regeneration of fatcells from myofibroblasts during …...report regeneration regeneration of...
TRANSCRIPT

REPORT◥
REGENERATION
Regeneration of fat cells frommyofibroblasts during wound healingMaksim V. Plikus,1,2* Christian F. Guerrero-Juarez,2 Mayumi Ito,3 Yun Rose Li,4
Priya H. Dedhia,5 Ying Zheng,1 Mengle Shao,6 Denise L. Gay,1,7 Raul Ramos,2
Tsai-Ching Hsi,2 Ji Won Oh,2,8 Xiaojie Wang,2 Amanda Ramirez,2 Sara E. Konopelski,2
Arijh Elzein,2 Anne Wang,1 Rarinthip June Supapannachart,1 Hye-Lim Lee,2
Chae Ho Lim,3 Arben Nace,1 Amy Guo,1 Elsa Treffeisen,1 Thomas Andl,9
Ricardo N. Ramirez,2 Rabi Murad,2 Stefan Offermanns,10 Daniel Metzger,11
Pierre Chambon,12 Alan D. Widgerow,13 Tai-Lan Tuan,14 Ali Mortazavi,2
Rana K. Gupta,6 Bruce A. Hamilton,15 Sarah E. Millar,1 Patrick Seale,4,16
Warren S. Pear,5 Mitchell A. Lazar,4,17 George Cotsarelis1*
Although regeneration through the reprogramming of one cell lineage to another occursin fish and amphibians, it has not been observed in mammals.We discovered in the mouse thatduring wound healing, adipocytes regenerate from myofibroblasts, a cell type thought to bedifferentiated and nonadipogenic. Myofibroblast reprogramming required neogenic hairfollicles, which triggered bone morphogenetic protein (BMP) signaling and then activation ofadipocyte transcription factors expressed during development. Overexpression of the BMPantagonist Noggin in hair follicles or deletion of the BMP receptor in myofibroblasts preventedadipocyte formation. Adipocytes formed from human keloid fibroblasts either when treatedwithBMPorwhenplacedwith humanhair follicles in vitro.Thus,we identify themyofibroblast asa plastic cell type that may be manipulated to treat scars in humans.
Wound healing in adult humans andmicegenerally results in a scar with excesscollagen and an absence of hair fol-licles and cutaneous fat. Recently, weand others discovered that a large skin
wound in an adult mouse often regenerates hairfollicles under the control of the Wnt and fibro-blast growth factor (FGF) pathways (fig. S1) (1–6).We then noticed new adipocytes within healedwounds that are indistinguishable from normalcutaneous fat cells in terms of size, density, anddepth from the skin surface (Fig. 1, A and B, andfig. S1C). The new adipocytes never form in thehairless part of the wound but develop exclusivelyaround new hair follicles (Fig. 1B and fig. S1C).Regenerated hair follicles begin to form around14 days postwounding, after reepithelialization(2). The first new adipocytes (orange) appear at23 days, immediately adjacent to the growing hairfollicle (blue), and their number and size increaseover the next few days (Fig. 1A and fig. S2). The
new adipocytes are classifiable as physiologicallymature and metabolically active white adiposecells because they express fat tissue–specifichormones, resistin and adiponectin, detected aslacZ-positive cells in Retn-lacZ mice (7) (n = 9)(figs. S2 and S3A) and Adipoq-Cre;R26R mice(n = 8) (fig. S3B), respectively.Given the close spatial and temporal associa-
tion between regenerated hair follicles and re-generated fat, we asked whether hair folliclesare necessary to establish adipocyte precursors.To test for such precursors, we placed dermalcells from wounds in a culture that promotesadipocyte differentiation (8). Dermal cells fromwounds with regenerated hair follicles differ-entiated into lipid-laden adipocytes, but dermalcells from wounds without hair follicles did not(Fig. 1C). Efficient in vitro differentiation andadipose gene up-regulation were consistentlyinduced from dermal cells that originated fromhair-bearing, but not hairless, wounds at different
postwounding time points, as early as 22 days(Fig. 1C and fig. S4).To determine the cellular origin of the new
adipocytes, we considered that during earlywoundrepair, dermal wound tissue containsmanymyo-fibroblasts expressing smoothmuscle actin.Myo-fibroblasts appear in large excisional wounds onday 5 and become abundant in the dermal scartissue by day 12. These cells largely cease to ex-press smooth muscle actin by day 17 (Fig. 2A).Cells expressing the adipogenic commitmentfactors ZFP423 (9, 10) (figs. S5B and S6A) andpCEBPb (figs. S5C and S6B) appear adjacent tothenewhair follicles at days 21 and24, respectively.This sequence suggests thatmyofibroblasts assumean alternative cell fate by converting to adipogenicprecursors. To test this, we examined the lineageidentity of new adipocytes in SM22-Cre;R26R andinducible SMA-CreERT2;R26Rmice, in which Creactivity turns on in wound myofibroblasts (Fig.2B and fig. S7, B to D). In normal depots of whitefat, SM22-Cre and SMA-CreERT2 are not activatedin adipocytes (Fig. 2C and fig. S7A) (11). However,in SM22-Cre;R26R (n= 12) and SMA-CreERT2;R26Rmice induced during wound healing (n = 4), mostnew adipocytes in wounds expressed lacZ, indi-cating their origin from myofibroblasts (figs. S8and S11).To functionally validate a myofibroblast-to-
