red rice (oryza sativa) cross-resistance to imidazolinone herbicides used in resistant rice...
TRANSCRIPT

Pesticide Biochemistry and Physiology 105 (2013) 177–183
Contents lists available at SciVerse ScienceDirect
Pesticide Biochemistry and Physiology
journal homepage: www.elsevier .com/locate /pest
Red rice (Oryza sativa) cross-resistance to imidazolinone herbicides usedin resistant rice cultivars grown in northern Greece
Nikolaos S. Kaloumenos a,1, Nieves Capote b, Ana Aguado b, Ilias G. Eleftherohorinos a,⇑a Laboratory of Agronomy, School of Agriculture, Aristotle University of Thessaloniki, 541 24 Thessaloniki, Greeceb IFAPA Las Torres-Tomejil Carretera Sevilla-Cazalla Km 12.2, 41200 Sevilla, Spain
a r t i c l e i n f o a b s t r a c t
Article history:Received 27 March 2012Accepted 29 January 2013Available online 8 February 2013
Keywords:ALS gene sequencingALS-inhibitor resistanceResistant red riceTarget-site mutationIntraspecific gene flow
0048-3575/$ - see front matter � 2013 Elsevier Inc. Ahttp://dx.doi.org/10.1016/j.pestbp.2013.01.008
Abbreviations: ALS, acetolactate synthase; Clearfietant rice; RF, resistance factor.⇑ Corresponding author. Fax: +30 2310 998634.
E-mail address: [email protected] (I.G. Elefther1 Present address: Research Project Biologist, Syngen
Research Centre, Bracknell, RG42 6EY, UK.
A putative resistant red rice (Oryza sativa) accession, an imidazolinone-resistant rice cultivar (Clearfield),a susceptible red rice accession and a susceptible rice cultivar were evaluated for cross-resistance toimazamox and imazethapyr in a whole-plant response experiment and seed bioassay. Additionally, a210-bp fragment of the ALS gene was sequenced to identify mutations responsible for resistance. Also,a 574 bp of the ALS gene was sequenced and PCR for detection of the ‘Clearfield allele’ was conductedby the Andalusian Institute of Agricultural Research and Training (IFAPA, Spain). In the whole-plantresponse experiment, the putative resistant red rice was >23 and >21 times more resistant to imazamoxand imazethapyr than the susceptible accession, respectively, whereas the respective resistance factorvalues based on seed bioassay were 86.4 and 141.7. Also, the respective resistance factor values forthe Clearfield rice cultivar were similar with those calculated for putative resistant red rice. Additionally,the sequence of the 210 bp ALS gene fragment from the putative resistant red rice and Clearfield rice cul-tivar revealed the same amino acid substitution of Ser653Asn in both alleles (homozygous). Furthermore,the sequence of a 574 bp ALS gene fragment and the PCR for detection of the ‘Clearfield allele’ confirmedthat the putative resistant red rice is homozygous mutant for the Ser653Asn mutation and provided addi-tional evidence that its genetic background matches that of Clearfield rice. These findings indicate clearlythat the insufficient control of the putative resistant red rice with imazamox was due to target-site resis-tance and particularly due to a point mutation at the Ser653 codon, which is positively identified as hav-ing been derived from the Clearfield rice cultivar.
� 2013 Elsevier Inc. All rights reserved.
1. Introduction
Rice (Oryza sativa) is one of the most important food crops inthe world and is widely cultivated in Greece [1]. Red rice is oneof the most important weeds in almost all regions of the worldwhere rice is grown including the southern United States, Japan,central and southern America, central Africa, India, China, northernAustralia and recently Italy and Greece [1–4]. Its presence in thefield results in great yield losses and reduction of the commercialquality of the rice grains [5,6].
Red rice and cultivated rice are both classified as O. sativa andare genetically similar, a fact that makes impossible red rice selec-tive control with herbicides in rice [5,7,8]. However, the introduc-tion of imidazolinone-resistant rice (Clearfield rice) in 2002 in the
ll rights reserved.
ld rice, imidazolinone-resis-
ohorinos).ta, Jealott’s Hill International
US, made selective control of red rice possible with the use of imi-dazolinone herbicides; potent inhibitors of the acetolactate syn-thase (ALS, EC 2.2.1.6), also referred as acetohydroxyacidsynthase (AHAS) [9–11]. It is worth noting that close to one-thirdof the rice cultivated area in the state of Arkansas was planted withClearfield rice in 2008 and about 22% of the rice cultivated area inCosta Rica, representing 60% of the weedy-rice-infested area, wasplanted with Clearfield rice in 2006/2007, only 2 years after itwas introduced [12].
Nevertheless, the Achilles’ heel of the imidazolinone herbicides,as of all ALS-inhibiting herbicides, is their propensity to rapidly se-lect for resistant weed biotypes or populations [13]. To date, resis-tant biotypes or populations to ALS-inhibiting herbicides havebeen identified in 129 weed species [14]. In most cases this isdue to point mutations in one of the following codons: Ala122,Pro197, Ala205, Asp376, Arg377, Trp574, Ser653, and Gly654 ofthe ALS gene, numbered based on the sequence of Arabidopsis tha-liana (L.) Heynh [15,16].
Red rice tolerance to imidazolinones is due to point mutationsin the codons Ala122, Ser653 or Gly654 of the ALS gene [7,17,18].

178 N.S. Kaloumenos et al. / Pesticide Biochemistry and Physiology 105 (2013) 177–183
In southern Brazil the predominant mechanism of resistance in redrice was the Gly654Glu substitution in the ALS enzyme, howeverthe substitutions of Ser653Asn and Ala122Thr were also present[18]. Likewise, the Gly654Glu and Val669Met substitutions wereobserved in two red rice accessions collected from Arkansas(USA) prior to the widespread of the imidazolinone-resistant ricecultivars [7]. However, in Italy the only mechanism of resistancein red rice grown in Clearfield rice was the Ser653Asn substitutionof the ALS enzyme [19].
