factor Oct4 in undifferentiated (a, d)embryonic stem cells, (b, e) embryonic germ cells, and (c, f) embryonic teratocarcinoma cells of the mouse. Scale, 100 μm.
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
REGULATION OF in vitro AND in vivo DIFFERENTIATION OF MOUSE 331
ers the initial stages of differentiation of ESCs, EGCs, andECCs are regulated by endogenous synthesized andsecreted factors of the TGFβ family, whose balance inlocal cellular microenvironment determines the directionof differentiation.
Study of differentiation of ESCs, EGCs and ECCs invivo. In experiments on the study of the mechanisms ofself-renewal and differentiation of ESCs, EGCs, andECCs in vivo in immunodeficient mice of the Nude strain,all developed teratomas and teratocarcinomas were sub-jected to histological and molecular-genetic analysis. Thestudy of the histological composition of the teratomasformed by ESCs and EGCs revealed diverse cellular struc-tures belonging to derivatives of three germ layers. Themesodermal derivatives were represented by cartilaginouscells, smooth and striated muscle cells, as well as connec-tive and adipose tissue cells. The ectodermal and endoder-mal derivatives were represented primarily by the tubularstructures formed by shedding keratinizing epithelium andsecretory and ciliated epithelium of the intestinal and res-piratory types, respectively (Fig. 7). Along with the differ-entiated cells, all teratomas also contained clusters ofundifferentiated cells of various sizes. The teratocarci-noma tumors formed by the ECCs of the F9 subline con-tained only the undifferentiated cells and were well vascu-larized (Fig. 7).
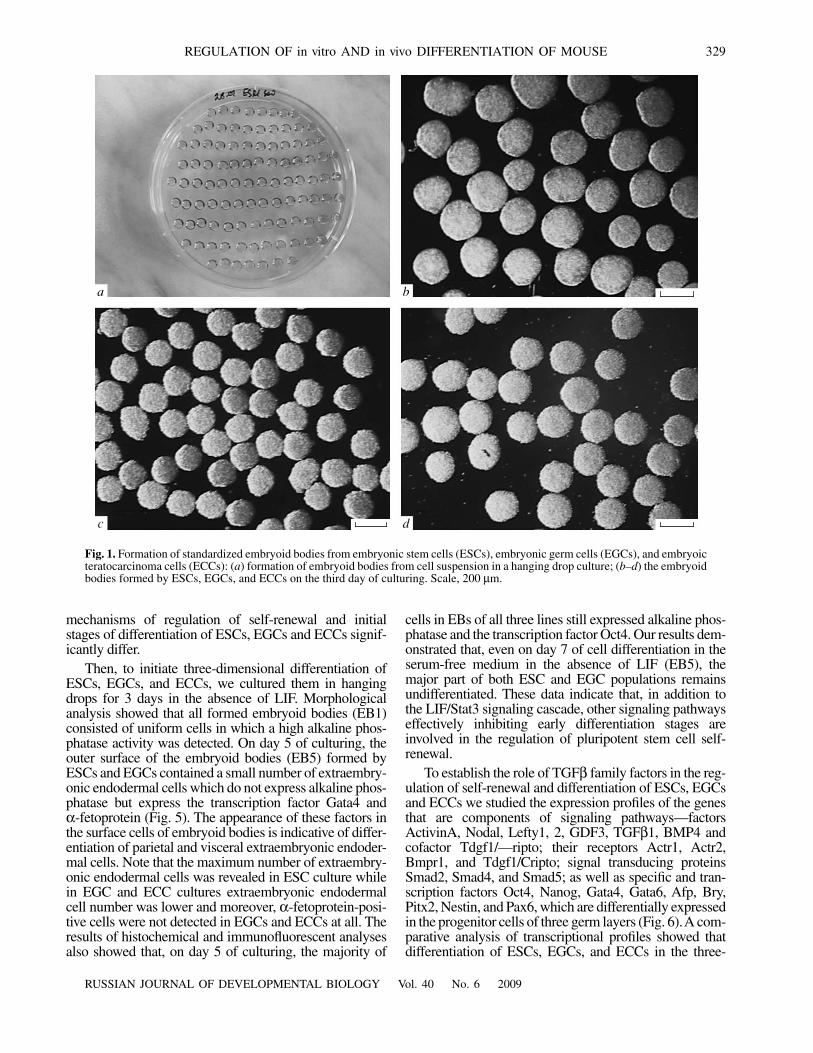
Our observations revealed certain differences in thegrowth dynamics of ESCs, EGCs and ECCs in the form ofsolid tumors with three-dimensional organization inimmunodeficient mice. For instance, the growth of ECCsin vivo was more intensive than the growth of ESCs andEGCs. On the third week of experiments, the teratocarci-nomas were sufficiently large whereas the teratomasformed by ESCs and EGCs only began to form and differ-entiate. This phenomenon can be explained by negativeregulation of proliferation of differentiating cells in terato-mas and cell growth arrest whereas the undifferentiatedcells in teratocarcinomas, whose cell cycle is much shorterthan that of differentiating cells, continue to actively pro-liferate.