adipocyte transformation, we generated the loss-of-function SM22-Cre;Pparg f lox/f lox and inducibleSMA-CreERT2;Pparg flox/floxmice.Wounds of SM22-Cre;Pparg f lox/f loxmice formedmany new hair fol-licles but were nearly devoid of new adipocytes[n = 7; new adipocyte/follicle ratio, 0.62 ± 0.2,versus 24.1 ± 6.8 in control mice (n = 7)] (fig. S7Fand materials and methods). The difference be-tween experimental and control groupswasnearly40-fold (fig. S7E and table S1). Importantly, inSM22-Cre;Pparg f lox/f lox mice, depots of whitefat outside the wound, both subcutaneous andelsewhere, were unaffected. Similarly, tamoxifeninduction of SMA-CreERT2;Pparg flox/flox mice dur-ing early time points after wounding largelyprevented regeneration of adipocytes [n = 6;new adipocyte/follicle ratio, 0.5 ± 0.07, versus22.7 ± 5.1 in control mice (n = 6)] (Fig. 2, D andE). Taken together, our lineage tracing studiesestablish myofibroblasts as the source for newregenerating adipocytes (fig. S9F).To evaluate the possible contribution of other
SM22- or SMA-positive cell populations—includingvascular smooth muscle cells, panniculus carnosus
RESEARCH
Plikus et al., Science 355, 748–752 (2017) 17 February 2017 1 of 5
1Department of Dermatology, Kligman Laboratories, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, PA 19104, USA. 2Department of Developmental and Cell Biology,Sue and Bill Gross Stem Cell Research Center, Center for Complex Biological Systems, University of California, Irvine, Irvine, CA 92697, USA. 3The Ronald O. Perelman Department ofDermatology, Department of Cell Biology, New York University School of Medicine, New York, NY 10016, USA. 4The Institute for Diabetes, Obesity and Metabolism, Perelman School of Medicineat the University of Pennsylvania, Philadelphia, PA 19104, USA. 5Department of Pathology and Laboratory Medicine, Abramson Family Cancer Research Institute, Institute for Medicine andEngineering, University of Pennsylvania, Philadelphia, PA 19104, USA. 6Touchstone Diabetes Center, Department of Internal Medicine, The University of Texas Southwestern Medical Center,Dallas, TX 75390, USA. 7INSERM U967, Commissariat à L’énergie Atomique et aux Énergies Alternatives, Institut de Radiobiologie Cellulaire et Moléculaire, 92265 Fontenay-aux-Roses Cedex,France. 8Department of Anatomy, School of Medicine, Kyungpook National University, Daegu, Korea. 9Burnett School of Biomedical Sciences, University of Central Florida, Orlando, FL 328116,USA. 10Department of Pharmacology, Max Planck Institute for Heart and Lung Research, Bad Nauheim 61231, Germany. 11Institut de Génétique et de Biologie Moléculaire et Cellulaire, CNRSUMR7104, INSERM U964, Université de Strasbourg, Illkirch 67404, France. 12Institut de Génétique et de Biologie Moléculaire et Cellulaire, CNRS UMR7104, INSERM U964, Institut d’EtudesAvancées de l’Université de Strasbourg, Collège de France, Illkirch 67404, France. 13Center for Tissue Engineering, Department of Plastic Surgery, University of California, Irvine, Irvine, CA 92868,USA. 14The Saban Research Institute of Children’s Hospital Los Angeles and Department of Surgery, Keck School of Medicine, University of Southern California, Los Angeles, CA 90027, USA.15Departments of Medicine and Cellular and Molecular Medicine, Moores Cancer Center and Institute for Genomic Medicine, University of California, San Diego, La Jolla, CA 92093, USA.16Department of Cell and Developmental Biology, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, PA 19104, USA. 17Division of Endocrinology, Diabetes, and Metabolism,Department of Medicine, Perelman School of Medicine at the University of Pennsylvania, Philadelphia, PA 19104, USA.*Corresponding author. Email: [email protected] (M.V.P.); [email protected] (G.C.)