Red rice resistance can be due to selection of pre-existing resis-tance alleles (spontaneous mutations) or due to outcrossing of redrice survivors with the herbicide-resistant rice [10,17,19,20]. Out-crossing is possible because both red rice and cultivated rice vari-eties are diploids (2n = 24) in the ‘AA’ genome group that allowsrice-red rice hybridization [21–23]. In addition, the herbicide resis-tance trait in the Clearfield rice is governed by a single dominantnuclear gene and thus the heterozygous F1 hybrid from a cross be-tween Clearfield rice and non-resistant rice lines or susceptible redrice accessions will produce resistant plants [24,25]. This was con-firmed in natural outcrossing experiments conducted in the RiceResearch and Extension Center (Stuttgart, AR, US) between red riceand Clearfield rice cultivars, where the Ser653Asn (existing in theClearfield rice cultivars) substitution was found in the red riceprogeny; evidence that pollen mediated gene flow from Clearfieldrice cultivars to wild red rice populations took place and resultedin resistant hybrid red rice plants [17]. In southern Brazil, the highfrequency of the amino acid substitution Ser653Asn found in resis-tant red rice accessions suggested the occurrence of gene flow fromClearfield rice cultivars to survivor red rice plants. On the contrary,the presence of the novel amino acid substitutions Gly654Glu andAla122Thr in the same area suggested that resistance alleles werepresent in the red rice populations prior to the introduction ofClearfield rice [18]. Similarly in Arkansas (US), the presence ofthe mutations Gly654Glu and Val669Met in resistant red riceaccessions from different geographical regions, which are not pres-ent in the Clearfield varieties grown in the area, support thehypothesis that ALS resistant red rice has been evolved as a conse-quence of the herbicide selection pressure [7].
During the 2010 growing period, a rice grower from northernGreece (county of Thessaloniki) noticed unsatisfactory red ricecontrol after the application of the recommended rate of imazamoxin paddy rice field cultivated with Clearfield rice cultivar. Survivorswere not removed from the field by the farmer because of the highlabor cost in the area. In the Clearfield rice system that was intro-duced in 2006 in Greece, red rice is sprayed with the ALS-inhibitingherbicide imazamox two times (split application is stated in thefarmers’ contract as compulsory) or once (this is not supportedalthough some farmers prefer this) in every growing season. There-fore, the purpose of the present work was (1) to determine if thereduced red rice control was due to the evolution of resistance toimazamox and possibly cross-resistance to imazethapyr, and (2)to clarify the mechanism of resistance and especially the possibleexistence of point mutations responsible for resistance.
2. Materials and methods
2.1. Seed source
The seeds from the putative resistant red rice accession, suscep-tible red rice accession, Clearfield rice cultivar ‘Sirio’ and imidazoli-none susceptible rice cultivar ‘Claudio’ were provided by BASFHellas. The putative resistant red rice seeds originated from a ricefield located in northern Greece (county of Thessaloniki) and, as re-ported earlier, the field was cultivated for four out of the five con-secutive years with Clearfield rice cultivar against stewardship
recommendations. The seeds of the susceptible red rice accessionalso originated from an adjacent rice field, which had been culti-vated continuously with conventional (imidazolinone susceptible)rice cultivars and consequently had never been treated with anyimidazolinone herbicide for red rice control. During seed collec-tion, care was taken to obtain a representative sample from eachfield (accession). The collected red rice seeds were initially placedin big plastic bags and then transferred to laboratory where theywere air-dried, threshed, and placed in paper bags. Afterwards,the collected seeds along with the rice cultivar seeds were storedat 3–5 �C.
2.2. Whole-plant pot experiment
The imazamox and imazethapyr rate response experimentswere conducted at the farm of Aristotle University of Thessalonikiduring 2011. Experiments were carried out using 10 � 10 � 9 cm(0.9 L) plastic pots filled with a peat:sand mixture (3:1 by volume).Each pot was seeded at a depth of 1 cm with approximately 15 redrice or rice seeds. When seedlings reached the two-leaf stage theywere carefully thinned to six per pot, which corresponds to a ricefield density of 600 plants m�2. Herbicide applications were per-formed when red rice or rice plants were at the 13–14 Zadoksgrowth stage (three-to four-leaf stage). Herbicide treatments wereapplied with a portable field plot sprayer (AZO-SPRAYERS, P.O. Box350-6710 BJ EDE, The Netherlands) using a 2.4 m wide boom fittedwith six 8002 flat-fan nozzles (Teejet Spray System Co., P.O. Box7900, Wheaton, IL 60188) and calibrated to deliver 300 L ha�1 ofwater at 280 kPa pressure. All pots were randomly placed outdoorsin a net protected area and watered as needed. Pots were random-ized each week in order to achieve uniform growth conditions forall plants. Imazamox (BAS 720-06H, 4 SL BASF, BASF Agro HellasLtd., Greece) and imazethapyr (BAS 685-08H, 10 SL BASF, BASFAgro Hellas Ltd., Greece) were applied at 40, 80, 160, 320,640 g ai ha�1 and 35, 70, 140, 280, 560 g ai ha�1, respectivelyagainst the putative resistant red rice accession and Clearfield ricecultivar. Imazamox is recommended in Greece to be applied at 35–50 g ai ha�1 (corresponding to 0.875–1.25 L ha�1 of Pulsar 4 SL)when rice and weed plants are at the 2–3 leaf stage, but the treat-ment has to be repeated after three weeks. Alternatively, it is rec-ommended to be applied at 80 g ai ha�1 (corresponding to2.0 L ha�1 of Pulsar 4 SL) when rice and grass weed plants are atthe beginning of tillering. Imazethapyr is not registered in Greeceand the selected rates were taken from published scientific papers.The respective rates of each herbicide applied against the S red riceaccession and S rice cultivar were 5, 10, 20, 40, 80 g ai ha�1 and 4.4,8.8, 70, 17.5, 35 g ai ha�1. All imazamox and imazethapyr treat-ments were applied in mixture with 0.5% v/v of the adjuvant DashHC (crop oil, 37% w/w methyl oleate palmitate + 5% w/w oleicacid + 22% w/w fat alcohol polyalkoxylate phosphate). Addition-ally, treatments with 125 g ai quizalofop ha�1 and 300 g ai cycloxy-dim ha�1 as chemical controls along with an untreated control foreach herbicide by red rice accession or herbicide by rice cultivarcombination were also included. Control of red rice accessions orrice cultivars was assessed by determining the survival rate andthe aboveground fresh weight of all survived plants in each pot4 weeks after treatment (WAT). Data were expressed as percentageof the untreated control for each red rice accession or rice cultivar.The experiment was conducted twice (two independent experi-ments replicated in time) using a completely randomized designwith four replications.
2.3. Seed bioassay
The putative resistant red rice accession, the susceptible red riceaccession, the Clearfield rice cultivar ‘Sirio’ and the imidazolinone

N.S. Kaloumenos et al. / Pesticide Biochemistry and Physiology 105 (2013) 177–183 179
susceptible rice cultivar ‘Claudio’ were tested in a 8.5-cm diameterplastic disposable Petri dish seed bioassay for cross-resistance toimazamox and imazethapyr. Eight uniform pre-germinated seedswere placed on top of a double-layer filter paper lined in Petridishes. The filter paper was then carefully moistened with 5 mLof imazamox or imazethapyr (0.01, 0.1, 1, 10, 100 mg ai L�1) freshlyprepared solution, whereas the untreated control Petri dishes weremoistened with 5 mL of distilled water. Petri dishes were shieldedwith plastic membrane to retain moisture and placed randomly indark plastic bags. Then, the bags were placed in a growth chamber(incubator) at 26–28 �C for 7 days. At the end of the incubation per-iod, the seedlings were carefully removed from the Petri dishes andthe root length of the seedlings was measured and expressed aspercentage of the untreated control for each red rice accession orrice cultivar. Four Petri dishes (replications) were used for eachherbicide concentration in a completely randomized design andeach seed bioassay was performed twice (two independent exper-iments replicated in time).