The study of the transcriptional profiles of teratomasand teratocarcinomas formed by ESCs, EGCs, and ECCsshowed that, with respect to the transcripts of the genes ofinterest, they corresponds to the transcriptional profiles ofembryoid bodies on day 7 of culturing in vitro (Fig. 6). Interatomas and teratocarcinoma, we detected the expres-sion of mRNA of Oct4 and Nanog, specific genes of pluri-potent stem and teratocarcinoma cells, which also con-firms the histological analysis data on the presence ofundifferentiated cells in all types of experimental tumors.With allowance for the fact that teratomas developed for6 weeks, the differentiation of pluripotent cells into differ-ent types of cells much advanced compared to in vitroexperiments. Therefore, the detection of expression ofgenes Gata4, Gata6, Pitx2, and Pax6 in teratomas mayindicate not only the presence of extraembryonic endoder-mal cells and early mesodermal cells but also the presenceof other types of differentiated cells expressing these
genes (e.g., cardiomyocyte progenitors and various neuralderivatives). Nevertheless, the results of analysis showedthat the majority of studied signaling factors of the TGFβfamily which were active in undifferentiated ESCs andEGCs and in embryoid bodies continued to express at laterstages of differentiation in vivo. However, like the case oftranscriptional factors, they may are expressed not only inthe undifferentiated cells but also in other cell types,including those that are difficult to identify morphologi-cally. The comparison of the transcriptional profiles of thegenes-components of the TGFβ family signaling path-ways in the embryoid bodies and teratomas formed byESCs and EGCs with the transcription profile of the totalmouse embryo at the early gastrulation stage (E7.5) alsodemonstrated their high similarity (Fig. 6).
The analysis of the transcriptional profiles of the tera-tocarcinoma showed that they are indistinguishable withthose of the undifferentiated ECCs and the embryoid bod-ies formed by these cells in vitro but not with the transcrip-tional profiles of the teratomas formed by ESCs andEGCs. The mRNAs of Afp, Bry, Nestin, Pitx2 and InhibinβA/ActivinA genes are expressed only at a low backgroundlevel in the teratocarcinoma cells developed in vivo, as inthe case of ECCs growing under serum-free conditions inthe three-dimensional culture system in vitro (Fig. 5). Ourstudy revealed no changes in the expression of specifictranscription factors and genes that are components of theTGFβ family signaling pathways in three-dimensionaltumor growth of mouse teratocarcinoma F9 cells trans-planted to immunodeficient mice.
× 104
160
140
120
100
80
60
40
20
01 2 3 4 5 6
*
*
Fig. 3. Growth of (1, 2) embryonic stem cells, (3, 4) embry-onic germ cells, and (5, 6) embryonic teratocarcinoma cellsin chemically defined medium in the (1, 3, 5) presence and(2, 4, 6) absence of the leukemia inhibiting factor LIF. Theordinate axis shows the number of cells on day 5 of cultur-ing. The asterisk marks the values that are statistically dif-ferent from respective compared values at p < 0.01.

332
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
GORDEEVA et al.