on April 1, 2020
http://science.sciencem
ag.org/D
ownloaded from

muscle, and dermal papillae of new hair follicles—to fat regeneration, we traced the progeny of thesecell populations by using relevant promoter sys-
tems.We found no contribution of these cell typesto new adipocytes (figs. S9 and S10 and supple-mentary text S1).
To comprehensively study themolecular natureof lineage reprograming of myofibroblasts to adi-pocytes in wounds in adult mice, we profiled the
Plikus et al., Science 355, 748–752 (2017) 17 February 2017 2 of 5
Fig. 1. New adipocytes onlyregenerate around new hairfollicles during woundhealing. (A) Histologicalsections (left two panels) andwhole-mount images (rightthree panels) of skin stained todetect follicular epithelium(blue) and adipocytes (orange)in K14-Cre;R26R mice at indi-cated postwounding days. Newadipocytes (arrowheads)increase in number and sizeover several days. (B) Skinviewed from the undersurface.New adipocytes form and per-sist exclusively around regener-ated hair follicles, which arise inthe center of the wound.(C) Cultured dermal cellsisolated from wounds withregenerated hair follicles differ-entiated into BODIPY-positive(green) adipocytes, whereascultured dermal cells fromwounds lacking follicles formedno adipocytes. Scale bars in (A)and (C), 20 mm; in (B), 1 mm.
Fig. 2. New adipo-cytes originate fromwound myofibro-blasts. (A) Smoothmuscle actin (SMA)–positive myofibro-blasts are present atthe wound edge andthen in the healingwound (arrows). Byday 17, dermal woundcells express very lit-tle SMA, but vascularsmooth muscle cellsremain labeled(arrow). (B) Lineagetracing of myofibro-blasts results in lacZ(blue)–expressingregenerated adipo-cytes (orange, whitearrowheads). (C) Adi-pocytes in normalskin are not labeled.(D and E) Deletion ofPparg in myofibro-blasts resulted innear-complete loss ofnew adipocytes,whereas normal cuta-neous adipocytes atthe wound edge remained intact. Scale bars in (A), 100 mm; in (B) (left), (D), and (E), 1 mm; in (B) (center), 200 mm; in (B) (right), 50 mm; in (C), 200 mm.