2.4. Statistical analysis
Data obtained from the whole-plant response experiment wereanalyzed (ANOVA) separately for the putative resistant red riceaccession and Clearfield rice cultivar and the susceptible red riceaccession or susceptible rice cultivar using a 2 � 5 � 2 [2 herbi-cides � 5 herbicide treatments (relative rate) � 2 (red rice acces-sion and rice cultivar)] factorial approach combined over time(two experiments). The quizalofop and cycloxydim treatmentswere not included in the ANOVA. For both experiments, the com-bined over two independent experiments ANOVA was performedbecause the Bartlett’s test indicated that the error variances ofthe data obtained from the two experiments (time replicated) werehomogeneous. Differences among treatment means were com-pared at the 5% level of significance using the LSD test. Becausethe ANOVAs indicated no significant treatment by time (twoexperiments) interaction, means were averaged over twoexperiments.
In the seed bioassay, a separate combined over two indepen-dent experiments 5 � 4 [5 herbicide concentrations � 4 (2 red riceaccessions plus 2 rice cultivars)] factorial approach was used forthe ANOVA of imazamox or imazethapyr. The combined over twoexperiments ANOVA was performed for each herbicide becausethe Bartlett’s test indicated that the error variances of the data ob-tained from the two experiments (time replicated) were homoge-neous. Because the ANOVAs indicated no significant treatment bytime (two experiments) interaction, data were averaged over twoexperiments.
The untransformed fresh weight and the root length data (ex-pressed as % of the untreated control) combined over the whole-plant response experiment and seed bioassay, respectively, wereused for regression analysis [26]. In particular, the y-axis data[fresh weight (g) or root length (cm)] was regressed against thex-axis data [herbicide rate (g ai ha�1) or concentration (mg ai L�1)]in order to estimate the amount of each herbicide required for 50%growth reduction (GR50) of red rice accession or rice cultivar freshweight or root length. The original fresh weight or root length datawere subjected to nonlinear regression analysis using the log–logistic equation 1 [27].
y ¼ C þ D� C1þ expfb½logðxÞ � logðGR50Þ�g
ð1Þ
where C = the lower limit, D = the upper limit, b = the slope at theGR50, and GR50 = the herbicide rate (g ai ha�1) or herbicide concen-tration (mg ai L�1) required for 50% reduction of the fresh weight orroot length. In this regression equation, the herbicide rate or con-
centration was the independent variable (x) and the fresh weight(g) or root length (cm) was the dependent variable (y). Additionally,the random distribution of the residuals was checked using the runstest, which indicated no violations of randomness at P < 0.05. Thelevels of resistance for all red rice accessions and rice cultivars toeach herbicide were determined by the resistance factor (RF) value,which was calculated as the GR50 of the putative resistant red riceaccession or Clearfield rice cultivar divided by the respective valueof the susceptible red rice accession or susceptible rice cultivar.
2.5. Amplification and sequencing of the 210 bp ALS gene fragment
DNA was extracted from 100 mg ground leaf tissue (four indi-vidual plants for each accession or cultivar) using the Cetyl trime-thylammonium bromide (CTAB) method according to the protocoloutlined in the NucleoSpin� Plant II kit (MACHEREY NAGEL GmbH& Co. KG. Postfach 10 13 52. D-52313 Düren, Germany). The testedPR red rice accession and IR rice cultivar plants used for plant tis-sue collection had survived the treatment of imazamox at the rec-ommended rate (80 g ai ha�1). For the S red rice accession and ricecultivar, plant tissue was collected from untreated plants. Theamplification of the entire ALS gene (2101 bp) from the genomicDNA extracted from PR red rice, R rice cultivar and S plants wasachieved using the forward 50-GTAAGAACCACCAGCGACACC-30
(Up-01) and the reverse 50-GATGCATATGCCTACGGAAAAC-30
(Down-01) primers. The polymerase chain reaction (PCR) mixtureconsisted of 0.25 mM of each primer, 200 mM of each deoxyribo-nucleotide triphosphate (dNTP), 1.5 mM MgCl2, 5% dimethyl sulf-oxide (DMSO), 4 mL of the supplied 10� thermophilic buffer,2 mL DNA sample, and 1 enzyme units (U) Thermus aquaticus(Taq) polymerase (DNA polymerase, recombinant, HyTest Ltd.20520 Turku, Finland) in 40 mL. Amplifications were carried outin a heated lid PCR machine (MJ Research, model PTC-200), withthe following cycle: denature at 95 �C for 5 min, denature at95 �C for 30 s, anneal at 60 �C for 30 s, elongate at 72 �C for 120 s,and cycle to second denaturation step 34 more times. A final elon-gation step was performed at 72 �C for 5 min. The resulting prod-ucts (named P1) were stored at �20 �C to be used as a templatefor subsequent PCR. The amplification of the ALS gene fragment(210 bp) from the PCR product P1 was achieved using the forward50-CCTGCAGTCCGTGTAACAAA-30 (Up-02) and the reverse 50-GTGCTTTGCCAACATACAGATT-30 (Down-02) primers. The ampli-fied ALS gene fragment contained the Ser653 and Gly654 codonswhere mutations responsible for red rice resistance to imidazoli-none herbicides have been reported. Also, the Ser653Asn aminoacid substitution is the resistant-bearing mutation in the Clear-field™ varieties commercialized in the area. The polymerase chainreaction (PCR) mixture consisted of 0.25 lM of each primer,200 lM of each deoxyribonucleotide triphosphate (dNTP),1.5 mM MgCl2, 5% dimethyl sulfoxide (DMSO), 4 ll of the supplied10� thermophilic buffer, 4 ll DNA sample, and 1 enzyme units (U)T. aquaticus (Taq) polymerase in 120 (3 � 40 mL). Amplificationswere carried out in a heated lid PCR machine (MJ Research, modelPTC-200), with the following cycle: denature at 95 �C for 5 min,denature at 95 �C for 30 s, anneal at 59 �C for 30 s, elongate at72 �C for 45 s, and cycle to second denaturation step 34 moretimes. A final elongation step was performed at 72 �C for 5 min.Afterwards, the PCR product, was separated on 1.3% agarose gelsand purified according to the protocol outlined in the NucleoSpin�
Extract II kit (MACHEREY NAGEL GmbH & Co. KG. D-52313 Düren,Germany). The purified DNA was sent immediately for sequencingto the University of Thessaly, School of Medicine, Department ofImmunology and Histocompatibility (Medical school campus, P.C.411 10 Greece). Each PCR product was sequenced twice, once withthe forward primer Up-02 and once with the reverse primer Down-02. The sequencing chromatograms were visualized and edited

180 N.S. Kaloumenos et al. / Pesticide Biochemistry and Physiology 105 (2013) 177–183
using BioEdit 7. The same software was used to translate the DNAsequences to the respective peptide sequences. Finally, the DNAand peptide sequences were aligned and compared using ClustalX2 [28].