DISCUSSION
Pluripotent stem and teratocarcinoma cell lines areunique cellular models which provide an opportunity toelucidate the role of signaling and specific transcriptionfactors in the regulation of pluripotent state maintenanceand cell fate determination of pluripotent cells and theirmalignant counterparts. The study of these issues in earlydevelopment is methodically restricted by the low cellnumber in early mammalian embryos. In our experimentswe used pluripotent stem cell lines of diverse origin, ESCsand EGCs, differing in the ontogenetic and epigeneticstates of progenitor cells, as well as the embryonic terato-carcinoma line with genetic failures. Our research wasfocused on studying the signaling pathways which areactivated in these lines when cells begin to differentiate.We performed a comparative analysis of transcriptionalprofiles to reveal the main regulatory principles of differ-ent branches of the TGFβ family signaling pathwaysinvolved in the pluripotent state maintenance and the in theinitiation of differentiation. To correctly interpret theexperimental data, we used serum-free standard media toexclude the effect of exogenous factors of fetal serumpresent in the conventional culture systems. We also usedthree-dimensional systems for culturing in vitro andgrowth in vivo, because the early embryogenesis events
are reproduced most adequately in such systems. Themodeling of differentiation in vitro and in vivo are a com-plementary approaches, because the possibilities of mod-eling of differentiation of pluripotent cells in vitro are lim-ited; however, in vitro differentiation is regulated moreprecisely in experiments. At the same time, the study offormation of experimental teratomas provides an opportu-nity to investigate cell differentiation at more advancedstages. Teratogenesis initiated by the transplantation ofESCs and EGCs into immunodeficient animals is a modelof incomplete and discoordinated embryogenesis, whichdemonstrates that pluripotent cells differentiate asynchro-nously, with preserving a certain number of undifferenti-ated cells whose nature has been studied insufficiently. Itwas assumed that pluripotent cells remaining undifferenti-ated during differentiation in vitro and in vivo in teratomasare genetically abnormal cells undergoing transformationand malignization. Another standpoint supports thehypothesis on a long-term self-sustaining population ofpluripotent cells that differentiate very slowly. To clarifythis issue, we studied transcription profiles, includingthose of the specific transcriptional factors—markers forearly embryonic cells, both undifferentiated ESCs, EGCs,and ECCs, as well as the pluripotent cells differentiating invitro and in vivo. In analyzing the expression of these
d e f
a b c
Fig. 4. Alkaline phosphatase activity in (a, d) ESCs, (b, e) EGCs, and (c, f) ECCs growing in chemically defined medium in the(a, b, c) presence and (d, e, f) absence of the leukemia inhibiting factor LIF. Scale, 100 μm.
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
REGULATION OF in vitro AND in vivo DIFFERENTIATION OF MOUSE 333
j k l
g h i
d e f
a b c
Fig. 5. Expression of specific factors in the embryoid bodies formed by (a, d, g, j) ESCs, (b, e, h, k) EGCs, and (c, f, i, l) ECCs onday 5 of culturing in three-dimensional serum-free culture system (EB5). Designations: (a–c), alkaline phosphatase activity;(d−f), expression of the Oct4 transcription factor; (g–i), expression of the Gata4 transcription factor; and (j–l), expression of α-feto-protein. Scale, 100 μm.

334
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
GORDEEVA et al.
Oct4
Nanog
Gata6
Gata4
Afp
Bry
Pitx2
Nestin
Pax6
ActivinA
Nodal
Lefty1
Lefty2
Tdgf
Tgfb1
Bmp4
Gdf3
Acvr1b
Acvr2b
Tgfbr1
Bmpr1a
Smad2
Smad4
Smad5
Hprt
ESCs
ESCs EBs1 EBs5
EGCs
EGCs EBs1 EBs5
ETCs
ETCs EBs1 EBs5 T(ESCs) Ö7.5 MEFsT(EGCs)T(ETCs)
Fig. 6. Transcriptional profiles of ESCs, EGCs, and ECCs, the embryoid bodies formed by these cells in vitro, as well as the terato-mas and teratocarcinoma formed in vivo, including the genes that are components of the signaling pathways of the TGFβ familyfactors and the specific genes that are active in the extraembryonic endoderm and early progenitor cells of three germ layers. Des-ignations: EB1 and EB5, embryoid bodies formed in suspension culture on the first and fifth days of culturing, respectively;T(ESCs), T(EGCs), and T(ECCs), teratomas and teratocarcinomas formed by ESCs, EGCs, and ECCs, respectively, after transplan-tation under the renal capsule of immunodeficient mice; E7.5, mouse embryo cDNA at the stage of day 7.5 of development (positivecontrol); MEFs, mouse embryonic fibroblasts (negative control).

RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
REGULATION OF in vitro AND in vivo DIFFERENTIATION OF MOUSE 335
genes, we performed simultaneous monitoring of changesin expression of signaling ligands of the TGFβ family,their receptors, and intracellular signal transducers toreveal correlations between them.