RESEARCH | REPORTon A
pril 1, 2020
http://science.sciencemag.org/
Dow
nloaded from
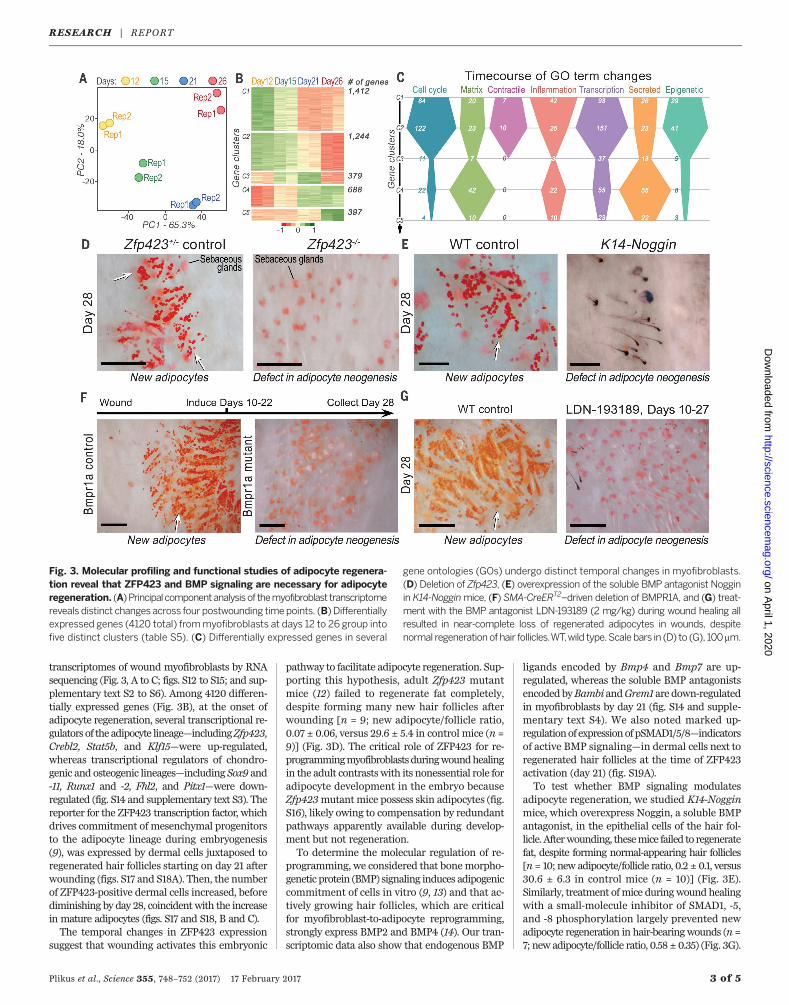
transcriptomes of wound myofibroblasts by RNAsequencing (Fig. 3, A to C; figs. S12 to S15; and sup-plementary text S2 to S6). Among 4120 differen-tially expressed genes (Fig. 3B), at the onset ofadipocyte regeneration, several transcriptional re-gulators of the adipocyte lineage—includingZfp423,Crebl2, Stat5b, and Klf15—were up-regulated,whereas transcriptional regulators of chondro-genic and osteogenic lineages—including Sox9 and-11, Runx1 and -2, Fhl2, and Pitx1—were down-regulated (fig. S14 and supplementary text S3). Thereporter for the ZFP423 transcription factor, whichdrives commitment of mesenchymal progenitorsto the adipocyte lineage during embryogenesis(9), was expressed by dermal cells juxtaposed toregenerated hair follicles starting on day 21 afterwounding (figs. S17 and S18A). Then, the numberof ZFP423-positive dermal cells increased, beforediminishing by day 28, coincidentwith the increasein mature adipocytes (figs. S17 and S18, B and C).The temporal changes in ZFP423 expression
suggest that wounding activates this embryonic
pathway to facilitate adipocyte regeneration. Sup-porting this hypothesis, adult Zfp423 mutantmice (12) failed to regenerate fat completely,despite forming many new hair follicles afterwounding [n = 9; new adipocyte/follicle ratio,0.07 ± 0.06, versus 29.6 ± 5.4 in control mice (n =9)] (Fig. 3D). The critical role of ZFP423 for re-programmingmyofibroblastsduringwoundhealingin the adult contrastswith its nonessential role foradipocyte development in the embryo becauseZfp423mutant mice possess skin adipocytes (fig.S16), likely owing to compensation by redundantpathways apparently available during develop-ment but not regeneration.To determine the molecular regulation of re-
programming, we considered that bonemorpho-genetic protein (BMP) signaling induces adipogeniccommitment of cells in vitro (9, 13) and that ac-tively growing hair follicles, which are criticalfor myofibroblast-to-adipocyte reprogramming,strongly express BMP2 and BMP4 (14). Our tran-scriptomic data also show that endogenous BMP
ligands encoded by Bmp4 and Bmp7 are up-regulated, whereas the soluble BMP antagonistsencodedbyBambi andGrem1 are down-regulatedin myofibroblasts by day 21 (fig. S14 and supple-mentary text S4). We also noted marked up-regulationof expressionofpSMAD1/5/8—indicatorsof active BMP signaling—in dermal cells next toregenerated hair follicles at the time of ZFP423activation (day 21) (fig. S19A).To test whether BMP signaling modulates
adipocyte regeneration, we studied K14-Nogginmice, which overexpress Noggin, a soluble BMPantagonist, in the epithelial cells of the hair fol-licle.Afterwounding, thesemice failed to regeneratefat, despite forming normal-appearing hair follicles[n= 10; new adipocyte/follicle ratio, 0.2 ± 0.1, versus30.6 ± 6.3 in control mice (n = 10)] (Fig. 3E).Similarly, treatment ofmice duringwound healingwith a small-molecule inhibitor of SMAD1, -5,and -8 phosphorylation largely prevented newadipocyte regeneration in hair-bearingwounds (n =7; new adipocyte/follicle ratio, 0.58 ± 0.35) (Fig. 3G).