2.6. Sequence of a 574 bp ALS gene fragment and PCR for detection ofthe Clearfield allele
The two ALS gene analyses were performed by IFAPA for bothour putative resistant and susceptible red rice accessions.
2.6.1. Red rice and rice samplesSamples consisted of seeds from ‘Puntal’ (wild rice-susceptible
reference line), ‘Sirio’ (Clearfield reference line, Ser653Asn), Greeksusceptible red rice accession, and Greek putative resistant red riceaccession were used. After removing the husk, rice and red riceseeds were homogenized in a mortar without liquid nitrogen.DNA extractions were performed from 100 mg of homogenizedmaterial using the Isolate Plant DNA minikit (Bioline). DNA con-centrations were measured in the Nanodrop.
2.6.2. Sequence of a 574 bp ALS gene fragmentThe polymerase chain reaction (PCR) mixture contained 200 ng
DNA, 1� HiFi reaction buffer, 1 mM deoxyribonucleotide triphos-phate (dNTP), 2 mM MgCl2, 0.3% dimethyl sulfoxide (DMSO),0.5 lM primer OS653F, 0.5 lM primer OS653R, 1 U velocity DNApolymerase (Bioline). The PCR amplification consisted of one cycleof 7 min at 98 �C, followed by 35 cycles of 30 s at 98 �C, 30 s at60 �C, 30 s at 72 �C and a final extension of 7 min at 72 �C. ThePCR products were run in 1% agarose gel. The corresponding bands(574 bp) were excised and purified using the FavorPrep™ Gel/PCRPurification kit (FAVORGEN). DNA concentrations of the purifiedbands were measured in the Nanodrop. The PCR products were se-quenced using the OS653F and OS653R primers, respectively bySTABVIDA company (Portugal). Sequences were analyzed by MEGA5.0 program.
2.6.3. PCR for detection of the ‘Clearfield allele’Extraction of DNA from the seed samples was made as de-
scribed above. The PCR reaction mixture contained 25 ng DNA,1� immolase reaction buffer, 0.2 mM dimethyl sulfoxide (dNTP),2 mM MgCl2, 0.5 lM primer OS653F, 0.5 lM primer OS653R,0.5 lM primer OS653WF or OS653MF, 0.5 U immolase DNA poly-merase (Bioline). The PCR amplification consisted of one cycle of7 min at 94 �C, followed by 35 cycles of 30 s at 94 �C, 30 s at60 �C, 1 min at 72 �C and a final extension of 7 min at 72 �C. ThePCR products were run in 1% agarose gel. Distilled water was usedas negative control.
Table 1Whole-plant response. Estimated imazamox and imazethapyr GR50 and resistance factor (Raccession (PR-RR) and one Clearfield rice cultivar (Cl-R) compared with the respective sus
Herbicide Accession or cultivar GR50a (95% CL)
—g ai ha�1—Imazamox PR-RR >640 (26% growth r
S-RR 27.8 (25.7–29.3)Cl-R >640 (27% growth rS-R 19.5 (18.4–20.6)
Imazethapyr PR-RR >560 (8% growth reS-RR 26.8 (24.2–29.8)Cl-R 560 (13% growth reS-R 26.5 (25.5–27.8)
a GR50, herbicide concentration for 50% reduction of red rice accession or rice cultivarb RF (resistance factor) = GR50 (PR-RR or Cl-R) � GR50(respective S-RR or S-R).
3. Results and discussion
3.1. Whole-plant response
The fresh weight of susceptible red rice accession and suscepti-ble rice cultivar was reduced by 96% and 95% due to recommendedrate of imazamox, whereas the respective reduction due to imaz-ethapyr was 94% and 95% (data not shown). Additionally, the sur-vival rate for both susceptible red rice accession and susceptiblerice cultivar due to herbicides was lower than 2% (data not shown).Nevertheless, the same rates of imazamox and imazethapyr re-sulted in unsatisfactory control of the putative resistant red riceaccession and the Clearfield rice cultivar. In particular, the applica-tion of the recommended rate of imazamox reduced their freshweight only by 6.4% and 15.8% respectively, whereas the respectivefresh weight reduction by the recommended rate of imazethapyrwas only 4.1% and 3.0%. The application of the eight times higherthan the recommended rate of imazamox reduced fresh weightof putative resistant red rice accession and Clearfield rice cultivarby 25.7% and 27.5%, respectively. Also, the eight times higher thanthe recommended rate of imazethapyr resulted in fresh weightreduction by 7.9% and 12.6% in putative resistant red rice accessionand Clearfield rice cultivar, respectively. In all cases, the survivalrate for the putative resistant red rice accession and Clearfield ricecultivar was similar to the untreated control (data not shown).
The use of the nonlinear regression analysis indicated that freshweight and herbicide concentration were described with the high-er approximation efficacy (judged by the low residual mean squarevalues), suggesting that log–logistic equation was the best fit forthe data related with the red rice accession or rice cultivars andherbicide concentrations (Table 1). In particular, the calculatedGR50 values [herbicide application rate (g ai ha�1) required for50% reduction of fresh weight] for the susceptible red rice acces-sion and susceptible rice cultivar were 27.8 and 19.5 g ai ha�1 forimazamox whereas for imazethapyr the respective values were26.8 and 26.5 g ai ha�1. For the putative resistant red rice accessionand the Clearfield rice cultivar the respective values could not beaccurately calculated and were higher than 640 g ai ha�1 forimazamox (this rate reduced the fresh weight of the putative resis-tant red rice accession and the Clearfield rice cultivar only by 26%and 27%, respectively) and higher than 560 g ai ha�1 for imazetha-pyr (this rate reduced the fresh weight of the putative resistant redrice accession and the Clearfield rice cultivar only by 8% and 13%,respectively) (Table 1). The RF values for the putative resistantred rice accession and the Clearfield rice cultivar were higher than23 and 33 for imazamox, whereas the respective RF values forimazethapyr were higher than 21 for both putative resistant redrice accession and Clearfield rice cultivar.
These data reveal that the putative resistant red rice accessionevaluated in the whole-plant response experiment has developed
F) values, as well as residual mean squares (Res. MS) for one putative resistant red riceceptible red rice accession (S-RR) and susceptible rice cultivar (S-R).