The revealed similarity between the transcriptionalprofiles of the cell lines at different stages of differentia-tion and the expression profile of the mouse embryo at theearly gastrulation stage E7.5 indicates that the earliestectodermal, endodermal, and mesodermal progenitor cellsdifferentiate in the embryoid bodies of ESCs and EGCs inthe first week of culturing in the serum-free medium in theabsence of LIF.
It should be noted that, despite the similarity of tempo-ral parameters, the morphogenetic processes during theformation of embryoid bodies in this culture system pro-ceeded much more slowly than in the embryo. However,
no changes in cell morphology and transcriptional profileswere observed at this stage in the embryoid bodies formedby ECCs. As was mentioned above, expression profiles ofstudied genes in teratomas and teratocarcinomas corre-sponded to embryoid bodies on day 7 of culturing (Fig. 5,EB5) and the mouse embryo profile at stage E7.5,although the level of expression of Gata4, Afp, Bry andPitx2 genes was significantly higher in teratomas than inthe embryoid bodies formed by ESCs and EGCs. Withallowance for the fact that the duration of development ofteratomas was almost twice longer than that of mouseembryos (6 weeks versus 20 days), the presence in terato-mas of undifferentiated Oct4- and Nanog-positive cells aswell as early progenitor cells of three germ layers andextraembryonic structures indicates that differentiation ofpluripotent cells at ectopic sites in vivo is accompanied bya decrease of their growth rate, although they continue to
g h i
d e f
a b c
Fig. 7. Histological analysis of teratomas and teratocarcinomas formed by (a–c, g) ESCs, (d–f, h) EGCs, and (i) ECCs after thetransplantation under the renal capsule of immunodeficient mice. Undifferentiated cells were present in the teratomas formed by(g) ESCs, (h) EGCs, and (i) ETCs. Various three-dimensional structures belonging to derivatives of three germ layers developed inthe teratomas formed by ESCs, and EGCs. The mesodermal derivatives were represented by (f) cartilaginous cells, (c, e, f) smoothand striated muscle cells, and (a, b, d, e) connective and adipose tissue cells. The ectodermal and endodermal derivatives were rep-resented by the tubular structures formed by (b, d) shedding keratinizing epithelium or (a, e) secretory and ciliated epithelium ofthe intestinal and respiratory types. Scale, 100 μm.

336
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
GORDEEVA et al.
actively grow. The mRNAs for all signaling proteins of theTGFβ family, such as ActivinA, Nodal, Lefty1, Lefty2,GDF3, TGFβ1, BMP4, and cofactor TDGF1/Cripto; theirreceptors Actr1, Actr2, Bmpr1, and Tβr1; and signal trans-ducers Smad2, Smad4, and Smad5 continue to express atlevels comparable to those of undifferentiated cells. This isindicative of interference of the profiles of undifferentiatedand differentiated cells.
It was shown earlier that the maintenance of pluripo-tency is ensured not only by the transcription factors Oct4and Nanog and the LIF/JAK/STAT3 signaling pathwaysbut also various branches of signaling pathways of factorsof the TGFβ family. For instance, it is known that theBMP4 factor can be regarded as an antagonist of the neu-roectoderm development in mouse embryo. Possibly, thisexplains the fact that the cooperation of BMP4 and LIF inmouse ESCs potentiates the effect of differentiation sup-pression (Ying et al., 2003). It was assumed that theirinteraction results in an increase in the pool of Smad4 pro-teins, which, in turn, activate the expression of genesencoding differentiation inhibitors of the Id family. Thiseffect is enhanced in the presence of fetal serum factors.However, in the absence of LIF, BMP4 activates predom-inantly the intracellular flow of signal transducers Smad1,Smad3, and Smad5 and enhances differentiation of pluri-potent cells into ectodermal and mesodermal progenitorcells (Watanabe and Miyazono, 2008).