Plikus et al., Science 355, 748–752 (2017) 17 February 2017 3 of 5
Fig. 3. Molecular profiling and functional studies of adipocyte regenera-tion reveal that ZFP423 and BMP signaling are necessary for adipocyteregeneration. (A)Principalcomponentanalysisof themyofibroblast transcriptomereveals distinct changes across four postwounding timepoints. (B) Differentiallyexpressed genes (4120 total) frommyofibroblasts at days 12 to 26 group intofive distinct clusters (table S5). (C) Differentially expressed genes in several
gene ontologies (GOs) undergo distinct temporal changes in myofibroblasts.(D) Deletion of Zfp423, (E) overexpression of the soluble BMP antagonist Nogginin K14-Nogginmice, (F) SMA-CreERT2–driven deletion of BMPR1A, and (G) treat-ment with the BMP antagonist LDN-193189 (2 mg/kg) during wound healing allresulted in near-complete loss of regenerated adipocytes in wounds, despitenormal regeneration of hair follicles.WT,wild type.Scale bars in (D) to (G), 100 mm.
RESEARCH | REPORTon A
pril 1, 2020
http://science.sciencemag.org/
Dow
nloaded from
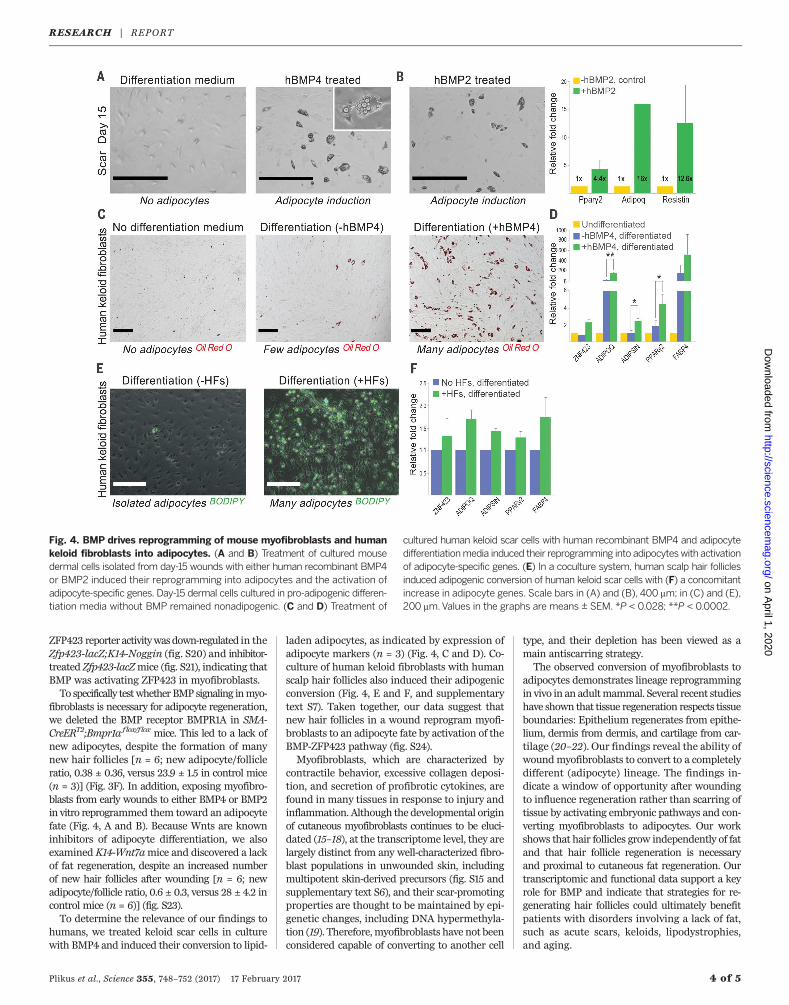
ZFP423 reporter activitywasdown-regulated in theZfp423-lacZ;K14-Noggin (fig. S20) and inhibitor-treated Zfp423-lacZmice (fig. S21), indicating thatBMP was activating ZFP423 in myofibroblasts.To specifically testwhetherBMP signaling inmyo-
fibroblasts is necessary for adipocyte regeneration,we deleted the BMP receptor BMPR1A in SMA-CreERT2;Bmpr1a flox/f lox mice. This led to a lack ofnew adipocytes, despite the formation of manynew hair follicles [n = 6; new adipocyte/follicleratio, 0.38 ± 0.36, versus 23.9 ± 1.5 in control mice(n = 3)] (Fig. 3F). In addition, exposing myofibro-blasts from early wounds to either BMP4 or BMP2in vitro reprogrammed them toward an adipocytefate (Fig. 4, A and B). Because Wnts are knowninhibitors of adipocyte differentiation, we alsoexamined K14-Wnt7amice and discovered a lackof fat regeneration, despite an increased numberof new hair follicles after wounding [n = 6; newadipocyte/follicle ratio, 0.6 ± 0.3, versus 28 ± 4.2 incontrol mice (n = 6)] (fig. S23).To determine the relevance of our findings to
humans, we treated keloid scar cells in culturewith BMP4 and induced their conversion to lipid-
laden adipocytes, as indicated by expression ofadipocyte markers (n = 3) (Fig. 4, C and D). Co-culture of human keloid fibroblasts with humanscalp hair follicles also induced their adipogenicconversion (Fig. 4, E and F, and supplementarytext S7). Taken together, our data suggest thatnew hair follicles in a wound reprogram myofi-broblasts to an adipocyte fate by activation of theBMP-ZFP423 pathway (fig. S24).Myofibroblasts, which are characterized by