Slope (SE) RFb Res. MS
eduction) >235.28 (0.33) – 16.1
eduction) >334.77 (0.25) – 14.6
duction) >214.39 (0.36) – 24.0
duction) >216.65 (0.52) – 18.0
fresh weight.

N.S. Kaloumenos et al. / Pesticide Biochemistry and Physiology 105 (2013) 177–183 181
resistance to imazamox and cross-resistance to imazethapyr. Sim-ilarly, in other studies the whole-plant response method showedthat 56% of the uncontrolled populations of red rice from rice fieldscultivated under the Clearfield system in the state of Rio Grande doSul were resistant to the herbicide mixture imazethapyr + imaza-pic (75 + 25 g ai L�1), but the determined RF values were lowerthan the respective imazethapyr RF values of this study and rangedfrom 5.2 to 11.6 [29]. Also, in large scale screening experimentscarried out in Arkansas, three out of 130 red rice accessions werefound 9.0 to 21.4 times more resistant to imazamox and alsocross-resistant to imazethapyr (RF values 16.5 to 48), imazapyr(RF values 6.3 to 25.7), and imazaquin (RF values 63.7 to 166.1)[30]. These RF values were lower than the respective values forthe Clearfield rice cultivars that ranged for imazamox from 13.2to 107.3, for imazethapyr from 41.3 to >176, for imazapyr from69 to 274, and for imazaquin from 27.8 to 461.1. The differentmechanism of resistance could be the reason for these differencesas the evaluated red rice accessions had been collected from fieldsnever planted with Clearfield rice cultivars and thus no gene flowcould have taken place [30]. On the contrary, in the present study,the similar resistance factor values observed in the whole-plant re-sponse experiment for the putative resistant red rice accession andthe Clearfield rice cultivar suggest that these two plant speciescould possess the same mechanism of resistance to imidazolinoneherbicides, which may have resulted from a possible gene flowfrom the Clearfield rice cultivars to red rice accession.
3.2. Seed bioassay
The use of the nonlinear regression analysis indicated that rootlength and herbicide concentration were described with the higherapproximation efficacy (judged by the low residual mean squarevalues), suggesting that log–logistic equation was the best fit forthe data related with the red rice accession or rice cultivars andherbicide concentrations (Table 2). The use of the 95% confidencelimits showed differences in the GR50 values calculated from thisequation. In particular, the imazamox GR50 values for the putativeresistant red rice accession and Clearfield rice cultivar were 28.5and 34.6 mg ai L�1, which were significantly higher than therespective GR50 values for the susceptible red rice accession andsusceptible rice cultivar (0.33 and 0.38 mg ai L�1). The imazethapyrGR50 values for the putative resistant red rice accession and Clear-field rice cultivar were 14.7 and 17.0 mg ai L�1, which were alsosignificantly higher than the respective GR50 values (0.10 and0.12 mg ai L�1) for the susceptible red rice accession and suscepti-ble rice cultivar. The calculated resistance factor (RF) values forimazamox were 86.4 and 91.1 for the respective putative resistantred rice accession and Clearfield rice cultivar, whereas the respec-tive RF values for imazethapyr were 147.0 and 141.7 (Table 2).
Table 2Seed bioassay. Estimated imazamox and imazethapyr GR50 and resistance factor (RF) vaaccession (PR-RR) and one Clearfield rice cultivar (Cl-R) compared with the respective sus
Herbicide Accession or cultivar GR50a (95% CL)
—mg ai L�1—Imazamox PR-RR 28.5 (24.4–33.6
S-RR 0.33 (0.21–0.5Cl-R 34.6 (29.1–39.4S-R 0.38 (0.23–0.6
Imazethapyr PR-RR 14.7 (11.8–17.8S-RR 0.10 (0.07–0.2Cl-R 17.0 (14.4–20.5S-R 0.12 (0.06–0.2
a GR50, herbicide concentration for 50% reduction of red rice accession or rice cultivarb RF (resistance factor) = GR50 (PR-RR or Cl-R) � GR50(respective S-RR or S-R).
These seed bioassay data confirmed the whole-plant responseexperiment outcomes and clearly showed that the putative resis-tant red rice accession was resistant to imazamox and cross-resis-tant to imazethapyr. Similar seed bioassay studies conducted byothers [30] indicated RF values for three red rice accessions lower(15.8 to 30.9) than the respective RF values in our study, whichcould be attributed to different mechanism of resistance involved.Likewise, analogous seed bioassays were also used successfully todiscriminate rice cultivars with resistance to the imidazolinoneherbicide mixture imazethapyr + imazapic [31]. Additionally, aseed bioassay based on shoot length of each seedling measured10 days after treatment showed that Clearfield rice cultivar resis-tance factor to imazethapyr was 63.8 [24].
The similar GR50 and RF values calculated for both putativeresistant red rice accession and Clearfield rice cultivar using seedbioassays, along with the respective similar whole-plant responseexperiment values suggest similar ALS point mutation resistancefor red rice that may have resulted from a possible Clearfield geneflow.
3.3. Amplification and sequencing of the ALS gene fragment
The 210 bp fragment of ALS sequenced from each individualplant was aligned with the susceptible rice (O. sativa L., japonicacultivar-group) ALS gene obtained from GenBank (Accession num-ber: AY885674) and with the respective resistant A. thaliana ALSgene (Accession number: X51514). The obtained analysis of theDNA sequence chromatograms for the susceptible red rice acces-sion and susceptible rice cultivar revealed a Ser (AGT) at position653 in the plants examined (Table 3). In contrast, the DNA se-quence analysis for the putative resistant red rice accession andthe Clearfield rice cultivar revealed a substitution of the secondbase guanine (G) by adenine (A) at the same codon (Ser653), whichresulted in a substitution of the amino acid Ser653 (AGT) by Asn(AAT). Moreover, the fact that the point mutation was present inboth alleles, suggests for both putative resistant red rice accessionand Clearfield rice cultivar homozygous plant genotypes (R/R). Fur-ther analysis of the 210-bp fragment sequence alignment did notreveal any other nucleotide polymorphisms (data not shown),whereas search for other nucleotide polymorphisms in other re-gions of the ALS gene was not made.
The sequence of the 574 bp ALS fragment conducted by IFAPAwas aligned with the susceptible rice (O. sativa L., japonica culti-var-group) ALS gene obtained from GenBank (Accession number:AY885674), the respective resistant A. thaliana ALS gene (Accessionnumber: X51514), the Clearfield reference line AHAS gene(‘93AS3510’ with Ser653-Asn and Gly654-Glu) and the red ricewild type ALS gene (susceptible reference). The obtained analysisof the DNA sequence chromatograms for the Greek susceptiblered rice accession and susceptible rice cultivar (‘Puntal’) revealed
lues, as well as residual mean squares (Res. MS) for one putative resistant red riceceptible red rice accession (S-RR) and susceptible rice cultivar (S-R).