Another branch of TGFβ family signaling pathways isinitiated by Activin, Nodal, Lefty1, and Lefty2 factors aswell as by the Tdgf1/Cripto coreceptor, which is involvedin the formation of a complex with activin receptorsActRII and ActRI for the activation of the Nodal signalingpathway or for its inhibition by factors Lefty1 and Lefty2.Transforming growth factors (TGFβ1, TGFβ2, andTGFβ3) bind to their own receptors Tβ1RI and RII, and,similarly to Activin/Nodal, activate the intracellular signalflows of Smad2 and Smad3 proteins by phosphorylatingthem; the latter form a complex with Smad4 and are trans-located into the cell nuclei, where they initiate the expres-sion of target genes (Shi and Massague, 2003). For thepluripotent cells of the inner cell mass of mouse blasto-cyst, it was shown that the absence of transcription of thegenes encoding proteins Smad2 and Smad3 and the block-ade of phosphorylation of these factors by the syntheticinhibitor SB 431542 leads to a decrease in the level ofOct4 expression and subsequent reduction of the epiblast(Dunn et al., 2004). Although the inhibition of the pool ofsignal transducers Smad2 and Smad3 did not change sig-nificantly the level of expression of the transcription factorOct4 and morphological changes in the population ofmouse ESCs (Sato et al., 2004; Ogawa et al., 2007), it con-siderably decreased the proliferation level. On the otherhand, it was established that the addition of factors Activinor Nodal and FGF to the culture medium for human ESCsresulted in the inhibition of their differentiation and stablemaintenance of the pluripotent state (Vallier et al., 2005;Xu et al., 2005). Conversely, the inhibition of signals fromthe Activin/Nodal receptors stimulated differentiation ofhuman ESCs and decreased the number of Oct4-positive
cells even in the presence of FGF (Vallier et al., 2005;James et al., 2005) in the same manner as factors BMP,TGFβ, and Cripto influenced human ESCs. It was shownthat signaling cascades of FGF and TGFβ/Activin factorsexhibit synergism in the inhibition of BMP4 signalingpathways and are able to maintain a high expression levelof OCT4, NANOG and SOX2 genes, because proteinsSMAD2 and SMAD3 can bind to the proximal promoterof the NANOG gene and activate its expression (Xu et al.,2008). All these data demonstrate that various factors ofthe TGFβ family are involved in the regulation of mainte-nance of the pluripotent state of cells both in early embryoand in mouse and human
ESCs. In this study we also established that culturingwithin 5–7 days in the serum-free culture system in theLIF absence resulted in a considerable suppression of thegrowth of ESCs and EGCs and expression of alkalinephosphatase in these cells as well as a considerabledecrease in the expression of factor ActivinA, whereas theexpression of other factors of the TGFβ family, theirreceptors, and proteins Smad2, Smad4, and Smad5 almostdoes not change. These results indicate that, as in humanESCs, factor ActivinA plays an important role in main-taining the pluripotent status of mouse ESCs and, mostlikely, is involved in the regulation of pluripotency inmouse ESCs in cooperation with LIF. At the same time, interatocarcinoma cells, factor ActivinA is not expressed atall and, similarly to LIF, is not involved in the regulationof their self-renewal. Apparently, other signaling path-ways play the key role in the regulation of proliferation ofthe nullipotent teratocarcinoma, which stably inhibit theirdifferentiation into the cells of three germ layers but notinto the primary extraembryonic endoderm.
It is known that factor ActivinA not only inhibits differ-entiation of pluripotent cells but, in cooperation with otherfactors, effectively stimulates their differentiation. Forinstance, ActivinA together with Nodal and Wnt regulatethe formation of the primitive streak at the gastrulationstage and differentiation of early mesodermal progenitorsfrom ESCs (Nostro et al., 2008). In addition, human ESCsdifferentiate into the mesoendoderm and definitive endo-derm as a result of stimulation of the signaling pathwaysof ActivinA/Nodal factors, though only if the phosphati-dylinositol 3-kinase (PI3K) signaling pathway, induced byinsulin or the insulin growth factor (IGF), is inhibited(McLean et al., 2007). It was also shown that the in vitrodifferentiation of ESCs into early hemopoietic cells isinduced only under the influence of BMP and further acti-vation of the ActivinA/Nodal and Wnt signaling pathwaysonly at a certain stage (Nostro et al., 2008).
The analysis of the transcriptional profiles of ESCs,EGCs, and ECCs at early stages of their differentiation invitro as well as in teratomas and teratocarcinoma showedthat all factors of the TGFβ family required for inductionof differentiation into different cell types are expressed inESCs and EGCs and almost all these factors, except forActivinA, are present in ECCs. However, despite this fact,the differentiation of ESCs and EGCs even for 6 weeks of

RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
REGULATION OF in vitro AND in vivo DIFFERENTIATION OF MOUSE 337
growth in vivo remains incomplete, because teratomascontain not only differentiated but also residual undiffer-entiated cells. Our results demonstrated that ownresources of ESCs and EGCs which express and secretethe signaling factors of the TGFβ family are insufficientfor normal complete differentiation of cells and morpho-genesis of definitive structures. The growth and differenti-ation factors present in the blood serum of the recipientmice, which are delivered to teratomas and teratocarcino-mas owing to intensive vascularization of tumors, are alsounable to compensate for this deficiency. Apparently, invivo, serum factors even accelerate the proliferation of ter-atocarcinoma cells and stimulate tumor growth rather thanstimulate their differentiation.