contractile behavior, excessive collagen deposi-tion, and secretion of profibrotic cytokines, arefound in many tissues in response to injury andinflammation. Although the developmental originof cutaneous myofibroblasts continues to be eluci-dated (15–18), at the transcriptome level, they arelargely distinct from any well-characterized fibro-blast populations in unwounded skin, includingmultipotent skin-derived precursors (fig. S15 andsupplementary text S6), and their scar-promotingproperties are thought to be maintained by epi-genetic changes, including DNA hypermethyla-tion (19). Therefore,myofibroblasts have not beenconsidered capable of converting to another cell
type, and their depletion has been viewed as amain antiscarring strategy.The observed conversion of myofibroblasts to
adipocytes demonstrates lineage reprogrammingin vivo in an adultmammal. Several recent studieshave shown that tissue regeneration respects tissueboundaries: Epithelium regenerates from epithe-lium, dermis from dermis, and cartilage from car-tilage (20–22). Our findings reveal the ability ofwoundmyofibroblasts to convert to a completelydifferent (adipocyte) lineage. The findings in-dicate a window of opportunity after woundingto influence regeneration rather than scarring oftissue by activating embryonic pathways and con-verting myofibroblasts to adipocytes. Our workshows that hair follicles grow independently of fatand that hair follicle regeneration is necessaryand proximal to cutaneous fat regeneration. Ourtranscriptomic and functional data support a keyrole for BMP and indicate that strategies for re-generating hair follicles could ultimately benefitpatients with disorders involving a lack of fat,such as acute scars, keloids, lipodystrophies,and aging.
Plikus et al., Science 355, 748–752 (2017) 17 February 2017 4 of 5
Fig. 4. BMP drives reprogramming of mouse myofibroblasts and humankeloid fibroblasts into adipocytes. (A and B) Treatment of cultured mousedermal cells isolated from day-15 wounds with either human recombinant BMP4or BMP2 induced their reprogramming into adipocytes and the activation ofadipocyte-specific genes. Day-15 dermal cells cultured in pro-adipogenic differen-tiation media without BMP remained nonadipogenic. (C and D) Treatment of
cultured human keloid scar cells with human recombinant BMP4 and adipocytedifferentiationmedia induced their reprogramming into adipocyteswith activationof adipocyte-specific genes. (E) In a coculture system, human scalp hair folliclesinduced adipogenic conversion of human keloid scar cells with (F) a concomitantincrease in adipocyte genes. Scale bars in (A) and (B), 400 mm; in (C) and (E),200 mm.Values in the graphs are means ± SEM. *P < 0.028; **P < 0.0002.