Slope (SE) RFb Res. MS
) 1.53 (0.13) 86.4 23.99) 1.01 (0.15) – 93.7) 1.57 (0.07) 91.1 4.90) 0.95 (0.11) – 40.8) 1.82 (0.16) 147.0 27.41) 1.51 (0.82) – 91.7) 1.72 (0.13) 141.7 9.60) 1.09 (0.24) – 41.3
root length.
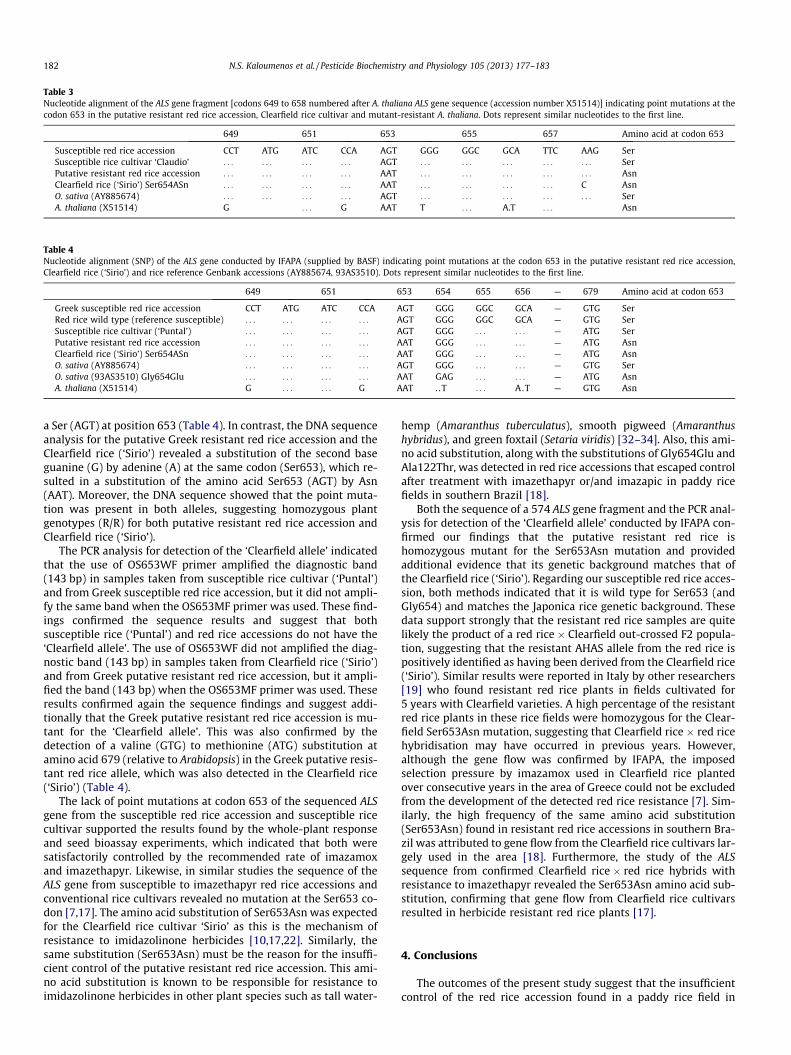
Table 3Nucleotide alignment of the ALS gene fragment [codons 649 to 658 numbered after A. thaliana ALS gene sequence (accession number X51514)] indicating point mutations at thecodon 653 in the putative resistant red rice accession, Clearfield rice cultivar and mutant-resistant A. thaliana. Dots represent similar nucleotides to the first line.
649 651 653 655 657 Amino acid at codon 653
Susceptible red rice accession CCT ATG ATC CCA AGT GGG GGC GCA TTC AAG SerSusceptible rice cultivar ‘Claudio’ . . . . . . . . . . . . AGT . . . . . . . . . . . . . . . SerPutative resistant red rice accession . . . . . . . . . . . . AAT . . . . . . . . . . . . . . . AsnClearfield rice (‘Sirio’) Ser654ASn . . . . . . . . . . . . AAT . . . . . . . . . . . . C AsnO. sativa (AY885674) . . . . . . . . . . . . AGT . . . . . . . . . . . . . . . SerA. thaliana (X51514) G . . . G AAT T . . . A.T . . . Asn
Table 4Nucleotide alignment (SNP) of the ALS gene conducted by IFAPA (supplied by BASF) indicating point mutations at the codon 653 in the putative resistant red rice accession,Clearfield rice (‘Sirio’) and rice reference Genbank accessions (AY885674, 93AS3510). Dots represent similar nucleotides to the first line.
649 651 653 654 655 656 — 679 Amino acid at codon 653
Greek susceptible red rice accession CCT ATG ATC CCA AGT GGG GGC GCA — GTG SerRed rice wild type (reference susceptible) . . . . . . . . . . . . AGT GGG GGC GCA — GTG SerSusceptible rice cultivar (‘Puntal’) . . . . . . . . . . . . AGT GGG . . . . . . — ATG SerPutative resistant red rice accession . . . . . . . . . . . . AAT GGG . . . . . . — ATG AsnClearfield rice (‘Sirio’) Ser654ASn . . . . . . . . . . . . AAT GGG . . . . . . — ATG AsnO. sativa (AY885674) . . . . . . . . . . . . AGT GGG . . . . . . — GTG SerO. sativa (93AS3510) Gly654Glu . . . . . . . . . . . . AAT GAG . . . . . . — ATG AsnA. thaliana (X51514) G . . . . . . G AAT . .T . . . A.T — GTG Asn
182 N.S. Kaloumenos et al. / Pesticide Biochemistry and Physiology 105 (2013) 177–183
a Ser (AGT) at position 653 (Table 4). In contrast, the DNA sequenceanalysis for the putative Greek resistant red rice accession and theClearfield rice (‘Sirio’) revealed a substitution of the second baseguanine (G) by adenine (A) at the same codon (Ser653), which re-sulted in a substitution of the amino acid Ser653 (AGT) by Asn(AAT). Moreover, the DNA sequence showed that the point muta-tion was present in both alleles, suggesting homozygous plantgenotypes (R/R) for both putative resistant red rice accession andClearfield rice (‘Sirio’).
The PCR analysis for detection of the ‘Clearfield allele’ indicatedthat the use of OS653WF primer amplified the diagnostic band(143 bp) in samples taken from susceptible rice cultivar (‘Puntal’)and from Greek susceptible red rice accession, but it did not ampli-fy the same band when the OS653MF primer was used. These find-ings confirmed the sequence results and suggest that bothsusceptible rice (‘Puntal’) and red rice accessions do not have the‘Clearfield allele’. The use of OS653WF did not amplified the diag-nostic band (143 bp) in samples taken from Clearfield rice (‘Sirio’)and from Greek putative resistant red rice accession, but it ampli-fied the band (143 bp) when the OS653MF primer was used. Theseresults confirmed again the sequence findings and suggest addi-tionally that the Greek putative resistant red rice accession is mu-tant for the ‘Clearfield allele’. This was also confirmed by thedetection of a valine (GTG) to methionine (ATG) substitution atamino acid 679 (relative to Arabidopsis) in the Greek putative resis-tant red rice allele, which was also detected in the Clearfield rice(‘Sirio’) (Table 4).