Drawing an analogy with embryonic development, itcan be assumed that the main cause of incomplete differ-entiation of pluripotent stem cells in vitro and in vivo is thealteration of the spatiotemporal pattern of gene expressionin “artificial” pluripotent cell systems with disturbance ofnatural concentration gradients of signaling factors. Denovo creation of defined combinations and gradients ofsignaling inducers in culture medium, within a certaintemporal framework and in certain sequences, makes itpossible to reconstruct some histogenetic events andobtain required cellular elements for basic and appliedresearch.
ACKNOWLEDGMENTS
This study was supported by the Russian Founda-tion for Basic Research (project no. 08-04-01307a).
REFERENCES
Adewumi, O., Aflatoonian, B., Ahrlund-Richter, L., et al.,Characterization of Human Embryonic Stem Cell Lines bythe International Stem Cell Initiative, Nat. Biotech., 2007,vol. 25, pp. 803–816. Allegrucci, C. and Young, L.E., Differences between HumanEmbryonic Stem Cell Lines, Hum. Repr. Update, 2007,vol. 13, no. 2, pp. 103–120. Andrews, P.W., From Teratocarcinomas to Embryonic StemCells, Philos. Trans. R. Soc. L. B. Biol. Sci., 2002, vol. 357,pp. 405–417. Aoi, T., Yae, K., Nakagawa, M., et al., Generation of Pluripo-tent Stem Cells from Adult Mouse Liver and Stomach Cells,Science, 2008, vol. 321, pp. 699–702. Beddington, R.S.P. and Robertson, E.J., Axis Developmentand Early Asymmetry in Mammals, Cell, 1999, vol. 96,pp. 195–209. Blelloch, R.H., Hochedlinger, K., Yamada, Y., et al., NuclearCloning of Embryonal Carcinoma Cells, Proc. Natl. Acad.Sci. USA, 2004, vol. 101, pp. 13985–13990. Brandenberger, R., Wei, H., Zhang, S., et al., TranscriptomeCharacterization Elucidates Signaling Networks That Con-trol Human ES Cell Growth and Differentiation, Nat. Bio-technol., 2004, vol. 22, pp. 707–716. Calhoun, J.D., Rao, R.R., Warrenfeltz, S., et al., Transcrip-tional Profiling of Initial Differentiation Events in Human
Embryonic Stem Cells, Biochem. Biophys. Res. Commun.,2004, vol. 323, pp. 453–464. Cibelli, J.B., Grant, K.A., Chapman, K.B., et al., Parthenoge-netic Stem Cells in Nonhuman Primates, Science, 2002,vol. 295, p. 819. Derynck, R. and Miyazono, K., TGF-β and the TGF-β Fam-ily, in The TGF-β Family, Derynck, R. and Miyazono, K.,Eds., New York: Cold Spring Harbor Lab. Press, 2008,pp. 29–43. Dunn, N.R., Vincent, S.D., Oxburgh, L., et al., CombinatorialActivities of Smad2 and Smad3 Regulate Mesoderm Forma-tion and Patterning in the Mouse Embryo, Development,2004, vol. 131, pp. 1717–1728. Evans, M.J. and Kaufman, M.H., Establishment in Culture ofPluripotential Cells from Mouse Embryos, Nature, 1981,vol. 292, pp. 154–156. Garcia-Gonzalo, F.R., Izpisua Belmonte J.C., Albumin-Associated Lipids Regulate Human Embryonic Stem CellSelf-Renewal, PlosOne, 2008, vol. 1, p. e1384. Gordeeva, O.F, Pluripotent Cells in Embryogenesis and inTeratoma Formation, in Stem Cells and Cancer,Parsons, D.W., Ed., New York: Nova Sci. Publ. Ink., 2007,pp. 62–85. Guan, K., Nayernia, K., Maier, L.S., et al., Pluripotency ofSpermatogonial Stem Cells from Adult Mouse Testis,Nature, 2006, vol. 440, pp. 1199–1203. James, D., Levine, A.J., Besser, D., and Hemmati-Brivanlou, A.,TGFβ/Activin/Nodal Signaling is Necessary for the Mainte-nance of Pluripotency in Human Embryonic Stem Cells, Devel-opment, 2005, vol. 132, pp. 1273–1282. Kanatsu-Shinohara, M., Inoue, K., Lee, J., et al., Generationof Pluripotent Stem Cells from Neonatal Mouse Testis, Cell,2004, vol. 119, pp. 1001–1012. Katagiri, T., Suda, T., and Miyazono, K., The Bone Morpho-genetic Proteins, in The TGF-β Family, Derynck, R. andMiyazono, K., Eds., New York: Cold Spring Harbor Lab.Press, 2008, pp. 121–149. Kimura, T., Suzuki, A., Fujita, Y., et