RESEARCH | REPORTon A
pril 1, 2020
http://science.sciencemag.org/
Dow
nloaded from

REFERENCES AND NOTES
1. D. Gay et al., Nat. Med. 19, 916–923 (2013).2. M. Ito et al., Nature 447, 316–320 (2007).3. A. W. Seifert et al., Nature 489, 561–565 (2012).4. X. Wang et al., Regeneration 2, 169–181 (2015).5. C. Breedis, Cancer Res. 14, 575–579 (1954).6. R. E. Billingham, P. S. Russell, Nature 177, 791–792 (1956).7. R. R. Banerjee et al., Science 303, 1195–1198 (2004).8. J. Ishibashi et al., Mol. Cell. Biol. 32, 2289–2299 (2012).9. R. K. Gupta et al., Nature 464, 619–623 (2010).10. S. Kang et al., PLOS Biol. 10, e1001433 (2012).11. W. Tang et al., Science 322, 583–586 (2008).12. W. A. Alcaraz et al., Proc. Natl. Acad. Sci. U.S.A. 103,
19424–19429 (2006).13. A. Hammarstedt et al., Proc. Natl. Acad. Sci. U.S.A. 110,
2563–2568 (2013).14. H. Kulessa, G. Turk, B. L. Hogan, EMBO J. 19, 6664–6674
(2000).15. R. R. Driskell et al., Nature 504, 277–281 (2013).16. S. Dulauroy, S. E. Di Carlo, F. Langa, G. Eberl, L. Peduto,
Nat. Med. 18, 1262–1270 (2012).17. H. Suga et al., Stem Cells 32, 1347–1360 (2014).18. Y. Rinkevich et al., Science 348, aaa2151 (2015).19. M. V. Plikus, C. F. Guerrero-Juarez, E. Treffeisen, D. L. Gay,
Exp. Dermatol. 24, 167–170 (2015).20. J. A. Lehoczky, B. Robert, C. J. Tabin, Proc. Natl. Acad. Sci. U.S.A.
108, 20609–20614 (2011).21. Y. Rinkevich, P. Lindau, H. Ueno, M. T. Longaker, I. L. Weissman,
Nature 476, 409–413 (2011).
22. T. H. Leung, E. R. Snyder, Y. Liu, J. Wang, S. K. Kim, Genes Dev.29, 2097–2107 (2015).
ACKNOWLEDGMENTS
Funding is provided by U.S. NIH grant R01-AR055309, NIH SkinDiseases Research Core grant P30-AR057217, and the Edward andFannie Gray Hall Center for Human Appearance. M.V.P. issupported by a pilot grant from the Diabetes and EndocrinologyResearch Center (University of Pennsylvania), a DermatologyFoundation research grant, an Edward Mallinckrodt Jr.Foundation grant, a Pew Charitable Trust grant, and NIH grantsR01-AR067273 and R01-AR069653. M.A.L. is supported by NIHgrant DK49210, M.I. by NIH grant R01-AR066022, S.E.M. by NIHgrant R37-AR047709 and Penn Skin Biology and DiseasesResource-based Core grant P30-AR069589, W.S.P. by NIH grantR01-AI047833, R.K.G. by NIH grant R01-DK104789, T.-L.T. by NIHgrant R01-GM095821, B.A.H. by NIH grant R01-NS05487, R.R. byCalifornia Institute for Regenerative Medicine training grant TG2-01152, C.F.G.-J. by the NSF Graduate Research Fellowship Program(DGE-1321846) and a training grant from MBRS-IMSD (Initiativefor Maximizing Student Development; GM055246), X.W. by aCanadian Institutes of Health Research postdoctoral fellowship(MFE-123724), J.W.O. by National Research Foundation of Korea grant2016R1C1B1015211, C.H.L. by the Cutaneous Biology and SkinDisease training program (T32-AR064184), Y.R.L. by a NIH NationalResearch Service Award F30 training grant and a Paul and DaisySoros Fellowship for New Americans, H.-L.L. by NIH T32 training grantT32-CA009054-37, and M.S. by American Heart Associationpostdoctoral fellowship 16POST26420136. Retn-lacZ mice were
generated with the Transgenic Mouse Core of the University ofPennsylvania Diabetes Research Center (NIH grant DK19525). Wethank Y. Mishina for providing Bmpr1aflox mice, C.-M. Chuong forproviding K14-Noggin mice, V. Scarfone and C. Tu for their assistancewith fluorescence-activated cell sorting and tissue culture, Z. Yang fortechnical assistance, and P. Sterling for reviewing the manuscript.SMA-CreERT2 mice are available from P.C. under a material transferagreement with the University of California, Irvine. P.C. and D.M. areinventors on patents EP 1 692 936 B1 and US 7112715 B2, held byGIE-CERBM (Centre Européen de Recherche en Biologie et Médecine),that cover the method for generating conditional DNA recombinationin mice by using the Cre-ERT2 fusion protein. M.V.P., C.F.G.-J., andG.C. are co-inventors on a patent application filed through the U.S.Patent and Trademark Office by the University of Pennsylvaniadescribing the BMP pathway as a target for promoting neogenic fatformation, among other claims.