The lack of point mutations at codon 653 of the sequenced ALSgene from the susceptible red rice accession and susceptible ricecultivar supported the results found by the whole-plant responseand seed bioassay experiments, which indicated that both weresatisfactorily controlled by the recommended rate of imazamoxand imazethapyr. Likewise, in similar studies the sequence of theALS gene from susceptible to imazethapyr red rice accessions andconventional rice cultivars revealed no mutation at the Ser653 co-don [7,17]. The amino acid substitution of Ser653Asn was expectedfor the Clearfield rice cultivar ‘Sirio’ as this is the mechanism ofresistance to imidazolinone herbicides [10,17,22]. Similarly, thesame substitution (Ser653Asn) must be the reason for the insuffi-cient control of the putative resistant red rice accession. This ami-no acid substitution is known to be responsible for resistance toimidazolinone herbicides in other plant species such as tall water-
hemp (Amaranthus tuberculatus), smooth pigweed (Amaranthushybridus), and green foxtail (Setaria viridis) [32–34]. Also, this ami-no acid substitution, along with the substitutions of Gly654Glu andAla122Thr, was detected in red rice accessions that escaped controlafter treatment with imazethapyr or/and imazapic in paddy ricefields in southern Brazil [18].
Both the sequence of a 574 ALS gene fragment and the PCR anal-ysis for detection of the ‘Clearfield allele’ conducted by IFAPA con-firmed our findings that the putative resistant red rice ishomozygous mutant for the Ser653Asn mutation and providedadditional evidence that its genetic background matches that ofthe Clearfield rice (‘Sirio’). Regarding our susceptible red rice acces-sion, both methods indicated that it is wild type for Ser653 (andGly654) and matches the Japonica rice genetic background. Thesedata support strongly that the resistant red rice samples are quitelikely the product of a red rice � Clearfield out-crossed F2 popula-tion, suggesting that the resistant AHAS allele from the red rice ispositively identified as having been derived from the Clearfield rice(‘Sirio’). Similar results were reported in Italy by other researchers[19] who found resistant red rice plants in fields cultivated for5 years with Clearfield varieties. A high percentage of the resistantred rice plants in these rice fields were homozygous for the Clear-field Ser653Asn mutation, suggesting that Clearfield rice � red ricehybridisation may have occurred in previous years. However,although the gene flow was confirmed by IFAPA, the imposedselection pressure by imazamox used in Clearfield rice plantedover consecutive years in the area of Greece could not be excludedfrom the development of the detected red rice resistance [7]. Sim-ilarly, the high frequency of the same amino acid substitution(Ser653Asn) found in resistant red rice accessions in southern Bra-zil was attributed to gene flow from the Clearfield rice cultivars lar-gely used in the area [18]. Furthermore, the study of the ALSsequence from confirmed Clearfield rice � red rice hybrids withresistance to imazethapyr revealed the Ser653Asn amino acid sub-stitution, confirming that gene flow from Clearfield rice cultivarsresulted in herbicide resistant red rice plants [17].
4. Conclusions
The outcomes of the present study suggest that the insufficientcontrol of the red rice accession found in a paddy rice field in

N.S. Kaloumenos et al. / Pesticide Biochemistry and Physiology 105 (2013) 177–183 183
northern Greece after the application of imazamox was due toevolved target-site resistance to this herbicide. The presence ofthe same point mutation in the ALS gene (corresponds ALS enzymewith Ser653Asn and Val679Met substitutions) extracted from bothClearfield rice cultivar (‘Sirio’) and Greek resistant red rice acces-sion suggests the occurrence of gene flow from the Clearfield rice(‘Sirio’) grown in the area to the putative resistant red rice acces-sion studied. However, as the identity of the rice plant cannot beconfirmed as red rice based solely on the sequence informationpresent, the phenotypic characteristics of the plant or molecularfingerprinting would be needed. These findings highlight a seriousthreat against the sustainability of the Clearfield technology in thearea. Therefore, in order to ensure the continued success of thistechnology in controlling red rice in paddy rice fields, the steward-ship measures mandatory in the use of Clearfield technology arerequired as obligation to be adhered by the Clearfield rice growersin Greece in order to reduce the occurrence in rice fields of suchresistant red rice hybrids and in the case of their presence mea-sures should be taken for their removal before seed maturity. Morespecifically, the new growers of Clearfield cultivar in rice monocul-ture fields should remove by hands or by glyphosate rope-wickapplicator all the red rice survived plants during the first and sec-ond growing season after imazamox application, as well as adhereto the strict split application technique of imazamox herbicide asdefined in the Clearfield rice stewardship protocol of BASF. In thethird growing season, the farmers should grow conventional ricecultivar in the same rice fields but in combination with false seed-ing for red rice control (cycloxydim, clethodim or glyphosate couldbe used for this purpose). In the fourth and fifth growing seasons,Clearfield rice cultivar could be grown in the same fields but againperforming the same procedure used in the first two growing sea-sons. Concerning the rice areas where crop rotation could be ap-plied, this practice should become compulsory. Moreover, inaddition to the previously mentioned measures, field monitoringfor red rice occurrence and further testing for resistance develop-ment should also become an obligatory measure.
Acknowledgments
The authors would like to thank BASF Hellas, 449 MesogeionAvenue, 153 43 Agia Paraskevi – Athens, Greece, for kindly provid-ing the seeds of the red rice accessions, the commercial rice culti-vars, the herbicides used, the red rice data of the sequence analysisconducted by IFAPA and for funding this research.
References
[1] I.G. Eleftherohorinos, K.V. Dhima, Red rice (Oryza sativa) control in rice withpreemergence and postemergence herbicides, Weed Technol. 16 (2002) 537–540.
[2] N.R. Burgos, J.K. Norsworthy, R.C. Scott, K.L. Smith, Red rice (Oryza sativa)status after 5 years of imidazolinone-resistant rice technology in Arkansas,Weed Technol. 22 (2008) 200–208.
[3] V.K. Shivrain, N.R. Burgos, M.M. Anders, S.N. Rajguru, J. Moore, M.A. Sales, Geneflow between Clearfield™ rice and red rice, Crop Prot. 26 (2007) 349–356.
[4] R.J. Smith, Weed thresholds in southern US rice Oryza sativa, Weed Technol. 2(1988) 232–241.