al., Conditional Loss ofPTEN Leads to Testicular Teratoma and Enhances Embry-onic Germ Cell Production, Development, 2003, vol. 130,pp. 1691–1700. Lin, G., OuYang, Q., Zhou, X., et al., A Highly Homozygousand Parthenogenetic Human Embryonic Stem Cell LineDerived from a One-Pronuclear Oocyte Following in vitroFertilization Procedure, Cell Res., 2007, vol. 17, pp. 999–1007. Maherali, N., Sridharan, R., Xie, W., et al., Directly Repro-grammed Fibroblasts Show Global Epigenetic Remodellingand Widespread Tissue Contribution, Cell Stem Cell, 2007,vol. 1, pp. 55–70. Martin, G.R., Isolation of Pluripotent Cell Line from EarlyMouse Embryo Cultured in Medium Conditioned by Terato-carcinoma Stem Cells, Proc. Natl. Acad. Sci. USA, 1981,vol. 78, pp. 7634–7638. Matsui, Y., Zsebo, K., and Hogan, B.L., Derivation of Pluri-potential Embryonic Stem Cells from Murine PrimordialGerm Cells in Culture, Cell, 1992, vol. 70, pp. 841–847. McLean, A.B., Kevin, A.D., Amour, K.A., et al., ActivinAEfficiently Specifies Definitive Endoderm from HumanEmbryonic Stem Cells Only when Phosphatidylinositol3-Kinase Signaling is Suppressed, Stem Cells, 2007, vol. 25,pp. 29–38.

338
RUSSIAN JOURNAL OF DEVELOPMENTAL BIOLOGY Vol. 40 No. 6 2009
GORDEEVA et al.
Miura, S., Whitman, M., and Mishina, Y., TGF-β Family Sig-naling in Early Postimplantation Development of the Mouse,in The TGF-β Family, Derynck, R. and Miyazono, K., Eds.,New York: Cold Spring Harbor Lab. Press, 2008, pp. 461–492. Nagy, A., Rossant, J., Nagy, R., et al., Embryonic Stem CellsAlone Are Able to Support Fetal Development in the Mouse,Proc. Natl. Acad. Sci. USA, 1993, vol. 90, pp. 8424–8428. Nostro, C.M., Cheng, X., Keller, G.M., and Gadue, P., Wnt,Activin, and BMP Signaling Regulate Distinct Stages in theDevelopmental Pathway from Embryonic Stem Cells toBlood, Cell Stem Cell, 2008, vol. 2, pp. 60–71. Ogawa, K., Saito, A., Matsui, H., et al., Activin-Nodal Sig-naling is Involved in Propagation of Mouse Embryonic StemCells, J. Cell Sci., 2007, vol. 120, pp. 55–65. Okita, K., Ichisaka, T., and Yamanaka, S., Generation of Ger-mline-Competent Induced Pluripotent Stem Cells, Nature,2007, vol. 448, pp. 313–317. Price, P.J., Goldsborough, M.D., and Tilkins, M.L., Embry-onic Stem Cell Serum Replacement, Internat. Patent Appli-cation WO, 1998, no. 98/30679. Resnick, J.L., Bixler, L.S., Cheng, L., and Donovan, P.J.,Long-Term Proliferation of Mouse Primordial Germ Cells inCulture, Nature, 1992, vol. 359, pp. 550–551. Rohwedel, J., Sehlmeyer, U., Shan, J., et al., PrimordialGerm Cell-Derived Mouse Embryonic Germ (EG) Cells invitro Resemble Undifferentiated Stem Cells with Respect toDifferentiation Capacity and Cell Cycle Distribution, CellBiol. Internat., 1996, vol. 20, pp. 579–587. Sato, N., Meijer, L., Skaltsounis, L., et al., Maintenance ofPluripotency in Human and Mouse Embryonic Stem CellsThrough Activation of Wnt Signaling by a PharmacologicalGSK-3-Specific Inhibitor, Nat. Med., 2004, vol. 10,pp. 55−63. Schilling, S.H., Hjilmeland, A.B., Rich, J.N., and Wang, X.-F.,TGF-β: a Multipotential Cytokine, in The TGF-β Family,Derynck, R. and Miyazono, K., Eds., New York: Cold SpringHarbor Lab. Press, 2008, pp. 45–77. Shamblott, M.J., Axelman, J., Wang, S., et al., Derivation ofPluripotent Stem Cells from Cultured Human PrimordialGerm Cells, Proc. Natl. Acad. Sci. USA, 1998, vol. 95,pp. 13726–13731. Sharova, L.V., Sharov, A.A., Piao, Y., et al., Global GeneExpression Profiling Reveals Similarities and Differencesamong Mouse Pluripotent Stem Cells of Different Originsand Strains, Devel. Biol., 2007, vol. 307, pp. 446–459. Shi, Y. and Massague, J., Mechanisms of TGF-β Signalingfrom Cell Membrane to the Nucleus, Cell, 2003, vol. 13,pp. 685–700.