SUPPLEMENTARY MATERIALS
www.sciencemag.org/content/355/6326/748/suppl/DC1Materials and MethodsSupplementary TextFigs. S1 to S24Tables S1 to S5References (23–109)Data S1
24 August 2016; accepted 19 December 2016Published online 5 January 201710.1126/science.aai8792
Plikus et al., Science 355, 748–752 (2017) 17 February 2017 5 of 5
RESEARCH | REPORTon A
pril 1, 2020
http://science.sciencemag.org/
Dow
nloaded from

Regeneration of fat cells from myofibroblasts during wound healing
CotsarelisMortazavi, Rana K. Gupta, Bruce A. Hamilton, Sarah E. Millar, Patrick Seale, Warren S. Pear, Mitchell A. Lazar and GeorgeRicardo N. Ramirez, Rabi Murad, Stefan Offermanns, Daniel Metzger, Pierre Chambon, Alan D. Widgerow, Tai-Lan Tuan, Ali Wang, Rarinthip June Supapannachart, Hye-Lim Lee, Chae Ho Lim, Arben Nace, Amy Guo, Elsa Treffeisen, Thomas Andl,L. Gay, Raul Ramos, Tsai-Ching Hsi, Ji Won Oh, Xiaojie Wang, Amanda Ramirez, Sara E. Konopelski, Arijh Elzein, Anne Maksim V. Plikus, Christian F. Guerrero-Juarez, Mayumi Ito, Yun Rose Li, Priya H. Dedhia, Ying Zheng, Mengle Shao, Denise
originally published online January 5, 2017DOI: 10.1126/science.aai8792 (6326), 748-752.355Science
, this issue p. 748; see also p. 693Scienceformation after wounding by adding BMP.
scar(BMP) signaling pathway and adipocyte transcription factors in the myofibroblast. Thus, it may be possible to reduce adipocytes (see the Perspective by Chan and Longaker). The hair follicles activated the bone morphogenetic proteinthat hair follicles in both mice and humans can convert myofibroblasts, the predominant dermal cell in a wound, into
now showet al.Wounding can leave scars, which are characterized by a lack of hair follicles and cutaneous fat. Plikus Although some animals easily regenerate limbs and heal broken flesh, mammals are generally not so gifted.
Hair follicles: Secret to prevent scars?
ARTICLE TOOLS http://science.sciencemag.org/content/355/6326/748
MATERIALSSUPPLEMENTARY http://science.sciencemag.org/content/suppl/2017/01/04/science.aai8792.DC1
CONTENTRELATED
http://stke.sciencemag.org/content/sigtrans/10/506/eaao1822.fullhttp://science.sciencemag.org/content/sci/355/6326/693.fullhttp://stke.sciencemag.org/content/sigtrans/9/457/ra118.full
REFERENCES
http://science.sciencemag.org/content/355/6326/748#BIBLThis article cites 109 articles, 40 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
is a registered trademark of AAAS.ScienceScience, 1200 New York Avenue NW, Washington, DC 20005. The title (print ISSN 0036-8075; online ISSN 1095-9203) is published by the American Association for the Advancement ofScience
Copyright © 2017, American Association for the Advancement of Science
on April 1, 2020
http://science.sciencem
ag.org/D
ownloaded from