[5] I.G. Eleftherohorinos, K.V. Dhima, I.B. Vasilakoglou, Interference of red rice inrice grown in Greece, Weed Sci. 50 (2002) 167–172.
[6] R.J. Smith, Control of red rice (Oryza sativa) in water-seeded rice (O. sativa),Weed Sci. 29 (1981) 663–666.
[7] M.A. Sales, V.K. Shivrain, N.R. Burgos, Y.I. Kuk, Amino acid substitutions in theacetolactate synthase gene of red rice (Oryza sativa) confer resistance toImazethapyr, Weed Sci. 56 (2008) 485–489.
[8] D.J. Pantone, J.B. Baker, Reciprocal yield analysis of red rice (Oryza sativa)competition in cultivated rice, Weed Sci. 39 (1991) 42–47.
[9] L.A. Avila, D.J. Lee, S.A. Senseman, G.N. McCauley, J.M. Chandler, J.T. Cothern,Assessment of acetolactate synthase (ALS) tolerance to imazethapyr in red riceecotypes (Oryza spp.) and imidazolinone tolerant/resistant rice (Oryza sativa)varieties, Pest Manag. Sci. 61 (2005) 171–178.
[10] V.K. Shivrain, N.R. Burgos, M.A. Sales, Y.I. Kuk, Polymorphisms in the ALS geneof weedy rice (Oryza sativa L.) accessions with different tolerance toimazethapyr, Crop Prot. 29 (2010) 336–341.
[11] E.P. Webster, J.A. Masson, Acetolactate synthase-inhibiting herbicides onimidazolinone-tolerant rice, Weed Sci. 49 (2001) 652–657.
[12] J. Gressel, B.E. Valverde, A strategy to provide long-term control of weedy ricewhile mitigating herbicide resistance transgene flow, and its potential use forother crops with related weeds, Pest Manag. Sci. 65 (2009) 723–731.
[13] P.J. Tranel, T.R. Wright, Resistance of weeds to ALS-inhibiting herbicides: whathave we learned?, Weed Sci 50 (2002) 700–712.
[14] I. Heap, International survey of herbicide resistant weeds. <http://www.weedscience.org/>, 2010 (accessed 14.09.12).
[15] D. Massa, B. Krenz, R. Gerhards, Target-site resistance to ALS-inhibitingherbicides in Apera spica-venti populations is conferred by documented andpreviously unknown mutations, Weed Res. 51 (2011) 294–303.
[16] P.J. Tranel, T.R. Wright, I.M. Heap, ALS mutations from herbicide-resistantweeds. <http://www.weedscience.org/>, 2012 (accessed 14.09.12).
[17] S.N. Rajguru, N.R. Burgos, V.K. Shivrain, J.McD. Steward, Mutations in the redrice ALS associated with resistance to imazethapyr, Weed Sci. 53 (2005) 567–577.
[18] A.C. Roso, A. Meroto, C.A. Delatorre, V.G. Menezes, Regional scale distributionof imidazolinone herbicide-resistant alleles in red rice (Oryza sativa L.)determined through SNP markers, Field Crops Res. 119 (2010) 175–182.
[19] M. Busconi, D. Rossi, C. Lorenzoni, G. Baldi, C. Fogher, Spread of herbicide-resistant weedy rice (red rice, Oryza sativa L.) after 5 years of Clearfield ricecultivation in Italy, Plant Biol. 14 (2012) 751–759.
[20] V.K. Shivrain, N.R. Burgos, M.A. Sales, A. Mauromoustakos, D.R. Gealy, K.L.Smith, H.L. Black, M. Jia, Factors affecting the outcrossing rate betweenClearfield™ rice and red rice (Oryza sativa), Weed Sci. 57 (2009) 394–403.
[21] D.R. Gealy, D.H. Mitten, J.N. Rutger, Gene flow between red rice (Oryza sativa)and herbicide resistant rice (O. sativa): implications for weed management,Weed Technol. 17 (2003) 627–645.
[22] S. Kadaru, W. Zhang, A. Yadav, J.H. Oard, Development and application ofallele-specific PCR assays for imazethapyr resistance in rice (Oryza sativa),Euphytica 160 (2008) 431–438.
[23] F. Wang, Q.H. Yuan, L. Shi, A large-scale field study of transgene flow formcultivated rice (Oryza sativa) to common wild rice (O. rufipogon) andbarnyardgrass (Echinochloa crus-galli), Plant Biotechnol. J. 4 (2006) 667–676.
[24] I. Wenefrida, H.S. Utomo, M.M. Meche, J.L. Nash, Inheritance of herbicideresistance in two germplasm lines of Clearfield rice (Oryza sativa L.), Can. J.Plant Sci. 87 (2007) 659–669.
[25] W. Zhang, S.D. Linscombe, E. Webster, Risk assessment of the transfer ofimazethapyr herbicide tolerance from Clearfield rice to red rice (Oryza sativa),Euphytica 152 (2006) 75–86.
[26] S.Z. Knezevic, J.S. Streibig, C. Ritz, Utilizing R software package for dose-response studies: the concept and data analysis, Weed Technol. 21 (2007)840–848.
[27] S.S. Seefeldt, J.E. Jensen, E.P. Fuerst, Log-logistic analysis of herbicide dose-response relationships, Weed Technol. 9 (1995) 218–227.
[28] J.D. Thompson, T.J. Gibson, F. Plewniak, F. Jeanmougin, D.G. Higgins, TheClustalX windows interface. flexible strategies for multiple sequencealignment aided by quality analysis tools, Nucleic Acids Res. 25 (1997)4876–4882.
[29] V.G. Menezes, C.H.P. Mariot, A. Kalsing, I.C.G.R. Goulart, Red rice (Oryza sativa)resistant to the herbicides imidazolinones, Planta Daninha 27 (2009) 1047–1052.
[30] Y.I. Kuk, N.R. Burgos, V.K. Shivrain, Natural tolerance to imazethapyr in red rice(Oryza sativa), Weed Sci. 56 (2008) 1–11.
[31] A.C. Roso, A. Merotto, C.A. Delatorre, Bioassays for diagnosis of resistance tothe herbicides imidazolinones in rice plants, Planta Daninha 28 (2010) 411–419.
[32] J. Laplante, I. Rajcan, F.J. Tardif, Multiple allelic forms of acetohydroxyacidsynthase are responsible for herbicide resistance in Setaria viridis, Theor. Appl.Genet. 119 (2009) 577–585.
[33] W.L. Patzoldt, P.J. Tranel, Multiple ALS mutations confer herbicide resistance inwaterhemp (Amaranthus tuberculatus), Weed Sci. 55 (2007) 421–428.
[34] C.M. Whaley, H.P. Wilson, J.H. Westwood, ALS resistance in several smoothpigweed (Amaranthus hybridus) biotypes, Weed Sci. 54 (2006) 828–832.