Siegel, P.M. and Massague, J., Growth Control by TGF-β:Mechanisms Controlling Cell Cycle Progression and Apop-tosis, in The TGF-β Family, Derynck, R. and Miyazono, K.,Eds., New York: Cold Spring Harbor Lab. Press, 2008,pp. 333–362.
Takahashi, K. and Yamanaka, S., Induction of PluripotentStem Cells from Mouse Embryonic and Adult FibroblastCultures by Defined Factors, Cell, 2006, vol. 126, pp. 663–676.
Takahashi, K., Tanabe, K., Ohnuki, V., et al., Induction ofPluripotent Stem Cells from Adult Human Fibroblasts byDefined Factors, J. High Resolut. Chromatogr. Chromatogr.Commun., 2007, vol. 131, pp. 861–872.
Thomson, J.A., Itskovitz-Eldor, J., Shapiro, S.S., et al.,Embryonic Stem Cell Lines Derived from Human Blasto-cysts, Science, 1998, vol. 282, pp. 1145–1147.
Vallier, L., Alexander, M., and Pedersen, R.A., Activin/Nodaland FGF Pathways Cooperate to Maintain Pluripotency ofHuman Embryonic Stem Cells, J. Cell Sci., 2005, vol. 118,pp. 4495–4509.
Vincent, S.D., Dunn, N.R., Hayashi, S., et al., Cell Fate Deci-sions within the Mouse Organizer are Governed by GradedNodal Signals, Genes Devel., 2003, vol. 17, pp. 1646–1662.
Watanabe, T and Miyazono, K., TGF-β Family Signaling inStem Cell Renewal and Differentiation, in The TGF-β Fam-ily, Derynck, R. and Miyazono, K., Eds., New York: ColdSpring Harbor Lab. Press, 2008, pp. 585–611.
Wernig, M., Meissner, A., Foreman, R., et al., In vitro Repro-gramming of Fibroblasts into a Pluripotent ES-Cell-LikeState, Nature, 2007, vol. 448, pp. 318–24.
Wiater, E. and Vale, W., Activins and Inhibins, in The TGF-βFamily, Derynck, R. and Miyazono, K., Eds., New York:Cold Spring Harbor Lab. Press, 2008, pp. 79–120.
Xu, R.-H., Sampsell-Barron, T.L., Gu, F., et al., Nanog is aDirect Target of TGF/Activin-Mediatewd SMAD Signalingin Human ESCs, Cell Stem Cell, 2008, vol. 3, pp. 196–206.
Xu, R.H., Peck, R.M., Li, D.S., et al., Basic FGF and Sup-pression of BMP Signaling Sustain Undifferentiated Prolif-eration of Human ES Cells, Nat. Methods, 2005, vol. 2,pp. 185–190.
Ying, Q.L., Nichols, J., Chambers, I., and Smith, A., BMPInduction of Id Proteins Suppresses Differentiation and Sus-tains Embryonic Stem Cell Self-Renewal in Collaborationwith STAT3, Cell, 2003, vol. 115, pp. 281–292.
Yu, J., Vodyanik, M., Smuga-Otoo, K., et al., Induced Pluri-potent Stem Cell Lines Derived from Human Somatic Cells,Science, 2007, vol. 318, pp. 1917–1920.